Embed Size (px)
Citation preview

Cardiovascular Research 58 (2003) 399–409www.elsevier.com/ locate/cardiores
C ardiomyocytes derived from embryonic stem cells resemblecardiomyocytes of the embryonic heart tube
Arnoud C. Fijnvandraat, Antoni C.G. van Ginneken, Piet A.J. de Boer, Jan M. Ruijter,*Vincent M. Christoffels, Antoon F.M. Moorman, Ronald H. Lekanne Deprez
Experimental and Molecular Cardiology Group, Academic Medical Center, University of Amsterdam, Meibergdreef 15, 1105AZ Amsterdam,The Netherlands
Received 12 September 2002; accepted 11 February 2003
Abstract
Objective: After formation of the linear heart tube a chamber-specific program of gene expression becomes active that underlies theformation of the chamber myocardium. To assess whether this program is recapitulated in in vitro differentiated embryonic stem cells, weperformed qualitative and quantitative analyses of cardiogenesis in vivo and in vitro.Methods: Gene expression profiles were made by insitu hybridisation and real-time PCR and electrophysiological profiles by patch clamp analyses of cardiomyocytes derived from timeseries of differentiating HM1 mouse embryonic stem cells and from embryonic and adult mouse hearts.Results: In embryoid bodies thein situ patterns of expression ofa-myosin heavy chain, myosin light chain 2a and sarcoendoplasmic reticulum calcium ATPase 2a weresimilar to that of the heart muscle-specific marker gene cardiac troponin I. Myosin light chain 2v was expressed in part of the cardiactroponin I-expressing area, indicating heterogeneity within the cardiac cell population. Atrial natriuretic factor expression, indicative ofthe chamber-type program, could only very occasionally be detected by in situ hybridisation. Quantitative reverse transcriptase PCRshowed that all cardiac genes, most notably atrial natriuretic factor, were expressed at relatively low levels, similar to those in embryonichearts at embryonic day 8.75–9. Analysis of the electrophysiological characteristics of embryonic stem cell-derived cardiomyocytesshowed an increase of the upstroke velocity and a shorter duration of the action potential during prolonged differentiation in vitro. Whenembryonic mouse heart compartments of embryonic day 12.5 were used as a reference, the electrophysiological characteristics of asubstantial part of the embryonic stem cell-derived cardiomyocytes were most reminiscent to those observed in the embryonic outflowtract.Conclusion: Together, these data suggest that most cardiomyocytes acquired by differentiation of embryonic stem cells maintain aphenotype reminiscent of that of the cardiomyocytes of the primary heart tube, and hardly any myocytes develop a chamber myocardialphenotype. 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.
Keywords: Embryonic stem cells; Developmental biology; Embryology; Gene expression; Membrane currents
1 . Introduction of atrial natriuretic factor (Anf) mRNA in the ventriclesand the atria[1]. Differentiation of embryonic stem (ES)
The vertebrate heart is formed in vivo as a linear tube of cells to the cardiac lineage provides an important model toso-called primary myocardium. At embryonic day (E)8.25 investigate mechanisms and genes involved in the earliestchamber formation is initiated, accompanied by expression steps of cardiac development[2]. Moreover, production of
cardiomyocytes in an in vitro system offers a potentiallyunlimited source of donor material for grafting therapies[3]. Both applications of the ES cell differentiation model
*Corresponding author. Department of Anatomy and Embryology, require proper characterisation of the formed heart cells. AAcademic Medical Centre, University of Amsterdam, Meibergdreef 15,1105 AZ Amsterdam, The Netherlands. Tel.:131-20-566-5415; fax:131-20-697-6177.
E-mail address: [email protected](R.H. Lekanne Deprez). Time for primary review 33 days.
0008-6363/03/$ – see front matter 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.doi:10.1016/S0008-6363(03)00282-7
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

400 A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409
developmental in vivo standard enables discrimination 310) days of incubation in hanging drops, EBs werebetween the initial primary myocardial cells and later brought in a floating culture, one EB per culture welldeveloping ventricular and atrial chamber myocytes. (24-well plate, COSTAR, diameter ca. 1.5 cm) coated with
It is the focus of many research groups to analyse how a layer of 1% sterile agar in differentiation medium toin vitro differentiation of ES cells relates to in vivo prevent attachment of the EB to the culture well. Differen-cardiogenesis. Besides an inventory of gene expression in tiation medium contains ISCOVES MDM culture mediumES cell-derived cardiomyocytes, these studies often in- (Gibco-BRL) enriched with 20% foetal calf serumclude electrophysiological characteristics. In these studies (Bodinco, cat. no. 39454), 13 amino acids and 13the expression of myosin light chain (Mlc)2v[3–10], Anf penicillin /streptavidin (both from Gibco-BRL). Attach-[5,9] and Mlc2a mRNA[3,10] has been used to ascribe a ment cultures were established by plating EBs on gelatin-ventricular or atrial phenotype to ES cell-derived car- coated culture wells after 4 days in a floating culture[16].diomyocytes. A confusing factor in these studies is that, Medium was changed every other day.although these genes are confined to their nominal com- FVB mouse embryos were obtained from timed-preg-partments in the heart after birth[11,12], none of them is nant animals and used for RNA isolation, electrophysiolo-restricted to these compartments at the early embryonic gy and in situ hybridisation. Noon of the day of detectionstages (before E13). Moreover, quantitative data are of the vaginal plug was considered E0.5. The investigationscarce. An additional complicating factor is the scarcity of conforms with theGuide for the Care and Use ofin vivo electrophysiological data on mouse cardiac de- Laboratory Animals published by the US National Insti-velopment that can be used as a standard to characterise tutes of Health (NIH Publication NO. 85-23, revisedES cell-derived cardiomyocytes. In the present study we 1996). The experiments were approved by The Committeeassessed the expression of a number of cardiac genes for Experiments on Animals (DEC) of the University ofamong which those that have been used in ES cell studies Amsterdam, The Netherlands.to define the phenotypes of ES cell-derived car-diomyocytes to analyse the progress of cardiac differentia- 2 .1.1. RNA isolationtion. We performed a quantitative and qualitative com- RNA was isolated from undifferentiated ES cells, fromparison between ES cell-derived cardiomyocytes and car- pools of seven to 10 beating EBs cultured for 314 untildiomyocytes derived from embryonic mice, based on in 3130 days in floating culture or from 3125 until 3139situ hybridisation (ISH), quantitative reverse transcriptase days cultured in attachment culture, and from hearts from(RT)-PCR and electrophysiology. The characteristics of E8.5 to 19, neonatal day (N)1, N7 and N14 and adult (3the ES cell-derived cardiomyocytes are largely similar to months) FVB mice. For E8.5–15 two pools of eight to 13those observed in embryonic cardiomyocytes at E8.75–9. hearts per time point were studied. For E19–N14 twoWe conclude that the development of ES cell-derived pools of two hearts were used and four adult hearts werecardiomyocytes includes hardly any formation of chamber used. Total RNA was isolated using an RNA isolation kitmyocardium and therefore does not significantly progress (RNeasy, Qiagen) according to the instructions of thebeyond that of cardiomyocytes of the embryonic heart tube manufacturer. To remove contaminating genomic DNA,stage in vivo. the RNA preparation (10–100mg) was subsequently
incubated with 10 units RQ1 RNase-free DNase (Promega)for 30 min at 378C, extracted with phenol and chloroform
2 . Methods and finally precipitated and dissolved in 3 mM Tris–HCl,pH 7.5–0.2 mM EDTA. Total RNA concentration was
Mouse locus names according to Mouse Genome Infor- determined spectrophotometrically at 260 nm.matics (http: / /www.informatics.jax.org) are Atp2a1 (sar-coendoplasmic reticulum calcium ATPase),Myh6 (a- 2 .1.2. Myocyte isolation for electrophysiologymyosin heavy chain),Mylc2a (myosin light chain 2a), Isolation of cardiomyocytes from EBs was performed asMylpc (myosin light chain 2v),Nppa (atrial natriuretic described by Maltsev et al.[15]. Myocytes from E12.5factor), Slc8a1 (sodium calcium exchanger 1), andTnni3 mouse hearts were prepared as follows. Pregnant mice(cardiac troponin I). were killed by cervical dislocation. The uterus was quickly
dissected, rinsed once with Tyrode’s solution 1, containing2 .1. Cell culturing, animals and tissue preparation in mM: NaCl 135.1, NaHCO 4.3, KCl 4.7, CaCl 2.6,3 2
MgCl 2.0, glucose 11 and Hepes 16.8 (pH was adjusted2
Culturing of undifferentiated mouse ES cells of the line to 7.3 with NaOH), opened, and the exposed embryos wereHM1 [13] was performed as described before[14]. Cardiac decapitated. The hearts of the embryos were dissected anddifferentiation was evoked by culturing the cells in aggre- separated into three parts: right1left atrium, apical part ofgates called embryoid bodies (EBs), using the hanging the left ventricle, and outflow tract, collected in Eppendorfdrop assay, essentially as described by Maltsev et al.[15] tubes and rinsed once with ice-cold Tyrode’s solution 1.with some small modifications. After 3 (indicated as day This solution was replaced by dissociation buffer, consist-
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409 401
ing of Tyrode’s solution 1 from which CaCl was omitted time points between 4 and 35 days of differentiation in2
and 1 mg/ml collagenase B (Roche; Mannheim, Germany) floating cultures. Independent experiments showed similarwas added, and incubated at 378C for 30 min. Every 10 patterns.min the tissue was gently triturated for 3 s until the tissuewas dissociated into single cells and small clusters. 2 .3. Immunohistochemistry
Cells and clusters were allowed overnight to adhere to3-aminopropyltriethoxysilane-coated coverslips before Mounted sections were deparaffinated and hydrated inbeing used the next day. xylene and a graded series of ethanol, and washed in PBS.
