Embed Size (px)
Citation preview

Brain Research, 280 (1983) 287-298 287 Elsevier
Responses of Semicircular Canal and Otolith Afferents to Small Angle Static Head Tilts
in the Gerbil
ADRIAN A. PERACHIO and MANNING J. CORREIA
Departments of Otolaryngology, and Physiology and Biophysics, The University of Texas Medical Branch, Galveston, TX 77550-2 778 (U.S.A.)
(Accepted April 5th, 1983)
Key words: vestibular afferents - - anesthesia - - decerebration - - linear forces - - gerbils
The discharge activity of first-order vestibular neurons was recorded in anesthetized or decerebrated gerbils from the post-ganglion- ic fibers of the vestibular nerve. Semicircular canal afferents were distinguished from otolith afferents on the basis of their responses to linear and angular head acceleration. In decerebrated preparations, canal afferents exhibited significantly faster discharge activity (average = 87.8 impulses/s) than that of canal afferents in anesthetized preparations (average = 66.2 impulses/s), when the head was held to position the lateral semicircular canals coplanar with the earth horizontal plane (standard position).
The effects of changes in linear forces on vestibular afferent activity were determined by statically tilting the head + 10 ° about either the fore-aft and/or left-right head axes. A change in activity, from that recorded in the standard position, of 10% or greater was consid- ered significant. Using this criterion, significant changes in the tilt response in anesthetized animals were observed in both anterior (23 of 48 neurons, 48%) and lateral (22 of 31, 71%) canal afferents as well as otolith (18 of 25, 72%) afferents. In decerebrated prepara- tions for tilts around the pitch (left-right) axis, comparable effects were measured in (19 of 36, 53%) anterior and (17 of 30, 57%) lat- eral canal afferents. Neurons with irregular firing activity were more likely than regularly firing canal afferents to change their average discharge rate during static tilt. No significant differences in response magnitude to + 10 ° head tilt were found between canal and oto- lith afferents in anesthetized animals. Mechanisms to account for the responses to linear acceleration of canal afferents are discussed.
INTRODUCTION
Several lines of evidence have been deve loped that
support the proposa l that the semicircular canals may
respond to l inear accelerat ion of the head. In a vari- ety of species and types of preparations2,10,16,22, 25, it
has been noted that fibers within the ampul lary
nerves exhibit a l terat ions in their firing pat terns as a
consequence of changes in static head position.
Among the earl iest of such findings were those re-
por ted by Ledoux 22 for recordings made from semi-
circular canal afferents in the frog. In those studies,
the prepara t ion was viable for a l imited t ime since the
head was severed and the otic capsule was opened to
gain access to the nerves. Al though the effects were
observed ear ly in the course of the exper iments , they
could have been due to a pos t -mor tem phenomenon.
Lowenstein 25 observed similar effects in semicircular
canal afferent activity in the isolated e lasmobranch
labyrinth but qualif ied his in terpre ta t ion of the data
since in the exper imenta l p repara t ion the b lood sup-
ply to the labyrinth had been in ter rupted and the per-
i lymphatic space was opened. Ei ther of these manip-
ulations could have produced changes in the ionic
composi t ion of the labyrinthine fluids or a physical
change in the cupulal6, 2s. Subsequent to those stud-
ies, similar findings were repor ted for mammal ian
semicircular canal afferents in the cat 11 and squirrel
monkey 16. In both investigations, the post-ganglionic
fibers of the vest ibular nerve were exposed by aspi-
ration of overlying brain tissue. In a set of control ex-
per iments , Goldberg and Fernandez 16 demonst ra ted
in squirrel monkeys that in such prepara t ions , a ther-
mal gradient could be created across the labyrinth
which was of sufficient magni tude to produce a sensi-
tivity of the canals to l inear accelerat ion due to ther-
mally induced density differences. In those same ex-
periments , when the anter ior semicircular canal was
plugged, responses of anter ior canal ampul lary nerve
fibers to angular accelerat ions in the plane of that ca-
nal were abolished; however , they remained respon-
sive to l inear accelerat ion. Thus, al though much of
0006-8993/83/$03.00 © 1983 Elsevier Science Publishers B.V.

288
the data concerning the responses of semicircular ca- nal primary afferents to linear acceleration must be
interpreted with caution in light of several sources of
experimental artifacts, there remains an intriguing
possibility that such phenomena can be appropriately
considered as physiological in nature.
The primary purpose of the present investigation was to re-examine this problem utilizing a prepara-
tion in which the integrity of the labyrinth and inter-
nal auditory meatus was maintained along with that
of surrounding neural structures. We report here the
finding that a significant number of semicircular ca-
nal primary afferents in both the anesthetized and the
unanesthetized decerebrated gerbil exhibit a tonic
alteration of their discharge pattern during small an-
gle static head tilts. The resting discharge of each
identified afferent was determined in order to char-
acterize the variability of its firing rate. Brief and
preliminary descriptions of a portion of these results have been reported previously 6,2s.
MATERIALS AND METHODS
In the first set of experiments, recordings were
made from the post-ganglionic portion of the vestibu-
lar nerve in 17 anesthetized male gerbils (Meriones unguiculatus). Anesthesia consisted of a combination
of ketamine hydrochloride (40 mg/kg, i.m.) and ure-
thane (500 mg/kg, i.p,) As required, supplementary
doses of ketamine (20 mg/kg) were administered
throughout the experiment. The animals were placed in a stereotaxic frame with the incisor bar secured 8
mm below the zero, ear bar plane of the instrument. This placed the head in a position, which will be re-
ferred to as the standard position, that aligned the major plane of the lateral (horizontal) semicircular
canals approximately coplanar with the horizontal
stereotaxic plane. Alignment of the lateral canals was verified by opening the bulla and exposing the labyrinth following the end of most experiments.
In the second set of experiments, an additional group of 13 male gerbils were used to control for the potential effects of anesthesia and thermal changes in the temporal bone due to the presence of stereotaxic ear bars placed in the external auditory canal. The
animals were decerebrated by a precollicular tran- section according to the method of Grill and Nor- gren TM. First the gerbils were anesthetized either with
a short-acting barbiturate (Brevital, 50 mg/kg, i.p.) alone or in combination with ketamine (25 mg/kg,
i.m.). After placement in the sterotaxic frame all in-
cisions and pressure points were infiltrated with a lo-
cal anesthetic (xylocaine, 1.0%). An L-shaped
bracket, secured to an electrode carrier, was ce-
mented to the cranium at bregma. This allowed the ear bars to be removed with no displacement of the
animal from the standard position.
