Embed Size (px)
Citation preview

Molecular Cell, Vol. 11, 1189–1200, May, 2003, Copyright 2003 by Cell Press
The F-Box Protein Skp2 Participatesin c-Myc Proteosomal Degradation and Actsas a Cofactor for c-Myc-Regulated Transcription
tion by c-Myc: the N-terminal transactivation domain(TAD) and the C-terminal basic-helix-loop-helix-leucinezipper (bHLH-Zip). The bHLH-Zip domain mediates di-merization with the cofactor Max and DNA binding toE-box elements at target gene promoters. Myc-box 2
Natalie von der Lehr,1 Sara Johansson,1
Siqin Wu,1 Fuad Bahram,1 Alina Castell,1
Cihan Cetinkaya,1 Per Hydbring,1
Ingrid Weidung,1 Keiko Nakayama,2
Keiichi I. Nakayama,2 Ola Soderberg,3
(MB2) within the TAD interacts with the coactivatorTom K. Kerppola,4 and Lars-Gunnar Larsson1,*TRRAP, a component of histone acetyl transferase (HAT)1Department of Plant Biology and Forest Geneticscomplexes, which participates in the regulation of chro-Swedish University of Agricultural Sciencesmatin structure (McMahon et al., 1998, 2000; Bouchard750 07 Uppsalaet al., 2001; Frank et al., 2001; for review see Amati etSwedenal., 2001). A growing number of target genes regulated2 Department of Molecular Geneticsby c-Myc have been identified, including genes involvedMedical Institute of Bioregulationin the cell cycle, cell growth, apoptosis, metabolism,Kyushu Universitydifferentiation, and immortalization (for review seeFukuoka 812-8582Grandori et al., 2000).Japan
The ubiquitin/proteasome pathway involves ubiquitin3 Department of Genetics and Pathologymodification of substrates and degradation of the sub-The Rudbeck Laboratorystrates by the proteasome. This pathway controls theUniversity of Uppsalaturnover rate of many regulatory proteins (for review see751 85 UppsalaHershko and Ciechanover, 1998), including c-Myc (FlinnSwedenet al., 1998; Salghetti et al., 1999; Sears et al., 1999;4 Howard Hughes Medical InstituteBahram et al., 2000; Gregory and Hann, 2000). UbiquitinDepartment of Biological Chemistryconjugation is catalyzed by enzymes designated E1, E2,University of Michigan School of Medicineand E3. The E3 ubiquitin ligases determine the substrateAnn Arbor, Michigan 48109specificity. E3 complexes of the SCF subfamily typicallycontain at least four subunits: Skp1, a Cullin, a Ring-finger protein, and a member of a large family of F-boxSummaryadaptor proteins involved in specific substrate recogni-tion (for review see Weissman, 2001). One such SCFThe transcription regulatory oncoprotein c-Myc con-complex is SCFSkp2 (Lisztwan et al., 1998), which containstrols genes involved in cell growth, apoptosis, andthe F-box protein Skp2, involved in the turnover of manyoncogenesis. c-Myc is turned over very quicklycell cycle regulatory substrates, including p27Kip1 (Car-through the ubiquitin/proteasome pathway. The pro-rano et al., 1999; Sutterluty et al., 1999; for review seeteins involved in this process are still unknown. WeDeSalle and Pagano, 2001). Skp2 is overexpressed inhave found that Skp2 interacts with c-Myc and partici-many tumors and has transforming capacity (Gstaigerpates in its ubiquitylation and degradation. The inter-et al., 2001; Latres et al., 2001), and is therefore classifiedaction between Skp2 and c-Myc occurs during theas an oncogene.G1 to S phase transition of the cell cycle in normal
Recent studies have suggested a close link betweenlymphocytes. Surprisingly, Skp2 enhances c-Myc-the activator function of many transcription factors andinduced S phase transition and activates c-Myc targettheir ubiquitylation/degradation (for reviews see Cona-genes in a Myc-dependent manner. Further, Myc-way et al., 2002; Ottosen et al., 2002; Muratani and
induced transcription was shown to be Skp2 depen-Tansey, 2003). The potencies of activation domains of-
dent, suggesting interdependence between c-Myc and ten correlate with their stabilities. Further, sequencesSkp2 in activation of transcription. Moreover, Myc- targeted by the degradation machinery (“degrons”) anddependent association of Skp2, ubiquitylated proteins, those required for transcriptional activation often over-and subunits of the proteasome to a c-Myc target lap and can in fact be functionally equivalent (Molinaripromoter was demonstrated in vivo. The results sug- et al., 1999; Salghetti et al., 2000). Both proteolytic andgest that Skp2 is a transcriptional cofactor for c-Myc nonproteolytic roles of ubiquitylation mediated by theand indicates a close relationship between transcrip- SCFMet30 E3 ligase for the transcriptional function of Met4tion activation and transcription factor ubiquitination. or the VP16 TAD in yeast have been suggested (Kaiser
et al., 2000; Salghetti et al., 2001; Kuras et al., 2002).Introduction Moreover, many proteins involved in ubiquitylation of
transcriptional regulators are integral parts of preinitia-The MYC protooncogene family encodes short-lived tion complexes and/or the RNA polII holoenzyme (Chitranscription factors that play important roles in cell et al., 2001; Brower et al., 2002). Finally, a nonproteolyticproliferation, apoptosis, and cancer development (for role of the AAA proteins independent of 20S (APIS)review see Grandori et al., 2000). Two regions are required ATPases of the 19S regulatory proteasome particle infor transcription regulation and oncogenic transforma- transcription has been suggested (Ferdous et al., 2001;
Gonzalez et al., 2002; for reviews see Conaway et al.,2002; Ottosen et al., 2002; Muratani and Tansey, 2003).*Correspondence: [email protected]

Molecular Cell1190
Although c-Myc is a well-known transcription factor c-Myc Interacts with Skp2 through the MB2and the HLH-Zip Domainsthat regulates many aspects of cell function, little is
known about the mechanisms that control its ubiquitin/ To identify the regions of c-Myc responsible for thec-Myc/Skp2 interaction, a series of c-Myc deletion deri-proteasome-mediated turnover and the impact that this
may have on its functions as a transcription factor. We vates (Figure 2A) were expressed in cells. Deletion ofMB1 did not affect Myc-Skp2 complex formation in co-here provide evidence that the SCFSkp2 E3 ligase pro-
motes ubiquitylation and proteasome-mediated degra- transfected cells, whereas deletion of MB2 reduced butdid not abolish the interaction (Figure 2B, left upperdation of c-Myc. Our results further suggest that SCFSkp2
may function as a cofactor for Myc-induced transcrip- panel). Deletion of the N-terminal 205 aa did not affectbinding to Skp2 as much as deletion of the C-terminaltion and that Skp2 is recruited to c-Myc target promoters
in a Myc-dependent manner. 224 residues (Figure 2B, middle panel). Deletion of boththe 224 C-terminal aa and MB2 abolished binding toSkp2. Further mapping showed that the first 148 aa of
Results c-Myc (mutant �C148–439) still bound, whereas mutant�95–435 did not (right panel of Figure 2B). The shortest
c-Myc Binds to the F-Box Protein Skp2 C-terminal fragment of c-Myc (mutant �2–366) was stillIn Vivo and In Vitro able to bind to Skp2, suggesting that aa residues 367–To investigate the possible involvement of an SCF E3 439, comprising the HLH-Zip domain of c-Myc, are suffi-ligase complex in c-Myc ubiquitylation and degradation, cient for Skp2 binding.we examined whether c-Myc could interact with Cul1- In vitro binding studies showed that deletion of MB2containing SCF complexes. Figure 1A (upper panel) alone or in combination with MB1 reduced binding ofshows that Cul1 coimmunoprecipitated with c-Myc after IVT c-Myc to GST-Skp2, whereas deletion of MB1 alonecotransfection into Cos7 cells. This suggested that had no effect (Figure 2C, left panel). Deletion of thec-Myc and Cul1 were present in the same complex, C-terminal aa 215–439 (mutant �C) reduced but did notpossibly through interactions with an endogenous F-box eliminate binding, whereas the combined �C��MB2protein(s). deletion abolished the GST-Skp2 interaction. These re-
To identify potential F-box proteins linking c-Myc to sults are thus in agreement with the data obtained usingSCF-complexes, we tested binding of c-Myc to known transfected cells (Figure 2B). Taken together, these re-Cul1-interacting F-box proteins. We found that c-Myc sults suggest that the interaction between c-Myc andcoimmunoprecipitated with the F-box protein Skp2 from Skp2 requires an intact MB2 and a region in the HLH-cotransfected cells, whereas only negligible binding to Zip domain of c-Myc. These regions of Myc are alsothe F-box protein �-TRCP and the transcription factor crucial for both transcription regulation and cell transfor-Stat-1 was observed (Figure 1B). A low level of binding mation.was observed to the F-box proteins NFB42 and Fbl5.Whereas the binding to NFB42 seems to be due to over-expression (data not shown), the interaction with the c-Myc and Skp2 Interact in the NucleusSkp2 family member Fbl5 was detected reproducibly. To visualize the intracellular location of the c-Myc/Skp2Since the Skp2/c-Myc interaction was stronger and interaction in living cells we utilized bimolecular fluores-since the effect of Skp2 on Myc degradation was much cence complementation (BiFC) (Hu et al., 2002). Thismore potent than that of Fbl5 and two other Skp2 family technique is based on reconstitution of yellow fluores-F-box proteins (see below Figure 3A), we concentrated cent protein (YFP) from nonfluorescent N-terminal (YN)our work on Skp2. and C-terminal (YC) YFP fragments when they are
The c-Myc/Skp2 interaction was substantiated by re- brought together by two interacting proteins fused tociprocal coimmunoprecipitation, demonstrating that the fragments. Figure 2D (left section, second left-handc-Myc coimmunoprecipitated with Skp2 antiserum and panel from the top) shows fluorescence in cells coex-vice versa (Figure 1C, first and third panels from the top, pressing MycYC and Skp2YN fusion constructs, similarrespectively) from cells cotransfected with c-Myc and to cells expressing Myc fused to EGFP (Myc-GFP) (leftSkp2. Further, endogenous c-Myc coimmunoprecipi- section, top left-hand panel). The fluorescence was lo-tated with Skp2 in untransfected HeLa cells (Figure 1D). calized almost exclusively to the cell nucleus as con-No precipitation of c-Myc was observed using unrelated firmed by DAPI staining (Figure 2D, left section, topcontrol antibodies. In the reciprocal experiment, a faint right-hand panel). No or very weak fluorescence wasband corresponding to the size of Skp2 was coimmuno- observed in cells expressing Skp2YN and Myc�bHLH-precipitated with two different c-Myc antibodies but not ZipYC (with reduced binding to Skp2, see Figure 2B),with control antibodies. thus demonstrating the specificity of the MycYC-
c-Myc and Skp2 also bound each other in vitro as Skp2YN interaction. Further, no or very weak fluores-shown by GST pull-downs (Figure 1E). Both the N-ter- cence was observed in control cells expressingminal 262 and the C-terminal 176 amino acid residues MycYC � I�B�YN, MycYC � p50YN, or Skp2YN �of c-Myc fused to GST interacted with IVT Skp2 but p65YC. When p50YN � p65YC and p65YC � I�B�YNnot with IVT p27Kip1 used as a control. In the reverse were cotransfected, nuclear and cytoplasmic fluores-
cence, respectively, were observed as previously re-experiment, bacterially expressed GST-Skp2 interactedwith IVT full-length c-Myc. No binding of either of the ported (Hu et al., 2002). These results suggested that
the c-Myc/Skp2 interaction takes place in the nucleus.IVT proteins to GST alone was observed.
