
MARINE ECOLOGY PROGRESS SERIES
Mar Ecol Prog Ser
Vol. 429: 291–301, 2011
doi: 10.3354/meps09126Published May 16
INTRODUCTION
Pelagic seabirds search widely for food in marineecosystems and are temporally- and energetically- constrained to exploit resources within a restrictedarea around their nest sites while breeding (Orians &Pearson 1979, Young et al. 2009). Most pelagic seabirdspecies breed in colonies located on remote islands(Warham 1990), and intra- and inter-specific competi-
tion for food might promote trophic resource depletionand ultimately affect breeding success, survival orrecruitment (Furness & Birkhead 1984, Hunt et al.1986, Grémillet et al. 2004). Recent studies have shownthat trophic partitioning exists among sympatric sea-bird populations and is influenced by the distribution,quality and availability of prey, as well as breedingpopulation size (Grémillet et al. 2004, Forero et al.2005). Trophic partitioning can operate through habi-
© Inter-Research 2011 · www.int-res.com*Email: [email protected]
Exploiting the closest productive area: geographicalsegregation of foraging grounds in a critically
endangered seabird
M. Louzao1, 2,*, J. Navarro3, M. G. Forero4, J. M. Igual1, M. Genovart1,
K. A. Hobson5, D. Oro1
1Institut Mediterrani d’Estudis Avançats IMEDEA (CSIC-UIB), Miquel Marquès 21, 07190 Esporles, Mallorca, Spain.2Helmholtz Centre for Environmental Research-UFZ, Permoserstraße 15, 04318 Leipzig, Germany
3Institut de Ciències del Mar (CSIC), Passeig Marítim de la Barceloneta 37–49, 08003 Barcelona, Spain4Departamento de Biología de la Conservacíon, Estación Biológica de Doñana (CSIC), Apartado 1056, 41013 Sevilla, Spain
5Environment Canada, 11 Innovation Blvd., Saskatoon, Saskatchewan S7N 3H5, Canada
ABSTRACT: While breeding, seabirds are limited to exploiting resources within a restricted areaaround their breeding site and should exploit the closest productive marine areas within their distri-bution range. We investigated this hypothesis in one of the most endangered European seabirds, theBalearic shearwater Puffinus mauretanicus (ca. 3200 breeding pairs), restricted to the BalearicIslands. Our aims were (1) to assess whether isotopic evidence (i.e. stable isotopes of δ13C and δ15N)of foraging habitat partitioning occurs among northern, central and southern populations, (2) to geo-graphically locate population-specific potential foraging grounds along the Iberian continental shelf,and (3) to assess whether oceanographic conditions could explain observed patterns of stable iso-topes (SI). SI values showed a latitudinal gradient, with birds from the northern population havinglower δ15N and δ13C values than central and southern populations. Potential foraging grounds ofnorthern, central and southern populations were centred in Cape Creus, Ebro Delta and Cape LaNao, respectively, results which were supported by habitat models. Oceanographic conditions ineach potential foraging ground were different; the northern population used richer, colder anddeeper waters compared to the central and southern populations. Chlorophyll a was the mainoceanographic variable that explained variation in SI values. We hypothesised that SI differencesamong Balearic shearwater populations might be a consequence of differences in baseline isotopicvalues among potential foraging grounds rather than real differences in diet. Our comprehensivestudy also provides important information for management strategies to conserve this criticallyendangered shearwater.
KEY WORDS: Balearic shearwater · Geographical foraging ground segregation · Habitat modelling ·Mediterranean Sea · Oceanography · Puffinus mauretanicus · Stable isotopes
Resale or republication not permitted without written consent of the publisher

Mar Ecol Prog Ser 429: 291–301, 2011
tat segregation or selection of different prey within thesame habitat, with individual factors such as sex andage promoting spatial segregation of foraging grounds(Forero et al. 2005).
Unravelling spatial distribution differences withinand among species not only helps to understand theecological links between the components of marineecosystems; this also helps to identify conservationgaps relative to human threats (Boersma 2008, Phillipset al. 2009). For example, Young et al. (2009) showedhow Laysan albatrosses Phoebastria immutabilis
breeding in 2 distant breeding populations in theNorth Pacific showed differences in patterns of plasticingestion relative to geographical segregation of forag-ing grounds. However, studying the at-sea distributionand foraging ecology of marine top predators is diffi-cult because they forage over vast areas, and logisticalconstraints limit the spatial and temporal coverage ofsampling programs. Recent ecological tools such asanalysis of stable isotope (SI) analysis (SIA), habitatmodelling and ocean remote sensing (Inger & Bearhop2008, Louzao et al. 2009) provide new opportunities toinvestigate the foraging ecology of marine predators.Levels of SIs of nitrogen (δ15N) and carbon (δ13C) in tis-sues are good indicators of the trophic level and forag-ing habitats, respectively, of marine predators (Forero& Hobson 2003). Moreover, the isotopic variabilityfound in marine species can be explained byspecific oceanographic conditions due to habitatpreferences (c.f. Phillips et al. 2009) since pro-ductive areas related with terrestrial inputs (e.g.river run-offs) might present different isotopicsignatures compared to marine-driven produc-tivity areas (e.g. upwelling). Similarly, habitatuse models help explain species– habitat rela-tionships through robust mathematical descrip-tions and can indicate the oceanographic fea-tures primarily associated with seabirds (Louzaoet al. 2006a). Furthermore, habitat models canbe used to predict the most likely pelagic habitats used in both time and space (Guisan& Zimmermann 2000).
In the present study, we focused on one ofthe most endangered European seabirds, theBalearic shearwater Puffinus mauretanicus,which breeds only on the Balearic Islands in thewestern Mediterranean Sea. Due to its small(ca. 3200 breeding pairs) and declining popula-tion (7.4% decrease per year; Oro et al. 2004),this pelagic species is currently listed as Criti-cally Endangered on the IUCN Red List (BirdLifeInternational 2008, Arcos 2011). Balearic shear-waters breed in scattered colonies, ranging fromca. 5 to 300 breeding pairs (Ruiz & Martí 2004).During breeding, the foraging range of the spe-
cies comprises the productive Iberian continental shelfand is influenced by oceanographic processes drivingthe distribution and availability of their natural prey(Louzao et al. 2006a, Bellido et al. 2008).