Micropipettes filled with 2 M NaCL, (10-18 Mr2 im- pedance) were driven through an opening (maximum
2 mm anterior-posterior by 4 mm medial/lateral) in the cranium into the intact cerebellum at an angle of
25 ° off the sagittal plane. In the decerebrated prepa-
rations, additional openings of 1 × 5 mm were made
over each hemisphere for passage of the spatula.
There was no retraction of muscle tissue or exposure
of the bullae of the temporal bone. Dental cement
covered the exposed cranium up to the opening over
the cerebellum. Prior to penetration of the post-gan-
glionic vestibular nerve fibers, neurons responsive to sound stimuli were usually encountered in the coch-
lear nucleus which lies dorsal to the root fibers of the
vestibular nerve. Verification of the recording site
was obtained in several animals by deposition of A1- cian blue dye ionophoreticaily23 (see Fig. 1) through
the recording pipettes at a locus where responsive
neurons were detected. In virtually all preparations,
primary afferents were identified at nearly identical stereotaxic coordinate locations.
Identification criteria Each vestibular afferent was identified according
to a set of physiological response characteristics. These were based on a modification of criteria de- rived originally by Estes and co-workers for cat vesti-
bular primary afferents u~. This procedure involved a
measurement of the firing activity of each neuron during a sinusoidai head angular acceleration at ap- proximately 1.0 Hz about an earth vertical axis while the animal's head was held in each static tilt position. The turntable to which the stereotaxic frame had been affixed was rotated manually or by a velocity- servo-controlled DC torque motor coupled to the central axis of the base of the table. Afferents inner- vating the vertical canals of the gerbil are responsive to angular acceleration about the yaw head axis when the head is held in the standard position27. Thus, af-

Fig. 1. Transverse section through the medulla illustrating the electrode tract (ET) of a micropipette that was driven through the intact cerebellum and brainstem into the post-ganglionic fi- bers of the vestibular nerve medial to Scarpa's ganglion (SG). Inset: magnified view of a portion of the same section con- taining SG cell bodies and fibers of the vestibular nerve. The site from which the activity of a semicircular canal afferent re- sponsive to static head tilt was recorded is marked by an Alcian blue dye spot (DS) ionophoretically deposited through the re- cording pipette.
ferents of the anterior semicircular canal could be
distinguished from other canal afferents by their re- sponse of decreased firing activity when the head was accelerated toward the recording side. Both lateral
and posterior canal afferents increased firing activity
with ipsilateral head acceleration. Further distinction among canal afferents was made possible by retesting
and noting each afferent 's response to acceleration
about the yaw axis after the head had been reposi- tioned to a roll tilt angle of +20 ° or greater. With a static roll tilt of the head to the ipsilateral side, a re- versal in the polarity of the response of vertical canal afferents was observed, i.e. anterior canal afferents were excited by ipsilateral yaw acceleration and post- erior canal afferents were inhibited. Although re-
289
duced in magnitude, the response of lateral canal af-
ferents when the head was tilted +20 ° was unaltered
in polarity. An additional group of fibers were classi-
fied as otolith afferents since they exhibited a system-
atic alteration in activity that was related to head tilt
position, but did not respond at any static head tilt
position to yaw angular acceleration t6.
Test paradigm The activity of each afferent was recorded while
the head of the animal was held in standard position
and after the head had been statically tilted through a
10 ° angle around either the longitudinal (roll) or
transverse (pitch) cardinal head axes. Recordings
were made both during repositioning of the head and
for a minimum of one-and-a-half minutes following
the tilt. The head was tilted slowly from one position
to another to minimize the cross coupling effects of
angular acceleration. The analyzed data were taken
from samples of neuronal activity that occurred from
30 to 60 s following cessation of head movement . In
decerebrated preparations, an abbreviated paradigm
was used. Static head tilts were restricted to the pitch axis and only canal afferents were systematically
tested.
Data analysis Magnetic tape recordings of the vestibular afferent
activity were analyzed to determine the average fir-
ing activity and the variability of discharge rate for
each neuron. Segments of data including 512 to 1024
interspike intervals were used for these calculations.
The coefficient of variation (CV = standard devia- tion/mean) was used as a measure of the regularity of
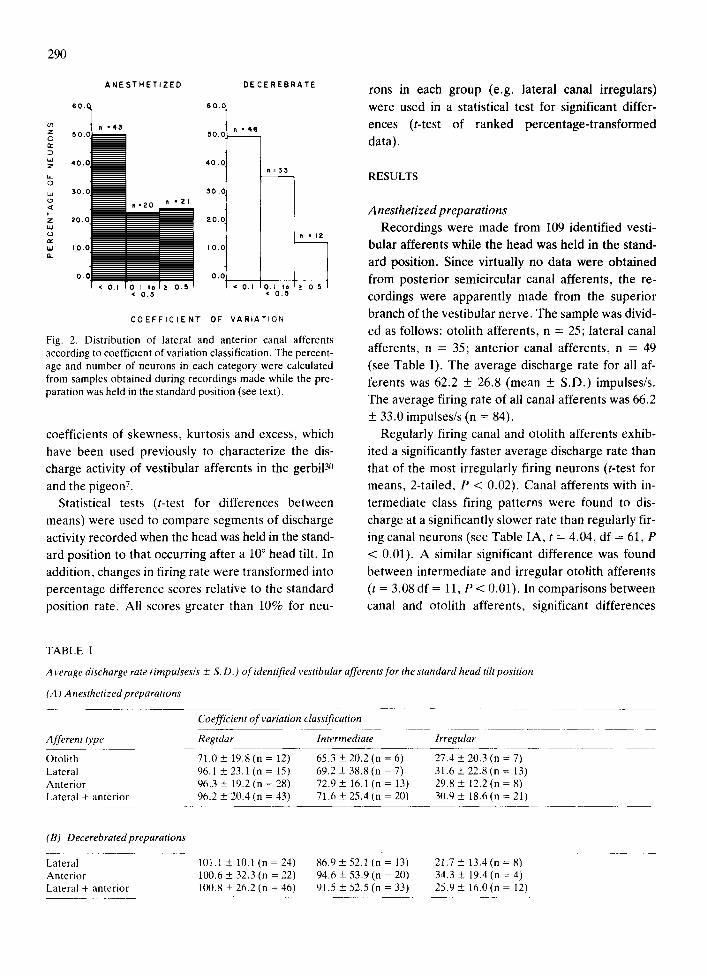
neuronal firing. A post-hoc analysis of all neurons classified as semicircular canal afferents (Fig. 2) indi-
cated that approximately half of the sample from
anesthetized preparations was composed of neurons that were highly regular in their firing rate with CVs
less than 0.1. The remainder of the sample could be
further divided in two roughly equal categories with
CVs falling into an intermediate range from 0.1 to 0.5 or in the range of highly irregularly firing neurons with CVs greater than 0.5. These classifications were based solely on data obtained from anesthetized ger- bils under the standard position test condition. Addi- tional first-order statistical properties of the dis- charge activity of all afferent neurons included the
290
ANESTHETIZED D E C E R E B R A T E
co Z 0
uJ (.9 <I
t~J o C~ UJ I1.