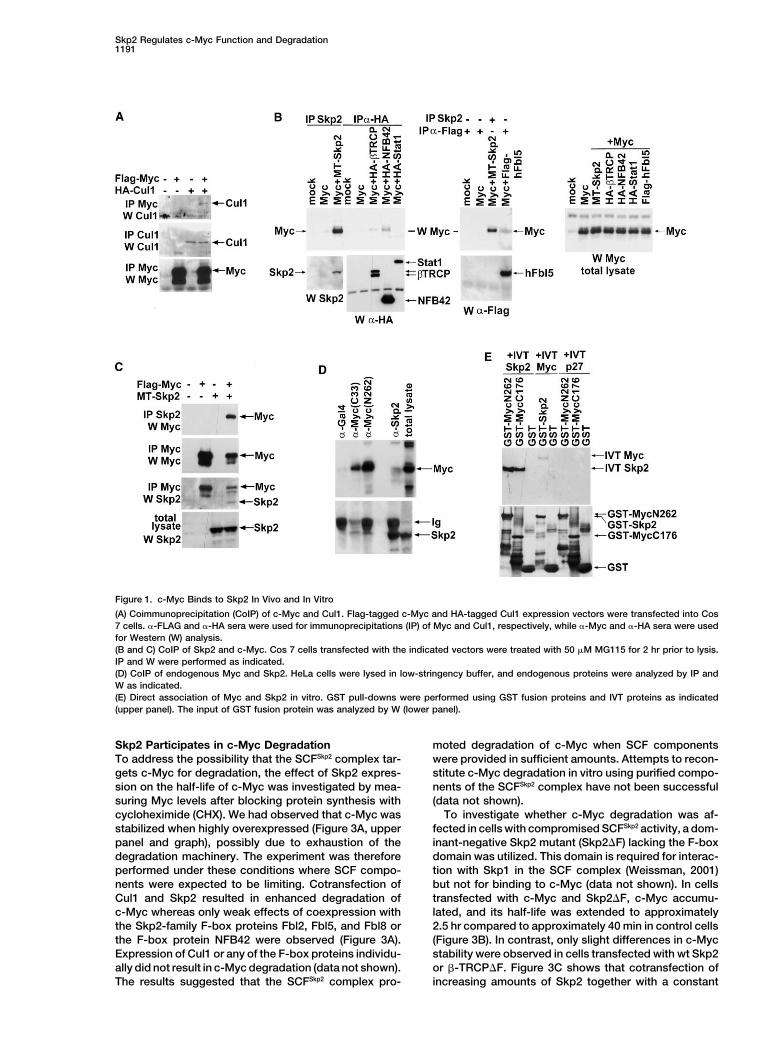
Skp2 Regulates c-Myc Function and Degradation1191
Figure 1. c-Myc Binds to Skp2 In Vivo and In Vitro
(A) Coimmunoprecipitation (CoIP) of c-Myc and Cul1. Flag-tagged c-Myc and HA-tagged Cul1 expression vectors were transfected into Cos7 cells. �-FLAG and �-HA sera were used for immunoprecipitations (IP) of Myc and Cul1, respectively, while �-Myc and �-HA sera were usedfor Western (W) analysis.(B and C) CoIP of Skp2 and c-Myc. Cos 7 cells transfected with the indicated vectors were treated with 50 �M MG115 for 2 hr prior to lysis.IP and W were performed as indicated.(D) CoIP of endogenous Myc and Skp2. HeLa cells were lysed in low-stringency buffer, and endogenous proteins were analyzed by IP andW as indicated.(E) Direct association of Myc and Skp2 in vitro. GST pull-downs were performed using GST fusion proteins and IVT proteins as indicated(upper panel). The input of GST fusion protein was analyzed by W (lower panel).
Skp2 Participates in c-Myc Degradation moted degradation of c-Myc when SCF componentswere provided in sufficient amounts. Attempts to recon-To address the possibility that the SCFSkp2 complex tar-
gets c-Myc for degradation, the effect of Skp2 expres- stitute c-Myc degradation in vitro using purified compo-nents of the SCFSkp2 complex have not been successfulsion on the half-life of c-Myc was investigated by mea-
suring Myc levels after blocking protein synthesis with (data not shown).To investigate whether c-Myc degradation was af-cycloheximide (CHX). We had observed that c-Myc was
stabilized when highly overexpressed (Figure 3A, upper fected in cells with compromised SCFSkp2 activity, a dom-inant-negative Skp2 mutant (Skp2�F) lacking the F-boxpanel and graph), possibly due to exhaustion of the
degradation machinery. The experiment was therefore domain was utilized. This domain is required for interac-tion with Skp1 in the SCF complex (Weissman, 2001)performed under these conditions where SCF compo-
nents were expected to be limiting. Cotransfection of but not for binding to c-Myc (data not shown). In cellstransfected with c-Myc and Skp2�F, c-Myc accumu-Cul1 and Skp2 resulted in enhanced degradation of
c-Myc whereas only weak effects of coexpression with lated, and its half-life was extended to approximately2.5 hr compared to approximately 40 min in control cellsthe Skp2-family F-box proteins Fbl2, Fbl5, and Fbl8 or
the F-box protein NFB42 were observed (Figure 3A). (Figure 3B). In contrast, only slight differences in c-Mycstability were observed in cells transfected with wt Skp2Expression of Cul1 or any of the F-box proteins individu-
ally did not result in c-Myc degradation (data not shown). or �-TRCP�F. Figure 3C shows that cotransfection ofincreasing amounts of Skp2 together with a constantThe results suggested that the SCFSkp2 complex pro-
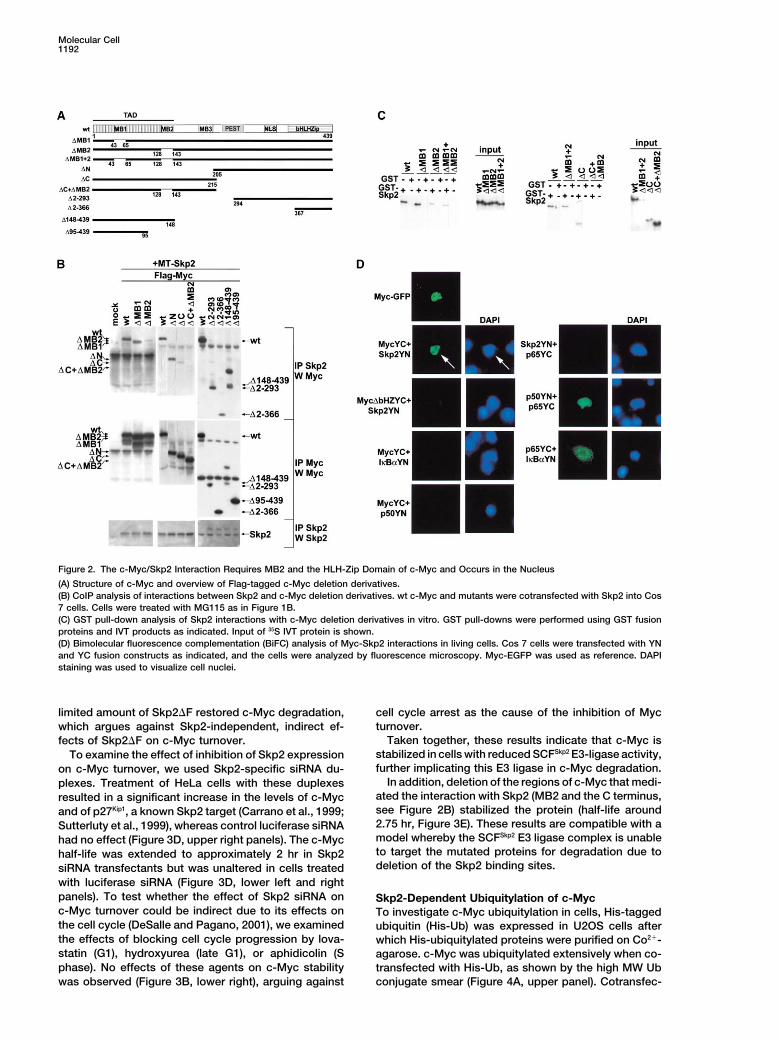
Molecular Cell1192
Figure 2. The c-Myc/Skp2 Interaction Requires MB2 and the HLH-Zip Domain of c-Myc and Occurs in the Nucleus
(A) Structure of c-Myc and overview of Flag-tagged c-Myc deletion derivatives.(B) CoIP analysis of interactions between Skp2 and c-Myc deletion derivatives. wt c-Myc and mutants were cotransfected with Skp2 into Cos7 cells. Cells were treated with MG115 as in Figure 1B.(C) GST pull-down analysis of Skp2 interactions with c-Myc deletion derivatives in vitro. GST pull-downs were performed using GST fusionproteins and IVT products as indicated. Input of 35S IVT protein is shown.(D) Bimolecular fluorescence complementation (BiFC) analysis of Myc-Skp2 interactions in living cells. Cos 7 cells were transfected with YNand YC fusion constructs as indicated, and the cells were analyzed by fluorescence microscopy. Myc-EGFP was used as reference. DAPIstaining was used to visualize cell nuclei.