Our main aim was to assist conservation efforts byelucidating spatial connections between breeding andforaging grounds. Specifically, our objectives were(1) to assess whether habitat partitioning occurs alongthe breeding range (northern, central and southernpopulations) based on SIA (δ13C, δ15N), (2) to locatepopulation-specific potential foraging grounds alongthe Iberian continental shelf, and (3) to assess whetheroceanographic conditions could explain observed pat-terns of SIs. We expected that Balearic shearwatersmake foraging trips between the less productivewaters around their breeding colonies and the highlyproductive waters on the Iberian Mediterranean conti-nental shelf (Louzao et al. 2006a, Louzao et al. 2009).However, marine resources are patchily distributedalong this continental shelf system and different meso-scale oceanographic features result in productivityhotspots in the Gulf of Lions, Cape Creus, offshoreBarcelona, Ebro Delta and Cape La Nao (Fig. 1; Louzaoet al. 2009). Within this context, we predicted thatnorthern, central and southern populations would eachexploit the closest productive marine areas within theIberian continental shelf. We also expected different SI
292
Fig. 1. Puffinus mauretanicus. Study area showing the locations (blackcircles) of the 4 breeding colonies studied (1 northern, 2 central popu-lations and 1 southern), the presence probability based on the habitatmodel (Louzao et al. 2006a), and the limits of the potential foraginggrounds (PFGs) of northern (solid lines), central (dashed lines) andsouthern (dotted lines) populations. Depth is represented by the iso-baths of 200 m at the limit of the continental shelf (thin solid line) and
2000 m (thin dotted line)

Louzao et al.: Geographical segregation of foraging Balearic shearwaters
signatures of individuals feeding in different areas,mainly related to water mass characteristics influenc-ing baseline isotopic values (Barnes et al. 2009). Theworkflow of the present comprehensive ecologicalstudy is described in Fig. S1 Supplement 1 at www. int-res. com/ articles/ suppl/ m429 p291 _ supp. pdf. Finally, wediscussed the implications of our results for the con -servation of this species.
MATERIALS AND METHODS
Sampling procedures. During the breeding period of2003, we sampled blood from several adults at 4 breed-ing populations covering the worldwide breedingrange of the Balearic shearwater (Fig. 1, Table 1): onenorthern population (Mola de Maó on Menorca, MOL),2 central populations (Sa Cella and Conills on Mal-lorca, CEL and CON respectively, ca. 18 km apart) andone southern population (Conillera in Eivissa, EIV).The linear distance between northern and central pop-ulations was larger (ca. 200 km) than the distance be -tween central and southern populations (ca. 120 km).We sampled a total of 29 males and 21 females once atthe end of the incubation period (Table 1). For eachindividual, we took 0.5 ml of blood from the brachialvein. The blood was stored in 70% ethanol and wasused for both molecular sexing and SIA of δ15N andδ13C. The extraction of lipids was considered unneces-sary for blood samples because the lipid componentin blood is generally very low (Deuel 1955). For amedium-sized bird such as the Balearic shearwater,
the isotope values of the whole blood integrate thedietary information of the previous 3 to 4 wk (Hobson& Clark 1992). Thus, SI values of adults integrated thediet information over the incubation period.
To test for differences in SI values among breedingpopulations, we used General Linear Models (GLMs)including SIs as response variables, sex and breedingpopulations as explanatory variables, and their inter-actions within the R environment (R DevelopmentCore Team 2009). Assuming that both isotopes fol-lowed a normal distribution, models were fitted with aGaussian error distribution (identity link) and wereranked based on their Akaike Information Criteria val-ues corrected for small sample sizes (AICc). The modelwith the lowest AICc is considered the best compro-mise between model deviance and model complexity.When models are within 4 points of AICc, they are con-sidered statistically equivalent (Williams et al. 2001).In this case, we chose the most parsimonious modelhaving the lowest number of parameters.
Identifying population-specific po tential foraging
grounds. To determine whether any isotopic evidenceof foraging habitat partitioning was present, wefirst identified population-specific potential foraginggrounds over the Iberian continental shelf based on atheoretical search radius. Second, potential foraginggrounds were validated by means of predictive habitatmodels and those potential foraging grounds werecharacterised oceanographically. Finally, we applied aresampling procedure to link oceano graphic conditionsto SI values.
Potential foraging grounds: We established a poten-tial search radius of 250 km, given the dispersal capac-ities of other Puffinus species. For instance, the maxi-mum flying speed of the closely related Manxshear water Puffinus puffinus is 50 km h–1 (Guilford etal. 2008), which means that Balearic shearwaters wouldonly need 5 h to travel 250 km from their nests. More-over, the maximum foraging range of other Puffinus
species does not exceed 300 km (Powell 2009). The coreof each potential foraging ground was established bythe minimum distance between each breeding popula-tion (MOL, CEL and EIV) and the Iberian shelf edge(200 m depth) within a potential maximum foragingrange of 250 km. Only the geographical location of CELwas considered as representative for central popula-tions, given the short distance between this breedingsite and CON. The latitude at which the distance be-tween each breeding population and the shelf edge exceeded 250 km defined the limits of each potentialforaging ground.
Oceanographic habitat models: We overlapped thelimits of the potential foraging grounds and predictionsof the habitat model of Balearic shearwaters (Louzaoet al. 2006a) to assess whether potential foraging
293
n δ15N δ13C
MOL 15 9.33 ± 0.19 –19.31 ± 0.04
Males 9 9.19 ± 0.31 –19.37 ± 0.05
Females 6 9.55 ± 0.35 –19.19 ± 0.06
CEL 21 9.91 ± 0.12 –18.78 ± 0.10
Males 12 9.76 ± 0.16 –18.86 ± 0.11
Females 9 10.12 ± 0.19 –18.68 ± 0.17
CON 8 9.98 ± 0.16 –19.35 ± 0.12
Males 3 10.37 ± 0.31 –19.01 ± 0.08
Females 5 9.74 ± 0.12 –19.55 ± 0.11
EIV 6 10.47 ± 0.33 –18.67 ± 0.28
Males 5 10.61 ± 0.37 –18.54 ± 0.31
Females 1 9.81 –19.34
Table 1. Puffinus mauretanicus. Summary of δ15N and δ13Cvalues (mean ± SD) in blood of adult Balearic shearwatersduring the incubation period of 2003. MOL: northern popula-tion from Mola de Maó; CEL: central population from SaCella; CON: central population from Conills; EIV: southern
population from Conillera

Mar Ecol Prog Ser 429: 291–301, 2011294
grounds were highly suitable habitats for each breed-ing population. The oceanographic habitat of theBalearic shear water was characterised during thechick-rearing period (May to June) in a previous studyalong the Mediterranean coast of the Iberian Peninsula(details in Louzao et al. 2006a). Observations ofBalearic shearwaters were made during 1999, 2000,and 2002 using standardised strip-transect techniqueswith a 300 m strip-width transect band and snapshotcounts of flying birds. All Balearic shearwaters ob -served within the survey transect were recorded. Weused a hierarchical modelling approach to identifythose environmental variables that most accurately re -flected the oceanographic habitat of this species by de -lineating its foraging range using presence/absencedata. The foraging range comprised the frontal sys-tems along the eastern Iberian continental shelf waters(depth > 200 m) and areas close to the breeding colo -nies in the Balearic Islands. Finally, predictive habitatmodels were used to estimate shearwater occurrenceprobability in the entire western Mediterranean basin(Fig. 1). Predictive models matched the foraging rangedescribed by the observed data and also identified thewaters close to the breeding colonies in the BalearicIslands as potentially suitable habitat for the Balearicshearwater. We assumed that Balearic shearwatersvisit the same foraging grounds as Cory’s shearwatersCalonectris dio medea (Louzao et al. 2009) during incu-bation and chick-rearing stages.