6 0 . (
5 0 . C
4 0 . C
3 0 . C
2 0 . 0
I 0 , 0
0 . ¢
6 0 , 0
n ' 4 G 50.C
4 0 .q
210 ,,
2 0 . 0
I O . C
0 . 0
< 0 .1
. = 3 3
I O . I to < 0 . 5 < 0 . 5
. - 1 2
COEFFICIENT OF VARIATION
Fig. 2. Distribution of lateral and anterior canal afferents according to coefficient of variation classification. The percent- age and number of neurons in each category were calculated from samples obtained during recordings made while the pre- paration was held in the standard position (see text).
coeff icients of skewness , kurtosis and excess, which
have been used prev ious ly to charac ter ize the dis-
charge activity of ves t ibular af ferents in the gerb iP 0
and the p igeon 7.
Statistical tests (t-test for d i f ferences be tween
means) were used to c o m p a r e segment s of discharge
activity r eco rded when the head was held in the stand-
ard posi t ion to that occur r ing af ter a 10 ° head tilt. In
addi t ion, changes in firing rate were t r ans fo rmed into
percen tage d i f ference scores re la t ive to the s tandard
posi t ion rate. All scores g rea te r than 10% for neu-
rons in each g roup (e.g. la teral canal i r regulars)
were used in a statistical test for s ignif icant differ-
ences (t-test of r anked p e r c e n t a g e - t r a n s f o r m e d
data) .
RESULTS
Anes the t i zed preparat ions
Record ings were m a d e f rom 109 ident i f ied vesti-
bular afferents whi le the head was he ld in the s tand-
ard posi t ion. Since v i r tua l ly no da ta were ob t a ined
f rom pos te r io r semic i rcu lar canal af ferents , the re-
cordings were apparen t ly m a d e f rom the super io r
branch of the ves t ibular ne rve . T h e sample was divid-
ed as follows: o to l i th af ferents , n = 25; la tera l canal
afferents , n = 35; an te r io r canal af ferents , n = 49
(see Tab le I). The ave rage discharge ra te for all af-
ferents was 62.2 + 26.8 (mean + S .D . ) impulses/s .
The average firing rate of all canal af ferents was 66.2
+ 33.0 impulses/s (n = 84).
Regula r ly firing canal and o to l i th af ferents exhib-
ited a significantly faster ave rage d ischarge ra te than
that of the most i r regular ly firing neurons (t-test for
means , 2- tai led, P < 0.02). Canal af ferents with in-
t e rmed ia t e class firing pa t te rns were found to dis-
charge at a significantly s lower rate than regular ly fir-
ing canal neurons (see Tab le I A , t = 4.04, df = 61, P
< 0.01). A similar s ignif icant d i f fe rence was found
be tween in t e rmed ia t e and i r regular o to l i th af ferents
(t = 3.08 df = 11, P < 0.01). In compar i sons b e t w e e n
canal and otol i th af ferents , s ignificant d i f ferences
TABLE I
Average discharge rate ( impulses/s + S. D. ) of identified vestibular afferents for the standard head tih position
(A ) Anesthetized preparations
Coefficient of variation classification
Afferent type Regular Intermediate Irregular
Otolith 71.0 + 19.8 (n = 12) 65.3 + 20.2 (n = 6) Lateral 96.1 + 23.1 (n = 15) 69.2 + 38.8 (n = 7) Anterior 96.3+ 19.2 (n =28) 72.9+ 16.1 (n = 13) Lateral + anterior 96.2 + 20.4 (n = 43) 71.6 + 25.4 (n = 20)
27.4 + 20.3 (n = 7) 31.6 + 22.8 (n = 13) 29.8 + 12.2 (n = 8) 30.9 ___ 18.6 (n = 21)
(B) Decerebrated preparations
Lateral Anterior Lateral + anterior
101.1 _+ 10.1 (n =24) 100.6 + 32.3 (n = 22) 100.8 + 26.2 (n = 46)
86.9 + 52.1 (n = 13) 94.6 + 53.9 (n = 20) 91.5 --- 52.5 (n = 33)
21.7 _+ 13.4 (n = 8) 34.3 + 19.4 (n = 4) 25.9 + 16,0 (n = 12)
were found for regularly firing neurons when tested
in the s tandard posi t ion; otol i th afferents fired at a
significantly slower rate than canal afferents (otoli th
vs lateral , t = 2.87, df = 25, P < 0.01; otoli th vs ante-
rior, t = 3.68, df = 38, P < 0.01). Fur the rmore , post-
hoc analyses comparing average discharge rate and
coefficients of skewness and excess for samples of af-
ferent activity for the s tandard posi t ion, be tween ca-
nal neurons that were subsequent ly found to respond
to l inear accelerat ion and the rest of the canal affer-
ents (within each CV category) , revea led no signifi-
cant differences.
A f f e r e n t activity - - static head tilt
Not all neurons could be tested for all head tilt con-
ditions; however , except for 5 canal afferents, data
were obta ined for each afferent (n = 104) for the con-
ditions of + 10 ° head tilt a round ei ther the pitch or roll
cardinal head axis. Neurons that were classified as
otol i th afferents (i.e. those that did not respond to
angular head accelerat ion for any head tilt posi t ion)
were usually tested by head tilts in whichever of the
two vertical planes was closest to their plane of maxi-
mal sensitivity. There was no a t tempt to maximize
the o to l i tha f fe ren t responses by defining their polar-
ization vectors 11.
