Embed Size (px)
Citation preview

Journal of the Neurological Sciences, 121 (1994) 79-89 79 © 1994 Elsevier Science Publishers B.V. All rights reserved 0022-510X/94/$07.00
JNS 04183
A new gerbil model of hindbrain ischemia by extracranial occlusion of the bilateral vertebral arteries
R y u j i H a t a a,., M a s a y a s u M a t s u m o t o b,c, K a z u o K i t a g a w a b, T o m o h i r o M a t s u y a m a b,
T o s h i h o O h t s u k i b, M a s a f u m i T a g a y a b, N o b u o H a n d a b, M i c h i o N i i n o b e d,
K a t s u h i k o M i k o s h i b a d, T s u n e h i k o N i s h i m u r a a, T a k e h i k o Y a n a g i h a r a c
a n d T a k e n o b u K a m a d a b
a Division of Tracer Kinetics, Biomedical Research Center b First Department of Internal Medicine, ¢ Department of Neurology, School of Medicine, and a Division of Regulation of Macromolecular Function, Institute for Protein Research, Osaka University, 2-2 Yamadaoka, Suita, Osaka 565, Japan
(Received 3 February, 1993) (Revised, received 19 July, 1993)
(Accepted 28 July, 1993)
Key words: Hindbrain ischemia; Gerbil; Cerebral ischemic response; Immunohistochemistry; Microtubule-associated protein 2
Summary
A new gerbil model of hindbrain ischemia was induced by extracranial occlusion of the bilateral vertebral arteries just before their entry into the transverse foramen of the cervical vertebra. Carbon black studies, performed at 5 min after occlusion, revealed that the pons-medulla oblongata, and the cerebellum were quite ischemic in all animals. Cardiovascular changes in mean arterial blood pressure (MABP) and heart rate were recorded until 30 min after occlusion, and revealed that the typical cerebral ischemic response (i.e., abrupt increase in MABP, bradycardia, and apnea) was elicited in all animals (n = 10). Thirty minutes after occlusion, animals (n = 4) were decapitated and immersion-fixed. Brain sections were stained with hematoxylin-eosin (HE) and also immunostained for microtubule-associated protein 2 in order to evaluate ischemic neuronal damage from 30 min of ischemia. By HE staining, ischemic lesions were detected bilaterally in the oculomotor, the trigeminal motor, the lateral vestibular, and the cerebeilar interpositus nucleus. In addition, immunostaining revealed ischemic lesions in several other hindbrain areas. In conclusion, we could successfully establish a new gerbil model of hindbrain ischemia. Carbon black perfusion and hemodynamic studies revealed that severe and reproducible hindbrain ischemia was produced. By histopathological examination, we could also clearly demonstrate symmetrical ischemic lesions in several hindbrain areas.
Introduction
To investigate the pathophysiology of cerebral is- chemia, many forebrain ischemia models have been developed in rodents (Levine et al. 1966; Pulsinelli et al. 1979; Smith et al. 1984; Kameyama et al. 1985). However, owing to technical difficulties, only a few models of hindbrain ischemia have been developed so far. A few examples include a gerbil model involving occlusion of the vertebro-basilar junction (Yamada et al. 1984), a rat model using basilar artery ligation (Wojak et al. 1991), and a SHR model employing cauterization of the bilateral vertebral arteries and also by imposing systemic hypotension (Yao et al. 1990). However, surgical approaches using basilar artery liga-
* Corresponding author. Osaka 565, Japan. Tel +816-875-7384 Fax + 816-875-7387
tion disturb the cerebro-spinal fluid and result in injury to the intracranial contents, while systemic hypotension can cause generalized metabolic disruption of several major organs including the brain. We therefore ex- plored the feasibility of extracranial occlusion of the vertebral arteries in the gerbil using less invasive surgi- cal procedures. We occluded the vertebral arteries just before their entry into the transverse foramen of the cervical vertebrae and examined the physiological and histological consequences of bilateral vertebral occlu- sion (BVO). A part of the present work has been reported in abstract form (Hata et al. 1991).
Materials and methods
General surgical procedures
Adult Mongolian gerbils of both sexes, weighing 60-80 g, which had been permitted free access to food
SSDI 0022-510X(93)E0193-D

80
and water until the day of the experiment, were lightly anesthetized with ketamine hydrochloride (50 m g / k g i.p.) and placed in the supine position on a table tilted at approximately 30 ° to the horizontal. An anterior midline cervical incision was made, and the operation was performed using a surgical microscope. After dis- secting the musculus longus colli, both vertebral arter- ies were exposed at the point just before entry into the transverse foramen of the cervical vertebra and loosely looped with silk sutures, without interrupting the verte- bral blood flow (Fig. 1). Afterwards, the incision was closed and, while still under anesthesia, a polyethylene tube was inserted into the femoral artery for recording mean arterial blood pressure (MABP). Animals were then allowed to recover from anesthesia for about 8 h until they became indistinguishable from normal ani- mals. During this recovery period, gerbils were fasted but were allowed free access to water. Again under light ether anesthesia, the neck incision was reopened, and the trachea was intubated. The sutures around each vertebral artery were pulled by 5g weights to occlude the circulation (Fig. 2). Cardiovascular changes in MABP and H R were recorded for 30 min following occlusion (n = 10), and animals were afterwards decap-
Fig. 1. The view of surgical field in the gerbil looped with silk sutures around the right vertebral arteries (white arrows) just before entering
the transverse foramen of the cervical vertebra ( × 10).
Fig. 2. Diagram of procedure for bilateral vertebral occlusion in gerbils.
itated for histological procedures (n = 4). In another group of animals (n = 10), 1.5 ml /100 g of carbon black (Bokusai, Fueki, Tokyo) was hand-injected tran- scardially (Matsuyama et al. 1983) at a rate of approxi- mately 0.8 m l / m i n 5 min after BVO. After allowing the carbon black to circulate for a few minutes, the animals were decapitated. The brains were then re- moved and placed into a 10% formalin solution prior to macroscopic observation. During ischemia, body temperature was monitored and maintained at 37°C by a heating lamp and a heating pad connected to a rectal thermistor (ATB-1100, Nihon Kohden, Tokyo). Artifi- cial ventilation with room air was initiated immediately after apnea was elicited during ischemia. In a separate study, we also checked the survival rate after transient hindbrain ischemia for 5, 10, 15, 30 rain (n = 10, in each group).
