
Uppkomst och spridning avstreptomycinresistens hos bakterier
Sofia Nordin
Independent Project in BiologySjälvständigt arbete i biologi, 15 hp, höstterminen 2017Institutionen för biologisk grundutbildning, Uppsala universitet

1
Uppkomst och spridning av streptomycinresistens hos bakterier Sofia Nordin Självständigt arbete i biologi 2017 Sammandrag Antibiotika är ett samlingsnamn för ämnen som används för att hämma eller döda olika typer av bakterier. Streptomycin är en typ av antibiotika som upptäcktes under tidigt 1940-tal av Selman Waksman. Den klassificeras som en aminoglykosid, vilket är en grupp av antibiotika som påverkar bakteriers proteinsyntes. Detta gör den genom att ingå i specifika interaktioner med två delar av ribosomens mindre subenhet, 30S. Dessa specifika delar är proteinet S12 och 16S rRNA. Det har tidigare visats att mutationer av komponenternas motsvarande gener, rpsL respektive 16S rRNA gener, kan inge resistens hos bakterien. Även gener som kodar för proteiner och enzymer som interagerar med 30S subenheten kan ge resistens. Exempel på dessa gener är gidB, aaDa, strA-strB och whiB7. Alla dessa gener kodar för enzymer eller proteiner som på olika sätt hindrar antibiotikans funktioner. Spridningen av streptomycinresistens mellan bakterier kan ske via fyra olika mekanismer; arv från en bakterie till dess avkomma, konjugation, transduktion och transformation. Hos bakterier ärvs alltid gener direkt till avkomman eftersom de inte har någon sexuell förökning. Konjugation är mekanism som innebär att två bakterier kopplas samman via en kanal som kallas pilus och DNA förs mellan cellerna. Transduktion betyder att virus som kallas bakteriofager infekterar en bakterie men råkar i processen ta upp bakteriellt DNA och föra det vidare till andra celler. Transformation innebär att bakterier själva tar upp DNA från omgivningen. Detta kan de göra om de är i ett tillstånd som kallas kompetens, vilket kan induceras av exempelvis stressförhållanden. Alla dessa mekanismer har experimentellt visats vara inblandade i spridningen av streptomycinresistenta bakterier, då någon av de gener som ger resistens kan överföras. Streptomycinresistens kan alltså uppkomma hos en bakterie relativt lätt, via spontana mutationer eller överföring av DNA mellan celler. På grund av detta har antibiotikaresistens blivit ett allt större problem inom sjukvården, eftersom resistenta bakterier lätt sprids. Den snabba spridningen av antibiotikaresistens via de olika mekanismerna har blivit ett globalt problem där sjukvården står handfallna för infektioner och sjukdomar orsakade av resistenta bakterier. Arbetet för att minska spridningen av antibiotikaresistens pågår världen över men för att verkligen lösa problemet kan nya former av läkemedel behövas. Forskare har upptäckt nya metoder, exempelvis CRISPR-Cas9, som eventuellt i framtiden kan minska resistensproblemen. Inledning Antibiotika är ett samlingsnamn för olika typer av ämnen som antingen hämmar eller eliminerar bakteriers tillväxt. Detta sker ofta genom interaktioner mellan läkemedlet och bakterien som stör processer viktiga för cellens överlevnad (Davies & Davies 2010). Antibiotika delas vanligtvis in i olika klasser beroende på hur dess mekanism påverkar bakterien. Några exempel är ß-laktamer som skadar cellväggen, rifampicin som stör RNA-syntesen och aminoglykosider som skadar proteinsyntesen eller cellväggen (van Hoek et al. 2011). De flesta antibiotikum som idag finns på marknaden produceras naturligt av bakterier, svampar och andra mikroorganismer. Den naturliga funktionen är att hämma andra mikroorganismers tillväxt och därmed konkurrera ut dem (Clardy et al. 2009).

2
Historien bakom antibiotika och streptomycin Den första medicinska antibiotikan, penicillin, upptäcktes av Alexander Flemming år 1928 när den isolerades från mögelsvampen Penicillum (Aminov 2010). Antibiotikan blev snabbt ett populärt läkemedel och användes bland annat av soldater under andra världskriget (Ventola 2015) Det var dock inte förrän 1945 som penicillin började produceras och säljas för allmänt läkemedelsbruk (Aminov 2010). Trots att penicillin var en effektiv medicin för att bota infektioner påträffades pencillinresistenta bakterier redan under 1950-talet, vilket ledde till en läkemedelskris (Ventola 2015). Som en följd började flera nya antibiotikum att upptäckas och de kommande årtiondena hade flera nya klasser av antibiotika kommit ut på marknaden, däribland streptomycin. Denna antibiotika upptäcktes egentligen redan under tidigt 1940-tal, när den isolerades från den jordlevande bakterien Streptomyces griseus (S. griseus) av forskaren Selman Waksman (van Hoek et al. 2011). S. griseus tillhör klassen aktinobakterier, vilka har varit av stor kommersiell vikt då de under sin reproduktiva fas producerar antibiotika (Chater 2006). Streptomycin är ett läkemedel av stor betydelse då det var den första antibiotika som användes mot bakterien Mycobacterium tuberculosis, som orsakar sjukdomen tuberkulos hos människor (Ruiz et al. 2002). Streptomycins verkningsmekanismer Streptomycin tillhör antibiotikafamiljen aminoglykosider, vilket betyder att dess verkningsmekanism påverkar bakteriers translation, det vill säga proteinsyntesen (van Hoek et al. 2011). Translationen sker i ribosomen och är en noggrann process där tre olika typer av RNA är inblandade. Den första heter budbärar-RNA, eller mRNA, vilket är en enkelsträngad kopia av DNA som för vidare den genetiska informationen