Sections were incubated in 0.5 M NH Cl with 0.25%4
2 .1.3. Tissue processing for histology Triton X-100 for 30 min to increase permeability of theFor preparation for ISH and immunohistochemistry, tissue, washed twice with PBS and blocked for 30 min
beating EBs were harvested from suspension culture, with freshly prepared bovine serum albumin (BSA) 5% inrinsed twice in PBS and fixed in 4% formaldehyde in PBS PBS at RT to reduce non-specific binding. After removalfor 30 min at RT. Groups of EBs were embedded in 1% of the BSA, a polyclonal primary antibody against Serca2asterile agarose (ultraPURE�, BRL, 5510UB) in PBS at (kind gift of F. Wuytack, Department of Physiology, K.U.50 8C and stored in 70% ethanol at 48C until further Leuven, Leuven B-3000, Belgium) was applied 1:3000 intreatment. Agarose blocks were dehydrated in a graded 0.8% BSA in PBS at RT overnight in a moist incubationethanol series and butanol, and embedded in paraplast. For chamber. The next day, sections were washed 4 times withpreparation for ISH on sections, embryos were fixed for PBS, blocked with 3% goat serum and 0.8% BSA in PBS4–16 h in freshly prepared 4% formaldehyde in phosphate- for 30 min at RT. To detect bound primary antibody,buffered saline (PBS, 150 mM NaCl, 10 mM Na-phos- sections were incubated with goat anti-rabbit antibodiesphate, pH 7.4) by rocking at 48C [17]. Embryos were conjugated to alkaline phosphatase (Dako) 1:200 in 0.8%dehydrated in a graded ethanol series and in butanol, and BSA for 2 h at RT in a moist chamber and washed 4 timesembedded in paraplast. in PBS. NBT/BCIP (Roche) diluted 1:50 in NTM-T
Serial sections of 15mm of EBs and embryos were (containing in mM: Tris-9.5 100, NaCl 100, MgCl 50,2
mounted onto microscope slides coated with 3-amino- Tween-20 0.05%) was used as a chromogenic substrate.propyltriethoxysilane. The staining reaction was ended by washing with bidistil-
led water after about 12 min. Sections were dehydrated in2 .2. Non-radioactive in situ hybridisation a series of ethanol and xylene, and embedded in Entallan.
Digoxigenin-labeled probes were made according to the 2 .4. Primer design, reverse transcription and real-timemanufacturers specifications, using Dig-UTP (Roche; polymerase chain reactionMannheim, Germany). Isoform-specific probes were usedcomplementary to the mRNA coding for cardiac troponin Complementary DNA PCR primers for mouse were(cTnI) [21], Mlc2v [11], Anf [22], Mlc2a [12], sarcoen- designed using Oligo primer analysis (version 4.1, Nationaldoplasmic reticulum calcium ATPase (Serca)2[23] and Biosciences) and Primer Express (version 1.0, PE Applieda-myosin heavy chain (aMhc) [24] (seeTable 1). Biosystems) software from DNA and RNA sequences
ISH on sections of EBs and embryos was performed as obtained from GenBank (Table 1). All primer sets had adetailed elsewhere[14,25]. Because of low RNA expres- calculated annealing temperature of 588C (nearest neigh-
1 21sion levels in ES cell-derived cardiomyocytes, the signal bour method). The sense primer for Na –Ca exchangerdetection of hybridisation on sections of EBs was en- (Ncx)1 was designed in a cardiac-specific part of thehanced using a tyramide-mediated amplification kit (NEN, transcript[18]. Primers were obtained from Biolegio (TheBoston). Expression patterns were studied in EBs derived Netherlands).from several independent differentiation experiments at First-strand complementary DNA was synthesized from
T able 1Primer characteristics used for RT-PCR
Gene Genbank Accession Forward Reverse Amplicona aNumber primer primer size
18S X00686 899–918 1049–1029 151cTnI NM 009406.1 511–527 613–596 103
]Serca2a AJ223584 3177–3197 3297–3277 121Ncx1 cardiac AF004666 2050–2072 2203–2187 154Mlc2v M91602 398–418 548–527 151Anf K02718 593–612 831–809 136
a Coordinates according to Genbank.
