Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
Contents lists available at SciVerse ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Application of non-grass phytoliths in reconstructing deltaic environments: A studyfrom the Indian Sunderbans
Sayantani Das a, Ruby Ghosh b, Subir Bera a,⁎a Centre of Advanced study, Palaeobotany-Palynology Laboratory, Department of Botany, University of Calcutta, 35, Ballygunge Circular Road, Kolkata-700019, Indiab Birbal Sahni Institute of Palaeobotany, Lucknow-226007, U.P., India
⁎ Corresponding author at: Centre of Advanced study, Dof Calcutta, Palaeobotany-Palynology Laboratory, 35, Ball700019, India. Tel.: +91 33 2461 4959; fax: +91 33 2461
E-mail address: [email protected] (S. Bera).
0031-0182/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.palaeo.2013.02.017
a b s t r a c t
a r t i c l e i n f oArticle history:Received 1 August 2012Received in revised form 8 February 2013Accepted 18 February 2013Available online 28 February 2013
Keywords:Deltaic sub-environmentsNon-grass phytolithsIndian SunderbansTaxonomic implicationEnvironment reconstructionQuantitative methods
Phytoliths have been proved as reliable indicators of an ecosystem's structure in both modern days as well asin the past. Quantitative phytolith studies from modern plants in India are negligible and most of the datacomes from grasses excepting few pteridophytes. Data on non-grass angiospermous phytoliths in India ishowever completely lacking. In order to interpret the fossil phytolith assemblages and their depositionalsub-environments from the deltaic Sunderbans, the largest mangrove vegetation of the world, modern ana-logue is a prerequisite. The objectives of this study are to establish a relationship between modern deltaicnon-grass plant communities of the Indian part of the Sunderbans along salinity gradient with their predom-inant phytolith contents and this study is the first step in identifying the environment associated with man-grove development from phytoliths in ancient sediments and further studies would ascertain which ofthese phytoliths would survive erosion and burial. Modern phytolith assemblages were studied from differentparts of 33 non-grass species representing 27 genera under 23 families collected from different deltaicsub-environments of the study area. Retrieved leaf phytolith assemblages were subjected to Principal Compo-nents Analysis and Cluster Analysis. Elemental analysis of some of these morphotypes done using an EnergyDispersive X-ray Spectrometer (EDX) to authenticate chemical composition of some newly described formsof phytoliths. Twenty five phytolith morphotypes identified under light and Scanning Electron microscopesrepresent a modern reference non-grass phytolith database of deltaic plant communities of the Sunderbans.Extracted morphotypes are described and their taxonomic and environmental implications are interpretedconsidering quantitative data and statistical analyses. The taxonomic implications of these morphotypes aremostly poor as observed through both Principal Components Analysis and Cluster Analysis except for mono-cotyledonous and few dicotyledonous families which produce distinctivemorphotypes. However, their role inenvironmental interpretations could be established using this modern phytolith data. The data would providea basis for identification of deltaic vegetation of an area using non-grass phytoliths.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Phytoliths are hydrated silica (SiO2, nH2O) particles of plant origindeposited in the tissues of many vascular plants. They are precipitatedas opal bodies in plant cells or in the intercellular spaces of plants andthese deposits can be cast from the cells, from the infilling of the celllumen or by the silicification of the cell walls (their shape resemblingthe outline of the original cell). Since phytoliths are resistant to decayand destruction in highly oxidative environment, they have an advan-tage over pollen grains during palaeoenvironmental reconstruction ofan area. In many species these non-crystalline, bio-mineralized silicaprecipitates are a major mineral plant constituent (Epstein, 1994)
epartment of Botany, Universityygunge Circular Road, Kolkata-4849.
rights reserved.
containing trace elements like Al, Fe, Cu, N, P, etc. Depending upon theextent of carbon coating, phytoliths may be clear, brown-pigmentedor even totally opaque (Prychid et al., 2004).
Phytolith morphologies observed in different geographical loca-tions reveal a general continuity of patterns within family, therebysuggesting a high level of genetic control over phytolith production.However differences in results seem to indicate that environmentalfactors must also be considered which play an important role in de-termining the nature of phytolith production (Wallis, 2003). Phyto-lith production pattern also depends on a number of factors like theenvironment of growth of the plant, physical and chemical nature ofthe soil, amount of water present in the soil, age of the plant andthe taxonomic affinity of the plant taxa, hence production patternvaries among different plant taxa belonging to different taxonomicgroups and also in different parts of a plant.
To interpret soil-borne phytolith assemblages a documentedinventory relevant for the study area is a prerequisite (Piperno, 1988;Bozarth, 1993; Lu and Liu, 2003; Mercader et al., 2009, 2010). However,

49S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
in India only few studies have been conducted onmodern plants (espe-cially from grasses) to explore their potential in palaeoenvironmentalstudies. Krishnan (2000) studied the morphology of phytoliths ofabout 100 species of modern grasses from the plains of Tamil Nadu,India, and discussed their value in identifying extinct and extant grassesat different taxonomic levels and in exploring past vegetation and cli-mate. Later Chauhan et al. (2011) studied the diversity, distributionand frequency based attributes of phytoliths of Arundo donax L. Somestudies on occurrence of opaline silica in pteridophytes were alsoconducted by Majumdar and Mukhopadhyay (2009a,b, 2011).Tripathi et al. (2012) had investigated the general pattern of phytolithdeposition and distribution in 2 species of Sorghum i.e. S. bicolorand S. halepense which would further help in tracing the history ofcultivation of Sorghum in Allahabad, UP, India because only thesetwo species grow in this region. The only study of phytoliths fromdeltaic Sunderbans was done by Ghosh et al. (2011) to check reliabilityof grass phytoliths in reconstructing deltaic environments. Virtually nodata of phytoliths from non-grass species of these unique deltaic re-gions of the Sunderbans are available however reliability of non-grassphytoliths in environmental reconstruction is well established (Wallis,2003; Mercader et al., 2010).
Fig. 1. The inset depicts the geographical map of West Bengal with the study area in the mwhere plant collection was made.
The ‘Sundarbans’ is the largest mangrove forest in the world aswell as one of the World Heritage Sites, where about 170 species oftrees, shrubs and herbs are grown in the Indian part (Fig. 1).
Input of both fresh water from the river Ganges and saline waterfrom Bay of Bengal results into such a unique dense mangrove forestwith variety of ecological habitats i.e. swampy mangrove or intertidalmangrove zone (river flats and river-slopes below the high tide leveland submerged by tidal water twice a day), the tidal mangrove zone(covered by spring tide and also during storms), the truemangrove de-cline zone (absence of tidal water only), the zone for colonization ofnon-littoral species (where the salinity of the soil is reduced by floodingrain and the soil condition supports themesophytic vegetation) and xe-rophytic non-mangrove-dry evergreen forest zones (Naskar and GuhaBakshi, 1987).
We present here a reference collection of a selected range of non-grass plant communities native to the deltaic environment of theSunderbans. More precisely, we attempt to determine the taxonomicsignificance of phytoliths produced by the indigenous trees, shrubsand herbs of this part of Ganga-Bhramputra delta and hypothesize thephytolithic signal that may be left behind by these plants in the geolog-ical record.
arked zone. The main map shows the study sites in the Indian part of the Sundarbans

50 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
This paper also gives insights to whether difference in ecologicalcondition influences the phytolith production of plants from differenteco-climatic zones along the salinity gradient in the deltaic plains ofthe Sunderbans, India. We used CA (Cluster Analysis) to check whetherthe taxonomic groups can be distinguished on the basis of phytolithtypes within the non-grass plant communities of the Sunderbans. PCA(Principal Component Analysis) was also attempted on the modernphytolith data. This can help to identify the characteristic phytolithmorphotypes that can be used in discriminating different deltaicsub-environments. This data may further be used in the interpretationof fossil phytolith assemblages frompast deltaic environments. Becausethe physiological role of silicon in vascular plants is still in infancy andecological conditions may strongly influence the degree of silicificationof plant individuals and populations, we used a large data set. Further,elemental analysis using Energy Dispersive X-ray Spectrometer (EDX)was done for some of the phytolith morphotypes derived from pneu-matophores to authenticate their biogenic origin.
2. Materials and methods
2.1. Collection of living plant specimens
Plant specimens for thepresent studywere collected fromLothian Is-land, Bhagabatpur, Sudhanyakhali, Sajnekhali, Dutta Lahiri, Pakhiralayaand Kakdwip (between lat. 21° 31/ N–22° 30/ N and long. 88°10/ E–89° 51/ E) located in the Indian part of the Sunderbans (Fig. 1).The ‘Sunderbans’ is situated in the estuarine mouths of deltaic plainsof Ganga–Brahmaputra–Meghna in both India and Bangladesh.Thedense mangrove forest of this exclusive ecosystem is world re-nowned and extends between long. 88° 10/–92° 15/ E and latitude 21°00/–22° 31/ N and estimated to cover about 1.48–1.73 million acres.
Table 1Plants used for phytolith extraction from the Indian part of the Sunderbans with their habi
Taxon Order Family Habit
Acanthus ilicifolius L. Lamiales Acanthaceae Shrub/tall hAcrostichum aureum L. Polypodiales Pteridaceae Herb
Aegialitis rotundifolia Roxb. Caryophyllales Plumbaginaceae ShrubAegiceras corniculatum L. Ericales Myrsinaceae Shrub / treeAmoora cucullata Roxb. Sapindales Meliaceae TreeAvicennia alba Bl. Lamiales Acanthaceae ShrubBruguiera gymnorrhiza L. Malpighiales Rhizophoraceae TreeBruguiera cylindrica Bl. Malpighiales Rhizophoraceae TreeCeriops tagal Perr. Malpighiales Rhizophoraceae TreeClerodendrum inerme L. Lamiales Verbenaceae ShrubDerris scandens Benth. Fabales Fabaceae Woody climExcoecaria agallocha L. Malpighiales Euphorbiaceae Tree
Grangea maderaspatana (Pers.) K.Schum. Asterales Asteraceae HerbHeliotropium curassavicum L. unplaced Boraginaceae HerbHeritiera fomes Buch. Ham. Malvales Sterculiaceae TreeKandelia candel L. Malpighiales Rhizophoraceae TreeLumnitzera racemosa Willd. Myrtales Combreatceae TreeNypa fruticans Wurmb. Arecales Arecaceae TreePandanus fascicularis Lamk. Pandanales Pandanaceae ShrubPedilanthus tithymaloides L. Malpighiales Euphorbiaceae ShrubPhoenix paludosa Roxb. Arecales Arecaceae TreeRhizophora apiculata Bl. Malpighiales Rhizophoraceae TreeSarcolobus globosus Wall. Gentianales Asclepiadaceae ShrubScirpus articulatus L. Poales Cyperaceae HerbSonneratia apetala Ham. Myrtales Lythraceae TreeSonneratia caseolaris L. Myrtales Lythraceae TreeSuaeda nudiflora Moq. Caryophyllales Chenopodiaceae HerbTamarix gallica L. Caryophyllales Tamaricaceae Shrub/smalThespesia populnea (L.)Sol.ex.Corr. Malvales Malvaceae Small treeThevetia peruviana (Pers.) K.Schum. Gentianales Apocynaceae Shrub/smalVitis trifolia L. Vitales Vitaceae ClimberXylocarpus granatum Koenig. Sapindales Meliaceae TreeXylocarpus moluccensis (L.) Roem. Sapindales Meliaceae Tree
Mangrove plants thrive well in muddy, wet soil andmarshy areas intropical tidal waters. The mangrove vegetation of the Sunderbans canbe categorized into shrubs, trees, ground ferns or palms depending onthe ratio of salt water and fresh water that the roots intake. Theseman-grove trees can grow well in a combination of salt water and freshwater. Mostly chloride and sulphate of sodium, magnesium, calciumand traces of bicarbonate salts are present in the soil. Plant taxa growingpredominantly in different deltaic sub-environments (Naskar and GuhaBakshi, 1987) were collected from all the sites for phytolith extraction.