Oceanographic characterisation of potential forag-
ing grounds: To characterise the foraging grounds, theoceanographic variables that were most relevant froma biological point of view were extracted for the west-ern Mediterranean from online databases providedby the Environmental Research Division, SouthwestFisheries Science Center and US National MarineFisheries Service (http:// coastwatch. pfel. noaa. gov/ coast watch/ CWBrowserWW180.jsp). Dynamic oceano-graphic variables such as sea surface temperature(SST,°C; proxy for water mass distribution) and chloro-phyll a concentration (chl a, mg m–3; surrogate ofmarine productivity) were extracted from SEAWIFSand PATHFINDER data, respectively. Dynamic vari-ables were obtained for March 2003 to match the SIdata for the incubation period that year. Static vari-ables such as bathymetry (m; proxy for coastal versuspelagic domains) were extracted once. All variableswere ag gregated to match the standard grid of 0.1° cellsize. Additionally, we estimated their spatial gradientsby estimating their proportional change (PC) within asurrounding 3 × 3 cell (0.1° × 0.1°) grid using a movingwindow as follows: PC = [(maximum value – minimumvalue) × 100] / (maximum value). This dimensionlessmetric expresses the magnitude of change in eachhabitat variable, scaled to the maximum value (e.g.
Louzao et al. 2009). The spatial gradients of chl a andSST indicate the presence of frontal systems, whereasthe gradient of bathymetry reflects the presence oftopographic features (e.g. shelf break or seamount).Finally, the 6 environmental variables were extractedover the Iberian continental shelf corresponding to thelatitudinal limits of each potential foraging ground. Wetested whether the oceanographic conditions of thepotential foraging grounds were similar based on thenon-parametric Kruskal-Wallis test.
Links between SI values and oceano graphy: Welinked oceanographic variables to the SI values of 15,21 and 6 shearwaters sampled from MOL, CEL andEIV, respectively. Only CEL was included as represen-tative of central populations given the relatively shortdistance between CEL and CON. First, we randomlyselected 25 values of the 6 oceanographic variables(SST, chl a, bathymetry and their spatial gradients) ofthe corresponding potential foraging ground (see Sup-plement 2 at www. int-res.com/ articles/ suppl/ m429p291 _ supp. pdf). This value (25) was selected as theminimum sample size necessary to reproduce median values of oceanographic variables in each potentialforaging ground (see Supplement 2). Then, we esti-mated the median value of each oceanographic vari-able. This first step was repeated, as several SI valueswere available for each breeding population (e.g. 15shearwaters were sampled from the northern popula-tion, and each shearwater was represented by 2 iso-tope values, δ15N and δ13C, totalling 30 values). Sec-ond, we linked the median values of oceanographicvariables to values of both isotopes of the correspond-ing population. This procedure was applied to each ofthe northern, central and southern populations.
We assessed the influence of oceanographic vari-ables on the values of δ15N and δ13C of each breedingpopulation based on GLMs. Oceanographic variableswere normalised, and strongly ‘correlated’ (| rs | > 0.5)predictors were identified by means of pair-wiseSpearman rank correlation coefficients for avoidingcolinearity and related problems with parameter esti-mations (Zuur et al. 2007). When pairs of predictorvariables were strongly correlated, we discarded oneof the redundant variables by ranking models withonly one variable (fitted with a Gaussian error dis -tribution and identity link) by their AICc value. Toassess the effect of oceanographic variables and popu-lation on SIs, we used GLMs including SI values asresponse variables and oceanographic variables asexplanatory variables. We also included the quadraticterm of oceanographic variables to account for po -tential non-linear relationships between SIs and ex -planatory variables, typically found in nature (Quinn& Keough 2002). Models were ranked based on theirAICc values.
Louzao et al.: Geographical segregation of foraging Balearic shearwaters
RESULTS
Isotopic evidence of foraging habitat partitioning
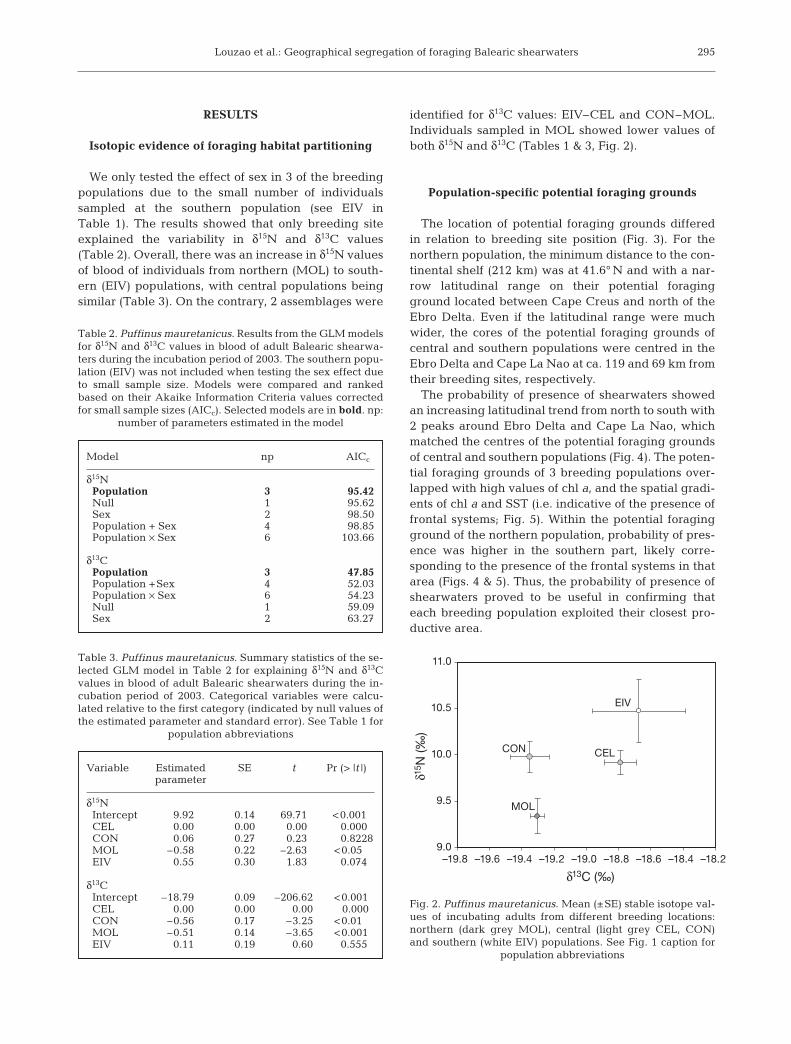
We only tested the effect of sex in 3 of the breedingpopulations due to the small number of individualssampled at the southern population (see EIV inTable 1). The results showed that only breeding siteex plained the variability in δ15N and δ13C values(Table 2). Overall, there was an increase in δ15N valuesof blood of individuals from northern (MOL) to south-ern (EIV) populations, with central populations beingsimilar (Table 3). On the contrary, 2 assemblages were
identified for δ13C values: EIV–CEL and CON–MOL.Individuals sampled in MOL showed lower values ofboth δ15N and δ13C (Tables 1 & 3, Fig. 2).
