Embed Size (px)
Citation preview

M I N I R E V I E W
Anupdate onPseudomonasaeruginosa bio¢lmformation,tolerance, and dispersalMorten Harmsen1, Liang Yang2, Sunje J. Pamp3 & Tim Tolker-Nielsen1
1Department of International Health, Immunology and Microbiology, Faculty of Health Sciences, University of Copenhagen, Copenhagen, Denmark;2Department of Systems Biology, Technical University of Denmark, Lyngby, Denmark; and 3Department of Microbiology and Immunology, Stanford
University, School of Medicine, Palo Alto, CA, USA
Correspondence: Tim Tolker-Nielsen,
Department of International Health,
Immunology and Microbiology, Faculty of
Health Sciences, University of Copenhagen,
Blegdamsvej 3B, DK-2200 Copenhagen N,
Denmark. Tel.: 145 353 26 656; fax: 145
353 27 853; e-mail: [email protected]
Received 5 January 2010; revised 16 March
2010; accepted 21 April 2010.
Final version published online 18 May 2010.
DOI:10.1111/j.1574-695X.2010.00690.x
Editor: Gianfranco Donelli
Keywords
biofilm formation; tolerance; dispersal;
Pseudomonas aeruginosa.
Abstract
We review the recent advances in the understanding of the Pseudomonas aeruginosa
biofilm lifestyle from studies using in vitro laboratory setups such as flow chambers
and microtiter trays. Recent work sheds light on the role of nutrients, motility, and
quorum sensing in structure formation in P. aeruginosa biofilms. The second
messenger, c-di-GMP, is established as an important regulator of the synthesis of
polysaccharide and protein components of the biofilm matrix. Extracellular DNA
is shown to be an essential component of the biofilm matrix. It has become
apparent that biofilm formation involves interactions between different subpopu-
lations. The molecular mechanisms underlying the tolerance of biofilm bacteria to
antimicrobial agents are beginning to be unraveled, and new knowledge has been
obtained regarding the environmental cues and regulatory mechanisms involved
in biofilm dispersal.
Introduction
Microbial biofilms have been subject to intense study during
the last decade mainly for two reasons. First, it is of basic
scientific interest to understand how bacteria form and live
in multicellular communities. Second, biofilm formation
causes considerable problems in medical and industrial
settings, because bacteria in biofilms can resist antibiotic
treatment, host immune responses, and biocide treatment.
Knowledge of the environmental cues, genetic elements, and
molecular mechanisms that are involved in biofilm forma-
tion is necessary for a rational design of strategies to
eliminate biofilms or to prevent biofilm formation.
A substantial part of the studies of microbial biofilms
conducted during the last decade has involved in vitro
laboratory setups such as microtiter trays and flow cham-
bers. Although the model biofilms grown in these setups
most likely differ from biofilms formed in nature, experi-
ments with these biofilms have provided important insights
regarding the fundamental processes of biofilm formation,
tolerance development, and biofilm dispersal that may be of
relevance outside the laboratory systems. In the present
review, we present an update on biofilm formation, toler-
ance, and dispersal based on in vitro studies with the
opportunistic pathogen Pseudomonas aeruginosa, which is a
model organism for biofilm research. Of the numerous
factors involved in the biofilm formation process, we
presently focus on attachment, motility, matrix production,
quorum sensing, and subpopulation interactions.
Attachment
Transport of P. aeruginosa bacteria to a surface before
attachment is assumed to involve diffusive, convective, and
active flagellum-driven transport (van Loosdrecht et al.,
1990). A variety of components including flagella (O’Toole
& Kolter, 1998; Sauer et al., 2002), type IV pili (O’Toole &
Kolter, 1998; Deziel et al., 2001; Chiang & Burrows, 2003),
Cup fimbria (Vallet et al., 2001), extracellular DNA (eDNA)
(Whitchurch et al., 2002), and Psl polysaccharide (Ma et al.,
2009) have been shown to play a role in the attachment of P.
aeruginosa to surfaces in microtiter trays and flow chambers.
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
IMM
UN
OLO
GY
& M
EDIC
AL
MIC
ROBI
OLO
GY

However, conditions under which lack of flagella and/or
type IV pili did not affect surface attachment in flow
chambers have also been reported (Klausen et al., 2003b).
After P. aeruginosa has attached to a surface, it either
detaches, remains attached at the position of attachment,
or moves along the surface by means of type IV pili or
flagella (Singh et al., 2002; Klausen et al., 2003a, b; Singh,
2004; Shrout et al., 2006). The gene sadB has been shown to
be involved in the regulation of the frequency by which P.
aeruginosa cells attach to and detach from surfaces (Caiazza
& O’Toole, 2004). Synthesis of the attachment regulator
SadB is regulated by the intracellular level of the second
messenger molecule cyclic diguanosine-50-monophosphate
(c-di-GMP) (Merritt et al., 2007).
Motility
Experiments in flow-chamber setups have suggested that
surface-associated motility is an integrated part of P. aerugi-
nosa biofilm formation (e.g. Singh et al., 2002; Klausen et al.,
2003a, b; Singh, 2004; Shrout et al., 2006; Patriquin et al.,
2008). The pattern of motility occurring in P. aeruginosa
biofilms appears to be dependent on the prevailing condi-
tions. With glucose as a carbon source, for example, the
attached bacteria differentiate initially into a nonmotile
subpopulation and a motile subpopulation (Klausen et al.,
2003a). The nonmotile subpopulation forms microcolonies
that serve as the ‘stalks’ of ‘mushroom’-shaped multicellular
structures that are formed when bacteria from the motile
subpopulation colonize the stalks and subsequently form
mushroom ‘caps’ upon the stalks (Klausen et al., 2003a).
With citrate as a carbon source, the entire bacterial popula-
tion is motile in the initial phase of biofilm formation and
microcolonies that could serve as mushroom stalks are not
formed, and consequently, a flat biofilm is formed (Klausen
et al., 2003b). Pseudomonas aeruginosa pilA mutants (defi-
cient in the biogenesis of type IV pili) formed protruding
microcolonies in biofilms irrigated with a citrate medium,
indicating that the motility occurring in the initial phase of
biofilm formation in citrate-grown P. aeruginosa wild-type
biofilms is driven by type IV pili and thus most likely is
similar to twitching motility (Klausen et al., 2003b). As
described below, P. aeruginosa twitching motility is stimu-
lated by iron limitation. When citrate is used as a carbon
source, it is present at a much higher concentration than
iron in the medium. Because citrate chelates iron, it is
possible that the use of citrate as a carbon source in flow-
chamber systems reduces the level of iron available to the
bacteria, which may explain why the entire population is
motile initially in P. aeruginosa biofilms irrigated with citrate
medium.
Twitching motility has been shown to be induced in
P. aeruginosa biofilms by iron limitation (Singh et al., 2002;
Singh, 2004; Patriquin et al., 2008). In accordance, P.
aeruginosa wild-type bacteria, which formed mushroom-
shaped structures in flow cells irrigated with iron-replete
medium, formed flat biofilms in flow chambers irrigated
with an iron-depleted medium (Patriquin et al., 2008).
Evidence was provided that iron-limitation-promoted
twitching motility is dependent on a functional Rhl quor-
um-sensing system, and a P. aeruginosa rhlI mutant was
shown to form microcolonies (lacking mushroom caps) in
flow chambers irrigated with an iron-depleted medium
(Patriquin et al., 2008). A role of the Rhl quorum-sensing
system in an iron-limitation-induced phenotype is in accor-
dance with numerous studies showing that the quorum-
sensing systems in P. aeruginosa are induced under iron
limitation, but repressed in the presence of high levels of
iron (e.g. Bollinger et al., 2001; Kim et al., 2005; Duan &
Surette, 2007). Spent medium from iron-limited, but not
iron-replete, P. aeruginosa wild-type cultures could induce
iron-limitation-promoted twitching motility of P. aerugino-
sa rhlI mutants, suggesting that some soluble factor, which is
low-iron-inducible and RlhI dependent, positively influ-
enced twitching motility (Patriquin et al., 2008). The avail-
able evidence suggested that this soluble factor is the
biosurfactant rhamnolipid, but it was not investigated
whether spent medium from P. aeruginosa rhlA cultures
(deficient in the biogenesis of rhamnolipid) failed to induce
iron-limitation-promoted twitching motility of P. aerugino-
sa rhlI mutants (Patriquin et al., 2008). In agreement with
the suggestion that increased levels of rhamnolipid may
stimulate twitching motility in iron-limited biofilms, Pamp
& Tolker-Nielsen (2007) reported that in addition to the
well-known role of rhamnolipid in facilitating swarming
motility (e.g. Kohler et al., 2000), the biosurfactant can also
facilitate twitching motility. If increased levels of rhamnoli-
pid produced in iron-deplete P. aeruginosa biofilms stimu-
late twitching motility and thereby prevent microcolony
formation, it would be expected that a P. aeruginosa rhlA
mutant would be able to form microcolonies under iron-
limited conditions. However, Pamp & Tolker-Nielsen (2007)
provided evidence that rhamnolipid is necessary for initial
microcolony formation in P. aeruginosa biofilms. In support
of a role of rhamnolipid in initial microcolony formation, it
has been shown that low concentrations of rhamnolipid
enhance the hydrophobicity of P. aeruginosa bacteria by
causing a release of lipopolysaccharide (LPS) from the cell
surface (Zhang & Miller, 1994; Al-Tahhan et al., 2000). An
increase in cell surface hydrophobicity may increase the
adhesiveness of the bacteria to a level that is critical for
initial microcolony formation in biofilms. It appears that
rhamnolipid, depending on the quantity produced, plays
multiple roles in P. aeruginosa biofilm formation. It is
necessary for initial microcolony formation (Pamp & Tolk-
er-Nielsen, 2007); it may prevent initial microcolony
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
254 M. Harmsen et al.

formation in iron-limited biofilms by stimulating twitching
motility (Patriquin et al., 2008); it facilitates bacterial
migration and thereby the formation of mushroom caps
(Pamp & Tolker-Nielsen, 2007); it prevents colonization of
the channels between the mushroom-shaped structures
(Davey et al., 2003); and it plays a role in biofilm dispersal
(Schooling et al., 2004; Boles et al., 2005).
Evidence has been presented that the formation of the
mushroom caps in P. aeruginosa biofilms requires both type
IV pili and flagella (Klausen et al., 2003a; Barken et al.,
2008). Cap formation evidently requires flagellum-driven
surface-associated motility, whereas the dependence of cap
formation on type IV pili may be due to the binding of these
pili to eDNA that is abundant on the microcolonies that
become colonized during mushroom-structure formation
(Whitchurch et al., 2002; Allesen-Holm et al., 2006; Barken
et al., 2008). Alternatively, a kind of migration that requires
both flagella and type IV pili might be occurring in the later
phase of P. aeruginosa biofilm formation. In support of this
suggestion, it has been reported that P. aeruginosa swarming
motility in addition to flagella and rhamnolipid may also
require type IV pili under some conditions (Kohler et al.,
2000).
As described in the next section, evidence is accruing that
the shift between migrating and sessile surface behavior is
regulated via proteins with diguanylate cyclase or phospho-
diesterase activities that control levels of the second messen-
ger molecule c-di-GMP (Hickman et al., 2005; Kuchma
et al., 2007; Merritt et al., 2007). Low intracellular levels of
c-di-GMP promote motility, whereas high intracellular c-
di-GMP levels induce the formation of cell–cell intercon-
necting matrix components and promote microcolony
formation.
Matrix production
The extracellular polymeric substance (EPS) matrix serves as
the ‘house for biofilm cells’. The matrix plays a role in
numerous processes including attachment, cell-to-cell inter-
connection, interactions between subpopulations, tolerance,
and exchange of genetic material (Molin & Tolker-Nielsen,
2003; Friedman & Kolter, 2004a; Jackson et al., 2004; Ma
et al., 2009; Yang et al., 2009). The P. aeruginosa EPS matrix
contains mainly polysaccharides, proteins, and nucleic acids
(e.g. Whitchurch et al., 2002; Friedman & Kolter, 2004a;
Jackson et al., 2004; Borlee et al., 2010). The composition of
the matrix depends on the environmental conditions, the
age of the biofilm, and the particular P. aeruginosa strain
forming the biofilm. Evidence has been provided that
P. aeruginosa induces the synthesis of matrix components
in response to environmental signals sensed by the sensor
kinase/response regulators LadS, RetS, and GacS (Goodman
et al., 2004; Ventre et al., 2006; Goodman et al., 2009).