A large percentage of the total tested sample of ca-
nal afferents exhibi ted a change in average firing rate
~ I ~ NOSE UP I0 °
STANDARD POSI TION
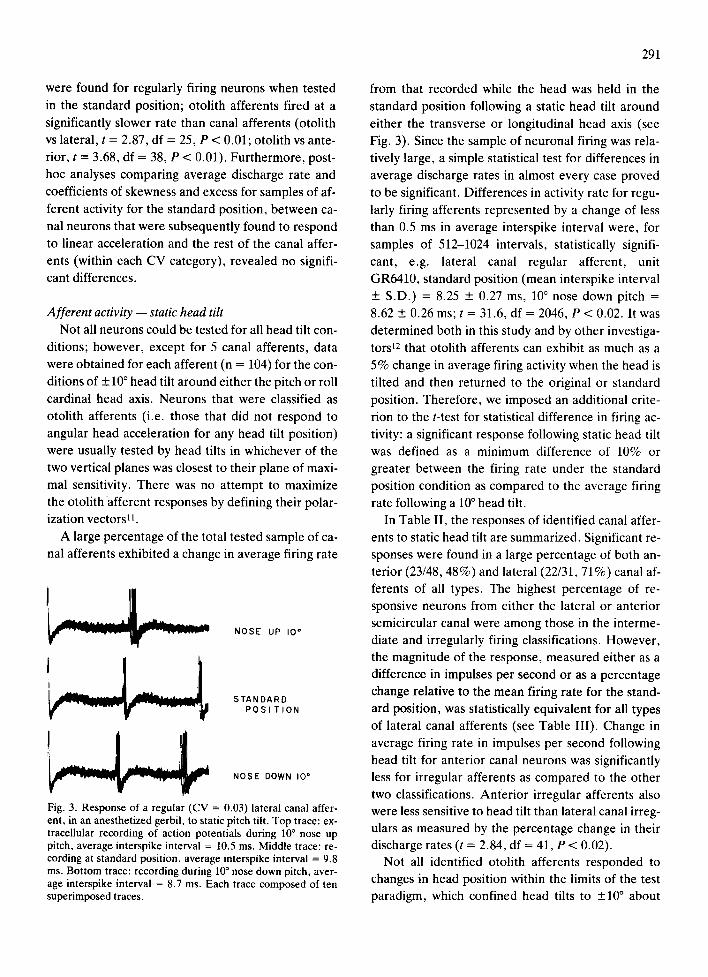
Fig. 3. Response of a regular (CV = 0.03) lateral canal after- ent, in an anesthetized gerbil, to static pitch tilt. Top trace: ex- tracellular recording of action potentials during 10 ° nose up pitch, average interspike interval = 10.5 ms. Middle trace: re- cording at standard position, average interspike interval = 9.8 ms. Bottom trace: recording during 10 ° nose down pitch, aver- age interspike interval = 8.7 ms. Each trace composed of ten superimposed traces.
291
from that recorded while the head was held in the
s tandard posi t ion following a static head tilt a round
ei ther the transverse or longitudinal head axis (see
Fig. 3). Since the sample of neuronal firing was rela-
tively large, a simple statistical test for differences in
average discharge rates in almost every case proved
to be significant. Differences in activity rate for regu-
larly firing afferents represented by a change of less
than 0.5 ms in average interspike interval were, for
samples of 512-1024 intervals, statistically signifi-
cant, e.g. lateral canal regular afferent , unit
GR6410, s tandard posit ion (mean interspike interval
_+ S.D.) = 8.25 _+ 0.27 ms, 10 ° nose down pitch =
8.62 + 0.26 ms; t = 31.6, df = 2046, P < 0.02. It was
de te rmined both in this study and by o ther investiga-
tors 12 that otoli th afferents can exhibit as much as a
5% change in average firing activity when the head is
t i l ted and then re turned to the original or s tandard
position. Therefore , we imposed an addit ional crite-
rion to the t-test for statistical difference in firing ac-
tivity: a significant response following static head tilt
was defined as a minimum difference of 10% or
greater between the firing rate under the s tandard
posit ion condit ion as compared to the average firing
rate following a 10 ° head tilt.
In Table II , the responses of identif ied canal affer-
ents to static head tilt are summarized. Significant re-
sponses were found in a large percentage of both an-
ter ior (23/48, 48%) and lateral (22/31, 71%) canal af-
ferents of all types. The highest percentage of re-
sponsive neurons from ei ther the lateral or anter ior
semicircular canal were among those in the interme-
diate and irregularly firing classifications. However ,
the magni tude of the response, measured either as a
difference in impulses per second or as a percentage
change relat ive to the mean firing rate for the stand-
ard posit ion, was statistically equivalent for all types
of lateral canal afferents (see Table III) . Change in
average firing rate in impulses per second following
head tilt for anter ior canal neurons was significantly
less for i rregular afferents as compared to the other
two classifications. An te r io r i rregular afferents also
were less sensitive to head tilt than lateral canal irreg-
ulars as measured by the percentage change in their
discharge rates (t = 2.84, df = 41, P < 0.02).
Not all identif ied otoli th afferents responded to
changes in head posit ion within the limits of the test
paradigm, which confined head tilts to +10 ° about
292
TABLE II
(A) Semicircular canal afferents in anesthetized animals that responded* to linear acceleration
Afferent type CV class Pitch Roll
No. tested % respond. No. tested % respond.
Anterior
Lateral
Regular 27 15% 13 23% Intermed. 13 62% 8 100% Irregular 7 86% 5 80%
Regular 13 31% 2 0% Intermed. 6 100% 2 50% Irregular 12 100% 10 80%
(B) C•mparis•n •f the percentage •f latera• and anteri•r semi•ircu•ar cana• afferents in anesthetized and decerebrated preparati•ns that responded to pitch tilt
Decerebrated Anesthetized
CV class No. tested % respond. No. tested % respond.
Regular 31 26% 40 20% Intermed. 27 74% 19 74% Irregular 8 88% 19 95%
* Difference of 10% or greater in mean firing rate after 10 ° static tilt from standard position.