Histological procedures Animals were anesthetized with ether and their
brains were removed immediately after decapitation. The infratentorial structures were then separated, di- vided into coronal sections, and immersed in ethanol containing 5% acetic acid at 4°C. After fixation for 4 -5 h, each tissue block was dehydrated, embedded in paraffin, and sliced into 5-/.~m sections. Serial brain sections were immunostained every 200 p~m for micro- tubule-associated protein 2 (MAP2) (Huber et al. 1984).
The peroxidase-antiperoxidase method for light mi- croscopy was used for the immunohistochemical proce- dure, as previously described (Kitagawa et al. 1989). Briefly, each deparaffinized tissue section (5 t~m) was

incubated with 10% non-immunized goat serum, with anti-MAP2 rabbit serum (1:200) for 1 h, with the secondary antiserum (1:20) for 30 min, and with the peroxidase-antiperoxidase complex (1 : 100) for 30 min. The peroxidase reaction was accomplished by incuba- tion in the presence of 0.05% 3,3'-diaminobenzidine tetrahydrochloride and 0.01% H20 2. Control sections were incubated with absorbed primary antiserum. Cell nuclei were visualized by counterstaining with Harris' hematoxylin. For comparison, adjacent sections were also stained with hematoxylin-eosin (HE).
Characterization of the primary antiserum for MAP2 was reported elsewhere (Niinobe et al. 1988).
Results
Carbon black perfusion Figs. 3 and 4 show representative photographs of
the brain surfaces and their serial coronal sections 5 min after BVO. As summarized in Fig. 5, all 10 ani- mals showed dark staining in the telencephalon and no staining in the pons-medulla oblongata, or the cerebel-
81
lum. Three gerbils showed no staining in ventro-medial parts of the diencephalon, and 7 gerbils showed no staining in the upper midbrain. No gerbil showed stain- ing in the ventro-medial tegmentum of the lower mid- brain.
Neurological and respiratory status In all animals, apnea was observed between 30 sec
and 1.0 min after occlusion, and consequent convul- sions were observed in all four limbs for about 60 sec. No other convulsions were observed during ischemia. Immediately following respiratory arrest, artificial res- piration was initiated to prevent systemic anoxia. After convulsions had ceased, all gerbils became unrespon- sive and lost their corneal reflex (Table 1, Fig. 6). All animals could survive during hindbrain ischemia with mechanical ventilation. After 5 min hindbrain is- chemia, spontaneous breathing reappeared within 5 min, and mean arterial blood pressure and pulse rate returned to pre-ischemic level within 10 min, and all gerbils could survive more than one week. After 10-30 min hindbrain ischemia, spontaneous breathing reap- peared. But all these animals died within 4 h because
Fig. 3. Dorsal (A) and ventral (B) view of a gerbil brain perfused with carbon black during hindbrain ischemia. Note lack of staining of the cerebellar surface.
82
E5~3 0 [-=~ < 25 %
~ < 5 0 % ~< 75%
< i 0 0 %
I lOO %
Fig. 5. Schematic distribution of carbon black staining in 10 gerbil brains during progressive hindbrain ischemia. White (0), dotted ( < 25), hatched ( < 50), meshed ( < 70), cross-hatched ( < 100) and black (100). Numbers in the parentheses indicate percentages of carbon
black staining based on 10 gerbils.
o f r e s p i r a t o r y f a i l u r e , if m e c h a n i c a l v e n t i l a t i o n w a s
c e a s e d .
Cardiovascular status B V O p r o d u c e d t h e w e l l - k n o w n c a r d i o v a s c u l a r
c h a n g e s w h i c h a r e g e n e r a l l y a s c r i b e d to t h e c e r e b r a l
Fig. 4. Coronal sections of the gerbil brain after bilateral vertebral occlusion and transcardial perfusion with carbon black. Note the marked pallor in the cerebellum and the pons-medulla oblongata.
TABLE 1
PHYSIOLOGICAL VARIABLES IN CONTROL ANIMALS A N D IN ANIMALS SUBJECTED TO 30 MIN OF HINDBRAIN IS- CHEMIA
Experimental PaO 2 PaCO z pH RT MABP group (mm Hg) (mm Hg) (°C) (mm Hg)
Control (n = 8) 86.3 ± 1.7 35.9 ± 1.7 7.32 ± 0.01 37.1 ± 0.2 84 ± 3 30 rain after
BVO ( n = 8 ) 89.4±4.8 35.0±2.3 7.32±0.02 37.0±0.2 6 0 ± 8 *
Since removal of a large volume of blood may affect systemic and cerebral circulation in these small animals, the arterial blood pH, PaO 2, and PaCO 2 before ischemia were measured in one group, and after 30 min of hindbrain ischemia in another group. MABP, mean arterial blood pressure; RT, rectal temperature; All values are means ± SEM, n = number of animals. * indicates significantly (P <0.01) lower than control by Mann-
Whitney U-test.
4 0 0
3 5 0
3 O 0
¢
\ 2 5 0
c~
P 200 <
=
[.,
~: 1 5 0 <
I O 0
,d, %++... O:BP
A l l v l l o l e i are mean ± S . [ . M
A P N E A
I I I I I I I I I I - 7 0 I 2 3 4 5 1 0 1,5 2 0 2 5 3 0
a:
- - 1 4 0 E v
u2 a:
1 2 0 .~
1 O 0 m
0 8 0 O
6 0 ~ <
¢
4 0 ~.
I =
2 O Z <
IE
0
T I M E ( m i n - )
Fig. 6. Changes in mean arterial blood pressure (BP), heart rate (HR), and respiration in gerbils with an occlusion of the bilateral vertebral artery (BVO). Respiration ceased in all animals at between
30 sec - 1.0 min after occlusion. For details, see text,

ischemic response (CIR) (an increase in systemic arte- rial pressure, bradycardia, and apnea) (Guyton et al. 1986). In response to BVO, MABP increased to 180% of the control value (84 5- 3 mmHg) 50 sec after occlu- sion. MABP returned to the pre-ischemic level at 3.5 min and gradually decreased to 70% of the control value by 30 rain after the occlusion. HR decreased by 4% from the control value (371 5- 12 beats/min) 20 sec after occlusion, but then increased to the maximum level (386 5- 12 beat/min) 1.5 min after occlusion, be- fore gradually decreasing again to 79% of the control value 30 min after occlusion (Fig. 6).
