Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/foodchemtox
Food and Chemical Toxicology 46 (2008) 270–279
An investigation of the effects of methylmercury in rats feddifferent dietary fats and proteins: Testicular steroidogenic
enzymes and serum testosterone levels
Mark J. McVey a, Gerard M. Cooke a,b,c,*, Ivan H.A. Curran a, Hing Man Chan d,Stan Kubow e, Eric Lok a, Rekha Mehta a
a Toxicology Research Division, Health Products and Foods Branch, Food Directorate, Health Canada, Sir Fredrick G. Banting Research Centre,
2202D1 Tunney’s Pasture, Ottawa, ON, Canada K1A 0L2b Department of Cellular and Molecular Medicine, Reproductive Biology Unit, University of Ottawa, Canada
c Department of Obstetrics and Gynecology, University of Ottawa, Canadad Centre for Indigenous Peoples’ Nutrition and Environment (CINE), Macdonald Campus of McGill, University 21, 111 Lakeshore Road,
Ste-Anne-de-Bellevue, Quebec, Canada H9X 3V9e School of Dietetics and Human Nutrition, Macdonald Campus of McGill, University 21, 111 Lakeshore Road,
Ste-Anne-de-Bellevue, Quebec, Canada H9X 3V9
Received 15 March 2007; accepted 6 August 2007
Abstract
Methylmercury (MeHg) is a testicular toxicant causing reduced steroidogenic enzyme activity, reduced serum testosterone (T) andabnormal spermatogenesis in mammals and fowl. It is also known that certain diets can alter androgen metabolism in rats. Previouslywe have shown that diets used in the current study impact circulating androgen levels and testicular steroidogenic enzyme activities inSprague Dawley rats in the absence of MeHg. In the present study, we have investigated the impact of imposing an environmental con-taminant (MeHg) commonly found in marine mammals and fish onto the rats’ dietary intake of different proteins and lipids in order todetermine if the different diets could modify MeHg toxicity in rats. Therefore, we examined the effects of MeHg on testicular steroido-genic enzymes and serum testosterone in rats fed diets containing either different protein sources (casein, fishmeal, whey) or different lipidsources (soybean oil, docosahexaenoic acid (DHA), seal oil, fish oil, lard). Male rats 42–45 days of age (18 per group) were assigned todifferent experimental diets for 28 days after which 6 rats in each group were gavaged daily with 0, 1 or 3 mg/kg body weight (BW)/dayMeHg chloride in 5 mM Na2CO3 solution for 14 days while being maintained on their diets. On the 43rd day of dosing, rats weresacrificed and blood plasma and testes frozen (�80 �C) until analysis. Microsomal steroidogenic enzyme activities (3b-HSD, 17-OHase,C-17, 20-lyase, 17b-HSD) were measured radiometrically. Serum testosterone was determined using ELISA kits. Testis weights were notaffected by MeHg. MeHg at 3 mg/kg BW/day caused a reduction (>50%) in the activity of C-17, 20-lyase in all three protein diets andsimilar reductions in 17-OHase activity were seen in the casein and whey protein fed rats. At 3 mg/kg BW/day, MeHg reduced 17-OHaseactivity in the DHA diet but had no effect on 3b-HSD activity and no inhibitory effects on 17b-HSD activity. MeHg (3 mg/kg BW/day)caused significant reductions in serum T in the whey, soybean oil and fish oil groups. Interestingly, fishmeal protein but not fish oiloffered some protection with respect to maintaining steroidogenic enzyme activities and serum T levels in rats dosed with MeHg. In con-clusion, these studies show that different lipid diets can alter the toxic effects of MeHg on male rat steroidogenesis in terms of serumtestosterone and steroidogenic enzyme activities.Crown Copyright � 2007 Published by Elsevier Ltd. All rights reserved.
0278-6915/$ - see front matter Crown Copyright � 2007 Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.fct.2007.08.004
* Corresponding author. Address: Toxicology Research Division, Health Products and Foods Branch, Food Directorate, Health Canada, Sir FredrickG. Banting Research Centre, 2202D1 Tunney’s Pasture, Ottawa, ON, Canada K1A 0L2. Tel.: +1 613 957 0990; fax: +1 613 941 6959.
E-mail address: [email protected] (G.M. Cooke).

M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279 271
Keywords: Methylmercury; Rat; Testis; Steroidogenesis; Serum testosterone; Dietary proteins and lipids
1. Introduction
Mercury is an industrial pollutant from coal-fired utilityplants and waste incinerators. Methyl mercury (MeHg) hasbeen used as a fungicide, topical antiseptic, disinfectantand as an intermediate in the production of other chemi-cally valuable compounds. The human health-risks associ-ated with MeHg exposure were tragically made evidentwith poisoning epidemics such as contaminated fish inJapan and contaminated grain in Iraq, where exposuresled to neurological abnormalities, motor damage, ataxia,seizures and developmental retardation (Marsh et al.,1987; Harada, 1995).
Besides acute poisoning by inhalation or ingestion,humans tend to encounter mercury (Hg) converted byaquatic-bacteria to MeHg, which accumulates in foodssuch as seafood and marine mammals (National ResearchCouncil, 2000; Wagemann et al., 2000). There have beennumerous studies where the levels of MeHg in humansconsuming contaminated fish have been determined (Chanet al., 1995; Grandjean and White, 1999; Wagemann et al.,2000; Dolbec et al., 2001; Davidson et al., 2001; Muckleet al., 2001a,b; Myers et al., 2003; Stern, 2005) revealingthat northern populations such as the Inuit, may beexposed to higher levels of mercury from their diet. Fish,beluga fat/muktuk, seal meat and fat are commonly con-sumed foods in northern communities and are sources ofboth harmful compounds such as MeHg and beneficialnutrients such as proteins, oils, minerals and vitamins. Lev-els of mercury above Health Canada consumer guidelines(0.5 lg/g wet weight) have been reported in terrestrialfreshwater and marine animals that are important to thetraditional Inuit diet in the Canadian North (Chan et al.,1995; Chan, 1998; Crump et al., 1998) and advisories havebeen issued to the general Canadian population regardingthe consumption of fish contaminated with MeHg(www.hc-sc.gc.ca/ewh-semt/pubs/contaminants/risk-ris-que/mercur/q12-q17_e.html#q-14). Longitudinal studies offish-eating populations exposed to MeHg have been con-ducted in New Zealand and the Faroe Islands and cogni-tive and neuromotor deficits were found in children thatwere exposed prenatally (Crump et al., 1998; Grandjeanet al., 1997). In contrast, this was not observed in a studyconducted in the Republic of Seychelles (Myers et al.,2003).
Mercury has been shown to be toxic to cells of the testis,resulting in impaired production of sex steroids (Ng andLiu, 1990). Specifically, mercury lowers the activity of Ley-dig cell enzymes involved with steroidogenesis and mem-brane function (Vachhrajani and Chowdhury, 1990;Ramalingam and Vimaladevi, 2002). Sex steroids areneeded for spermatogenesis and mercury and MeHg have
been shown to have negative impacts on sperm productionin fowl and on serum T levels and spermatozoa in rats(Burton and Meikle, 1980; Chowdhury et al., 1985,1989a,b; Maretta et al., 1995; Ramalingam et al., 2001,2003). Experiments in vitro with human sperm haverevealed decreased motility after acute contact with20 lM MeHg (Ernst and Lauritsen, 1991).
Steroidogenesis in the male rat is regulated by hypotha-lamic gonadotropin releasing hormone (GnRH) whichstimulates the release of pituitary luteinizing hormone(LH), which in turn binds to Leydig cell LH surface recep-tors (LHR) and up-regulates androgen production. LHoffers longer-term regulation of androgen production bytranscriptional regulation of genes important for the pro-duction of sex steroids such as enzymes and transport pro-teins (Payne and Youngblood, 1995). The biosynthesis ofandrogens in the rat testis involves a stepwise pathway ofenzyme-dependent modifications of cholesterol in Leydigcells (for review see Payne and Youngblood, 1995),involving transport proteins such as steroidogenic acuteregulatory protein (STAR) and steroidogenic enzymes(reviewed in Miller and Strauss, 1999). Pregnenolone,formed from cholesterol by the mitochondrial cholesterolside chain cleavage enzyme (P450 SCC), is converted totestosterone by enzymes in the smooth endoplasmic reticu-lum; 3b-hydroxysteroid dehydrogenase (3b-HSD), 17-hydroxylase/C-17,20 lyase (17OHase/C17,20-lyase) and17b-hydroxysteroid dehydrogenase (17b-HSD). In the rattestis, pregnenolone is converted sequentially to progester-one, 17-hydroxyprogesterone, androstenedione and finallyT (Hojbjerg et al., 1992). Testosterone is important forforming the male reproductive tract, spermatogenesis,maintenance of secondary sex characteristics and other sex-ual parameters such as libido (Huhtaniemi, 1995; Sprandoand Zirkin, 1997; Hoeben et al., 1999; Parker et al., 1999;McLachlan et al., 2002). Impairment in the production ofT can adversely impact reproduction by impairing spermproduction among other effects (Kim et al., 2004).