2.2. Extraction of phytoliths
33 plant taxa belonging to 27 genera and 33 species under 23 fam-ilies of the non-grass mangrove plant communities collected from dif-ferent deltaic sub-environments from the Indian Sundarbans wereused for phytolith study (Table 1). The analyzed plant parts (mostlyleaf, fruit, seed, flower, spathe and pneumatophore) were selectedrandomly from the selected area on the basis of their availability.
Collected fresh plant tissues were placed in a beaker followed bywashing with distilled water in an ultrasonic water bath to removethe superficial debris adhering to the surface of the tissues. In caseof pneumatophore, its outer layer was peeled and washed followingthe same procedure as before. The fresh plant tissues were dried ina hot air oven overnight at 80 °C. The dried tissues were then oxi-dized with Schulze's solution (conc. HNO3 + NaClO3 in 3:1 ratio) inhot water bath until the reaction ceased. Researchers have reportedproblems with acid extraction, like differential rates of digestion fordifferent plant parts, loss of phytoliths during processing, and the un-desirability of using noxious chemicals (Pearsall, 1989; Parr et al.,2001b). Keeping all these in mind the oxidation time for the differentplant parts were calibrated for the maximum yield of phytoliths
t and habitat (after Naskar and Guha Bakshi, 1987).
Habitat
erb Swampy/ intertidal mangrove zone and tidal mangrove zoneTrue Mangrove Decline zone and zone for colonization of non- littoralspecies zoneTidal Mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ Intertidal Mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneTidal mangrove zone and zone for colonization of non- littoral species zone
ber True Mangrove decline zone and zone for colonization of non- littoral speciesTidal mangrove zone, true mangrove decline zone and zone for colonizationof non- littoral species zoneXerophytic non- mangrove and dry evergreen forestsTrue mangrove decline zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ Intertidal Mangrove zoneSwampy/ intertidal mangrove zoneZone for colonization of non- littoral speciesXerophytic non- mangrove and dry evergreen forestsSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneTidal mangrove zone and true mangrove decline zoneTidal mangrove zone and true mangrove decline zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zone
l tree Colonization of non- littoral species zoneTrue mangrove decline zone and colonization of non- littoral species zone
l tree True mangrove decline zone and colonization of non- littoral species zoneXerophytic non- mangrove and dry evergreen forestsSwampy/ intertidal mangrove zone and tidal mangrove zoneSwampy/ intertidal mangrove zone and tidal mangrove zone
Table 2List of plant taxa and plant tissues from which phytoliths were extracted along with brief description and acronyms of the morpotypes [P = Present; A = Absent].
Taxon Phytolith morphotypes Acronym Presence ofDiagnosticCharacters
Size Range
PteridaceaeAcrostichum aureumLeaf Perforated plate with circular pores having undulating margin PER CIRCULAR P 20–100 μm
Stomate STO 40–60 μmAnticlinal plate ANT 150–270 μm
SterculiaceaeHeritiera fomesLeaf Multicellular hair cell base comprising
of 5–6 surrounding cells.MULT HAIR BASE P 20–40 μm
Sclereids SCL 150–175 μmStomate STO 40–60 μm
Seed coat Globular facetate bodies. GLO FACET P 20–42 μmPneumatophore Blocky polyhedral bodies BLOCK POLY P 9–24 μm
LythraceaeSonneratia caseolarisLeaf Globular psilate bodies GLO PSI P 4–14 μm
Sclereids SCL 100–120 μmCylindric sulcate tracheids CYLIN SUL 72–80 μm
Pneumatophore Cymbiform porous bodies: Boat shapedbodies with folded outer surface and theperiphery of the inner surface with minutepores with striations on its inner surface.
CYMB POR P 9–12 μm
Blocky polyhedral bodies BLOCK POLY 2. 9–36 μm
Sonneratia apetalaLeaf Globular psilate bodies GLO PSI P 1.4–14 μm
Sclereids SCL 100–120 μmCylindric sulcate tracheids CYLIN SUL 72–80 μm
Pneumatophore Cymbiform porous bodies: Boat shapedbodies with folded outer surface and theperiphery of the inner surface with minutepores with striations on its inner surface.
CYMB POR P 9–12 μm
Blocky polyhedral bodies BLOCK POLY 9–36 μm
CombretaceaeLumnitzera racemosaLeaf Irregular scrobiculate bodies. IRR SCROBI P 12–30 μm
AsclepiadaceaeSarcolobus globosusLeaf Globular psilate bodies (almost smooth
surface texture but seldom slight grainy).GLO PSI P 4–14 μm
AcanthaceaeAcanthus ilicifoliusLeaf Multicellular hair cell base. MULT HAIR BASE P 36–40 μm
Cylindric sulcate tracheids CYLIN SUL 72–80 μmEpidermal cells EPI 120–240 μm
Seed coat Verrucate spheroids (with foldings on theupper surface and lower surface almost smooth).
VER SPH P 16–30 μm
Fruit coat Verrucate spheroids (with foldings on the uppersurface and lower surface almost smooth).
VER SPH P 24–38 μm
Sclereids SCL 100–150 μm
Avicennia albaLeaf Globular granulate bodies (rough wrinkled/undulating surface). GLO GRAN P 2–6 μm
TamaricaceaeTamarix gallicaLeaf Globular granulate bodies (rough wrinkled/
undulating surface).GLO GRAN P 8–28 μm
EuphorbiaceaeExcoecaria agallochaLeaf Stomate STO A 40–60 μm
Cylindric sulcate tracheids CYLIN SUL 72–80 μmSclereids SCL 120–140 μm
Pedilanthus tithymaloidesLeaf Globular granulate bodies (rough wrinkled/
undulating surface).GLO GRAN P 8–21 μm
(continued on next page)
51S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
Table 2 (continued)
Taxon Phytolith morphotypes Acronym Presence ofDiagnosticCharacters
Size Range
RhizophoraceaeBruguiera gymnorrhizaLeaf Globular granulate bodies (rough wrinkled surface). GLO GRAN P 8–21 μmFlower Unicellular hair cell: Hair cell with thick
margin and narrow lumen in between.UNI HAIR P 180–220 μm
Pneumatophore Blocky polyhedral bodies BLOCK POLY P 9–24 μm
Bruguiera cylindricaLeaf Epidermal cell EPI A 100–240 μm
Stomate STO 40–44 μmCylindric sulcate tracheids CYLIN SUL 72–80 μm
Flower Irregular spheres with surface folding. IRR SPH P 14–22 μm
Ceriops tagalLeaf Globular lacunose bodies (with surface
perforations/ depressions).GLO LAC P 4–18 μm
Flower Irregular spheres with surface folding. IRR SPH P 8–22 μm
Rhizophora apiculataLeaf Globular lacunose bodies (with surface
perforations/ depressions).GLO LAC P 4–18 μm
Flower Irregular spheres with surface folding. IRR SPH P 14–34 μmSclereids SCL 150–175 μm
Kandelia candelLeaf Epidermal cell EPI A 44–80 μm
Stomate STO 40–60 μmMeliaceaeXylocarpus granatumLeaf Epidermal cell EPI A 44–80 μm
Sclereids SCL 80–100 μm
Xylocarpus moluccensisLeaf Stomate STO A 20–28 μm
Cylindric sulcate tracheid CYLIN SUL 40–80 μmEpidermal cell EPI 44–80 μm
Amoora cucullataLeaf Stomate STO A 24–36 μm
MalvaceaeThespesia populneaLeaf No phytoliths found
ApocynaceaeThevetia peruvianaLeaf No phytoliths found
MyrsinaceaeAegiceras corniculatumLeaf Globular lacunose bodies (with surface
perforations/ depressions).GLO LAC P 6–18 μm
PlumbaginaceaeAegialitis rotundifoliaLeaf Globular granulate bodies (rough wrinkled/
undulating surface).GLO GRA P 8–20 μm
Large irregularly folded perforated plate. IRR PER 12–60 μm
BoraginaceaeHeliotropium curassavicumLeaf Hair cell base with concentric ring pattern
and undulating surface.CONC HAIR BASE P 12–28 μm
AsteraceaeGrangea maderaspatanaLeaf Hair cell-multicellular hair segmented with a swollen
notch and surface covered with blunt spines.Smooth walled hair cells also present.
MULT HAIR P 32–44 μm
Flower Hair cell-multicellular hair segmented with a swollennotch and surface covered with blunt spines.Smooth walled hair cells also present.
MULT HAIR P 32–44 μm
52 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65

Table 2 (continued)
Taxon Phytolith morphotypes Acronym Presence ofDiagnosticCharacters
Size Range
FabaceaeDerris scandensLeaf Stomate STO A 40–48 μm
Epidermal cell. EPI A 120–240 μm
VerbenaceaeClerodendrum inermeLeaf Epidermal cell. EPI A 120–240 μm
Cylindric sulcate tracheids CYLIN SUL 72–80 μm
VitaceaeVitis trifoliaLeaf Cylindric sulcate tracheids CYLIN SUL A 72–80 μm
ChenopodiaceaeSuaeda nudifloraLeaf No phytoliths found
CyperaceaeScirpus articulatusLeaf Perforated plate with elliptical pores at regular intervals. PER ELLIP P 80–150 μm
Sinuate elongated bodies with conical projections. SINU ELON 24–82 μm
PandanaceaePandanus fascicularisLeaf Papillae base around Stomata. PAP P 40–48 μm
ArecaceaePhoenix paludosaLeaf Globular echinate bodies: Spherical in shape of varying
size with spinulose projection over the entire surfaceGLO ECH P 4–8 μm
Nypa fruticansLeaf Hat shaped conical bodies: spheroidal in shape from top
view with spinulose projection.HAT P 4–8 μm
Spathe Hat shaped conical bodies: spheroidal in shape from topview with spinulose projection, similar to those found in leaves.
HAT P 4 – 8 μm
53S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
owing to their differential rates of digestion. For the harder plantparts like pneumatophores, reaction was more vigorous and took lon-ger time to cease than the softer parts like leaf, flower or fruit. It wasthen followed by repeated washing with distilled water. The residuecontaining phytoliths were collected in water and stored in vials.Slides were then prepared with minimum amount of residuecontaining phytoliths using polyvinyl alcohol and mounted in DPXfor photomicrography, and in distilled water for counting.
The total number of phytoliths counted was 15,231. Combiningall tissues, an average of 354 phytoliths was counted from every spe-cies. Study and photomicrography of different phytolith morphotypewere done using a Zeiss Axioskop microscope at × 450 magnifica-tion. Scanning Electron Microscopy of distinctive phytolithmorphotypes of each taxon was done using a Scanning Electron Mi-croscope (Carl Zeiss – Evo 40) at GSI (Geological Survey of India),Kolkata. A number of new phytolith morphotypes are reported anddescribed using the International Code for Phytolith Nomenclature(ICPN) of Madella et al., 2005. When describing previously reportedphytolith morphotypes we consulted Wallis (2003) and Kaelhoferand Piperno.