Histochemical observation The distribution of MAP2 in various regions of the
normal gerbil brain has been reported elsewhere (Kitagawa et al. 1989). Representative micro-photo- graphs of ischemic lesions with reaction for MAP2 and HE are shown in Figs. 7-11. The frequency and distri- bution of lesions demonstrated with reaction for MAP2
83
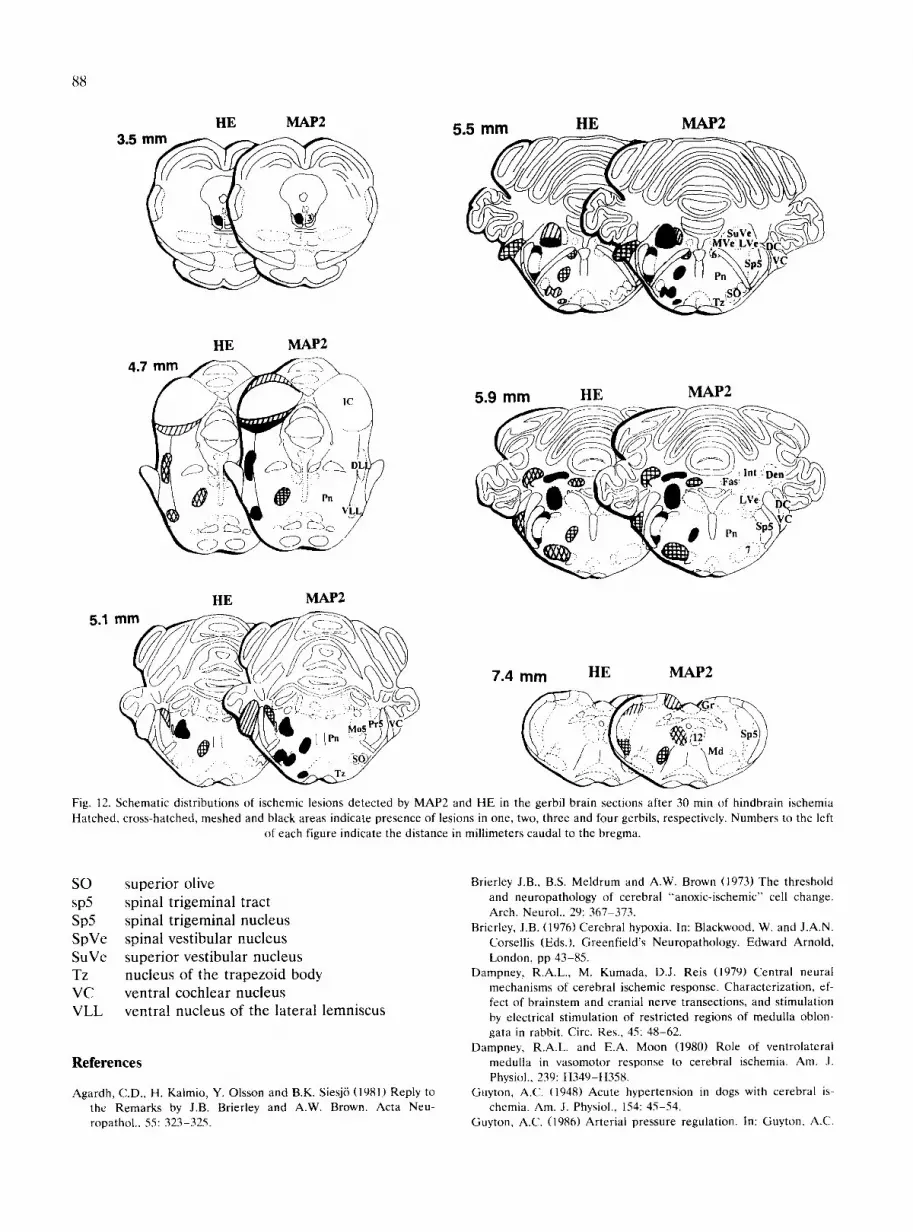
and HE after 30 rain of hindbrain ischemia are shown in Fig. 12. Each brain section was examined by two investigators; whenever there was any uncertainty, a third investigator examined the specimen without any prior information. The lesions were mapped by a single investigator.
By HE staining, "hydropic cell changes" (Brierley et al. 1973) like pattern (Jenkins et al. 1979; Agardh et al. 1981; Kalmio et al. 1981) characterized by pale staining and moderate swelling of the neuronal cytoplasm, were seen in the oculomotor, the motor trigeminal nucleus, the lateral vestibular, and the cerebellar interpositus nucleus in all 4 gerbils; in the abducens nucleus, and the ventral cochlear nucleus in 3 animals; in the nu- cleus of the lateral lemniscus, the facial nucleus, the cerebellar fastigial nucleus, and the cerebellar dentate nucleus in 2 animals; in the superior vestibular nucleus, the superior olive, the nucleus of the trapezoid body, and the spinal vestibular nucleus in one animal. Small and scattered lesions were seen in the dorsal and
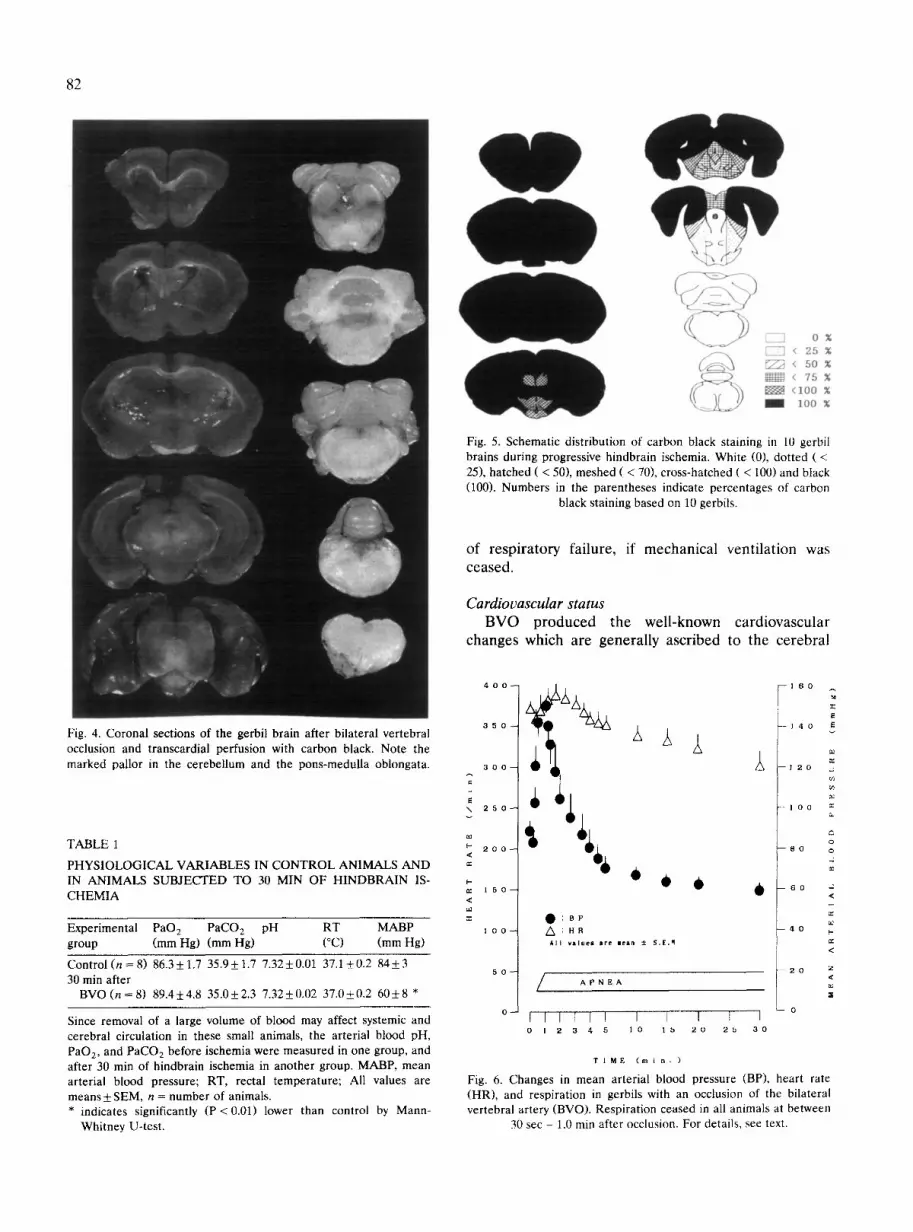
Fig. 7. Representative micro-photographs of the midbrain section (corresponding to the stereotaxic section (Loskota et al. 1974), -3.5 mm rostral to the bregma) stained with MAP2 antiserum in sham-operated control (A) and after 30 min of hindbrain ischemia (C). (B,D) Schematic drawings of microphotographs in A and C, respectively. Cross-hatched areas indicate ischemic lesions (D). For details, see text. Scale
bars = 1 mm.
84
ventral margin of the nucleus of the spinal trigeminal tract in all four gerbils; in the central parts of the pontine reticular nucleus in 3 animals; in the ventral margin of the nucleus of the inferior colliculus, and the dorsomedial part of the principal sensory trigeminal nucleus in one animal. These ischemic lesions were detected bilaterally and could not be found in the sham-operated control group.