limited amount of Skp2�F restored c-Myc degradation, cell cycle arrest as the cause of the inhibition of Mycturnover.which argues against Skp2-independent, indirect ef-
Taken together, these results indicate that c-Myc isfects of Skp2�F on c-Myc turnover.stabilized in cells with reduced SCFSkp2 E3-ligase activity,To examine the effect of inhibition of Skp2 expressionfurther implicating this E3 ligase in c-Myc degradation.on c-Myc turnover, we used Skp2-specific siRNA du-
In addition, deletion of the regions of c-Myc that medi-plexes. Treatment of HeLa cells with these duplexesated the interaction with Skp2 (MB2 and the C terminus,resulted in a significant increase in the levels of c-Mycsee Figure 2B) stabilized the protein (half-life aroundand of p27Kip1, a known Skp2 target (Carrano et al., 1999;2.75 hr, Figure 3E). These results are compatible with aSutterluty et al., 1999), whereas control luciferase siRNAmodel whereby the SCFSkp2 E3 ligase complex is unablehad no effect (Figure 3D, upper right panels). The c-Mycto target the mutated proteins for degradation due tohalf-life was extended to approximately 2 hr in Skp2deletion of the Skp2 binding sites.siRNA transfectants but was unaltered in cells treated
with luciferase siRNA (Figure 3D, lower left and rightpanels). To test whether the effect of Skp2 siRNA on Skp2-Dependent Ubiquitylation of c-Mycc-Myc turnover could be indirect due to its effects on To investigate c-Myc ubiquitylation in cells, His-taggedthe cell cycle (DeSalle and Pagano, 2001), we examined ubiquitin (His-Ub) was expressed in U2OS cells afterthe effects of blocking cell cycle progression by lova- which His-ubiquitylated proteins were purified on Co2�-statin (G1), hydroxyurea (late G1), or aphidicolin (S agarose. c-Myc was ubiquitylated extensively when co-phase). No effects of these agents on c-Myc stability transfected with His-Ub, as shown by the high MW Ub
conjugate smear (Figure 4A, upper panel). Cotransfec-was observed (Figure 3B, lower right), arguing against
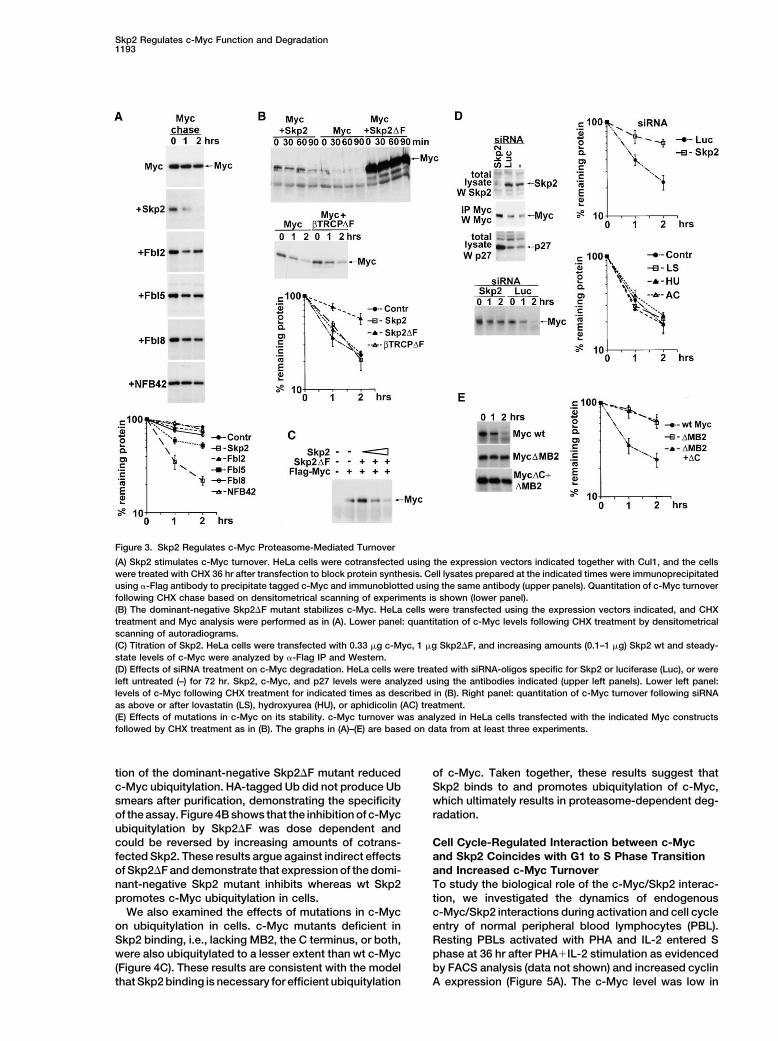
Skp2 Regulates c-Myc Function and Degradation1193
Figure 3. Skp2 Regulates c-Myc Proteasome-Mediated Turnover
(A) Skp2 stimulates c-Myc turnover. HeLa cells were cotransfected using the expression vectors indicated together with Cul1, and the cellswere treated with CHX 36 hr after transfection to block protein synthesis. Cell lysates prepared at the indicated times were immunoprecipitatedusing �-Flag antibody to precipitate tagged c-Myc and immunoblotted using the same antibody (upper panels). Quantitation of c-Myc turnoverfollowing CHX chase based on densitometrical scanning of experiments is shown (lower panel).(B) The dominant-negative Skp2�F mutant stabilizes c-Myc. HeLa cells were transfected using the expression vectors indicated, and CHXtreatment and Myc analysis were performed as in (A). Lower panel: quantitation of c-Myc levels following CHX treatment by densitometricalscanning of autoradiograms.(C) Titration of Skp2. HeLa cells were transfected with 0.33 �g c-Myc, 1 �g Skp2�F, and increasing amounts (0.1–1 �g) Skp2 wt and steady-state levels of c-Myc were analyzed by �-Flag IP and Western.(D) Effects of siRNA treatment on c-Myc degradation. HeLa cells were treated with siRNA-oligos specific for Skp2 or luciferase (Luc), or wereleft untreated (–) for 72 hr. Skp2, c-Myc, and p27 levels were analyzed using the antibodies indicated (upper left panels). Lower left panel:levels of c-Myc following CHX treatment for indicated times as described in (B). Right panel: quantitation of c-Myc turnover following siRNAas above or after lovastatin (LS), hydroxyurea (HU), or aphidicolin (AC) treatment.(E) Effects of mutations in c-Myc on its stability. c-Myc turnover was analyzed in HeLa cells transfected with the indicated Myc constructsfollowed by CHX treatment as in (B). The graphs in (A)–(E) are based on data from at least three experiments.
tion of the dominant-negative Skp2�F mutant reduced of c-Myc. Taken together, these results suggest thatSkp2 binds to and promotes ubiquitylation of c-Myc,c-Myc ubiquitylation. HA-tagged Ub did not produce Ub
smears after purification, demonstrating the specificity which ultimately results in proteasome-dependent deg-radation.of the assay. Figure 4B shows that the inhibition of c-Myc
ubiquitylation by Skp2�F was dose dependent andcould be reversed by increasing amounts of cotrans- Cell Cycle-Regulated Interaction between c-Myc
and Skp2 Coincides with G1 to S Phase Transitionfected Skp2. These results argue against indirect effectsof Skp2�F and demonstrate that expression of the domi- and Increased c-Myc Turnover
To study the biological role of the c-Myc/Skp2 interac-nant-negative Skp2 mutant inhibits whereas wt Skp2promotes c-Myc ubiquitylation in cells. tion, we investigated the dynamics of endogenous
c-Myc/Skp2 interactions during activation and cell cycleWe also examined the effects of mutations in c-Mycon ubiquitylation in cells. c-Myc mutants deficient in entry of normal peripheral blood lymphocytes (PBL).