In addition, sensor kinases and response regulators encoded
by the bfiSR, bfmSR, and mifSR genes are evidently involved
in further P. aeruginosa biofilm formation (Petrova & Sauer,
2009), although the functions affected by these signal
transduction systems have not been identified. Below, we
describe the involvement of the matrix components Pel, Psl,
alginate, CdrA protein, Cup fimbria, type IV pili, lectins,
and eDNA in P. aeruginosa biofilm formation.
The glucose-rich Pel polysaccharide, encoded by the pel
cluster, was initially reported to be required for the forma-
tion of P. aeruginosa liquid–air interface pellicles and sur-
face-associated biofilms (Friedman & Kolter, 2004b).
Evidence has been presented that the transmembrane pro-
tein PelD binds c-di-GMP, and that there is a strict correla-
tion between c-di-GMP binding and the synthesis of the Pel
polysaccharide (Lee et al., 2007; Lory et al., 2009). Hickman
& Harwood (2008) presented data showing that c-di-GMP
binds to the transcriptional regulator FleQ and thereby
derepresses the transcription of the pel genes. The pel locus
is furthermore regulated by the sensor kinase-response
regulator hybrids RetS and LadS, which repress and activate
its expression, respectively (Goodman et al., 2004; Ventre
et al., 2006). This regulation is mediated through the small-
RNAs, RsmZ/Y, which alter the level of the free RsmA post-
transcriptional regulator (Goodman et al., 2004; Ventre
et al., 2006). The two-component system GacA/S interacts
with the RetS and LadS regulatory network, and recent data
suggest that RetS can modulate the phosphorylation state of
GacS by forming heterodimers that block the GacS autop-
hosphorylation, leading to a reduced rsmZ expression, and
subsequently free RsmA, thereby repressing exopolysacchar-
ide synthesis and promoting the translation of genes neces-
sary for acute infection (Goodman et al., 2009). Recent work
showed that rugose small-colony variants (RSCVs) of
P. aeruginosa isolated from laboratory biofilms and cystic
fibrosis (CF) patients had increased expression of the pel
locus compared with clonally related wild types, and it was
suggested that RSCVs may partially contribute to increased
persistence of biofilms in the airways of CF lungs (Starkey
et al., 2009). It was further shown that c-di-GMP signaling
regulated the increased expression of the pel locus in the
RSCVs (Starkey et al., 2009).
The mannose-rich Psl polysaccharide, encoded by the psl
cluster, is highly conserved in many P. aeruginosa strains
(Friedman & Kolter, 2004a; Ryder et al., 2007). The Psl
polysaccharide mediates cell-to-surface and cell-to-cell
interactions, which are essential for P. aeruginosa biofilm
formation and maintenance (Overhage et al., 2005; Ma et al.,
2006, 2009). In a recent study, Ma et al. (2009) showed that
Psl forms a helical structure around P. aeruginosa cells that
increase cell-to-surface and cell-to-cell interactions neces-
sary for biofilm formation. By lectin staining, Psl was shown
to be located predominantly in the peripheries of the
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
255P. aeruginosa biofilm formation, tolerance, and dispersal

mushroom-shaped structures, whereas it was evenly distrib-
uted in flat biofilms (Ma et al., 2009). As for the Pel
polysaccharide, Psl expression is regulated by LadS and RetS
(Goodman et al., 2004; Ventre et al., 2006) and by c-di-GMP
(Hickman et al., 2005). A newly characterized protein,
CdrA, which shows increased expression with elevated c-
di-GMP levels, is suggested to either crosslink Psl polymers
and/or tether Psl to the P. aeruginosa cell wall (Borlee et al.,
2010). This protein is the first to be identified as a structural
component of the P. aeruginosa biofilm matrix. Borlee et al.
(2010) found that the cdrAB complex is elevated in biofilms,
and that overexpression of both proteins induces auto-
aggregation if Psl is present. The second protein in the
complex, CdrB, works as an outer-membrane transporter of
CdrA. Evidence was provided that CdrA binds directly to the
mannose part of Psl, and it was suggested that CdrA is a
multivalent adhesin with the ability to recognize multiple
carbohydrates (Borlee et al., 2010).
Alginate is an acetylated polysaccharide composed of
nonrepetitive monomers of b-1,4-linked L-guluronic and
D-mannuronic acids (Govan & Deretic, 1996), and is mainly
produced by P. aeruginosa in chronic infections of the lungs
of CF patients (Hoiby, 1974; Govan & Deretic, 1996). Its
special physical and chemical properties play multiple roles
in protecting P. aeruginosa cells (Govan & Deretic, 1996).
Biofilms formed by an alginate-overproducing strain were
shown to exhibit a highly structured architecture and were
significantly more resistant to the antibiotic tobramycin
than biofilms formed by an isogenic nonmucoid strain
(Hentzer et al., 2001). This might indicate that alginate acts
as a physical barrier for antibiotics because alginate lyase
treatment has been shown to enhance the diffusion of
aminoglycosides through the EPS of biofilms formed by
mucoid P. aeruginosa strains (Hatch & Schiller, 1998). A
recent study showed that a supermucoid P. aeruginosa strain
was strongly impaired in attachment compared with the
respective mucoid or nonmucoid strains and formed a
thicker biofilm with large extended mushroom-like micro-
colonies (Hay et al., 2009a). Recently, it has been found that
post-transcriptional regulation plays a role in the biosynth-
esis of alginate in P. aeruginosa. The membrane-anchored
protein, Alg44, which is essential for alginate biosynthesis,
contains a c-di-GMP-binding PilZ domain (Amikam &
Galperin, 2006; Remminghorst & Rehm, 2006b; Merighi
et al., 2007; Oglesby et al., 2008; Lory et al., 2009). Regula-
tion of Alg44 activity via the PilZ domain has recently been
suggested to occur through a new membrane-anchored
diguanylate cyclase, MucR, which has been identified as a
positive regulator of alginate biosynthesis (Hay et al.,
2009b). It was suggested that MucR possibly acts by the
localized production of c-di-GMP in the vicinity of Alg44.
The PilZ domain of Alg44 in turn responds to the high local
concentration of c-di-GMP and promotes the production of
alginate. The glycosyltransferase Alg8 was characterized
recently and shown to be critical for alginate production
(Remminghorst & Rehm, 2006a; Oglesby et al., 2008). It was
shown that the Alg44 and Alg8 proteins were required in
combination for the polymerization reactions leading to
alginate production (Oglesby et al., 2008).
Recently, eDNA was recognized as one of the major
matrix components of bacterial biofilms (Whitchurch et al.,
2002; Qin et al., 2007; Rice et al., 2007). It was shown that
DNAse treatment led to the dispersal of young P. aeruginosa
biofilms (Whitchurch et al., 2002). However, DNAse did not
disperse a flow-chamber-grown P. aeruginosa mature bio-
film, probably due to increasing amounts of other EPS
materials being produced during biofilm formation. South-
ern and RAPD PCR analysis provided evidence that the
eDNA in the P. aeruginosa biofilm matrix is similar to
chromosomal P. aeruginosa DNA (Allesen-Holm et al.,
2006). In addition to a small amount of eDNA that is
present and necessary in the initial phase of P. aeruginosa
biofilm formation (Whitchurch et al., 2002), the release of a
large amount of eDNA occurs during subsequent P. aerugi-
nosa biofilm formation, evidently through lysis of a small
subpopulation of the bacteria regulated via the P. aeruginosa
quinolone signal (PQS) quorum-sensing system (Allesen-
Holm et al., 2006). In agreement with a role of quorum
sensing in cell lysis, D’Argenio et al. (2002) reported that
mutants that could not produce the PQS quorum-sensing
signal molecule did not show autolysis, whereas mutants
that overproduced PQS displayed high levels of autolysis. In
addition, Heurlier et al. (2005) presented evidence that P.
aeruginosa quorum-sensing mutants, unlike the wild type,
did not undergo cell lysis in stationary-phase cultures.
Moreover, Yang et al. (2007) presented evidence that high
levels of iron suppressed P. aeruginosa pqs gene expression,
DNA release, and structural biofilm development. Quino-
lone compounds have previously been shown to induce
prophages in bacteria (Phillips et al., 1987; Froshauer et al.,
1996), and studies by Webb et al. (2003) and Hentzer et al.
(2004) have suggested that quorum-sensing-regulated DNA
release might be linked to bacteriophage induction in
biofilms. PQS was shown to be necessary for membrane
vesicle formation in P. aeruginosa (Mashburn & Whiteley,
2005; Nakamura et al., 2008), and membrane vesicles
produced by P. aeruginosa might also play a role in DNA
release. The membrane vesicles released by P. aeruginosa
have bacteriolytic effects and contain DNA (Kadurugamuwa
& Beveridge, 1996; Renelli et al., 2004). eDNA might be
released either from vesicles that eventually lyse or through
the bacteriolytic activity of the vesicles that might lyse a
small subpopulation of P. aeruginosa bacteria. The eDNA
appears to be organized in distinct patterns in P. aeruginosa
biofilms (Allesen-Holm et al., 2006). In flow-chamber-
grown P. aeruginosa biofilms, which contain mushroom-
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
256 M. Harmsen et al.
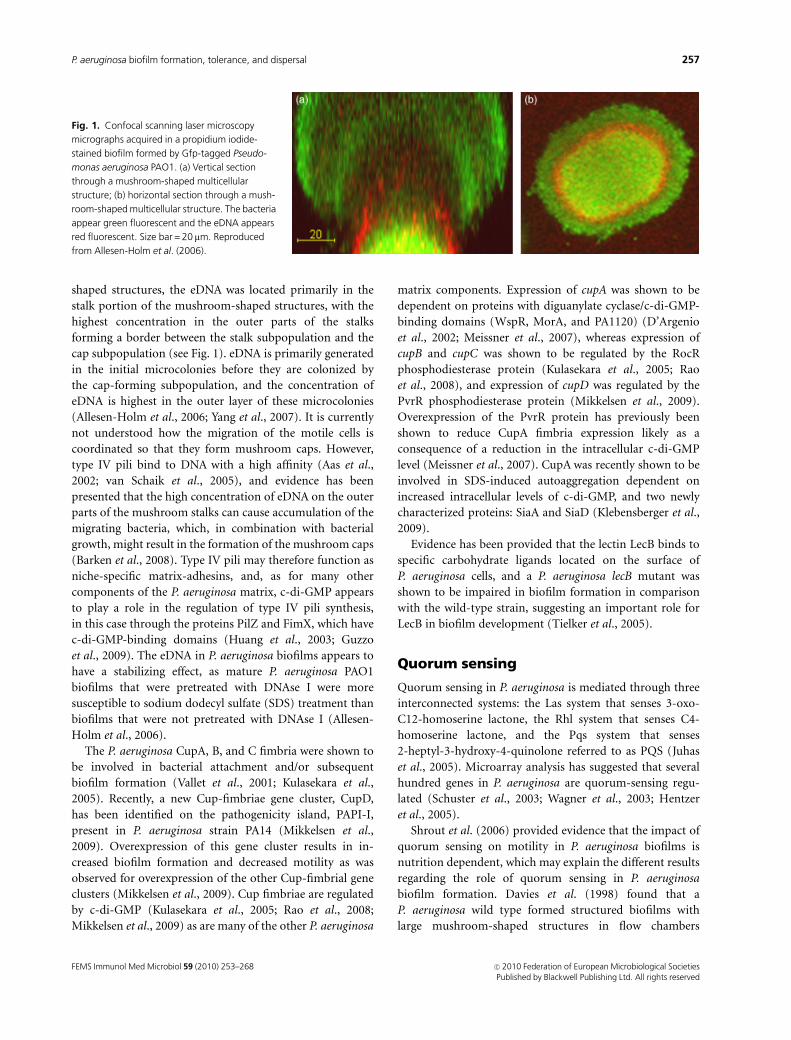
shaped structures, the eDNA was located primarily in the
stalk portion of the mushroom-shaped structures, with the
highest concentration in the outer parts of the stalks
forming a border between the stalk subpopulation and the
cap subpopulation (see Fig. 1). eDNA is primarily generated
in the initial microcolonies before they are colonized by
the cap-forming subpopulation, and the concentration of
eDNA is highest in the outer layer of these microcolonies
(Allesen-Holm et al., 2006; Yang et al., 2007). It is currently
not understood how the migration of the motile cells is
coordinated so that they form mushroom caps. However,
type IV pili bind to DNA with a high affinity (Aas et al.,
2002; van Schaik et al., 2005), and evidence has been
presented that the high concentration of eDNA on the outer
parts of the mushroom stalks can cause accumulation of the
migrating bacteria, which, in combination with bacterial
growth, might result in the formation of the mushroom caps
(Barken et al., 2008). Type IV pili may therefore function as
niche-specific matrix-adhesins, and, as for many other
components of the P. aeruginosa matrix, c-di-GMP appears
to play a role in the regulation of type IV pili synthesis,
in this case through the proteins PilZ and FimX, which have
c-di-GMP-binding domains (Huang et al., 2003; Guzzo
et al., 2009). The eDNA in P. aeruginosa biofilms appears to
have a stabilizing effect, as mature P. aeruginosa PAO1
biofilms that were pretreated with DNAse I were more
susceptible to sodium dodecyl sulfate (SDS) treatment than
biofilms that were not pretreated with DNAse I (Allesen-
Holm et al., 2006).