till proteinsyntesen. Den andra kallas för transfer-RNA, eller tRNA, som under proteinsyntesen bär de aminosyror som ska bilda proteinet. Den sista RNA-molekylen kallas ribosom-RNA, eller rRNA, som utgör en del av själva ribosomen (Campbell et al. 2014). I proteinsyntesen avkodas mRNA och aminosyror används för att bygga upp ett funktionellt protein. Varje aminosyra kodas av tre stycken kvävebaser som tillsammans bildar ett kodon. Kodon från mRNA:t läses och matchas med motsvarande antikodon som bärs av en tRNA molekyl. TRNA bär också på de aminosyror som inuti ribosomen kopplas ihop till ett fullständigt protein. Ribosomen består av en större och en mindre subenhet som i bakterier kallas för 50S respektive 30S (Cooper 2000). De kallas detta på grund av deras sedimentationshastighet i sockerlösning. S:et i deras namn står för enheten Svedberg som reflekterar subenheternas storlek, massa och densitet (Klug et al. 2014). Den mindre subenheten, 30S, är viktig för streptomycins verkningsmekanism, eftersom det är med denna subenhet som antibiotikan interagerar (Carter et al. 2000). 30S subenheten består av 16S rRNA och ett flertal proteiner, däribland ett protein som kallas S12. (Cooper 2000). Translationsprocessen i en bakterie och de viktiga molekylerna inblandade kan ses i Figur 1. I translationen har 30S-subenheten två primära funktioner för att proteinsyntesen ska vara funktionell och specifik. Det första är att den är involverad i initiering av translationen där 16S rRNA binder till mRNA:s startkodon. Det andra är att den är med och koordinerar rörelsen av mRNA och tRNA genom ribosomen. Antibiotikan streptomycin verkar genom att binda till 16S rRNA via starka vätebindingar och skapa vissa interaktioner med en aminosyra i proteinet S12 (Carter et al. 2000). Streptomycins interaktioner leder till strukturella förändringar av 30S-subenheten, vilket leder till att avkodningen rubbas. Som följd blir ribosomen mer benägen till att göra fel och proteinerna som bildas blir därför felaktiga eller ofullständiga. En sådan allvarlig störning av proteinsyntesen leder till celldöd då bakterien inte kan fungera normalt (Sharma et al. 2007). Bakterier delas upp i två olika typer av bakterier beroende på hur cellmembranet ser ut. Dessa två typer kallas för gram-positiva respektive gram-negativa bakterier. Skillnaden är att gram-positiva bakterier har ett cellmembran täckt av ett tjockt lager peptidoglykan medan gram-

3
negativa har ett inre och ett yttre membrans separerade av ett tunt lager peptidoglykan (Brown et al. 2015). Streptomycin räknas som en antibiotika med brett spektrum. Detta innebär att den är verksam mot både gram-positiva och gram-negativa bakterier samt att den kan användas mot flera olika familjer av bakterier (NCBI, Pubchem).
Varför har antibiotikaresistens uppkommit? Trots att streptomycin har varit en väldigt effektiv antibiotika, har en resistensutveckling skett hos flera arter av bakterier (Barker 1999). Detta beror till stor del på bakteriers flexibilitet när det kommer till att hantera sin omgivning. En hög mutationsfrekvens i kombination med deras snabba generationstid har gett dem en stark evolutionär fördel för att kunna hantera skiftande förutsättningar (Denamur & Matic 2006). Detta är en av orsakerna till varför resistens mot inte bara streptomycin, utan flera antibiotikum, har skett så pass snabbt de senaste årtiondena. Antibiotika ökar selektionstrycket hos bakterien eftersom det eliminerar antibiotikakänsliga konkurrenter och endast de resistenta bakterierna överlever för att reproducera sig (Ventola 2015). Antibiotika har länge varit sjukvårdens mest effektiva sätt att behandla olika infektioner och läkemedlen har räddat miljontals liv världen över (Chater 2006). Antibiotikaresistenta bakterier skapar emellertid problem inom sjukvården då inte alla sjukdomar och infektioner längre kan behandlas med de medel som de tidigare gjorts. Att antibiotikaresistens har blivit ett globalt problem beror på flera faktorer, bland annat överanvändning, felaktig preskribering och användandet av antibiotika i boskapsdjur. Redan år 1945 varnade upptäckaren av penicillin, Alexander Flemming, för konsekvenserna av överanvändning av antibiotika. Detta hörsammades inte och idag är det fortfarande många länder som säljer antibiotika utan preskribering, vilket ökar risken för överdriven användning (Ventola 2015). Felaktig preskribering har också bidragit till antibiotikaresistens då val av antibiotika, dos eller tidsomfattning för behandling är inkorrekt i upp till 50% av alla receptutskrivningar i USA (CDC 2013). Antibiotika används inte bara på människor, utan också på boskapsdjur för att hålla dem friska så att de kan producera kött i större kvantiteter och av bättre kvalité. Detta leder till att antibiotikaresistenta bakterier kan spridas till människor vid konsumtion av köttet, vilket kan skapa svårbehandlade infektioner hos konsumenter. Användandet av antibiotika på boskapsdjur påverkar även mikroorganismer i
Figur 1: Hur translationen sker i den bakteriella ribosomen. Ribosomen består av en 50S och en 30S subenhet. 30S subenheten innehåller de komponenter som streptomycin interagerar med, 16S rRNA och protein S12. MRNA strängen avkodas i ribosomen genom att tRNA bär motsvarande antikodon till ribosomen. TRNA för även med sig den aminosyra som kodonet kodar för. Aminosyrorna sätts sedan ihop till ett fullständigt protein. Figuren är omritad efter Campbell et al. 2014.