After quantification, the leaf phytolith data were subjected to sta-tistical analyses. Leaves were found to be more consistent producersof phytoliths than other plant parts; therefore, we only used phytolithdata from leaves for statistical analysis. Cluster analysis (CA) and PCAanalysis were employed using SPSS 16 to test if taxonomically relatedspecies could be identified and grouped on the basis of the presenceand frequency of occurrence of different types. These two statisticalmethods reduce the number of variables to a smaller set of principal
components to reveal its internal structure, group type variablesand detect covariance (Kim and Mueller, 1978; Basilevsky, 1994;Jolliffe, 2002). The phytolith data for PCA were not normalized andran on the original correlation matrix. Ward’s method and thesquared Euclidean distance with the variables rescaled to 0–1 wereused for the CA. Both the results of PCA and CA were found similarin nature.
3. Results
3.1. Pattern of phytolith production
Out of the 33 non-grass plants analyzed, 30 taxa (91%) have beenproved to be phytolith producers of which only 42% have been foundto be significant phytolith producers. Descriptions of the recoveredphytolith morphotypes along with their acronyms and anatomical or-igin in the analyzed taxa are represented in Table 2. 25 phytolithmorphotypes are obtained from different parts (i.e., leaf, flower,fruit coat, seed coat, spathe and pneumatophore) of the studiedplant community (Figs. 2, 3, 4). 17 morphotypes are recovered fromangiosperm leaves i.e. globular lacunose (GLO LAC), globular granu-late (GLO GRAN), globular psilate (GLO PSI), large irregularly foldedperforated plate (IRR PER), papillae base (PAP), perforate plate withelliptical pores (PER ELLIP), sinuate elongate bodies (SINU ELON),multicellular hair cell base (MULTI HAIR BASE), hair cell base withconcentric rings (CONC HAIR BASE), globular echinate (GLO ECH),hat-shaped conical bodies (HAT), multicellular hair (MULTI HAIR), ir-regular scrobiculate bodies (IRR SCROBI), epidermal cell (EPI),
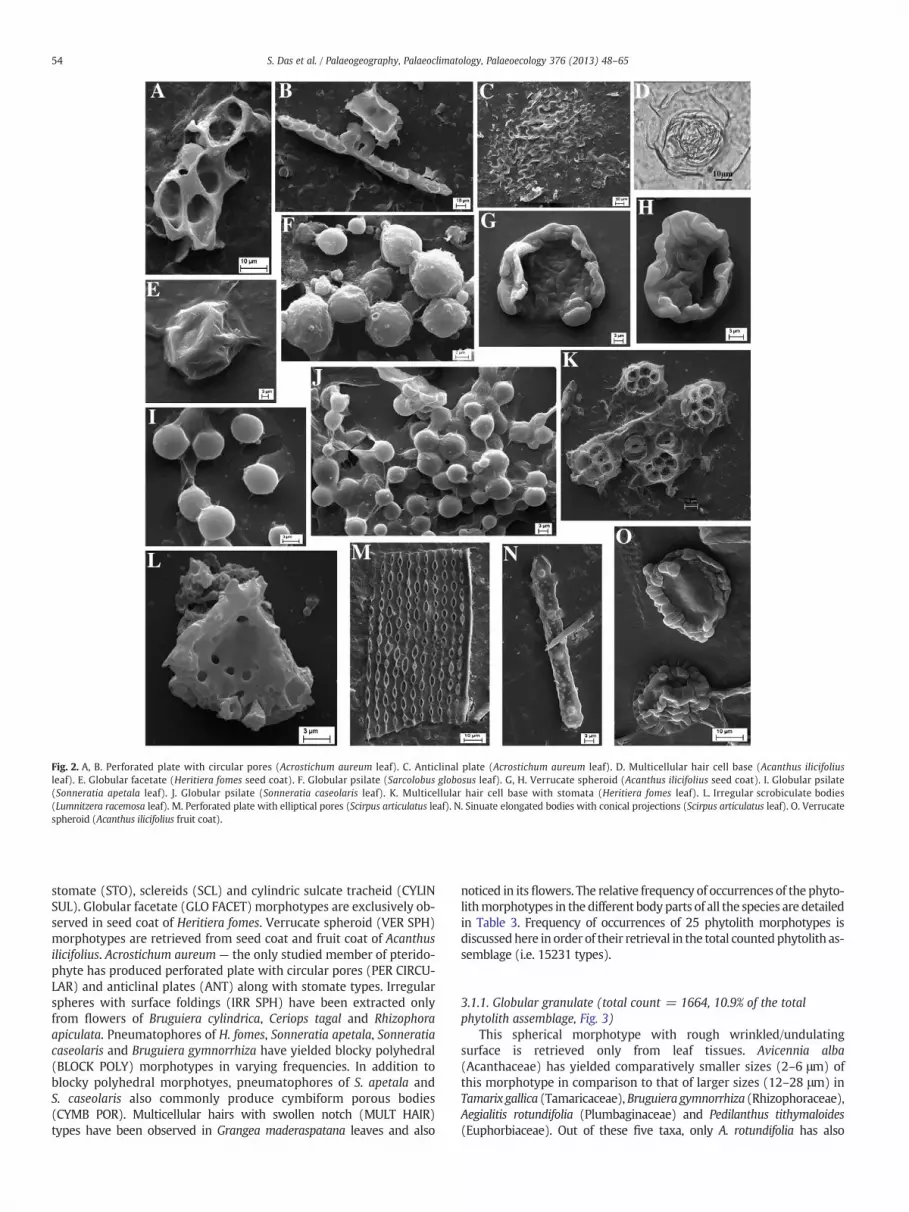
Fig. 2. A, B. Perforated plate with circular pores (Acrostichum aureum leaf). C. Anticlinal plate (Acrostichum aureum leaf). D. Multicellular hair cell base (Acanthus ilicifoliusleaf). E. Globular facetate (Heritiera fomes seed coat). F. Globular psilate (Sarcolobus globosus leaf). G, H. Verrucate spheroid (Acanthus ilicifolius seed coat). I. Globular psilate(Sonneratia apetala leaf). J. Globular psilate (Sonneratia caseolaris leaf). K. Multicellular hair cell base with stomata (Heritiera fomes leaf). L. Irregular scrobiculate bodies(Lumnitzera racemosa leaf). M. Perforated plate with elliptical pores (Scirpus articulatus leaf). N. Sinuate elongated bodies with conical projections (Scirpus articulatus leaf). O. Verrucatespheroid (Acanthus ilicifolius fruit coat).
54 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
stomate (STO), sclereids (SCL) and cylindric sulcate tracheid (CYLINSUL). Globular facetate (GLO FACET) morphotypes are exclusively ob-served in seed coat of Heritiera fomes. Verrucate spheroid (VER SPH)morphotypes are retrieved from seed coat and fruit coat of Acanthusilicifolius. Acrostichum aureum — the only studied member of pterido-phyte has produced perforated plate with circular pores (PER CIRCU-LAR) and anticlinal plates (ANT) along with stomate types. Irregularspheres with surface foldings (IRR SPH) have been extracted onlyfrom flowers of Bruguiera cylindrica, Ceriops tagal and Rhizophoraapiculata. Pneumatophores of H. fomes, Sonneratia apetala, Sonneratiacaseolaris and Bruguiera gymnorrhiza have yielded blocky polyhedral(BLOCK POLY) morphotypes in varying frequencies. In addition toblocky polyhedral morphotyes, pneumatophores of S. apetala andS. caseolaris also commonly produce cymbiform porous bodies(CYMB POR). Multicellular hairs with swollen notch (MULT HAIR)types have been observed in Grangea maderaspatana leaves and also
noticed in its flowers. The relative frequency of occurrences of the phyto-lithmorphotypes in the different body parts of all the species are detailedin Table 3. Frequency of occurrences of 25 phytolith morphotypes isdiscussedhere in order of their retrieval in the total countedphytolith as-semblage (i.e. 15231 types).
3.1.1. Globular granulate (total count = 1664, 10.9% of the totalphytolith assemblage, Fig. 3)
This spherical morphotype with rough wrinkled/undulatingsurface is retrieved only from leaf tissues. Avicennia alba(Acanthaceae) has yielded comparatively smaller sizes (2–6 μm) ofthis morphotype in comparison to that of larger sizes (12–28 μm) inTamarix gallica (Tamaricaceae),Bruguiera gymnorrhiza (Rhizophoraceae),Aegialitis rotundifolia (Plumbaginaceae) and Pedilanthus tithymaloides(Euphorbiaceae). Out of these five taxa, only A. rotundifolia has also
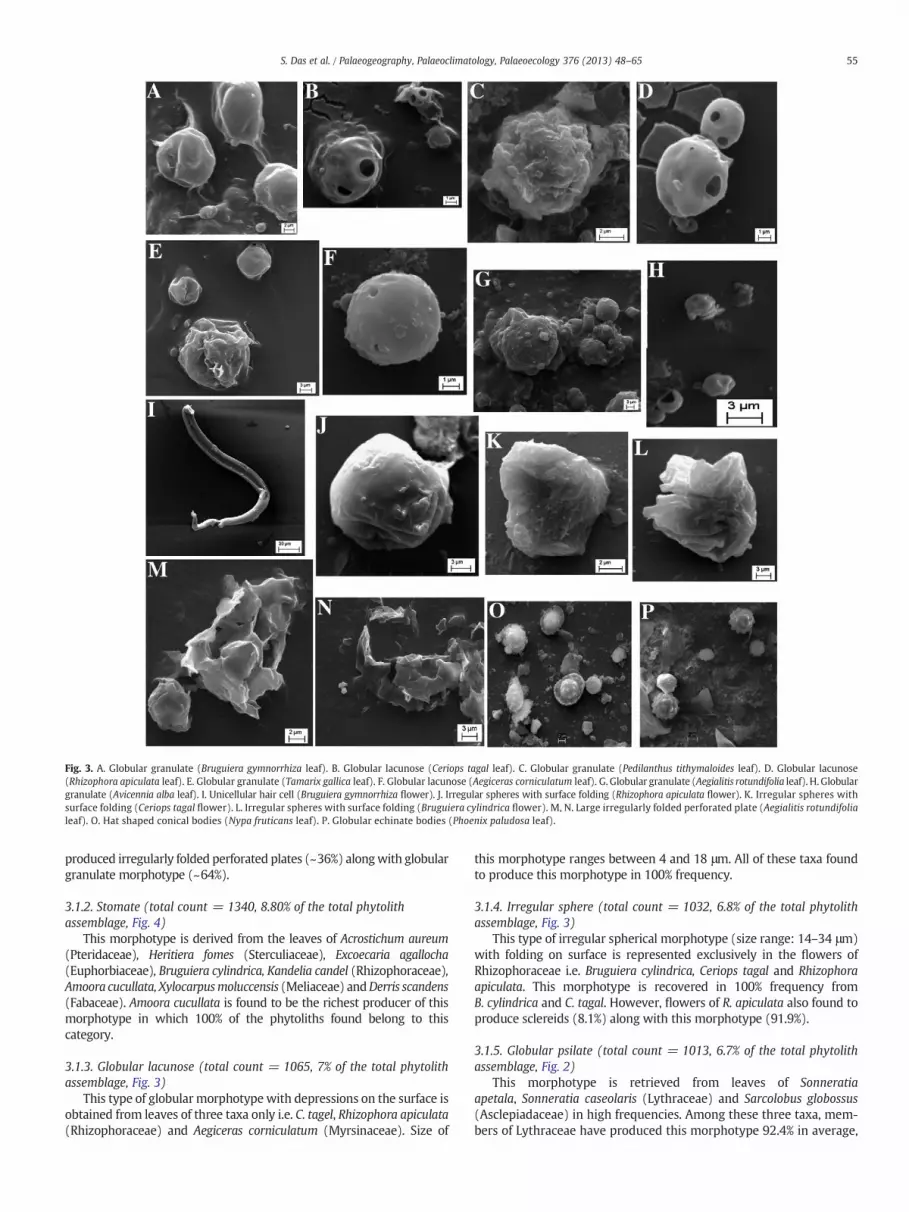
Fig. 3. A. Globular granulate (Bruguiera gymnorrhiza leaf). B. Globular lacunose (Ceriops tagal leaf). C. Globular granulate (Pedilanthus tithymaloides leaf). D. Globular lacunose(Rhizophora apiculata leaf). E. Globular granulate (Tamarix gallica leaf). F. Globular lacunose (Aegiceras corniculatum leaf). G. Globular granulate (Aegialitis rotundifolia leaf). H. Globulargranulate (Avicennia alba leaf). I. Unicellular hair cell (Bruguiera gymnorrhiza flower). J. Irregular spheres with surface folding (Rhizophora apiculata flower). K. Irregular spheres withsurface folding (Ceriops tagal flower). L. Irregular spheres with surface folding (Bruguiera cylindrica flower). M, N. Large irregularly folded perforated plate (Aegialitis rotundifolialeaf). O. Hat shaped conical bodies (Nypa fruticans leaf). P. Globular echinate bodies (Phoenix paludosa leaf).