TABLE III
Afferent type Regular Intermediate Irregular
(A ) Average percentage change (+ S.D.) in discharge rate of responsive neurons, in anesthetized animals following 10 ° static head tilt Otolith 15.3 + 7.4 (13)* 20.4 + 13.0 (8) 26.6 + 14.2 (9) Lateral 11.2 + 0.4 (4) 20.5 + 12.0 (12) 36.2 + 26.1 (29) Anterior 13.2 + 3.0 (8) 16.3 _+ 6.0 (20) 19.4 + 13.9 (14)
(B) Average change in discharge rate of responsive neurons in anesthetized animals in impulses per second (+ S.D.) Otolith 10.0 + 2.9 9.8 + 3.1 6.1 + 3.0 Lateral 9.0 + 1.9 9.8 + 8.0 7.4 + 4.9 Anterior 9.9 + 3.4 10.1 + 4.3 4.8 + 3.1
* Total number of responses for all tilt positions in parentheses; some neurons represented for multiple tests.
the pitch and roll head axes. No signif icant differ-
ences were found be tween the 18 responsive otol i th
afferents and the responsive lateral and an te r ior ca-
nal afferents in te rms of the magn i tude of the change
of firing rate (see Tab le III) . A m o n g otol i th neu rons ,
regularly firing afferents differed significantly from
irregularly firing neu rons . In te rms of percentage
change, i r regular otol i th afferents exhibi ted a signifi-
cantly greater response to static head tilt; however ,
the difference in firing rate measu red in impulses per
second was significantly grea ter for regular ly firing
otoli th neurons .
Directional sensitivity
Al though a n u m b e r of canal afferents exhibi ted
systematic responses re la ted to the direct ion of head
tilt, no single rule appl ied to all neurons . This was in
part because the full test pa rad igm could no t be ap-
plied to the ent i re sample. The fol lowing results per-
tain to anes thet ized animals : of the lateral canal af-
ferents that were responsive to backward pitch tilt
(15/31), all exhibi ted a decrease in discharge activity.
More were affected by forward pitch; of these, 70%
(12/17) r e sponded with an increase in discharge rate.
Of the lateral canal neu rons that r e sponded to roll tilt

to the contralateral side, approximately 80% (5/6)
were inhibited; however, ipsilateral roll tilts were ap-
proximately equally effective for inducing excitatory
or inhibitory effects. Less consistent response pat-
terns were observed for anterior canal afferents.
One-third of the tested anterior canal neurons
(16/48) exhibited a decrease in their firing rate for
any tested direction of head tilt. Of all anterior canal
afferents tested by head pitch, approximately one- half (18/37) responded. Most of these neurons
(11/18) showed inhibitory responses. Ten anterior
canal afferents were tested with ipsilateral roll head
tilt; all exhibited inhibitory responses.
In most cases, the magnitudes of the responses for
any given plane of head tilt were asymmetric. Ap-
proximately 32% (7/22) of responsive lateral canal afferents were sensitive to both forward and back-
ward 10 ° head tilts, increasing their discharge rate in
the nose down position and decreasing their dis-
charge rate in the nose up position, Among this small
number, there was no significant difference in the av-
erage response or percentage change in neuronal ac-
tivity for forward as compared to backward pitch;
however, for individual neurons, the response mag-
nitude was in some cases as much as 3-fold more sen-
sitive for the nose up tilt condition. Similarly, of all
responsive anterior canal afferents, approximately
30% (7/23) had a significant bidirectional response to
roll or pitch static head tilt. However , unlike the bidi- rectionally responsive lateral canal afferents, no con-
sistency was found in response directionality across
anterior canal neurons. Most canal afferents were
found to be sensitive for only one direction of tilt for a given head plane. Thus, for the tilt planes and angles
used in this study, a vector of maximum sensitivity could not be defined for the directionality and magni-
tude of the responsive canal afferents to linear acce- leration.
Decerebrated preparations Following experiments on anesthetized animals, a
second set of experiments was conducted on decere- brated animals, specifically to test whether the pres- ence of centrally acting anesthetics and/or the place-
ment of restraining ear bars in the external auditory canal have any significant influence on vestibular af- ferents' spontaneous activity or the response of canal afferents to static head tilt. Data were obtained on
293
the spontaneous activity of lateral (n = 45) and ante-
rior (n = 46) canal afferents from 13 decerebrated
gerbils. The average discharge rate compared to that
of canal afferents in anesthetized animals was signifi-
cantly faster, i.e. 87.6 + 44.2 impulses/s in decere-
brated gerbils vs 66.2 + 33.0 impulses/s in anesthe- tized animals (t = 3.50, df = 173, P < 0.001). In both
anesthetized and decerebrated preparations, a sig-
nificant inverse correlation existed between the CV
and average discharge rate of canal neurons: lateral and anterior canal afferents in anesthetized gerbils, r
+ ~0 .83 , n = 84, P < 0.001; lateral and anterior ca-
nal afferents in decerebrated gerbils, r = ----0.48, n =
91, P < 0.01. The reduced correlation for decere-
brated preparations was related in part to the in-
crease of the average firing activity of intermediate
neurons, e.g. 0.1 ~< CV ~< 0.5. In contrast to the anes- thetized preparations, there was no significant differ-
ence in the average discharge rate of regular vs inter-
mediate canal neurons (see Table IB, t = 1.02, df =
77, P > 0.05). The response of canal neurons to static head tilt
was tested in the decerebrated preparations by pitch
tilts about the left-right head axis alone. A compara-
ble percentage of canal afferents in decerebrated ani-
mals to that found in anesthetized gerbils responded
with a change of 10% or greater in discharge rate
when the head was pitched +10 ° nose up or down from the major plane of the lateral semicircular canal
(see Table liB). As in anesthetized animals, there
was no consistent directional sensitivity that typified those responses.
A post-hoc assessment of the relationship between percentage change in firing activity relative to that of the standard position, for +10 ° pitch tilt, was made
for data obtained from all tested canal afferents in
both types of preparations. A significant correlation between CV and percentage change (independent of
sign) was found for both anesthetized and decere- brated samples of canal afferents with the exception
of anterior canal afferents in decerebrated animals
(see Fig. 4). Upon inspection of the data, it appears that the a priori criterion of a 10% change in firing ac- tivity as a minimum limit for defining a significant
change compares favorably with the results of a lin- ear regression analysis for the middle CV range. For regular and irregular neurons, a 10% criterion may have been an over or under estimation, respectively,

294
A N E S T H E T I Z E D DECEREBRATE
W (.9
Z <~ "I-
W (.9 <~
Z W
rr" W (3.