Resting PBLs activated with PHA and IL-2 entered SSkp2 binding, i.e., lacking MB2, the C terminus, or both,were also ubiquitylated to a lesser extent than wt c-Myc phase at 36 hr after PHA�IL-2 stimulation as evidenced
by FACS analysis (data not shown) and increased cyclin(Figure 4C). These results are consistent with the modelthat Skp2 binding is necessary for efficient ubiquitylation A expression (Figure 5A). The c-Myc level was low in
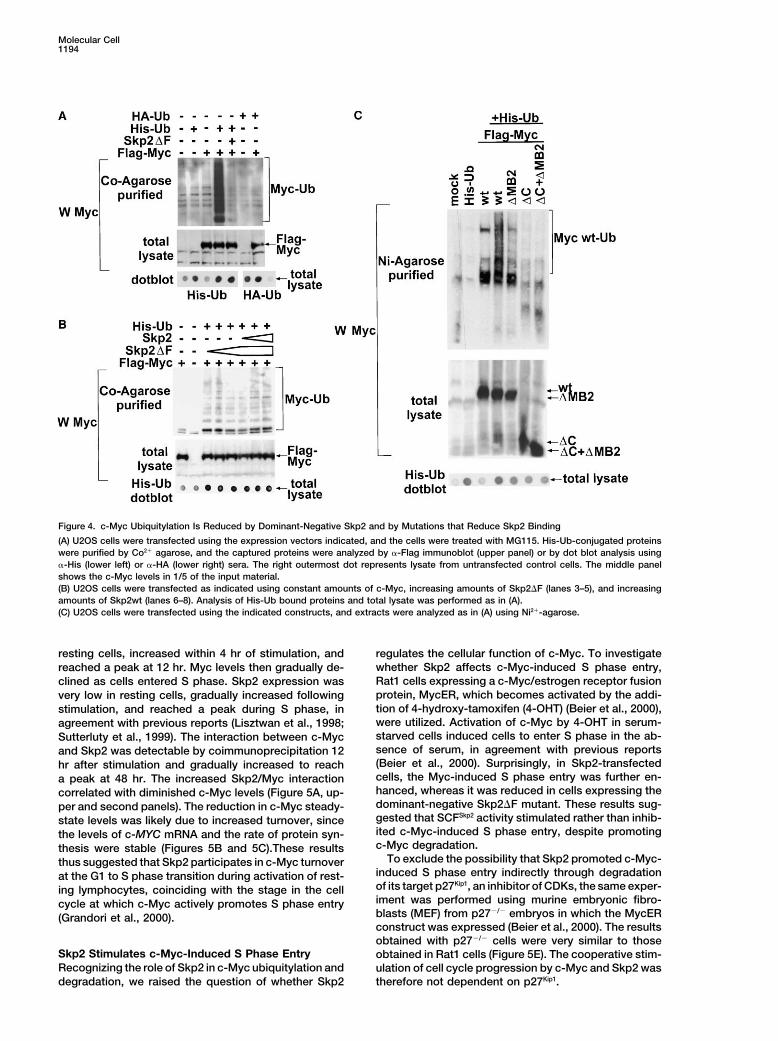
Molecular Cell1194
Figure 4. c-Myc Ubiquitylation Is Reduced by Dominant-Negative Skp2 and by Mutations that Reduce Skp2 Binding
(A) U2OS cells were transfected using the expression vectors indicated, and the cells were treated with MG115. His-Ub-conjugated proteinswere purified by Co2� agarose, and the captured proteins were analyzed by �-Flag immunoblot (upper panel) or by dot blot analysis using�-His (lower left) or �-HA (lower right) sera. The right outermost dot represents lysate from untransfected control cells. The middle panelshows the c-Myc levels in 1/5 of the input material.(B) U2OS cells were transfected as indicated using constant amounts of c-Myc, increasing amounts of Skp2�F (lanes 3–5), and increasingamounts of Skp2wt (lanes 6–8). Analysis of His-Ub bound proteins and total lysate was performed as in (A).(C) U2OS cells were transfected using the indicated constructs, and extracts were analyzed as in (A) using Ni2�-agarose.
resting cells, increased within 4 hr of stimulation, and regulates the cellular function of c-Myc. To investigatewhether Skp2 affects c-Myc-induced S phase entry,reached a peak at 12 hr. Myc levels then gradually de-Rat1 cells expressing a c-Myc/estrogen receptor fusionclined as cells entered S phase. Skp2 expression wasprotein, MycER, which becomes activated by the addi-very low in resting cells, gradually increased followingtion of 4-hydroxy-tamoxifen (4-OHT) (Beier et al., 2000),stimulation, and reached a peak during S phase, inwere utilized. Activation of c-Myc by 4-OHT in serum-agreement with previous reports (Lisztwan et al., 1998;starved cells induced cells to enter S phase in the ab-Sutterluty et al., 1999). The interaction between c-Mycsence of serum, in agreement with previous reportsand Skp2 was detectable by coimmunoprecipitation 12(Beier et al., 2000). Surprisingly, in Skp2-transfectedhr after stimulation and gradually increased to reachcells, the Myc-induced S phase entry was further en-a peak at 48 hr. The increased Skp2/Myc interactionhanced, whereas it was reduced in cells expressing thecorrelated with diminished c-Myc levels (Figure 5A, up-dominant-negative Skp2�F mutant. These results sug-per and second panels). The reduction in c-Myc steady-gested that SCFSkp2 activity stimulated rather than inhib-state levels was likely due to increased turnover, sinceited c-Myc-induced S phase entry, despite promotingthe levels of c-MYC mRNA and the rate of protein syn-c-Myc degradation.thesis were stable (Figures 5B and 5C).These results
To exclude the possibility that Skp2 promoted c-Myc-thus suggested that Skp2 participates in c-Myc turnoverinduced S phase entry indirectly through degradationat the G1 to S phase transition during activation of rest-of its target p27Kip1, an inhibitor of CDKs, the same exper-ing lymphocytes, coinciding with the stage in the celliment was performed using murine embryonic fibro-cycle at which c-Myc actively promotes S phase entryblasts (MEF) from p27�/� embryos in which the MycER(Grandori et al., 2000).construct was expressed (Beier et al., 2000). The resultsobtained with p27�/� cells were very similar to those
Skp2 Stimulates c-Myc-Induced S Phase Entry obtained in Rat1 cells (Figure 5E). The cooperative stim-Recognizing the role of Skp2 in c-Myc ubiquitylation and ulation of cell cycle progression by c-Myc and Skp2 was
therefore not dependent on p27Kip1.degradation, we raised the question of whether Skp2
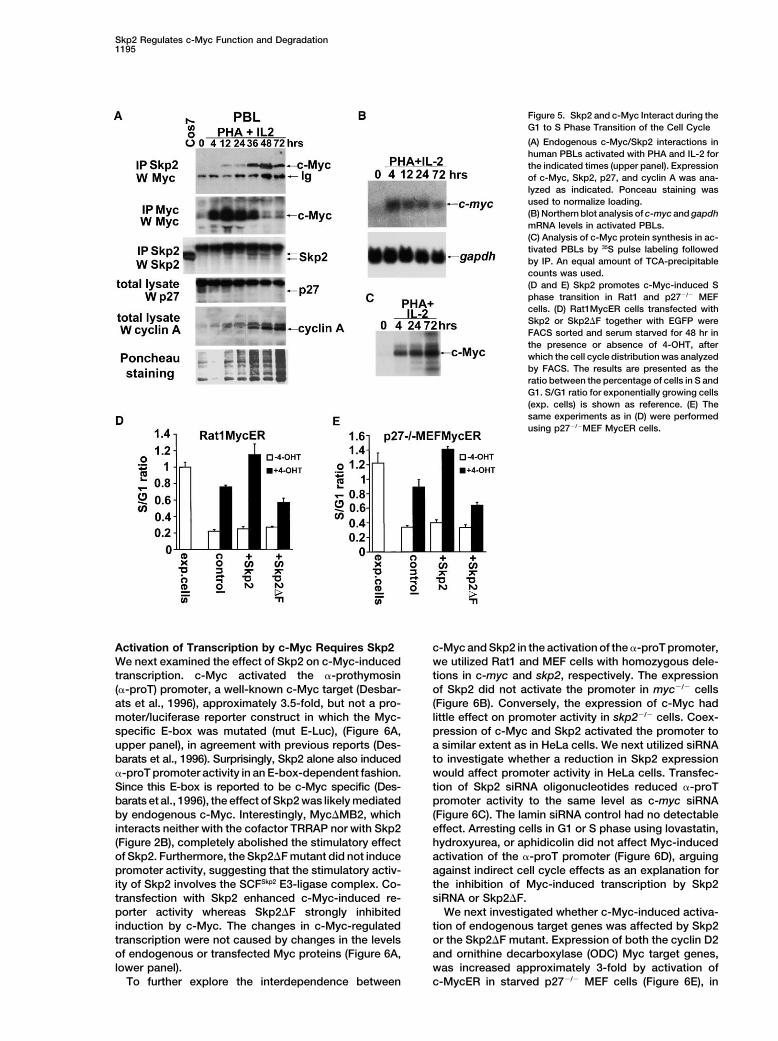
Skp2 Regulates c-Myc Function and Degradation1195
Figure 5. Skp2 and c-Myc Interact during theG1 to S Phase Transition of the Cell Cycle
(A) Endogenous c-Myc/Skp2 interactions inhuman PBLs activated with PHA and IL-2 forthe indicated times (upper panel). Expressionof c-Myc, Skp2, p27, and cyclin A was ana-lyzed as indicated. Ponceau staining wasused to normalize loading.(B) Northern blot analysis of c-myc and gapdhmRNA levels in activated PBLs.(C) Analysis of c-Myc protein synthesis in ac-tivated PBLs by 35S pulse labeling followedby IP. An equal amount of TCA-precipitablecounts was used.(D and E) Skp2 promotes c-Myc-induced Sphase transition in Rat1 and p27�/� MEFcells. (D) Rat1MycER cells transfected withSkp2 or Skp2�F together with EGFP wereFACS sorted and serum starved for 48 hr inthe presence or absence of 4-OHT, afterwhich the cell cycle distribution was analyzedby FACS. The results are presented as theratio between the percentage of cells in S andG1. S/G1 ratio for exponentially growing cells(exp. cells) is shown as reference. (E) Thesame experiments as in (D) were performedusing p27�/�MEF MycER cells.
