Embed Size (px)
Citation preview

Caveolin-1 is regulated by c-myc and suppresses c-myc-induced apoptosis
Terry L Timme1, Alexei Goltsov1, Salahaldin Tahir1, Likun Li1, Jianxiang Wang1,Chengzhen Ren1, Randal N Johnston3 and Timothy C Thompson*,1,2
1Scott Department of Urology, Baylor College of Medicine, 6560 Fannin St., Suite 2100, Houston, Texas, TX 77030, USA;2Departments of Cell Biology and Radiology, Baylor College of Medicine, One Baylor Plaza, Houston, Texas, TX 77030, USA;3Department of Biochemistry and Molecular Biology, University of Calgary, Calgary, Alberta, Canada, T2N 4N1
Recent data indicating that overexpression of caveolin-1as well as c-myc are relatively common features ofadvanced prostate cancer prompted us to test forpotential cooperative interactions between caveolin-1and c-myc that would be consistent with malignantprogression. We used the well-characterized Rat1Amy-cERTM cells to show that the caveolin-1 gene is down-regulated at the level of transcription by c-myc. Bymaintaining relatively high levels of caveolin-1 with anadenoviral vector or in stably transfected clones we showthat caveolin-1 can suppress c-myc-induced apoptosis.Further we established human prostate cancer cell lineswith the mycERTM construct and show that clones withincreased caveolin-1 are more resistant to myc-inducedapoptosis and have increased capacity for growth in softagar when c-myc is activated. Oncogene (2000) 19,3256 ± 3265.
Keywords: caveolin; c-myc; apoptosis; prostate cancer
Introduction
Genetic alterations that lead to increased c-myc levelsare exceedingly common in human malignancies (Marcuet al., 1992). The c-MYC protein is a transcriptionfactor that can dimerize with Max to regulate theexpression of numerous target genes (Dang, 1999;Grandori et al., 1997). The induced or repressedactivities of speci®c c-myc target genes involve theregulation of cell proliferation versus apoptosis whichare conditional responses to c-myc expression (Dang,1999; Evan and Littlewood, 1998). The regulation of c-myc-induced apoptosis is of critical importance to therole of c-myc in malignant progression as neutralizationof the apoptotic e�ects of c-myc is necessary forproliferative functions of c-myc to be fully manifest(Evan and Littlewood, 1998; Prendergast, 1999).Speci®c oncogenes such as bcl-2 can cooperate with c-myc in certain malignancies through their anti-apoptotice�ects (Adams and Cory, 1998).
We have recently identi®ed and characterized theexpression of a speci®c gene, caveolin-1 (Yang et al.,1998b), that appears to manifest some properties of bcl-2family genes within the context of prostate cancer(Thompson et al., 1999a,b). Caveolin-1 is a major
structural component of caveolae, invaginations of theplasma membrane and trans-Golgi network that havebeen implicated in sphingolipid-cholesterol transportand speci®c signal transduction pathways (Parton, 1996).We determined that caveolin-1 is overexpressed in bothmouse and human prostate cancer (Yang et al., 1998b)and is predictive of recurrence following surgery forpresumed localized disease (Yang et al., 1999). Using aseries of metastatic androgen-resistant mouse prostatecancer cells, clones that were stably transfected withantisense caveolin-1 we demonstrated reduced levels ofcaveolin-1 protein triggered apoptosis following with-drawal of androgen in vitro and in vivo (Nasu et al.,1998). Reintroduction of caveolin-1 with an adenovirusin a representative antisense clone substituted for thesurvival e�ects of testosterone in vitro (Nasu et al., 1998).Two independent reports have correlated overexpressionof caveolin-1 with a multidrug resistant phenotypeindependent of P-glycoprotein in human cancer cell linesfrom several di�erent tumor types (Lavie et al., 1998;Yang et al., 1998a). The recent reports associatingcaveolin-1 with drug resistance are generally consistentwith the survival promoting functions we have demon-strated in prostate cancer cells. Many drugs includingtaxol can activate speci®c signal transduction pathwaysand recent reports also indicate that induction ofapoptosis by agents that lead to intracellular Ca2+
elevation also activate speci®c signal transductioncascades (Srivastava et al., 1999). Numerous studieshave shown that caveolin-1/caveolae may regulatemultiple signal transduction pathways including mitogenand Ca2+ activated kinases in a cell- and context-speci®cfashion (reviewed by Shaul and Anderson, 1998).Therefore it's conceivable that apoptotic signals gener-ated by extracellular stimuli or potential oncogenesintersect with caveolin-1 mediated activities at the levelof signal transduction.
Since overexpression and ampli®cation of c-myc havebeen shown to be critical determinants of prostatecancer progression (Bubendorf et al., 1999; Jenkins etal., 1997; Nupponen et al., 1998; Qian et al., 1997; Satoet al., 1999; Thompson et al., 1989) we tested forpossible interactions between caveolin-1 and c-myc thatwould be relevant to prostate cancer progression. Theresults of our studies using multiple ®broblast modelsindicated that caveolin-1 is down-regulated by c-myc atthe level of transcription. This down-regulationrequired active protein synthesis suggesting an indirectmechanism. Further studies revealed that enforcedexpression of caveolin-1 suppressed c-myc-inducedapoptosis in di�erent cell types and under multipleconditions. Finally using a human prostate cancer cellmodel we show that caveolin-1 and c-myc cooperate to
Oncogene (2000) 19, 3256 ± 3265ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: TC Thompson, Scott Department of Urology,Baylor College of Medicine, 6560 Fannin St., Suite 2100, Houston,Texas, TX 77030, USAReceived 21 December 1999; revised 3 May 2000; accepted 3 May2000

generate increased numbers of soft agar colonies invitro.
Results
Caveolin-1 is down-regulated by c-myc
We ®rst considered whether caveolin-1 was regulatedby c-myc. The c-mycERTM fusion protein is a well-established system for identifying c-myc target genes(Eilers et al., 1991) which are either up- or down-regulated by c-myc. As reviewed by Dang (1999), morethan 30 c-myc regulated genes have been identi®ed andcorrelated with a myc-mediated phenotype such asapoptosis, proliferation or transformation. The numberof genes identi®ed as up-regulated by c-myc throughspeci®c DNA binding regions for MYC-MAX dimers(E-boxes) have been more numerous than thoserepressed by c-myc. However, it has been suggestedthat genes that are suppressed by c-myc may actuallybe more important in regard to myc-mediated pro-liferation and transformation than the genes that areup-regulated (Claassen and Hann, 1999; Facchini andPenn, 1998) yet, the mechanism(s) that underlie c-mycgene repression are poorly de®ned.