Population-specific potential foraging grounds
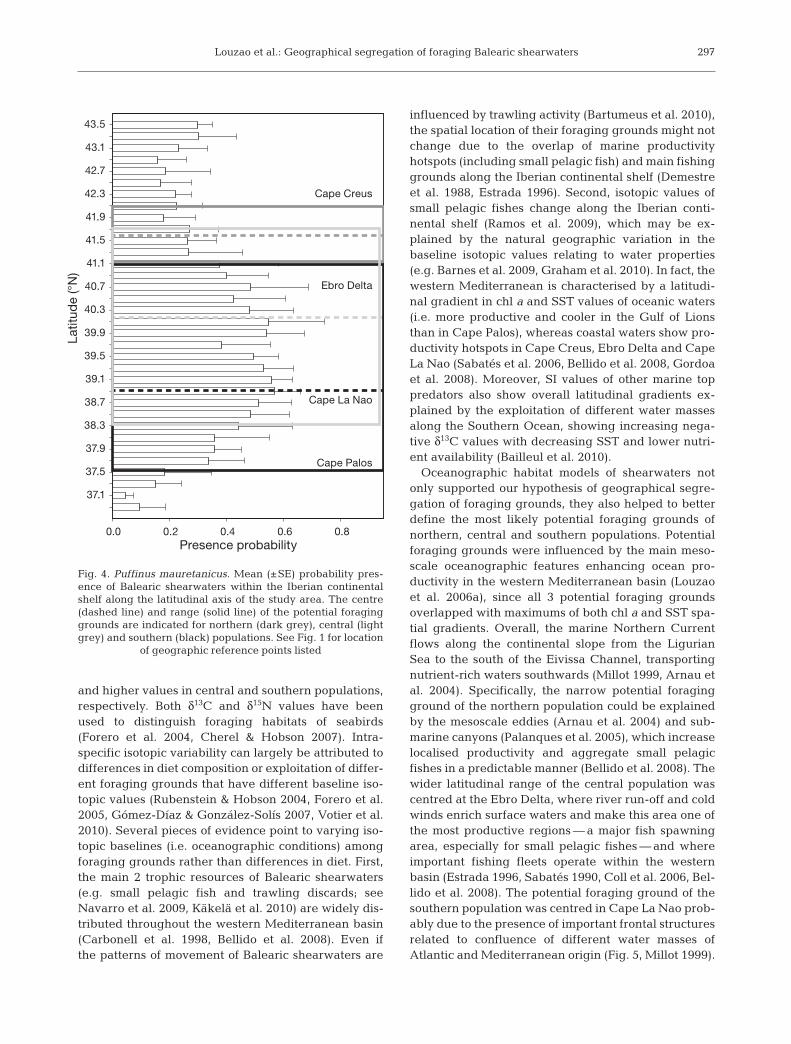
The location of potential foraging grounds differedin relation to breeding site position (Fig. 3). For thenorthern population, the minimum distance to the con-tinental shelf (212 km) was at 41.6° N and with a nar-row latitudinal range on their potential foragingground located between Cape Creus and north of theEbro Delta. Even if the latitudinal range were muchwider, the cores of the potential foraging grounds ofcentral and southern populations were centred in theEbro Delta and Cape La Nao at ca. 119 and 69 km fromtheir breeding sites, respectively.
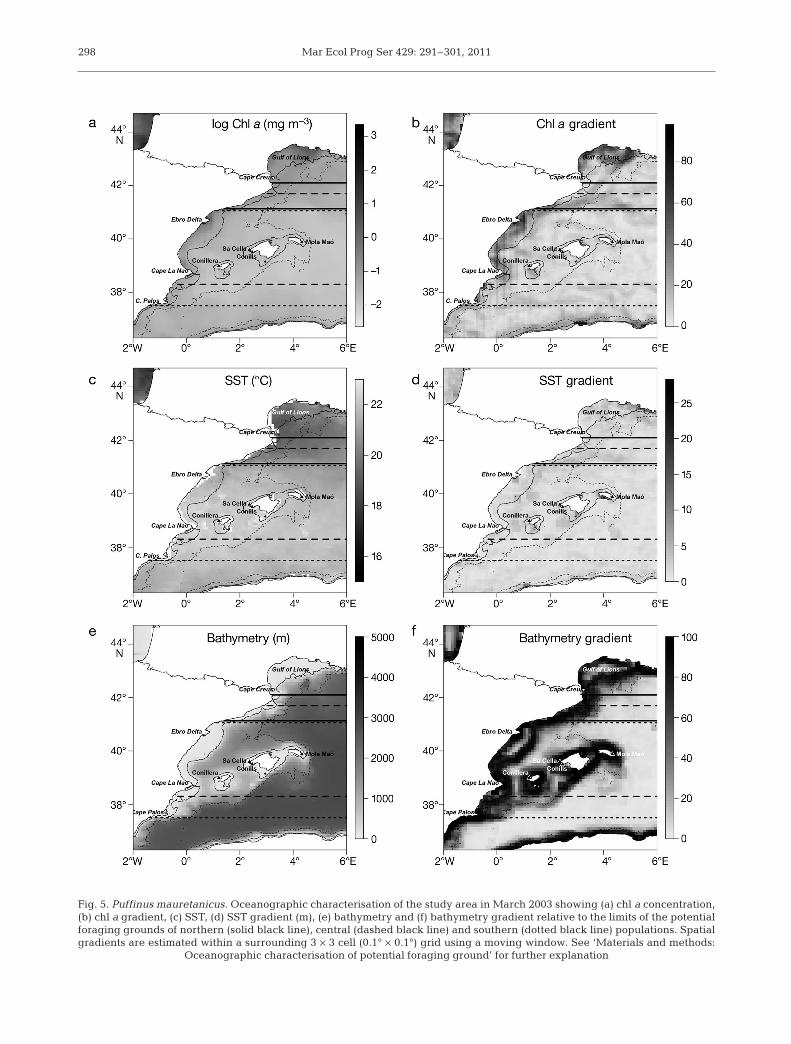
The probability of presence of shearwaters showedan increasing latitudinal trend from north to south with2 peaks around Ebro Delta and Cape La Nao, whichmatched the centres of the potential foraging groundsof central and southern populations (Fig. 4). The poten-tial foraging grounds of 3 breeding populations over-lapped with high values of chl a, and the spatial gradi-ents of chl a and SST (i.e. indicative of the presence offrontal systems; Fig. 5). Within the potential foragingground of the northern population, probability of pres-ence was higher in the southern part, likely corre-sponding to the presence of the frontal systems in thatarea (Figs. 4 & 5). Thus, the probability of presence ofshearwaters proved to be useful in confirming thateach breeding population exploited their closest pro-ductive area.