Previously we investigated the effects of altering dietarylipids and proteins on Sprague Dawley (SD) rat testicularandrogen production and circulating serum T levels(McVey et al., 2008). This work was inspired by other stud-ies where altered male rat androgen metabolism was causedby dietary lipids (Gromadzka-Ostrowska et al., 2002).There have been few efforts made to determine how differ-ent dietary lipids and proteins impact the toxicity of envi-ronmental pollutants such as MeHg, which is known todecrease circulating androgens and alter testicular function(Burton and Meikle, 1980; Vachhrajani and Chowdhury,1990; Homma-Takeda et al., 2001; McLachlan et al.,2002). We have tested the hypothesis that different lipidand protein diets would alter the toxicity of MeHg (0, 1

272 M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279
or 3 mg/kg body weight (BW)/day) with respect to serumtestosterone levels and steroidogenic enzyme activities.This hypothesis is based on research suggesting that a vari-ety of dietary factors such as selenium, zinc, cysteine, pro-teins, fats, fibers and vitamins modulate MeHg toxicity(Parizek and Ostadalova, 1967; Chapman and Chan,2000). Animal studies have shown that different dietary oilsand proteins cause variable MeHg retention and toxicity(Kling and Soares, 1981; Rowland et al., 1986; Hojbjerget al., 1992). Modulating the lipids in diets can alter sex-ratios of offspring, altered hypothalamic–pituitary–adrenalsignalling, testicular steroidogenic enzyme activities,steroid levels and growth regulation in animal studies(Staples et al., 1998; Gromadzka-Ostrowska et al., 2002;Rosenfeld et al., 2003; Taylor et al., 2003; Funston, 2004;La Fleur et al., 2005). Furthermore, correlations betweenfatty diets and steroid hormone levels in human serum havealso been reported (Dorgan et al., 2003). To date, no stud-ies have examined the impact of MeHg on testicular steroi-dogenesis and circulating androgens in rats fed dietscontaining different lipids and proteins. The potential risksor benefits of different proteins (casein, whey and fishmeal)as well as different lipids (soybean oil, DHA, seal oil, fishoil and lard) in diets were examined previously in termsof the reproductive toxicological endpoints of testisweights, serum testosterone levels, and microsomal steroi-dogenic enzyme activities in rats (McVey et al., 2008).Here, we now investigate the effects of these dietary compo-nents on MeHg toxicity (0, 1 or 3 mg/kg BW/day). Thedoses of MeHg used were similar to concentrations ofMeHg found in seafoods consumed by humans, as manyfish contain roughly 1 to 3 mg/kg BW MeHg (Kosatskyet al., 2000; Muckle et al., 2001a; Sing et al., 2003; Johns-son et al., 2004), and certain populations consume largequantities of such fish. The following study intended toinvestigate if different lipids and proteins in rat diets wouldalter the toxicity of MeHg at either 0, 1, 3 mg/kg BW/day.We hypothesize that the composition of the diets fed to ratswill alter toxic effects of MeHg on serum testosterone levelsand steroidogenic enzyme activities based on our previousexamination of these parameters in the absence of MeHgshowing changes in serum testosterone and steroidogenicenzyme activities (McVey et al., 2008). The findings willbe useful for characterisation of the health risk of mercuryin the Inuit and other diets containing large amounts of fishand marine mammals.
2. Experimental
2.1. Materials
Unlabelled steroids were purchased from Steraloids Inc. (Newport,RI). Aquasol and labelled steroids: [1,2,6,7-3H] DHEA (60.0 Ci/mmol),[1,2,6,7-3H] progesterone (114.4 Ci/mmol), [1,2,6,7-3H], [1,2,6,7-3H] 17-hydroxy-progesterone (57.0 Ci/mmol) and [1,2,6,7-3H] androstenedione(74.0 Ci/mmol) were bought from Dupont/NEN (Boston, MA). Organicsolvents were purchased from EM Science Merck KgaA (Darmstadt,Germany). Plastic coated WhatmanTM PE SIL G silica gel chromatography
plates were bought from Chromatographic Specialties (Montreal, QC).TRIS, nicotinamide cofactors (NADPH and NAD+), and b-mercap-toethanol were purchased from Sigma Chemical Co. (St Louis, MO).Sucrose was purchased from BDH Inc., (Toronto, ON). Fish oil andomega protein silver herring low ash menhaden fish meal were obtainedfrom Omega-protein (Hammond, Louisiana, USA), DHA oil from Mar-tek Corporation (Columbia, MD, USA), Seal oil from Atlantis MarineInc. (St. John’s, NF), methyl mercury chloride CAS# 115-09-3 (>95%pure) purchased from Alfa Aesar (Ward Hill, MA), vitamin free caseinprotein bought from ICN Biochemicals Inc. (Cleveland, OH) and wheyprotein bought from Avonmore Waterford (Provon 190 whey proteinisolate) were combined to form the experimental diets.
2.2. Animals
Animal care and handling conformed to the guidelines of The Cana-dian Council on Animal Care and the protocols were approved by theHealth Canada Ottawa Animal Care Committee. Male SD rats (42–45days of age) (Charles River Canada, St. Constant, Quebec, Canada) wereacclimatized on a starch-based, semi-purified basal diet containing cornstarch and casein for 5–10 days, before being placed on experimental dietsin glove box style Isotec units (Harlan Sprague Dawley Inc., Indiana)under negative pressure with exhausted ventilation. After acclimatization,rats were randomly assigned to eight different diet groups of 18 animals.Throughout the study, each group of rats was fed an isocaloric dietcontaining 15% by weight (15%/wt) of different fats (lard, fish oil, seal oil,DHA) added to the basal starch-based, semi-purified corn starch andvitamin free casein diet, or different protein diets with corn oil, soybeanoil, lard and butter lipids (McVey et al., 2008). Peroxide value, vitamin Econtent (mg/100 ml), selenium content (mg/kg), mercury levels (ppm), andfatty acid composition (SFA, MUFA, PUFA, omega 3 and 6 content,EPA and DHA) (g/100 g lipid) were determined previously (Jin et al.,2007). For inter-diet comparisons, the casein diet was chosen as a controldiet. This casein diet was designed according to guidelines recommendedby the National Research Council (NRC) nutrient requirement for labo-ratory rats (NRC, 1995) with modifications in fat to mimic human diets(Atwal et al., 1997). The experimental lipid diets used in the present studycontained large amounts of a single lipid source, while the casein controldiet was created with a combination of various lipid sources as opposed toa single type of lipid simulating typical commercial casein based rat chowdiets such as AIN-93G. All diets contained a minimal level of soy oil (1.6%of the total diet) to provide essential fatty acid requirements for rats(NRC, 1995). The current study was a satellite study to a larger project,which was the reason why the casein, whey and fishmeal diets containing70 g soy oil/kg diet as opposed to the 16 g soy oil/kg diet seen in the lipidgroups. Commercial casein based rat chows, such as AIN-93G, containapproximately 70 g soy oil/kg diet and have 7% total lipids.
Rats were given free access to food and water ad libitum throughoutthe study. Food and water consumption were monitored twice a week,while body weights were recorded daily. On the 28th day of exposure tothe experimental diets, rats within each diet group were randomly assignedto 3 separate treatment groups of 6 animals and were gavaged daily for 14days with 5 mM Na2CO3, solution containing sufficient MeHg chloride togenerate a final oral dose of 0, 1 or 3 mg MeHg/kg BW/day. On the 43rdday following the commencement of the experimental diets, rats werebetween 89 and 97 days of age, and were sacrificed following abdominalaorta exsanguination under isoflurane anaesthesia. Serum was collectedand stored at �80 �C until analysis and testes were weighed and imme-diately frozen in liquid nitrogen at time of necropsy and stored at �80 �Cuntil assayed.
