Embed Size (px)
Citation preview

Insects that feed on toxic plants (adaptation)
Prepared by: Hael Saeed Ahmed Raweh (436107622)
Ph.D. 2nd SemesterCollege of Food and Agriculture SciencesKing Saud University, Riyadh, KSA

Mechanisms of Plant Defense Against Insect Herbivores
Phenolics
phenolic
glycosides
bound
phenolics
condensed
tannins
lignin
Terpenes
Monoterpenes
diterpene
acids
Secondary Metabolites
Defense-related
proteins
perxoidases
polyphenol oxidase
hydrolysable
tannins
N-containing
Alkaloids
Mustard oils
Physical
Defenses
Spines, thorns
cutins
waxes
suberins

• Plants evolve defense chemicals in response to attacks by
insects
• While insects counter-evolve detoxification systems.

Resistance Mechanisms In Insects
Survival insects employ a variety of resistance mechanisms, including:
• Detoxification, target site modifications, and nerve insensitivity.
• Detoxification occurs when toxins are modified by reduction, oxidation, and conjugation reactions,
resulting in the excretion of modified toxin molecules.
• The metabolic defense system in insects generally involves three main groups of enzymes acting in
three phases against a number of insecticides and phytochemicals.
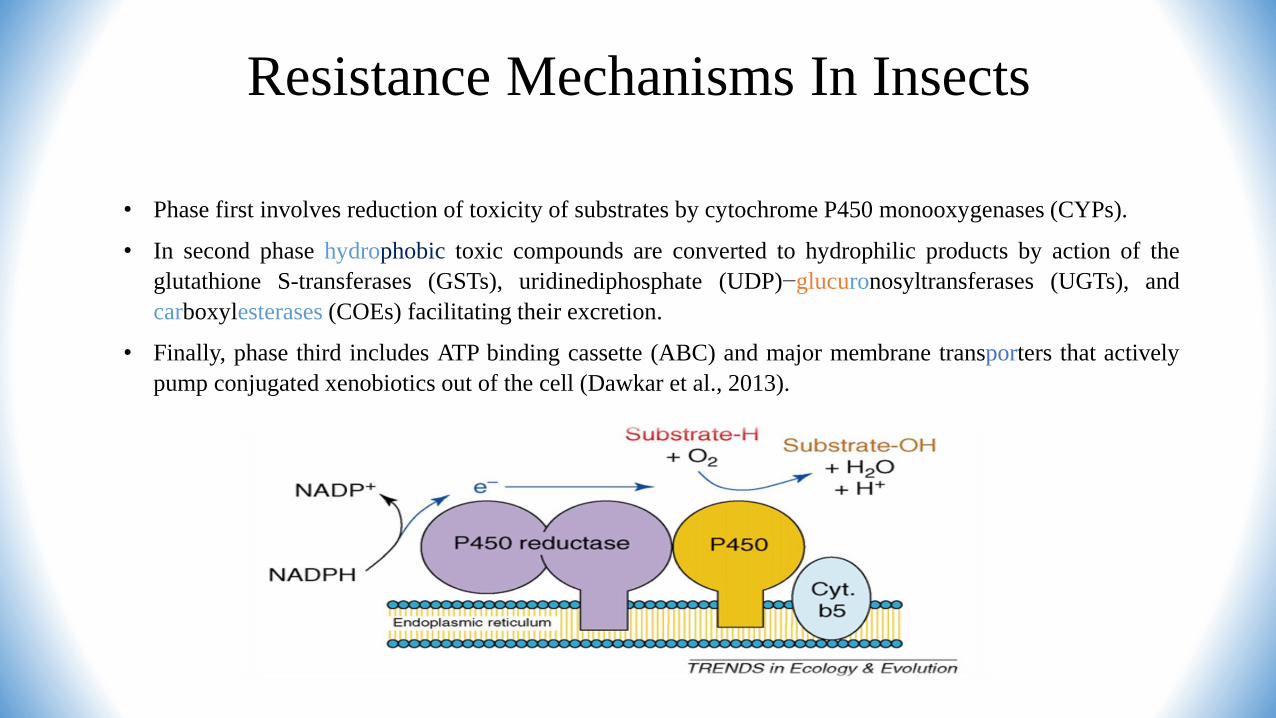
Resistance Mechanisms In Insects
• Phase first involves reduction of toxicity of substrates by cytochrome P450 monooxygenases (CYPs).
• In second phase hydrophobic toxic compounds are converted to hydrophilic products by action of the
glutathione S-transferases (GSTs), uridinediphosphate (UDP)−glucuronosyltransferases (UGTs), and
carboxylesterases (COEs) facilitating their excretion.
• Finally, phase third includes ATP binding cassette (ABC) and major membrane transporters that actively
pump conjugated xenobiotics out of the cell (Dawkar et al., 2013).