55S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
produced irregularly folded perforated plates (~36%) alongwith globulargranulate morphotype (~64%).
3.1.2. Stomate (total count = 1340, 8.80% of the total phytolithassemblage, Fig. 4)
This morphotype is derived from the leaves of Acrostichum aureum(Pteridaceae), Heritiera fomes (Sterculiaceae), Excoecaria agallocha(Euphorbiaceae), Bruguiera cylindrica, Kandelia candel (Rhizophoraceae),Amoora cucullata, Xylocarpusmoluccensis (Meliaceae) andDerris scandens(Fabaceae). Amoora cucullata is found to be the richest producer of thismorphotype in which 100% of the phytoliths found belong to thiscategory.
3.1.3. Globular lacunose (total count = 1065, 7% of the total phytolithassemblage, Fig. 3)
This type of globular morphotype with depressions on the surface isobtained from leaves of three taxa only i.e. C. tagel, Rhizophora apiculata(Rhizophoraceae) and Aegiceras corniculatum (Myrsinaceae). Size of
this morphotype ranges between 4 and 18 μm. All of these taxa foundto produce this morphotype in 100% frequency.
3.1.4. Irregular sphere (total count = 1032, 6.8% of the total phytolithassemblage, Fig. 3)
This type of irregular spherical morphotype (size range: 14–34 μm)with folding on surface is represented exclusively in the flowers ofRhizophoraceae i.e. Bruguiera cylindrica, Ceriops tagal and Rhizophoraapiculata. This morphotype is recovered in 100% frequency fromB. cylindrica and C. tagal. However, flowers of R. apiculata also found toproduce sclereids (8.1%) along with this morphotype (91.9%).
3.1.5. Globular psilate (total count = 1013, 6.7% of the total phytolithassemblage, Fig. 2)
This morphotype is retrieved from leaves of Sonneratiaapetala, Sonneratia caseolaris (Lythraceae) and Sarcolobus globossus(Asclepiadaceae) in high frequencies. Among these three taxa, mem-bers of Lythraceae have produced this morphotype 92.4% in average,

Fig. 4. A, B. Multicellular hair cell- partly broken (Grangea maderaspatana leaf). C, D. Hair cell base with concentric rings (Heliotropium curassavicum leaf). E. Papillae base aroundstomata (Pandanus fascicularis leaf). F. Cymbiform porous bodies (Sonneratia apetala pneumatophore). G. Cymbiform porous bodies (Sonneratia caseolaris pneumatophore). H. Blockypolyhedral bodies (Sonneratia apetala pneumatophore). I. Blocky polyhedral bodies (Sonneratia caseolaris pneumatophore). J. Blocky polyhedral bodies (Bruguiera gymnorrhizaleaf). K. Blocky polyhedral bodies (Avicennia alba pneumatophore). L. Blocky polyhedral bodies (Heritiera fomes pneumatophore). M. Stomate. N, O. Sclereids. P. Cylindric sulcatetracheids. Q. Epidermal cell.
56 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
wherein S. globosus, this type is found to occur in 100% frequency. Sizeof this morphotype ranges between 4 and 14 μm.
3.1.6. Blocky polyhedral (total count = 920, 6% of the total phytolithassemblage, Fig. 4)
This morphotype is derived only from pneumatophores of someunrelated families i.e. Sterculiaceae (Heritiera fomes), Lythraceae(Sonneratia apetala and Sonneratia caseolaris) and Rhizophoraceae(Bruguiera gymnorrhiza). Of these four taxa, the highest yield is ob-served in H. fomes and B. gymnorrhiza (100%) followed by S. caseolaris(79.7%) and S. apetala (60.5%).
3.1.7. Cylindric sulcate tracheids (total count = 891, 5.8% of the totalphytolith assemblage, Fig. 4)
This morphotype is obtained from the tracheids of the leaf tissuesof seven taxa i.e. Sonneratia apetala, Sonneratia caseolaris(Lythraceae), Acanthus ilicifolius (Acanthaceae), Excoecaria agallocha
(Euphorbiaceae), Bruguiera cylindrica (Rhizophoraceae), Clerodendruminerme (Verbenaceae) and Vitis trifolia (Vitaceae). The average frequen-cy per species is 36.6% (range: 2.2–100%). The top most producer isV. trifoliawith 100% frequency of recovery.
3.1.8. Multicellular hair (total count = 706, 4.6% of the total phytolithassemblage, Fig. 4)
Morphotype like multicellular segmented hair with a swollennotch and blunt spines on the surface is observed solitarily in theleaf and flower of Grangea maderaspatana (Asteraceae).
3.1.9. Hat shaped (total count = 706, 4.6% of the total phytolithassemblage, Fig. 3)
Leaf and spathe of Nypa fruticans (Arecaceae) only produce thishat shaped conical morphotype. In both of the cases hat shapedmorphotypes are recovered in 100% frequencies.
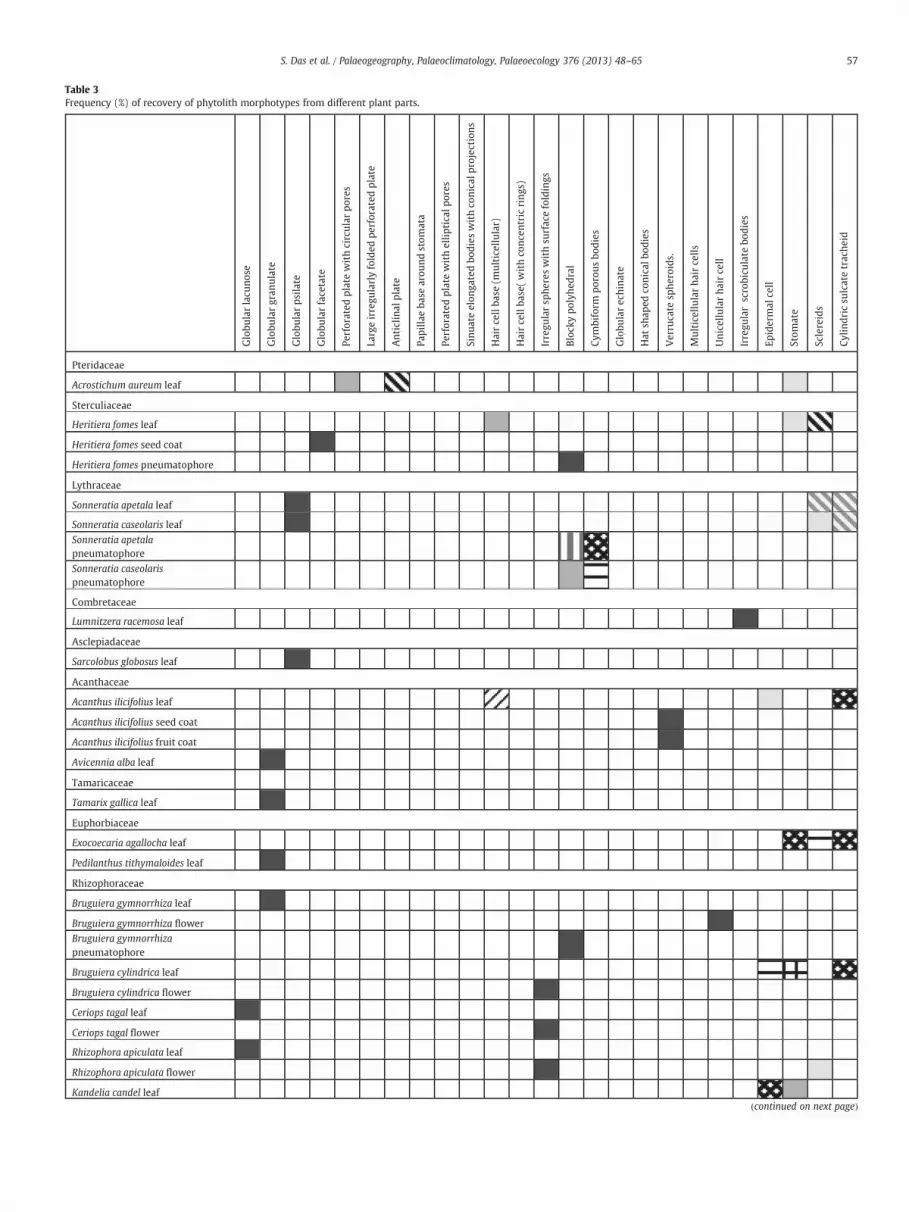
Table 3Frequency (%) of recovery of phytolith morphotypes from different plant parts.
Glo
bu
lar
lacu
no
se
Glo
bu
lar
gra
nu
late
Glo
bu
lar
psi
late
Glo
bu
lar
face
tate
Pe
rfo
rate
d p
late
wit
h c
ircu
lar
po
res
Larg
e i
rre
gu
larl
y f
old
ed
pe
rfo
rate
d p
late
An
ticl
ina
l p
late
Pa
pil
lae
ba
se a
rou
nd
sto
ma
ta
Pe
rfo
rate
d p
late
wit
h e
llip
tica
l p
ore
s
Sin
ua
te e
lon
ga
ted
bo
die
s w
ith
co
nic
al
pro
ject
ion
s
Ha
ir c
ell
ba
se (
mu
ltic
ell
ula
r)
Ha
ir c
ell
ba
se(
wit
h c
on
cen
tric
rin
gs)
Irre
gu
lar
sph
ere
s w
ith
su
rfa
ce f
old
ing
s
Blo
cky
po
lyh
ed
ral
Cy
mb
ifo
rm p
oro
us
bo
die
s
Glo
bu
lar
ech
ina
te
Ha
t sh
ap
ed
co
nic
al
bo
die
s
Ve
rru
cate
sp
he
roid
s.