Activation of Transcription by c-Myc Requires Skp2 c-Myc and Skp2 in the activation of the �-proT promoter,we utilized Rat1 and MEF cells with homozygous dele-We next examined the effect of Skp2 on c-Myc-induced
transcription. c-Myc activated the �-prothymosin tions in c-myc and skp2, respectively. The expressionof Skp2 did not activate the promoter in myc�/� cells(�-proT) promoter, a well-known c-Myc target (Desbar-
ats et al., 1996), approximately 3.5-fold, but not a pro- (Figure 6B). Conversely, the expression of c-Myc hadlittle effect on promoter activity in skp2�/� cells. Coex-moter/luciferase reporter construct in which the Myc-
specific E-box was mutated (mut E-Luc), (Figure 6A, pression of c-Myc and Skp2 activated the promoter toa similar extent as in HeLa cells. We next utilized siRNAupper panel), in agreement with previous reports (Des-
barats et al., 1996). Surprisingly, Skp2 alone also induced to investigate whether a reduction in Skp2 expressionwould affect promoter activity in HeLa cells. Transfec-�-proT promoter activity in an E-box-dependent fashion.
Since this E-box is reported to be c-Myc specific (Des- tion of Skp2 siRNA oligonucleotides reduced �-proTpromoter activity to the same level as c-myc siRNAbarats et al., 1996), the effect of Skp2 was likely mediated
by endogenous c-Myc. Interestingly, Myc�MB2, which (Figure 6C). The lamin siRNA control had no detectableeffect. Arresting cells in G1 or S phase using lovastatin,interacts neither with the cofactor TRRAP nor with Skp2
(Figure 2B), completely abolished the stimulatory effect hydroxyurea, or aphidicolin did not affect Myc-inducedactivation of the �-proT promoter (Figure 6D), arguingof Skp2. Furthermore, the Skp2�F mutant did not induce
promoter activity, suggesting that the stimulatory activ- against indirect cell cycle effects as an explanation forthe inhibition of Myc-induced transcription by Skp2ity of Skp2 involves the SCFSkp2 E3-ligase complex. Co-
transfection with Skp2 enhanced c-Myc-induced re- siRNA or Skp2�F.We next investigated whether c-Myc-induced activa-porter activity whereas Skp2�F strongly inhibited
induction by c-Myc. The changes in c-Myc-regulated tion of endogenous target genes was affected by Skp2or the Skp2�F mutant. Expression of both the cyclin D2transcription were not caused by changes in the levels
of endogenous or transfected Myc proteins (Figure 6A, and ornithine decarboxylase (ODC) Myc target genes,was increased approximately 3-fold by activation oflower panel).
To further explore the interdependence between c-MycER in starved p27�/� MEF cells (Figure 6E), in
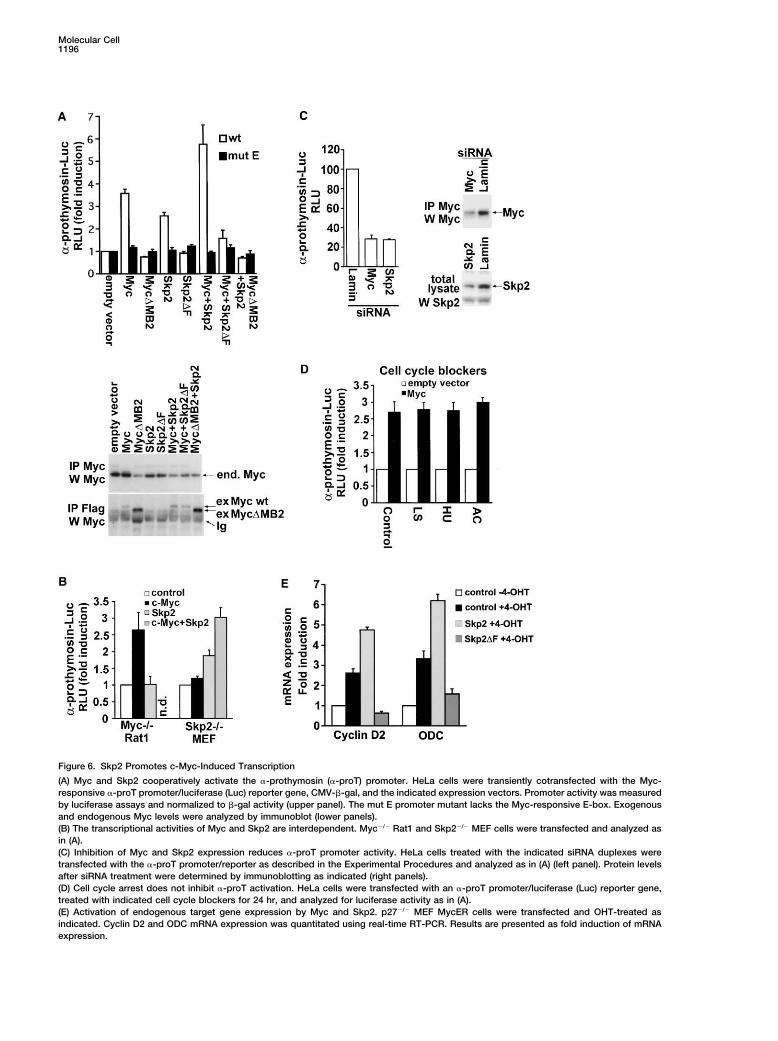
Molecular Cell1196
Figure 6. Skp2 Promotes c-Myc-Induced Transcription
(A) Myc and Skp2 cooperatively activate the �-prothymosin (�-proT) promoter. HeLa cells were transiently cotransfected with the Myc-responsive �-proT promoter/luciferase (Luc) reporter gene, CMV-�-gal, and the indicated expression vectors. Promoter activity was measuredby luciferase assays and normalized to �-gal activity (upper panel). The mut E promoter mutant lacks the Myc-responsive E-box. Exogenousand endogenous Myc levels were analyzed by immunoblot (lower panels).(B) The transcriptional activities of Myc and Skp2 are interdependent. Myc�/� Rat1 and Skp2�/� MEF cells were transfected and analyzed asin (A).(C) Inhibition of Myc and Skp2 expression reduces �-proT promoter activity. HeLa cells treated with the indicated siRNA duplexes weretransfected with the �-proT promoter/reporter as described in the Experimental Procedures and analyzed as in (A) (left panel). Protein levelsafter siRNA treatment were determined by immunoblotting as indicated (right panels).(D) Cell cycle arrest does not inhibit �-proT activation. HeLa cells were transfected with an �-proT promoter/luciferase (Luc) reporter gene,treated with indicated cell cycle blockers for 24 hr, and analyzed for luciferase activity as in (A).(E) Activation of endogenous target gene expression by Myc and Skp2. p27�/� MEF MycER cells were transfected and OHT-treated asindicated. Cyclin D2 and ODC mRNA expression was quantitated using real-time RT-PCR. Results are presented as fold induction of mRNAexpression.