4
naturen eftersom upp till 90% av den använda antibiotikan utsöndras i djurens urin. Dessutom används vissa antibiotikum, i synnerhet streptomycin, som bekämpningsmedel på fruktträd i USA. Detta leder till att streptomycin släpps ut i naturen och kan orsaka resistensutveckling hos jordlevande bakterier (Ventola 2015). Eftersom antibiotikaresistens är ett av den medicinska världens största problem, vill jag med min uppsats utreda mekanismerna bakom resistens samt hur resistensen kan spridas mellan bakterier. Jag har valt att fokusera på streptomycin, då det är en äldre antibiotika som har varit ett vanligt och väl använt läkemedel. Streptomycinresistens uppkommer via specifika gener Ett vanligt sätt för bakterier att bli resistenta är genom spontana mutationer av gener som är inblandade i antibiotikans verkningsmekanism, eftersom interaktionerna ofta är väldigt specifika (Springer et al. 2006). Det har sedan tidigare varit känt att resistens mot streptomycin hos många bakterier är associerat med mutationer i genen som kodar för protein S12. Dessa mutationer ger oftast hög resistens mot streptomycin, vilket innebär att bakterierna tål höga koncentrationer av antibiotikan. Genen som kodar för S12 kallas för rpsL, vilket står för ”ribosomal subunit protein S12” (Keseler et al. 2017). Det har visats att det räcker med en enda mutation i denna gen för att bakterien ska bli resistent och mutationen ofta är av typen missens (Honoré & Cole 1994). Missens är en typ av mutation där nukleotid i DNA-strängen blir utbytt. Detta leder till att fel aminosyra kodas under translationen och proteinet kan som följd få en annan form. När detta händer blir S12 inte längre specifikt för streptomycin och antibiotikan kan inte längre interagera med proteinet (Klug et al. 2014). Eftersom streptomycin även interagerar med 16S rRNA via vätebindningar kan mutationer av gener som kodar för denna molekyl också ge streptomycinresistens. Experiment har visat att ändringar av specifika kvävebaser i 16S rRNA till följd av punktmutationer kan göra streptomycin oförmögen till att binda till 30S subenheten (Springer et al. 2006). En annan gen som kan ge hög streptomycinresistens kallas för aadA. Denna gen sitter oftast inte på bakteriens egna kromosom utan återfinns vanligtvis på så kallade plasmider (Hollingshead & Vapnek 1985). En plasmid är cirkulärt extrakromosomalt DNA som kan replikera oberoende av kromosomen (Klug et al. 2014). Genen aadA kodar för ett enzym som kallas streptomycin adenyltransferas. Enzymet inhiberar antibiotikan genom att modifiera streptomycin och därmed påverka dess förmåga interagera med ribosomens 30S subenhet. Genen aadA finns på flera olika typer av plasmider, även på de som redan innehåller gener för resistens (Hollingshead & Vapnek 1985). Flera arter av bakterier har påträffats med denna gen, exempelvis E. coli, Klebsiella pneumonia och Agrobacterium tumefaciens. Även i bakterien Salmonella choseraesuis fanns enzymet, men skiljde sig markant från enzymet i de andra bakterierna (Leung et al. 1992). Ett experiment av Ohnuki och Imanaka (1985) visade att två länkade gener som kallas strA-strB kodar för enzymer i S. griseus som är involverade i streptomycinresistens. Bakterien har dessa gener för att tåla antibiotikan som den själv producerar (Onuki & Imanaka 1985). Dessa två gener kodar för enzymer som kallas fosfotransferas, vilka inaktiverar streptomycin. Detta sker via en mekanism där enzymerna fosforylerar streptomycin, det vill säga adderar en fosfatgrupp, vilket gör att antibiotikan inte längre kan fungera (Thompson et al. 1996). Generna strA och strB befinner sig inte i bakteriens egna genom, utan finns på plasmider i bakteriens cytoplasma (Pezella et al. 2004). Ett experiment av Sunde och Norström (2005) visade ytterligare att generna strA-strB kan återfinnas i tarmbakterien Escherichia coli, där de

5
ger hög streptomycinresistens. I deras experiment undersöktes 136 olika stammar av E. coli som samlats in från köttprodukter. Av de 136 stammarna innehöll 110 av dessa strA-strB generna. De upptäckte även att flera stammar innehöll aadA genen. Bakterierna med en kombination av strA-strB och aadA var mer resistenta mot streptomycin än de bakterier som bara innehöll en av dessa gener (Sunde & Norström 2005). Det finns ytterligare en identifierad gen som kan ge streptomycinresistens efter mutation. Ett experiment av Okamoto och kollegor (2007) klargjorde att en gen som heter gidB är ansvarig för svag streptomycinresistens när den muteras. Flera av deras bakteriestammar visade att mutationer i denna gen sker väldigt frekvent. De visade även att mekanismen bakom resistens beror på att proteinet som kodas av genen, GidB, är involverad i metylering av 16S rRNA. I experimentet upptäcktes det att mutation av gidB-genen är orsaken till 33% av alla fall av resistensutveckling hos M. tuberculosis. Bakterier med mutationer i gidB har mer än 2000 gånger större sannolikhet att utveckla mutationer i rpsL och 16S rRNA, vilket ger hög streptomycinresistens (Okamoto et al. 2007). I ett liknande experiment av Spies och kollegor (2011) visade att cirka 27% av streptomycinresistenta bakteriestammar endast har en mutation i gidB-genen, vilket ytterligare bevisar genens betydelse för resistens. De flesta av deras resistenta stammar uppvisade endast svag resistens mot antibiotikan (Spies et al. 2011). Experiment med en annan stam visade att borttagning av gidB i bakterien Salmonella gav hög resistens mot streptomycin (Mikheil et al. 2012). Ett protein som kallas WhiB7 har visats kunna bidra till resistens mot aminoglykosider, streptomycin inkluderat. Detta verkar vara ett genetiskt välbevarat protein eftersom det kan hittas i nästan alla arter av