Mu
ltic
ell
ula
r h
air
ce
lls
Un
ice
llu
lar
ha
ir c
ell
Irre
gu
lar
scr
ob
icu
late
bo
die
s
Ep
ide
rma
l ce
ll
Sto
ma
te
Scl
ere
ids
Cy
lin
dri
c su
lca
te t
rach
eid
Pteridaceae
Acrostichum aureum leaf
Sterculiaceae
Heritiera fomes leaf
Heritiera fomes seed coat
Heritiera fomes pneumatophore
Lythraceae
Sonneratia apetala leaf
Sonneratia caseolaris leaf
Sonneratia apetala
pneumatophore
Sonneratia caseolaris
pneumatophore
Combretaceae
Lumnitzera racemosa leaf
Asclepiadaceae
Sarcolobus globosus leaf
Acanthaceae
Acanthus ilicifolius leaf
Acanthus ilicifolius seed coat
Acanthus ilicifolius fruit coat
Avicennia alba leaf
Tamaricaceae
Tamarix gallica leaf
Euphorbiaceae
Exocoecaria agallocha leaf
Pedilanthus tithymaloides leaf
Rhizophoraceae
Bruguiera gymnorrhiza leaf
Bruguiera gymnorrhiza flower
Bruguiera gymnorrhiza
pneumatophore
Bruguiera cylindrica leaf
Bruguiera cylindrica flower
Ceriops tagal leaf
Ceriops tagal flower
Rhizophora apiculata leaf
Rhizophora apiculata flower
Kandelia candel leaf
(continued on next page)
57S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65

Glo
bu
lar
lacu
no
se
Glo
bu
lar
gra
nu
late
Glo
bu
lar
psi
late
Glo
bu
lar
face
tate
Pe
rfo
rate
d p
late
wit
h c
ircu
lar
po
res
Larg
e i
rre
gu
larl
y f
old
ed
pe
rfo
rate
d p
late
An
ticl
ina
l p
late
Pa
pil
lae
ba
se a
rou
nd
Sto
ma
ta
Pe
rfo
rate
d p
late
wit
h e
llip
tica
l p
ore
s
Sin
ua
te e
lon
ga
ted
bo
die
s w
ith
co
nic
al
pro
ject
ion
s
Ha
ir c
ell
ba
se (
mu
ltic
ell
ula
r)
Ha
ir c
ell
ba
se(
wit
h c
on
cen
tric
rin
gs)
Irre
gu
lar
sph
ere
s w
ith
su
rfa
ce f
old
ing
s
Blo
cky
po
lyh
ed
ral
Cy
mb
ifo
rm p
oro
us
bo
die
s
Glo
bu
lar
ech
ina
te
Ha
t sh
ap
ed
co
nic
al
bo
die
s
Ve
rru
cate
sp
he
roid
s.
Mu
ltic
ell
ula
r h
air
ce
lls
Un
ice
llu
lar
ha
ir c
ell
Irre
gu
lar
scr
ob
icu
late
bo
die
s
Ep
ide
rma
l ce
ll
Sto
ma
te
Scl
ere
ids
Cy
lin
dri
c su
lca
te t
rach
eid
Myrsinaceae
Aegiceras corniculatum leaf
Plumbaginaceae
Aegialitis rotundifolia leaf
Boraginaceae
Heliotropium curassavicum leaf
Asteraceae
Grangea maderaspatana leaf
Grangea maderaspatana flower
Meliaceae
Amoora cucullata leaf
Xylocarpus granatum leaf
Xylocarpus moluccensis leaf
Fabaceae
Derris scandens leaf
Verbenaceae
Clerodendrum inerme leaf
Vitaceae
Vitis trifolia leaf
Arecaceae
Phoenix paludosa leaf
Nypa fruticans leaf
Nypa fruticans spathe
Cyperaceae
Scirpus articulatus leaf
Pandanaceae
Pandanus fascicularis leaf
Legends
<5% 5-10% 10-20% 20-30% 30-40% 90-100%
40-50% 50-60% 60-70% 70-80% 80-90%
Table 3 (continued)
58 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
3.1.10. Epidermal cell (total count = 678, 4.5% of the total phytolithassemblage, Fig. 4)
Epidermal cell with polygonal outline has been observed in Acanthusilicifolius (Acanthaceae), Excoecaria agallocha (Euphorbiaceae), Bruguieracylindrica, Kandelia candel (Rhizophoraceae), Xylocarpus granatum,Xylocarpus moluccensis (Meliaceae), Derris scandens (Fabaceae) and
Vitis trifolia (Vitaceae). Mean production represented is 27.4% perspecies. This morphotype has been observed only in the leaf tissuesand the highest abundance has seen in Clerodendrum inerme(43.8%) followed by D. scandens (35.5%), X. granatum (34%), K. candel(28.5%), B. cylindrica (23.4%), X. moluccensis (17%) and A. ilicifolius(9.4%).

Fig. 5. Principal Components Analysis (PCA) ordination diagram (SPSS 16) of the leafphytoliths showing the interrelationship of the morphotypes.
59S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
3.1.11. Verrucate spheroids (total count = 721, 4.7% of the totalphytolith assemblage, Fig. 2)
This morphotype is produced solitarily in fruit and seed coats ofAcanthus ilicifolius (Acanthaceae) and in both of the cases productionfrequency is 100%.
3.1.12. Sclereids (total count = 593, 3.9% of the total phytolithassemblage, Fig. 4)
This morphotype is produced by leaves of the true mangrove plantsviz., Xylocarpus granatum (66%), Excoecaria agallocha (29.6%), Heritierafomes (17.1%), Sonneratia caseolaris (5.9%) and Sonneratia apetala (4.6%)in varying frequencies. The morphotype is also extracted from theflowers of Rhizophora apiculata although in negligible frequency (8.1%).
3.1.13. Cymbiform porous (total count = 497, 3.3% of the total phytolithassemblage, Fig. 4)
This morphotype is only recovered from pneumatophores of themembers of Lythraceae i.e. Sonneratia apetala and Sonneratia caseolaris.The morphotype is best represented in S. apetala (39.6%) than inS. caseolaris (20.2%).
3.1.14. Multicellular hair cell base (total count = 476, 3.1% of the totalphytolith assemblage, Fig. 2)
Leaves of only two taxa have produced this morphotype viz.,Heritiera fomes (Sterculiaceae) and Acanthus ilicifolius (Acanthaceae).The average representation per species is 67.4% and H. fomes (74.7%)is the richest producer.
3.1.15. Globular facetate (total count = 360, ~2.4% of the total phytolithassemblage, Fig. 2)
This type of facetate body ranges in size from 20 to 42 μm and areretrieved in 100% frequency from seed coat of Heritiera fomes only.
3.1.16. Hair base (with concentric rings) (total count = 357, ~2.3% ofthe total phytolith assemblage, Fig. 4)
Leaves of Heliotropium curassavicum are found to produce thismorphotype solitarily and the frequency of recovery is 100%. Themorphotype varies in size between12 and 28 μm.
3.1.17. Irregular scrobiculate (total count = 355, ~2.3% of the totalphytolith assemblage, Fig. 2)
This morphotype is obtained only in leaves of Lumnitzera racemosa(Combretaceae) and the frequency of occurrence is 100%.
3.1.18. Globular echinate (total count = 353, ~2.3% of the total phytolithassemblage, Fig. 3)
This morphotype has been recovered exclusively from leaves ofPhoenix paludosa (Arecaceae) and the frequency of retrieval is 100%.
3.1.19. Papillae base (total count = 350, ~2.3% of the total phytolithassemblage, Fig. 4)
This morphotype is only represented in leaf tissues of Pandanusfascicularis (Pandanaceae) and is obtained in 100% frequency.
3.1.20. Unicellular hair cell (total count = 348, ~2.3% of the totalphytolith assemblage, Fig. 3)
This type represents hair cell with a thick cell wall with a size rangeof 180–220 μm was found only in flower of Bruguiera gymnorrhiza(Rhizophoraceae) in 100% frequency.
3.1.21. Perforated plate with circular pores (total count = 270, 1.8% ofthe total phytolith assemblage, Fig. 2) and anticlinal plate (totalcount = 55, 0.4% of the total phytolith assemblage, Fig. 2)
These two morphotypes are produced exclusively in leaf tissues ofAcrostichum aureum (Pteridaceae). The rates of recovery of these twomorphotypes are 76.5% and 15.6% respectively.
3.1.22. Perforated plate with elliptical pores (total count = 224, 1.5%of the total phytolith assemblage, Fig. 2) and sinuate elongate (totalcount = 130, 0.9% of the total phytolith assemblage, Fig. 2)
Scirpus articulatus (Cyperaceae) is the only producer of thesemorphotypes and their rates of recovery are 69.3% and 30.7%respectively.
3.1.23. Irregular perforated (total count = 127, 0.8% of the totalphytolith assemblage, Fig. 3)
This morphotype is produced by leaves of Aegialitis rotundifoliaand the rate of occurrence is ~36%.
3.2. Cluster analysis and principle components analysis: classificationand grouping
The PCA and CA were applied on the leaf phytolith data set to testthe taxonomic significance of these phytolith morphotypes in identi-fying related taxa and also to test reliability of these morphotypes inenvironmental interpretations.
PCA classification is a multivariate ordination technique appliedhere to examine the degree of correspondence between phytolithmorphotypes and taxa. The first two axes of the PCA account for11.2% of the total variance. Such a low percentage can be explainedby the relatively low taxonomic diagnostic power of the non-grassmangrove phytolith morphotypes excepting the unique types. It hasbeen noticed that 19 morphotypes have created 13 principal compo-nents. In the biplot of principal component loadings for 19 phytolithmorphotypes, PER ELLIP (perforated plate with elliptical pore) andSINU ELON (sinuate elongated bodies with conical projections)morphotypes have accounted for the highest loadings followed byPER CIRCULAR (perforated plate with circular pore) and ANT (anticli-nal plate), whereas GLO GRAN (globular granulate), GLO ECH (globu-lar echinate), GLO LAC (globular lacunose), GLO PSI (globular psilate)and HAT (hat shaped conical bodies) showed intermediate positionson axis 1 (Fig. 5). Interestingly EPI (epidermal cell), STO (stomata),SCL (sclereid) and CYLIN SUL (cylindric sulcate) have shown highestnegative loadings on axis 2.
Considering linkage distance and associations of the phytolithmorphotypes, the clustering of leaf morphotypes has been conducted.16 clusters have been identified (Fig. 6, Table 4) and cluster numbers1, 2, 4, 5, 6 and 7 have been found to be separated from the rest showingshort rescaled linkage distance (b1) indicating that taxonomically

60 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
unrelated families like, Rhizophoraceae, Meliaceae, Euphorbiaceae,Acanthaceae, Asclepiadaceae, Sterculiaceae, Lythraceae, Fabaceae andVitaceae produce similar phytolithmorphotypes hence having least tax-onomic value. In contrast, thewider dissimilarity coefficient seen in restof the 10 clusters point towards their uniqueness as displayed by dicot-yledonous members like Asteraceae, Plumbaginaceae, Combretaceae,Boraginaceae and monocotyledons like Arecaceae, Pandanaceae,Cyperaceae and pteridophyte i.e. Pteridaceae.
Qualitative data from Energy Dispersive X ray Spectroscopy (EDX)analysis of some of these phytolith morphotypes clearly indicates thepresence of silicon as the principle elemental constituent in the spec-trum region. Since the peak for oxygen is distinct in the spectrum re-gion, it signifies that the phytoliths are mainly composed of silicondioxide, earlier proved by Jones and Milne (1963). This EDX dataalso indicates the presence of aluminium, magnesium, calcium,
Fig. 6. Dendrograms from cluster analysis (Ward's method, Euclidean distance, by SPSS 16Numbers on the far left indicate the cluster number.
sodium, potassium, iron and carbon (Fig. 7). Presence of carbon isthought to be organic in nature as the phytoliths has been extractedfrom the living tissues and thus indicating biogenic origin of the opal-ine silica particles. The quantitative data showing the relative amountof elements present in the phytoliths are given in Table 5.