Skp2 Regulates c-Myc Function and Degradation1197
Figure 7. Association of the c-Myc-Respon-sive Cyclin D2 Promoter with Skp2, Ubiqui-tylated Proteins, and Proteasome Compo-nents Is Myc Dependent
(A) Skp2 and Cul1 colocalize with c-Myc,ubiquitylated proteins, and proteasomal sub-units on the cyclin D2 promoter. ChIP assayswere performed using chromatin preparedfrom fixed HeLa cells using the indicated anti-sera or preimmune (p.i.) serum. PCR primersspecific for the cyclin D2 promoter (upperpanels) or the p21 coding region (lower pan-els) were used.(B) Binding of Skp2, Sug1, and Ub to thecyclin D2 promoter is Myc dependent. ChIPassays were performed as in (A) using theindicated antibodies, and chromatin was pre-pared from fixed myc�/� and myc�/� Rat1cells. PCR primers specific for the promotersindicated were used.(C) Expression of Skp2 in Rat1 cells. myc�/�
and myc�/� Rat1 cells were analyzed for ex-pression of Skp2 by IP and Western. Skp2overexpressed in Cos7 cells was included ascontrol.
agreement with previous reports (Bouchard et al., 2001; that Skp2 recruitment to the cyclin D2 promoter requiredc-Myc.Frank et al., 2001). Transfection of Skp2 enhanced
c-Myc-induced expression of both cyclin D2 and ODC The presence of two subunits of SCFSkp2 at the cyclinD2 promoter led us to the question of whether protea-gene expression, whereas the Skp2�F mutant inhibited
their induction by c-Myc. some subunits and/or ubiquitylated proteins could bedetected at the promoter. Indeed, Sug1/Rpt6 (FigureTaken together, these results suggest that Myc-
induced transcription of target promoters requires Skp2 7A, middle upper panel) and Rpt3 (right upper panel),both subunits of the hexameric ring of AAA ATPasesexpression and activity. Conversely, Skp2-induced acti-
vation of the same promoters requires c-Myc. (APIS) of the 19S regulatory particle, were present onthe cyclin D2 promoter. Weaker signals, which wereclearly above background as shown by three control
Myc-Dependent Association of Skp2, Ubiquitylated antibodies, were detected using antibodies directedProteins, and Proteasomal Subunits against the non-ATPase subunit Rpn7 of the 19S particlewith the Cyclin D2 Promoter and the �2 subunit of the 20S proteasome core particle.The results above and the nuclear localization of the Furthermore, antibodies directed against mono- and poly-c-Myc/Skp2 complex (Figure 2D) prompted us to raise ubiquitylated proteins precipitated this chromosomal re-the question of whether Skp2 acts as a coactivator with gion (Figure 7A, middle upper panel). None of the anti-Myc at target promoters. Chromatin immunoprecipita- sera immunoprecipitated chromatin from the codingtion (ChIP) assays using HeLa cells demonstrated the region of p21Cip1, used as a control (Figure 7A, lowerpresence of c-Myc and Max at the E-box region of the panels). Interestingly, neither Skp2, Sug1, nor ubiqui-cyclin D2 promoter (Figure 7A, left upper panel), pre- tylated proteins were detected at the promoter in Rat1viously shown to bind c-Myc (Bouchard et al., 2001). myc�/� cells (Figure 7B, left upper panel). These resultsInterestingly, Skp2 and Cul1 were also detected at the demonstrate that this cyclin D2 promoter region is asso-promoter. Although both Skp2 and c-Myc bound to the ciated with Skp2, ubiquitylated proteins, and severalcyclin D2 promoter in Rat1 myc�/� cells, neither c-Myc subunits of the proteasome in a Myc-dependentnor Skp2 was present at the promoter in Rat1 myc�/�
manner.cells (Figure 7B, upper panels). Binding of the E2F4transcription factor to the B23 promoter was observedin both Rat1 myc�/� and myc�/� cells (Figure 7B, lower Discussionpanels) in agreement with previous reports (Zeller et al.,2001), demonstrating that the quality of the chromatin Previous studies have shown that the c-Myc oncopro-
tein, like many other transcription factors, is rapidlypreparation from myc�/� cells was satisfactory. Further,similar expression of Skp2 was observed in Rat1 myc�/� turned over via the ubiquitin/proteasome pathway and
that regions of c-Myc important for this turnover coin-and myc�/� cells (Figure 7C). These results suggested

Molecular Cell1198
cide with regions necessary for its transcription activa- ubiquitylation and degradation of c-Myc occurs at thetion function (Kato et al., 1990; Flinn et al., 1998; Salghetti promoter, it is intriguing that not only Skp2 but alsoet al., 1999; Sears et al., 1999; Bahram et al., 2000; ubiquitylated proteins and subunits of the proteasomeGregory and Hann, 2000). The proteins responsible for are detected at the promoter in a c-Myc-dependentrecognition and targeting of c-Myc for ubiquitylation and manner. Further, our observations that the Skp2�F mu-proteasomal degradation have remained unidentified, tant, which binds and stabilizes c-Myc but does notand their roles in transcription activation by c-Myc are recruit SCF, not only has lost coactivator activity butunknown. The work presented provides evidence that represses c-Myc-induced transcription, and that Cul1Skp2 interacts with c-Myc in vivo and promotes its ubiq- is also detected at promoter, suggest that the E3 ligaseuitylation and degradation. The interaction between activity of Skp2 is essential for its coactivator function.c-Myc and Skp2 seems to be direct since it was also In what way could E3 ligase activity stimulate c-Myc-demonstrated in vitro. The role of Skp2 is further empha- induced transcription? One possibility is that SCFSkp2
sized by the requirement for Skp2 expression and func- ubiquitylates and degrades negative regulators of tran-tion as well as for the Skp2-interacting regions of c-Myc scription at the promoter. This could also be part of anfor efficient c-Myc ubiquitylation and turnover. We have autoregulatory loop, where the Myc activator proteinnot, however, been able to demonstrate SCFSkp2-depen- needs to be eliminated at some step in order to completedent ubiquitylation/degradation of c-Myc in vitro, possi- the transcription cycle. Another possibility is that ubiqui-bly due to requirements for additional cofactors or modi- tin modifications of c-Myc or other substrates at thefications. We cannot, therefore, exclude the possibility promoter play a nonproteolytic function in, for instance,that another E3 ligase mediates the ubiquitylation of protein-protein interactions of importance for transcrip-c-Myc. Our observations also do not exclude other path- tion, as has been suggested for Met30-directed ubiqui-ways of c-Myc degradation independent of SCFSkp2 in tylation of the Met4 transcription factor (Kaiser et al.,different cell types or in response to different signals. 2000; Kuras et al., 2002). Further, recent reports haveIndications supporting this view are that only a subset suggested that certain proteasomal subunits such asof previously described degrons in c-Myc (Flinn et al., the APIS complex, which is recruited through binding1998; Salghetti et al., 1999; Bahram et al., 2000; Gregory to ubiquitylated proteins, may carry out nonproteolyticand Hann, 2000) seem to play a role in Skp2-induced functions at the promoter, either alone or as part of theturnover, and that the turnover rate of c-Myc is compara- proteasome (Ferdous et al., 2001; Gonzalez et al., 2002;tively high also in, e.g., the early G1 phase of the cell for reviews see Conaway et al., 2002; Ottosen et al.,cycle, when c-Myc and Skp2 do not seem to interact 2002; Muratani and Tansey, 2003). These APIS ATPasesstrongly. have been implicated in the transcription process
What is the biological role of the Skp2/c-Myc interac- through their chaperone-like or remodeling activity. Ittion? Its dependence of the MB2 and bHLH-Zip do- also remains to be determined whether degradation ofmains, which are the most conserved and functionally c-Myc is a necessary step for activation of transcription.important regions of c-Myc, indicates that this interac- This general model has been proposed in the “licensing”tion may be essential. The timing of the interaction event hypothesis (Salghetti et al., 2001), linking transcriptionat the G1 to S phase transition during activation of nor- factor activity to their destruction in order to maintainmal lymphocytes is intriguing, since this is the time at stringent control of transcription activation in cells.which c-Myc is known to be required for S phase entry. Studies of transcription regulatory proteins during re-One possibility is that degradation is required to prevent cent years have suggested that a major function of manypersistent c-Myc activity, which can be damaging to the of these factors, including c-Myc, is to recruit coactiva-cell. However, our data demonstrate that Skp2 rather tor complexes with enzymatic activities such as HAT-than dampening c-Myc activity instead promotes c-Myc- and chromatin remodeling complexes to promotersinduced S phase entry. The reason for this is apparently
(Amati et al., 2001). The work presented here suggeststhat Skp2 expression and activity not only promotes but
an additional important function of Myc, namely the re-also is required for c-Myc-induced transcription of its
cruitment of coactivators with E3 ubiquitin ligase activitytarget genes. Conversely, our data suggest that Skp2-to its target promoters. Further studies will be requiredinduced transcription of c-Myc target genes requiresto clarify the role of this enzymatic activity in the regula-c-Myc expression and activity. This interdependencetion of transcription.seems to require regions of c-Myc interacting with Skp2.