The activation of c-myc with 4-hydroxytamoxifen(OHT) in con¯uent serum-starved Rat1AmycERTM
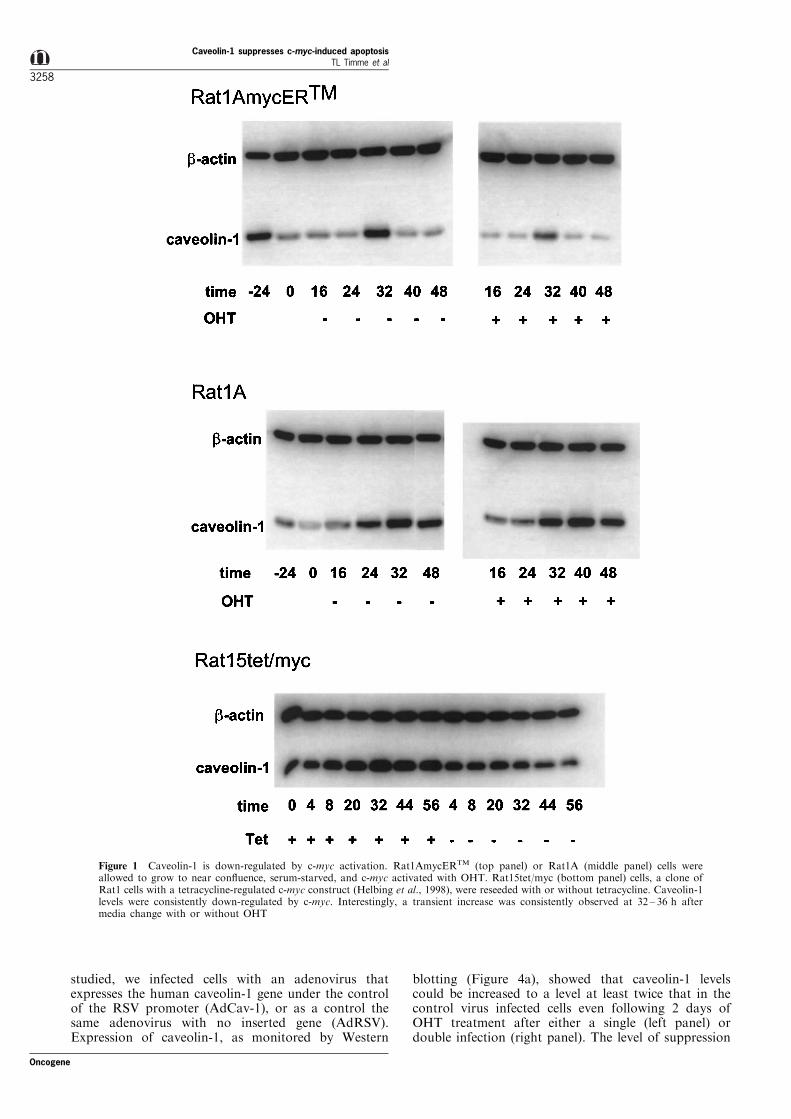
cells produced a time dependent decrease of caveolin-1 protein (Figure 1, top right panel). Serum deprivationwithout OHT also led to decreased levels of caveolin-1protein but to a lesser extent than with OHT (Figure 1,top left panel). Interestingly, a transient increase wasconsistently observed at 32 ± 36 h after media changewith or without OHT. The c-myc speci®city of thisresponse was con®rmed in parental Rat1A cells whichwhen treated similarly had no decrease in caveolin-1protein (Figure 1a, middle panel). In a parallelexperiment, induction of expression of c-myc from atetracycline repressed promoter in Rat15TetMyc cells(Helbing et al., 1998) also led to a time dependentdownregulation of caveolin-1 protein (Figure 1, bottompanel). These results con®rm that c-myc expressionleads to down-regulation of caveolin-1 protein. Theyfurther demonstrate that caveolin-1 protein levels areexquisitely sensitive to serum levels and nutrientconditions. Previous studies have also shown thatcaveolin-1 protein levels are regulated by factorscontained in serum (Galbiati et al., 1998; Lee et al.,1998).
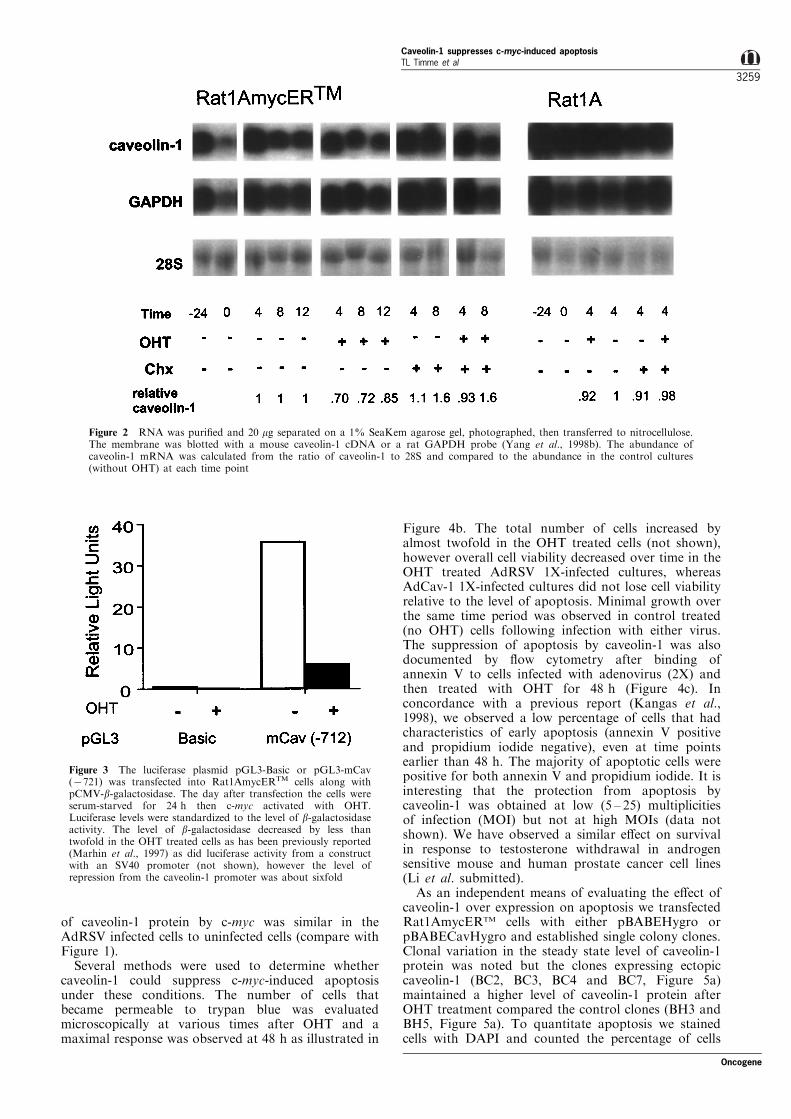
The identi®cation of c-myc target genes with theMyc-ER fusion protein is a well established system tocon®rm that candidate genes are activated or repressedat the mRNA level by c-myc in either a direct orindirect mechanism. In Figure 2 we document thatcaveolin-1 mRNA is down-regulated by c-myc instarved Rat1AmycERTM ®broblasts by activation of c-myc with OHT. Steady-state mRNA levels are low inserum starved cells (time 0) compared to cells nearcon¯uence in 10% serum (time 724) but transientlyincrease 4 h after replacing the media with fresh 0.1%serum, as observed with another gene transcriptionallydown-regulated by c-myc, H-ferritin (Wu et al., 1999).Caveolin-1 mRNA levels decrease at 8 and 12 hwithout or with OHT however treatment with OHTreduced the caveolin-1 mRNA levels by an additional
30% after 4 h relative to that in parallel cultureswithout OHT. The c-myc-induced down-regulation ofcaveolin-1 was dependent on new protein synthesissince cycloheximide treatment blocked OHT-induced c-myc down-regulation of caveolin-1 mRNA. Extendedtreatments (48 h) with cycloheximide were notanalysed due to extensive cell death consistent withprevious reports in Rat1AmycERTM cells (Evan et al.,1992; Ulrich et al., 1998).