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

402 A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409
1 mg total RNA as described[19] by priming with a spontaneous interval. Of each recording the parameters ofmixture of 2 pmol gene-specific reverse and 125 pmol at least five APs were averaged.oligo-dT primer (Biolegio, The Netherlands) in a total In voltage-clamp, calcium current (I ) was measured14VN Ca,L
volume of 25ml. For each sample two separate cDNA in a series of 500-ms clamp steps ranging from2100 tosynthesis reactions were performed. 160 mV, increment 10 mV, interval 5 s. Holding potential
Quantitative RT-PCR was performed as described[19] (V ) was240 mV to inactivate fast sodium currents. Thehold
using the fluorescent dye SYBR green I, the Light Cycler difference between maximum inward peak and current atInstrument (Roche) and software version 3.0 (Roche). the end of the steps was taken as the amplitude ofI .Ca,L
Absolute copy numbers were estimated using standard Similarly, the amplitude of the fast sodium currentI wasNa
curves made from dilution series of amplicon sequence taken as the difference between inward current peak andsolutions of known concentration[19]. Approximate aver- steady state current recorded during steps from aV ofhold
age mRNA copy numbers per cell were calculated assum-280 mV. The change in slope during current pulses,ing a 1:1 relationship between RNA and cDNA molecules applied in current clamp was used to calculate membranemade during reverse transcription, and that each cell capacitanceC using C 5 I /dV/dt. Membrane currentsm m
contains the same total amount, 45 pg[20], of RNA. 18S were expressed as current densities by dividingI throughm
expression levels were used to correct for variations in C . Data are expressed as mean6S.E.M., ANOVA on eachm
RNA input. Results are expressed as mean6standard electrophysiological parameter was used for testing differ-deviation. ences between differentiation stages. Cluster analysis
Differences in gene expression between time points (SPSS, version 11.0.1) was used to divide ES cell data intowere tested with a one-way ANOVA. The Student–New- homogeneous subgroups (Fig. 6D).man –Keuls test was used to determine homogeneoussubsets of time points per gene. To determine equivalenceof in vitro and in vivo expression levels, per pair of in vitro 3 . Resultsand in vivo time points the squared difference of the valuesper gene was calculated and summed for all genes. The The aggregation of ES cells into EBs appears to be thecombination of time points with the least squared differ- trigger for differentiation towards a plethora of phenotypesence was considered the combination at which the gene[5], including beating cardiomyocytes. After at least 2 daysexpression patterns in vitro and in vivo correlated best. To in a floating culture, contractions were observed betrayingavoid undue influence of differences in expression level the presence of cardiomyocytes. After 4 days in a floatingbetween genes (a 10 000 times difference between Mlc2v culture up to 96% of the EBs shows vigorous beatingand cardiac troponin I (cTnI) mRNA was observed) the activity throughout the entire EB, owing to an approxi-logarithms of the values were used in this calculation. mately 30% cardiomyocyte content (unpublished data).Statistics was performed using SPSS (SPSS Inc. version11.0.1). 3 .1. Gene expression patterns
2 .5. Electrophysiology To define whether cTnI mRNA expression can be usedas a marker for all cardiomyocytes in EBs, the in vivo
Coverslips containing the attached differentiated ES pattern of expression in the mouse heart was analysed. Incells were mounted in a chamber on the stage of an Xenopus [26] and chicken[27] cTnI mRNA expressioninverted microscope (Nikon Diaphot) and superfused at a marks specifically all cardiomyocytes from early stagesrate of approximately 1 ml /min with Tyrode’s solution 2 onward. In rat, cTnI mRNA is expressed in the embryonicof 33–358C, containing in mM: NaCl 140, KCl 5.4, CaCl heart as well[21]. Ubiquitous expression was observed in2
1.8, MgCl 1.0, glucose 5.5 and Hepes 5.0 (pH set at 7.4 the chamber myocardium of the atria and ventricles, and in2
with NaOH). Action potentials (APs) and membrane the primary myocardium of the atrioventricular canal (Fig.currents were recorded using whole-cell patch clamp, 1A) and outflow tract (not shown) of a E9.5 mouse heart.digitised at 2–10 kHz and stored on the hard disk of a cTnI mRNA expression was found to be restricted to thecomputer (Apple Macintosh Quadra 650), equipped with a myocardium. In vitro, cTnI mRNA expression colocalisesdata-acquisition board (National Instruments). Recording with Serca2a, another gene expressed in all car-pipettes had resistances of 5–10 MV when filled with a diomyocytes in vivo[28], at the protein (Fig. 2C,A) andsolution containing in mM: K-gluconate 140, KCl 10, the mRNA level (Fig. 2D,F). This corroborates the as-Hepes 5.0 (pH set at 7.2 with KOH). Series resistance was sumption that cTnI mRNA expression can be used as acompensated for 60–80%. general marker for cardiomyocytes in the EB. Moreover,
From spontaneous and stimulated APs the following the absence of the skeletal muscle-specific transcriptionparameters were measured: AP amplitude (APA), maxi- factor MyoD mRNA (data not shown) demonstrates themum diastolic potential (MDP), upstroke velocity (V ), absence of skeletal muscle differentiation and thereby themax
AP duration (APD) at 20, 50 and 80% repolarisation and cardiac specificity of Serca2a expression in EBs. During
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409 403
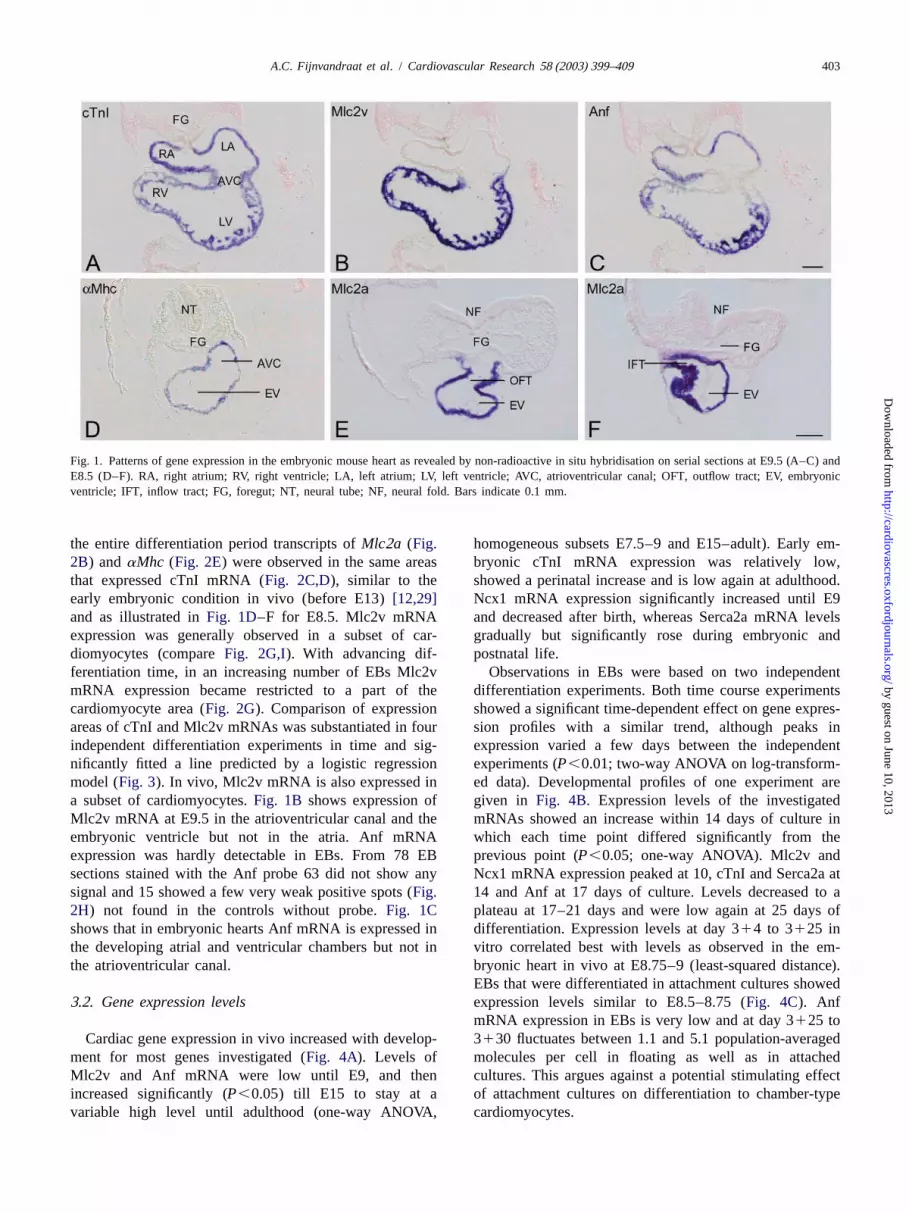
Fig. 1. Patterns of gene expression in the embryonic mouse heart as revealed by non-radioactive in situ hybridisation on serial sections at E9.5 (A–C) andE8.5 (D–F). RA, right atrium; RV, right ventricle; LA, left atrium; LV, left ventricle; AVC, atrioventricular canal; OFT, outflow tract; EV, embryonicventricle; IFT, inflow tract; FG, foregut; NT, neural tube; NF, neural fold. Bars indicate 0.1 mm.