The P. aeruginosa CupA, B, and C fimbria were shown to
be involved in bacterial attachment and/or subsequent
biofilm formation (Vallet et al., 2001; Kulasekara et al.,
2005). Recently, a new Cup-fimbriae gene cluster, CupD,
has been identified on the pathogenicity island, PAPI-I,
present in P. aeruginosa strain PA14 (Mikkelsen et al.,
2009). Overexpression of this gene cluster results in in-
creased biofilm formation and decreased motility as was
observed for overexpression of the other Cup-fimbrial gene
clusters (Mikkelsen et al., 2009). Cup fimbriae are regulated
by c-di-GMP (Kulasekara et al., 2005; Rao et al., 2008;
Mikkelsen et al., 2009) as are many of the other P. aeruginosa
matrix components. Expression of cupA was shown to be
dependent on proteins with diguanylate cyclase/c-di-GMP-
binding domains (WspR, MorA, and PA1120) (D’Argenio
et al., 2002; Meissner et al., 2007), whereas expression of
cupB and cupC was shown to be regulated by the RocR
phosphodiesterase protein (Kulasekara et al., 2005; Rao
et al., 2008), and expression of cupD was regulated by the
PvrR phosphodiesterase protein (Mikkelsen et al., 2009).
Overexpression of the PvrR protein has previously been
shown to reduce CupA fimbria expression likely as a
consequence of a reduction in the intracellular c-di-GMP
level (Meissner et al., 2007). CupA was recently shown to be
involved in SDS-induced autoaggregation dependent on
increased intracellular levels of c-di-GMP, and two newly
characterized proteins: SiaA and SiaD (Klebensberger et al.,
2009).
Evidence has been provided that the lectin LecB binds to
specific carbohydrate ligands located on the surface of
P. aeruginosa cells, and a P. aeruginosa lecB mutant was
shown to be impaired in biofilm formation in comparison
with the wild-type strain, suggesting an important role for
LecB in biofilm development (Tielker et al., 2005).
Quorum sensing
Quorum sensing in P. aeruginosa is mediated through three
interconnected systems: the Las system that senses 3-oxo-
C12-homoserine lactone, the Rhl system that senses C4-
homoserine lactone, and the Pqs system that senses
2-heptyl-3-hydroxy-4-quinolone referred to as PQS (Juhas
et al., 2005). Microarray analysis has suggested that several
hundred genes in P. aeruginosa are quorum-sensing regu-
lated (Schuster et al., 2003; Wagner et al., 2003; Hentzer
et al., 2005).
Shrout et al. (2006) provided evidence that the impact of
quorum sensing on motility in P. aeruginosa biofilms is
nutrition dependent, which may explain the different results
regarding the role of quorum sensing in P. aeruginosa
biofilm formation. Davies et al. (1998) found that a
P. aeruginosa wild type formed structured biofilms with
large mushroom-shaped structures in flow chambers
Fig. 1. Confocal scanning laser microscopy
micrographs acquired in a propidium iodide-
stained biofilm formed by Gfp-tagged Pseudo-
monas aeruginosa PAO1. (a) Vertical section
through a mushroom-shaped multicellular
structure; (b) horizontal section through a mush-
room-shaped multicellular structure. The bacteria
appear green fluorescent and the eDNA appears
red fluorescent. Size bar = 20 mm. Reproduced
from Allesen-Holm et al. (2006).
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
257P. aeruginosa biofilm formation, tolerance, and dispersal

irrigated with a glucose medium, while the corresponding
lasI quorum-sensing mutant formed flat and undifferen-
tiated biofilms. The flat biofilms formed by the lasI mutant
were susceptible to treatment with the detergent SDS, while
the structured biofilms formed by the wild type were
tolerant. Heydorn et al. (2002) reported that a P. aeruginosa
wild type and lasI mutant both formed flat biofilms in flow
chambers irrigated with citrate medium. Purevdorj et al.
(2002) showed that in flow chambers irrigated with dilute
Luria–Bertani (LB) under high-flow conditions, both the P.
aeruginosa wild type and the lasI mutant formed biofilms
containing large aggregates, although the biofilms differed
slightly in microscopic appearance. Hentzer et al. (2002)
demonstrated that homoserine lactone signal analogues
called furanones, known to inhibit P. aeruginosa quorum
sensing, affected biofilm development when added to the
growth medium consisting of diluted LB. Patriquin et al.
(2008) showed that the P. aeruginosa wild type formed
mushroom-shaped structures in flow chambers irrigated
with dilute tryptic soy broth, whereas an rhlI quorum-
sensing mutant formed microcolonies lacking the mush-
room caps. Similarly, Yang et al. (2009) showed that the P.
aeruginosa wild type formed mushroom-shaped structures
in flow chambers irrigated with glucose medium, whereas a
pqsA quorum-sensing mutant could only form small micro-
colonies lacking the mushroom cap. In addition, evidence
was provided that chemical inhibition of the Pqs system
caused P. aeruginosa to form microcolonies on which mush-
room caps were not subsequently formed (Yang et al., 2007,
2009).
The above-mentioned studies indicate that quorum sen-
sing is necessary for the formation of the cap portion of the
mushroom-shaped structures in P. aeruginosa biofilms.
Interestingly, quorum sensing is, however, shown to occur
mainly in the stalk portion of the mushroom-shaped
structures in P. aeruginosa biofilms (de Kievit et al., 2001;
Yang et al., 2009). Pseudomonas aeruginosa uses quorum
sensing to regulate numerous factors including the produc-
tion of rhamnolipid (Ochsner & Reiser, 1995) and eDNA
(Allesen-Holm et al., 2006). As mentioned in the previous
section, Pamp & Tolker-Nielsen (2007) provided evidence
that rhamnolipid plays a role in mushroom cap formation
by promoting motility occurring in the later phase of P.
aeruginosa biofilm formation. As described in more detail in
the following section, evidence was recently presented that
PQS quorum sensing in the initial microcolonies leads to the
production of eDNA, which plays an important role in the
subsequent formation of the mushroom caps (Allesen-
Holm et al., 2006; Barken et al., 2008; Yang et al., 2009).
In addition to regulating the production of rhamnolipid
and eDNA, quorum sensing evidently regulates a number of
other factors involved in P. aeruginosa biofilm formation.
Sakuragi & Kolter (2007) presented evidence that transcrip-
tion of the pel operon is considerably reduced in P. aerugi-
nosa lasI and rhlI mutants, suggesting that quorum-sensing
signaling regulates the production of Pel polysaccharide
during P. aeruginosa biofilm formation. On the contrary,
however, Ueda & Wood (2009) recently reported that Las
quorum-sensing represses Pel production in P. aeruginosa.
The tyrosine phosphatase TpbA was shown to be positively
regulated by Las quorum sensing, and evidence was pre-
sented that TpbA activity results in decreased levels of
c-di-GMP and thereby a reduction in Pel production (Ueda
& Wood, 2009). Transcriptome analysis has indicated that
the cupA3 and cupB5 genes are subject to quorum-sensing
control in P. aeruginosa biofilms (Hentzer et al., 2004),
suggesting that the expression of CupA and CupB fimbria
may be quorum sensing regulated during P. aeruginosa
biofilm formation. In addition, expression of the lectins
LecA and LecB was shown to be regulated via quorum
sensing (Winzer et al., 2000). The production of both lectins
was found to be directly dependent on the rhl locus, while,
in a lasR mutant, the onset of lectin synthesis was delayed,
but not abolished.
Unlike the previous examples, the last example of an
effect of quorum sensing on P. aeruginosa biofilm develop-
ment is related to the central metabolism in the bacteria.
Evidence has been presented that anaerobic nitrate respira-
tion may play an important role in P. aeruginosa biofilm
development in clinical settings, and that the rhlRI system is
necessary to prevent accumulation of toxic nitric oxide
during the process (Worlitzsch et al., 2002; Yoon et al.,
2002). In agreement, the nirCMSQ and napEF genes, which
are required for respiratory nitrate reduction, were found to
be strongly upregulated in P. aeruginosa during biofilm
growth (Hentzer et al., 2005).
Subpopulation interactions
Pseudomonas aeruginosa bacteria release several compounds
into their surrounding environment, for example iron side-
rophores, biosurfactants, and EPS. A common feature of
these compounds is that they are costly to synthesize and
able to benefit the entire population and therefore can be
regarded as ‘public goods’ (West et al., 2007). Many extra-
cellular public goods play a role in P. aeruginosa biofilm
formation. For example, the iron siderophore pyoverdine is
necessary for the formation of structured P. aeruginosa
biofilms (Banin et al., 2005). The biosurfactant rhamnolipid
plays multiple roles in P. aeruginosa biofilm formation, and
is, among other things, required for biofilm channel main-
tenance as well as for migration of motile subpopulations
(Davey et al., 2003; Pamp & Tolker-Nielsen, 2007). eDNA
released via quorum sensing plays a role as EPS material
required for the formation of structured P. aeruginosa
biofilms (Allesen-Holm et al., 2006).
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
258 M. Harmsen et al.

The bacteria in P. aeruginosa biofilms exist in various
physiological states dependent on their spatial localization.
Pseudomonas aeruginosa biofilms contain subpopulations
that produce public goods and subpopulations that do not
produce public goods. For example the pyoverdine synthesis
genes, rhamnolipid synthesis genes, and quorum-sensing
genes were all reported to be expressed specifically in the
stalk portion of the mushroom-shaped structures in P.
aeruginosa biofilms (De Kievit et al., 2001; Lequette &
Greenberg, 2005; Kaneko et al., 2007; Yang et al., 2007,
2009).
Interactions between producers and nonproducers of
public goods may play a role in P. aeruginosa biofilm
formation. Yang et al. (2009) studied subpopulation inter-
actions in mixed P. aeruginosa biofilms containing a non-
motile subpopulation (pilA mutants) and a motile
subpopulation (wild type). In this model system, the non-
motile bacteria form the stalk portion and the motile
bacteria form the cap portion of mushroom-shaped biofilm
structures. In the study, the role of pyoverdine in subpopu-
lation interactions during P. aeruginosa biofilm formation
was investigated. Using a pvdA<gfp fluorescent reporter, the
pyoverdine synthesis genes were found to be expressed
specifically in the pilA stalk-forming subpopulation of the
mushroom-shaped structures formed in pilA/wild-type
mixed biofilms. In agreement with the study of Banin et al.