60 .q
50 .q
4 0 . 0 .
3o.1
20.~
l O,i .
o . o 0~0
6 0 . (
5 0 . (
4 0 . (
3 0 . (
2 0 . (
LATERAL
o * o
÷ . . . . . - ~ o
. . - - " ' o
. - - - - ' " ' o o o
. '2 . '4 . '6 .'8 I'.0
4 1 4 C V * 3 4
0 ~ ,321 c v e 4 4
A N T E R I O R
o oo4 • **
a o i c .~ 6 _ ~ ~ ~. . .=: - : . :2- - - _
o.( ~ : .; 9. o •
0 : 6 , '2 ' 4 [ 6 I e I~ 0
n -48 t • 0 . 32" ÷ A'/~ • 8.ZCV + 4 . 0
1~ , 4 6 r 10 .50 , o A % , 1 2 . 7 c v + 4 I
LATERAL
6 0 .
5 0 . ,
4 0 J
o o
3 0 H
2 0 . ( o * ................
, o 0 : L " "
0 . 0 i ~ ~ °
Om*O [ "m~ r *]4 ' . '6 ' . 'e ' i '.o
÷ * , 28 r ,O .g3" A % , 2 5 . 0 C V + 4 2
o { ~ ; . . . , • o . . • 22 .3CV + 3. l
6 0 . 0
5 0 .I
4 0 J
3 0 . 0
2 0 . 0
I 0 . 0
0 . 0
0 . 0
ANTERIOR
o o
o
. I 2 I . ' 4 I 1 6 I r 8 I i 1,0
~ 135 [ I 0 Z7 + & % i 1 4 I O C V + 5 , (
n ,33 r "010 ] 0 { ~ , % , 5 . 3 C V + I I . 5
COEFFICIENT OF VARIATION
Fig. 4. Relationship, in canal afferents in anesthetized and decerebrated gerbils, between coefficient of variation and percentage change (A%) in average interspike interval relative to the standard position for 10% nose up (+) or nose down (O) static head tilt. Criterion level of 10% change (see text) is indicated by long dashed line. Correlation coefficients (r) were computed for each canal type for each condition (*P < 0.01, **P < 0.05). Note that the 4 highest values for lateral canal afferents in anesthetized preparations were greater than 60% change in interspike interval. Linear least-squares best-fit lines and equations are shown for each test condi- tion: solid line for nose up tilt, short dashed line for nose down tilt.
of significant changes. Nevertheless, the overall per-
centage of responsive neurons would not have been
different from that determined on the basis of the
original criterion. That is 48% (23 of 38 neurons) of
anterior afferents in anesthetized preparations and
53% (19 of 36 neurons) in decerebrated preparations
responded with a 10% change. Comparable figures
for lateral canal afferents in anesthetized gerbils
were 71% (22 of 31 neurons) and 57% (17 of 30 neu-
rons) in decerebrated gerbils.
DISCUSSION
Responses of vestibular primary afferents to linear ac- celeration
A large percentage of vestibular semicircular canal

afferents responded to changes in linear acceleration produced by static head tilts. The results do no t ap- pear to be artifactual since, in the types of prepara- tions used in this study, the labyrinth and brain re- mained intact and vestibular nerve and petrous bone were not surgically exposed, thus presumably avoid- ing both ionic imbalances that might produce fluid/ tissue density differences and thermal gradients across exposed or deeper lying portions of the lab- yrinth15, 25. Moreover, in the decerebrated prepara-
tions, the ear bars were removed. Although decere- brated preparations differed from the anesthetized animals, in terms of spontaneous activity of the ves- tibular afferents, the response of canal afferents to static head tilt was still observed. It also appears un- likely that the findings can be accounted for by a me- chanical effect produced by movement of the ner- vous tissue with respect to the electrode tip. Within the same preparation and for the same electrode pen- etration, successively encountered neurons were ei- ther unresponsive or responsive to head tilting with different directional sensitivities, i.e. anterior canal neurons differed in the directionality of their re- sponses while responsive lateral canal neurons exhib- ited a fairly consistent bidirectional response. Aside from action potentials, the only other monitored physiological parameter was body core temperature; thus, it was not possible to definitively determine whether other physiological functions varied over the course of the experiment. Such events are also un- likely to account for the responses to linear accelera- tion since positive and comparable effects were ob- tained in all preparations, at all stages of the experi- ments.
Possible mechanisms for canal neuron responses to
linear acceleration
Four features of the labyrinth suggest possible mechanisms that may account for responses of physi- ologically identified semicircular canal neurons to changes in static head tilt position. First, Caston and GribenskP. 4 have proposed, on the basis of physio- logical studies, that the saccule and semicircular ca- nals in the frog are directly related by receptor-to-re- ceptor fibers that apparently form connections be- tween neurons within Scarpa's ganglion 4. Their find- ings suggest that the mechanism for the responses of canal neurons to gravity might act through an addi-
295
tional input afferent from otolith receptors. Al-
though this proposal offers an interesting possibility, Caston and GribenskP, 4 reported that 21% (12/57 of tested neurons) of anterior canal afferents exhibited an inhibitory response to electrical stimulation of the saccule. This figure is smaller than our sample of tilt- sensitive anterior canal afferents (23/48, 48%). Moreover, there is limited anatomical evidence for intrinsic connections between neurons within Scar- pa's ganglion 13.
A second anatomical feature of primary vestibular neurons that may imply a mechanism for connection of individual neurons to different receptors in the lab- yrinth is the recently reported finding of a number (39/928) of multipolar cells with unmyelinated den- dritic processes in the vestibular ganglion of the cat 5. At present, the receptor innervation of multipolar neurons has not been identified; however, it has been established that the multipolar neurons are primary vestibular afferents that project to the vestibular nu- clei. The extent of dendritic branching in those neu- rons, particularly with regard to their distribution to more than one receptor within the labyrinth, is not known; nor has it been established whether intrinsic synaptic connections exist between those neurons. However, such arrangements could offer a morpho- logical basis for preganglionic or intraganglionic in- teractions between the labyrinthine neurons of linear (otolith organs) and angular (semicircular canal) ac- celeration sensors.