The fact that the interaction takes place in the nucleusExperimental Proceduresand that Skp2 binds specifically to the Myc-responsive
E-box region of the cyclin D2 promoter only in the pres-Cell Culture, Transfections, and Cell Cycle Analysis
ence of c-Myc suggests that Skp2 is recruited to target Cos 7, HeLa, U2OS, Rat1, and MEF cells were maintained in Dulbec-promoters as a coactivator of c-Myc-induced transcrip- co’s modified essential medium (DMEM) supplemented with 10%tion. Our data, therefore, support the hypothesis that fetal bovine serum and antibiotics. Peripheral blood mononuclearthere is a close relationship between the activities of cells (PBMNC) were purified from buffy coats by Ficoll separation
and activated by addition of 1 �g/ml phytohemagglutinin (PHA,transcription factors and their ubiquitylation and degra-Sigma) and 10 ng/ml IL-2 (Amersham). Skp2�/� MEFs have beendation, based on observations during recent years (Moli-described (Nakayama et al., 2000). Rat1 MycER and MEF p27�/�
nari et al., 1999; Kaiser et al., 2000; Salghetti et al., 2000,MycER cells (Beier et al., 2000) were kindly provided by C. Bouchard
2001; Chi et al., 2001; Ferdous et al., 2001; Gonzalez and M. Eilers; Rat1 HO 15.19 (myc�/�) and Rat1 TGR (myc�/�) cellset al., 2002; Kuras et al., 2002 for recent reviews see (Mateyak et al., 1997) were kindly provided by J.M. Sedivy. TransientConaway et al., 2002; Ottosen et al., 2002; Muratani and transfections were carried out using FuGene (Roche). Cells trans-
fected with EGFP were sorted by FACSVantage SE (Becton Dickin-Tansey, 2003). Although we do not have evidence that

Skp2 Regulates c-Myc Function and Degradation1199
son). For cell cycle analysis, MycER cells were serum starved for or lamin were used. For luciferase assays, cells were pretreated withsiRNA duplexes for 18 hr followed by transfection of the luciferase48 hr in the presence or absence of 200 nM 4-hydroxy-tamoxifen
(4-OHT) (Sigma). The cell cycle distribution was analyzed using construct. Six hours posttransfection the cells were siRNA afterwhich cells were analyzed 24 hr later.FACSort (Becton Dickinson), and the obtained data were analyzed
with CellQuest (Becton Dickinson) and ModFit LT 3.0 (Verity Soft-ware House Inc.). For blocking the HeLa cell cycle in G1 and in S Bimolecular Fluorescence Complementationphase, lovastatin (40 �M), hydroxyurea (2.5 mM), or aphidicolin (5 BiFC was performed essentially as described (Hu et al., 2002). Cos�g/ml), respectively, was applied to the cultures for 24 hr. 7 cells were transfected with YN and YC fusion constructs alone or
in combination and incubated for 24 hr followed by further incuba-Plasmids tion at 30�C overnight. Cell nuclei were visualized by DAPI staining.pCDNA3-FlagMyc wt, �MB1 and �MB2, and the pGEX construc- Fluorescence emissions were observed in living cells using an in-tions were kindly provided by B. Luscher; pCDNA3-MT-Skp2, verted microscope (Nikon, Opthiphot, Japan), and images were cap-pCDNA3-Flag-Skp1, and pCDNA3-HA-Cul1 by W. Krek; CMV-His6- tured with a Nikon FX-35 WA camera. YFP fluorescence was mea-Ub and CMV-HA-Ub by D. Bohmann; and pCIneo-HA-Stat1 by A. sured by excitation at 513 nm and emission at 527 nm.Dimberg. �-TRCP wt and �F were kindly provided by R. Benarous;NFB42 by R. Pittman, Myc-GFP, and pEYFP by A. Wright; and RT-PCR and Northern BlotpEGFP by H. Jernberg-Wiklund. The Fbl2 and Fbl8 constructs were Four micrograms total RNA was converted to cDNA with RevertAidobtained from RZPD (Berlin), and Fbl5 was kindly provided by T. M-MuLV Reverse Transcriptase (Fermenta). Primers and probes forImamura. Constructs containing �-prothymosin-luciferase (wt and RT-PCR were designed by the software Primer Express (Perklinmutant) have been described (Desbarats et al., 1996) and were kindly Elmer Applied Biosystems). Sequences are provided upon request.provided by M. Eilers. pCIneo-Myc wt has been described previously As a reference GAPDH was analyzed by using Taqman Rodent(Bahram et al., 2000). Details for the construction of Skp2-YFP(1–154) GAPDH Control Reagents (VIC) (Applied Biosystems). RT-PCR reac-(Skp2YN), Myc(342–439)YFP(155–238), Myc�bHZYC, I�B�YN, p50YN, tions were performed using the Taqman PCR Core Reagent kit (Ap-p65YC, pCIneo-p27, pCDNA3-Flag-Myc�MB1��MB2, pCDNA3-Flag- plied Biosystems) in triplicates with the ABI PRISM 7700 SequenceMyc�N, �N, �C, �C��MB2, �2–293, �2–366, �148–439, �95–439, Detector (Perklin Elmer Applied Biosystems). The Northern blot pro-and pCDNA3-MT-Skp2�F are available upon request. cedure and probe for human c-MYC were as described (Bahram et
al., 1999) using 15 �g of total RNA per lane.GST Fusion Protein Purification, In Vitro Translation,and GST Pull-Down
AcknowledgmentsPurification of GST-fusion proteins from overnight cultures of bacte-ria and GST-pull-downs were performed essentially as described
We thank B. Luscher, N. Malek, D. Bohmann, A. Wright, W. Krek,(Gonzalez et al., 2002). In vitro translations were performed usingA. Dimberg, H. Jernberg-Wiklund, L. Hengst, S. Elledge, R. Pittman,the TNT T7 Coupled Reticulocyte Lysate System kit (Promega). TheR. Benarous, J. Weinstein, P. Chambon, J.M. Sedivy, T. Imamura,GST pull-down reaction was performed using 2 �g GST fusion pro-M. Henriksson, C. Bouchard, M. Eilers, and C.-D. Hu for reagents.tein and 3 �l prewashed in vitro translated protein.We also thank C. Gillberg, H. Claerhout, J. Grawe, and Ge11 biotechstudents for technical assistance. We are grateful to B. Luscher, J.
Chromatin Immunoprecipitation and Reporter Gene AssaysVervoorts, M. Thompson, S. Frank, B. Amati, N. Malek, C. Svensson,
HeLa cells were lysed and crosslinked with formaldehyde as de-H. Ronne, D. Balcunias, M. Thelander, J. Eriksson, E. Gronroos, L.
scribed (Bouchard et al., 2001; Frank et al., 2001). Chromatin DNAProkunina, T. Stromberg, A-S Hoglund, A. Blokzijl, and C. Eberstark
was sonicated to an average size of 300–500 bp. Immunoprecipita-for valuable help and discussions and B. Luscher for critically read-
tions were carried out in RIPA buffer with the following Abs: �-IG0ing the manuscript. The FACSvantage SE equipment was financed
(preimmune serum), polyclonal �-pan-Myc (IG9), �-Skp2 H-435,by the Wallenberg Foundation. This work was supported by the
�-Max C-17X, and �-E2F4 A20 (all St. Cruz), �-Cul1 ZL-18 (Zymed),Swedish Cancer Foundation, Swedish Children Cancer Foundation,
�-Sug1/Rpt6, �-ubiquitin clone FK2, �-Rpt3, �-proteasome subunitThe Lovisa & Thielman Foundation, and Agrifungen.
�2, and �-Rpn7 (all Affiniti). Immunoprecipitates were collected withprotein G-sepharose beads, washed, and eluted in TE, 1% SDS.
Received: September 19, 2002Purified DNA samples were analyzed by PCR. PCR primer se-Revised: May 2, 2003quences are provided upon request. Transient luciferase andAccepted: May 5, 2003�-galactosidase reporter gene assays were performed as describedPublished: May 22, 2003(Desbarats et al., 1996).
References35S-Labeling, Cell Lysis, Immunoprecipitations,Immunoblot, and Ubiquitin Assays
Amati, B., Frank, S.R., Donjerkovic, D., and Taubert, S. (2001). Func-In vivo labeling of cells by 35S-methionine, cell lysis, immunoprecipi-tion of the c-Myc oncoprotein in chromatin remodeling and tran-tations, and immunoblot were performed as described (Bahram etscription. Biochim. Biophys. Acta 1471, M135–M145.al., 1999). One hundred �g/ml cycloheximide (Sigma) was used for
blocking protein synthesis in protein chase experiments. Ubiquitin Bahram, F., Wu, S., Oberg, F., Luscher, B., and Larsson, L.G. (1999).assays were performed as described (Treier et al., 1994; Bahram et Posttranslational regulation of Myc function in response to phorbolal., 2000). Cells were treated with the proteasome inhibitor Z-Leu- ester/interferon-gamma-induced differentiation of v-Myc-trans-Leu-Leu-H aldehyde (MG115) (Peptides International) at a concen- formed U-937 monoblasts. Blood 93, 3900–3912.tration of 50 �M for 2 hr prior to cell lysis. Abs used for immunoblot
Bahram, F., von der Lehr, N., Cetinkaya, C., and Larsson, L.G. (2000).were �-c-Myc (C33X), �-Myc (N262), �-9E10HRP, �-GST (Z5), �-p27c-Myc hot spot mutations in lymphomas result in inefficient ubiquiti-(C-19), �-His (H-15), �-cyclin A (BF683X) (all Santa Cruz), �-Flag M2nation and decreased proteasome-mediated turnover. Blood 95,and M5 (Sigma), and �-HA 3F10 (Roche).2104–2110.