2.3. Serum androgen quantitation
Serum T assays on 4–6 rats were conducted in duplicate for each dietgroup and dose of MeHg using commercial ELISA kits (IBL, HamburgCat# RE521-51). The cross-reactivity of dihydrotestosterone (DHT) in the

M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279 273
T assay and inter- and intra-assay coefficients of variation (CV%) forserum T were reported previously (McVey et al., 2008).
2.4. Testicular microsomal enzyme activities
Testicular microsomal (3b-HSD, 17-OHase, C-17, 20-lyase and 17b-HSD) steroidogenic enzyme activities were assayed according to themodified methods of Cooke et al. (1997, 1998) from testes of each diet andMeHg dose. Testicular microsomes were prepared and assayed identicallyas reported previously (McVey et al., 2008). Microsomal proteinconcentrations were determined using the method of Lowry et al. (1951).
2.5. Statistical analysis
Intra- and inter-assay coefficients of variation (CV%) for ELISAassays were experimentally derived from the results (standard deviation/mean · 100%). Linear and non-linear regressions were carried out usingGraphPad Prism V3.02 1994–2000. (Graphpad Software Inc., San Diego,CA). One-way ANOVA was used to compare the effects of proteins andlipid diets on enzyme activities and microsomal protein concentrations(independent of dose of MeHg), followed by Bonferroni’s tests (usingcasein as the control). Two-way ANOVA was used for analysis ofmicrosomal protein concentrations at different doses of MeHg, andactivities of 3b-HSD, 17-OHase, C-17,20 lyase and 17b-HSD, followed byBonferroni’s t-tests used for multiple comparisons (Sigma Stat V2.031992–1997 for windows (SPSS Inc., Chicago, IL)). Tests for normality(Kolmogorov–Smirnov) and equal variance (Levene median Test) forserum testosterone and microsomal steroidogenic enzymes were passed for
Effects of Methyl Mercury on Testosterone
0.0 1.0 3.0 0.0 1.0 30
1
2
3
Me-Hg (mg/Kg Bw
Seru
m T
esto
ster
one
(ng/
ml)
Effects of Methyl Mercury onTestosterone
0 1 3 0 1 3 0 1 3 00
1
2
3
4
*
a
Me-Hg (mg/Kg B
Seru
m T
esto
ster
one
(ng/
ml)
Fig. 1. The effects of MeHg (0, 1 or 3 mg/kg body weight (BW)/day) on serfishmeal and whey protein based diets (a) or soybean oil, DHA, seal, fish aBonferoni’s t-tests) showed a significant decrease in serum T for 3 mg/kg BW/d(P < 0.05). Six animals were sampled per group with the exceptions of 3 mg/kg1 mg/kg BW (DHA and fish oil) and 3 mg/kg BW/day (casein and lard) n = 5
the present study. The Grubb’s test was used for the detection of outliersfor serum androgen levels (Grubbs, 1969).
3. Results
3.1. Body and testis weights
There were no differences in body weights or testisweights caused by MeHg (P > 0.05). MeHg had no effecton microsomal protein concentration except for the soy-bean oil diet, where the dose of 1.0 mg/kg BW/day was sig-nificantly higher than both 0.0 and 3.0 mg/kg/day but thesedoses were not significantly different from each other(28.6 ± 2.0, 35.1 ± 1.7 and 27.1 ± 1.9 mg/ml, respectively)(P < 0.05).
3.2. Serum androgens
Serum T findings in the absence of MeHg are reportedelsewhere (McVey et al., 2008). For the protein and lipiddiets examined all 1 mg/kg BW/day MeHg serum T levelswere not significantly changed from 0 mg/kg. At 3.0 mg/kg BW/day, MeHg significantly reduced serum T in thewhey, soybean oil and fish oil groups (Fig. 1).
Rat Serum
.0 0.0 1.0 3.0
Fishmeal
Whey
Casein
*
t/day)
Rat Serum
1 3 0 1 3 0 1 3
Soy
DHA
Casein
Seal
Fish
Lard
*
wt/day)
um testosterone (T) (ng/ml, mean ± SE) in male rats consuming casein,nd lard diet groups (b). Statistical analysis (2 way-ANOVA followed byay compared with 0 mg/kg BW/day (casein, whey, soy, DHA and seal) (*)BW/day (fishmeal) n = 4; 0 mg/kg BW/day (fishmeal, whey and fish oil),and 0 mg/kg BW/day (lard) n = 7.

274 M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279
3.3. 3b-hydroxysteriod dehydrogenase activity
The 3b-HSD activities in the different diet groups arepresented in Figs. 2a and 3a. There were no significanteffects of MeHg on 3b-HSD activity at any of the dosesexamined.
3.4. 17-OHase activity
MeHg at 3 mg/kg BW/day caused a significant reduc-tion (>50%) in 17-OHase activities in the casein and wheygroups (Fig. 2b). Within the different oil diets, MeHg at3 mg/kg BW/day reduced 17-OHase activity in the DHAdiet only (Fig. 4a).
3.5. C-17,20-lyase activity
There were no differences in C-17,20 lyase activitiesbetween the casein, fishmeal and whey protein diets exceptat 3 mg/kg BW/day, where MeHg caused a significantreduction (>50%) in activity (Fig. 2c). The effect of MeHgin the different lipid diets was minimal and no significanteffects were evident (Fig. 4b).
3.6. 17b-hydroxysteroid dehydrogenase activity
There were no MeHg dependent changes in 17b-HSDactivities reported for the protein diets (Fig. 2d). No inhib-
Effects of Methyl Mercury on Rat TestisMicrosomal 3ß-HSD Activity
0.0 1.0 3.0 0.0 1.0 3.0 0.0 1.0 3.00
200
400
600
800 FishmealWhey
Casein
Me-Hg (mg/Kg Bwt/day)
3β-H
SD A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Effects of Methyl Mercury on Rat TestisMicrosomal C17,20-Lyase Activity
0.0 1.0 3.0 0.0 1.0 3.0 0.0 1.0 3.00
10
20
30FishmealWhey
Casein
* * *
Me-Hg (mg/Kg Bwt/day)
C17
,20-
Lyas
e A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Fig. 2. (a) 3b-Hydroxysteroid dehydrogenase (3b-HSD) activity (pmol DHEAmin/mg protein), (c) C17,20-lyase (pmol androstenedione/min/mg protein)testosterone/min/mg protein) (mean ± SE) in testicular microsomes from rashowed a significant decrease in enzyme activity for: 17-OHase 3 mg/kg bodywell as 3 mg/kg BW/day (whey) compared with 0 mg/kg BW/day (whey) (*); anday within each dietary protein group (casein, fishmeal and whey) (*), (P < 0.05day (casein and fishmeal) [3b-HSD, 17-OHase, C17,20-lyase, 17b-HSD] n = 4kg BW/day (fishmeal) [C17,20-lyase], 0 mg/kg BW/day (whey) [17-OHase], 0 mday (whey) [17b-HSD] n = 5.
itory effects of MeHg were observed for lipid diet fed ratsbut, in fact, increased 17b-HSD activity was evident inthe fish and lard diets at 0 and 1 mg/kg BW/day MeHg,and in the DHA diet at 3.0 mg/kg BW/day (Fig. 3b).
4. Discussion
MeHg is a prominent contaminant in fish from varioussources and has a half-life of approximately 45–55 days inhumans (reviewed in Smith and Farris, 1996). Conse-quently, dietary exposure to MeHg is a serious health con-cern particularly among fish-eating communities and hasbeen linked to adverse neurological and reproductiveeffects (Grandjean et al., 1997; Crump et al., 1998). Thecurrent study has examined the effects of different dietaryprotein and lipid sources on the toxicity of MeHg towardstesticular weights, serum T levels and testicular steroido-genic enzyme activities in male rats.