aktinobakterier, exempelvis hela familjen Mycobacterium. Proteinet bildas när den motsvarande genen whiB7 aktiveras till följd av att bakterien utsätts för antibiotika. Resistensen uppkommer eftersom proteinet fungerar som en transkriptionsfaktor och aktiverar andra gener som inger antibiotikaresistens (Burian et al. 2012). Dessa kan exempelvis vara gener som är involverade i skyddandet av ribosomen samt gener som kodar för så kallade effluxpumpar (Morris et al. 2005). Effluxpumpar är proteinkomplex som aktivt kan utsöndra substanser, exempelvis skadliga antibiotika, ur en bakterie (Blanco et al. 2016). Eftersom genen whiB7 verkar finnas i väldigt många olika typer av bakterier, antyder detta att proteinet har en stor betydelse för jordlevande aktinobakterier (Burian et al. 2012). Hur sprids streptomycinresistens hos bakterier? De mutationer som kan inducera streptomycinresistens sprids ofta direkt från förälder till avkomma eftersom bakterier delar sig utan någon sexuell förökning (Newcombe & Nyholm 1950). Detta är dock inte det enda sättet som streptomycinresistensen kan spridas. Spridning mellan bakterier både inom eller mellan arter sker ofta via horisontell genöverföring (HGT), även om inte alla arter av bakterier gör detta. Horisontell genöverföring har spelat en nyckelroll i den evolutionära framgången hos bakterier, eftersom fördelaktiga gener snabbt kan fixeras i en population (Klug et al. 2014). HGT kan ske med tre olika mekanismer som heter konjugation, tranduktion och transformation (Burmeister 2015). Spridning via konjugation Konjugation är en bakteriell process där genetiskt material överförs från en bakterie till en annan via cell-till-cellkontakt. För att överföring av DNA ska kunna ske krävs en fertilitetsfaktor, eller F-faktor som den också kallas. F-faktorn är en plasmid som finns i bakteriens cytoplasma (Klug et al. 2014). Dessa plasmider kan ha antingen en brett eller ett smalt spektrum för värdceller. Detta innebär att plasmiden antingen kan överföras till vilken

6
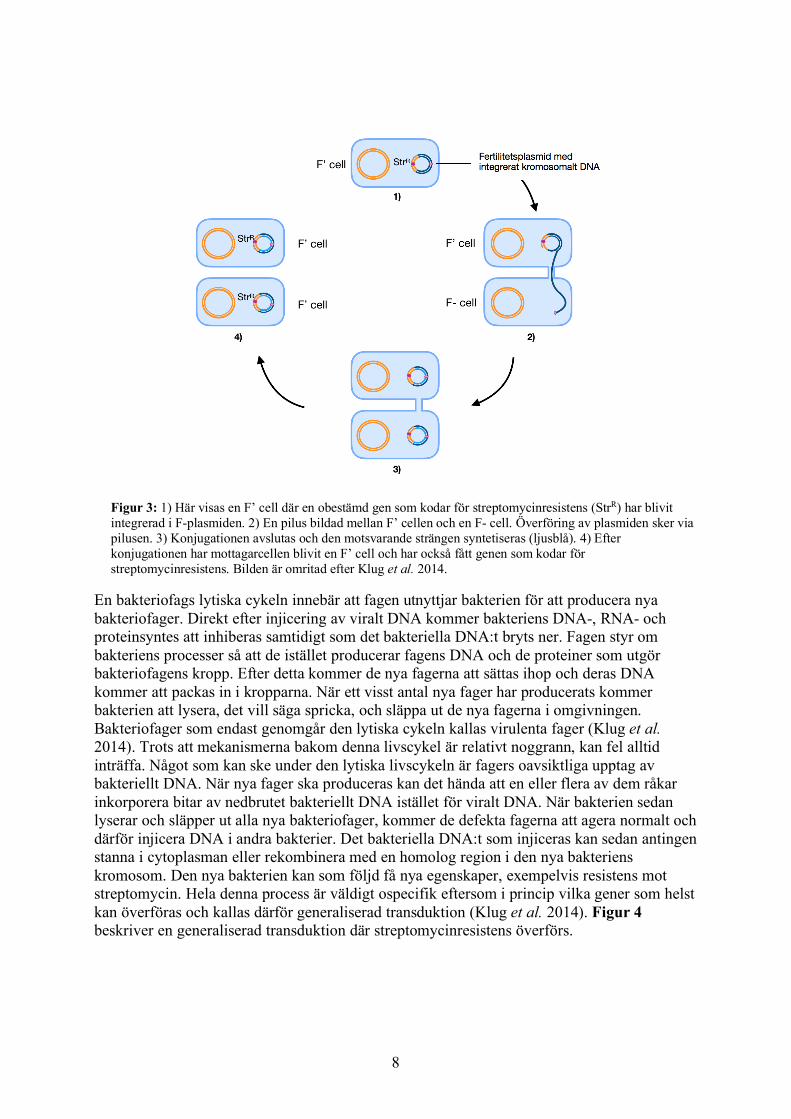
bakterie som helst respektive att den är begränsad till liknande bakteriearter (Bennett 2008). En av F-plasmidens uppgifter är att initiera formandet av en slags ”brygga” mellan två konjugerande celler. Denna brygga kallas F-pilus eller sex-pilus, och skapar kontakt mellan cellerna via en por, där det genetiska materialet kan överföras. För att överföring ska kunna ske mellan två celler måste den ena bakterier ha en F-faktor medan den andra bakterien måste vara utan. Bakterien med F-faktorn i cytoplasman kommer under konjugationen att fungera som ”donator” medan bakterien utan F-faktor kommer att fungera som ”mottagare”. Dessa namnges ofta som F+ respektive F-, där bakterien om kallas F+ har en fertilitetsplasmid och bakterien som kallas F- saknar plasmiden. (Klug et al. 2014). Vid konjugation kommer F-plasmidens dubbelsträngade DNA att separeras till två separata DNA-strängar varav den ena kommer att föras över till mottagaren. Den komplementära DNA-strängen kommer sedan att syntetiseras i både donatorn och mottagaren. Efter konjugationen har mottagarcellen fått en F-faktor och kan i fortsättningen agera som en donatorcell. Donatorcellen förlorar inte sin fertilitetsfaktor, då endast en kopia har förts över till mottagaren (Griffiths et al. 2000). Konjugation mellan bakterier sker väldigt snabbt vilket leder till att en F-faktor kan spridas inom en population under mycket kort tid (Griffiths et al. 1999). I vissa fall kan en bakteries genom innehålla något som kallas för IS-element, vilket står för ”införingssekvens” (eng: insertion sequence). Ett IS-element är segment av DNA som kan röra sig inom bakteriens genom eller mellan genom och plasmid. Om ett IS-element även finns på F-plasmiden öppnar det upp för möjligheten till en process som kallas homolog rekombination (Griffiths et al. 1999). Detta är en process där två likartade DNA-strängar utbyter DNA-segment mellan varandra, vilket leder till fler genetiska kombinationer och ökar den genetiska variationen (Kuzminov 2011). Om rekombination sker mellan bakteriens genom och F-plasmiden kan bakterien få F-faktorn integrerad i sitt DNA. Som en följd kan hela eller en större del av bakteriens genom överföras till en annan bakterie vid konjugation. Väl inne i mottagarcellen kan rekombination ske och mottagarcellen får därför gener från donatorcellen integrerade i sin kromosom. Då celler med integrerad F-faktor i sitt genom alltid överför bakteriella gener till en annan bakterie, kallas dessa för Hfr, vilket står för ”hög frekvens av rekombination”. Ett event där streptomycinresistens överförs mellan bakterier via detta sätt kan ses i Figur 2. Det kan även hända att F-faktorn lämnar kromosomen och återgår till att vara en plasmid. I dessa fall följer ibland några bakteriella gener med och när plasmiden överförs till mottagaren får cellen donatorns gener. En bakterie i ett sådant tillstånd kallas för F’ (F-prim). Väl inne i mottagarcellen syntetiseras den komplementära strängen och bakterien har på så sätt fått nya egenskaper. Figur 3 visas ett exempel på en konjugation där streptomycinresistens förs till en annan bakterie via en fertilitetsplasmid med integrerat bakteriellt DNA. Genom dessa olika typer av konjugation kan gener från en donatorcell effektivt föras över och integreras i en mottagarcell (Griffiths et al. 1999). Om donatorcellen har fått mutationer som ger streptomycinresistens, exempelvis en muterad rpsL gen, kan även detta överföras. Ett experiment av Watanabe och Fukasawa (1961) visade att resistens mot streptomycin kan överföras via konjugation mellan olika arter av bakterier, från stammen Shigella till arten E. coli (Watanabe & Fukasawa 1961).