To determine if the down-regulation of caveolin-1was the result of transcriptional regulation we cloned721 bases of DNA upstream of the mouse caveolin-1gene from a PAC library into a luciferase reporterplasmid and transfected it into Rat1AmycERTM cells.As shown in Figure 3 induction of c-myc in serumstarved cells leads to dramatically decreased promoteractivity in the caveolin-1 promoter-luciferase construct.The level of suppression of the caveolin-1 promoterwas about sixfold when normalized for transfectione�ciency with a co-transfected CMV-b-galactosidaseconstruct. Preliminary sequence analysis of the mousecaveolin-1 promoter region did not reveal a canonicalE-box and the published sequence of the humancaveolin-1 promoter (Bist et al., 1997) also does notcontain an E-box. One potential DNA sequenceinvolved in repression of genes by c-myc has beentermed Inr (Roy et al., 1993; Li et al., 1994), with theweak consensus sequence YYCAYYYYY. There is atleast one such site in the mouse caveolin-1 promoterhowever its functional signi®cance remains to beresolved. The e�ect of cycloheximide on caveolin-1mRNA levels suggests an indirect mechanism ofrepression by c-myc, however regulation of thecaveolin-1 gene is likely to be complex and regulatedby multiple factors in addition to c-myc. Furtherstudies will be required for elucidation of themechanism of repression.
It is of interest that decreased steady state levels ofcaveolin-1 protein have been reported in NIH3T3 cellstransformed with the oncogenes ras or abl (Koleske etal., 1995), neu (Engelman et al., 1997), or ras+myc(Yang et al., 1998b). In the case of ras-mediated down-regulation of caveolin-1 it is conceivable that theseactivities occur through increased stabilization of mycas has been demonstrated in REF52 cells (Sears et al.,1999). Overall, the data could suggest that down-regulation of caveolin-1 by c-myc creates a permissivestate for induction of apoptosis. Conversely if caveolin-1 levels were up-regulated as observed in prostatecancer (Nasu et al., 1998; Yang et al., 1998b, 1999) c-myc-induced apoptosis may be suppressed.
Caveolin-1 suppresses c-myc-induced apoptosis
To initially evaluate the potential role of caveolin-1 inapoptosis resistance we also used the well-characterizedRat1AmycERTM cells in which apoptosis can bereproducibly induced in con¯uent, serum-starved cellsby treatment with OHT leads to c-myc-inducedapoptosis (Eilers et al., 1989). In this system speci®ccytokines, insulin-like growth factor and plateletderived growth factor, have been shown to be able tosuppress apoptosis independent of their e�ects on cellcycle progression (Harrington et al., 1994). To ensurethat caveolin-1 expression could be enforced on cellsduring a time frame in which apoptosis could be
Oncogene
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3257
studied, we infected cells with an adenovirus thatexpresses the human caveolin-1 gene under the controlof the RSV promoter (AdCav-1), or as a control thesame adenovirus with no inserted gene (AdRSV).Expression of caveolin-1, as monitored by Western
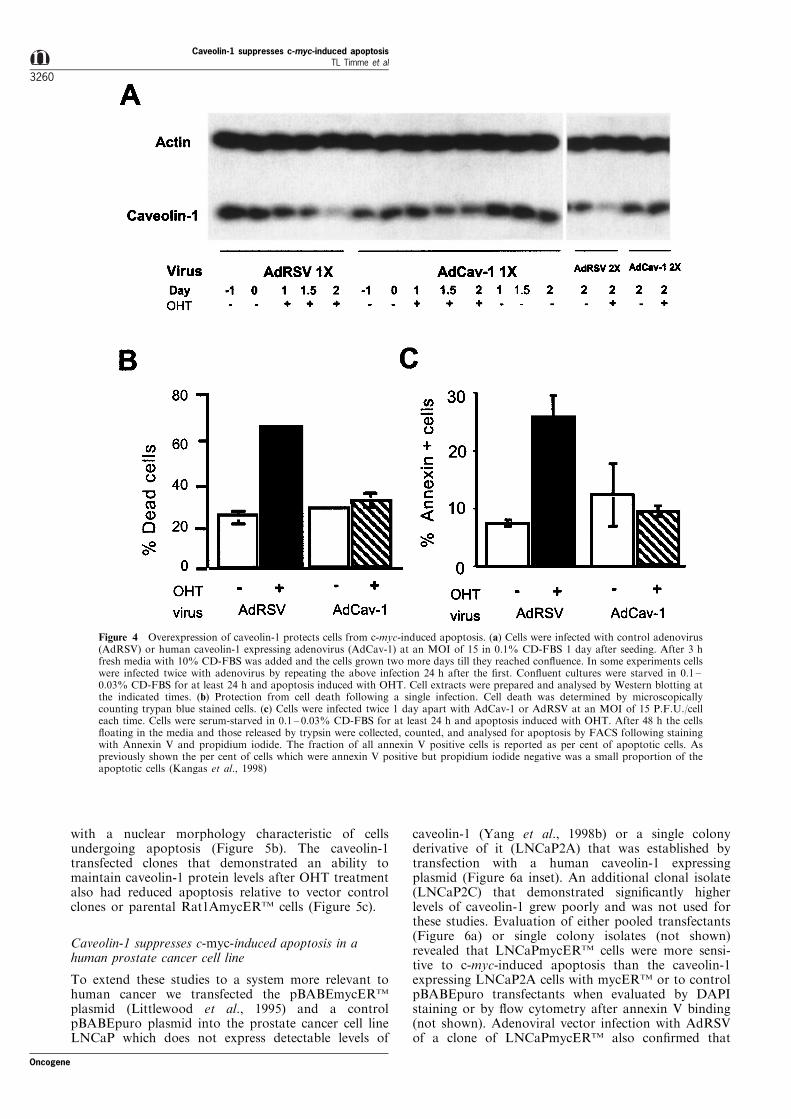
blotting (Figure 4a), showed that caveolin-1 levelscould be increased to a level at least twice that in thecontrol virus infected cells even following 2 days ofOHT treatment after either a single (left panel) ordouble infection (right panel). The level of suppression
Figure 1 Caveolin-1 is down-regulated by c-myc activation. Rat1AmycERTM (top panel) or Rat1A (middle panel) cells wereallowed to grow to near con¯uence, serum-starved, and c-myc activated with OHT. Rat15tet/myc (bottom panel) cells, a clone ofRat1 cells with a tetracycline-regulated c-myc construct (Helbing et al., 1998), were reseeded with or without tetracycline. Caveolin-1levels were consistently down-regulated by c-myc. Interestingly, a transient increase was consistently observed at 32 ± 36 h aftermedia change with or without OHT
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3258
Oncogene
of caveolin-1 protein by c-myc was similar in theAdRSV infected cells to uninfected cells (compare withFigure 1).