Insect adaptation strategies
• Behavioral adaptation
• Detoxification of plant chemical defense compounds
• Sequestration

Behavioral adaptation
Bursera squirts toxic
compounds (terpenes)
when the vein is cut
Blephara beetles cut the
veins before they start
feeding

Detoxification of Plant Chemical Defense Compounds
• General detoxification enzymes: P450's,
• Specific detoxification enzymes: example (Nitrile Specifier Protein) NSP.
What happens when a Pieris rapae larvae feeds on a brassicales plant?
By NSP to convert it to a non-toxic form.

Senecio jacobaea
contain
Pyrrolizidine
Alkaloids (PA's)
PA's sequestered
by larvae of
tigermoths
Tyria jacobaea
(Arctiidae)
• Sequestration
PA's transferred
to adults
- Males produce aphrodisiacs from PA's
- Males transfer PA's to females by mating
(nuptial gift)
- Females are better protected against predators
- Females transfer PA's to eggs for protection

Examples of adaptive:2
• Scientists at the Max Planck Institute for Chemical Ecology in Jena, Germany, now found the opposite
mechanism: a defensive compound of the wild tobacco species Nicotiana attenuata which is toxic with sugar
molecules bound to it and a glycosidase in the gut of the tobacco hornworm Manduca sexta which removes
one sugar from this toxin to convert it to a non-toxic form.
• This is the first time that the role of deglycosylation in detoxification as an insect counter-adaptation.
• Scientists showed that removing a sugar molecule from a plant’s defensive compound can also result in
detoxification. (Poreddy et al., 2015).
Lyc4 RGHGL
Lyc4

Another way to avoid contact with the toxin is by suppressing plant defences.
• The induction of plant defences by chewing insects is well documented, and usually involves salivary elicitors.
• Oral secretions of several caterpillar species have recently been shown to suppress or reduce host-plant
defences.
• The principal component of the tobacco earworm Helicoverpa zea (Lepidoptera) saliva is a glucose oxidase
that decreases the level of nicotine induced in leaves from tobacco Nicotiana tabacum (Musser, R.O. et al.,
2002).
Helicoverpa zea that decreases the level of nicotine induced in leaves from
tobacco Nicotiana tabacumglucose oxidase

• The only well documented example is from two unrelated lineages, the specialist monarch butterfly and
two leaf beetle (Chrysochus sp) species, which are all resistant to ouabain, a toxic cardenolide found in
their milkweed host plants.
• In this case, a single amino-acid substitution in the target site of ouabain (the Na+, K+-ATPase) is
responsible for resistance (Labeyrie & Dobler., 2004).
Cardenolides Nervous system (depressing activity);
Na+, K+-ATPase (specific inhibitor)Target-site mutation

• Danaus plexippus (Lepidoptera) Late-instar larvae deactivate the defence of milkweeds by cutting veins
before feeding.
Cardenolides Nervous system (depressing activity);
Na+, K+-ATPase (specific inhibitor)Canal trenching behaviour