7
Ett annat sätt som streptomycinresistens kan överföras är genom något som kallas för resistens-faktor eller R-faktor (Griffiths et al. 2000). Detta är, precis som F-faktorn, extracellulärt DNA i form av en plasmid som kan replikera oberoende av kromosomens funktioner. R-plasmiden består av två olika komponenter; en resistance transfer factor (RTF) och en eller flera r-determinanter. RTF-komponenten innehåller gener som är nödvändiga för överföring av plasmiden, exempelvis en F-faktor som möjliggör bildandet av en pilus. R-determinanter är specifika gener som kodar för olika typer av antibiotikaresistens. För att överföring av dessa plasmider ska kunna ske måste både RTF och r-determinanten finnas i bakterien. Resistens mot streptomycin, tillsammans med flera andra typer av antibiotikaresistens sprids ofta genom överföring av R-plasmider. Resistensfaktorn kan innehålla flera r-determinanter och bakterier som får plasmiden via konjugation blir ofta multiresistenta, det vill säga resistenta mot flera olika antibiotikum. Spridning av antibiotikaresistens via R-plasmider upptäcktes redan under 1950-talet i Japan när multiresistenta patogena bakterier från familjen Shigella påträffades i sjukhusmiljöer (Klug et al. 2014). Spridning via transduktion Bakterier behöver inte utföra cell-till-cellkontakt för att överföra genetiskt material. De kan också göra det med hjälp av bakteriofager i en process som kallas transduktion. Bakteriofager är virus som infekterar bakterier, vilket vanligtvis resulterar i värdcellens död. Viruset är ofta specifikt för en eller flera liknande bakterietyper. En bakteriofag infekterar en bakterie genom att fästa sig vid cellens utsida och sedan injicerar sitt virala DNA i bakteriens cytoplasma. Efter detta moment kan fagen genomgå en av två olika livscykler; antingen den lytiska eller den lysogena cykeln (Klug et al. 2014).
Figur 2: Ett exempel på hur en obestämd gen som kodar för streptomycinresistens (kallas här StrR) kan överföras till en bakterie som är känslig för streptomycin (kallas här StrK). 1) En Hfr cell har fått fertilitetsfaktorn integrerad i sin kromosom. 2) Cellen skapar kontakt med en F- cell via en pilus och överföring av DNA börjar. 3) Eftersom genen på F- cellens kromosom är homolog till Hfr cellens kan rekombination mellan generna ske. 4) F- cellen får en del av Hfr cellens gener i sin kromosom. Detta inkluderar genen som ger bakterien streptomycinresistens. Bilden är omritad från Klug et al. 2014.
8
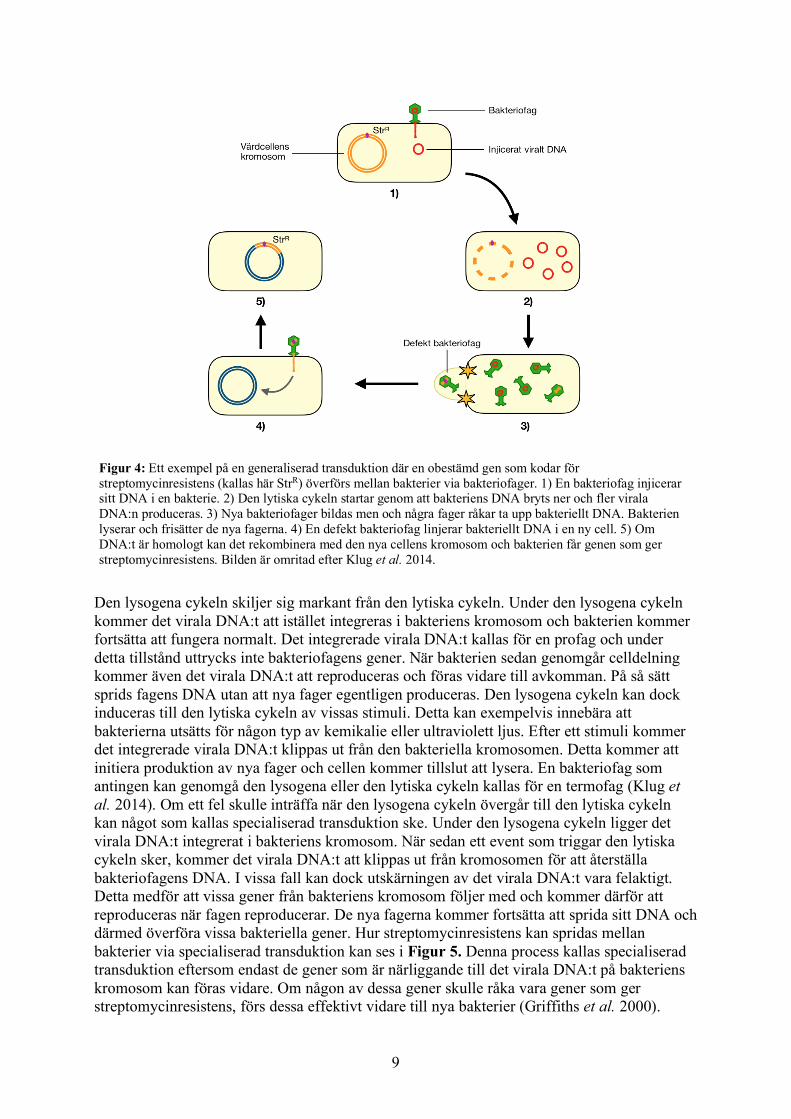
En bakteriofags lytiska cykeln innebär att fagen utnyttjar bakterien för att producera nya bakteriofager. Direkt efter injicering av viralt DNA kommer bakteriens DNA-, RNA- och proteinsyntes att inhiberas samtidigt som det bakteriella DNA:t bryts ner. Fagen styr om bakteriens processer så att de istället producerar fagens DNA och de proteiner som utgör bakteriofagens kropp. Efter detta kommer de nya fagerna att sättas ihop och deras DNA kommer att packas in i kropparna. När ett visst antal nya fager har producerats kommer bakterien att lysera, det vill säga spricka, och släppa ut de nya fagerna i omgivningen. Bakteriofager som endast genomgår den lytiska cykeln kallas virulenta fager (Klug et al. 2014). Trots att mekanismerna bakom denna livscykel är relativt noggrann, kan fel alltid inträffa. Något som kan ske under den lytiska livscykeln är fagers oavsiktliga upptag av bakteriellt DNA. När nya fager ska produceras kan det hända att en eller flera av dem råkar inkorporera bitar av nedbrutet bakteriellt DNA istället för viralt DNA. När bakterien sedan lyserar och släpper ut alla nya bakteriofager, kommer de defekta fagerna att agera normalt och därför injicera DNA i andra bakterier. Det bakteriella DNA:t som injiceras kan sedan antingen stanna i cytoplasman eller rekombinera med en homolog region i den nya bakteriens kromosom. Den nya bakterien kan som följd få nya egenskaper, exempelvis resistens mot streptomycin. Hela denna process är väldigt ospecifik eftersom i princip vilka gener som helst kan överföras och kallas därför generaliserad transduktion (Klug et al. 2014). Figur 4 beskriver en generaliserad transduktion där streptomycinresistens överförs.