(2005), it was found that a P. aeruginosa pvdA mutant could
form small microcolonies, but was unable to form mush-
room-shaped structures (see Fig. 2a), suggesting that pyo-
verdine production is necessary for mushroom structure
formation. In pilA/pvdA mixed biofilms, the pvdA mutant
was able to associate with stalks formed by the pilA mutant
and form the cap portion of the mushroom-shaped biofilm
structures (Fig. 2b), indicating that pyoverdine production
in the cap is not necessary for mushroom-structure forma-
tion. In contrast, in pilApvdA/wild-type mixed biofilms, the
wild type was not able to associate with the pilApvdA
microcolonies (Fig. 2c), suggesting that pyoverdine synth-
esis in the stalk is necessary for the cap to be formed.
Additional experiments provided evidence that even though
the cap-forming subpopulation in the mushroom-shaped
structures did not synthesize pyoverdine; it used the major
pyoverdine receptor FpvA for uptake of pyoverdine synthe-
sized by the stalk-forming subpopulation. Yang et al. (2009)
also studied the role of PQS-mediated DNA release in
subpopulation interactions during P. aeruginosa biofilm
formation. Use of a pqsA<gfp fluorescent reporter provided
evidence that the pqs genes are expressed specifically in the
pilA stalk-forming subpopulation of the mushroom-shaped
structures formed in pilA/wild-type mixed biofilms. A P.
aeruginosa pqsA mutant could form small microcolonies,
but was unable to form mushroom-shaped structures,
suggesting that pqs expression is necessary for mushroom
structure formation. In a pilA/pqsA mixed biofilm, the pqsA
mutant was able to associate with the pilA stalks and form
the caps of the mushroom-shaped structures, indicating that
pqs expression in the cap is not necessary for mushroom-
structure formation. In contrast, in a pilApqsA/wild-type
mixed biofilm, the wild type did not associate with the
pilApvdA microcolonies, but formed independent mush-
room-shaped structures, suggesting that pqs expression in
the stalk is necessary for the cap to be formed. Evidence was
presented that the stalk-forming subpopulation produces
eDNA via PQS quorum-sensing, and that the motile bacteria
bind to the eDNA using type IV pili and thereby initiate cap
formation. The PQS quorum-sensing system is probably
expressed in the stalk microcolonies in P. aeruginosa bio-
films due to the high cell concentration in this location.
Because PQS chelates iron and can thereby induce the
synthesis of pyoverdin and its receptors (Bredenbruch
Fig. 2. Confocal scanning laser microscopy (CSLM) micrographs acquired in biofilms formed by Pseudomonas aeruginosa pvdA (a), a pilA/pvdA mixture
(b), and a pilApvdA/wild-type mixture (c). The wild type and pvdA mutant were tagged with Yfp (yellow), whereas the pilA mutant and the pilApvdA
mutant were tagged with Cfp (blue). The central pictures show horizontal CLSM optical sections, and the flanking pictures show vertical CLSM optical
sections. Size bars = 20 mm. Reproduced from Yang et al. (2009).
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
259P. aeruginosa biofilm formation, tolerance, and dispersal

et al., 2006; Diggle et al., 2007), it is possible that the
observed distributed expression of the Pvd system in P.
aeruginosa biofilms is linked to the distributed expression of
the PQS system.
The study by Yang et al. (2009) is one of the first to
indicate that the formation of heterogeneous biofilms by P.
aeruginosa might occur through mechanisms that involve
complex interactions between subpopulations. The pilA,
pvdA, and pqsA mutants, which alone are deficient in the
formation of mushroom-shaped biofilm structures, are
shown in this study to interact with each other and together
build mature mushroom-shaped biofilm structures.
Tolerance
One of the most important features of microbial biofilms is
their tolerance to antibiotics and components of the host
immune system. Although antimicrobial agents may de-
crease the number of bacteria in biofilms, they will not
completely eradicate the bacteria, which may have impor-
tant clinical consequences in the form of relapses of infec-
tions. Tolerance to antimicrobial agents is a physiological
condition that does not involve mutation and allows the
bacteria to survive, but not necessarily grow, in the presence
of the antimicrobial agent. Investigations of P. aeruginosa
biofilms have revealed that biofilm tolerance is multifactor-
ial. The mechanisms that contribute to tolerance include
restricted antimicrobial diffusion, differential physiological
activity, induction of specific tolerance mechanisms, and
persister cell formation.
The available evidence suggests that biofilm matrices in
general do not inhibit diffusion of antibiotics, but penetra-
tion of some antimicrobial compounds appears to be
delayed. Penetration of ciprofloxacin and levofloxacin
through P. aeruginosa flow-chamber biofilms was found
not to be significantly delayed (Vrany et al., 1997). In
support of this, Walters et al. (2003) found that penetration
of ciprofloxacin was also not significantly delayed in a P.
aeruginosa colony biofilm; however, penetration of tobra-
mycin was somewhat retarded, but eventually penetrated the
biofilm completely. Whereas most antimicrobials might
diffuse readily through biofilms formed by wild-type P.
aeruginosa strains, it appears that alginate produced by
mucoid P. aeruginosa strains can retard the diffusion of
some antimicrobials (e.g. piperacillin, amikacin, gentami-
cin), whereas others penetrate readily (e.g. ciprofloxacin,
levofloxacin, sparfloxacin, ofloxacin) (Hoyle et al., 1992;
Shigeta et al., 1997). Recently, evidence was provided that
the activity of an antibiotic on mucoid P. aeruginosa biofilms
can be significantly enhanced by addition of alginate lyase,
and DNase, suggesting that alginate and eDNA can function
as an antibiotic barrier (Alipour et al., 2009). Cochran et al.
(2000) found that wild-type P. aeruginosa cells attached to
alginate beads were significantly less susceptible to disinfec-
tion by hydrogen peroxide than planktonic cells of the same
microorganism, although diffusion of hydrogen peroxide
was not significantly delayed, indicating that mechanisms
other than diffusion barriers contribute to the tolerance.
Biofilms contain distinct subpopulations of cells that
exhibit differential physiological states. An increasing body
of evidence suggests that the prevailing physiological states
of biofilm cell subpopulations directly relate to their sus-
ceptibility and tolerance phenotypes dependent on the
antimicrobial compound used. Evidence has been provided
that P. aeruginosa flow-chamber biofilms, as well as colony
biofilms and biofilms established in drip flow reactors, and
capillary glass tubes, are composed of at least two distinct
physiological subpopulations: a cell subpopulation close to
the substratum (e.g. the stalk portion in biofilms with
mushroom-shaped structures) that exhibits low metabolic
activity and a cell subpopulation on top (e.g. the cap portion
in biofilms with mushroom-shaped structures) that exhibits
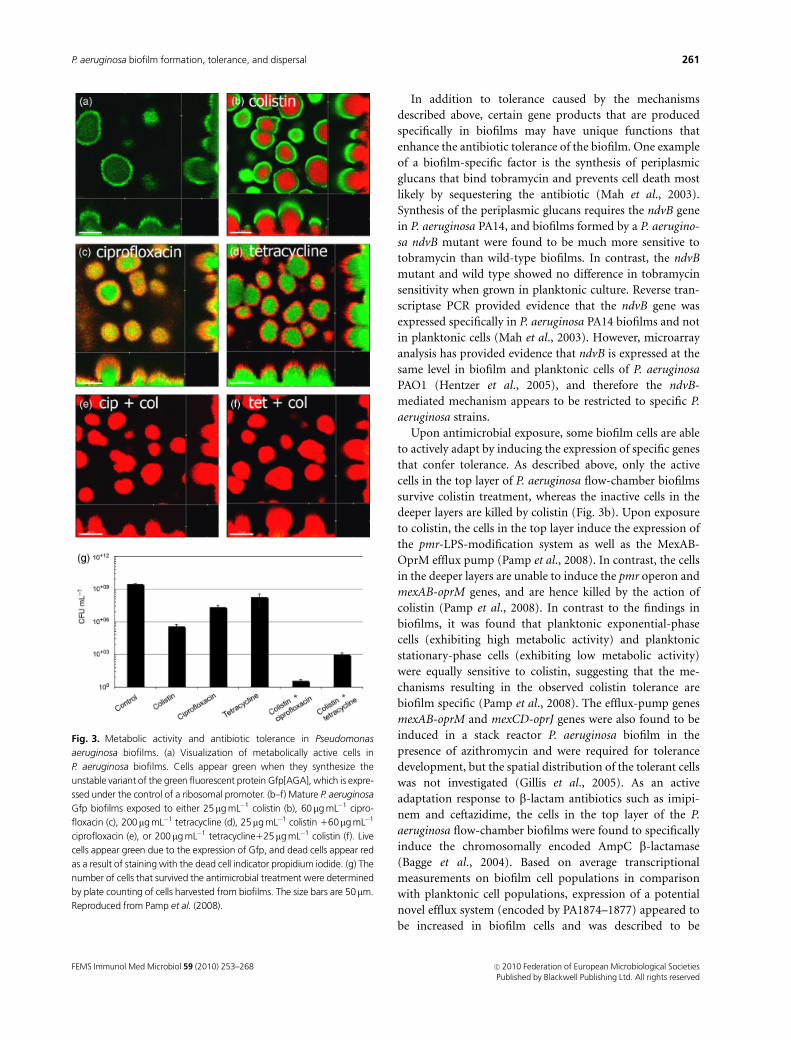
high metabolic activity (see Fig. 3a) (Xu et al., 1998; Walters
et al., 2003; Werner et al., 2004; Pamp et al., 2008). This
spatial distribution of bacteria with low and high metabolic
activity is prevailing due to microscale chemical gradients
that are generated by the relative diffusion and consumption
rates of chemicals as can be revealed by microelectrode
measurements (e.g. Xu et al., 1998; Werner et al., 2004). For
example, oxygen and nutrient concentrations are high in the
bulk liquid and top layer of the biofilms, whereas concentra-
tions are low in the deeper layers of the biofilm. Conven-
tional antimicrobial agents that are known to interfere with
fundamental physiological processes of bacterial cells, such
as replication (e.g. ciprofloxacin), or translation (e.g. tetra-
cycline, tobramycin, gentamicin), were found to specifically
kill the metabolically active cells in the top layer of biofilms,
whereas cells of low metabolic activity survived the treat-
ment (see Fig. 3c and d) (Hentzer et al., 2003; Walters et al.,
2003; Banin et al., 2006; Pamp et al., 2008). In contrast,
antimicrobial agents that interfere with bacterial membrane
structure/function, such as colistin, EDTA, and SDS, were
found to kill the cells in the deeper layer, whereas cells of
high metabolic activity in the top layer survived the treat-
ment (see Fig. 3b) (Banin et al., 2006; Haagensen et al., 2007;
Pamp et al., 2008). In addition, DFO-gallium (a post-
transition metal in complex with a siderophore), which is
known to interfere with cellular iron metabolism, was found
to preferentially kill bacteria in the deeper layers of P.
aeruginosa biofilm (Kaneko et al., 2007; Banin et al., 2008).
Systematic combined antimicrobial treatments simulta-
neously targeting physiologically distinct subpopulations,
for example using ciprofloxacin1colistin (see Fig. 3e),
tetracycline1colistin (see Fig. 3f), or gentamicin1DFO-
gallium, enables killing of almost all the bacteria in P.
aeruginosa biofilms (Banin et al., 2008; Pamp et al., 2008).
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
260 M. Harmsen et al.
In addition to tolerance caused by the mechanisms
described above, certain gene products that are produced
specifically in biofilms may have unique functions that
enhance the antibiotic tolerance of the biofilm. One example
of a biofilm-specific factor is the synthesis of periplasmic
glucans that bind tobramycin and prevents cell death most
likely by sequestering the antibiotic (Mah et al., 2003).