• In many insects, Schistocerca americana (Orthoptera); Hypera brunneipennis
(Coleoptera) Zygaena sp. (Lepidoptera); Clossiana euphrosyne (Lepidoptera)
Heliconius sara (Lepidoptera) a large proportion of the accumulated toxic plant
compounds can be excreted, or lost with exuvia during the moult (Zagrobelny,
M. et al., 2004).
Cyanogenic glycosides Electron transport (inhibition
of mitochondrial cytochrome
oxidase)
Ingestion avoidance;sequestration anddetoxification

• In the aphid Myzus persicae, the overproduction of the glutathione S-transferases (GSTs)
is probably responsible for the adaptation of the insect to glucosinolates and
isothiocyanates contained in its Brassicaceae host plants (Francis, F. et al., 2005).
Glucosinolates Respiration (inhibition) Detoxification by GSTs

• Another interesting example is the response of the diamondback moth Plutella xylostella (Lepidoptera) to the
defence system of its crucifer host plants, the ‘mustard oil bomb’.
• This system relies on the co-secretion of glucosinolates and a specific enzyme (myrosinase), each stored in
separate cell compartments.
• Following tissue damage, glucosinolates are hydrolysed by myrosinase into highly toxic products, such as
isothiocyanate.
• The diamondback moth has developed an original adaptive strategy based on the modification of ingested plant
glucosinolates by a sulfatase gut enzyme, preventing their hydrolysis by plant myrosinase, thus disarming the
‘mustard oil bomb’ (Ratzka, A. et al.,2002).
Glucosinolates Respiration (inhibition) Detoxification by a glucosinolate sulfatase

Formation of nitriles
instead of isothiocyanate
• The cabbage white butterfly Pieris rapae (Lepidoptera) has developed a different adaptive mechanism by
redirecting toxic isothiocyanate formation toward non-toxic nitrile formation with a specific gut protein
(Wittstock, U. et al., 2004).
• Cyanide in P. rapae is detoxified by β-cyanoalanine synthase and rhodanese activity into nontoxic β-
cyanoalanine and rhodanide (Stauber et al., 2012).

• *Another example is the adaptation of cactophilic Drosophila species from the
Sonoran desert to the specific allelochemicals contained in their host plants, where
several unrelated P450-encoding genes are induced by different isoquinoline
alkaloids, suggesting the exploitation of evolutionarily distant P450 enzymes in the
adaptation of the various Drosophila species to their different host cacti (Fogleman,
J.C. 2000).
Detoxification by P450s

• When the polyphagous arctiid moth Estigmene acrea and Tyria jacobaeae (Lepidoptera) feeds on
species of Asteraceae, it not only sequesters pyrrolizidine alkaloids (PAs) contained in those plants,
but also detoxifies them by N-oxidation catalysed by a specific flavin-dependent monooxygenase
(Hartmann, T. et al., 2005).
Detoxification by N-oxidation
and sequestration

• Larvae of the cowpea bruchid Callosobruchus maculatus (Coleoptera) fed on a diet containing the soybean
cysteine protease inhibitor soyacystatin N (scN) activate an array of counterdefence genes to adapt to this
toxin.
• Using microarray technology, Monn et al. 2004, suggested that the overexpression of cathepsin-like cysteine
proteases has a key role in the adaptation of this insect to ingested scN by saturating the protease inhibitor
molecules.
Overexpression of insensitive proteaseProtease inhibitors Digestive system (inhibition of protease)

• The glucose oxidase enzyme in the saliva of the beet armyworm Spodoptera exigua decreases the
transcript level of key regulatory genes involved in the early steps of the defence pathways of the
Medicago truncatula plant (Bede, J.C. et al., 2006).
Terpenoids
Nervous system (inhibition of acetylcholine
esterase); feeding (deterrent owing to physical
barrier and bitterness); growth and development
inhibitor (pheromone analog)
Repression of genes involved
in biosynthetic pathways

Ostrinia furnacalis maize contain cyclic hydroxamic
acids (cHx) Feeding
UDP-glucosyltransferases (UGT)
activity in the midgut and excretion of
non-toxic glucosides
Enzyme
• A number of gramineous plants such as maize contain cyclic hydroxamic acids (cHx) that are toxic to many
herbivores such as Ostrinia furnacalis (Kojiima et al., 2010).
.