4. Discussion
In this study we have analyzed phytolith spectra produced by 33non-grass taxa from the Indian part of the Sunderbans, one of theworld's heritage sites and unique for its deltaic sub-environments andmangrove ecosystem. Present study creates an inventory of phytolithsfrom native mangrove plants of the Sunderbans and sheds light onwhether thesephytolithmorphotypes impart any taxonomic significance,
). Taxa clustered on the basis of specific morphotype frequency (leaf phytoliths only).
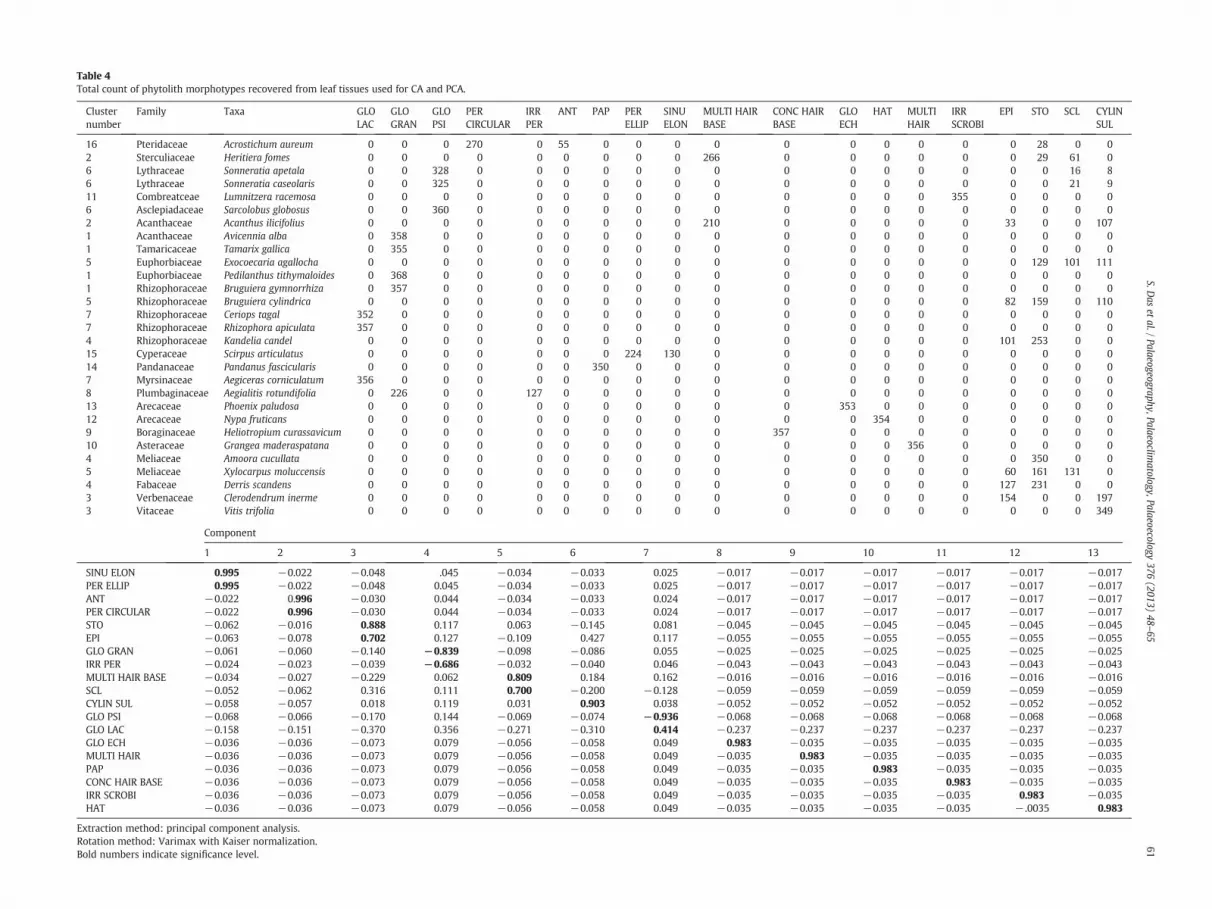
Table 4Total count of phytolith morphotypes recovered from leaf tissues used for CA and PCA.
Clusternumber
Family Taxa GLOLAC
GLOGRAN
GLOPSI
PERCIRCULAR
IRRPER
ANT PAP PERELLIP
SINUELON
MULTI HAIRBASE
CONC HAIRBASE
GLOECH
HAT MULTIHAIR
IRRSCROBI
EPI STO SCL CYLINSUL
16 Pteridaceae Acrostichum aureum 0 0 0 270 0 55 0 0 0 0 0 0 0 0 0 0 28 0 02 Sterculiaceae Heritiera fomes 0 0 0 0 0 0 0 0 0 266 0 0 0 0 0 0 29 61 06 Lythraceae Sonneratia apetala 0 0 328 0 0 0 0 0 0 0 0 0 0 0 0 0 0 16 86 Lythraceae Sonneratia caseolaris 0 0 325 0 0 0 0 0 0 0 0 0 0 0 0 0 0 21 911 Combreatceae Lumnitzera racemosa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 355 0 0 0 06 Asclepiadaceae Sarcolobus globosus 0 0 360 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 02 Acanthaceae Acanthus ilicifolius 0 0 0 0 0 0 0 0 0 210 0 0 0 0 0 33 0 0 1071 Acanthaceae Avicennia alba 0 358 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 01 Tamaricaceae Tamarix gallica 0 355 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 05 Euphorbiaceae Exocoecaria agallocha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 129 101 1111 Euphorbiaceae Pedilanthus tithymaloides 0 368 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 01 Rhizophoraceae Bruguiera gymnorrhiza 0 357 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 05 Rhizophoraceae Bruguiera cylindrica 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 82 159 0 1107 Rhizophoraceae Ceriops tagal 352 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 07 Rhizophoraceae Rhizophora apiculata 357 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 04 Rhizophoraceae Kandelia candel 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 101 253 0 015 Cyperaceae Scirpus articulatus 0 0 0 0 0 0 0 224 130 0 0 0 0 0 0 0 0 0 014 Pandanaceae Pandanus fascicularis 0 0 0 0 0 0 350 0 0 0 0 0 0 0 0 0 0 0 07 Myrsinaceae Aegiceras corniculatum 356 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 08 Plumbaginaceae Aegialitis rotundifolia 0 226 0 0 127 0 0 0 0 0 0 0 0 0 0 0 0 0 013 Arecaceae Phoenix paludosa 0 0 0 0 0 0 0 0 0 0 0 353 0 0 0 0 0 0 012 Arecaceae Nypa fruticans 0 0 0 0 0 0 0 0 0 0 0 0 354 0 0 0 0 0 09 Boraginaceae Heliotropium curassavicum 0 0 0 0 0 0 0 0 0 0 357 0 0 0 0 0 0 0 010 Asteraceae Grangea maderaspatana 0 0 0 0 0 0 0 0 0 0 0 0 0 356 0 0 0 0 04 Meliaceae Amoora cucullata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 350 0 05 Meliaceae Xylocarpus moluccensis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 60 161 131 04 Fabaceae Derris scandens 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 127 231 0 03 Verbenaceae Clerodendrum inerme 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 154 0 0 1973 Vitaceae Vitis trifolia 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 349
Component
1 2 3 4 5 6 7 8 9 10 11 12 13
SINU ELON 0.995 −0.022 −0.048 .045 −0.034 −0.033 0.025 −0.017 −0.017 −0.017 −0.017 −0.017 −0.017PER ELLIP 0.995 −0.022 −0.048 0.045 −0.034 −0.033 0.025 −0.017 −0.017 −0.017 −0.017 −0.017 −0.017ANT −0.022 0.996 −0.030 0.044 −0.034 −0.033 0.024 −0.017 −0.017 −0.017 −0.017 −0.017 −0.017PER CIRCULAR −0.022 0.996 −0.030 0.044 −0.034 −0.033 0.024 −0.017 −0.017 −0.017 −0.017 −0.017 −0.017STO −0.062 −0.016 0.888 0.117 0.063 −0.145 0.081 −0.045 −0.045 −0.045 −0.045 −0.045 −0.045EPI −0.063 −0.078 0.702 0.127 −0.109 0.427 0.117 −0.055 −0.055 −0.055 −0.055 −0.055 −0.055GLO GRAN −0.061 −0.060 −0.140 −0.839 −0.098 −0.086 0.055 −0.025 −0.025 −0.025 −0.025 −0.025 −0.025IRR PER −0.024 −0.023 −0.039 −0.686 −0.032 −0.040 0.046 −0.043 −0.043 −0.043 −0.043 −0.043 −0.043MULTI HAIR BASE −0.034 −0.027 −0.229 0.062 0.809 0.184 0.162 −0.016 −0.016 −0.016 −0.016 −0.016 −0.016SCL −0.052 −0.062 0.316 0.111 0.700 −0.200 −0.128 −0.059 −0.059 −0.059 −0.059 −0.059 −0.059CYLIN SUL −0.058 −0.057 0.018 0.119 0.031 0.903 0.038 −0.052 −0.052 −0.052 −0.052 −0.052 −0.052GLO PSI −0.068 −0.066 −0.170 0.144 −0.069 −0.074 −0.936 −0.068 −0.068 −0.068 −0.068 −0.068 −0.068GLO LAC −0.158 −0.151 −0.370 0.356 −0.271 −0.310 0.414 −0.237 −0.237 −0.237 −0.237 −0.237 −0.237GLO ECH −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 0.983 −0.035 −0.035 −0.035 −0.035 −0.035MULTI HAIR −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 −0.035 0.983 −0.035 −0.035 −0.035 −0.035PAP −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 −0.035 −0.035 0.983 −0.035 −0.035 −0.035CONC HAIR BASE −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 −0.035 −0.035 −0.035 0.983 −0.035 −0.035IRR SCROBI −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 −0.035 −0.035 −0.035 −0.035 0.983 −0.035HAT −0.036 −0.036 −0.073 0.079 −0.056 −0.058 0.049 −0.035 −0.035 −0.035 −0.035 − .0035 0.983
Extraction method: principal component analysis.Rotation method: Varimax with Kaiser normalization.Bold numbers indicate significance level.