Synthesis of the periplasmic glucans requires the ndvB gene
in P. aeruginosa PA14, and biofilms formed by a P. aerugino-
sa ndvB mutant were found to be much more sensitive to
tobramycin than wild-type biofilms. In contrast, the ndvB
mutant and wild type showed no difference in tobramycin
sensitivity when grown in planktonic culture. Reverse tran-
scriptase PCR provided evidence that the ndvB gene was
expressed specifically in P. aeruginosa PA14 biofilms and not
in planktonic cells (Mah et al., 2003). However, microarray
analysis has provided evidence that ndvB is expressed at the
same level in biofilm and planktonic cells of P. aeruginosa
PAO1 (Hentzer et al., 2005), and therefore the ndvB-
mediated mechanism appears to be restricted to specific P.
aeruginosa strains.
Upon antimicrobial exposure, some biofilm cells are able
to actively adapt by inducing the expression of specific genes
that confer tolerance. As described above, only the active
cells in the top layer of P. aeruginosa flow-chamber biofilms
survive colistin treatment, whereas the inactive cells in the
deeper layers are killed by colistin (Fig. 3b). Upon exposure
to colistin, the cells in the top layer induce the expression of
the pmr-LPS-modification system as well as the MexAB-
OprM efflux pump (Pamp et al., 2008). In contrast, the cells
in the deeper layers are unable to induce the pmr operon and
mexAB-oprM genes, and are hence killed by the action of
colistin (Pamp et al., 2008). In contrast to the findings in
biofilms, it was found that planktonic exponential-phase
cells (exhibiting high metabolic activity) and planktonic
stationary-phase cells (exhibiting low metabolic activity)
were equally sensitive to colistin, suggesting that the me-
chanisms resulting in the observed colistin tolerance are
biofilm specific (Pamp et al., 2008). The efflux-pump genes
mexAB-oprM and mexCD-oprJ genes were also found to be
induced in a stack reactor P. aeruginosa biofilm in the
presence of azithromycin and were required for tolerance
development, but the spatial distribution of the tolerant cells
was not investigated (Gillis et al., 2005). As an active
adaptation response to b-lactam antibiotics such as imipi-
nem and ceftazidime, the cells in the top layer of the P.
aeruginosa flow-chamber biofilms were found to specifically
induce the chromosomally encoded AmpC b-lactamase
(Bagge et al., 2004). Based on average transcriptional
measurements on biofilm cell populations in comparison
with planktonic cell populations, expression of a potential
novel efflux system (encoded by PA1874–1877) appeared to
be increased in biofilm cells and was described to be
Fig. 3. Metabolic activity and antibiotic tolerance in Pseudomonas
aeruginosa biofilms. (a) Visualization of metabolically active cells in
P. aeruginosa biofilms. Cells appear green when they synthesize the
unstable variant of the green fluorescent protein Gfp[AGA], which is expre-
ssed under the control of a ribosomal promoter. (b–f) Mature P. aeruginosa
Gfp biofilms exposed to either 25mg mL�1 colistin (b), 60mg mL�1 cipro-
floxacin (c), 200mg mL�1 tetracycline (d), 25mg mL�1 colistin 160mg mL�1
ciprofloxacin (e), or 200mg mL�1 tetracycline125mg mL�1 colistin (f). Live
cells appear green due to the expression of Gfp, and dead cells appear red
as a result of staining with the dead cell indicator propidium iodide. (g) The
number of cells that survived the antimicrobial treatment were determined
by plate counting of cells harvested from biofilms. The size bars are 50mm.
Reproduced from Pamp et al. (2008).
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
261P. aeruginosa biofilm formation, tolerance, and dispersal

involved in the tolerance toward tobramycin, gentamicin,
and ciprofloxacin (Zhang & Mah, 2008). In addition,
planktonic P. aeruginosa cells appear to use an adaptation
strategy, which involves upregulation of c-di-GMP-depen-
dent genes that increase protective biofilm formation in the
presence of subinhibitory concentrations of antimicrobial
agents (Hoffman et al., 2005).
Mulcahy et al. (2008) presented evidence that eDNA may
create a cation-limited environment in P. aeruginosa bio-
films that result in induction of the pmr-LPS-modification
system conferring tolerance toward antimicrobial peptides
and aminoglycosides. However, Pamp et al. (2008) did not
observe induction of the pmr genes in regions of P. aerugi-
nosa flow-chamber biofilms shown by Allesen-Holm et al.
(2006) to have high concentrations of eDNA.
From the observations described above, it appears that
the majority of biofilm cells can be killed by combined
antimicrobial treatment targeting the major physiologically
distinct cell subpopulations. However, it has been reported
that even in combined treatments involving two antimicro-
bials, a small number of bacteria did survive (see Fig. 3g)
(Pamp et al., 2008). Offspring of the bacteria that survived
the combined antimicrobial treatment did not exhibit
increased resistance to the antimicrobial agents used (Pamp
et al., 2008). It might be speculated that these few surviving
cells represent so-called persister cells. Persister cells are
dormant or slowly dividing bacteria that are less vulnerable
to antibiotics than the majority of the cell population
(Brooun et al., 2000; Lewis, 2001; De Groote et al., 2009).
Mutant screens in P. aeruginosa have provided evidence that
a number of genes (e.g. rpoS, spoT, relA, dksA, dinG, spuC,
algR, pilH, ycgM, pheA) are involved in persister formation,
suggesting that the persister phenotype can be reached
through multiple pathways (Murakami et al., 2005; Viducic
et al., 2006; De Groote et al., 2009). In addition to the
various physiologically distinct cell subpopulations in bio-
films, the generation of genetic variant cells within biofilms
has been described. Rough/wrinkled and small colony
variants in P. aeruginosa biofilms can emerge in the absence
or presence of an antimicrobial agent, whereof some of the
variants can exhibit reduced susceptibility toward antimi-
crobial compounds (e.g. H2O2) (e.g. Drenkard & Ausubel,
2002; Boles et al., 2004; Kirisits et al., 2005).
Recent reports have demonstrated that rhamnolipid
production by P. aeruginosa plays a role in the tolerance of
P. aeruginosa biofilms toward immune cells (Bjarnsholt
et al., 2005; Jensen et al., 2007; Alhede et al., 2009). Purified
P. aeruginosa rhamnolipids were shown to be able to destroy
polymorphonuclear neutrophilic leukocytes (PMNs) via
necrosis (Jensen et al., 2007). Moreover, it was reported that
biofilm cells of P. aeruginosa respond to the presence of
PMNs by upregulating the synthesis of rhamnolipid (Alhede
et al., 2009). The available evidence suggests that the
rhamnolipids stick to the biofilm bacteria and function as a
shield that eliminates immune cells upon contact (Alhede
et al., 2009).
Dispersal
In addition to the mechanisms involved in biofilm forma-
tion, bacteria also possess mechanisms to disperse from
biofilms. These mechanisms involve a reduction of bacterial
adhesiveness, and breakdown or modulation of the biofilm
matrix. Emigration of cells from biofilm communities is
necessary to spawn novel communities at new locations, and
it may be induced if the biofilm cells face unfavorable
conditions (e.g. Tolker-Nielsen et al., 2000; Gjermansen
et al., 2005).
In P. aeruginosa biofilms grown in flow chambers irri-
gated with LB medium, local dispersal was observed as a
hollowing out of some microcolonies (Purevdorj-Gage
et al., 2005). In the initial phase of the dispersal process, a
wall-forming subpopulation of nonmotile cells constituted
the outer part of the microcolonies, whereas a motile rapidly
moving subpopulation was present inside the microcolo-
nies. The motile subpopulation eventually found its way out
of the microcolony, which resulted in microcolonies with a
central void. This dispersal phenomenon was shown to be
dependent on the microcolonies reaching a critical size.
Dispersal of P. aeruginosa biofilms in response to shifts in
carbon availability was reported by Sauer et al. (2004).
Pseudomonas aeruginosa biofilms grown in flow chambers
on glutamate medium responded to an abrupt upshift in
carbon availability by initiating a dispersion process that led
to the majority of the biomass being released from the
biofilm. The extent of dispersion was dependent on the
carbon source and was associated with increased expression
of flagella and downregulation of twitching motility. In a
subsequent study by Morgan et al. (2006), a gene product
involved in sensing environmental cues that trigger P.
aeruginosa biofilm dispersal was identified. The gene encod-
ing this sensor was denoted bdlA for biofilm dispersion
locus, and through sequence analysis and phenotypic com-
parison of the P. aeruginosa wild type and bdlA mutant, the
BdlA protein was suggested to be a chemotaxis regulator
that affects the intracellular level of c-di-GMP. As described
in a previous section, evidence is accruing that the produc-
tion of matrix components such as Pel/Psl polysaccharides
and Cup fimbriae is regulated via proteins that contain
diguanylate cyclase or phosphodiesterase activities and con-
trol the intracellular level of c-di-GMP (Hickman et al.,
2005; Gjermansen et al., 2006; Meissner et al., 2007). It
appears that in general, high intracellular c-di-GMP levels
upregulate matrix production and biofilm formation,
whereas low intracellular c-di-GMP levels downregulate
matrix production and induce a planktonic lifestyle. Carbon
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
262 M. Harmsen et al.

starvation and nitric oxide signaling were shown to induce
the dispersal of P. aeruginosa biofilms via the induction of
phosphodiesterase activity, causing decreased intracellular
c-di-GMP levels (Barraud et al., 2009; Schleheck et al.,
2009). Evidence was presented that the above-mentioned
BdlA chemotaxis regulator is involved in nitric oxide-
mediated biofilm dispersal (Barraud et al., 2009).
As described in a previous section, rhamnolipid appears
to play multiple roles in the P. aeruginosa biofilm develop-
ment cycle, one of them being that the production of large
amounts may lead to the dispersal of cells from the biofilms
(Schooling et al., 2004; Boles et al., 2005). Ryan et al. (2009)
recently presented evidence that rhamnolipid-mediated dis-
persal in P. aeruginosa biofilms may involve c-di-GMP.
Evidence was provided that the PA2572 protein has a
degenerate inactive c-di-GMP phosphodiesterase domain
that may play a regulatory role, and a P. aeruginosa PA2572
mutant was found to overproduce rhamnolipid.
Recent work by Davies & Marques (2009) provides
evidence that the compound cis-2-decenoic acid produced
by P. aeruginosa is capable of inducing the dispersal of
established biofilms and of inhibiting biofilm development.
When added exogenously to P. aeruginosa biofilms at a
native concentration, cis-2-decenoic acid was shown to
induce the dispersal of biofilm microcolonies. This molecule
was also shown to induce the dispersal of biofilms, formed
by Escherichia coli, Klebsiella pneumoniae, Proteus mirabilis,
Streptococcus pyogenes, Bacillus subtilis, Staphylococcus aur-
eus, and the yeast Candida albicans. The authors suggested
that cis-2-decenoic acid is produced continuously by P.
aeruginosa during growth in biofilms, and that small micro-
colonies do not disperse because cis-2-decenoic acid is
removed through diffusive and advective transport; how-
ever, dispersal from larger microcolonies occurs because the
rate of production of cis-2-decenoic acid exceeds the rate of
diffusion.
A recent publication by Gjermansen et al. (2010) de-
scribes a mechanism involved in the dispersal of Pseudomo-
nas putida biofilms that may also be of relevance for P.
aeruginosa biofilms. In P. putida, the large adhesive outer-
membrane protein, LapA, mediates attachment to surfaces
and to matrix components. Release of LapA from the cell
surface results in biofilm dispersal and is mediated through
the activity of the periplasmic protease LapG. The activity of
the LapG protease is controlled by the transmembrane
protein LapD, which contains a c-di-GMP-binding domain,
and represses LapG when the concentration of c-di-GMP is
high, but derepressses LapG when the concentration of c-di-
GMP is low. The available evidence (Hinsa et al., 2003;
Hinsa & O’Toole, 2006; Monds et al., 2007; Newell et al.,
2009) suggests that a similar system is operating in Pseudo-
monas fluorescens. Pseudomonas aeruginosa encodes a num-
ber of large adhesive proteins, but it does not have a
homolog of lapA. However, P. aeruginosa does have homo-
logs of lapD and lapG, and therefore a mechanism similar to
the P. putida mechanism could potentially be involved in P.
aeruginosa biofilm dispersal.