Various fats present in the diets used in these studieshave been shown to be advantageous in terms of their pro-ven roles in development (DHA, fishmeal, soybeans)(Sakamoto et al., 2004), or in altering the onset or symp-toms of various diseases and cancers (fish oils, soybean oils,soy proteins and whey proteins) (Simophoulos, 2002; Car-roll and Roth, 2002; Guallar et al., 2002; Rowlands et al.,2002; Li, 2003; Dorfman et al., 2005; Xiao et al., 2005).Alternatively, lard has been shown to cause heightened riskof cancer and cardiac disease (Reddy, 1987; Kesterloot
Effects of Methyl Mercury on Rat TestisMicrosomal 17-OHase Activity
0.0 1.0 3.0 0.0 1.0 3.0 0.0 1.0 3.00
10
20
30
40
50FishmealWhey
Casein
* *
Me-Hg (mg/Kg Bwt/day)
17-O
Has
e A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Effects of Methyl Mercury on Rat TestisMicrosomal 17β-HSD Activity
0.0 1.0 3.0 0.0 1.0 3.0 0.0 1.0 3.00
10
20 FishmealWhey
Casein
Me-Hg (mg/Kg Bwt/day)
17β-
HSD
Act
ivity
pmol
/min
/mg
prot
ein
/min/mg protein), (b) 17-OHase activity (pmol 17-hydroxyprogesterone/and (d) 17b-Hydroxysteroid dehydrogenase (17b-HSD) activity (pmol
ts. Statistical analysis (2 way-ANOVA followed by Bonferroni’s t-tests)weight (BW)/day (casein) compared with 0 mg/kg BW/day (casein) (*) asd C17,20-lyase activity for 3 mg/kg BW/day compared with 0 mg/kg BW/). Six animals were sampled per group with the exceptions of 3 mg/kg BW/; 0 mg/kg BW/day (fishmeal) [17-OHase, C17,20-lyase, 17b-HSD], 1 mg/g/kg BW/day + 3 mg/kg BW/day (whey) [C17,20-lyase] and 0 mg/kg BW/
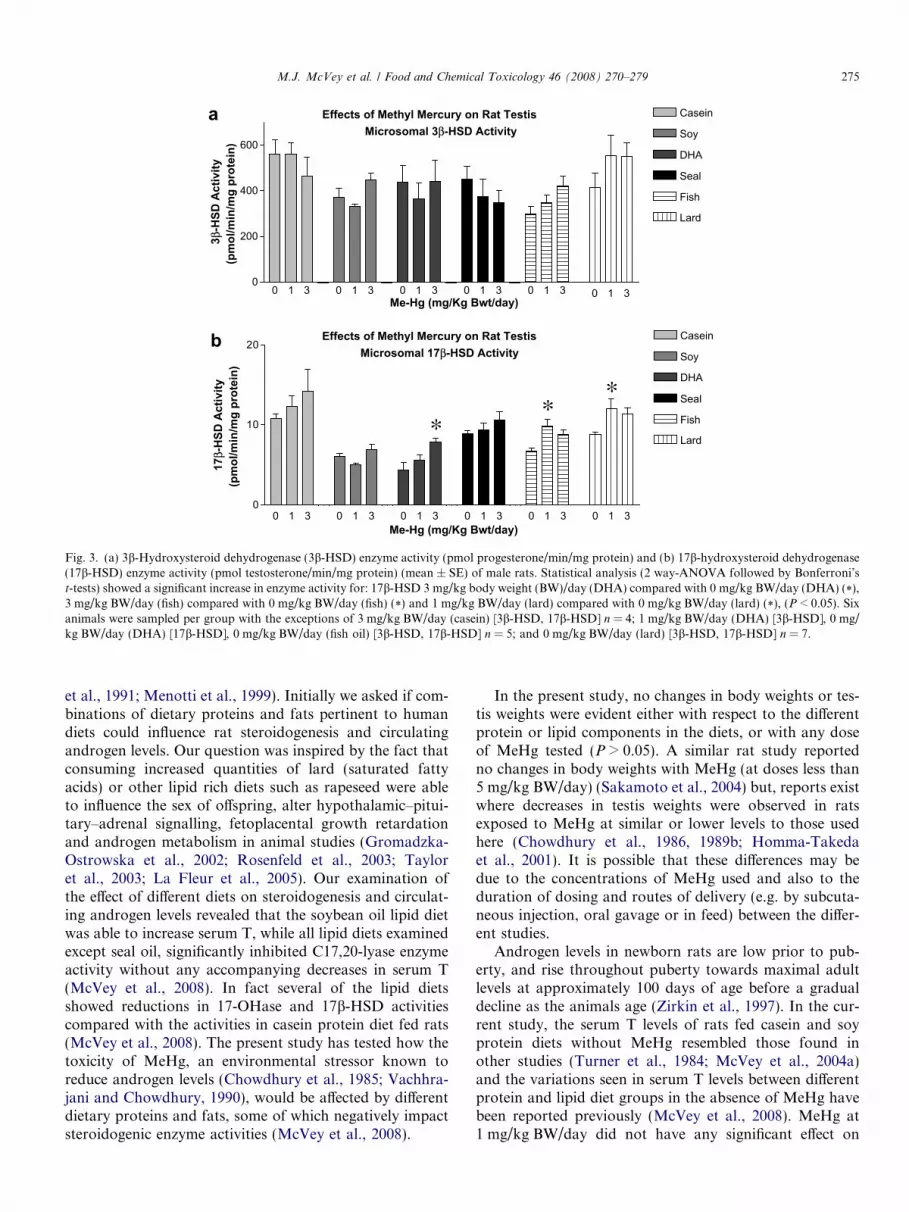
Effects of Methyl Mercury on Rat TestisMicrosomal 3β-HSD Activity
0 1 3 0 1 3 0 1 3 0 1 3 0 1 3 0 1 30
200
400
600Soy
DHA
Casein
Seal
Fish
Lard
Me-Hg (mg/Kg Bwt/day)
3β-H
SD A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Effects of Methyl Mercury on Rat TestisMicrosomal 17β-HSD Activity
0 1 3 0 1 3 0 1 3 0 1 3 0 1 3 0 1 30
10
20Soy
DHA
Casein
*Seal
Fish
Lard
**
Me-Hg (mg/Kg Bwt/day)
17β-
HSD
Act
ivity
(pm
ol/m
in/m
g pr
otei
n)
Fig. 3. (a) 3b-Hydroxysteroid dehydrogenase (3b-HSD) enzyme activity (pmol progesterone/min/mg protein) and (b) 17b-hydroxysteroid dehydrogenase(17b-HSD) enzyme activity (pmol testosterone/min/mg protein) (mean ± SE) of male rats. Statistical analysis (2 way-ANOVA followed by Bonferroni’st-tests) showed a significant increase in enzyme activity for: 17b-HSD 3 mg/kg body weight (BW)/day (DHA) compared with 0 mg/kg BW/day (DHA) (*),3 mg/kg BW/day (fish) compared with 0 mg/kg BW/day (fish) (*) and 1 mg/kg BW/day (lard) compared with 0 mg/kg BW/day (lard) (*), (P < 0.05). Sixanimals were sampled per group with the exceptions of 3 mg/kg BW/day (casein) [3b-HSD, 17b-HSD] n = 4; 1 mg/kg BW/day (DHA) [3b-HSD], 0 mg/kg BW/day (DHA) [17b-HSD], 0 mg/kg BW/day (fish oil) [3b-HSD, 17b-HSD] n = 5; and 0 mg/kg BW/day (lard) [3b-HSD, 17b-HSD] n = 7.
M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279 275
et al., 1991; Menotti et al., 1999). Initially we asked if com-binations of dietary proteins and fats pertinent to humandiets could influence rat steroidogenesis and circulatingandrogen levels. Our question was inspired by the fact thatconsuming increased quantities of lard (saturated fattyacids) or other lipid rich diets such as rapeseed were ableto influence the sex of offspring, alter hypothalamic–pitui-tary–adrenal signalling, fetoplacental growth retardationand androgen metabolism in animal studies (Gromadzka-Ostrowska et al., 2002; Rosenfeld et al., 2003; Tayloret al., 2003; La Fleur et al., 2005). Our examination ofthe effect of different diets on steroidogenesis and circulat-ing androgen levels revealed that the soybean oil lipid dietwas able to increase serum T, while all lipid diets examinedexcept seal oil, significantly inhibited C17,20-lyase enzymeactivity without any accompanying decreases in serum T(McVey et al., 2008). In fact several of the lipid dietsshowed reductions in 17-OHase and 17b-HSD activitiescompared with the activities in casein protein diet fed rats(McVey et al., 2008). The present study has tested how thetoxicity of MeHg, an environmental stressor known toreduce androgen levels (Chowdhury et al., 1985; Vachhra-jani and Chowdhury, 1990), would be affected by differentdietary proteins and fats, some of which negatively impactsteroidogenic enzyme activities (McVey et al., 2008).