61S.D
aset
al./Palaeogeography,Palaeoclim
atology,Palaeoecology376
(2013)48
–65
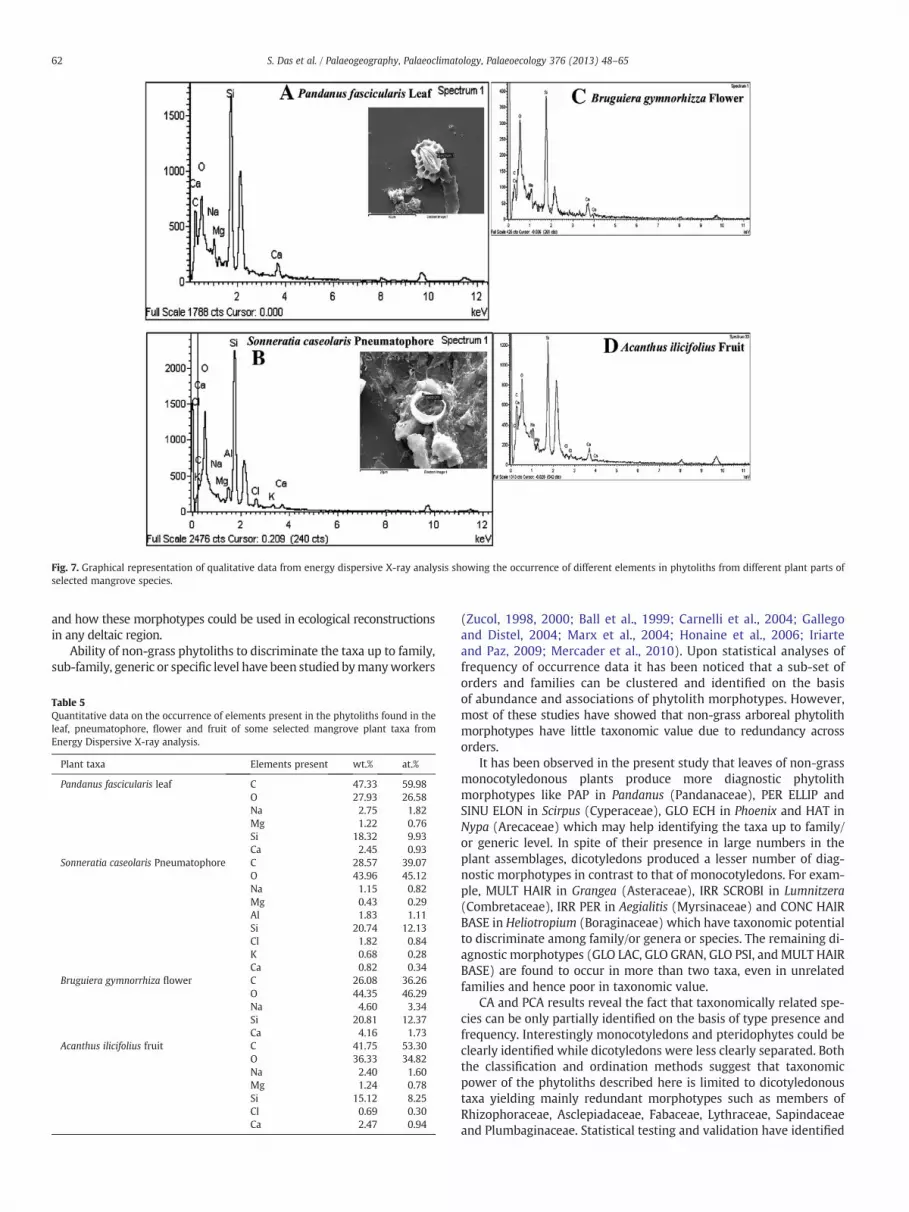
Fig. 7. Graphical representation of qualitative data from energy dispersive X-ray analysis showing the occurrence of different elements in phytoliths from different plant parts ofselected mangrove species.
62 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
and how these morphotypes could be used in ecological reconstructionsin any deltaic region.
Ability of non-grass phytoliths to discriminate the taxa up to family,sub-family, generic or specific level have been studied bymanyworkers
Table 5Quantitative data on the occurrence of elements present in the phytoliths found in theleaf, pneumatophore, flower and fruit of some selected mangrove plant taxa fromEnergy Dispersive X-ray analysis.
Plant taxa Elements present wt.% at.%
Pandanus fascicularis leaf C 47.33 59.98O 27.93 26.58Na 2.75 1.82Mg 1.22 0.76Si 18.32 9.93Ca 2.45 0.93
Sonneratia caseolaris Pneumatophore C 28.57 39.07O 43.96 45.12Na 1.15 0.82Mg 0.43 0.29Al 1.83 1.11Si 20.74 12.13Cl 1.82 0.84K 0.68 0.28Ca 0.82 0.34
Bruguiera gymnorrhiza flower C 26.08 36.26O 44.35 46.29Na 4.60 3.34Si 20.81 12.37Ca 4.16 1.73
Acanthus ilicifolius fruit C 41.75 53.30O 36.33 34.82Na 2.40 1.60Mg 1.24 0.78Si 15.12 8.25Cl 0.69 0.30Ca 2.47 0.94
(Zucol, 1998, 2000; Ball et al., 1999; Carnelli et al., 2004; Gallegoand Distel, 2004; Marx et al., 2004; Honaine et al., 2006; Iriarteand Paz, 2009; Mercader et al., 2010). Upon statistical analyses offrequency of occurrence data it has been noticed that a sub-set oforders and families can be clustered and identified on the basisof abundance and associations of phytolith morphotypes. However,most of these studies have showed that non-grass arboreal phytolithmorphotypes have little taxonomic value due to redundancy acrossorders.
It has been observed in the present study that leaves of non-grassmonocotyledonous plants produce more diagnostic phytolithmorphotypes like PAP in Pandanus (Pandanaceae), PER ELLIP andSINU ELON in Scirpus (Cyperaceae), GLO ECH in Phoenix and HAT inNypa (Arecaceae) which may help identifying the taxa up to family/or generic level. In spite of their presence in large numbers in theplant assemblages, dicotyledons produced a lesser number of diag-nostic morphotypes in contrast to that of monocotyledons. For exam-ple, MULT HAIR in Grangea (Asteraceae), IRR SCROBI in Lumnitzera(Combretaceae), IRR PER in Aegialitis (Myrsinaceae) and CONC HAIRBASE in Heliotropium (Boraginaceae) which have taxonomic potentialto discriminate among family/or genera or species. The remaining di-agnostic morphotypes (GLO LAC, GLO GRAN, GLO PSI, and MULT HAIRBASE) are found to occur in more than two taxa, even in unrelatedfamilies and hence poor in taxonomic value.
CA and PCA results reveal the fact that taxonomically related spe-cies can be only partially identified on the basis of type presence andfrequency. Interestingly monocotyledons and pteridophytes could beclearly identified while dicotyledons were less clearly separated. Boththe classification and ordination methods suggest that taxonomicpower of the phytoliths described here is limited to dicotyledonoustaxa yielding mainly redundant morphotypes such as members ofRhizophoraceae, Asclepiadaceae, Fabaceae, Lythraceae, Sapindaceaeand Plumbaginaceae. Statistical testing and validation have identified

63S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
only 4 diagnostic morphotypes, which can be used in taxonomicdiscrimination.
In spite of the low taxonomic value of the recovered phytolithmorphotypes they show a high eco-environmental signal. It hasbeen observed that abundance and production pattern of phytolithmorphotypes to some extent depends on the salinity gradient alongthe Sundarbans delta. Regular inundation by the tidal water resultsin high salinity in the Sundarbans delta and consequently its absencelowers the salinity level. Salinity gradient decreases with increasingdistance from the tidal mudflat as less frequency and duration oftidal inundation can be a possible reason for low salinity at the land-ward sites (Joshi and Ghose, 2003). Thus based on the tidal activityand subsequent salinity levels in the Sunderbans delta, the ecologicalsuccession of themangrove swamps has been divided into five distinctphases: 1. swampy mangroves or intertidal mangrove zones (riverflats and river-slopes below the high tide level and submerged bytide water twice a day; within 0.0 m–5.0 m from sea level), 2. tidalmangrove zone (covered by spring tide and also during storms;within5.0 m–8.0 m from sea level), 3. true mangrove decline zone (absenceof tidal water only; above 8.0 m from sea level), 4. zone for coloniza-tion of non- littoral species (where the salinity of soil is reduced bythe rain flashing and the soil condition improves the mesophytic veg-etation) and 5. the xerophytic non- mangrove and dry evergreen for-est zone (above 9 m sea level) (Naskar and Guha Bakshi, 1987). Thetrue mangrove species along with the characteristic viviparous germi-nation and pneumatophores are strictly restricted within the swampymangrove forests or on the tidal mangrove forests.
Generally most of the woody plants are known to produce globulargranulate type of phytoliths. Interestingly our study has revealed thatmangrove plants inhabiting mostly the swampy mangrove zones ortidal mangrove zones (such as Acanthus ilicifolius, Aegialitis rotundifolia,Aegiceras corniculatum, Avicennia alba, Bruguiera gymnorrhiza, Ceriopstagal, Phoenix paludosa, Rhizophora apiculata, Sarcolobus globosus andSonneratia caseolaris) produce globular phytoliths with varied surfaceornamentations like GLO LAC, GLO GRAN, GLO FACET, GLO PSI andGLO ECH in abundance. Hence recovery of these morphotypes fromany sediment may be an indicator of true mangrove ecosuccessionalzones i.e. swampy and/or tidal mangrove zones (Fig. 8). A study onphytosuccessions of the Sunderbans area revealed that Sonneratiaapetala, A. alba, A. corniculatum were among the first non grass plantspecies to appear in the newly formed lands of Sunderbans delta(Naskar and Guha Bakshi, 1987). Again, phytolith type i.e. hair cellbase comprising of 5–6 surrounding cells (MULT HAIR BASE) producedin the leaves ofHeritieramay be an indicator of the truemangrove zonesas it inhabits mostly the swampy/intertidal mangrove and tidal man-grove zones (Fig. 8). Phytolith types i.e. blocky polyhedral (BLOCKPOLY) and cymbiform porous bodies (CYMB POR) found in pneumato-phores of mangrove taxa likeHeritiera fomes, S. apetala, S. caseolaris andB. gymnorrhiza may also indicate a true mangrove environment aspneumatophores are strictly restricted in areas of swampy/intertidalmangrove and tidal mangrove zones with high salt concentration toaid the plants in respiration in physiologically dry soil (Fig. 8). Phytolithtypes like sinuate elongated bodies with conical projections (SINUELON) and perforated plate with elliptical pores (PER ELLIP) whichare chiefly produced in Scirpus articulatusmay be an additional indica-tor of tidal mangrove zone as Scirpus sp. dominates this zone (Fig. 8).Moreover, hat shaped conical bodies (HAT) diagnostic of Nypa fruticansleaves may suggest a tidal mangrove environment as Nypa sp. havebeen known to inhabit mostly this zone (Fig. 8). Perforated plate withcircular pores (PER CIRCULAR) and anticlinal plate (ANT) types inAcrostichum aureum leaves may be suggestive of an environment prev-alent in true mangrove decline zone and zone for colonization of non-littoral species, as Acrostichum is seen to abundantly inhabit these twozones (Fig. 8). Moreover, phytolith types i.e. papillae base around sto-mata (PAP)which are diagnostic of Pandanus fascicularis leaves growingmostly in zone for colonization of non- littoral species may act as an
indicator of this zone (Fig. 8). Multicellular hair cells (MULT HAIR)found in Grangea maderaspatana, if found prevalently in any assem-blage may act as an indicator of xerophytic non-mangrove and dry ev-ergreen forest as Grangea sp. inhabits mostly these regions (Fig. 8).
It is noteworthy that the phytolith production pattern of northwestAustralian flora studied by Wallis (2003) revealed non production ofphytoliths in Acanthaceae, Asteraceae, Sterculiaceae and Pandanaceae.However, interestingly phytolith morphopypes were found in abun-dance in some studied plant taxa belonging to these families inhabitingthe swampy mangrove habitats of the Sundarbans. So it is assumedthat environmental factors may play an important role in phytolithproduction.
We have also considered the frequency of occurrence of differentphytolith morphotypes in different plant parts, which may help under-standing the difference in production pattern of phytoliths in the leaves,fruits, seeds, flower, spathe and pneumatophores of the plant taxa be-longing to different taxonomic groups. These differences may be dueto variation in morphology in different parts of a plant or depend onthe complex process of silica uptake and deposition by plants. These dif-ferential productions of phytoliths in different body parts of a plant alsoinfluence their subsequent dispersion in sediment after decaying of thetissue of their origin. Studying the reason behind this bias is not withinthe objectives of this study and will be discussed elsewhere.