Final remarks
Recent work has provided knowledge about the environ-
mental cues, genetic elements, and molecular mechanism
involved in biofilm formation, tolerance, and dispersal.
Based on this research, potential antibiofilm strategies can
be envisioned, for example enzymatic digestion of matrix
components, blocking of c-di-GMP regulated matrix synth-
esis, treatment with multiple antibiotics that target different
subpopulations, and blocking of quorum sensing. However,
the molecular understanding of the biofilm lifestyle ob-
tained from in vitro studies should be used in future in vivo
studies using animal models that mimic the complex inter-
actions between the biofilm and the host. A more detailed
understanding of the multicellular nature of microbial life
will ultimately enable us to develop efficient treatments
against biofilm-related infections.
Acknowledgement
This work was supported by grants from the Danish Council
for Independent Research.
References
Aas FE, Wolfgang M, Frye S, Dunham S, Lovold C & Koomey M
(2002) Competence for natural transformation in Neisseria
gonorrhoeae: components of DNA binding and uptake linked
to type IV pilus expression. Mol Microbiol 46: 749–760.
Alhede M, Bjarnsholt T, Jensen PO et al. (2009) Pseudomonas
aeruginosa recognizes and responds aggressively to the
presence of polymorphonuclear leukocytes. Microbiology 155:
3500–3508.
Alipour M, Suntres ZE & Omri A (2009) Importance of DNase
and alginate lyase for enhancing free and liposome
encapsulated aminoglycoside activity against Pseudomonas
aeruginosa. J Antimicrob Chemoth 64: 317–325.
Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS,
Kjelleberg S, Molin S, Givskov M & Tolker-Nielsen T (2006) A
characterization of DNA release in Pseudomonas aeruginosa
cultures and biofilms. Mol Microbiol 59: 1114–1128.
Al-Tahhan RA, Sandrin TR, Bodour AA & Maier RM (2000)
Rhamnolipid-induced removal of lipopolysaccharide from
Pseudomonas aeruginosa: effect on cell surface properties and
interaction with hydrophobic substrates. Appl Environ Microb
66: 3262–3268.
Amikam D & Galperin MY (2006) PilZ domain is part of the
bacterial c-di-GMP binding protein. Bioinformatics 22: 3–6.
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
263P. aeruginosa biofilm formation, tolerance, and dispersal

Bagge N, Hentzer M, Andersen JB, Ciofu O, Givskov M & Hoiby
N (2004) Dynamics and spatial distribution of beta-lactamase
expression in Pseudomonas aeruginosa biofilms. Antimicrob
Agents Ch 48: 1168–1174.
Banin E, Vasil ML & Greenberg EP (2005) Iron and Pseudomonas
aeruginosa biofilm formation. P Natl Acad Sci USA 102:
11076–11081.
Banin E, Brady KM & Greenberg EP (2006) Chelator-induced
dispersal and killing of Pseudomonas aeruginosa cells in a
biofilm. Appl Environ Microb 72: 2064–2069.
Banin E, Lozinski A, Brady KM, Berenshtein E, Butterfield PW,
Moshe M, Chevion M & Greenberg EP (2008) The potential of
desferrioxamine-gallium as an anti-Pseudomonas therapeutic
agent. P Natl Acad Sci USA 105: 16761–16766.
Barken KB, Pamp SJ, Yang L, Gjermansen M, Bertrand JJ, Klausen
M, Givskov M, Whitchurch CB, Engel JN & Tolker-Nielsen T
(2008) Roles of type IV pili, flagellum-mediated motility and
extracellular DNA in the formation of mature multicellular
structures in Pseudomonas aeruginosa biofilms. Environ
Microbiol 10: 2331–2343.
Barraud N, Schleheck D, Klebensberger J, Webb JS, Hassett DJ,
Rice SA & Kjelleberg S (2009) Nitric oxide signaling in
Pseudomonas aeruginosa biofilms mediates phosphodiesterase
activity, decreased cyclic diguanosine-5 0-monophosphate
levels and enhanced dispersal. J Bacteriol 191: 7333–7342.
Bjarnsholt T, Jensen PO, Burmolle M et al. (2005) Pseudomonas
aeruginosa tolerance to tobramycin, hydrogen peroxide and
polymorphonuclear leukocytes is quorum-sensing dependent.
Microbiology 151: 373–383.
Boles BR, Thoendel M & Singh PK (2004) Self-generated
diversity produces ‘insurance effects’ in biofilm communities.
P Natl Acad Sci USA 101: 16630–16635.
Boles BR, Thoendel M & Singh PK (2005) Rhamnolipids mediate
detachment of Pseudomonas aeruginosa from biofilms. Mol
Microbiol 57: 1210–1223.
Bollinger N, Hassett DJ, Iglewski BH, Costerton JW &
McDermott TR (2001) Gene expression in Pseudomonas
aeruginosa: evidence of iron override effects on quorum
sensing and biofilm-specific gene regulation. J Bacteriol 183:
1990–1996.
Borlee BR, Goldman A, Murakami K, Samudrala R, Wozniak DJ
& Parsek MR (2010) Pseudomonas aeruginosa uses a cyclic-di-
GMP-regulated adhesin to reinforce the biofilm extracellular
matrix. Mol Microbiol 75: 827–842.
Bredenbruch F, Geffers R, Nimtz M, Buer J & Haussler S (2006)
The Pseudomonas aeruginosa quinolone signal (PQS) has an
iron-chelating activity. Environ Microbiol 8: 1318–1329.
Brooun A, Liu S & Lewis K (2000) A dose–response study of
antibiotic resistance in Pseudomonas aeruginosa biofilms.
Antimicrob Agents Ch 44: 640–646.
Caiazza NC & O’Toole GA (2004) SadB is required for the
transition from reversible to irreversible attachment
during biofilm formation by Pseudomonas aeruginosa PA14.
J Bacteriol 186: 4476–4485.
Chiang P & Burrows LL (2003) Biofilm formation by
hyperpiliated mutants of Pseudomonas aeruginosa. J Bacteriol
185: 2374–2378.
Cochran WL, McFeters GA & Stewart PS (2000) Reduced
susceptibility of thin Pseudomonas aeruginosa biofilms to
hydrogen peroxide and monochloramine. J Appl Microbiol 88:
22–30.
D’Argenio DA, Calfee MW, Rainey PB & Pesci EC (2002)
Autolysis and autoaggregation in Pseudomonas aeruginosa
colony morphology mutants. J Bacteriol 184: 6481–6489.
Davey ME, Caiazza NC & O’Toole GA (2003) Rhamnolipid
surfactant production affects biofilm architecture in
Pseudomonas aeruginosa PAO1. J Bacteriol 185: 1027–1036.
Davies DG & Marques CN (2009) A fatty acid messenger is
responsible for inducing dispersion in microbial biofilms.
J Bacteriol 191: 1393–1403.
Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW &
Greenberg EP (1998) The involvement of cell-to-cell signals in
the development of a bacterial biofilm. Science 280: 295–298.
De Groote VN, Verstraeten N, Fauvart M, Kint CI, Verbeeck AM,
Beullens S, Cornelis P & Michiels J (2009) Novel persistence
genes in Pseudomonas aeruginosa identified by high-
throughput screening. FEMS Microbiol Lett 297: 73–79.
De Kievit TR, Parkins MD, Gillis RJ, Srikumar R, Ceri H, Poole K,
Iglewski BH & Storey DG (2001) Multidrug efflux pumps:
expression patterns and contribution to antibiotic resistance in
Pseudomonas aeruginosa biofilms. Antimicrob Agents Ch 45:
1761–1770.
Deziel E, Comeau Y & Villemur R (2001) Initiation of biofilm
formation by Pseudomonas aeruginosa 57RP correlates with
emergence of hyperpiliated and highly adherent phenotypic
variants deficient in swimming, swarming, and twitching
motilities. J Bacteriol 183: 1195–1204.
Diggle SP, Matthijs S, Wright VJ et al. (2007) The Pseudomonas
aeruginosa 4-quinolone signal molecules HHQ and PQS play
multifunctional roles in quorum sensing and iron entrapment.
Chem Biol 14: 87–96.
Drenkard E & Ausubel FM (2002) Pseudomonas biofilm
formation and antibiotic resistance are linked to phenotypic
variation. Nature 416: 740–743.
Duan K & Surette MG (2007) Environmental regulation of
Pseudomonas aeruginosa PAO1 Las and Rhl quorum-sensing
systems. J Bacteriol 189: 4827–4836.
Friedman L & Kolter R (2004a) Genes involved in matrix
formation in Pseudomonas aeruginosa PA14 biofilms. Mol
Microbiol 51: 675–690.
Friedman L & Kolter R (2004b) Two genetic loci produce distinct
carbohydrate-rich structural components of the Pseudomonas
aeruginosa biofilm matrix. J Bacteriol 186: 4457–4465.
Froshauer S, Silvia AM, Chidambaram M, Sharma B & Weinstock
GM (1996) Sensitization of bacteria to danofloxacin by
temperate prophages. Antimicrob Agents Ch 40: 1561–1563.
Gillis RJ, White KG, Choi KH, Wagner VE, Schweizer HP &
Iglewski BH (2005) Molecular basis of azithromycin-resistant
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
264 M. Harmsen et al.

Pseudomonas aeruginosa biofilms. Antimicrob Agents Ch 49:
3858–3867.
Gjermansen M, Ragas P, Sternberg C, Molin S & Tolker-Nielsen T
(2005) Characterization of starvation-induced dispersion in
Pseudomonas putida biofilms. Environ Microbiol 7: 894–906.
Gjermansen M, Ragas P & Tolker-Nielsen T (2006) Proteins with
GGDEF and EAL domains regulate Pseudomonas putida
biofilm formation and dispersal. FEMS Microbiol Lett 265:
215–224.
Gjermansen M, Nilsson M, Yang L & Tolker-Nielsen T (2010)
Characterization of starvation-induced dispersion in
Pseudomonas putida biofilms: genetic elements and molecular
mechanisms. Mol Microbiol 75: 815–826.
Goodman AL, Kulasekara B, Rietsch A, Boyd D, Smith RS & Lory
S (2004) A signaling network reciprocally regulates genes
associated with acute infection and chronic persistence in
Pseudomonas aeruginosa. Dev Cell 7: 745–754.
Goodman AL, Merighi M, Hyodo M, Ventre I, Filloux A & Lory S
(2009) Direct interaction between sensor kinase proteins
mediates acute and chronic disease phenotypes in a bacterial
pathogen. Gene Dev 23: 249–259.
Govan JR & Deretic V (1996) Microbial pathogenesis in cystic
fibrosis: mucoid Pseudomonas aeruginosa and Burkholderia
cepacia. Microbiol Rev 60: 539–574.
Guzzo CR, Salinas RK, Andrade MO & Farah CS (2009) PilZ
protein structure and interactions with PilB and the FimX EAL
domain: implications for control of type IV pilus biogenesis.
J Mol Biol 393: 848–866.
Haagensen JA, Klausen M, Ernst RK, Miller SI, Folkesson A,
Tolker-Nielsen T & Molin S (2007) Differentiation and
distribution of colistin and sodium dodecyl sulfate-tolerant
cells in Pseudomonas aeruginosa biofilms. J Bacteriol 189:
28–37.
Hatch RA & Schiller NL (1998) Alginate lyase promotes diffusion
of aminoglycosides through the extracellular polysaccharide of
mucoid Pseudomonas aeruginosa. Antimicrob Agents Ch 42:
974–977.
Hay ID, Gatland K, Campisano A, Jordens JZ & Rehm BH
(2009a) Impact of alginate overproduction on attachment and
biofilm architecture of a supermucoid Pseudomonas
aeruginosa strain. Appl Environ Microb 75: 6022–6025.
Hay ID, Remminghorst U & Rehm BH (2009b) MucR, a novel
membrane-associated regulator of alginate biosynthesis in
Pseudomonas aeruginosa. Appl Environ Microb 75: 1110–1120.