After immunostaining for MAP2, loss of reactivity for MAP2 in the neuropil, nerve cell body, and den- drites was used as criteria for the presence of ischemic lesions. The findings were compared with sham-oper- ated animals. Ischemic lesions were detected in the oculomotor, the nucleus of the lateral lemniscus, the motor trigeminal, the superior olive, the nucleus of the trapezoid body, the superior vestibular nucleus, the lateral vestibular nucleus, the spinal vestibular nucleus, and the cerebellar interpositus nucleus in all four ger- bils; in the ventral cochlear nucleus, the abducens nucleus, the facial nucleus, the cerebellar fastigial nu- cleus, and the cerebellar dentate nucleus in three ani- mals; in the hypoglossal nucleus in two animals; in the medial vestibular nucleus, and the gracile nucleus in
one animal. Small and scattered lesions were seen in the ventral margin of the nucleus of the inferior col- liculus, the dorsal and ventral margin of the nucleus of the spinal trigeminal tract, and the central parts of the pontine reticular nucleus in all four gerbils; in the dorsomedial part of the principal sensory trigeminal nucleus, and the central parts of the medullary reticu- lar nucleus in three animals; in the dorsal margin of the nucleus of the inferior colliculus in one animal.
Discussion
The Mongolian gerbil (Meriones unguiculatus) is un- usually susceptible to cerebral infarction following oc- clusion of one or both common carotid arteries (Levine et al. 1966). This is attributed to the lack of the posterior communicating artery (Levine et al. 1969). In the present study, we applied this anatomical advan- tage to produce hindbrain ischemia. There were some possibilities that blood flow to the brainstem and cere- bellum, after bilateral vertebral occlusion, was supplied
s5 \ J MoSj
!1
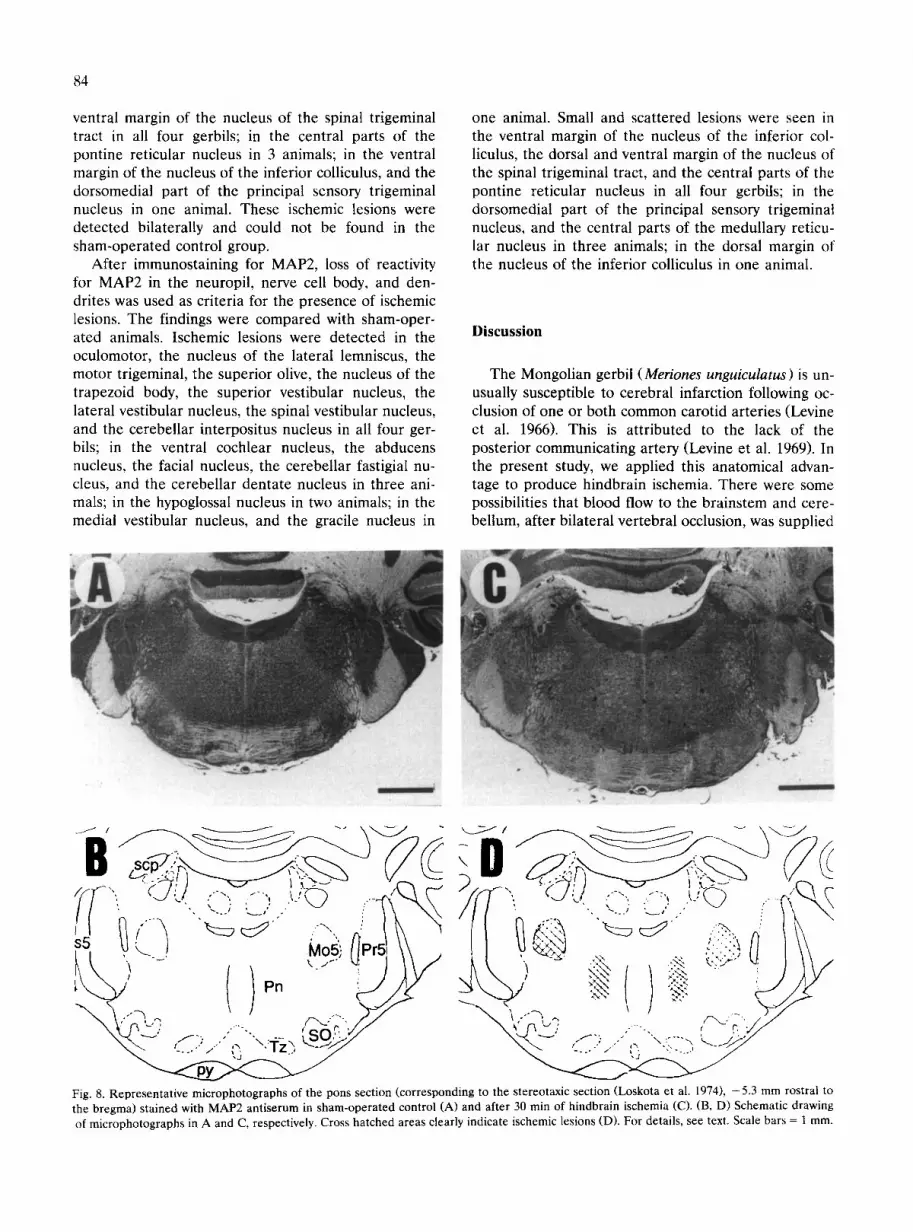
Fig. 8. Representative microphotographs of the pons section (corresponding to the stereotaxic section (Loskota et al. 1974), -5.3 mm rostral to the bregma) stained with MAP2 antiserum in sham-operated control (A) and after 30 min of hindbrain ischemia (C). (B, D) Schematic drawing of microphotographs in A and C, respectively. Cross hatched areas clearly indicate ischemic lesions (D). For details, see text. Scale bars = 1 mm.
by the anterior spinal artery (Pulsinelli et al. 1979) or small vessels linking the basilar artery with the poste- rior cerebral arteries (Levy et al. 1974). To determine whether these communicating vessels function in vivo, we performed an intravascular injection of carbon black on the anesthetized animals. Since carbon black could circulate for a few minutes without a drop in blood pressure, this experimental manipulation would not cause a decrease in cerebral blood flow. Regions of the brain that were stained with intravenous carbon black could not be completely ischemic in vivo. Our results clearly demonstrate that severe and reproducible hind- brain ischemia was produced in the gerbil by extracra- nial occlusion of the bilateral vertebral arteries.