In the present study, no changes in body weights or tes-tis weights were evident either with respect to the differentprotein or lipid components in the diets, or with any doseof MeHg tested (P > 0.05). A similar rat study reportedno changes in body weights with MeHg (at doses less than5 mg/kg BW/day) (Sakamoto et al., 2004) but, reports existwhere decreases in testis weights were observed in ratsexposed to MeHg at similar or lower levels to those usedhere (Chowdhury et al., 1986, 1989b; Homma-Takedaet al., 2001). It is possible that these differences may bedue to the concentrations of MeHg used and also to theduration of dosing and routes of delivery (e.g. by subcuta-neous injection, oral gavage or in feed) between the differ-ent studies.
Androgen levels in newborn rats are low prior to pub-erty, and rise throughout puberty towards maximal adultlevels at approximately 100 days of age before a gradualdecline as the animals age (Zirkin et al., 1997). In the cur-rent study, the serum T levels of rats fed casein and soyprotein diets without MeHg resembled those found inother studies (Turner et al., 1984; McVey et al., 2004a)and the variations seen in serum T levels between differentprotein and lipid diet groups in the absence of MeHg havebeen reported previously (McVey et al., 2008). MeHg at1 mg/kg BW/day did not have any significant effect on
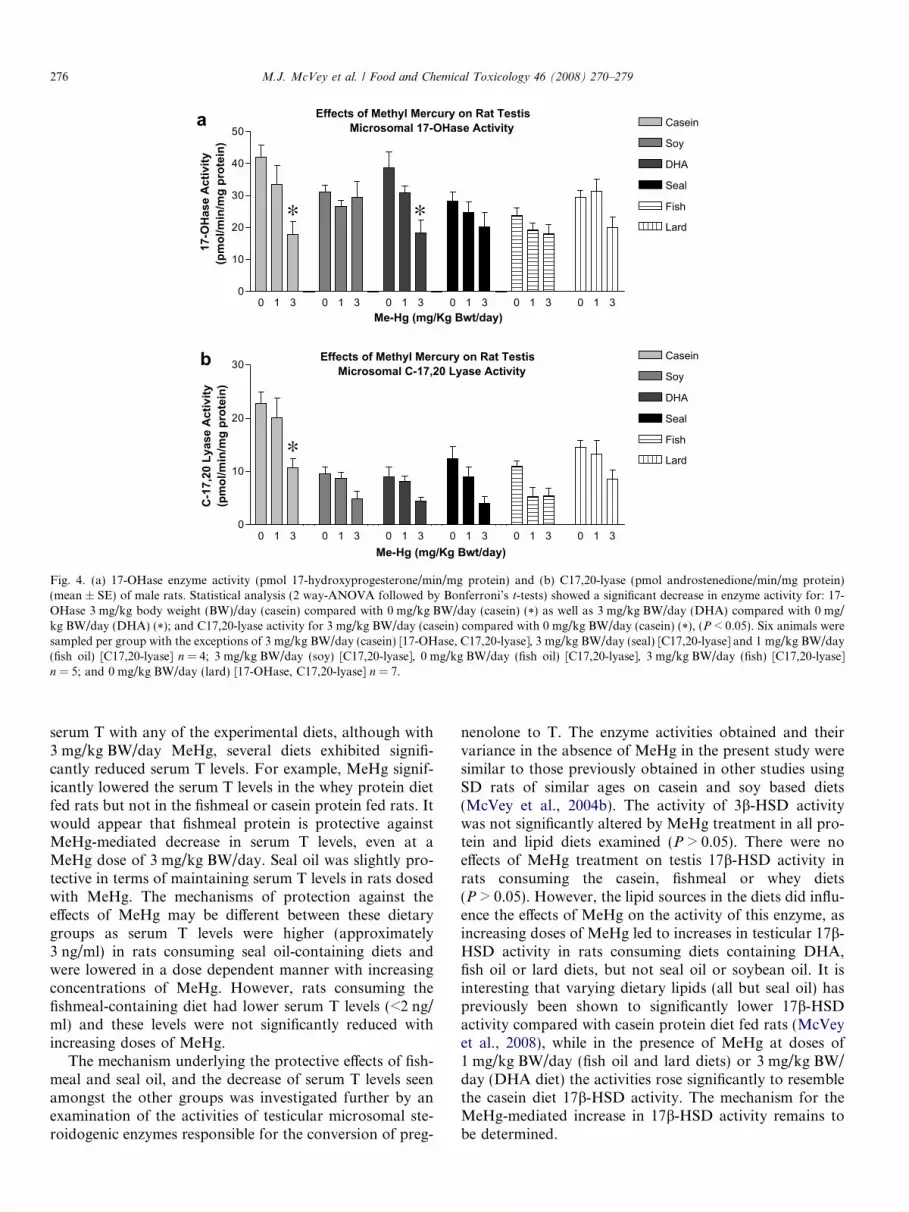
Effects of Methyl Mercury on Rat TestisMicrosomal 17-OHase Activity
0 1 3 0 1 3 0 1 3 0 1 3 0 1 3 0 1 30
10
20
30
40
50Soy
DHA
Casein
*Seal
Fish
Lard*
Me-Hg (mg/Kg Bwt/day)
17-O
Has
e A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Effects of Methyl Mercury on Rat TestisMicrosomal C-17,20 Lyase Activity
0 1 3 0 1 3 0 1 3 0 1 3 0 1 3 0 1 30
10
20
30Soy
DHA
Casein
*
Seal
Fish
Lard
Me-Hg (mg/Kg Bwt/day)
C-1
7,20
Lya
se A
ctiv
ity(p
mol
/min
/mg
prot
ein)
Fig. 4. (a) 17-OHase enzyme activity (pmol 17-hydroxyprogesterone/min/mg protein) and (b) C17,20-lyase (pmol androstenedione/min/mg protein)(mean ± SE) of male rats. Statistical analysis (2 way-ANOVA followed by Bonferroni’s t-tests) showed a significant decrease in enzyme activity for: 17-OHase 3 mg/kg body weight (BW)/day (casein) compared with 0 mg/kg BW/day (casein) (*) as well as 3 mg/kg BW/day (DHA) compared with 0 mg/kg BW/day (DHA) (*); and C17,20-lyase activity for 3 mg/kg BW/day (casein) compared with 0 mg/kg BW/day (casein) (*), (P < 0.05). Six animals weresampled per group with the exceptions of 3 mg/kg BW/day (casein) [17-OHase, C17,20-lyase], 3 mg/kg BW/day (seal) [C17,20-lyase] and 1 mg/kg BW/day(fish oil) [C17,20-lyase] n = 4; 3 mg/kg BW/day (soy) [C17,20-lyase], 0 mg/kg BW/day (fish oil) [C17,20-lyase], 3 mg/kg BW/day (fish) [C17,20-lyase]n = 5; and 0 mg/kg BW/day (lard) [17-OHase, C17,20-lyase] n = 7.
276 M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279
serum T with any of the experimental diets, although with3 mg/kg BW/day MeHg, several diets exhibited signifi-cantly reduced serum T levels. For example, MeHg signif-icantly lowered the serum T levels in the whey protein dietfed rats but not in the fishmeal or casein protein fed rats. Itwould appear that fishmeal protein is protective againstMeHg-mediated decrease in serum T levels, even at aMeHg dose of 3 mg/kg BW/day. Seal oil was slightly pro-tective in terms of maintaining serum T levels in rats dosedwith MeHg. The mechanisms of protection against theeffects of MeHg may be different between these dietarygroups as serum T levels were higher (approximately3 ng/ml) in rats consuming seal oil-containing diets andwere lowered in a dose dependent manner with increasingconcentrations of MeHg. However, rats consuming thefishmeal-containing diet had lower serum T levels (<2 ng/ml) and these levels were not significantly reduced withincreasing doses of MeHg.