295
Model np AICc
δ15NPopulation 3 95.42Null 1 95.62Sex 2 98.50Population + Sex 4 98.85Population × Sex 6 103.66
δ13CPopulation 3 47.85Population +Sex 4 52.03Population × Sex 6 54.23Null 1 59.09Sex 2 63.27
Table 2. Puffinus mauretanicus. Results from the GLM modelsfor δ15N and δ13C values in blood of adult Balearic shearwa-ters during the incubation period of 2003. The southern popu-lation (EIV) was not included when testing the sex effect dueto small sample size. Models were compared and rankedbased on their Akaike Information Criteria values correctedfor small sample sizes (AICc). Selected models are in bold. np:
number of parameters estimated in the model
Variable Estimated SE t Pr (> |t |)parameter
δ15NIntercept 9.92 0.14 69.71 <0.001CEL 0.00 0.00 0.00 0.000CON 0.06 0.27 0.23 0.8228MOL –0.58 0.22 –2.63 <0.05EIV 0.55 0.30 1.83 0.074
δ13CIntercept –18.790 0.09 –206.62 <0.001CEL 0.00 0.00 0.00 0.000CON –0.56 0.17 –3.25 <0.01MOL –0.51 0.14 –3.65 <0.001EIV 0.11 0.19 0.60 0.555
Table 3. Puffinus mauretanicus. Summary statistics of the se-lected GLM model in Table 2 for explaining δ15N and δ13Cvalues in blood of adult Balearic shearwaters during the in-cubation period of 2003. Categorical variables were calcu-lated relative to the first category (indicated by null values ofthe estimated parameter and standard error). See Table 1 for
population abbreviations
–19.8 –19.6 –19.4 –19.2 –19.0 –18.8 –18.6 –18.4 –18.2
δ15N
(‰
)
9.0
9.5
10.0
10.5
11.0
CELCON
MOL
EIV
δ13C (‰)
Fig. 2. Puffinus mauretanicus. Mean (±SE) stable isotope val-ues of incubating adults from different breeding locations:northern (dark grey MOL), central (light grey CEL, CON)and southern (white EIV) populations. See Fig. 1 caption for
population abbreviations

Mar Ecol Prog Ser 429: 291–301, 2011296
Oceanographically, each breeding population ex -ploited distinct potential foraging grounds, highlightedby chl a (Kruskal-Wallis test, H2, 669 = 86.29, p < 0.001),SST (H2, 669 = 134.63, p < 0.001) and its gradient (H2, 669
= 18.23, p < 0.001), and bathymetry (H2, 669 = 27.21, p <0.001) and its spatial gradient (H2, 669 = 12.82, p < 0.01;Fig. 6). However, the chl a gradient did not differamong potential foraging grounds. Oceanographicconditions of each potential foraging ground differedshowing a latitudinal gradient; the northern popula-tion exploited richer (high chl a concentration), colderand deeper waters compared to central and southernpopulations.
When linking SIs with oceanographic variables, wedid not consider the chl a gradient since it did not differamong potential foraging grounds. The uncorrelatedvariable for explaining both SIs was only chl a and wedeveloped all possible combinations of models. Wefound that only the linear positive effect of chl a was im-
portant in explaining the observed variation in δ15N andδ13C values (models with lowest AICc value; Table 4).Individuals of northern, central and southern popula-tions exploited potential foraging grounds of higher, intermediate and lower chl a values, respectively(Table 5, Fig. 6 and Fig. S3 in Supplement 3 at www. int-res. com/ articles/ suppl/ m429 p291 _ supp. pdf).
DISCUSSION
By comparing SI values, habitat models and oceano -graphy, we provided important information on the for-aging ecology of the Balearic shearwater. SI values ofshearwaters varied with latitude in relation to the loca-tion of the breeding site, which suggested that eachpopulation exploited geographically separated forag-ing grounds; birds from the northern population hadlower δ15N and δ13C values, followed by intermediate
0 200 400 600
Latitu
de (ºN
)
36.937.137.337.537.737.938.138.338.538.738.939.139.339.539.739.940.140.340.540.740.941.141.341.541.741.942.142.342.542.742.943.1
Distance to continental shelf (km)
0 200 400 600 0 200 400 600
Northern Central Southern
41.6°N
40.2°N
38.9°N
42.1°N
41.1°N
41.7°N
38.3°N
41.1°N
37.5°NCape Palos
Cape La Nao
Ebro Delta
Cape Creus
Cape Palos
Cape La Nao
Ebro Delta
Cape Creus
Cape Palos
Cape La Nao
Ebro Delta
Cape Creus
Fig. 3. Puffinus mauretanicus. Mean (±SE) distance between northern, central and southern populations and the limit of the con-tinental shelf at a depth of 200 m along the latitudinal axis of the study area. Only CEL was included as representative of centralpopulations given the relatively short distance between CEL and CON. The centre of the potential foraging ground was identi-fied by the minimum distance (grey dashed line) and its range by the limit of the search radius of 250 km (black dashed lines).
See Fig. 1 for locations of geographic reference points listed
Louzao et al.: Geographical segregation of foraging Balearic shearwaters
and higher values in central and southern populations,respectively. Both δ13C and δ15N values have beenused to distinguish foraging habitats of seabirds(Forero et al. 2004, Cherel & Hobson 2007). Intra-specific isotopic variability can largely be attributed todifferences in diet composition or ex ploitation of differ-ent foraging grounds that have different baseline iso-topic values (Rubenstein & Hobson 2004, Forero et al.2005, Gómez-Díaz & González-Solís 2007, Votier et al.2010). Several pieces of evidence point to varying iso-topic baselines (i.e. oceano graphic conditions) amongforaging grounds rather than differences in diet. First,the main 2 trophic resources of Balearic shearwaters(e.g. small pelagic fish and trawling discards; seeNavarro et al. 2009, Käkelä et al. 2010) are widely dis-tributed throughout the western Mediterranean basin(Carbonell et al. 1998, Bellido et al. 2008). Even ifthe patterns of movement of Balearic shearwaters are
influenced by trawling activity (Bartumeus et al. 2010),the spatial location of their foraging grounds might notchange due to the overlap of marine productivityhotspots (including small pelagic fish) and main fishinggrounds along the Iberian continental shelf (Demestreet al. 1988, Estrada 1996). Second, isotopic values ofsmall pelagic fishes change along the Iberian conti-nental shelf (Ramos et al. 2009), which may be ex -plained by the natural geographic variation in thebaseline isotopic values relating to water properties(e.g. Barnes et al. 2009, Graham et al. 2010). In fact, thewestern Mediterranean is characterised by a latitudi-nal gradient in chl a and SST values of oceanic waters(i.e. more productive and cooler in the Gulf of Lionsthan in Cape Palos), whereas coastal waters show pro-ductivity hotspots in Cape Creus, Ebro Delta and CapeLa Nao (Sabatés et al. 2006, Bellido et al. 2008, Gordoaet al. 2008). Moreover, SI values of other marine toppredators also show overall latitudinal gradients ex -plained by the exploitation of different water massesalong the Southern Ocean, showing increasing nega-tive δ13C values with decreasing SST and lower nutri-ent availability (Bailleul et al. 2010).
Oceanographic habitat models of shearwaters notonly supported our hypothesis of geographical segre-gation of foraging grounds, they also helped to betterdefine the most likely potential foraging grounds ofnorthern, central and southern populations. Potentialforaging grounds were influenced by the main meso-scale oceanographic features enhancing ocean pro-ductivity in the western Mediterranean basin (Louzaoet al. 2006a), since all 3 potential foraging groundsoverlapped with maximums of both chl a and SST spa-tial gradients. Overall, the marine Northern Currentflows along the continental slope from the LigurianSea to the south of the Eivissa Channel, transporting nutrient- rich waters southwards (Millot 1999, Arnau etal. 2004). Specifically, the narrow potential foragingground of the northern population could be explainedby the mesoscale eddies (Arnau et al. 2004) and sub-marine canyons (Palanques et al. 2005), which increaselocalised productivity and aggregate small pelagicfishes in a predictable manner (Bellido et al. 2008). Thewider latitudinal range of the central population wascentred at the Ebro Delta, where river run-off and coldwinds enrich surface waters and make this area one ofthe most productive regions — a major fish spawningarea, especially for small pelagic fishes — and whereimportant fishing fleets operate within the westernbasin (Estrada 1996, Sabatés 1990, Coll et al. 2006, Bel-lido et al. 2008). The potential foraging ground of thesouthern population was centred in Cape La Nao prob-ably due to the presence of important frontal structuresrelated to confluence of different water masses ofAtlantic and Mediterranean origin (Fig. 5, Millot 1999).