Following BVO all animals spontaneously ceased to breath and lost their corneal reflex during ischemia, suggesting that the lower brain stem was adversely affected. Furthermore, the typical CIR was elicited just after BVO and according to Guyton, the CIR was observed in complete cerebral ischemia (Guyton 1948). Many investigators have reported that the center re- sponsible for the CIR is located in the brain stem,
85
especially in the medulla (Dampney et al. 1979, 1980; Hasel ton et al. 1985). Guyton also reported that the CIR often became exhausted, sometimes within only a few minutes when ischemia in the medulla became very intense (Guyton 1948). This resulted from an almost complete lack of metabolic nutrients reaching the ac- tive neuronal cells of the medulla. We observed the typical CIR until its exhaustion in all gerbils undergo- ing BVO. These findings also suggest that severe hind- brain ischemia is produced after BVO.
The detection of morphologic damage in early cere- bral ischemia proved to be difficult using only conven- tional light microscopic examination. Therefore, we also immunostained the brain sections for MAP2 (Ni- inobe et al. 1988). MAP2 is abundant in the brain and is present in large amounts in the dendrites, where this protein is associated with dendritic microtubules and postsynaptic dendrites, both in shaft and spine synapses (Huber et al. 1984). We have previously demonstrated that immunoreactivity to MAP2 clearly visualized the dendrites, the neuronal perikarya, and the neuropil in various regions of the gerbil brain, although axon fibers
B i , " ~ ' ~ . . . . . . ~ ~ ' - ~ - - ' " " " . . . . "" / ( f . - " :. . . . . t ; . = - - - : . In t , . D e n - ~ "
,,.. ~. . - ~ " [ " , ~ q - a s ; . . . . . . :, . -
)"-' i~'i';[ "-" >:';~~lJ" "-~"SuVe') "~ Px(~ ; , c p ~. ~-- . . . . . . .___. ,
- / f / ) --. _,o,-.:~.':>,,--,t . .--",-- '~ \~. / " ' - g 7 . J ~ ~ . . c ~ - ... ~ ~1"-.
D / - 1 . . . . . ~ . ~ ,-'z'.-?;,.- ~, ,, . . . . , . ,',,-,TrO.'.X*. i I .... 5X~,,/,,, . . . . L...'~zc:;z~'~;:-:~'----__/~C.~ ~,,~...'
.....
/V-~-,~<S~'-~<.'-!, <-,,~ ,, f / ;~~L~ ' ,A. : ' , - ; r -xk~x? ' , - r--'~ \'~.. ' / 2 Z--~..--~-~,--,"~-~ \ \ "~/-. i (.14::..i~4..-) \; I/i
'-U \LJtY \ h . - , . , .... / X . . . . .