Several methods were used to determine whethercaveolin-1 could suppress c-myc-induced apoptosisunder these conditions. The number of cells thatbecame permeable to trypan blue was evaluatedmicroscopically at various times after OHT and amaximal response was observed at 48 h as illustrated in
Figure 4b. The total number of cells increased byalmost twofold in the OHT treated cells (not shown),however overall cell viability decreased over time in theOHT treated AdRSV 1X-infected cultures, whereasAdCav-1 1X-infected cultures did not lose cell viabilityrelative to the level of apoptosis. Minimal growth overthe same time period was observed in control treated(no OHT) cells following infection with either virus.The suppression of apoptosis by caveolin-1 was alsodocumented by ¯ow cytometry after binding ofannexin V to cells infected with adenovirus (2X) andthen treated with OHT for 48 h (Figure 4c). Inconcordance with a previous report (Kangas et al.,1998), we observed a low percentage of cells that hadcharacteristics of early apoptosis (annexin V positiveand propidium iodide negative), even at time pointsearlier than 48 h. The majority of apoptotic cells werepositive for both annexin V and propidium iodide. It isinteresting that the protection from apoptosis bycaveolin-1 was obtained at low (5 ± 25) multiplicitiesof infection (MOI) but not at high MOIs (data notshown). We have observed a similar e�ect on survivalin response to testosterone withdrawal in androgensensitive mouse and human prostate cancer cell lines(Li et al. submitted).
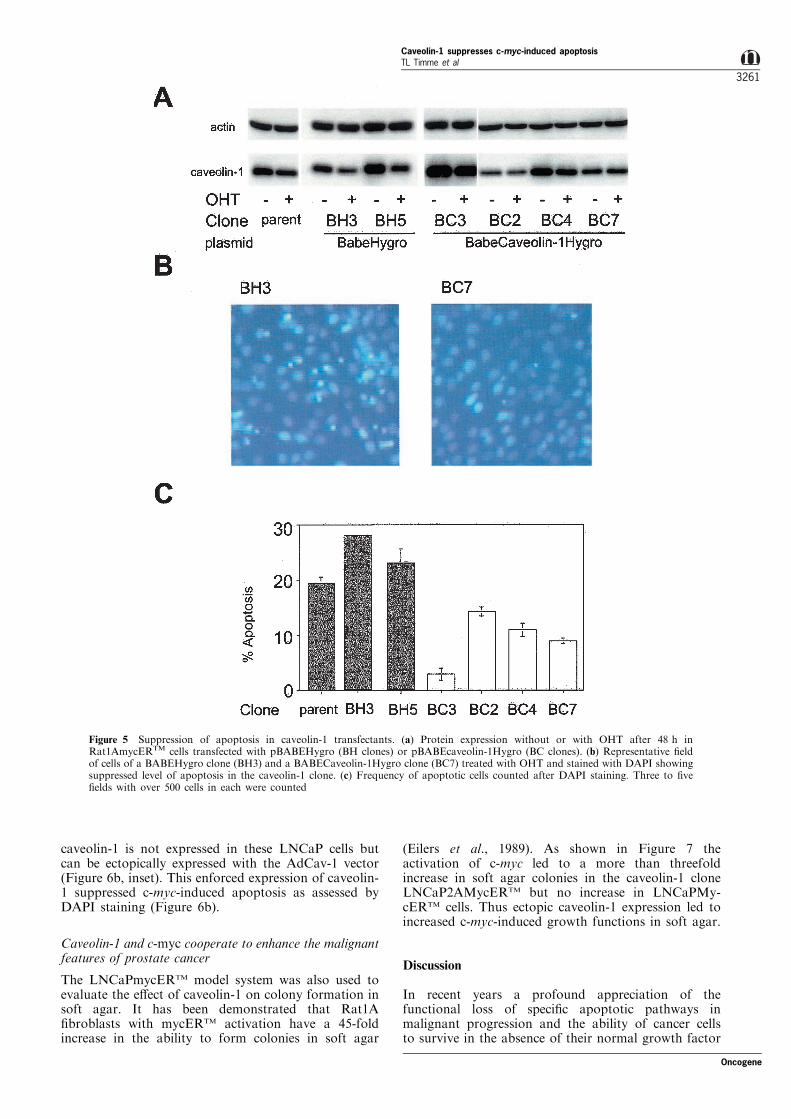
As an independent means of evaluating the e�ect ofcaveolin-1 over expression on apoptosis we transfectedRat1AmycERTM cells with either pBABEHygro orpBABECavHygro and established single colony clones.Clonal variation in the steady state level of caveolin-1protein was noted but the clones expressing ectopiccaveolin-1 (BC2, BC3, BC4 and BC7, Figure 5a)maintained a higher level of caveolin-1 protein afterOHT treatment compared the control clones (BH3 andBH5, Figure 5a). To quantitate apoptosis we stainedcells with DAPI and counted the percentage of cells
Figure 2 RNA was puri®ed and 20 mg separated on a 1% SeaKem agarose gel, photographed, then transferred to nitrocellulose.The membrane was blotted with a mouse caveolin-1 cDNA or a rat GAPDH probe (Yang et al., 1998b). The abundance ofcaveolin-1 mRNA was calculated from the ratio of caveolin-1 to 28S and compared to the abundance in the control cultures(without OHT) at each time point
Figure 3 The luciferase plasmid pGL3-Basic or pGL3-mCav(7721) was transfected into Rat1AmycERTM cells along withpCMV-b-galactosidase. The day after transfection the cells wereserum-starved for 24 h then c-myc activated with OHT.Luciferase levels were standardized to the level of b-galactosidaseactivity. The level of b-galactosidase decreased by less thantwofold in the OHT treated cells as has been previously reported(Marhin et al., 1997) as did luciferase activity from a constructwith an SV40 promoter (not shown), however the level ofrepression from the caveolin-1 promoter was about sixfold
Oncogene
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3259
with a nuclear morphology characteristic of cellsundergoing apoptosis (Figure 5b). The caveolin-1transfected clones that demonstrated an ability tomaintain caveolin-1 protein levels after OHT treatmentalso had reduced apoptosis relative to vector controlclones or parental Rat1AmycERTM cells (Figure 5c).