A third characteristic of the anatomy of the lab- yrinth was identified by Estes and colleagues10 as a possible mechanism to account for the gravity sensiti- vity of semicircular canal afferent neurons. They noted that the base of the crista of all 3 canals pro- jects into the vestibule and is supported by the endo- lymphatic membrane which also underlies the ultri- cular macula. They suggested that the mass of the utricule can exert varying degrees of tension, de- pending on the head tilt angle, along the membrane thus producing movement of the crista with respect to the cupula. This morphological feature was also cited to account for the polarization of the response of anterior and lateral canal afferents to roll tilt. Our findings, based on a large sample of neurons, were more equivocal concerning the directionality of the canal roll tilt response. However, one finding consis- tent with the prediction of the proposal of Estes et

296
al. m was that static roll head tilt toward the ipsilateral
ear produced an inhibition in all responsive anterior
canal neurons. It is more difficult to determine the
mechanical effects of the membranous coupling be-
tween the crista and the utricle during pitching mo-
tions of the head. However, assuming that a back-
ward head pitch causes the crista to be pulled toward
the utricle, decreased firing activity in lateral canal
afferents would be predicted, a result we observed.
The centrally originating efferent innervation of
the vestibular receptors is a fourth anatomical entity
which may act as a potential mechanism to modify
the activity of the primary vestibular afferents in re-
sponse to a change in head tilt position. In mamma-
lian species, electrical stimulation of the efferent sys-
tem has been found to produce either inhibitory 29 or
excitatory ~7 effects on the resting activity of vestibu-
lar primary afferents. The excitatory effect, meas-
ured in squirrel monkeys, was of markedly greater
magnitude in the irregularly discharging afferents 17.
This finding may have a bearing on the finding of our
study that the greatest percentage of canal neurons responsive to static head tilt were irregularly firing.
The vestibular efferent system consists of relatively few neurons 14,17 that appear to branch extensively
and innervate all receptor organs of the labyrinth 24.
These physiological and anatomical findings suggest that the efferent vestibular system may operate in a
feedback manner to modify the discharge rate of ca-
nal afferents as the head is statically tilted. Since rela-
tively little is known about the efferent inputs and
their response characteristics to natural vestibular stimulation, especially in warm-blooded animals 2l,
the role of the efferents in the response of canal after- ents to linear forces remains speculative.
Relative sensitivity of semicircular canal and otolith afferents
Semicircular canal afferents are, according to our criteria for response, as sensitive as otolith afferents
to small angle head tilts restricted to the cardinal head axes. This restriction is an important qualifica- tion since head tilts only in the two orthogonal planes are not sufficient to determine the actual linear force sensitivity of vestibular neuronslJ. Nevertheless,
within the limits of our test procedures, there was a remarkable overlap in the range of sensitivity for all types of afferents and no significant average differ-
ence between the responses of presumed otolith af-
ferents and identified canal afferents. The most im-
portant distinction in the canal afferent response to
static tilt as compared to that observed in the tested otolith afferents was a lack of symmetry or direction-
ality of response related to the functional and mor-
phological polarization of the receptors. In lieu of
any direct explanation for this finding, it may only be
speculated that in addition to the possible mech-
anisms discussed above, the vectors of sensitivity of
canal neurons responsive to linear forces are difficult
to define because of the complex geometry of the
neuroepithelial surface of the crista ampullaris. Important issues for future study include the deter-
mination of: (1) whether linear acceleration-sensitive
canal afferents exhibit a polarization vector; (2) the
degree of consistency of their response directionali- ty; and (3) the symmetry of the tilt response meas-
ured over greater head tilt angles. Estes et al.10 re-
ported results from a few gravity-sensitive canal neu-
rons tested over larger head tilt angles, e.g.
180°-360 °, that exhibited a systematic and replicable
change in the magnitude and directionality of their
responses to changes in head position, especially with roll tilt. In our study, two lateral canal neurons,
tested with pitch tilt of up to +20 ° , similarly exhibited a significant relationship between head tilt angle and
discharge rate. Moreover, a significant inverse corre-
lation was found between their mean firing rate and
their CV. These responses to static linear force imply that canal neurons may also exhibit responses to dy-
namic linear acceleration. In addition, their dynamic response characteristics to angular acceleration may be altered (other than by changes in gain or sensitivi-
ty) as a function of head position with respect to the
gravity vector, since it has been demonstrated that the transfer characteristics of canal neurons are cor- related with the CV of their firing rates15, 30.
Implication for vestibular function The finding that the semicircular canals respond to
linear forces requires a re-interpretation of a number of experimental findings on labyrinthine function and may have important implications for clinical phe- nomena. Goldberg and Fernandez 16 have suggested
that this property of the canals may account for the clinically observed phenomenon of a positional nystagmus observed in some apparently normal sub-

297
jects 1. In squirrel monkeys ablation of the maculae
did not abolish a pre-existing idiopathic positional
nystagmus that can be reliably observed in individual
animals of that species t9. These and other phenome-
na that are sequelae to head t rauma 31 may be re-in-
terpreted .in terms of a contr ibut ion from a canal
gravity-sensitive mechanism.
A number of investigators have reported that neu-
rons within the vestibular nuclei apparently receive
converging inputs from the canals and the otolith or-
gans since these neurons respond to both angular and
linear accelerationS, 9. An assumption common to
both those studies was that slow or static tilting of the
head or other forms of l inear force would stimulate
only otolith related inputs to the vestibular nuclei
while angular head rotation would affect only canal
related afferents. Although canal/otolith conver-
gence on second- or higher-order vestibular neurons
is likely, it cannot be definitely proven by such lim-
ited neurophysiological criteria.
ACKNOWLEDGEMENTS
The authors wish to acknowledge the expert tech-
nical assistance of H. Hughes and S. Steffens and to
thank S. Rainey for typing the manuscript. The col-
laboration of Dr. T. Clegg on early portions of this
study and the helpful suggestions of Dr. D. J. Ander-
son concerning the post-hoc correlational analysis
are gratefully acknowledged. This research was sup-
ported by the National Aeronautics and Space Ad-
ministration through Grant NAG2-26 and Contracts
NAS9-14641 and NAS2-10502.