Hentzer M, Teitzel GM, Balzer GJ, Heydorn A, Molin S, Givskov
M & Parsek MR (2001) Alginate overproduction affects
Pseudomonas aeruginosa biofilm structure and function.
J Bacteriol 183: 5395–5401.
Hentzer M, Riedel K, Rasmussen TB et al. (2002) Inhibition of
quorum sensing in Pseudomonas aeruginosa biofilm bacteria
by a halogenated furanone compound. Microbiology 148:
87–102.
Hentzer M, Wu H, Andersen JB et al. (2003) Attenuation of
Pseudomonas aeruginosa virulence by quorum sensing
inhibitors. EMBO J 22: 3803–3815.
Hentzer M, Givskov M & Eberl L (2004) Quorum sensing in
biofilms:gossip in the slime world? Microbial Biofilms
(Ghannoum M & O’Toole GA, eds), pp. 118–140. ASM Press,
Washington, DC.
Hentzer M, Eberl L & Givskov M (2005) Transcriptome analysis
of Pseudomonas aeruginosa biofilm development: anaerobic
respiration and iron limitation. Biofilms 2: 37–61.
Heurlier K, Denervaud V, Haenni M, Guy L, Krishnapillai V &
Haas D (2005) Quorum-sensing-negative (lasR) mutants of
Pseudomonas aeruginosa avoid cell lysis and death. J Bacteriol
187: 4875–4883.
Heydorn A, Ersboll B, Kato J, Hentzer M, Parsek MR, Tolker-
Nielsen T, Givskov M & Molin S (2002) Statistical analysis of
Pseudomonas aeruginosa biofilm development: impact of
mutations in genes involved in twitching motility, cell-to-cell
signaling, and stationary-phase sigma factor expression. Appl
Environ Microb 68: 2008–2017.
Hickman JW & Harwood CS (2008) Identification of FleQ from
Pseudomonas aeruginosa as a c-di-GMP-responsive
transcription factor. Mol Microbiol 69: 376–389.
Hickman JW, Tifrea DF & Harwood CS (2005) A chemosensory
system that regulates biofilm formation through modulation
of cyclic diguanylate levels. P Natl Acad Sci USA 102:
14422–14427.
Hinsa SM & O’Toole GA (2006) Biofilm formation by
Pseudomonas fluorescens WCS365: a role for LapD.
Microbiology 152: 1375–1383.
Hinsa SM, Espinosa-Urgel M, Ramos JL & O’Toole GA (2003)
Transition from reversible to irreversible attachment during
biofilm formation by Pseudomonas fluorescens WCS365
requires an ABC transporter and a large secreted protein. Mol
Microbiol 49: 905–918.
Hoffman LR, D’Argenio DA, MacCoss MJ, Zhang Z, Jones RA &
Miller SI (2005) Aminoglycoside antibiotics induce bacterial
biofilm formation. Nature 436: 1171–1175.
Hoiby N (1974) Pseudomonas aeruginosa infection in cystic
fibrosis. Relationship between mucoid strains of Pseudomonas
aeruginosa and the humoral immune response. Acta Patholog
Microb 82: 551–558.
Hoyle BD, Alcantara J & Costerton JW (1992) Pseudomonas
aeruginosa biofilm as a diffusion barrier to piperacillin.
Antimicrob Agents Ch 36: 2054–2056.
Huang B, Whitchurch CB & Mattick JS (2003) FimX, a
multidomain protein connecting environmental signals to
twitching motility in Pseudomonas aeruginosa. J Bacteriol 185:
7068–7076.
Jackson KD, Starkey M, Kremer S, Parsek MR & Wozniak DJ
(2004) Identification of psl, a locus encoding a potential
exopolysaccharide that is essential for Pseudomonas aeruginosa
PAO1 biofilm formation. J Bacteriol 186: 4466–4475.
Jensen PO, Bjarnsholt T, Phipps R et al. (2007) Rapid necrotic
killing of polymorphonuclear leukocytes is caused by quorum-
sensing-controlled production of rhamnolipid by
Pseudomonas aeruginosa. Microbiology 153: 1329–1338.
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
265P. aeruginosa biofilm formation, tolerance, and dispersal

Juhas M, Eberl L & Tummler B (2005) Quorum sensing: the
power of cooperation in the world of Pseudomonas. Environ
Microbiol 7: 459–471.
Kadurugamuwa JL & Beveridge TJ (1996) Bacteriolytic effect of
membrane vesicles from Pseudomonas aeruginosa on other
bacteria including pathogens: conceptually new antibiotics.
J Bacteriol 178: 2767–2774.
Kaneko Y, Thoendel M, Olakanmi O, Britigan BE & Singh PK
(2007) The transition metal gallium disrupts Pseudomonas
aeruginosa iron metabolism and has antimicrobial and
antibiofilm activity. J Clin Invest 117: 877–888.
Kim EJ, Wang W, Deckwer WD & Zeng AP (2005) Expression of
the quorum-sensing regulatory protein LasR is strongly
affected by iron and oxygen concentrations in cultures of
Pseudomonas aeruginosa irrespective of cell density.
Microbiology 151: 1127–1138.
Kirisits MJ, Prost L, Starkey M & Parsek MR (2005)
Characterization of colony morphology variants isolated from
Pseudomonas aeruginosa biofilms. Appl Environ Microb 71:
4809–4821.
Klausen M, Aaes-Jorgensen A, Molin S & Tolker-Nielsen T
(2003a) Involvement of bacterial migration in the
development of complex multicellular structures in
Pseudomonas aeruginosa biofilms. Mol Microbiol 50: 61–68.
Klausen M, Heydorn A, Ragas P, Lambertsen L, Aaes-Jorgensen
A, Molin S & Tolker-Nielsen T (2003b) Biofilm formation by
Pseudomonas aeruginosa wild type, flagella and type IV pili
mutants. Mol Microbiol 48: 1511–1524.
Klebensberger J, Birkenmaier A, Geffers R, Kjelleberg S & Philipp
B (2009) SiaA and SiaD are essential for inducing
autoaggregation as a specific response to detergent stress in
Pseudomonas aeruginosa. Environ Microbiol 11: 3073–3086.
Kohler T, Curty LK, Barja F, van Delden C & Pechere JC (2000)
Swarming of Pseudomonas aeruginosa is dependent on cell-to-
cell signaling and requires flagella and pili. J Bacteriol 182:
5990–5996.
Kuchma SL, Brothers KM, Merritt JH, Liberati NT, Ausubel FM &
O’Toole GA (2007) BifA, a cyclic-di-GMP phosphodiesterase,
inversely regulates biofilm formation and swarming motility
by Pseudomonas aeruginosa PA14. J Bacteriol 189: 8165–8178.
Kulasekara HD, Ventre I, Kulasekara BR, Lazdunski A, Filloux A
& Lory S (2005) A novel two-component system controls the
expression of Pseudomonas aeruginosa fimbrial cup genes. Mol
Microbiol 55: 368–380.
Lee VT, Matewish JM, Kessler JL, Hyodo M, Hayakawa Y & Lory
S (2007) A cyclic-di-GMP receptor required for bacterial
exopolysaccharide production. Mol Microbiol 65: 1474–1484.
Lequette Y & Greenberg EP (2005) Timing and localization of
rhamnolipid synthesis gene expression in Pseudomonas
aeruginosa biofilms. J Bacteriol 187: 37–44.
Lewis K (2001) Riddle of biofilm resistance. Antimicrob Agents Ch
45: 999–1007.
Lory S, Merighi M & Hyodo M (2009) Multiple activities of
c-di-GMP in Pseudomonas aeruginosa. Nucleic Acids Symp Ser
53: 51–52.
Ma L, Jackson KD, Landry RM, Parsek MR & Wozniak DJ (2006)
Analysis of Pseudomonas aeruginosa conditional psl variants
reveals roles for the psl polysaccharide in adhesion and
maintaining biofilm structure postattachment. J Bacteriol 188:
8213–8221.
Ma L, Conover M, Lu H, Parsek MR, Bayles K & Wozniak DJ
(2009) Assembly and development of the Pseudomonas
aeruginosa biofilm matrix. PLoS Pathog 5: e1000354.
Mah TF, Pitts B, Pellock B, Walker GC, Stewart PS & O’Toole GA
(2003) A genetic basis for Pseudomonas aeruginosa biofilm
antibiotic resistance. Nature 426: 306–310.
Mashburn LM & Whiteley M (2005) Membrane vesicles traffic
signals and facilitate group activities in a prokaryote. Nature
437: 422–425.
Meissner A, Wild V, Simm R, Rohde M, Erck C, Bredenbruch F,
Morr M, Romling U & Haussler S (2007) Pseudomonas
aeruginosa cupA-encoded fimbriae expression is regulated by a
GGDEF and EAL domain-dependent modulation of the
intracellular level of cyclic diguanylate. Environ Microbiol 9:
2475–2485.
Merighi M, Lee VT, Hyodo M, Hayakawa Y & Lory S (2007) The
second messenger bis-(30-50)-cyclic-GMP and its PilZ
domain-containing receptor Alg44 are required for alginate
biosynthesis in Pseudomonas aeruginosa. Mol Microbiol 65:
876–895.
Merritt JH, Brothers KM, Kuchma SL & O’Toole GA (2007) SadC
reciprocally influences biofilm formation and swarming
motility via modulation of exopolysaccharide production and
flagellar function. J Bacteriol 189: 8154–8164.
Mikkelsen H, Ball G, Giraud C & Filloux A (2009) Expression of
Pseudomonas aeruginosa cupD fimbrial genes is
antagonistically controlled by RcsB and the EAL-containing
PvrR response regulators. PLoS ONE 4: e6018.
Molin S & Tolker-Nielsen T (2003) Gene transfer occurs with
enhanced efficiency in biofilms and induces enhanced
stabilisation of the biofilm structure. Curr Opin Biotech 14:
255–261.
Monds RD, Newell PD, Gross RH & O’Toole GA (2007)
Phosphate-dependent modulation of c-di-GMP levels
regulates Pseudomonas fluorescens Pf0-1 biofilm formation by
controlling secretion of the adhesin LapA. Mol Microbiol 63:
656–679.
Morgan R, Kohn S, Hwang SH, Hassett DJ & Sauer K (2006)
BdlA, a chemotaxis regulator essential for biofilm dispersion
in Pseudomonas aeruginosa. J Bacteriol 188: 7335–7343.
Mulcahy H, Charron-Mazenod L & Lewenza S (2008)
Extracellular DNA chelates cations and induces antibiotic
resistance in Pseudomonas aeruginosa biofilms. PLoS Pathog 4:
e1000213.
Murakami K, Ono T, Viducic D, Kayama S, Mori M, Hirota K,
Nemoto K & Miyake Y (2005) Role for rpoS gene of
Pseudomonas aeruginosa in antibiotic tolerance. FEMS
Microbiol Lett 242: 161–167.
Nakamura S, Higashiyama Y, Izumikawa K, Seki M, Kakeya H,
Yamamoto Y, Yanagihara K, Miyazaki Y, Mizuta Y & Kohno S
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
266 M. Harmsen et al.

(2008) The roles of the quorum-sensing system in the release
of extracellular DNA, lipopolysaccharide, and membrane
vesicles from Pseudomonas aeruginosa. Jpn J Infect Dis 61:
375–378.
Newell PD, Monds RD & O’Toole GA (2009) LapD is a bis-
(30,50)-cyclic dimeric GMP-binding protein that regulates
surface attachment by Pseudomonas fluorescens Pf0-1. P Natl
Acad Sci USA 106: 3461–3466.
Ochsner UA & Reiser J (1995) Autoinducer-mediated regulation
of rhamnolipid biosurfactant synthesis in Pseudomonas
aeruginosa. P Natl Acad Sci USA 92: 6424–6428.
Oglesby LL, Jain S & Ohman DE (2008) Membrane topology and
roles of Pseudomonas aeruginosa Alg8 and Alg44 in alginate
polymerization. Microbiology 154: 1605–1615.
O’Toole GA & Kolter R (1998) Flagellar and twitching motility
are necessary for Pseudomonas aeruginosa biofilm
development. Mol Microbiol 30: 295–304.