• Burnet moth larvae are able to sequester cyanogenic glucosides from their food plant as well as to carry
out de novo biosynthesis.
• Here we show that three genes (CYP405A2, CYP332A3 and UGT33A1) encode the entire biosynthetic
pathway of cyanogenic glucosides in the Burnet moth Zygaena filipendulae (Zagrobelny &Moller,. 2011).
Zygaena filipendulae Detoxification
Caterpillar defence droplets: poisoning the predators
by (β-cyanoalanine synthase) Sequestration

• The generalists cabbage moth Mamestra brassicae, cotton bollworm Helicoverpa armigera (both Lepidoptera,Noctuidae), fall armyworm, Egyptian armyworm and the cabbage looper, which feed on glucosinolate-containing plants, conjugate toxic isothiocyanates with glutathione by GST activity in their gut and subsequentlyexcrete non-toxic products with the frass (Schramm et al., 2012).
Mamestra brassicae and Helicoverpa armigera (GST) Glutathione S-transferase activity, probably in the gut and excrete non-toxic products with the frass

• Insect herbivores that are adapted to low to medium levels of defence compounds can regularly switch to other host
plants to avoid ingestion of lethal doses by dietary mixing a feeding behaviour mainly exhibited by generalists.
• Hostplant switching may dilute excessive detrimental effects of any single plant defence compound.
• For example, larvae of the lepidopteran generalists yellow woolly bear Spilosoma virginica and tiger moth Grammia
incorrupta (both Lepidoptera, Arctiidae) can feed on high iridoid-glucoside-containing plants like Plantago
lanceolata (Plantaginaceae) (Pankoke et al., 2012).
Spilosoma virginica, Grammia incorrupta Inducible reduction of endogenous insect BGD
(β-glucosidase;) activity in the midgut.Host-plant switching

Protection against predators & natural enemies• Insects to avoid plant poisons is by sequestering and deploying the poisons for their own pheromone
system and defense.
• Lepidoptera sequesters plant secondary metabolites such some terpenes, phenols and many nitrogen-
containing compounds and uses them as toxic or unpalatable to predators.
• Insects frequently detoxify or sequester plant defense compounds and, sometimes, they result in
protection against parasitoids and predators being used as toxic or unpalatable at defense.
• Sequestering specialists have developed the ability to incorporate these compounds with relative
impunity, ingesting, transporting and depositing the substances to be sequestered in parti- cular sites of
the larvae, adult body and even in the eggs.

An example of this adaptation is illustrated by the tobacco hornworm.
• This insect accumulates the nicotine synthesized by tobacco plants in its
own body which is toxic to most insects and uses it as a deterrent to
parasitoids.

• Monarch butterfly caterpillars have evolved the ability to store toxins known as cardenolides, obtained
from their milkweed diet, specifically to make themselves poisonous to birds, as has at least one other
species of milkweed-munching caterpillar.
• Sequestration of cardenolides in the body (as a defence against predators) is associated with the three
levels of Na+/K+-ATPase resistance.


Conclusions
• The co-evolution of plants and insects is very interesting.
• Plants have developed efficient mechanisms to protect them against
herbivory while insects have found diverse ways of avoiding negative effects
of their host plants defense mechanisms.
• Even though many workers have attempted to study plant-insect interaction,
our knowledge is still limited.
• The better understanding of this process will allow us to achieve more
effective methods for the biological control of insect pests with natural
products by the development of new plant varieties with enhanced chemical
defenses .