Figur 3: 1) Här visas en F’ cell där en obestämd gen som kodar för streptomycinresistens (StrR) har blivit integrerad i F-plasmiden. 2) En pilus bildad mellan F’ cellen och en F- cell. Överföring av plasmiden sker via pilusen. 3) Konjugationen avslutas och den motsvarande strängen syntetiseras (ljusblå). 4) Efter konjugationen har mottagarcellen blivit en F’ cell och har också fått genen som kodar för streptomycinresistens. Bilden är omritad efter Klug et al. 2014.
9
Den lysogena cykeln skiljer sig markant från den lytiska cykeln. Under den lysogena cykeln kommer det virala DNA:t att istället integreras i bakteriens kromosom och bakterien kommer fortsätta att fungera normalt. Det integrerade virala DNA:t kallas för en profag och under detta tillstånd uttrycks inte bakteriofagens gener. När bakterien sedan genomgår celldelning kommer även det virala DNA:t att reproduceras och föras vidare till avkomman. På så sätt sprids fagens DNA utan att nya fager egentligen produceras. Den lysogena cykeln kan dock induceras till den lytiska cykeln av vissas stimuli. Detta kan exempelvis innebära att bakterierna utsätts för någon typ av kemikalie eller ultraviolett ljus. Efter ett stimuli kommer det integrerade virala DNA:t klippas ut från den bakteriella kromosomen. Detta kommer att initiera produktion av nya fager och cellen kommer tillslut att lysera. En bakteriofag som antingen kan genomgå den lysogena eller den lytiska cykeln kallas för en termofag (Klug et al. 2014). Om ett fel skulle inträffa när den lysogena cykeln övergår till den lytiska cykeln kan något som kallas specialiserad transduktion ske. Under den lysogena cykeln ligger det virala DNA:t integrerat i bakteriens kromosom. När sedan ett event som triggar den lytiska cykeln sker, kommer det virala DNA:t att klippas ut från kromosomen för att återställa bakteriofagens DNA. I vissa fall kan dock utskärningen av det virala DNA:t vara felaktigt. Detta medför att vissa gener från bakteriens kromosom följer med och kommer därför att reproduceras när fagen reproducerar. De nya fagerna kommer fortsätta att sprida sitt DNA och därmed överföra vissa bakteriella gener. Hur streptomycinresistens kan spridas mellan bakterier via specialiserad transduktion kan ses i Figur 5. Denna process kallas specialiserad transduktion eftersom endast de gener som är närliggande till det virala DNA:t på bakteriens kromosom kan föras vidare. Om någon av dessa gener skulle råka vara gener som ger streptomycinresistens, förs dessa effektivt vidare till nya bakterier (Griffiths et al. 2000).
Figur 4: Ett exempel på en generaliserad transduktion där en obestämd gen som kodar för streptomycinresistens (kallas här StrR) överförs mellan bakterier via bakteriofager. 1) En bakteriofag injicerar sitt DNA i en bakterie. 2) Den lytiska cykeln startar genom att bakteriens DNA bryts ner och fler virala DNA:n produceras. 3) Nya bakteriofager bildas men och några fager råkar ta upp bakteriellt DNA. Bakterien lyserar och frisätter de nya fagerna. 4) En defekt bakteriofag linjerar bakteriellt DNA i en ny cell. 5) Om DNA:t är homologt kan det rekombinera med den nya cellens kromosom och bakterien får genen som ger streptomycinresistens. Bilden är omritad efter Klug et al. 2014.