Caveolin-1 suppresses c-myc-induced apoptosis in ahuman prostate cancer cell line
To extend these studies to a system more relevant tohuman cancer we transfected the pBABEmycERTM
plasmid (Littlewood et al., 1995) and a controlpBABEpuro plasmid into the prostate cancer cell lineLNCaP which does not express detectable levels of
caveolin-1 (Yang et al., 1998b) or a single colonyderivative of it (LNCaP2A) that was established bytransfection with a human caveolin-1 expressingplasmid (Figure 6a inset). An additional clonal isolate(LNCaP2C) that demonstrated signi®cantly higherlevels of caveolin-1 grew poorly and was not used forthese studies. Evaluation of either pooled transfectants(Figure 6a) or single colony isolates (not shown)revealed that LNCaPmycERTM cells were more sensi-tive to c-myc-induced apoptosis than the caveolin-1expressing LNCaP2A cells with mycERTM or to controlpBABEpuro transfectants when evaluated by DAPIstaining or by ¯ow cytometry after annexin V binding(not shown). Adenoviral vector infection with AdRSVof a clone of LNCaPmycERTM also con®rmed that
Figure 4 Overexpression of caveolin-1 protects cells from c-myc-induced apoptosis. (a) Cells were infected with control adenovirus(AdRSV) or human caveolin-1 expressing adenovirus (AdCav-1) at an MOI of 15 in 0.1% CD-FBS 1 day after seeding. After 3 hfresh media with 10% CD-FBS was added and the cells grown two more days till they reached con¯uence. In some experiments cellswere infected twice with adenovirus by repeating the above infection 24 h after the ®rst. Con¯uent cultures were starved in 0.1 ±0.03% CD-FBS for at least 24 h and apoptosis induced with OHT. Cell extracts were prepared and analysed by Western blotting atthe indicated times. (b) Protection from cell death following a single infection. Cell death was determined by microscopicallycounting trypan blue stained cells. (c) Cells were infected twice 1 day apart with AdCav-1 or AdRSV at an MOI of 15 P.F.U./celleach time. Cells were serum-starved in 0.1 ± 0.03% CD-FBS for at least 24 h and apoptosis induced with OHT. After 48 h the cells¯oating in the media and those released by trypsin were collected, counted, and analysed for apoptosis by FACS following stainingwith Annexin V and propidium iodide. The fraction of all annexin V positive cells is reported as per cent of apoptotic cells. Aspreviously shown the per cent of cells which were annexin V positive but propidium iodide negative was a small proportion of theapoptotic cells (Kangas et al., 1998)
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3260
Oncogene
caveolin-1 is not expressed in these LNCaP cells butcan be ectopically expressed with the AdCav-1 vector(Figure 6b, inset). This enforced expression of caveolin-1 suppressed c-myc-induced apoptosis as assessed byDAPI staining (Figure 6b).
Caveolin-1 and c-myc cooperate to enhance the malignantfeatures of prostate cancer
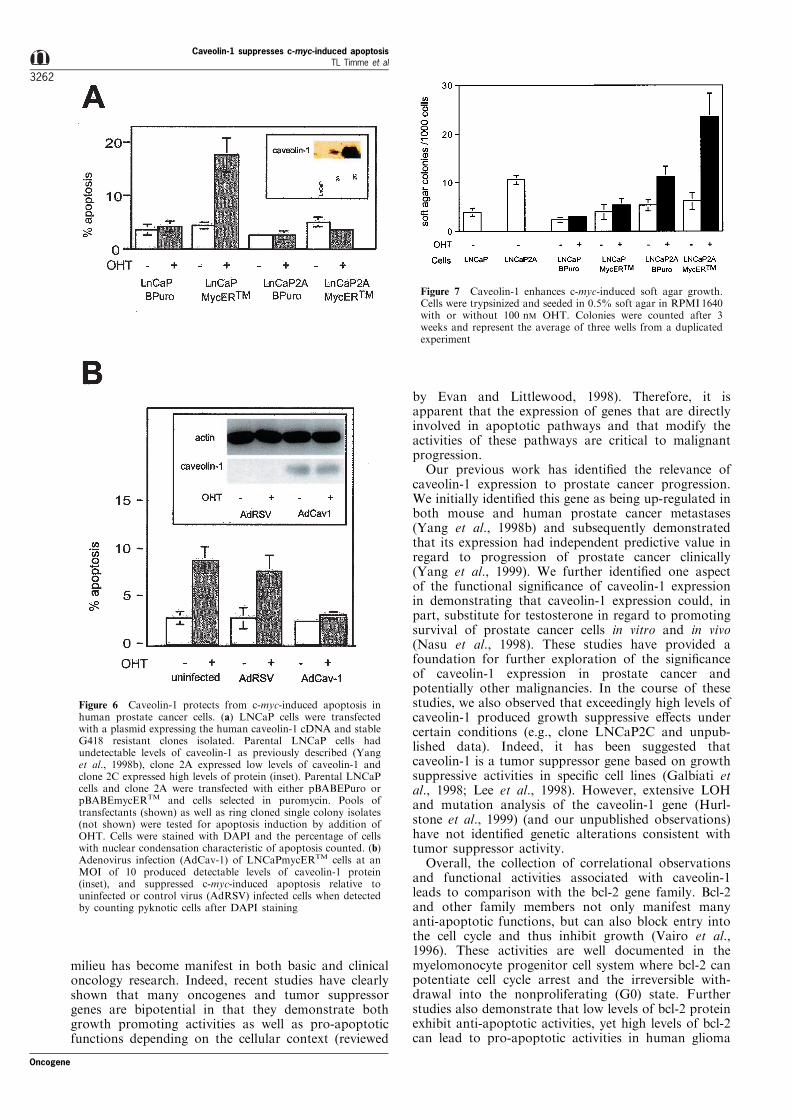
The LNCaPmycERTM model system was also used toevaluate the e�ect of caveolin-1 on colony formation insoft agar. It has been demonstrated that Rat1A®broblasts with mycERTM activation have a 45-foldincrease in the ability to form colonies in soft agar
(Eilers et al., 1989). As shown in Figure 7 theactivation of c-myc led to a more than threefoldincrease in soft agar colonies in the caveolin-1 cloneLNCaP2AMycERTM but no increase in LNCaPMy-cERTM cells. Thus ectopic caveolin-1 expression led toincreased c-myc-induced growth functions in soft agar.
Discussion
In recent years a profound appreciation of thefunctional loss of speci®c apoptotic pathways inmalignant progression and the ability of cancer cellsto survive in the absence of their normal growth factor
Figure 5 Suppression of apoptosis in caveolin-1 transfectants. (a) Protein expression without or with OHT after 48 h inRat1AmycERTM cells transfected with pBABEHygro (BH clones) or pBABEcaveolin-1Hygro (BC clones). (b) Representative ®eldof cells of a BABEHygro clone (BH3) and a BABECaveolin-1Hygro clone (BC7) treated with OHT and stained with DAPI showingsuppressed level of apoptosis in the caveolin-1 clone. (c) Frequency of apoptotic cells counted after DAPI staining. Three to ®ve®elds with over 500 cells in each were counted
Oncogene
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3261
milieu has become manifest in both basic and clinicaloncology research. Indeed, recent studies have clearlyshown that many oncogenes and tumor suppressorgenes are bipotential in that they demonstrate bothgrowth promoting activities as well as pro-apoptoticfunctions depending on the cellular context (reviewed
by Evan and Littlewood, 1998). Therefore, it isapparent that the expression of genes that are directlyinvolved in apoptotic pathways and that modify theactivities of these pathways are critical to malignantprogression.