REFERENCES
1 Barber, H. O. and Wright, G., Positional nystagmus in nor- mals, Advanc. Otorhinolaryngol., 19 (1973) 276-285.
2 Budelmann, B. U. and Wolff, H. G., Gravity response from angular acceleration receptors in Octopus vulgaris, J. Comp. Physiol., 85 (1973) 283-290.
3 Caston, J. and Gribenski, A., Influence of the saccule, la- gena and vertical posterior canal on the ipsilateral horizon- tal and vertical anterior semicircular canals in Rana escu- lenta: role of receptor-receptor fibres, Pflugers Arch. ges. Physiol., 372 (1977) 139-143.
4 Caston, J. and Gribenski, A., New findings about interrela- tions between vestibular receptors in the frog, J. Neuro- physiol., 47 (1982) 55-59.
5 Chat, M. and Sans, A., Multipolar neurons in the cat vesti- bular ganglion, Neuroscience, 4 (1979) 651-657.
6 Clegg, T., Perachio, A. A. and Correia, M. J., Tilt re- sponses of semicircular canal primary afferents, Otolaryn- gol. Head-Neck Surg., 90 (1982) 103-107.
7 Correia, M. J. and Landolt, J. P., A point process analysis of the spontaneous activity of anterior semicircular canal units in the anesthetized pigeon, Biol. Cybernet. 27 (1977) 199-213.
8 Curthoys, I. S. and Markham, C. H., Convergence of la- byrinthine influences on units in the vestibular nuclei of the cat. I. Natural stimulation, Brain Research, 35 (1971) 469-490.
9 Duensing, F. and Schaefer, K. P., Uber die Konyergenz verschiedener labyrintharer Afferenzen auf einzelne Neu- rone des Vestibulariskerngebietes, Arch. Psychiat. Nervenkr., 199 (1959) 345-371.
10 Ehrenbrand, V. F. and Wittemann, G., Uber synaptische Strukturen im Ganglion vestibulare der Maus, Anat. Anz. Bd., 126 (1970) 300-308.
11 Estes, M. D., Blanks, R. H. I. and Markham, C. M., Physi- ologic characteristics of vestibular first-order canal neurons in the cat. 1. Response plane determination and resting dis-
charge characteristics, J. Neurophysiol., 38 (1975) 1232-1249.
12 Fernandez, C. and Goldberg, J. M., Physiology of periph- eral neurons innervating otolith organs of the squirrel mon- key. 1. Response to static tilts and to long-duration centri- fugal force, J. Neurophysiol., 39 (1976) 970-984.
13 Fernandez, C., Goldberg, J. M. and Abend, W. K., Re- sponse to static tilts of peripheral neurons innervating oto- lith organs of the squirrel monkey, J. Neurophysiol., 35 (1972) 397-997.
14 Gacek, R. R. and Lyon, M., The localization of vestibular efferent neurons in the kitten with horseradish peroxidase, Acta otolaryngol., 77 (1974) 92-101.
15 Goldberg, J. M. and Fernandez, C., Physiology of periph- eral neurons innervating semicircular canals of the squirrel monkey. III. Variations among units in their discharge properties, J. Neurophysiol., 34 (1971) 676-684.
16 Goldberg, J. M. and Fernandez, C., Responses of periph- eral vestibular neurons to angular and linear acceleration in the squirrel monkey, Acta otolaryngol., 80 (1975) 101-110.
17 Goldberg, J. M. and Fernandez, C., Efferent vestibular system in the squirrel monkey: anatomical location and in- fluence on afferent activity, J. Neurophysiol., 43 (1980) 986-1025.
18 Grill, H. J. and Norgren, R., The taste reactivity test. II. Mimetic responses to gustatory stimuli in chronic thalamic and chronic decerebrate rats, Brain Research, 143 (1978) 281-297.
19 Kato, Y. M., Igarashi, M., Takahashi, M. and Wright, W. K., Effect of vestibular deafferentation upon positional nystagmus in the squirrel monkey, J. Laryngol. D. Otol., 40 (1976) 619-638.
20 Keller, E. L., Behavior of horizontal semicircular canal af- ferents in alert monkey during vestibular and optokinetic stimulation, Exp. Brain Res., 24 (1976) 459-471.
21 Klinke, R. and Galley, N., Efferent innervation of vestibu- lar and auditory receptors, Physiol. Rev., 54 (1974) 316-357.

298
22 Ledoux, A., Activite electrique des nerfs des cannaux semi- circulaires, du saccule et de l'utricule chez la grenouille, A cta otorhinolaryngol. (Belg.), 3 (1949) 335-349.
23 Lee, B. B., Mandl, G. and Stean, J. P. B., Micro-electrode tip position marking in nervous tissue: a new method, Elec- troenceph, clin. Neurophysiol., 27 (1969)610-613.
24 Lindeman, H. H., Studies on the morphology of the sensory regions of the vestibular apparatus, Advanc. Anat. Cell Biol., 42 (1969) 1-113.
25 Lowenstein, O., Physiology of vestibular receptors. In A. Brodal and O. Pompeiano (Eds.), Progress in Brain Re- search, Vol. 37, Basic Aspects of Central Vestibular Mech- anisms, Elsevier, Amsterdam, 1972, pp. 121-127.
26 Lowenstein, O., Comparative morphology and physiology, Ch. II. In H. H. Kornhuber (Ed.), Handbook of Sensory Physiology, Vol. VIII, Vestibular System, Part 1, Basic
Mechanisms, Springer-Verlag, New York, 1974, pp. 75-122.
27 Perachio, A. A. and Correia, M. J., Transfer characteris- tics of anterior semicircular canal afferents in the anesthe- tized gerbil, Soc. Neurosci. Abstr., 6 (1980) 558.
28 Perachio, A. A., Correia, M. J. and Clegg, T., Responses of semicircular canal and otolith afferents to linear accele- ration, Soc. Neurosci. Abstr., 7 (1981) 148.
29 Sala, O., The efferent vestibular system. Electrophysiolog- ical research, Acta otolaryngol., Suppl. 197 (1965) 1-34.
30 Schneider, L. W. and Anderson, D. J., Transfer character- istics of first and second order lateral canal vestibular neu- rons in gerbil, Brain Research, 112 (1976) 61-76.
31 Schuknecht, H. E., Pathology of the Ear, Harvard Univ. Press, Cambridge, MA, 1974.





