Overhage J, Schemionek M, Webb JS & Rehm BH (2005)
Expression of the psl operon in Pseudomonas aeruginosa PAO1
biofilms: PslA performs an essential function in biofilm
formation. Appl Environ Microb 71: 4407–4413.
Pamp SJ & Tolker-Nielsen T (2007) Multiple roles of
biosurfactants in structural biofilm development by
Pseudomonas aeruginosa. J Bacteriol 189: 2531–2539.
Pamp SJ, Gjermansen M, Johansen HK & Tolker-Nielsen T (2008)
Tolerance to the antimicrobial peptide colistin in Pseudomonas
aeruginosa biofilms is linked to metabolically active cells, and
depends on the pmr and mexAB-oprM genes. Mol Microbiol 68:
223–240.
Patriquin GM, Banin E, Gilmour C, Tuchman R, Greenberg EP &
Poole K (2008) Influence of quorum sensing and iron on
twitching motility and biofilm formation in Pseudomonas
aeruginosa. J Bacteriol 190: 662–671.
Petrova OE & Sauer K (2009) A novel signaling network essential
for regulating Pseudomonas aeruginosa biofilm development.
PLoS Pathog 5: e1000668.
Phillips I, Culebras E, Moreno F & Baquero F (1987) Induction of
the SOS response by new 4-quinolones. J Antimicrob Chemoth
20: 631–638.
Purevdorj B, Costerton JW & Stoodley P (2002) Influence of
hydrodynamics and cell signaling on the structure and
behavior of Pseudomonas aeruginosa biofilms. Appl Environ
Microb 68: 4457–4464.
Purevdorj-Gage B, Costerton JW & Stoodley P (2005) Phenotypic
differentiation and seeding dispersal in non-mucoid and
mucoid Pseudomonas aeruginosa biofilms. Microbiology 151:
1569–1576.
Qin Z, Ou Y, Yang Y, Zhu Y, Tolker-Nielsen T, Molin T & Qu D
(2007) Role of autolysin-mediated DNA release in biofilm
formation of Staphylococcus epidermidis. Microbiology 153:
2083–2092.
Rao F, Yang Y, Qi Y & Liang ZX (2008) Catalytic mechanism of
cyclic di-GMP-specific phosphodiesterase: a study of the EAL
domain-containing RocR from Pseudomonas aeruginosa.
J Bacteriol 190: 3622–3631.
Remminghorst U & Rehm BH (2006a) In vitro alginate
polymerization and the functional role of Alg8 in alginate
production by Pseudomonas aeruginosa. Appl Environ Microb
72: 298–305.
Remminghorst U & Rehm BH (2006b) Alg44, a unique protein
required for alginate biosynthesis in Pseudomonas aeruginosa.
FEBS Lett 580: 3883–3888.
Renelli M, Matias V, Lo RY & Beveridge TJ (2004) DNA-
containing membrane vesicles of Pseudomonas aeruginosa
PAO1 and their genetic transformation potential. Microbiology
150: 2161–2169.
Rice KC, Mann EE, Endres JL, Weiss EC, Cassat JE, Smeltzer MS
& Bayles KW (2007) The cidA murein hydrolase regulator
contributes to DNA release and biofilm development in
Staphylococcus aureus. P Natl Acad Sci USA 104: 8113–8118.
Ryan RP, Lucey J, O’Donovan K, McCarthy Y, Yang L, Tolker-
Nielsen T & Dow JM (2009) HD-GYP domain proteins
regulate biofilm formation and virulence in Pseudomonas
aeruginosa. Environ Microbiol 11: 1126–1136.
Ryder C, Byrd M & Wozniak DJ (2007) Role of polysaccharides in
Pseudomonas aeruginosa biofilm development. Curr Opin
Microbiol 10: 644–648.
Sakuragi Y & Kolter R (2007) Quorum-sensing regulation of the
biofilm matrix genes (pel) of Pseudomonas aeruginosa.
J Bacteriol 189: 5383–5386.
Sauer K, Camper AK, Ehrlich GD, Costerton JW & Davies DG
(2002) Pseudomonas aeruginosa displays multiple phenotypes
during development as a biofilm. J Bacteriol 184: 1140–1154.
Sauer K, Cullen MC, Rickard AH, Zeef LA, Davies DG & Gilbert
P (2004) Characterization of nutrient-induced dispersion in
Pseudomonas aeruginosa PAO1 biofilm. J Bacteriol 186:
7312–7326.
Schleheck D, Barraud N, Klebensberger J, Webb JS, McDougald
D, Rice SA & Kjelleberg S (2009) Pseudomonas aeruginosa
PAO1 preferentially grows as aggregates in liquid batch
cultures and disperses upon starvation. PLoS ONE 4: e5513.
Schooling SR, Charaf UK, Allison DG & Gilbert P (2004) A role
for rhamnolipid in biofilm dispersion. Biofilms 1: 91–99.
Schuster M, Lostroh CP, Ogi T & Greenberg EP (2003)
Identification, timing, and signal specificity of Pseudomonas
aeruginosa quorum-controlled genes: a transcriptome analysis.
J Bacteriol 185: 2066–2079.
Shigeta M, Tanaka G, Komatsuzawa H, Sugai M, Suginaka H &
Usui T (1997) Permeation of antimicrobial agents through
Pseudomonas aeruginosa biofilms: a simple method.
Chemotherapy 43: 340–345.
Shrout JD, Chopp DL, Just CL, Hentzer M, Givskov M & Parsek
MR (2006) The impact of quorum sensing and swarming
motility on Pseudomonas aeruginosa biofilm formation is
nutritionally conditional. Mol Microbiol 62: 1264–1277.
Singh PK (2004) Iron sequestration by human lactoferrin
stimulates P. aeruginosa surface motility and blocks biofilm
formation. Biometals 17: 267–270.
FEMS Immunol Med Microbiol 59 (2010) 253–268 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
267P. aeruginosa biofilm formation, tolerance, and dispersal

Singh PK, Parsek MR, Greenberg EP & Welsh MJ (2002) A
component of innate immunity prevents bacterial biofilm
development. Nature 417: 552–555.
Starkey M, Hickman JH, Ma L et al. (2009) Pseudomonas
aeruginosa rugose small-colony variants have adaptations that
likely promote persistence in the cystic fibrosis lung. J Bacteriol
191: 3492–3503.
Tielker D, Hacker S, Loris R, Strathmann M, Wingender J,
Wilhelm S, Rosenau F & Jaeger KE (2005) Pseudomonas
aeruginosa lectin LecB is located in the outer membrane and is
involved in biofilm formation. Microbiology 151: 1313–1323.
Tolker-Nielsen T, Brinch UC, Ragas PC, Andersen JB, Jacobsen
CS & Molin S (2000) Development and dynamics of
Pseudomonas sp. biofilms. J Bacteriol 182: 6482–6489.
Ueda A & Wood TK (2009) Connecting quorum sensing, c-di-
GMP, Pel polysaccharide, and biofilm formation in
Pseudomonas aeruginosa through tyrosine phosphatase TpbA
(PA3885). PLoS Pathog 5: e1000483.
Vallet I, Olson JW, Lory S, Lazdunski A & Filloux A (2001) The
chaperone/usher pathways of Pseudomonas aeruginosa:
identification of fimbrial gene clusters (cup) and their
involvement in biofilm formation. P Natl Acad Sci USA 98:
6911–6916.
van Loosdrecht MC, Norde W & Zehnder AJ (1990) Physical
chemical description of bacterial adhesion. J Biomater Appl 5:
91–106.
van Schaik EJ, Giltner CL, Audette GF, Keizer DW, Bautista DL,
Slupsky CM, Sykes BD & Irvin RT (2005) DNA binding: a
novel function of Pseudomonas aeruginosa type IV pili.
J Bacteriol 187: 1455–1464.
Ventre I, Goodman AL, Vallet-Gely I, Vasseur P, Soscia C, Molin
S, Bleves S, Lazdunski A, Lory S & Filloux A (2006) Multiple
sensors control reciprocal expression of Pseudomonas
aeruginosa regulatory RNA and virulence genes. P Natl Acad
Sci USA 103: 171–176.
Viducic D, Ono T, Murakami K, Susilowati H, Kayama S, Hirota
K & Miyake Y (2006) Functional analysis of spoT, relA and
dksA genes on quinolone tolerance in Pseudomonas aeruginosa
under nongrowing condition. Microbiol Immunol 50: 349–357.
Vrany JD, Stewart PS & Suci PA (1997) Comparison of
recalcitrance to ciprofloxacin and levofloxacin exhibited by
Pseudomonas aeruginosa biofilms displaying rapid-transport
characteristics. Antimicrob Agents Ch 41: 1352–1358.
Wagner VE, Bushnell D, Passador L, Brooks AI & Iglewski BH
(2003) Microarray analysis of Pseudomonas aeruginosa
quorum-sensing regulons: effects of growth phase and
environment. J Bacteriol 185: 2080–2095.
Walters MC III, Roe F, Bugnicourt A, Franklin MJ & Stewart PS
(2003) Contributions of antibiotic penetration, oxygen
limitation, and low metabolic activity to tolerance of
Pseudomonas aeruginosa biofilms to ciprofloxacin and
tobramycin. Antimicrob Agents Ch 47: 317–323.
Webb JS, Thompson LS, James S, Charlton T, Tolker-Nielsen T,
Koch B, Givskov M & Kjelleberg S (2003) Cell death in
Pseudomonas aeruginosa biofilm development. J Bacteriol 185:
4585–4592.
Werner E, Roe F, Bugnicourt A, Franklin MJ, Heydorn A, Molin
S, Pitts B & Stewart PS (2004) Stratified growth in
Pseudomonas aeruginosa biofilms. Appl Environ Microb 70:
6188–6196.
West SA, Diggle SP, Buckling A, Gardner A & Griffin AS (2007)
The social lives of microbes. Annu Rev Ecol Evol S 38:
53–77.
Whitchurch CB, Tolker-Nielsen T, Ragas PC & Mattick JS (2002)
Extracellular DNA required for bacterial biofilm formation.
Science 295: 1487.
Winzer K, Falconer C, Garber NC, Diggle SP, Camara M &
Williams P (2000) The Pseudomonas aeruginosa lectins PA-IL
and PA-IIL are controlled by quorum sensing and by RpoS.
J Bacteriol 182: 6401–6411.
Worlitzsch D, Tarran R, Ulrich M et al. (2002) Effects of reduced
mucus oxygen concentration in airway Pseudomonas
infections of cystic fibrosis patients. J Clin Invest 109: 317–325.
Xu KD, Stewart PS, Xia F, Huang CT & McFeters GA (1998)
Spatial physiological heterogeneity in Pseudomonas aeruginosa
biofilm is determined by oxygen availability. Appl Environ
Microb 64: 4035–4039.
Yang L, Barken KB, Skindersoe ME, Christensen AB, Givskov M
& Tolker-Nielsen T (2007) Effects of iron on DNA release and
biofilm development by Pseudomonas aeruginosa. Microbiology
153: 1318–1328.
Yang L, Nilsson M, Gjermansen M, Givskov M & Tolker-Nielsen
T (2009) Pyoverdine and PQS mediated subpopulation
interactions involved in Pseudomonas aeruginosa biofilm
formation. Mol Microbiol 74: 1380–1392.
Yoon SS, Hennigan RF, Hilliard GM et al. (2002) Pseudomonas
aeruginosa anaerobic respiration in biofilms: relationships to
cystic fibrosis pathogenesis. Dev Cell 3: 593–603.
Zhang L & Mah TF (2008) Involvement of a novel efflux system
in biofilm-specific resistance to antibiotics. J Bacteriol 190:
4447–4452.
Zhang Y & Miller RM (1994) Effect of a Pseudomonas
rhamnolipid biosurfactant on cell hydrophobicity and
biodegradation of octadecane. Appl Environ Microb 60:
2101–2106.
FEMS Immunol Med Microbiol 59 (2010) 253–268c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
268 M. Harmsen et al.





