Our previous work has identi®ed the relevance ofcaveolin-1 expression to prostate cancer progression.We initially identi®ed this gene as being up-regulated inboth mouse and human prostate cancer metastases(Yang et al., 1998b) and subsequently demonstratedthat its expression had independent predictive value inregard to progression of prostate cancer clinically(Yang et al., 1999). We further identi®ed one aspectof the functional signi®cance of caveolin-1 expressionin demonstrating that caveolin-1 expression could, inpart, substitute for testosterone in regard to promotingsurvival of prostate cancer cells in vitro and in vivo(Nasu et al., 1998). These studies have provided afoundation for further exploration of the signi®canceof caveolin-1 expression in prostate cancer andpotentially other malignancies. In the course of thesestudies, we also observed that exceedingly high levels ofcaveolin-1 produced growth suppressive e�ects undercertain conditions (e.g., clone LNCaP2C and unpub-lished data). Indeed, it has been suggested thatcaveolin-1 is a tumor suppressor gene based on growthsuppressive activities in speci®c cell lines (Galbiati etal., 1998; Lee et al., 1998). However, extensive LOHand mutation analysis of the caveolin-1 gene (Hurl-stone et al., 1999) (and our unpublished observations)have not identi®ed genetic alterations consistent withtumor suppressor activity.
Overall, the collection of correlational observationsand functional activities associated with caveolin-1leads to comparison with the bcl-2 gene family. Bcl-2and other family members not only manifest manyanti-apoptotic functions, but can also block entry intothe cell cycle and thus inhibit growth (Vairo et al.,1996). These activities are well documented in themyelomonocyte progenitor cell system where bcl-2 canpotentiate cell cycle arrest and the irreversible with-drawal into the nonproliferating (G0) state. Furtherstudies also demonstrate that low levels of bcl-2 proteinexhibit anti-apoptotic activities, yet high levels of bcl-2can lead to pro-apoptotic activities in human glioma
Figure 6 Caveolin-1 protects from c-myc-induced apoptosis inhuman prostate cancer cells. (a) LNCaP cells were transfectedwith a plasmid expressing the human caveolin-1 cDNA and stableG418 resistant clones isolated. Parental LNCaP cells hadundetectable levels of caveolin-1 as previously described (Yanget al., 1998b), clone 2A expressed low levels of caveolin-1 andclone 2C expressed high levels of protein (inset). Parental LNCaPcells and clone 2A were transfected with either pBABEPuro orpBABEmycERTM and cells selected in puromycin. Pools oftransfectants (shown) as well as ring cloned single colony isolates(not shown) were tested for apoptosis induction by addition ofOHT. Cells were stained with DAPI and the percentage of cellswith nuclear condensation characteristic of apoptosis counted. (b)Adenovirus infection (AdCav-1) of LNCaPmycERTM cells at anMOI of 10 produced detectable levels of caveolin-1 protein(inset), and suppressed c-myc-induced apoptosis relative touninfected or control virus (AdRSV) infected cells when detectedby counting pyknotic cells after DAPI staining
Figure 7 Caveolin-1 enhances c-myc-induced soft agar growth.Cells were trypsinized and seeded in 0.5% soft agar in RPMI1640with or without 100 nM OHT. Colonies were counted after 3weeks and represent the average of three wells from a duplicatedexperiment
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3262
Oncogene

cells (Shinoura et al., 1999). These observations arehighly similar to those that we and others haveobtained through overexpression of caveolin-1 inprostate cancer cells as well as in ®broblast systems.In the present study, we further explored comparisonsto bcl-2 in investigations that focused on interactionsbetween c-myc and caveolin-1.
We have demonstrated in multiple systems thatoverexpression of c-myc can lead to down-regulation ofcaveolin-1 through transcriptional regulation. Thistranscriptional down-regulation appeared to be indir-ect, as protein synthesis was required. Although thespeci®c elements within the caveolin-1 promoter thatmediate this inhibitory response have yet to be de®ned,there are potential sites that include an Inr elementwithin the proximal region of the caveolin-1 promoter.It appears that caveolin-1 gene regulation occurs atmultiple levels. Although there is clearly someregulation at the level of transcription that likelyresults in a reduction of steady state mRNA as we haveshown, there appears to be additional mechanisms ofregulating caveolin-1 mRNA that counteract thesee�ects since the reduction in steady state mRNA issmall compared to the suppression of caveolin-1promoter activities by c-myc observed by transienttransfection analysis. However, the level of reductionof caveolin-1 steady state mRNA (Figure 2, 30%reduction at 4 h,) and the level of c-myc repression ofcaveolin-1 promoter activities by transient transfectionanalysis (Figure 3, sixfold reduction at 24 h) cannot bedirectly compared due to fundamental di�erences inthe analyses. Unfortunately extended analysis ofcaveolin-1 steady state mRNA levels is precluded inthis model because of extensive apoptosis. As caveolin-1 has been shown previously to be regulated by variousserum factors (Galbiati et al., 1998; Lee et al., 1998 andsee Figure 1), it is possible that either the growth ofRat1AmycER cells stimulated by c-myc and/or serumfactors in the medium contribute to overall stability ofcaveolin-1 mRNA in this system. Caveolin-1 may alsobe regulated at the post-translational level by c-myc.
To further explore possible similarities with caveo-lin-1 and bcl-2, we demonstrated that overexpressionof caveolin-1 can block c-myc-induced apoptosis inboth ®broblast and prostate cancer cell line models.These e�ects led us to consider the potential forcooperative activities between caveolin-1 and c-myc inregard to the transformed phenotype. In additionalstudies, we demonstrated that caveolin-1 together withc-myc was capable of cooperating to generateenhanced numbers of colonies in soft agar in vitrorelative to expression of c-myc alone or caveolin-1alone. Overall, our studies establish interactive activ-ities between c-myc and caveolin-1 that are consistentwith cooperation during malignant progression inprostate cancer.
Additional studies will be required to clarify themechanisms that underlie the protective e�ects ofcaveolin-1 within the context of c-myc stimulatedapoptosis. With regard to bcl-2 the mechanism ofprotection from apoptosis induced by c-myc remainsunclear (Prendergast, 1999). It has been proposed thatbcl-2 and speci®c cytokines such as IGF-1 suppress theinitiation c-myc-induced apoptosis rather than a�ectingthe kinetics of apoptosis (McCarthy et al., 1997). Onecould speculate that caveolin-1 may interfere with the
expression or functions of p53 and/or ARF/p19 whichcan be up-regulated in response to c-myc expressionand have been shown to mediate c-myc-inducedapoptosis in ®broblasts (Hermeking and Eick, 1994;Wagner et al., 1994; Zindy et al., 1998). However, thecapacity of these c-myc-regulated e�ectors to ultimatelyinduce apoptosis likely depends on the capacity toactivate the caspase protease cascade. Previous studiesindicate that activation of PKB/Akt may regulate c-myc-induced apoptosis (Kau�mann-Zeh et al., 1997)raising the possibility that caveolin-1 a�ects PKB/Aktor other levels of the survival pathway in which thiskinase functions (Kau�mann-Zeh et al., 1997). A novelPKB/Akt-independent survival pathway for c-myc-induced apoptosis has also been reported for signalingthrough the nerve growth factor receptor TrkA (Ulrichet al., 1998).