297
Presence probability0.0 0.2 0.4 0.6 0.8
Latitu
de (°N
)
36.937.137.337.537.737.938.138.338.538.738.939.139.339.539.739.940.140.340.540.740.941.141.341.541.741.942.142.342.542.742.943.143.343.5
Cape Palos
Cape La Nao
Ebro Delta
Cape Creus
Fig. 4. Puffinus mauretanicus. Mean (±SE) probability pres-ence of Balearic shearwaters within the Iberian continentalshelf along the latitudinal axis of the study area. The centre(dashed line) and range (solid line) of the potential foraginggrounds are indicated for northern (dark grey), central (lightgrey) and southern (black) populations. See Fig. 1 for location
of geographic reference points listed
Mar Ecol Prog Ser 429: 291–301, 2011298
Fig. 5. Puffinus mauretanicus. Oceanographic characterisation of the study area in March 2003 showing (a) chl a concentration,(b) chl a gradient, (c) SST, (d) SST gradient (m), (e) bathymetry and (f) bathymetry gradient relative to the limits of the potentialforaging grounds of northern (solid black line), central (dashed black line) and southern (dotted black line) populations. Spatialgradients are estimated within a surrounding 3 × 3 cell (0.1° × 0.1°) grid using a moving window. See ‘Materials and methods:
Oceanographic characterisation of potential foraging ground’ for further explanation

Louzao et al.: Geographical segregation of foraging Balearic shearwaters
Breeding Balearic shearwaters likelyuse the same foraging grounds year af-ter year as the western Mediterraneanis characterised by a relatively stablecoastal circulation, and overall chl a
values may not differ greatly betweenyears (Gordoa et al. 2008). In fact, thelocation of resources at large and inter-mediate scales appears reasonablypredictable for pelagic seabirds overlong time periods in the Mediterraneanbecause they associate to the samemesoscale features along the Iberiancontinental shelf during different years(Arcos & Oro 2002, Abelló et al. 2003,Louzao et al. 2006a, Louzao et al. 2009).The use of tracking devices may linkforaging trip characteristics to fitnessparameters such as breeding perfor-mance. Individuals that have to travellonger distances may experience lowerreproductive success (Boersma & Reb-stock 2009).
Our comprehensive study also pro-vided important information for man-agement strategies to assist the conser-vation of this shearwater; we estimatedfor the first time the spatial location ofthe potential foraging grounds ofnorthern, central and southern popula-tions along the Iberian continentalshelf. Given the delicate status of thispopulation, marine zoning measuresmay greatly benefit the conservation ofthe Balearic shearwater by extendingprotective measures beyond thebreeding sites. In fact, potential forag-ing grounds, such as those centred in
Cape Creus, Ebro Delta and Cape La Nao, could con-stitute a network of Marine Protected Areas (MPAs),and they have been recently identified as marineImportant Bird Areas (IBAs) for the conservation of thewestern Mediterranean seabird community (Arcos etal. 2009). Within the potential foraging grounds, othermore diffuse protective measures could be imple-mented, such as a compulsory fishery observer pro-gram to record potential bycatch in bottom longlinefisheries and the study of potential impacts of windfarms over the continental shelf region. Evidenceexists that Balearic shearwaters are incidentallycaught in bottom longline, (Arcos et al. 2008, ICES2008, Louzao et al. in press), and trawling activitycould influence this probability (Laneri et al. 2010).Moreover, reducing fisheries bycatch in bottom long-lines is still the key factor in the conservation of the
299
Fig. 6. Puffinus mauretanicus. Oceanographic characterisation of the potentialforaging grounds of northern, central and southern populations over the Iberiancontinental shelf indicated by box plots of median (thick black horizontal lines),25–75% inter quartile range (horizontal limits of boxes), and non-outlier range(horizontal limits of dashed lines): (a) chl a concentration, (b) chl a gradient,
(c) SST, (d) SST gradient, (e) bathymetry, and (f) bathymetry gradient
Model np AICc
δ15NCHL 2 97.58CHL + CHL2 3 100.28Null 1 102.04
δ13CCHL 2 54.21CHL + CHL2 3 57.79Null 1 65.14
Table 4. Puffinus mauretanicus. Modelling δ15N and δ13C val-ues in blood of Balearic shearwaters to randomly assignedoceanographic variables based on GLMs. Models were com-pared and ranked based on their Akaike Information Criteriavalues corrected for small sample sizes (AICc). Selected modelsare in bold. CHL: chl a; np: number of parameters estimated in
the model

species (Oro et al. 2004, Louzao et al. 2006b), in addi-tion to removing threats related to introduced pre-dators in some accessible colonies. Unfortunately, notmuch effort has been directed at obtaining detailedlocal information on seabird mortality at national orEuropean levels and, in turn, the spatio-temporal win-dow of the interaction between bottom longlines andBalearic shearwaters is not well known. Thus, weemphasize the urgent need to assess the interaction ofshearwaters with longline fishery within their foraginggrounds during breeding in the western Mediter-ranean, and also recommend assessment during thepost-breeding period along the Mediterranean andNorth Atlantic.
Acknowledgements. We are especially grateful to I. Afán, J.Calvo, E. Cardona, R. Escandell, R. Fernández-Graña, O.Martínez, M. McMinn, A. Rodríguez, C. Santana, and R. Triayfor their help during the fieldwork. V. Rodríguez and M.Bauzà helped with molecular sex identification. J.M. Arcosand G. Bortolotti provided invaluable comments to first ver-sions of this manuscript. Permits were provided by Conselle-ria de Medi Ambient (Govern de les Illes Balears), CabreraNational Park, the Military Government of Menorca, ParcNatural de Cala d’Hort and Parc Natural de Ses Salines. Thisstudy was funded by Govern de les Illes Balears, DISCBIRD(QLRT-2000-00839) and LIFE projects (European Commis-sion) and the Spanish Ministries of Education and Science(ref. BOS2003-01960 and REN2002-00450) and of the Envi-ronment (ref. 024A/2002). M.L. was supported by fellowshipsfrom Govern de les Illes Balears and Institut Menorquí d’Es-tudis, as well as by a postdoctoral contract of the Spanish Min-istry of Education and Science (ref. EX-2007-1148) and MarieCurie Individual Fellowship (PIEF-GA-2008-220063). J.N. wassupported by a researcher contract from of the Spanish Min-istry of Science and Innovation (Juan de la Cierva program).M.G.F. and M.G. by Ramon y Cajal and I3P contracts, respec-tively, of the Spanish Ministry of Education and Science.Experiments comply with the current laws of the country inwhich the experiments were performed. We acknowledge theeditor and 4 anonymous referees for their help with bringingthe manuscript to its final state.