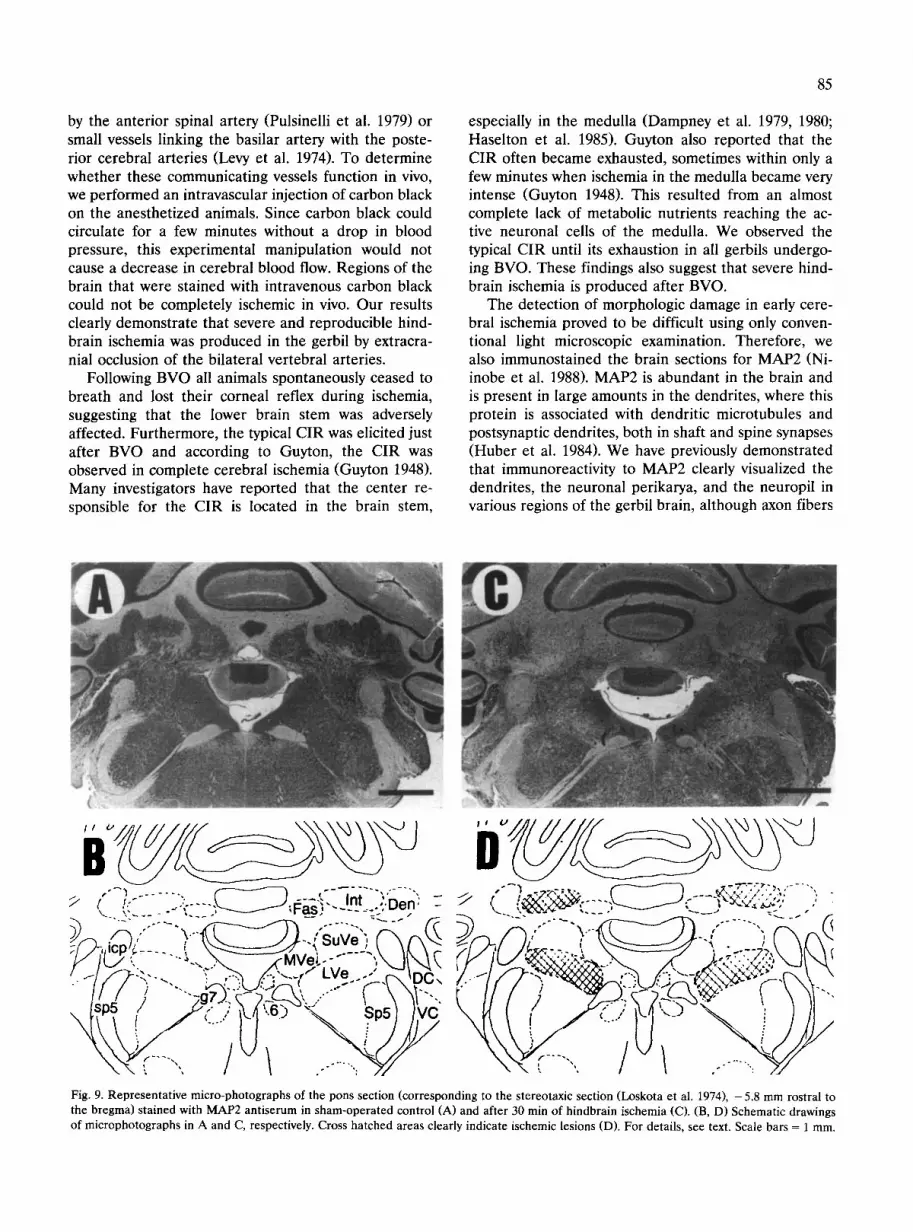
Fig. 9. Representative micro-photographs of the pons section (corresponding to the stereotaxic section (Loskota et al. 1974), - 5.8 mm rostral to the bregma) stained with MAP2 antiserum in sham-operated control (A) and after 30 min of hindbrain ischemia (C). (B, D) Schematic drawings of microphotographs in A and C, respectively. Cross hatched areas clearly indicate ischemic lesions (D). For details, see text. Scale bars = 1 mm.
86
and glial cells exhibited no staining (Kitagawa et al. 1989), MAP2 immunoreactivity was diminished in brain regions which were vulnerable to ischemia, suggesting that MAP2 was a sensitive marker protein for early detection of ischemic damage (Kitagawa et al. 1989). By using this immunohistochemical method, we demonstrated that some neurons in the brain stem lost staining to MAP2, whereas other neurons retained staining. This selectivity of staining may reflect differ- ences in the vulnerability of neuronal regions to is- chemia. It was an unexpected finding that the perikarya of the Purkinje cells, which have been reported to be vulnerable to ischemia (Brierley 1977), retained reactiv- ity to the antiserum during the entire 30 min of hind- brain ischemia. However, our results are in good agree- ment with a previous report by Yamada et al. (1985), who also observed selective vulnerability of brainstem neurons. Employing an immunohistochemical method with immunoreactivity to tubulin, large neurons in the vestibular nucleus were the most vulnerable to hind- brain ischemia, whereas the neuronal perikarya of the Purkinje ceils retained staining for up to 6 h of hind-
brain ischemia. Although the mechanisms underlying regional heterogeneity of ischemic damage are unclear, differences in cellular vulnerability to ischemia (Johan- sen et al. 1987), regional heterogeneity in focal cere- bral blood flow (Matsumoto et al. 1987) and energy metabolism (Mrsulja et al. 1986) are possible candi- dates. Therefore, investigation into the progression of neuronal damage, with respect to, quantitative mea- surements in local cerebral blood flow and energy metabolism are needed for further study.
In conclusion, we have successfully established a new hindbrain ischemia model in gerbils by extracra- nial occlusion of the bilateral vertebral arteries. Our model has the following advantages: (1) it avoids injury to the intracranial contents, (2) it produces severe and reproducible hindbrain ischemia, and (3) it allows reperfusion.
By immunostaining for MAP2, we have also clearly demonstrated several hindbrain areas that are vulnera- ble to cerebral ischemia. This model may be useful for the pathophysiological investigation of cerebral is- chemia.