10
Transduktion har visats vara ett enkelt sätt för streptomycinresistens att spridas, som demonstrerat i ett experiment av Kazuo Hashimoto. I försöket beskrivs hur streptomycinresistens överförs från resistenta till antibiotikakänsliga E. coli bakterier. Han beskriver även uppkomsten av revertanter, det vill säga bakterier som återgått till att vara känsliga mot streptomycin. Hashimoto nämner dock inga specifika gener som överförs (Hashimoto 1960). Spridning via transformation Transformation är ännu ett exempel på genetisk överföring hos bakterier som inte involverar cell-till-cellkontakt. Processen innebär att bakterier aktivt tar upp små bitar av DNA från omgivningen. DNA:t kan sedan exempelvis användas som material till DNA-uppbyggnad eller som näring för cellen. Den viktigaste aspekten är dock att transformation ger en möjlighet för bakterien att få nya genetiska egenskaper (Krüger & Stingl 2011). Fritt DNA i omgivningen kan hittas efter att en cell dör och lyserar, vilket frisätter DNA-segment och plasmider (Kloos et al. 1994). Fritt DNA är allmänt förekommande i flera olika bakteriella miljöer, speciellt i formationer av biofilm (Antonova & Hammer 2011). För att en bakterie ska kunna ta upp genetiskt material från omgivningen krävs att cellen är i ett tillstånd som kallas för kompetens (Klug et al. 2014). De flesta bakterier är inte kompetenta hela tiden, utan detta är ett tillstånd som regleras av miljön eller biokemiska faktorer Vad som kan inducera kompetens hos en bakterie kan inte konkret bestämmas eftersom det skiljer sig markant mellan arter. Det är känt att förekomsten av fritt DNA i omgivningen inte har någon effekt på inducering av tillståndet. Däremot kan avsaknaden eller förekommande av näringsämnen göra
Figur 5: Specialiserad transduktion där en obestämd gen som kodar för streptomycinresistens (kallas här StrR) överförs mellan bakterier via bakteriofager. 1) En bakteriofag injicerar viralt DNA i en bakterie som är resistent mot streptomycin. 2) Det virala DNA:t integreras i den bakteriella kromosomen, vilket kallas profag. Bakterien kan fortsätta fungera normalt och dela sig. 3) Ett stimuli, exempelvis UV-ljus, triggar utskärning av det virala DNA:t men utskärningen går fel och närliggande bakteriella gener följer med, även genen som ger streptomycinresistens. 4) Den lytiska cykeln induceras och nya fager produceras. Alla fager är delvis defekta och bär på bakteriella gener. 5) En defekt jag injicerar DNA i en bakterie. Detta integreras i bakteriens kromosom och cellen blir som följd resistent. Bilden är omritad efter Lang et al. 2012.

11
celler kompetenta (Mell & Redfield 2014). Bakterier som utsätts för vissa stressförhållanden har visats kunna ta upp fritt DNA. Exempelvis har det under experiment noterats att bakterien Streptococcus pneumoniae kan bli kompetent efter att den utsatts för vissa doser av antibiotika (Slager et al. 2014). Bakterien Haemophilus influenzae kan istället bli kompetent som en respons till avsaknaden av näringsämnen. Detta har visats inträffa under experiment där bakterien flyttats från ett näringsrikt till ett näringsfattigt tillväxtmedium (Redfield 1991). Möjligheten för en bakteriecell att vara kompetent och kunna ta upp DNA från omgivningen skiljer sig mellan olika arter. Bakterien S. pneumoniae är ett exempel på en patogen bakterie som har naturligt möjlighet att ta upp DNA från omgivningen (Slager et al. 2014). Den vanliga tarmbakterien Escherichia coli är ett exempel på en bakterie som inte är naturligt kompetent (Palchevskiy & Finkel 2006). Det har sedan länge varit känt att hos bakterier som saknar naturlig kompetens kan tillståndet kemiskt induceras med en behandling av kalciumklorid (Dagert & Ehrlich 1979). Hur transformationen sker skiljer sig något mellan gram-positiva och gram-negativa bakterier. Hur det ser ut när gram-positiva respektive gram-negativa bakterier gör när de tar upp extracellulärt DNA kan ses i Figur 6. Transformationens första steg är bindningen av extracellulärt dubbelsträngat DNA till cellens yta. Detta sker med en typ av pilus, som binder till DNA:t och drar det mot bakteriens yta. Denna pilus skiljer sig från den tidigare beskrivna F-pilusen, som används under konjugation. Transformationens pilus är istället indragbar, vilket betyder att pilusen kan dras in mot cellens yta (Filloux 2010). Hos gram-positiva bakterier finns ingen preferens för något specifikt DNA medan hos gram-negativa bakterier har det visats en preferens för specifika sekvenser av homologt DNA. Det andra steget av transformation är transport av det bundna DNA:t in i bakteriens cytoplasma. Hos gram-positiva bakterier, som saknar ett yttre membran, kan DNA:t direkt föras ner till en por där det kan dras in i cytoplasman av pilusen. Hos gram-negativa bakterier kommer pilusen att dra DNA:t genom en por i det yttre membranet och sedan genom en annan por i det inre membranet. För båda typer av bakterier kommer DNA:t att göras enkelsträngat istället för dubbelsträngat efter att det förts genom den inre poren (Stewart & Carlson 1986). Väl inne i cellens cytoplasma kan två saker hända med det genetiska materialet. Det första som kan hända är att DNA-segmentet bryts ner till mindre komponenter för att återanvändas av bakterien. Detta är ett energieffektivt sätt att tillhandahålla de molekyler som behövs för att bakterien ska kunna syntetisera nytt DNA. Egen produktion av dessa molekyler är ofta kostsamt för bakterien och transformation ger därför möjligheten till att spara på resurser. Det andra som kan hända när DNA:t befinner sig i cytoplasman är homolog rekombination. Om det upptagna materialet uppvisar någon form av homologi, det vill säga liknar bakteriens egna DNA, kan utbyte av gener ske. På så sätt kan bakterier få nya genetiska egenskaper, exempelvis resistens mot vissa antibiotikum (Mell & Redfield 2014). Transformation av bakterier till att bli streptomycinresistenta har experimentellt visats i flera arter. Bland annat har arterna Xanthomonas phaseoli, Streptococcus och Neisseria gonorrhoeae alla visats vara kapabla till att bli streptomycinresistenta efter transformation (Corey & Starr 1957, Perry & Slade 1962, Starling 1966).