Overall our results establish that caveolin-1 is down-regulated at the level of transcriptional repression by c-myc and that this down-regulation appears to beindirect. Further, when caveolin-1 is overexpressed invarious ways c-myc-induced apoptosis is suppressedand phenotypic characteristics consistent with malig-nant progression are enhanced. As caveolin-1 and c-myc are overexpressed in advanced prostate cancer our®ndings have obvious clinical relevance.
Materials and methods
Cells
The Rat1AmycERTM cell line and Rat1A cells weregenerously provided by Robert Eisenman of WashingtonUniversity, Seattle, WA, USA. They were routinely grown inphenol red-free DMEM with 10% charcoal/dextran treatedfetal bovine serum (CD-FBS) (Hyclone). Rat15tet/myc cells,a clone of Rat1 cells with a tetracycline-regulated c-mycconstruct (Helbing et al., 1998), were grown in DMEM with10% FBS in the presence of 2 mg/ml tetracycline to repress c-myc expression. LNCaP cells were obtained from theAmerican Type Culture Collection, grown in RPMI 1640with 10% FBS, and used at low passage (less than 60).LNCaP cells transfected with pBABEmycERTM or pBabe-Puro were grown in RPMI 1640 with 10% CD-FBS with1 mg/ml puromycin and 261078 M testosterone.
c-myc activation
To activate c-myc, Rat1AmycERTM cells were allowed togrow to near con¯uence then the serum concentrationreduced to 0.1 ± 0.03% for 24 ± 48 h. 4-hydroxytamoxifen(OHT) (Calbiochem) was dissolved in DMSO and added tomedia to give a ®nal concentration of 100 ± 250 nM. Controlplates received an equal amount of DMSO. Rat15tet/myccells were reseeded at 26104 cells/cm2 with or withouttetracycline. After 8 h the cells were washed twice withphosphate bu�ered saline and the media replaced withDMEM with 0.1% FBS with or without tetracycline. TheLNCaPmycERTM cells were seeded at 104 cells/cm2 in RPMI1640 with 10% FBS they were switched to RPMI 1640supplemented with 261078 M testosterone without serum for24 and then OHT added at 100 nM.
Plasmids and adenovirus
A fragment containing 721 nucleotides immediately upstreamof the ATG codon of the mouse caveolin-1 gene wassubcloned from a PAC clone. This mouse caveolin-1
Oncogene
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3263

promoter was inserted into the promoterless luciferaseplasmid pGL3-Basic (Promega) to create pGL3-mCav(7721). Luciferase plasmids (2.0 mg) were transfected intoRat1AmycERTM cells using lipofectamine (Life Technologies)along with 0.5 mg of a plasmid with b-galactosidase under thecontrol of the CMV promoter, pCMV-b-galactosidase. Theday after transfection the cells were serum-starved for 24 hthen c-myc activated with OHT. Cell lysates were preparedwith reporter lysis bu�er (Promega) and luciferase assayedwith LucLite (Packard Instruments) and standardized to thelevel of b-galactosidase activity. A mouse caveolin-1 cDNAwas inserted into pBABEHygro, transfected into Rat1Amy-cERTM cells using lipofectamine and stable clones selectedwith 300 mg/ml hygromycin. A human caveolin-1 cDNA wasinserted into pcDNA3.1 and transfected into LNCaP cellswith lipofectamine. Stable clones were selected with 400 mg/ml G418. pBABEPuro and pBABEmycERTM, also providedby Robert Eisenman, were transfected into LNCaP cells andstable colonies selected with 1 mg/ml puromycin.
An adenoviral vector with the human caveolin-1 cDNA(AdCav-1) was constructed as previously described (Nasu etal., 1998). The control adenoviral vector without a cDNAinsert (AdRSV) or AdCav-1 was used to infect Rat1Amy-cERTM cells 1 day after seeding at 36104 cells/cm2 by addingvirus at the indicated MOI to media without serum andleaving it on cells for 3 h. Additional media with serum wasthen added and the cells incubated for an additional day. Insome experiments cells were infected twice with adenovirus byrepeating the above infection 24 h after the ®rst. Forapoptosis the cells were allowed to reach con¯uence thenstarved for 1 day and then treated with OHT for 2 days.Infection of LNCaPmycERTM was essentially as describedabove except at an MOI of 10 was used.
Western and Northern blots
Cells were scraped from culture plates and collected bycentrifugation. The cell pellets were washed once with PBSand then lysed with TNES lysis bu�er (50 mM Tris (pH 7.5),2 mM EDTA, 100 mM NaCl, 1% NP40, 20 mg/ml aprotinin,20 mg/ml leupeptin and 1 mM PMSF) on ice for 45 min.Proteins were separated on a 15% polyacrylamide-SDS geland then electrophoretically transferred onto a nitrocellulosemembrane. Caveolin-1 was detected with polyclonal caveolin-1 antiserum (SC-894, Santa Cruz Biotech). A monoclonal b-actin antibody (A5441, Sigma) was used to detect b-actin for
loading control. Blots were visualized with Pierce Supersignaland the image captured with a Nucleovision camera. RNAwas isolated from con¯uent Rat1AmycERTM at the indicatedtimes. At time 724 the media was changed to 0.1% CD-FBS. At time 0 the media was changed again to 0.1% CD-FBS with or without OHT. RNA was isolated withUltraSpec (Biotecx Laboratories Inc) and 20 mg separatedon a 1% SeaKem agarose gel, photographed, then transferredto nitrocellulose. The membrane was blotted with a mousecaveolin-1 cDNA or a rat GAPDH probe (Yang et al.,1998b).
Apoptosis detection
The per cent of dead cells was determined by counting¯oating plus cells detached by trypsinization which wereunable to exclude trypan blue. Detection of annexin V andpropidium iodide stained cells was after 48 h of OHTtreatment. The cells ¯oating in the media and those releasedby trypsin were collected and counted. An aliquot of 56105
was analysed for apoptosis by FACS following staining withAnnexin V and propidium iodide (Oncogene Sciences). Directcounting of apoptotic cells was done following staining with10 mg/ml bisbenzimide (DAPI). Cells with pyknotic orfragmented nuclei were counted as apoptotic.
Soft agar growth
Cells were trypsinized, counted and diluted to yield 1000 ±2500 cells per 6 well. The cells were mixed with moltenagarose in RPMI 1640 with 10% CD-FBS to give a ®nalagarose concentration of 0.5%. They were immediatelylayered on a base of 2% agar containing medium. The softagar was allowed to solidify and then overlayed with anadditional of 0.5% agarose overlay. OHT was included in thesoft agar and overlaying media at 100 nM. After solidi®ca-tion, additional media was added and the plates incubatedfor 3 weeks.
AcknowledgmentsThis work was supported by grants from the NationalInstitutes of Health (SPORE P50-CA58204 and RO-1,CA68814 and CA50588) and the Department of Defense(DAMD 17-98-1-8575). We thank Drs Craig Chinault,Joann Trial and Yanfuei Yuan for technical assistance.