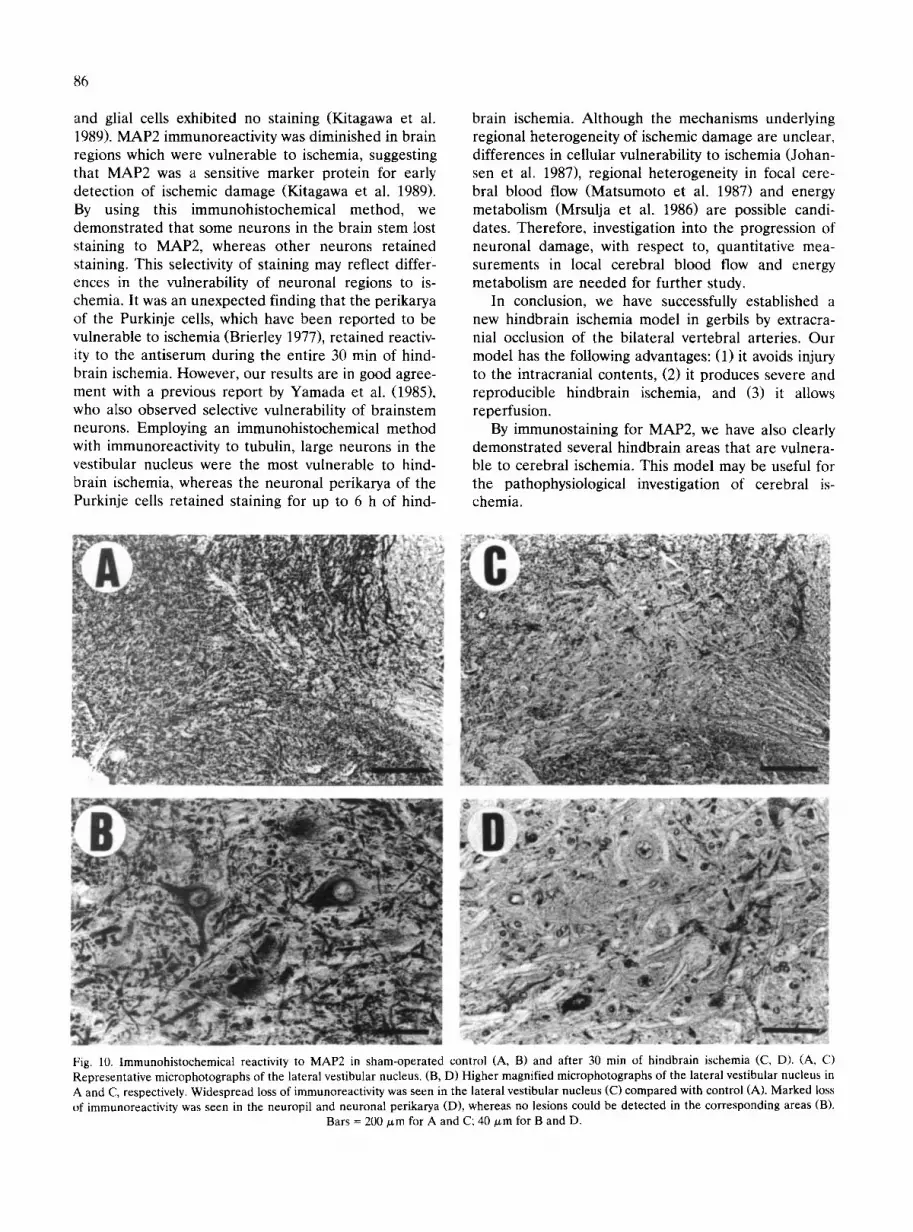
Fig. 10. Immunohistochemical reactivity to MAP2 in sham-operated control (A, B) and after 30 min of hindbrain ischemia (C, D). (A, C) Representative microphotographs of the lateral vestibular nucleus. (B, D) Higher magnified microphotographs of the lateral vestibular nucleus in A and C, respectively. Widespread loss of immunoreactivity was seen in the lateral vestibular nucleus (C) compared with control (A). Marked loss of immunoreactivity was seen in the neuropil and neuronal perikarya (D), whereas no lesions could be detected in the corresponding areas (B).
Bars = 200 ~,m for A and C; 40/zm for B and D.

87
Fig. 11. Coronal brain sections stained with HE in sham-operated control (A, B) and after 30 min of hindbrain ischemia (C, D). (A, C) Representative microphotographs of the lateral vestibular nucleus. (B, D) Higher magnified microphotographs of the lateral vestibular nucleus in A and C, respectively. Ischemic lesions could detect in the lateral vestibular nucleus (B) compared with sham-operated control (D). For details,
see text. Scale bars = 250 p~m for A and C; 50 p~m for B and D.
Acknowledgments We wish to thank Mr. K. Wakitani and H. Aishita (Research Institute, Ono Pharmaceutical, Osaka) for careful man- agement of animals, Ms S. Goi, Mr. A. Naito and H. Yoshimura for their technical assistance and Miss M. Sakai and K. Moriguchi for their secretarial assistance. Supported by a research grant for cardio- vascular disease (2A-2) from the Ministry of Health and Welfare and by a Grant-in-aid (#03670450) from the Ministry of Education, Science and Culture in Japan.
Abbreviations
MABP mean arterial blood pressure BVO MAP2 HE 3 6 7 12 DC
bilateral vertebral occlusion microtubule-associated protein 2 hematoxylin-eosin oculomotor nucleus abducens nucleus facial nucleus hypoglossal nucleus dorsal cochlear nucleus
D e n
DLL Fas g7 Gr icp IC Int LVe Md MG ml Mo5 MVe Pn Pr5 PY s5 scp
SN
cerebellar dentate nucleus dorsal nucleus of the lateral lemniscus cerebellar fastigial nucleus genu facial nerve gracile nucleus inferior cerebeUar peduncle inferior colliculus cerebellar interpositus nucleus lateral vestibular nucleus medullary reticular nucleus medial geniculate nucleus medial lemniscus motor trigeminal nucleus medial vestibular nucleus pontine reticular nucleus principal sensory trigeminal nucleus pyramidal tract sensory root trigeminal nerve superior cerebellar peduncle substantia nigra
5.5 mm HE MAP2 HE MAP2 3.5
88
HE MAP2
HE MAP2
5.1 mm 7.4 turn HE MAP2
l~ ~ - - - ~ . a - I / ' ~ i ,' ,~---,-:-:~--~ ~ ~ \ . ," :2 . . . . . . o . . _ % & . . ~ ', , ~ . ~ \ ~ ~ ( ~ ' , " " '~ ~'12" 'Sp5) )
Fig. 12. Schematic distributions of ischemic lesions detected by MAP2 and HE in the gerbil brain sections after 30 rain of hindbrain ischemia Hatched, cross-hatched, meshed and black areas indicate presence of lesions in one, two, three and four gerbils, respectively. Numbers to the left
of each figure indicate the distance in millimeters caudal to the bregma.
SO sp5 Sp5 SpVe SuVe Tz VC VLL
superior olive spinal trigeminal tract spinal trigeminal nucleus spinal vestibular nucleus superior vestibular nucleus nucleus of the trapezoid body ventral cochlear nucleus ventral nucleus of the lateral lemniscus
References
Agardh, C.D., H. Kalmio, Y. Olsson and B.K. Siesj6 (1981) Reply to the Remarks by J.B. Brierley and A.W. Brown. Acta Neu- ropathol.. 55: 323-325.
Brierley J.B., B.S. Meldrum and A.W. Brown (1973) The threshold and neuropathology of cerebral "anoxic-ischemic" cell change. Arch. Neurol., 29: 367-373.