The mechanism underlying the protective effects of fish-meal and seal oil, and the decrease of serum T levels seenamongst the other groups was investigated further by anexamination of the activities of testicular microsomal ste-roidogenic enzymes responsible for the conversion of preg-
nenolone to T. The enzyme activities obtained and theirvariance in the absence of MeHg in the present study weresimilar to those previously obtained in other studies usingSD rats of similar ages on casein and soy based diets(McVey et al., 2004b). The activity of 3b-HSD activitywas not significantly altered by MeHg treatment in all pro-tein and lipid diets examined (P > 0.05). There were noeffects of MeHg treatment on testis 17b-HSD activity inrats consuming the casein, fishmeal or whey diets(P > 0.05). However, the lipid sources in the diets did influ-ence the effects of MeHg on the activity of this enzyme, asincreasing doses of MeHg led to increases in testicular 17b-HSD activity in rats consuming diets containing DHA,fish oil or lard diets, but not seal oil or soybean oil. It isinteresting that varying dietary lipids (all but seal oil) haspreviously been shown to significantly lower 17b-HSDactivity compared with casein protein diet fed rats (McVeyet al., 2008), while in the presence of MeHg at doses of1 mg/kg BW/day (fish oil and lard diets) or 3 mg/kg BW/day (DHA diet) the activities rose significantly to resemblethe casein diet 17b-HSD activity. The mechanism for theMeHg-mediated increase in 17b-HSD activity remains tobe determined.

M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279 277
MeHg significantly decreased 17-OHase activity (casein,whey) and C17,20-lyase activity (casein, whey and fish-meal) by approximately 50%. The effects of MeHg on theseenzyme activities was less dramatic in rats consuming thedifferent lipid diets with significant reductions observed in17-OHase activity only the DHA diet group. C17,20-lyasewas not significantly altered by MeHg treatment in ratsconsuming any of the alternative lipid source diets. Previ-ously we showed this enzyme activity to be significantlydepressed for all lipid diets examined compared with caseincontrols by nearly 50% (McVey et al., 2008) and it ispossible that the lack of changes seen with the additionof MeHg could be attributable to a shared mechanism ofinhibition already maximally affected by a factor presentin certain lipid diets. It is interesting to note that thedecreased C17,20-lyase activity caused by MeHg at 3 mg/kg BW/day in the protein groups was similar to the levelin the lipid diet groups with no MeHg. However, sincethe serum T levels were similar between both the proteinand lipid diet groups, it would appear that the reducedC17,20-lyase activity was still sufficient to maintain normalserum androgen levels. In this context, the apparent protec-tion against MeHg effects on serum T levels offered by thefishmeal protein group would appear to not be directlyrelated to the microsomal steroidogenic enzymes responsi-ble for the conversion of pregnenolone to T.
Other researchers have examined different aspects ofinorganic mercury and MeHg effects on steroidogenic tes-ticular enzymes and have shown varying results. Short-term rat studies using comparable doses of MeHg to thepresent study have shown similar effects on serum T levels(Burton and Meikle, 1980; Vachhrajani and Chowdhury,1990) and also, reduced LH levels, possibly indicating thatMeHg may reduce T production via a non-testicular target(Ramalingam et al., 2003). In the current study the hypo-thalamic pituitary testicular axis was not investigated fur-ther due to limitations on access to tissues and serum.Rat studies with MeHg have shown that suppressed spermproduction is linked with low serum T and increased apop-tosis of germ cells at the spermatogenic stages that areuniquely sensitive to testosterone withdrawal (Homma-Takeda et al., 2001; McLachlan et al., 2002). However, astudy using monkeys showed that MeHg caused abnormal-ities in spermatozoa without any accompanying decrease inserum T (Mohamed et al., 1987). Sperm were not analyzedin the current study. MeHg has been shown to alter plasmamembrane enzymes involved with membrane permeabilityand integrity (Ramalingam and Vimaladevi, 2002), whichcould influence the activities of the steroidogenic enzymes(Machino et al., 1969; Cooke and Robaire, 1988; Cooke,1989). Experiments in fish, have shown increased 3b-HSD mRNA and enzyme activity resulting from the bind-ing of inorganic mercury to plasma membrane ATPases(Mondal et al., 1997). Furthermore, rodents dosed withMeHg or inorganic mercury exhibit reductions in the con-version of cholesterol to pregnenolone, indicative of inhibi-tion of P450 SCC activity (Burton and Meikle, 1980;
Chowdhury et al., 1986). Furthermore, inorganic mercurycan decrease the concentrations of microsomal cytochromeP-450s, which may have led to the steroidogenic enzymeinhibition seen in the present study (Veltman and Maines,1986). The diets used here were tested for inorganic mer-cury and only the seal oil and DHA diets contained Hgand these, at levels less than 2 ppm (Jin et al., 2007).P450 SCC was not examined in the current study, due todifficulties in obtaining suitably radiolabelled cholesterolsubstrates. Rats exposed to MeHg by intraperitoneal (IP)injection at 0.05–10 mg/kg/day for 90 days exhibited Ley-dig cell damage, decreased 3b-HSD activity and decreasedserum T levels (Chowdhury et al., 1985; Vachhrajani andChowdhury, 1990) suggesting that either the oral route ofadministration or the period of dosing in our study didnot allow MeHg to accumulate sufficiently to cause signif-icant inhibition of 3b-HSD activity. Further investigationsare needed to examine changes in factors such as liver Tmetabolism or ABG (androgen binding globulin)/SHBG(sex hormone binding globulin), which could account forthe decreased amounts of circulating T seen in the currentstudy due to MeHg. Additionally an investigation of intra-testicular T levels would be informative as to whetherMeHg affects testicular function directly or actsperipherally.
Based on our earlier studies of the effects of dietaryproteins and lipids on steroidogenic enzyme activities andserum T (McVey et al., 2008), we asked what would bethe effect of adding MeHg, a known contaminant of suchdiets, on enzyme activities and serum T levels. From thesestudies, we conclude that the only diet that offered someprotection against the inhibitory effects of MeHg withrespect to testicular steroidogenic enzyme activities andserum T levels was the diet containing fishmeal as themajor protein source.
Acknowledgements
The authors would like to thank Drs. Xiaolei Jin, MikeWade, Genevieve Bondy, Mrs. Susan Gurofsky and thetechnical staff of Toxicology Research Division and Ani-mal Resources Division of Health Canada.
References
Atwal, A.S., Kubow, S., Wolynetz, M., 1997. Effects of protein source andanimo acid supplementation on plasma cholesterol in guinea pigs. J.Vitam. Nutr. Res. 67, 192–195.
Burton, G.V., Meikle, A.W., 1980. Acute and chronic methyl mercurypoisoning impairs rat adrenal and testicular function. J. Toxicol.Environ. Health 6, 597–606.
Carroll, H.M., Roth, M.T., 2002. Evidence for the cardioprotective effectsof omega-3 fatty acids. Ann. Pharmacol. 36, 1950–1956.
Chan, H.M., 1998. A database for environmental contaminants intraditional foods in northern and arctic Canada. Development andapplication. Food Addit. Contam. 15, 127–134.
Chan, H.M., Kim, C.K., Receveur, O., Kuhnlein, H., 1995. Assessment ofdietary exposures to trace metals in Baffin Inuit food samples. Environ.Health Perspect. 103, 740–746.

278 M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279
Chapman, L., Chan, H.M., 2000. The influence of nutrition on methylmercury intoxication. Environ. Health Perspect. 108, 29–56.
Chowdhury, A.R., Vachharajani, K.D., Chatterjee, B.B., 1985. Inhibitionof 3b-hydroxysteroid dehydrogenase in rat testicular tissue by mercuricchloride. Toxicol. Lett. 27, 45–49.
Chowdhury, A.R., Vachhrajani, K.D., Makhija, S., Kashyaf, S.K., 1986.Histomorphometric and biochemical changes in the testicular tissuesof rats treated with mercuric chloride. Biomed. Biochim. Acta 7, 949–956.