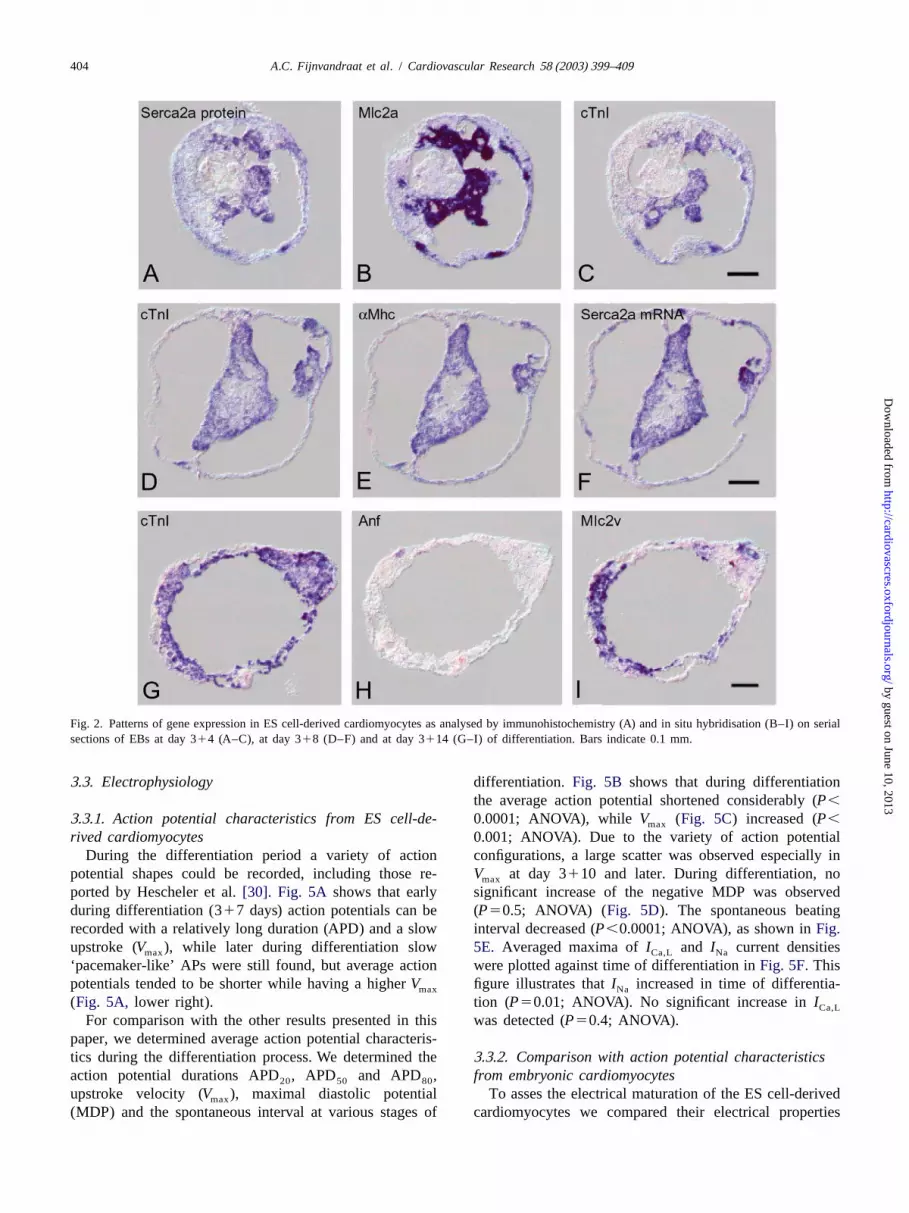
the entire differentiation period transcripts ofMlc2a (Fig. homogeneous subsets E7.5–9 and E15–adult). Early em-2B) andaMhc (Fig. 2E) were observed in the same areas bryonic cTnI mRNA expression was relatively low,that expressed cTnI mRNA (Fig. 2C,D), similar to the showed a perinatal increase and is low again at adulthood.early embryonic condition in vivo (before E13)[12,29] Ncx1 mRNA expression significantly increased until E9and as illustrated inFig. 1D–F for E8.5. Mlc2v mRNA and decreased after birth, whereas Serca2a mRNA levelsexpression was generally observed in a subset of car- gradually but significantly rose during embryonic anddiomyocytes (compareFig. 2G,I). With advancing dif- postnatal life.ferentiation time, in an increasing number of EBs Mlc2v Observations in EBs were based on two independentmRNA expression became restricted to a part of the differentiation experiments. Both time course experimentscardiomyocyte area (Fig. 2G). Comparison of expression showed a significant time-dependent effect on gene expres-areas of cTnI and Mlc2v mRNAs was substantiated in four sion profiles with a similar trend, although peaks inindependent differentiation experiments in time and sig- expression varied a few days between the independentnificantly fitted a line predicted by a logistic regression experiments (P,0.01; two-way ANOVA on log-transform-model (Fig. 3). In vivo, Mlc2v mRNA is also expressed in ed data). Developmental profiles of one experiment area subset of cardiomyocytes.Fig. 1B shows expression of given inFig. 4B. Expression levels of the investigatedMlc2v mRNA at E9.5 in the atrioventricular canal and the mRNAs showed an increase within 14 days of culture inembryonic ventricle but not in the atria. Anf mRNA which each time point differed significantly from theexpression was hardly detectable in EBs. From 78 EB previous point (P,0.05; one-way ANOVA). Mlc2v andsections stained with the Anf probe 63 did not show any Ncx1 mRNA expression peaked at 10, cTnI and Serca2a atsignal and 15 showed a few very weak positive spots (Fig. 14 and Anf at 17 days of culture. Levels decreased to a2H) not found in the controls without probe.Fig. 1C plateau at 17–21 days and were low again at 25 days ofshows that in embryonic hearts Anf mRNA is expressed in differentiation. Expression levels at day 314 to 3125 inthe developing atrial and ventricular chambers but not in vitro correlated best with levels as observed in the em-the atrioventricular canal. bryonic heart in vivo at E8.75–9 (least-squared distance).
EBs that were differentiated in attachment cultures showed3 .2. Gene expression levels expression levels similar to E8.5–8.75 (Fig. 4C). Anf
mRNA expression in EBs is very low and at day 3125 toCardiac gene expression in vivo increased with develop- 3130 fluctuates between 1.1 and 5.1 population-averaged
ment for most genes investigated (Fig. 4A). Levels of molecules per cell in floating as well as in attachedMlc2v and Anf mRNA were low until E9, and then cultures. This argues against a potential stimulating effectincreased significantly (P,0.05) till E15 to stay at a of attachment cultures on differentiation to chamber-typevariable high level until adulthood (one-way ANOVA, cardiomyocytes.
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
404 A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409
Fig. 2. Patterns of gene expression in ES cell-derived cardiomyocytes as analysed by immunohistochemistry (A) and in situ hybridisation (B–I) on serialsections of EBs at day 314 (A–C), at day 318 (D–F) and at day 3114 (G–I) of differentiation. Bars indicate 0.1 mm.
3 .3. Electrophysiology differentiation. Fig. 5B shows that during differentiationthe average action potential shortened considerably (P,
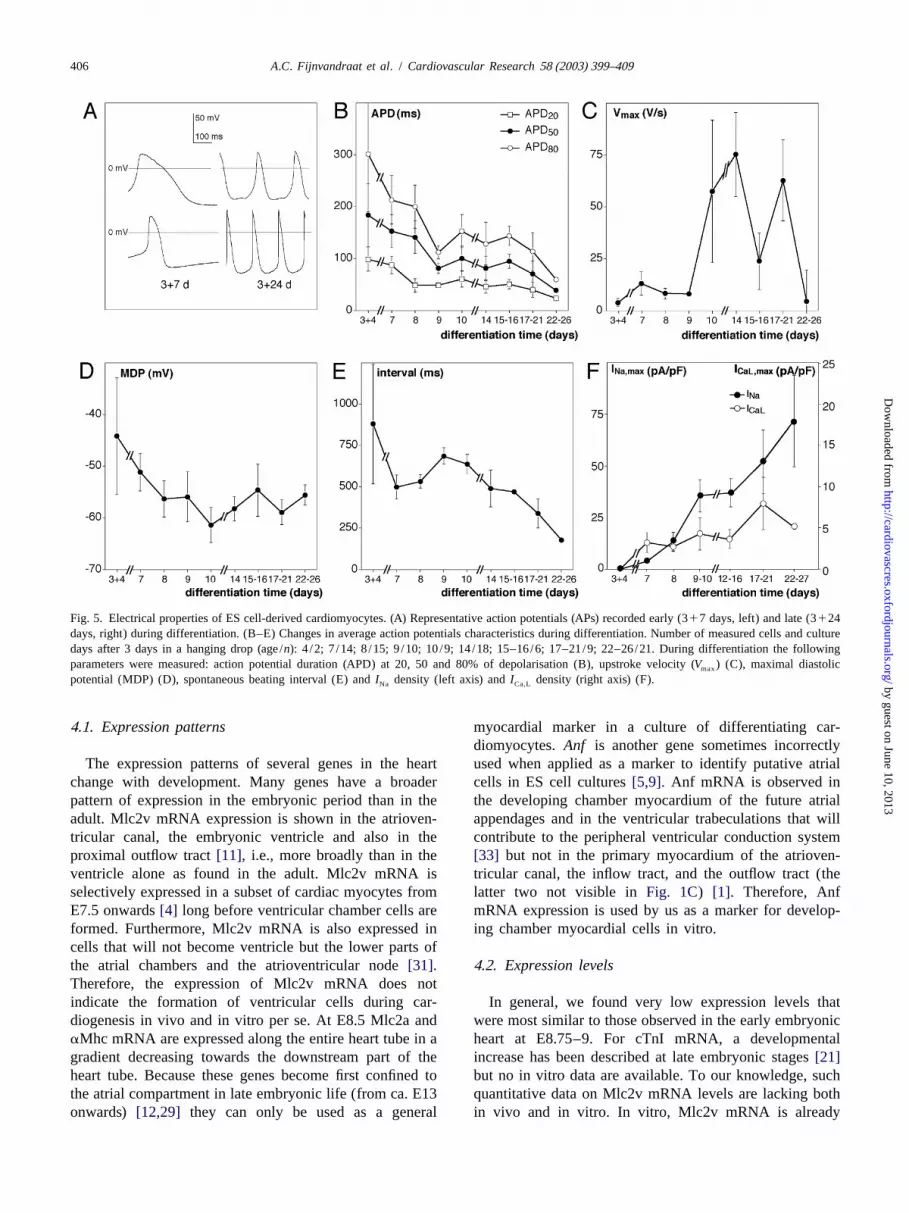