Brierley, J.B. (1976) Cerebral hypoxia. In: Blackwood, W. and J.A.N. Corsellis (Eds.), Greenfield's Neuropathology, Edward Arnold, London, pp 43-85.
Dampney, R.A.L., M. Kumada, D.J. Reis (1979) Central neural mechanisms of cerebral ischemic response. Characterization, ef- fect of brainstem and cranial nerve transections, and stimulation by electrical stimulation of restricted regions of medulla oblon- gata in rabbit. Circ. Res., 45: 48-62.
Dampney, R.A.L. and E.A. Moon (1980) Role of ventrolateral medulla in vasomotor response to cerebral ischemia. Am. J. Physiol., 239: H349-H358.
Guyton, A.C. (1948) Acute hypertension in dogs with cerebral is- chemia. Am. J. Physiol., 154: 45-54.
Guyton, A.C. (1986) Arterial pressure regulation, ln: Guyton, A.C.

(Ed.), Text Book of Medical Physiology, W.B. Saunders, Philadel- phia, PA, pp 244-256.
Haselton, J.R., C. Haselton, P. Vera, H.H. Ellenberger, W.G. Leblanc, N. Schneiderman and P.M. Mccabe (1985) Nucleus reticularis lateralis involvement in the pressor component of the cerebral ischemic response, Brain Res., 335: 315-320.
Hata, R., M. Matsumoto, K. Kitagawa, T. Matsuyama, N. Niinobe, K. Kuwabara, T. Ohtsuki, M. Tagaya, N. Handa, K. Kimura, K. Mikoshiba and T. Kamada (1991) A new gerbil model of hind- brain ischemia produced by bilateral vertebral occlusion and immunohistochemical investigation for differential vulnerability. J. Cereb. Blood Flow Metab., 11 (Suppl. 2): $535.
Huber, G. and A. Matus (1984) Differences in the cellular distribu- tions of two microtubule-associated proteins, MAP1 and MAP2, in rat brain. J. Neurosci. 4: 151-160.
Jenkins, L.W., J.T. Povlishock, D.P. Becker, J.D. Miller and H.G. Sullivan (1979) Complete cerebral ischemia - an ultrastructual study. Acta Nenropathol., 48: 113-125.
Johansen, F.F., J. Zimmer and N.H. Diemer (1987) Early loss of somatostatin neurons in dentate hilus after cerebral ischemia in the rat precedes CA-1 pyramidal cell loss. Acta Neuropathol., 73: 110-114.
Kalmio, H., S. Rehncrona, B. S6derfeldt, Y. Olsson and B.K. Siesj6 (1981) Brain lactic acidosis and ischemic cell damage. 2. Histopathology. J. Cereb. Blood Flow Metab., 1: 313-327.
Kameyama, M., J. Suzuki, R. Shirane and A. Ogawa (1985) A new model of bilateral hemispheric ischemia in the rat - - three vessel occlusion model. Stroke, 16: 489-493.
Kitagawa, K., M. Matsumoto, M. Niinobe, K. Mikoshiba, R. Hata, H. Ueda, N. Handa, R. Fukunaga, Y. Isaka, K. Kimura and T. Kamada (1989) Microtubule associated protein 2 as a sensitive marker for cerebral ischemic damage: immunohistochemical in- vestigation of dendritic damage. Neuroscience, 31: 401-411.
Levine, S. and H. Payan (1966) Effects of ischemia and other procedures on the brain and retina of the gerbils. Exp. Neurol., 16: 255-262.
Levine, S. and D. Sohn (1969) Cerebral ischemia in infant and adult gerbils. Relation to incomplete circle of Willis. Arch. Pathol., 87: 315-317.
Levy, D.E. and J.B. Brierley (1974) Communications between verte-
89
bro-basilar and carotid arterial circulations in the gerbil. Exp. Neurol., 45: 503-508.
Loskota, W.M., P. Lomax and M.A. Verity (1974) A stereotaxic atlas of the mongolian gerbil brain (Meriones unguiculatus). Ann Ar- bor Science, Ann Arbor, MI.
Matsumoto, M., T. Hatakeyama, K. Yamamoto and T. Yanagihara (1987) A combined method for measurement of cerebral blood flow and immunohistochemistry for investigation of cerebral is- chemia. Brain Res., 424: 231-238.
Matsuyama, T., M. Matsumoto, A. Fujisawa, N. Handa, K. Tanaka, S. Yoneda, K. Kimura and H. Abe (1983) Why are infant gerbils more resistant than adults to cerebral infarction after carotid ligation?. J. Cereb. Blood Flow Metab., 3: 381-385.
Mrsulja, B.B., Y. Ueki and W.D. Lust (1986) Regional metabolite profiles in early stages of global ischemia in the gerbil. Metab. Brain Dis., 1: 205-220.
Niinobe, M., N. Maeda, H. Ino and K. Mikoshiba (1988) Characteri- zation of microtubule-associated protein 2 from mouse brain and its localization in the cerebellar cortex. J. Neurochem. 51:1132- 1139.
Pulsinelli, W.A. and J.B. Brierley (1979) A new model of bilateral hemispheric ischemia in the unanesthetized rat. Stroke, 10: 267- 272.
Smith, M.L., G. Bendek, N. Dahlgren, I. Rosen, T. Wieloch and B.K. Siesj6 (1984) Models for studying long-term recovery following forebrain ischemia in the rat. 2. A 2-vessel occlusion model. Acta Neurol. Scand., 69: 385-401.
Wojak, J.C., V. DeCrescito and W. Young (1991) Basilar artery occlusion in rats. Stroke, 22: 247-252.
Yamada, K., T. Hayakawa, T. Yoshimine and Y. Ushio (1984) A new model of transient hindbrain ischemia in gerbils. J. Neurosurg., 60: 1054-1058.
Yamada, K., T. Yoshimine, T. Hayakawa, T. Yanagihara, J. Taguchi, K. Morimoto, Y. Ushio and H. Mogami (1985) Selective vulnera- bility of brainstem neurons to ischemia. J. Cereb. Blood Flow Metab., 5 (Suppl. 1): $403-404.
Yao, H., S. Sadoshima, Y. Okada, S. lbayashi and M. Fujishima (1990) Hindbrain ischemia produced by bilateral vertebral artery occlusion and moderate hypotension in spontaneously hyperten- sive rats. Angiology 41: 848-854.





