Chowdhury, A.R., Makhija, S., Vachhrajani, K.D., Gautam, A.K., 1989a.Methylmercury- and mercuric-chloride-induced alterations in ratepididymal sperm. Toxicology Lett. 47, 125–134.
Chowdhury, A.R., Makhija, S., Vachhrajani, K.D., 1989b. Methylmer-cury induced biochemical and histochemical alterations in rat testis.Indian J. Physiol. Pharmacol. 33, 218–222.
Cooke, G.M., 1989. Identification and mechanism of action of phospho-lipids capable of modulating rat testicular microsomal 3 beta-hydroxysteroid dehydrogenase-isomerase activity in vitro. Biol.Reprod. 41, 438–445.
Cooke, G.M., Robaire, B., 1988. Phospholipases modulate the rattesticular androgen biosynthetic pathway in vitro. Biol. Reprod. 39,329–339.
Cooke, G.M., Pothier, F., Murphy, B.D., 1997. The effects of progester-one, 4,16-androstadien-3-one and MK-434TM on the kinetics of pigtestis microsomal testosterone-4-ene-5a-reductase activity. J. SteroidBiochem. Molec. Biol. 60, 353–359.
Cooke, G.M., Price, C.A., Oko, R.J., 1998. Effects of In Utero andlactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)on serum androgens and steroidogenic enzyme activities in the male ratreproductive tract. J. Steroid Biochem. Molec. Biol. 67, 347–354.
Crump, K.S., Kjellstrom, T., Shipp, A.M., Silvers, A., Stewart, A., 1998.Influence of prenatal mercury exposure upon scholastic and psycho-logical test performance: benchmark analysis of a New Zealandcohort. Risk Anal. 18, 701–713.
Davidson, P.W., Kost, J., Myers, G.J., Cox, C., Clarkson, T.W.,Shamlaye, C.F., 2001. Methylmercury and neurodevelopment: reanal-ysis of the Seychelles Child Development Study outcomes at 66 monthsof age. JAMA 285, 1291–1293.
Dolbec, J., Mergler, D., Larribe, F., Roulet, M., Lebel, J., Lucotte, M.,2001. Sequential analysis of hair mercury levels in relation to fish dietof an Amazonian population. Sci. Total. Environ. 271, 87–97.
Dorfman, S.E., Wang, S., Vega-Lopez, S., Jauhiainen, M., Lichtenstein,A.H., 2005. Dietary fatty acids and cholesterol differentially modulateHDL cholesterol metabolism in golden-syrian hamsters. J. Nutr. 135,492–498.
Dorgan, J.F., Hunsberger, S.A., McMahon, R.P., Kwiterovich, P.O.,Lauer, R.M., Van Horn, L., Lasser, N.L., Stevens, V.J., Friedman,L.A., Yanovski, J.A., Greenhut, S.F., Chandler, D.W., Franklin, F.A.,Barton, B.A., Buckman, D.W., Snetselaar, L.G., Patterson, B.H.,Schatzkin, A., Taylor, P.R., 2003. Diet and sex hormones in girls:findings from a randomized controlled clinical trial. J. Natl. CancerInst. 95, 132–141.
Ernst, E., Lauritsen, J.G., 1991. Effect of organic and inorganic mercuryon human sperm motility. Pharmacol. Toxicol. 69, 440–444.
Funston, R.N., 2004. Fat supplementation and reproduction in beeffemales. J. Anim. Sci. 82, 154–161.
Grandjean, P., White, R.F., 1999. Effects of methylmercury exposure onneurodevelopment. JAMA 281, 896–897.
Grandjean, P., Weihe, P., White, R.F., Debes, F., Araki, S., Yokoyama,K., Murata, K., Sorensen, N., Dahl, R., Jorgensen, P.J., 1997.Cognitive deficit in 7-year-old children with prenatal exposure tomethylmercury. Neurotoxicol. Teratol. 6, 417–428.
Gromadzka-Ostrowska, J., Przepiorka, M., Romanowicz, K., 2002.Influence of dietary fatty acids composition, level of dietary fat andfeeding period on some parameters of androgen metabolism in malerats. Reprod. Biol. 2, 277–293.
Grubbs, F.E., 1969. Procedures for detecting outlying observations insamples. Technometrics 11, 1–21.
Guallar, E., Sandz-Gallardo, I., Van’t Veer, P., Bode, P., Aro, A., Gomez-Aracena, J., Kark, J.D., Riemersma, R.A., Martin-Moreno, J.M.,Kok, F.J., 2002. Mercury, fish oil, and the risk of mycocardialinfarction. New. Engl. J. Med. 22, 1747–1754.
Harada, M., 1995. Minamata disease: methylmercury poisoning in Japancaused by environmental pollution. Crit. Rev. Toxicol. 25, 1–24.
Hoeben, E., Swinnen, J.V., Heyns, W., Verhoeven, G., 1999. Heregulins orNeu differentiation factors and the interactions between PeritubularMyoid cells and Sertoli cells. Endocrinology 140, 2216–2223.
Hojbjerg, S., Neilsen, J.B., Anderson, O., 1992. Effects of dietary lipids onwhole-body retention and organ distribution of organic and inorganicmercury in mice. Food Chem. Toxicol. 30, 703–708.
Homma-Takeda, S., Kugenuma, Y., Iwamuro, T., Kumagai, Y., Shimojo,N., 2001. Impairment of spermatogenesis in rats by methylmercury:involvement of stage- and cell- specific germ cell apoptosis. Toxicology169, 25–35.
Huhtaniemi, I., 1995. Molecular aspects of the ontogeny of the pituitary-gonadal axis. Reprod. Fertil. Dev. 7, 1025–1035.
Jin, X., Lok, E., Bondy, G., Caldwell, D., Mueller, R., Kapal, K.,Armstrong, C., Taylor, M., Kubow, S., Mehta, R., Chan, H.-M., 2007.Modulating effects of dietary fats on methylmercury toxicity anddistribution in rats. Toxicology 230, 22–44.
Johnsson, C., Sallsten, G., Schutz, A., Sjors, A., Barregard, L., 2004. Hairmercury levels versus freshwater fish consumption in householdmembers of Swedish angling societies. Environ. Res. 96, 257–263.
Kesterloot, H., Lesaffre, E., Joossens, J.V., 1991. Dairy fat, saturatedanimal fat, and cancer risk. Prev. Med. 20, 226–236.
Kim, S., Lee, H.J., Yang, H., Kim, H.S., Yoon, Y.D., 2004. Prepubertalexposure to 4-tert-octylphenol induces apoptosis of testicular germcells in adult rat. Arch. Androl. 50, 427–441.
Kling, L.J., Soares, J.H., 1981. The effects of vitamin E and dietary linoleicacid on mercury toxicity. Nutr. Rep. Int. 24, 39–45.
Kosatsky, T., Przybysz, R., Armstrong, B., 2000. Mercury exposure inMontrealers who eat St. Lawrence River sportfish. Environ. Res. 84,36–43.
La Fleur, S.E., Houshyar, H., Roy, M., Dallman, M.F., 2005. Choice oflard, but not total lard calories, damps ACTH responses to restraint.Endocrinology 146, 2193–2199.
Li, D., 2003. Omega-3 fatty acids and non-communicable diseases. Chin.Med. J. 116, 453–458.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–271.
Machino, A., Nakano, H., Tamaoki, B., 1969. Influence of physical andchemical treatments upon the microsomal enzymes of testes related toandrogen biosynthesis. Endocrinol. Jpn. 16, 37–46.
Maretta, M., Marettova, E., Skrobanek, P., Ledec, M., 1995. Effect ofmercury on the seminiferous epithelium of the fowl testis. Acta Vet.Hung. 43, 153–161.
Marsh, D.O., Clarkson, T.W., Cox, C., Myers, G.J., Amin-Zaki, L., Al-Tikriti, S., 1987. Fetal methylmercury poisoning. Relationship betweenconcentration in single strands of maternal hair and child effects. Arch.Neurol. 44, 1017–1022.
McLachlan, R.I., O’Donnell, L., Meachem, S.J., Stanton, P.G., deKretser, D.M., Pratis, K., Robertson, D.M., 2002. Identification ofspecific sites of hormonal regulation in spermatogenesis in rats,monkeys and man. Rec. Prog. Horm. Res. 57, 149–179.