Reference
Haruta M, Major IT, Christopher ME, Patton JJ, Constabel CP (2001) A Kunitz trypsin inhibitor gene family from
trembling aspen (Populus tremuloides Michx.): cloning, functional expression, and induction by wounding and
herbivory. Plant Mol. Biol. 46:347-359.
Birkett MA, Campbell CAM, Chamberlain K, Guerrieri E, Hick AJ, Martin JL, Matthes M, Napier JA, Pettersson
J, Pickett JA, Poppy GM, Pow EM, Pye BJ, Smart LE, Wadhams GH, Wadhams LJ, Woodcock CM (2000)
New roles for cis-jasmone as an insect semiochemical and in plant defense. Proc. Natl Acad. Sci. USA
97:9329-9334.
Baldwin IT (2001) An ecologically motivated analysis of plant-herbivore interactions in native tobacco. Plant
Physiol. 127:1449-1458.
Fordyce JA, Agrawal AA (2001) The role of plant trichomes and caterpillar group size on growth and defense of
the pipevine swallowtail Battus philenor. J. Animal Ecol. 70:997-1005.
Scott JG, Wen ZM (2001) Cytochromes P450 of insects: the tip of the iceberg. Pest Manag. Sci. 57:958-967.
Silva CP, Terra WR, Grossi de Sل MF, Samuels RI, Isejima EM, Bifano TD, Almeida JS (2001) Induction of
digestive α-amylases in larvae of Zabrotes subfasciatus (Coleoptera: Bruchidae) in response to ingestion of
common bean α-amylase inhibitor 1. J. Insect Physiol. 47:1283-1290.
Nishida R (2002) Sequestration of defensive substances from plants by lepidoptera. Annu. Rev. Entomol. 47:57-
92.

Haruta M, Major IT, Christopher ME, Patton JJ, Constabel CP (2001) A Kunitz trypsin inhibitor gene
family from trembling aspen (Populus tremuloides Michx.): cloning, functional expression, and
induction by wounding and herbivory. Plant Mol. Biol. 46:347-359.
Birkett MA, Campbell CAM, Chamberlain K, Guerrieri E, Hick AJ, Martin JL, Matthes M, Napier
JA, Pettersson J, Pickett JA, Poppy GM, Pow EM, Pye BJ, Smart LE, Wadhams GH, Wadhams
LJ, Woodcock CM (2000) New roles for cis-jasmone as an insect semiochemical and in plant
defense. Proc. Natl Acad. Sci. USA 97:9329-9334.
Baldwin IT (2001) An ecologically motivated analysis of plant-herbivore interactions in native
tobacco. Plant Physiol. 127:1449-1458.
Fordyce JA, Agrawal AA (2001) The role of plant trichomes and caterpillar group size on growth and
defense of the pipevine swallowtail Battus philenor. J. Animal Ecol. 70:997-1005.
Scott JG, Wen ZM (2001) Cytochromes P450 of insects: the tip of the iceberg. Pest Manag. Sci.
57:958-967.
Silva CP, Terra WR, Grossi de Sل MF, Samuels RI, Isejima EM, Bifano TD, Almeida JS (2001)
Induction of digestive α-amylases in larvae of Zabrotes subfasciatus (Coleoptera: Bruchidae) in
response to ingestion of common bean α-amylase inhibitor 1. J. Insect Physiol. 47:1283-1290.
Nishida R (2002) Sequestration of defensive substances from plants by lepidoptera. Annu. Rev.
Entomol. 47:57-92.

Labeyrie, E., & Dobler, S. (2004). Molecular adaptation of Chrysochus leaf beetles to toxic compounds
in their food plants. Molecular Biology and Evolution, 21(2), 218–21. doi:10.1093/molbev/msg240.
Musser, R.O. et al. (2002) Herbivory: Caterpillar saliva beats plant defences - a new weapon emerges in
the evolutionary arms race between plants and herbivores. Nature 416, 599–600.
Helmus, M.R. and Dussourd, D.E. (2005) Glues or poisons: which triggers vein cutting by monarch
caterpillars? Chemoecology 15, 45–49.
Zagrobelny, M. et al. (2004) Cyanogenic glucosides and plant–insect interactions. Phytochemistry 65,
293–306.
Francis, F. et al. (2005) Glutathione S-transferases in the adaptation to plant secondary metabolites in
the Myzus persicae aphid. Arch. Insect Biochem. Physiol. 58, 166–174.
Ratzka, A. et al. (2002) Disarming the mustard oil bomb. Proc. Natl. Acad. Sci. U. S. A. 99, 11223–
11228.
Wittstock, U. et al. (2004) Successful herbivore attack due to metabolic diversion of a plant chemical
defense. Proc. Natl. Acad. Sci. U. S. A. 101, 4859–4864.