3 .3.1. Action potential characteristics from ES cell-de- 0.0001; ANOVA), while V (Fig. 5C) increased (P,max
rived cardiomyocytes 0.001; ANOVA). Due to the variety of action potentialDuring the differentiation period a variety of action configurations, a large scatter was observed especially in
potential shapes could be recorded, including those re-V at day 3110 and later. During differentiation, nomax
ported by Hescheler et al.[30]. Fig. 5A shows that early significant increase of the negative MDP was observedduring differentiation (317 days) action potentials can be (P50.5; ANOVA) (Fig. 5D). The spontaneous beatingrecorded with a relatively long duration (APD) and a slow interval decreased (P,0.0001; ANOVA), as shown inFig.upstroke (V ), while later during differentiation slow 5E. Averaged maxima ofI and I current densitiesmax Ca,L Na
‘pacemaker-like’ APs were still found, but average action were plotted against time of differentiation inFig. 5F.Thispotentials tended to be shorter while having a higherV figure illustrates thatI increased in time of differentia-max Na
(Fig. 5A, lower right). tion (P50.01; ANOVA). No significant increase inICa,L
For comparison with the other results presented in this was detected (P50.4; ANOVA).paper, we determined average action potential characteris-tics during the differentiation process. We determined the 3 .3.2. Comparison with action potential characteristicsaction potential durations APD , APD and APD , from embryonic cardiomyocytes20 50 80
upstroke velocity (V ), maximal diastolic potential To asses the electrical maturation of the ES cell-derivedmax
(MDP) and the spontaneous interval at various stages of cardiomyocytes we compared their electrical properties
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409 405
showed a considerably lowerV than that found in themax
other regions. Both outflow tract and ventricular cellsshowed a negative frequency dependence of the APD .50
We compared these three cell types with ES cell-derivedcardiomyocytes in a plot ofV versus APD, wherein themax
average data of the embryonic cardiomyocytes are plottedtogether with those from the individual ES cell-derivedcardiomyocytes (Fig. 6D). Cluster analysis indicated thatthe ES cell data formed three groups: (1) highV , shortmax
APD; (2) low V , short APD; and (3) lowV , longmax max
APD. In this panel the ES cell-data are grouped in ‘early’and ‘late’, illustrating that a combination of a lowV andmax
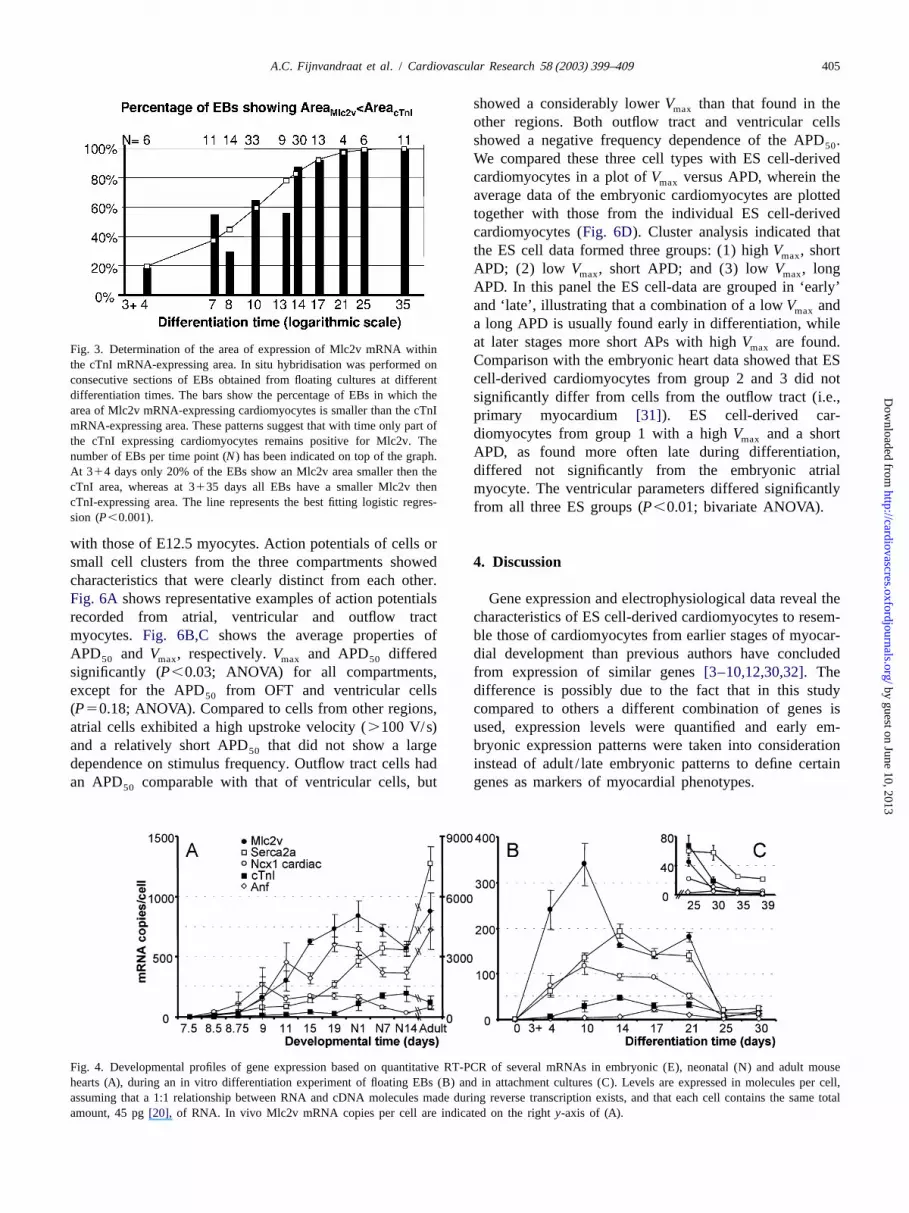
a long APD is usually found early in differentiation, whileat later stages more short APs with highV are found.maxFig. 3. Determination of the area of expression of Mlc2v mRNA withinComparison with the embryonic heart data showed that ESthe cTnI mRNA-expressing area. In situ hybridisation was performed oncell-derived cardiomyocytes from group 2 and 3 did notconsecutive sections of EBs obtained from floating cultures at different
differentiation times. The bars show the percentage of EBs in which the significantly differ from cells from the outflow tract (i.e.,area of Mlc2v mRNA-expressing cardiomyocytes is smaller than the cTnI primary myocardium [31]). ES cell-derived car-mRNA-expressing area. These patterns suggest that with time only part of
diomyocytes from group 1 with a highV and a shortmaxthe cTnI expressing cardiomyocytes remains positive for Mlc2v. TheAPD, as found more often late during differentiation,number of EBs per time point (N) has been indicated on top of the graph.differed not significantly from the embryonic atrialAt 314 days only 20% of the EBs show an Mlc2v area smaller then the
cTnI area, whereas at 3135 days all EBs have a smaller Mlc2v then myocyte. The ventricular parameters differed significantlycTnI-expressing area. The line represents the best fitting logistic regres- from all three ES groups (P,0.01; bivariate ANOVA).sion (P,0.001).