LITERATURE CITED
Abelló P, Arcos JM, Gil de Sola L (2003) Geographical patternsof seabird attendance to a research trawler along the IberianMediterranean coast. Sci Mar 67:69–75
Arcos JM (compiler) (2011) International species action planfor the Balearic shearwater, Puffinus mauretanicus. SEO/BirdLife & BirdLife International. Available at http:// ec.europa. eu/ environment/ nature/ conservation/ wildbirds/action_ plans/ per_ species_en.htm
Arcos JM, Oro D (2002) Significance of fisheries discards for athreatened Mediterranean seabird, the Balearic shear waterPuffinus mauretanicus. Mar Ecol Prog Ser 239: 209–220
Arcos JM, Louzao M, Oro D (2008) Fisheries ecosystem impactsand management in the Mediterranean: seabirds point ofview. In: Nielsen JL, Dodson JJ, Friedland K, Hamon TR,Musick J, Verspoor E (eds) Reconciling fisheries with con-servation. Proc 4th World Fish Cong. American FisheriesSociety, Bethesda, MD, p 1471–1479
Arcos JM, Bécares J, Rodrígez B, Ruiz A (2009) Áreas impor-tantes para la conservación de las aves marinas en España.LIFE04NAT/ES/000049-Sociedad Española de Ornitología(SEO/Bird-Life), Madrid
Arnau P, Liquete C, Canals M (2004) River mouth plume eventsand their dispersal in the northwestern Mediterranean Sea.Oceanography 17:22–31
Bailleul F, Authier M, Ducatez S, Roquet F, Charrassin JB, CherelY, Guinet C (2010) Looking at the unseen: combining animalbio-logging and stable isotopes to reveal a shift in the ecolog-ical niche of a deep diving predator. Ecography 33:709–719
Barnes C, Jennings S, Barry JT (2009) Environmental correlatesof large-scale spatial variation in the δ13C of marine animals.Est Coast Shelf Sci 81:368–374
Bartumeus F, Giuggioli L, Louzao M, Bretagnolle V, Oro D,Levin S (2010) Fishery discards impact on seabird move-ment patterns at regional scales. Curr Biol 20:215–222
Bellido JM, Brown AM, Valavanis VD, Giráldez A, Pierce GJ,Iglesias M, Palialexis A (2008) Identifying essential fishhabitat for small pelagic species in Spanish Mediterraneanwaters. Hydrobiologia 612:171–184
BirdLife International (2008) Species factsheet: Puffinus
mauretanicus. Available at: www.birdlife. org/ datazone/ speciesfactsheet. php?id=30026
Boersma PD (2008) Penguins as marine sentinels. Bioscience 58:597–607
Boersma PD, Rebstock GA (2009) Foraging distance affectsreproductive success in Magellanic penguins. Mar EcolProg Ser 375:263–275
Carbonell A, Martín P, de Ranieri S & WEDIS team 1 (1998) Discards of the western Mediterranean trawl fleets. RappComm Int Mer Médit 35:392–393
Cherel Y, Hobson KA (2007) Geographical variation in carbonstable isotope signatures of marine predators: a tool toinvestigate their foraging areas in the Southern Ocean. MarEcol Prog Ser 329:281–287
Coll M, Palomera I, Tudela S, Sarda F (2006) Trophic flows, eco-system structure and fishing impacts in the South CatalanSea, Northwestern Mediterranean. J Mar Syst 59: 63–96
Demestre M, Lleonart J, Martín P, Recasens L, Sánchez P (1988)La pesca en Cataluña. FAO rapport sur les pêches 395:101–103
Deuel HJ (1955) Biochemistry, digestion, absorption, transport,and storage. The lipids, Vol 2. Interscience, New York, NY
Estrada M (1996) Primary production in the northwestern Medi-terranean. Sci Mar 60:55–64
Forero MG, Hobson KA (2003) Using stable isotopes of nitrogenand carbon to study seabird ecology: applications in theMediterranean seabird community. Sci Mar 67:23–32
Mar Ecol Prog Ser 429: 291–301, 2011300
Variable Estimated SE t Pr (> |t |)parameter
δ15NIntercept 9.79 0.11 90.55 <0.001CHL –0.33 0.11 –3.03 <0.001
δ13CIntercept –18.96 0.06 –293.79 <0.001CHL –0.270 0.06 –4.16 <0.001
Table 5. Puffinus mauretanicus. Summary statistics of theselected GLM model in Table 4 for explaining δ15N and δ13Cvalues in blood of adult Balearic shearwaters to randomly
assigned oceanographic variables. CHL: chl aä
ä
ä
ä
ää
ä
ä
ä
ä
ä

Louzao et al.: Geographical segregation of foraging Balearic shearwaters
Forero MG, Bortolotti GR, Hobson KA, Donázar JA, BertellottiM, Blanco G (2004) High trophic overlap within the sea-bird community of Argentinean Patagonia: a multiscaleapproach. J Anim Ecol 73:789–801
Forero MG, González-Solís J, Hobson KA, Donázar JA, Bertel-lotti M, Blanco G, Bortolotti GR (2005) Stable isotopes revealtrophic segregation by sex and age in the southern giantpetrel in two different food webs. Mar Ecol Prog Ser 296:107–113
Furness RW, Birkhead TR (1984) Seabird colony distributionssuggest competition for food supplies during the breedingseason. Nature 311:655–665
Gómez-Díaz E, González-Solís J (2007) Geographic assignmentof seabirds to their origin: combining morphologic, genetic,and biogeochemical analyses. Ecol Appl 17: 1484–1498
Gordoa A, Illas X, Cruzado A, Velásques Z (2008) Spatio-tem-poral patterns in the north-western Mediterranean fromMERIS derived chlorophyll a concentration. Sci Mar 72:757–767
Graham BS, Koch PL, Newsome SD, McMahon KW, Aurioles D(2010) Using isoscapes to trace the movements and foragingbehavior of top predators in oceanic ecosystems. In: West JB,Bowen GJ, Dawson TE, Tu KP (eds) Isoscapes: understandingmovement, pattern and process on earth through iso tope mapping. Springer-Verlag, New York, NY, p 299–318
Grémillet D, Dell’Omo G, Ryan PG, Peters G, Ropert-Coudert Y,Weeks SJ (2004) Offshore diplomacy, or how seabirds miti-gate intra-specific competition: a case study based on GPStracking of Cape gannets from neighbouring colonies. MarEcol Prog Ser 268:265–279
Guilford TC, Meade J, Freeman R, Biro D and others (2008)GPS tracking of the foraging movements of Manx Shearwa-ters Puffinus puffinus breeding on Skomer Island, Wales.Ibis 150:462–473
Guisan A, Zimmermann NE (2000) Predictive habitat distribu-tion models in ecology. Ecol Model 135:147–186
Hobson KA, Clark RG (1992) Assessing avian diets using stableisotopes II: factors influencing diet-tissue fractionation. Condor 94:189–197
Hunt GL, Eppley ZA, Schneider DC (1986) Reproductive per-formance of seabirds: the importance of population andcolony size. Auk 103:306–317