Hartmann, T. et al. (2005) Specific recognition, detoxification and metabolism of pyrrolizidine alkaloids by the
polyphagous arctiid Estigmene acrea. Insect Biochem. Mol. Biol. 35, 391–411.
Fogleman, J.C. (2000) Response of Drosophila melanogaster to selection for P450-mediated resistance to
isoquinoline alkaloids.Chem. Biol. Interact. 125, 93–105.
Naumann, C. et al. (2002) Evolutionary recruitment of a flavindependent monooxygenase for the detoxification of
host plantacquired pyrrolizidine alkaloids in the alkaloid-defended arctiid moth Tyria jacobaeae. Proc. Natl.
Acad. Sci. U. S. A. 99, 6085–6090.
Moon, J. et al. (2004) Transcriptional regulation in cowpea bruchid guts during adaptation to a plant defence
protease inhibitor. Insect Mol. Biol. 13, 283–291.
Bede, J.C. et al. (2006) Caterpillar herbivory and salivary enzymes decrease transcript levels of Medicago truncatula
genes encoding earlyenzymes in terpenoid biosynthesis. Plant Mol. Biol. 60, 519–531.
Kojima, W., Fujii, T., Suwa, M., Miyazawa, M. & Ishikawa, Y. (2010). Physiological adaptation of the Asian corn
borer Ostrinia furnacalis to chemical defenses of its host plant, maize. Journal of Insect Physiology 56, 1349–
1355.
Jensen, N. B., Zagrobelny, M., Hjernø, K., Olsen, C. E., Houghton-Larsen, J., Borch, J., Møller, B. L. & Bak, S.
(2011). Convergent evolution in biosynthesis of cyanogenic defence compounds in plants and insects. Nature
Communications 2, art. no. 273.

Stauber, E. J., Kuczka, P., van Ohlen,M., Vogt, B., Janowitz, T., Piotrowski, M., Beuerle, T. & Wittstock, U.
(2012). Turning the ‘mustard oil bomb’ into a ‘cyanide bomb’: aromatic glucosinolate metabolism in a
specialist insect herbivore. PLoS One 7, e35545.
Schramm, K., Vass˜ao, D. G., Reichelt, M., Gershenzon, J. & Wittstock, U. (2012). Metabolism of
glucosinolate-derived isothiocyanates to glutathione conjugates in generalist lepidopteran herbivores.
Insect Biochemistry and Molecular Biology 42, 174–182.
Pankoke, H., Bowers, M. D. & Dobler, S. (2012). The interplay between toxinreleasing β-glucosidase and
plant iridoid glycosides impairs larval development in a generalist caterpillar, Grammia incorrupta
(Arctiidae). Insect Biochemistry and Molecular Biology.
Wittstock, U., Agerbirk, N., Stauber, E. J., Olsen, C. E., Hippler, M., Mitchell-olds, T., … Vogel, H. (2004).
Successful herbivore attack due to metabolic diversion of a plant chemical defense.
de Bruxelles GL, Roberts MR (2001) Signals regulating multiple responses to wounding and herbivores. Crit.
Rev. Plant Sci. 20:487-521.
Wittstock, U., & Gershenzon, J. (2002). Constitutive plant toxins and their role in defense against herbivores
and pathogens, 1–8.
Petschenka G, & Agrawal AA (2015). Milkweed butterfly resistance to plant toxins is linked to sequestration,
not coping with a toxic diet. Proceedings. Biological sciences / The Royal Society, 282 (1818).