Plants experience oxidative stress upon exposure to heavy metalsand lead to cellular damage. However, a number of experimentalstudies indicate that the addition of silicon in growth media mitigatesthe damaging effects of aluminum and manganese on plant growth(Epstein, 1994; Sangster et al., 2001). A co-deposition of solid silicaand aluminum in and around plant cells has been demonstrated fora wide variety of species and plant organs; hence, the mechanismfor detoxification may be a sequestration of aluminum by the silica(Sangster and Hodson, 2001; Sangster et al., 2001). Mangrove sedi-ments have a high capacity for absorbing and holding heavy metalsthereby preventing the spread of metal pollution in coastal areas.Silicon dioxide may ameliorate the toxic effects of aluminum andother heavy metals, such as manganese, which are ingested by plantsalong with other substances in the ground water (Hodson et al., 1997;Sangster et al., 2001). Therefore, it can be assumed that presence ofthis biogenic silica (phytoliths) in different parts of themangrove plantsmay help protect these plants to some extent by ameliorating the toxiceffects of these heavy metals.
5. Conclusions
The objectives of this study were to check the taxonomic reliabil-ity of the non-grass phytoliths of this unique deltaic environment andto test their efficacy in identifying different eco-phytosuccessionalhabitats resulting from differences in salinity in the deltaic regionof the Sunderbans. Earlier in a similar study Ghosh et al., 2011have used grass phytoliths as a tool to discriminate different deltaicsub-environments and established its future potential in deltaicpalaeoenvironment reconstructions.
Our study provides the first quantitative taxonomy of non-grassphytoliths from the deltaic mangrove environment of the Indian partof the Sunderbans and reports some new phytolith morphotypesfrom mangrove plant communities of this region. We have seen thatalthough these non-grass phytolith morphotypes possess a low taxo-nomic value but reflect a strong signal of change in salinity. It hasbeen noticed that differing tidal activities and changes in salinity gra-dient influence the community structure and distribution of plants inthe deltaic plains of the Sunderbans. Phytolith compositions of plantshave found to vary as observed by the variation in morphotypes alongsalinity gradient. It may be said that different types of globularmorphotypes and blocky polyhedral types if found together in any fos-sil sediment from this unique deltaic region may identify a swampyand tidal mangrove depositional environment. On the other hand,

Fig. 8. A hypothetical transect of deltaic sub-environments in the Indian part of the Sunderbans showing major vegetation zones and their representative phytolith assemblages.Bars indicate relative presence of each phytolith morphotype (modified after Sen and Banerjee, 1990 and Ghosh et al., 2011).
64 S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
phytolith types i.e. papillae base around stomata (PAP) diagnostic ofPandanus fascicularis leaves may identify a depositional environmentfor non- littoral species. In addition, multicellular hair cells (MULTHAIR) as observed in Grangea maderaspatana if found frequently inany fossil phytolith assemblage, may suggest a dry xerophytic envi-ronment of deposition in that region. EDX data also indicates the pres-ence of silicon dioxide along with aluminium, magnesium, calcium,sodium, potassium, iron and organic carbon to authenticate chemicalcomposition of some newly described forms of phytoliths.
This paper advocates for the role of non-grass phytoliths in envi-ronmental reconstruction and put forward a model, which may helpin future phytolith studies concerning fossil sediments from similardeltaic environments (Fig. 8).
Acknowledgements
The authors (SD and SB) would like to thank UGC-RFSMS forfinancial assistance. RG thankfully acknowledges the Director, BirbalSahni Institute of Palaeobotany, Lucknow for his encouragementand permission to publish this work. The authors would also like toacknowledge the authorities of Geological Survey of India (GSI) forSEM photographs and EDX analyses of the phytoliths studied. Thanks
are also due to Mr. Dipak Kumar Paruya for his help and untiringeffort during the collection of the studied plant taxa. We also thanktwo anonymous reviewers and the editor for their valuable sugges-tions to improve the quality of the article.
References
Ball, T.B., Gardner, J.S., Anderson, N., 1999. Identyfying inflorescence phytoliths fromselected species of wheat (Triticum monococcum, T. dicoccon, T. dicoccoides andT. aestivum) and barley (Hordeum vulgare and H. spontaneum) (Gramineae).American Journal of Botany 86, 1615–1623.
Basilevsky, A., 1994. Statistical Factor Analysis and Related Methods. Wiley- Interscience,New York.
Bozarth, S., 1993. Biosilicate assemblages of boreal forests and aspen parklands. In:Pearsall, D.M., Piperno, D.R. (Eds.), Current Research in Phytoliths Analysis:Applications in Archeology and Palaeoecology, vol. 10. University of Pennsylvania,Philadelphia, pp. 95–105.
Carnelli, A.L., Theurillat, J.-P., Madella, M., 2004. Phytolith types and type-frequencies insubalpine-alpine plant species of the European Alps. Review of Palaeobotany andPalynology 129, 39–65.
Chauhan, D.K., Tripathi, D.K., Kumar, Dharmendra, Kumar, Yashwant, 2011. Diversity,distribution and frequency based attributes of phytolith in Arundo donax L. Interna-tional Journal of Innovations in Biological and Chemical Sciences 1, 22–27.
Epstein, E., 1994. The anomaly of silicon in plant biology. Proceedings of the NationalAcademy of Sciences of the United States of America 91, 11–17.
Gallego, L., Distel, R.A., 2004. Phytolith assemblages in grasses native to Central Argentina.Annals of Botany 94, 865–874.

65S. Das et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 376 (2013) 48–65
Ghosh, R., Naskar, M., Bera, S., 2011. Phytolith assemblages of grasses from theSunderbans, India and their implications for the reconstruction of deltaic environ-ments. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 93–102.
Hodson, M.J., Williams, S.E., Sangster, A.G., 1997. Silica deposition in the needles of thegymnosperms. Ι. Chemical analysis and light microscopy. In: Pinilla, A., Juan-Tresserras, J., Machado, M. (Eds.), The State of the Art of Phytoliths in Soils andPlants.
Honaine, M., Zucol, A.F., Osterrieth, M., 2006. Phytolith assemblages and systematic as-sociations in grassland species of the south-eastern Pampean Plains, Argentina.Annals of Botany 98, 1155–1165.
Iriarte, J., Paz, E.A., 2009. Phytolith analysis of selected native plants and modern soilsfrom southeastern Uruguay and its implications for paleoenvironmental andarcheological reconstruction. Quaternary International 193, 99–123.
Jolliffe, I.T., 2002. Principal Component Analysis. Springer- Verlag, New York.Jones, L.H.P., Milne, A.A., 1963. Studies of silica in the oat plant:I. Chemical and physical
properties of silica. Plant and Soil 18, 207–220.Joshi, H., Ghose, M., 2003. Forest structure and species distribution along soil salinity
and pH gradient in mangrove swamps of the Sundarbans. Tropical Ecology 44(2), 197–206.
Kim, J., Mueller, C.W., 1978. Factor Analysis: Statistical Methods and Practical Issues.Sage Publications, Newbury Park.
Krishnan, S., 2000. Phytoliths of Indian grasses and their potential use in identification.Botanical Journal of the Linnean Society 132, 241–252.
Lu, H.Y., Liu, K.B., 2003. Morphological variations in lobate phytoliths from grasses inChina and the southeastern USA. Diversity and Distributions 9 (1), 73–87.
Madella, M., Alexander, A., Ball, T., 2005. International code for phytolith nomenclature1.0. Annals of Botany 96, 253–260.
Marx, R., Lee, D.E., Lloyd, K.M., Lee, W., 2004. Phytolith morphology and biogenic silicaconcentrations and abundance in leaves of Chionochloa (Danthonieae) and Festuca(Poaceae) in New Zealand. New Zealand Journal of Botany 42, 677–691.
Mazumdar, J., Mukhopadhyay, R., 2009a. Opal phytoliths in Three Indian ThelypteroidFerns. Bionature 29 (1), 11–15.
Mazumdar, J., Mukhopadhyay, R., 2009b. Phytoliths of some lycopods. Indian Fern Journal26, 132–136.
Mazumdar, J., Mukhopadhyay, R., 2011. Phytoliths of ferns IV: In some aquatic fernsand Chinese brake fern. Bioresearch Bulletin 6, 385–388.
Mercader, J., Bennett, T., Esselmont, C., Simpson, C., Walde, D., 2009. Phytoliths inwoody plants from the Miombo woodlands of Mozambique. Annals of Botany1–23.
Mercader, J., Astudillo, F., Barkworth, M., Bennett, T., Esselmont, C., Kinyanjui, R.,Grossman, D.L., Simpson, S., Walde, D., 2010. Poaceae phytoliths from the NiassaRift, Mozambique. Journal of Archaeological Science 37, 1953–1967.
Naskar, K.R., Guha Bakshi, D.N., 1987. Mangrove swamps of the Sunderbans (An Eco-logical Perspective). Naya Prokash, India.
Parr, J.F., Dolic, V., Lancaster, G., Boyd, W.E., 2001. A microwave digestion method forthe extraction of phytoliths from herbarium specimens. Review of Palaeobotanyand Palynology 116, 203–212.
Pearsall, D.M., 1989. Paleoethnobotany: A Handbook of Procedures. Academic Press,San Diego.
Piperno, D.R., 1988. Phytolith Analysis: An Archaeological and Geological Perspective.Academic Press, San Diego, p. 280.
Prychid, C.J., Rudall, P.J., Gregory, M., 2004. Systematics and biology of silica bodies inmonocotyledons. The Botanical Review 69, 377–440.
Sangster, A.G., Hodson, M.J., 2001. Silicon and aluminium codeposition in the cell wallof gymnosperm leaves. In: Meunier, J.D., Colin, F. (Eds.), Phytoliths: Applicationin Earth Sciences and Human History. A.A. Balkema Publishers, Lisse, Netherlands,pp. 343–355.
Sangster, A.G., Hodson, M.J., Tubb, H.J., 2001. Silicon deposition in higher plants. In:Datnoff, L.E., Snyder, G.H., Korndorfer, G.H. (Eds.), Silicon in Agriculture. Elsivier,Amsterdam, pp. 85–113.
Sen, P.K., Banerjee, M., 1990. Palyno-plankton stratigraphy and environmental changesduring the Holocene in the Bengal Basin, India. Review of Palaeobotany and Palynol-ogy 65, 25–35.
Tripathi, D.K., Mishra, S., Chauhan, D.K., Tiwari, S.P., Kumar, C., 2012. Typological andfrequency based study of opaline silica (phytolith) deposition in two commonIndian Sorghum L. species. Proceedings of the National Academy of Sciences,India Section B: Biological Sciences. http://dx.doi.org/10.1007/s40011-012-0066-5.
Wallis, L., 2003. An overview of leaf phytolith production patterns in selected north-west Australian flora. Review of Palaeobotany and Palynology 125, 201–248.
Zucol, A.F., 1998. Microfitolitos de las Poaceae Argentinas: II. Microfitolitos foliares dealgunas especies del genero Panicum (Poaceae, Paniceae) de la Provincia de EntreRios. Darwiniana 36, 29–50.
Zucol, A.F., 2000. Fitolitos de Poaceae de Argentina. III. Fitolitos foliares de especies delgenero Paspalum (Paniceae) en la Povincia de Entre Rios. Darwiniana 38, 11–32.





