ICES (International Council for the Exploration of the Sea)(2008) Report of the Working Group on Seabird Ecology(WGSE). 10–14 March 2008, Lisbon, Portugal. ICES CM2008/LRC:05. ICES, Copenhagen. Available at www. ices.dk/ reports/ LRC/ 2008/WGSE/WGSE2008.pdf
Inger R, Bearhop S (2008) Applications of stable isotope analy-ses to avian ecology. Ibis 150:447–461
Käkelä R, Käkelä A, Martínez-Abraín A, Sarzo B and others(2010) Fatty acid signature analysis confirms foragingresources of a globally endangered Mediterranean seabirdspecies: calibration test and application to the wild. MarEcol Prog Ser 398:245–258
Laneri K, Louzao M, Martínez-Abraín A, Arcos JM and others(2010) Trawling regime influences longline seabird bycatchin the Mediterranean: new insights from a small-scale fish-ery. Mar Ecol Prog Ser 420:241–252
Louzao M, Hyrenbach KD, Arcos JM, Abelló P, Gil de Sola L,Oro D (2006a) Oceanographic habitat of an endangeredMediterranean Procellariiform: implications for marine pro-tected areas. Ecol Appl 16:1683–1695
Louzao M, Igual JM, McMinn M, Aguilar JS, Triay R, Oro D(2006b) Small pelagic fish, trawling discards and breedingperformance of the critically endangered Balearic shearwa-ter: improving conservation diagnosis. Mar Ecol Prog Ser318:247–254
Louzao M, Bécares J, Rodríguez B, Hyrenbach KD, Ruiz A,
Arcos JM (2009) Combining vessel-based surveys andtracking data to identify key marine areas for seabirds. MarEcol Prog Ser 391:183–197
Louzao M, Arcos JM, Laneri K, Martínez-Abraín A and others(in press) Evidencias de la captura incidental de pardelabalear en el mar. Actas del VI Congreso del GIAM
Millot C (1999) Circulation in the Western Mediterranean Sea.J Mar Syst 20:423–442
Navarro J, Louzao M, Igual JM, Oro D and others (2009) Sea-sonal changes in the diet of a critically endangered seabirdand the importance of trawling discards. Mar Biol 156:2571–2578
Orians GH, Pearson NE (1979) On the theory of central placeforaging theory. In: Horn DJ, Mitchell RM, Stairs GR (eds)Analysis of ecological systems. Ohio State University Press,Columbus, OH
Oro D, Aguilar JS, Igual JM, Louzao M (2004) Modellingdemography and extinction risk in the endangered Balearicshearwater. Biol Conserv 116:93–102
Palanques A, García-Ladona E, Gomis D, Martín J and others(2005) General patterns of circulation, sediment fluxes andecology of the Palamós (La Fonera) submarine canyon,northwestern Mediterranean. Prog Oceanogr 66:89–119
Phillips RA, Bearhop S, Mcgill RAR, Dawson DA (2009) Stableisotopes reveal individual variation in migration strategiesand habitat preferences in a suite of seabirds during thenonbreeding period. Oecologia 160:795–806
Powell CDL (2009) Foraging movements and the migration tra-jectory of flesh-footed shearwaters Puffinus carneipes fromthe south coast of western Australia. Mar Ornithol 37: 115–120
Quinn GP, Keough MJ (2002) Experimental design and dataanalysis for biologists. Cambridge University Press, Cam-bridge
R Development Core Team (2009) R: a language and environ-ment for statistical computing. R Foundation for StatisticalComputing, Vienna
Ramos R, Ramírez F, Sanpera C, Jover L, Ruiz X (2009) Feedingecology of yellow-legged gulls Larus michahellis in thewestern Mediterranean: a comparative assessment usingconventional and isotopic methods. Mar Ecol Prog Ser377:289–297
Rubenstein DR, Hobson KA (2004) From birds to butterflies:animal movement patterns and stable isotopes. Trends EcolEvol 19:256–263
Ruiz A, Martí R (eds) (2004) La pardela balear. SEO/BirdLife-Conselleria de Medi Ambient del Govern de les IllesBalears, Madrid
Sabatés A (1990) Distribution pattern of larval fish populationsin the Northwestern Mediterranean. Mar Ecol Prog Ser59:75–82
Sabatés A, Martín P, Lloret J, Raya V (2006) Sea warming andfish distribution: the case of the small pelagic fish, Sardi-
nella aurita, in the western Mediterranean. Glob ChangeBiol 12:2209–2219
Votier SC, Bearhop S, Witt MJ, Inger R, Thompson D, Newton J(2010) Individual responses of seabirds to commercial fish-eries revealed using GPS tracking, stable isotopes and ves-sel monitoring systems. J Appl Ecol 47:487–497
Warham J (1990) The petrels: their ecology and breeding sys-tems. London Academic Press, London
Williams BK, Nichols JD, Conroy MJ (2002) Analysis and man-agement of animal populations. Academic Press, San Diego
Young LC, Vanderlip C, Duffy DC, Afanasyev V, Shaffer SA(2009) Bringing home the trash: Do colony-based differ-ences in foraging distribution lead to increased plasticingestion in Laysan albatrosses? PLoS ONE 4:e7623
Zuur AF, Ieno EN, Smith GM (2007) Analysing ecological data.Springer, New York, NY
301
Editorial responsibility: Matthias Seaman,
Oldendorf/Luhe, Germany
Submitted: June 30, 2010; Accepted: March 14, 2011
Proofs received from author(s): April 15, 2011
ä
ä
ä
ää
ä
ä
ä
ä
ä
ä
ä
ä
ä
ää
ä
ä
ä
ä
ä
ä
ää
ä
ää
ä
ä
ä
ää

1
The following supplements accompany the article
Exploiting the closest productive area: geographical segregation of foraging
grounds in a critically endangered seabird
M. Louzao1,2,*
, J. Navarro3, M. G. Forero
4, J. M. Igual
1, M. Genovart
1, K. A. Hobson
5, D. Oro
1
1Institut Mediterrani d’Estudis Avançats IMEDEA (CSIC-UIB), Miquel Marquès 21, 07190 Esporles, Mallorca, Spain.
2Helmholtz Centre for Environmental Research-UFZ, Permoserstraße 15, 04318 Leipzig, Germany
3Institut de Ciències del Mar (ICM, CSIC), Passeig Marítim de la Barceloneta 37–49, 08003 Barcelona, Spain
4Department of Conservation Biology, Estación Biológica de Doñana (CSIC), Apartado 1056, 41013 Sevilla, Spain
5Environment Canada, 11 Innovation Blvd., Saskatoon, Saskatchewan S7N 3H5, Canada
*Email: [email protected]
Marine Ecology Progress Series: 429: 291–301 (2011)
Supplement 1.
Fig. S1. Puffinus mauretanicus. Workflow of the comprehensive ecological study applied to identify population-specific potential
foraging grounds (PFGs).
Recommended
















![[2022] SGHC 55 - GROUNDS OF DECISION](https://img.dokumen.tips/doc/110x75/6327a8f9e491bcb36c0b6428/2022-sghc-55-grounds-of-decision.jpg)