with those of E12.5 myocytes. Action potentials of cells orsmall cell clusters from the three compartments showed 4 . Discussioncharacteristics that were clearly distinct from each other.Fig. 6A shows representative examples of action potentials Gene expression and electrophysiological data reveal therecorded from atrial, ventricular and outflow tract characteristics of ES cell-derived cardiomyocytes to resem-myocytes. Fig. 6B,C shows the average properties of ble those of cardiomyocytes from earlier stages of myocar-APD and V , respectively.V and APD differed dial development than previous authors have concluded50 max max 50
significantly (P,0.03; ANOVA) for all compartments, from expression of similar genes[3–10,12,30,32].Theexcept for the APD from OFT and ventricular cells difference is possibly due to the fact that in this study50
(P50.18; ANOVA). Compared to cells from other regions, compared to others a different combination of genes isatrial cells exhibited a high upstroke velocity (.100 V/s) used, expression levels were quantified and early em-and a relatively short APD that did not show a large bryonic expression patterns were taken into consideration50
dependence on stimulus frequency. Outflow tract cells had instead of adult / late embryonic patterns to define certainan APD comparable with that of ventricular cells, but genes as markers of myocardial phenotypes.50
Fig. 4. Developmental profiles of gene expression based on quantitative RT-PCR of several mRNAs in embryonic (E), neonatal (N) and adult mousehearts (A), during an in vitro differentiation experiment of floating EBs (B) and in attachment cultures (C). Levels are expressed in molecules per cell,assuming that a 1:1 relationship between RNA and cDNA molecules made during reverse transcription exists, and that each cell contains the same totalamount, 45 pg[20], of RNA. In vivo Mlc2v mRNA copies per cell are indicated on the righty-axis of (A).
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
406 A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409
Fig. 5. Electrical properties of ES cell-derived cardiomyocytes. (A) Representative action potentials (APs) recorded early (317 days, left) and late (3124days, right) during differentiation. (B–E) Changes in average action potentials characteristics during differentiation. Number of measured cells and culturedays after 3 days in a hanging drop (age/n): 4 /2; 7 /14; 8/15; 9/10; 10/9; 14/18; 15–16/6; 17–21/9; 22–26/21. During differentiation the followingparameters were measured: action potential duration (APD) at 20, 50 and 80% of depolarisation (B), upstroke velocity (V ) (C), maximal diastolicmax
potential (MDP) (D), spontaneous beating interval (E) andI density (left axis) andI density (right axis) (F).Na Ca,L
4 .1. Expression patterns myocardial marker in a culture of differentiating car-diomyocytes.Anf is another gene sometimes incorrectly
The expression patterns of several genes in the heart used when applied as a marker to identify putative atrialchange with development. Many genes have a broader cells in ES cell cultures[5,9]. Anf mRNA is observed inpattern of expression in the embryonic period than in the the developing chamber myocardium of the future atrialadult. Mlc2v mRNA expression is shown in the atrioven- appendages and in the ventricular trabeculations that willtricular canal, the embryonic ventricle and also in the contribute to the peripheral ventricular conduction systemproximal outflow tract[11], i.e., more broadly than in the [33] but not in the primary myocardium of the atrioven-ventricle alone as found in the adult. Mlc2v mRNA is tricular canal, the inflow tract, and the outflow tract (theselectively expressed in a subset of cardiac myocytes from latter two not visible inFig. 1C) [1]. Therefore, AnfE7.5 onwards[4] long before ventricular chamber cells are mRNA expression is used by us as a marker for develop-formed. Furthermore, Mlc2v mRNA is also expressed in ing chamber myocardial cells in vitro.cells that will not become ventricle but the lower parts ofthe atrial chambers and the atrioventricular node[31]. 4 .2. Expression levelsTherefore, the expression of Mlc2v mRNA does notindicate the formation of ventricular cells during car- In general, we found very low expression levels thatdiogenesis in vivo and in vitro per se. At E8.5 Mlc2a and were most similar to those observed in the early embryonicaMhc mRNA are expressed along the entire heart tube in a heart at E8.75–9. For cTnI mRNA, a developmentalgradient decreasing towards the downstream part of the increase has been described at late embryonic stages[21]heart tube. Because these genes become first confined to but no in vitro data are available. To our knowledge, suchthe atrial compartment in late embryonic life (from ca. E13 quantitative data on Mlc2v mRNA levels are lacking bothonwards) [12,29] they can only be used as a general in vivo and in vitro. In vitro, Mlc2v mRNA is already
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
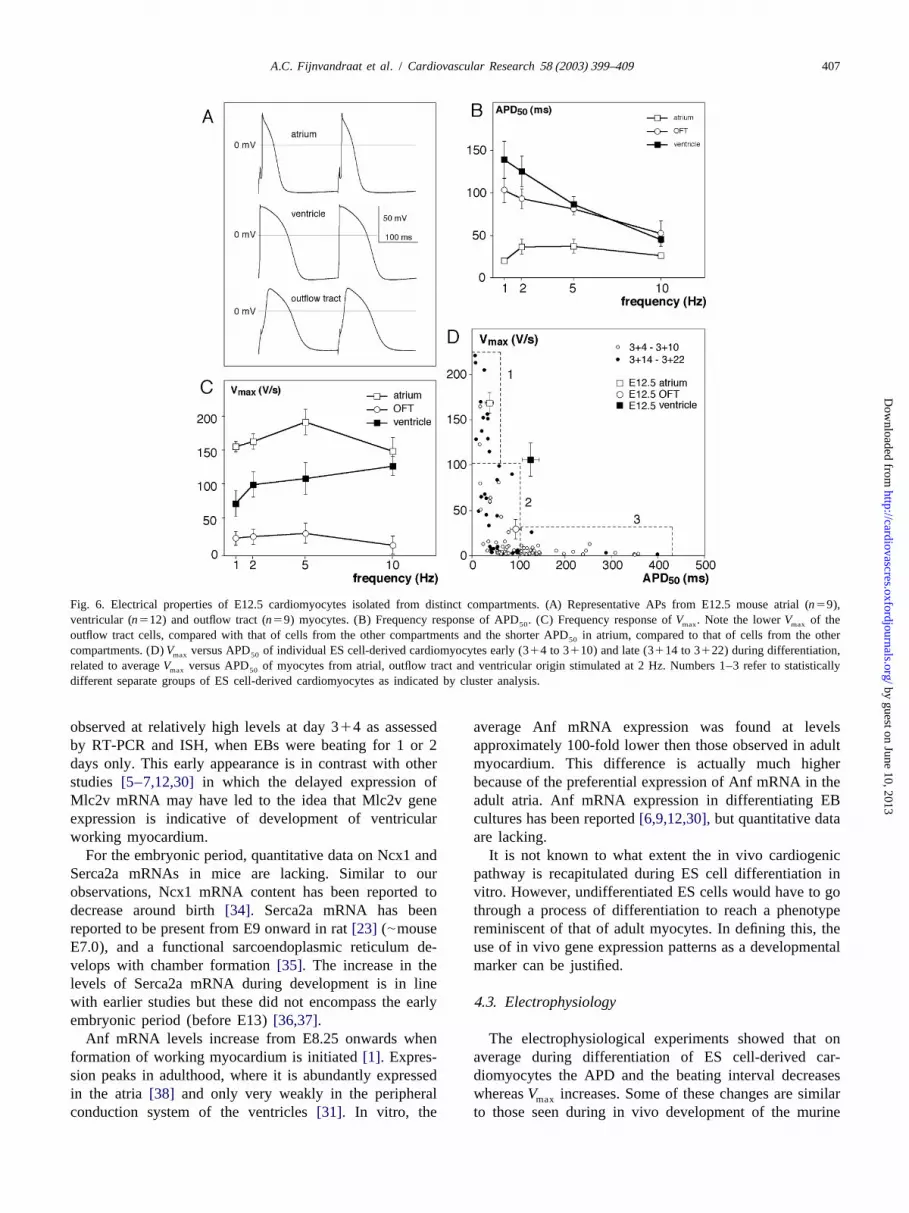
A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409 407
Fig. 6. Electrical properties of E12.5 cardiomyocytes isolated from distinct compartments. (A) Representative APs from E12.5 mouse atrial (n59),ventricular (n512) and outflow tract (n59) myocytes. (B) Frequency response of APD . (C) Frequency response ofV . Note the lowerV of the50 max max
outflow tract cells, compared with that of cells from the other compartments and the shorter APD in atrium, compared to that of cells from the other50
compartments. (D)V versus APD of individual ES cell-derived cardiomyocytes early (314 to 3110) and late (3114 to 3122) during differentiation,max 50
related to averageV versus APD of myocytes from atrial, outflow tract and ventricular origin stimulated at 2 Hz. Numbers 1–3 refer to statisticallymax 50
different separate groups of ES cell-derived cardiomyocytes as indicated by cluster analysis.