McVey, M.J., Cooke, G.M., Curran, I.H.A., 2004a. Increased serum andtesticular androgen levels in F1 rats with lifetime exposure to soyisoflavones. Reprod. Toxicol. 18, 677–685.
McVey, M.J., Cooke, G.M., Curran, I.H.A., 2004b. Altered testicularmicrosomal steroidogenic enzyme activities in rats with lifetimeexposure to soy isoflavones. J. Steroid Biochem. Mol. Biol. 92, 435–446.
McVey, M.J., Cooke, G.M., Curran, I.H.A., Chan, H.C., Kubow, S.,Lok, E., Mehta, R., 2008. Effects of dietary fats and proteins on rattesticular steroidogenic enzymes and serum testosterone levels. FoodChem. Toxicol. 46 (1), 259–269.

M.J. McVey et al. / Food and Chemical Toxicology 46 (2008) 270–279 279
Menotti, A., Kromhout, D., Blackburn, H., Fidanza, F., Buzina, R.,Nissien, A., 1999. Food intake patterns and 25-year mortality fromcoronary heart disease: cross-cultural correlations in the sevencountries study. The seven countries study research group. Eur. J.Epidemiol. 15, 507–515.
Miller, W.L., Strauss, J.F., 1999. Molecular pathology and mechanism ofaction of the steroidogenic acute regulatory protein, STAR. J. SteroidBiochem. Mol. Biol. 69, 131–141.
Mohamed, M.K., Burbacher, T.M., Mottet, N.K., 1987. Effects of methylmercury on testicular functions in Macaca fascicularis monkeys.Pharmacol. Toxicol. 60, 29–36.
Mondal, S., Mukhopadhyay, B., Bhattacharya, S., 1997. Inorganicmercury binding to fish oocyte plasma membrane induces steroido-genesis and translatable messenger RNA synthesis. Biometals 10, 285–290.
Muckle, G., Ayotte, P., Dewailly, E., Jacobson, S.W., Jacobson, J.L.,2001a. Determinants of polychlorinated biphenyls and methylmercuryexposure in Inuit women of childbearing age. Environ. HealthPerspect. 109, 957–963.
Muckle, G., Ayotte, P., Dewailly, E., Jacobson, S.W., Jacobson, J.L.,2001b. Prenatal exposure of the Northern Quebec Inuit infants toenvironmental contaminants. Environ. Health Perspect. 109, 1291–1299.
Myers, G.J., Davidson, P.W., Cox, C., Shamlaye, C.F., Palumbo, D.,Cernichiari, E., Sloane-Reeves, J., Wilding, G.E., Kost, J., Huang,L.S., Clarkson, T.W., 2003. Prenatal methylmercury exposure fromocean fish consumption in the Seychelles child development study.Lancet 361, 1686–1692.
National Research Council (NRC), 1995. Nutrient Requirements ofLaboratory Rats. Nutrient Requirements of Laboratory Animals,fourth ed. (revised). National Academy of Sciences, Washington, DC,pp. 11–79.
National Research Council, 2000. Toxicological Effects of Methylmer-cury. National Academy Press, Washington, DC.
Ng, T.B., Liu, W.K., 1990. Toxic effect of heavy metals on cells isolatedfrom the rat adrenal and testis. In Vitro Cell. Dev. Biol. 1, 24–28.
Parizek, J., Ostadalova, I., 1967. The protective effect of small amounts ofselenite in sublimate intoxication. Experientia 23, 142–143.
Parker, K.L., Schedl, A., Schimmer, B.P., 1999. Gene interactions ingonadal development. Annu. Rev. Physiol. 61, 417–433.
Payne, A.H., Youngblood, G., 1995. Regulation of expression ofsteroidogenic enzymes in Leydig cells. Biol. Reprod. 52, 217–225.
Ramalingam, V., Vimaladevi, V., 2002. Effect of mercuric chloridemembrane-bound enzymes in rat testis. Asian J. Androl. 4, 309–311.
Ramalingam, V., Panneerdoss, S., Girija, M., Ilango, S., 2001. Mercuricchloride induced changes in the histology of the testis and serumtestosterone in adult albino rats. Pollut. Res. 21, 7–11.
Ramalingam, V., Vimaladevi, V., Rajeswary, S., Suryavathi, V., 2003.Effect of mercuric chloride on circulating hormones in adult albinorats. J. Environ. Biol. 4, 401–404.
Reddy, B.S., 1987. Dietary fat and colon cancer: animal models. Prev.Med. 16, 460–467.
Rosenfeld, C.S., Grimm, K.M., Livingston, K.A., Brokman, A.M.,Lamberson, W.E., Roberts, R.M., 2003. Striking variation in the sexratio of pups born to mice according to whether maternal diet is highin fat or carbohydrate. Proc. Natl. Acad. Sci. USA 100, 4628–4632.
Rowland, I.R., Mallett, A.K., Flynn, J., Hargreaves, R.J., 1986. The effectof various dietary fibres on tissue concentration and chemical form ofmercury after methylmercury exposure in mice. Arch. Toxicol. 59, 94–98.
Rowlands, J.C., Hakkak, R., Ronis, M.J.J., Badger, T.M., 2002. Alteredmammary gland differentiation and progesterone receptor expressionin rats fed soy and whey proteins. Toxicol. Sci. 70, 40–45.
Sakamoto, M., Kakita, A., de Oliveira, R.B., Sheng Pan, H., Takahashi,H., 2004. Dose-dependent effects of methylmercury administeredduring neonatal brain spurt in rats. Brain Res. Dev. Brain Res. 152,171–176.
Simophoulos, A.P., 2002. Omega-3 fatty acids in inflammation autoim-mune diseases. J. Amer. Coll. Nut. 21, 495–505.
Sing, K.A., Hryhorczuk, D., Saffirio, G., Sinks, T., Paschal, D.C.,Sorensen, J., Chen, E.H., 2003. Organic mercury levels among theYanomama of the Brazilian Amazon Basin. Ambio 32, 434–439.
Smith, J.C., Farris, F.F., 1996. Methyl mercury pharmacokinetics in man:a reevaluation. Toxicol. Appl. Pharmacol. 137, 245–252.
Sprando, R.L., Zirkin, B.R., 1997. Is Leydig cell steroidogenic functionaffected by the germ cell content of the seminiferous tubules? J. Androl.18, 424–430.
Staples, C.R., Burke, J.M., Thatcher, W.W., 1998. Influence of supple-mental fats on reproductive tissues and performance of lactating cows.J. Dairy Sci. 81, 656–871.
Stern, A.H., 2005. A revised probabilistic estimate of the maternal methylmercury intake dose corresponding to a measured cord blood mercuryconcentration. Environ. Health Perspect. 113, 155–163.
Taylor, P.D., Khan, I.Y., Lakasing, L., Dekou, V., O’Brien-Coker, I.,Mallet, A.I., Hanson, M.A., Poston, L., 2003. Uterine artery functionin pregnant rats fed a diet supplemented with animal lard. Exp.Physiol. 88, 389–398.
Turner, T.T., Jones, C.E., Howards, S.S., Ewing, L.L., Zegeye, B.,Gunsalus, G.L., 1984. On the androgen microenvironment of maturingspermatozoa. Endocrinology 115, 1925–1932.
Vachhrajani, K.D., Chowdhury, A.R., 1990. Distribution of mercury andevaluation of testicular steroidogenesis in mercuric chloride andmethylmercury administered rats. Indian J. Exp. Biol. 28, 746–751.
Veltman, J.C., Maines, M.D., 1986. Alterations of heme, cytochrome P-450, and steroid metabolism by mercury in rat adrenal. Arch. BiochemBiophys. 248, 467–478.
Wagemann, R., Trebacz, E., Boila, G., Lockhart, W.L., 2000. Mercuryspecies in the liver of ringed seals. Sci Total Environ. 261, 21–32.
Xiao, R., Badger, T.M., Simmen, F.A., 2005. Dietary exposure to soy orwhey proteins alters colonic global gene expression profiles during ratcolon tumorigenesis. Mol. Cancer 4 (e-pub ahead of print).
Zirkin, B.R., Chen, H., Luo, L., 1997. Leydig cell steroidogenesis in agingrats. Exp. Gerontol. 32, 529–537.





