Beier, R., Burgin, A., Kiermaier, A., Fero, M., Karsunky, H., Saffrich,Silencing by Small Interfering RNAR., Moroy, T., Ansorge, W., Roberts, J., and Eilers, M. (2000). Induc-HeLa cells seeded onto 6-well plates at a confluence of 40%–50%tion of cyclin E-cdk2 kinase activity, E2F-dependent transcriptionwere transfected with siRNA duplexes using oligofectamine (In-and cell growth by Myc are genetically separable events. EMBO J.vitrogen) as described (Elbashir et al., 2001). Protein levels were19, 5813–5823.analyzed by immunoblot. The siRNA duplexes used for Skp2 and
c-Myc silencing were in general 21 bp synthetic oligonucleotides Bouchard, C., Dittrich, O., Kiermaier, A., Dohmann, K., Menkel, A.,Eilers, M., and Luscher, B. (2001). Regulation of cyclin D2 gene(Dharmacon Research); sequences are available on request. As con-
trols 21 nt siRNA duplexes for silencing of the firefly luciferase gene expression by the Myc/Max/Mad network: Myc-dependent TRRAP

Molecular Cell1200
recruitment and histone acetylation at the cyclin D2 promoter. Genes conjugating enzyme CDC34 with the F-box protein p45(SKP2): evi-dence for evolutionary conservation in the subunit composition ofDev. 15, 2042–2047.the CDC34-SCF pathway. EMBO J. 17, 368–383.Brower, C.S., Sato, S., Tomomori-Sato, C., Kamura, T., Pause, A.,
Stearman, R., Klausner, R.D., Malik, S., Lane, W.S., Sorokina, I., et Mateyak, M.K., Obaya, A.J., Adachi, S., and Sedivy, J.M. (1997).Phenotypes of c-Myc-deficient rat fibroblasts isolated by targetedal. (2002). Mammalian mediator subunit mMED8 is an Elongin BC-
interacting protein that can assemble with Cul2 and Rbx1 to recon- homologous recombination. Cell Growth Differ. 8, 1039–1048.stitute a ubiquitin ligase. Proc. Natl. Acad. Sci. USA 99, 10353–10358. McMahon, S.B., Van Buskirk, H.A., Dugan, K.A., Copeland, T.D.,
and Cole, M.D. (1998). The novel ATM-related protein TRRAP is anCarrano, A.C., Eytan, E., Hershko, A., and Pagano, M. (1999). SKP2is required for ubiquitin-mediated degradation of the CDK inhibitor essential cofactor for the c-Myc and E2F oncoproteins. Cell 94,
363–374.p27. Nat. Cell Biol. 1, 193–199.
Chi, Y., Huddleston, M.J., Zhang, X., Young, R.A., Annan, R.S., Carr, McMahon, S.B., Wood, M.A., and Cole, M.D. (2000). The essentialcofactor TRRAP recruits the histone acetyltransferase hGCN5 toS.A., and Deshaies, R.J. (2001). Negative regulation of Gcn4 and
Msn2 transcription factors by Srb10 cyclin-dependent kinase. c-Myc. Mol. Cell. Biol. 20, 556–562.Genes Dev. 15, 1078–1092. Molinari, E., Gilman, M., and Natesan, S. (1999). Proteasome-medi-
ated degradation of transcriptional activators correlates with activa-Conaway, R.C., Brower, C.S., and Conaway, J.W. (2002). Emergingroles of ubiquitin in transcription regulation. Science 296, 1254– tion domain potency in vivo. EMBO J. 18, 6439–6447.1258. Muratani, M., and Tansey, W.P. (2003). How the ubiquitin-protea-
some system controls transcription. Nat. Rev. Mol. Cell Biol. 4,DeSalle, L.M., and Pagano, M. (2001). Regulation of the G1 to Stransition by the ubiquitin pathway. FEBS Lett. 490, 179–189. 192–201.
Nakayama, K., Nagahama, H., Minamishima, Y.A., Matsumoto, M.,Desbarats, L., Gaubatz, S., and Eilers, M. (1996). Discriminationbetween different E-box-binding proteins at an endogenous target Nakamichi, I., Kitagawa, K., Shirane, M., Tsunematsu, R., Tsukiyama,
T., Ishida, N., et al. (2000). Targeted disruption of Skp2 results ingene of c-myc. Genes Dev. 10, 447–460.accumulation of cyclin E and p27(Kip1), polyploidy and centrosomeElbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K.,overduplication. EMBO J. 19, 2069–2081.and Tuschl, T. (2001). Duplexes of 21-nucleotide RNAs mediate RNA
interference in cultured mammalian cells. Nature 411, 494–498. Ottosen, S., Herrera, F.J., and Triezenberg, S.J. (2002). Transcrip-tion. Proteasome parts at gene promoters. Science 296, 479–481.Ferdous, A., Gonzalez, F., Sun, L., Kodadek, T., and Johnston, S.A.
(2001). The 19S regulatory particle of the proteasome is required Salghetti, S.E., Kim, S.Y., and Tansey, W.P. (1999). Destruction ofMyc by ubiquitin-mediated proteolysis: cancer-associated andfor efficient transcription elongation by RNA polymerase II. Mol. Cell
7, 981–991. transforming mutations stabilize Myc. EMBO J. 18, 717–726.
Salghetti, S.E., Muratani, M., Wijnen, H., Futcher, B., and Tansey,Flinn, E.M., Busch, C.M., and Wright, A.P. (1998). myc boxes, whichare conserved in myc family proteins, are signals for protein degra- W.P. (2000). Functional overlap of sequences that activate transcrip-
tion and signal ubiquitin-mediated proteolysis. Proc. Natl. Acad. Sci.dation via the proteasome. Mol. Cell. Biol. 18, 5961–5969.USA 97, 3118–3123.Frank, S.R., Schroeder, M., Fernandez, P., Taubert, S., and Amati,
B. (2001). Binding of c-Myc to chromatin mediates mitogen-induced Salghetti, S.E., Caudy, A.A., Chenoweth, J.G., and Tansey, W.P.(2001). Regulation of transcriptional activation domain function byacetylation of histone H4 and gene activation. Genes Dev. 15, 2069–
2082. ubiquitin. Science 293, 1651–1653.
Sears, R., Leone, G., DeGregori, J., and Nevins, J.R. (1999). RasGonzalez, F., Delahodde, A., Kodadek, T., and Johnston, S.A. (2002).Recruitment of a 19S proteasome subcomplex to an activated pro- enhances Myc protein stability. Mol. Cell 3, 169–179.moter. Science 296, 548–550. Sutterluty, H., Chatelain, E., Marti, A., Wirbelauer, C., Senften, M.,
Muller, U., and Krek, W. (1999). p45SKP2 promotes p27Kip1 degra-Grandori, C., Cowley, S.M., James, L.P., and Eisenman, R.N. (2000).The Myc/Max/Mad network and the transcriptional control of cell dation and induces S phase in quiescent cells. Nat. Cell Biol. 1,
207–214.behavior. Annu. Rev. Cell Dev. Biol. 16, 653–699.
Gregory, M.A., and Hann, S.R. (2000). c-Myc proteolysis by the Treier, M., Staszewski, L.M., and Bohmann, D. (1994). Ubiquitin-dependent c-Jun degradation in vivo is mediated by the delta do-ubiquitin-proteasome pathway: stabilization of c-Myc in Burkitt’s
lymphoma cells. Mol. Cell. Biol. 20, 2423–2435. main. Cell 78, 787–798.
Weissman, A.M. (2001). Themes and variations on ubiquitylation.Gstaiger, M., Jordan, R., Lim, M., Catzavelos, C., Mestan, J., Slinger-land, J., and Krek, W. (2001). Skp2 is oncogenic and overexpressed Nat. Rev. Mol. Cell Biol. 2, 169–178.in human cancers. Proc. Natl. Acad. Sci. USA 98, 5043–5048. Zeller, K.I., Haggerty, T.J., Barrett, J.F., Guo, Q., Wonsey, D.R., and
Dang, C.V. (2001). Characterization of nucleophosmin (B23) as aHershko, A., and Ciechanover, A. (1998). The ubiquitin system. Annu.Rev. Biochem. 67, 425–479. Myc target by scanning chromatin immunoprecipitation. J. Biol.
Chem. 276, 48285–48291.Hu, C.D., Chinenov, Y., and Kerppola, T.K. (2002). Visualization ofinteractions among bZIP and Rel family proteins in living cells usingbimolecular fluorescence complementation. Mol. Cell 9, 789–798.
Kaiser, P., Flick, K., Wittenberg, C., and Reed, S.I. (2000). Regulationof transcription by ubiquitination without proteolysis: Cdc34/SCF(Met30)-mediated inactivation of the transcription factor Met4.Cell 102, 303–314.
Kato, G.J., Barrett, J., Villa-Garcia, M., and Dang, C.V. (1990). Anamino-terminal c-myc domain required for neoplastic transforma-tion activates transcription. Mol. Cell. Biol. 10, 5914–5920.
Kuras, L., Rouillon, A., Lee, T., Barbey, R., Tyers, M., and Thomas, D.(2002). Dual regulation of the met4 transcription factor by ubiquitin-dependent degradation and inhibition of promoter recruitment. Mol.Cell 10, 69–80.
Latres, E., Chiarle, R., Schulman, B.A., Pavletich, N.P., Pellicer, A.,Inghirami, G., and Pagano, M. (2001). Role of the F-box protein Skp2in lymphomagenesis. Proc. Natl. Acad. Sci. USA 98, 2515–2520.
Lisztwan, J., Marti, A., Sutterluty, H., Gstaiger, M., Wirbelauer, C.,and Krek, W. (1998). Association of human CUL-1 and ubiquitin-





