observed at relatively high levels at day 314 as assessed average Anf mRNA expression was found at levelsby RT-PCR and ISH, when EBs were beating for 1 or 2 approximately 100-fold lower then those observed in adultdays only. This early appearance is in contrast with other myocardium. This difference is actually much higherstudies [5–7,12,30] in which the delayed expression of because of the preferential expression of Anf mRNA in theMlc2v mRNA may have led to the idea that Mlc2v gene adult atria. Anf mRNA expression in differentiating EBexpression is indicative of development of ventricular cultures has been reported[6,9,12,30],but quantitative dataworking myocardium. are lacking.
For the embryonic period, quantitative data on Ncx1 and It is not known to what extent the in vivo cardiogenicSerca2a mRNAs in mice are lacking. Similar to our pathway is recapitulated during ES cell differentiation inobservations, Ncx1 mRNA content has been reported to vitro. However, undifferentiated ES cells would have to godecrease around birth[34]. Serca2a mRNA has been through a process of differentiation to reach a phenotypereported to be present from E9 onward in rat[23] (|mouse reminiscent of that of adult myocytes. In defining this, theE7.0), and a functional sarcoendoplasmic reticulum de- use of in vivo gene expression patterns as a developmentalvelops with chamber formation[35]. The increase in the marker can be justified.levels of Serca2a mRNA during development is in linewith earlier studies but these did not encompass the early4 .3. Electrophysiologyembryonic period (before E13)[36,37].
Anf mRNA levels increase from E8.25 onwards when The electrophysiological experiments showed that onformation of working myocardium is initiated[1]. Expres- average during differentiation of ES cell-derived car-sion peaks in adulthood, where it is abundantly expressed diomyocytes the APD and the beating interval decreasesin the atria [38] and only very weakly in the peripheral whereasV increases. Some of these changes are similarmax
conduction system of the ventricles[31]. In vitro, the to those seen during in vivo development of the murine
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

408 A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409
ventricle. Initially at E9.5, ventricular cells have a rela- started. However, the propensity of increasing upstroketively low V and an APD of 100–200 ms. At E18, velocity of action potentials at later differentiation timesmax 50
V increases and the APD shortens to 74 ms[39,40] and could indicate that a first step towards more maturemax
at three neonatal days, APD shortens even further to 18 working myocardium has been made. It will be challeng-50
ms [41]. Other changes are different from the in vivo ing to induce further differentiation of chamber myocar-observations. At E9.5 ventricular cells beat spontaneously dium in vitro and unravel the underlying process using thiswith a regular interval of about 1 s, whereas from E18 culture system.onward the cells become silent at a negative restingpotential[40]. We showed that the average beating intervalof ES cell-derived cardiomyocytes decreased in time. This A cknowledgementssuggests that most ES cell-derived cardiomyocytes do notdevelop into a ventricular phenotype, but towards a nodal We express our gratitude to C.C. Verhoek-Pocock, C. dephenotype, which mostly resembles primary myocardium Gier-de Vries and M.A.W. Markman for expert technical[31]. assistance and Dr. P.E.M.H. Habets for providing the
Our observation thatI increases during in vitro embryos. Probes were kindly provided by Drs C.E. Seid-Na
differentiation agrees well with the findings of Maltsev et man (Anf), K.R. Chien (Mlc2v, MLC2a), S. Schiaffinoal. [15] and indicates that this increase is involved in the (cTnI) and K.R. Boheler (aMhc). This study was supportedincrease inV as observed during differentiation. by Netherlands Heart Foundation, grants 99.170 (ACF),max
and M96.002 (to AFMM, RHLD and VMC).
5 . Conclusion
R eferencesES cell-derived cardiomyocytes are not differentiated
towards fully mature chamber myocardium as yet. They [1] C hristoffels VM, Habets PEMH, Franco D, Campione M, de Jong F,rather show a phenotype comparable to young embryonic Lamers WH, Bao ZZ, Palmer S, Biben C, Harvey RP, Moorman
AFM. Chamber formation and morphogenesis in the developingcardiomyocytes in vivo. Several arguments support thismammalian heart. Dev Biol 2000;223:266–278.interpretation. Firstly, expression levels of genes in dif-
[2] B oheler KR, Czyz J, Tweedie D, Yang HT, Anisimov SV, Wobusferentiating ES cell cultures are similar to levels observedAM. Differentiation of pluripotent embryonic stem cells into
in the primary heart tube stage at E8.75–9. Secondly, cardiomyocytes. Circ Res 2002;91:189–201.markers that are chamber-specific in the late embryonic [3] K lug MG, Soonpaa MH, Koh GY, Field LJ. Genetically selected
cardiomyocytes from differentiating embryonic stem cells formand adult heart (aMhc andMlc2a) [12,29],and the generalstable intracardiac grafts. J Clin Invest 1996;98:216–224.cardiac markerscTnI andSerca2a [21,28] showed expres-
[4] L yons I, Parsons LM, Hartley L, Li R, Andrews JE, Robb L, Harveysion in the entire myocardial area of the EB. TheseRP. Myogenic and morphogenetic defects in the heart tubes of
observations indicate that at the level of regulation of murine embryos lacking the homeo box gene Nkx2-5. Gene DevMlc2a and aMhc the distinction between a primary or a 1995;9:1654–1666.chamber phenotype has not been made. Thirdly,Anf, the [5] G uan K, Rohwedel J, Wobus AM. Embryonic stem cells differentia-
tion models: cardiogenesis, myogenesis, neurogenesis, epithelial andonly currently known gene that is exclusively expressed invascular smooth muscle cell differentiation in vitro. Cytotechnologythe forming atrial and ventricular chambers[1], is ex-1999;30:211–226.
pressed at very low levels in differentiating EBs, indicating [6] M iller-Hance WC, LaCorbiere M, Fuller SJ, Evans SM, Lyons G,that hardly any chamber myocardium has developed. Schmidt C, Robbins J, Chien KR. In vitro chamber specificationFourthly, electrophysiological properties of early ES cell- during embryonic stem cell cardiogenesis. Expression of the ven-
tricular myosin light chain-2 gene is independent of heart tubederived cardiomyocytes were most reminiscent of cellsformation. J Biol Chem 1993;268:25244–25252.from the outflow tract and further differentiation resulted
[7] D oevendans PA, Kubalak SW, An RH, Becker KD, Chien KR, Kassin cells comparable to those of the embryonic atrium. RS. Differentiation of cardiomyocytes in floating embryoid bodies isHowever, it has to be assessed whether the atrial electro- comparable to fetal cardiomyocytes. Mol Cell Cardiol 2000;32:839–physiological characteristics are also represented in the 851.