References
Adams JM and Cory S. (1998). Science, 281, 1322 ± 1326.Bist A, Fielding PE and Fielding CJ. (1997). Proc. Natl.
Acad. Sci. USA, 94, 10693 ± 10698.Bubendorf L, Kononen J, Koivisto P, Schraml P, Moch H,
Gasser TC, Willi N, Mihatsch MJ, Sauter G andKallioniemi OP. (1999). Cancer Res., 59, 803 ± 806.
Claassen GF and Hann SR. (1999). Oncogene, 18, 2925 ±2933.
Dang CV. (1999). Mol. Cell. Biol., 19, 1 ± 11.Eilers M, Picard D, Yamamoto KR and Bishop JM. (1989).
Nature, 340, 66 ± 68.Eilers M, Schirm S and Bishop JM. (1991). EMBO J., 10,
133 ± 141.Engelman JA, Wyko� CC, Yasuhara S, Song KS, Okamoto
T and Lisanti MP. (1997). J. Biol. Chem., 272, 16374 ±16381.
Evan GI, Wyllie Ah, Gilbert GS, Littlewood TD, Land H,Brooks M, Waters CM, Penn LZ and Hancock DC.(1992). Cell, 69, 119 ± 128.
Evan G and Littlewood T. (1998). Science, 281, 1317 ± 1322.Facchini LM and Penn LZ. (1998). FASEB J., 12, 633 ± 651.
Galbiati F, Volonte D, Engelman JA, Watanabe G, Burk R,Pestell RG and Lisanti MP. (1998). EMBO J., 17, 6633 ±6648.
Grandori C and Eisenman RN. (1997). Trends Biochem. Sci.,22, 177 ± 181.
Harrington EA, Bennett MR, Fanidi A and Evan GI. (1994).EMBO J., 13, 3286 ± 3295.
Helbing CC, Wellington CL, Gogela-Spehar M, Cheng T,Pinchbeck GG and Johnston RN. (1998). Oncogene, 17,1491 ± 1501.
Hermeking H and Eick D. (1994). Science, 265, 2091 ± 2093.Hurlstone AF, Reid G, Reeves JR, Fraser J, Strathdee G,
Rahilly M, Parkinson EK and Black DM. (1999).Oncogene, 18, 1881 ± 1890.
Jenkins RB, Qian J, Lieber MM and Bostwick DG. (1997).Cancer Res., 57, 524 ± 531.
Kangas A, Nicholson DW and Hottla E. (1998). Oncogene,16, 387 ± 398.
Kau�mann-Zeh A, Rodriguez-Viciana P, Ulrich E, GilbertC, Co�er P, Downward J and Evan G. (1997).Nature, 385,544 ± 548.
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3264
Oncogene

Koleske AJ, Baltimore D and Lisanti MP. (1995). Proc. Natl.Acad. Sci. USA, 92, 1381 ± 1385.
Lavie Y, Fiucci G and Liscovitch M. (1998). J. Biol. Chem.,273, 32380 ± 32383.
Lee SW, Reimer CL, Oh P, Campbell DB and Schnitzer JE.(1998). Oncogene, 16, 1391 ± 1397.
Li LH, Nerlov C, Prendergast G, MacGregor D, and Zi� EB.(1994). EMBO J., 13, 4070 ± 4079.
Littlewood TD, Hancock DC, Danielian PS, Parker MG andEvan GI. (1995). Nucleic Acids Res., 23, 1686 ± 1690.
Marcu KB, Bossone SA and Patel AJ. (1992). Annu. Rev.Biochem., 61, 809 ± 860.
Marhin WW, Chen S, Facchini LM, Fornace Jr AJ and PennLZ. (1997). Oncogene, 14, 2825 ± 2834.
McCarthy NJ, Whyte MK, Gilbert CS and Evan GI. (1997).J. Cell Biol., 136, 215 ± 227.
Nasu Y, Timme TL, Yang G, Bangma CH, Li L, Ren C, ParkSH, DeLeon M, Wang J and Thompson TC. (1998). Nat.Med., 4, 1062 ± 1064.
Nupponen NN, Kakkola L, Koivisto P and Visakorpi T.(1998). Am. J. Pathol., 153, 141 ± 148.
Parton RG. (1996). Curr. Opin. Cell. Biol., 8, 542 ± 548.Prendergast GC. (1999). Oncogene, 18, 2967 ± 2987.Qian J, Jenkins RB and Bostwick DG. (1997). Mod. Pathol.,
10, 1113 ± 1119.Roy AL, Carruthers C, Gutjahr T and Roeder RG. (1993).
Nature, 365, 359 ± 361.Sato K, Qian J, Slezak JM, Lieber MM, Bostwick DG,
Bergstralh EJ and Jenkins RB. (1999). J. Natl. CancerInst., 91, 1574 ± 1580.
Sears R, Leone G, DeGregori J and Nevins JR. (1999). Mol.Cell, 3, 169 ± 179.
Shaul PW and Anderson RG. (1998). Am. J. Physiol., 275,L843 ± 851.
Shinoura N, Yoshida Y, Nishimura M, Muramatsu Y, AsaiA, Kirino T and Hamada H. (1999). Cancer Res., 59,4119 ± 4128.
Srivastava RK, Sollott SJ, Khan L, Hansford R, Lakatta EGand Longo DL. (1999). Mol. Cell. Biol., 19, 5659 ± 5674.
Thompson TC, Southgate J, Kitchener G and Land H.(1989). Cell, 56, 917 ± 930.
Thompson TC, Timme T, Li L, Goltsov A and Yang G.(1999a). Emerging Therapeutic Targets, 3, 337 ± 346.
Thompson TC, Timme TL, Li L and Goltsov A. (1999b).Apoptosis, 4, 233 ± 237.
Ulrich E, Duwel A, Kau�mann-Zeh A, Gilbert C, Lyon D,Rudkin B, Evan G and Martin-Zanca D. (1998).Oncogene, 16, 825 ± 832.
Vairo G, Innes KM and Adams JM. (1996). Oncogene, 13,1511 ± 1519.
Wagner AJ, Kokontis JM and Hay N. (1994). Genes Dev., 8,2817 ± 2830.
Wu KJ, Polack A and Dalla-Favera R. (1999). Science, 283,676 ± 679.
Yang CP, Galbiati F, Volonte D, Horwitz SB and LisantiMP. (1998a). FEBS Lett., 439, 368 ± 372.
Yang G, Truong LD, Timme TL, Ren C, Wheeler TM, ParkSH, Nasu Y, Bangma CH, Kattan MW, Scardino PT andThompson TC. (1998b). Clin. Cancer Res., 4, 1873 ± 1880.
Yang G, Truong LD, Wheeler TM and Thompson TC.(1999). Cancer Res., 59, 5719 ± 5723.
Zindy F, Eischen CM, Randle DH, Kamijo T, Cleveland JL,Sherr CJ and Roussel MF. (1998). Genes Dev., 12, 2424 ±2433.
Oncogene
Caveolin-1 suppresses c-myc-induced apoptosisTL Timme et al
3265





