¨[8] M uller M, Fleischmann BK, Selbert S, Ji GJ, Endl E, Middeler G,embryonic heart tube and as such would represent normal¨Muller OJ, Schlenke P, Frese S, Wobus AM, Hescheler J, Katus HA,variation of cardiomyocytes in this stage of heart develop-
Franz WM. Selection of ventricular-like cardiomyocytes from ESment. Finally, all cardiomyocytes analysed by patch clamp cells in vitro. FASEB J 2001;14:2540–2548.showed automaticity, a characteristic for all embryonic ¨[9] F assler R, Rohwedel J, Maltsev V, Bloch W, Lentini S, Guan K,cardiomyocytes and for nodal cells in the mature heart. Gullberg D, Hescheler J, Addicks K, Wobus AM. Differentiation and
integrity of cardiac muscle cells are impaired in the absence of beta1Taken together, the levels of expression of several keyintegrin. J Cell Sci 1996;109:2989–2999.cardiac genes and electrophysiological features strongly
[10] M eyer N, Jaconi M, Landopoulou A, Fort P, Puceat M. A fluores-suggest that the phenotypes of cardiomyocytes in differen- cent reporter gene as a marker for ventricular specification intiating EBs are most reminiscent of cardiomyocytes from ES-derived cardiac cells. FEBS Lett 2000;478:151–158.E8.75–9 mouse hearts where chamber formation has just[11] O ’Brien TX, Lee KJ, Chien KR. Positional specification of ventricu-
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

A.C. Fijnvandraat et al. / Cardiovascular Research 58 (2003) 399–409 409
lar myosin light chain 2 expression in the primitive murine heart [26] D rysdale TA, Tonissen KF, Patterson KD, Crawford MJ, Krieg PA.tube. Proc Natl Acad Sci USA 1993;90:5157–5161. Cardiac troponin I is a heart-specific marker in the Xenopus embryo
[12] K ubalak SW, Miller-Hance WC, O’Brien TX, Dyson E, Chien KR. expression during abnormal heart morphogenesis. Dev BiolChamber specification of atrial myosin light chain-2 expression 1994;165:432–441.precedes septation during murine cardiogenesis. J Biol Chem [27] H ouweling AC, Somi S, van den Hoff MJ, Moorman AFM,1994;269:16961–16970. Christoffels VM. The developmental pattern of ANF gene expres-
[13] M agin TM, McWhir J, Melton DW. A new mouse embryonic stem sion reveals a strict localization of cardiac chamber formation incell line with good germ line contribution and gene targeting chicken. Anat Rec 2002;266:93–102.frequency. Nucleic Acids Res 1992;20:3795–3796. ´[28] A nger M, Samuel JL, Marotte F, Wuytack F, Rappaport L, Lompre
[14] F ijnvandraat AC, de Boer PAJ, Lekanne Deprez RH, Moorman AM. In situ mRNA distribution of sarco(endo)plasmic reticulum21AFM. Non-radioactive in situ detection of mRNA in ES cell-derived Ca -ATPase isoforms during ontogeny in the rat. J Mol Cell
cardiomyocytes and in the developing heart. Microsc Res Techn Cardiol 1994;26:539–550.2002;58:387–394.
[29] L yons GE, Schiaffino S, Sassoon D, Barton P, Buckingham ME.[15] M altsev VA, Wobus AM, Rohwedel J, Bader M, Hescheler J.
Developmental regulation of myosin expression in mouse cardiacCardiomyocytes differentiated in vitro from embryonic stem cells
muscle. J Cell Biol 1990;111:2427–2437.developmentally express cardiac-specific genes and ionic currents.
[30] H escheler J, Fleischmann BK, Lentini S, Maltsev VA, Rohwedel J,Circ Res 1994;75:233–244.Wobus AM, Addicks K. Embryonic stem cells: a model to study[16] M etzger JM, Lin WI, Samuelson LC. Vital staining of cardiacstructural and functional properties in cardiomyogenesis. Cardiovascmyocytes during embryonic stem cell cardiogenesis in vitro. CircRes 1997;36:149–162.Res 1996;78:547–552.
[31] M oorman AFM, Christoffels VM. Cardiac chamber formation:[17] M oorman AFM, de Boer PAJ, Ruijter JM, Hagoort J, Franco D,development, genes and evolution. Physiol Rev 2003;in press.Lamers WH. Radio-isotopic in situ hybridization on tissue sections:
[32] W obus AM, Guan KM, Jin S, Wellner MC, Rohwedel J, Ji GJ,practical aspects and quantification. In: Tuan RS, Lo CW, editors,Fleischmann B, Katus HA, Hescheler T, Franz WM. Retinoic acidDevelopmental Biology Protocols, vol. III, Methods in Molecularaccelerates embryonic stem cell-derived cardiac differentiation andBiology, vol. 137, Totowa, NJ: Humana Press, 2000, pp. 97–115,enhances development of ventricular cardiomyocytes. J Mol Cellchapter 11.Cardiol 1997;29:1525–1539.[18] K ofuji P, Lederer WJ, Schulze DH. Mutually exclusive and cassette
exons underlie alternatively spliced isoforms of the Na/Ca ex- [33] M oorman AFM, de Jong F, Denyn MMFJ, Lamers WH. Develop-changer. J Biol Chem 1994;269:5145–5149. ment of the cardiac conduction system. Circ Res 1998;82:629–644.
[19] L ekanne Deprez RH, Fijnvandraat AC, Ruijter JM, Moorman AFM. [34] K oban MU, Moorman AFM, Holtz J, Yacoub MH, Boheler KR.Sensitivity and accuracy of quantitative real-time polymerase chain Expressional Analysis of the cardiac Na-Ca exchanger in ratreaction using SYBR green I depends on cDNA synthesis con- development and senescence. Cardiovasc Res 1998;37:405–423.ditions. Anal Biochem 2002;307:63–69. [35] N akanishi H, Okuda K, Kamati K, Abe K, Sekiguchi M, Takao A.
[20] A lberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD. Table Development of myocardial contractile system in the fetal rabbit.3-1, Approximate chemical compositions of a typical bacterium and Pediatr Res 1987;22:201–207.a typical mammalian cell. In: Alberts B, Bray D, Lewis J, Raff M, [36] R ibadeau-Dumas A, Boateng S, Schwartz K, Boheler KR. Sarco(en-
21Roberts K, Watson JD, editors, Molecular Biology of The Cell, 3rd do)plasmic reticulum Ca -ATPase (SERCA2) gene products areedition, New York: Garland Publishing,Inc, 1994, p. 90. regulated posttransciptionally during rat cardiac development. Car-
[21] A usoni S, de Nardi C, Moretti P, Gorza L, Schiaffino S. De- diovasc Res 1999;43:426–436.velopmental expression of rat cardiac troponin I mRNA. Develop-
´[37] L ompre AM, Lambert F, Lakatta EG, Schwarz K. Expression ofment 1991;112:1041–1051. 21sarcoplasmic reticulum Ca -ATPase and calsequestrin genes in rat
[22] S eidman CE, Duby AD, Choi E, Graham RM, Haber E, Homcy C,heart during ontogenic development and aging. Circ Res
Smith JA, Seidman JG. The structure of rat preproatrial natriuretic1991;69:1380–1388.
factor as defined by a complementary DNA clone. Science[38] Z eller R, Bloch KD, Williams BS, Arceci RJ, Seidman CE.1984;225:324–326.
Localized expression of the atrial natriuretic factor gene during[23] M oorman AFM, Vermeulen JLM, Koban MU, Schwartz K, Lamerscardiac embryogenesis. Gene Dev 1987;1:693–698.WH, Boheler KR. Patterns of expression of sarcoplasmic reticulum
21 [39] L iu W, Yasui K, Arai A, Kamiya K, Cheng J, Kodama I, Toyama J.Ca ATPase and phospholamban mRNAs during rat heart develop-21beta-adrenergic modulation of L-type Ca -channel currents inment. Circ Res 1995;76:616–625.
early-stage embryonic mouse heart. Am J Physiol 1999;276:H608–[24] B oheler KR, Chassagne C, Martin X, Wisnewsky C, Schwartz K.H613.Cardiac expressions of alpha- and beta-myosin heavy chains and
[40] Y asui K, Liu W, Opthof T, Kada K, Lee JK, Kamiya K, Kodama I.sarcomeric alpha-actins are regulated through transcriptional mecha-I(f) current and spontaneous activity in mouse embryonic ventricularnisms. J Biol Chem 1992;267:12979–12985.myocytes. Circ Res 2001;88:536–542.[25] M oorman AFM, Houweling AC, de Boer PAJ, Christoffels VM.
Sensitive nonradioactive detection of mRNA in tissue sections: [41] W ang L, Feng ZP, Kondo CS, Sheldon RS, Duff HJ. Developmentalnovel application of the whole-mount in situ hybridization protocol. changes in the delayed rectifier K1 channels in mouse heart. CircJ Histochem Cytochem 2001;49:1–8. Res 1996;79:79–85.
by guest on June 10, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from





