12
Diskussion Streptomycinresistens kan alltså uppkomma hos bakterier via fem olika mekanismer; arv från förälder till avkomma, spontana mutationer, konjugation mellan bakterier, transformation av fritt DNA eller tranduktion med bakteriofager. Bakterier är med andra ord väldigt duktiga på att anpassa sig till rådande förutsättningar. Det är fascinerande att resistens kan uppstå hos en population av bakterier med tanke på att uppkomsten av antibiotikaresistens är en helt slumpmässig process. Mutationer och överföring av gener via HGT sker spontant och de inblandade generna väljs alltså ut av slumpen. Av alla gener i en bakteriell kromosom är det därför just de gener som uppsatsen diskuterat; rpsL, gidB, strA-strB, gidB, whiB7, aadA eller 16S rRNA gener som måste muteras och sedan överföras. Chansen för att det här ska inträffa verkar liten men trots det händer detta ofta. En annan aspekt som verkar bero helt av slumpen är att uppkomsten av dessa mutationer sker innan bakterien utsätts för antibiotikan. Bakterier blir inte antibiotikaresistenta som följd av en antibiotikabehandling, utan det har sedan länge varit känt att resistenta bakterier uppkommer innan de utsätts för antibiotikumet (Griffiths et al. 2000). Bakterier kan dock inte förutse om de kommer utsättas för toxiska miljöer och de bakterier som blir resistenta blir det helt slumpmässigt. Det antibiotikan gör är istället att utsätta bakterierna för en selekterande miljö, där endast de resistenta överlever och kan reproduceras. Förmodligen är det bakteriers snabba generationstid som gör att vi människor
Figur 6: A) visar hur extracellulärt DNA förs in i den gram-positiva bakterien. B) visar motsvarande process i gram-negativa bakterier. I båda typer av bakterier sker processen i två steg; 1) bindning av DNA till cellens yta via en pilus. 2) Pilus drar ner DNA:t genom en respektive två porer och DNA:t görs enkelsträngat innan det transporteras in i cellens cytoplasma. Bilden är omritad från Martin et al. 2014.

13
upplever bakterier som så pass anpassningsbara. En skrämmande aspekt om antibiotikaresistens är spridningen av multiresistenta bakterier via konjugation av R-plasmider. Konjugationen går så pass snabbt och enkelt för bakterier, vilket gör att R-faktorn enkelt sprids i en population av bakterier. Det är speciellt skrämmande att dessa plasmider inte bara kan sprida resistens mot en antibiotika utan flera. Multiresistenta bakterier kan skapa stora problem inom sjukvården eftersom de antibiotikum som brukar verka på bakterierna inte kan användas. Sjuka personer som ej kan behandlas innebär också en smittorisk, där andra människor kan smittas av den resistenta bakterien. Detta kan på sikt leda till utbrott av epidemier av exempelvis influensa, där många människor riskerar att insjukna (WHO 2017). Konsekvenser av överanvändning inom sjukvård och lantbruk Att antibiotika har använts på de sätt som de har gjort de senaste årtiondena är en av de största bidragande orsakerna till spridning av antibiotikaresistens. Felaktig användning av antibiotika har minskat i flera länder men tyvärr sker oreglerat bruk fortfarande världen över. Globalt fortsätter antibiotikaanvändning på både människor, djur och inom jordbruk att öka. Hur allvarliga effekter som användning av antibiotika på djur och växter genererar är dock tvistat om. Scherer och medförfattare (2013) publicerade en artikel 2013 där de utfört en studie som visar en tydlig ökning av streptomycinresistenta E. coli och Staphylococcus i nosen och avföringen hos får. Fåren hade tillåtits beta på fält som sprejats med streptomycin i de koncentrationer som används i fruktträdgårdar. Ökningen av resistenta bakterier gick från 22,3% i kontrollgruppen till 39,9% i testgruppen (Scherer et al. 2013). En annan studie som publicerades 2014 av Duffy och kollegor visade däremot att användning av streptomycin som ett bekämpningsmedel på äppelträd inte påverkar förekomsten av resistens hos närlevande bakterier. Prover togs från träden samt omkringliggande jord från tre olika äppelodlingar under 2010, 2011 och 2012. Statistiska analyser visade inga signifikanta resultat för att streptomycinanvändningen skulle påverka resistensutveckling hos den bakteriella faunan (Duffy et al. 2014). Att antibiotikaresistens sprids i en ökad takt på grund av felaktig användning ett allvarligt problem. Detta beror delvis på lättillgängligheten av dessa via försäljning över internet. Att vissa nationer fortfarande inte reglerar sin användning av antibiotika påverkar inte bara landet i sig utan också alla andra. Tack vare bättre och effektivare förbindelser mellan länder sprids resistenta bakterier med resande människor eller transport av djur och grödor. Detta gör att antibiotikaresistenta bakterier enkelt sprids mellan länder och även hamnar i maten vi äter. När människor blir infekterade med dessa resistenta bakterier leder det till en ökad belastning på sjukvården oavsett var i världen personen bor. Patienten riskerar längre sjukdomstid, längre/fler besök på sjukhus, ökad risk för dödlig utgång och minskad säkerhet under operationer. Detta leder i sin tur till ökade kostnader, både för individen och för samhället (WHO 2015). Vad görs för att minska spridning av antibiotikaresistens i Sverige och globalt? Enligt Folkhälsomyndigheten har Sverige sedan flera år tillbaka haft en plan för långsiktig minskning av antibiotikaresistens. På nationell nivå finns nätverket Strama; Strategigruppen för rationell användning och minskad antibiotikaresistens, som bildades redan 1995. Strama-grupper finns i varje län och leds av en smittskyddsläkare. Deras främsta uppgift är att analysera data om resistens samt receptutskrivning av antibiotika. Sverige samarbetar även aktivt med både Europeiska Unionen (EU) och Världshälsoorganisationen (WHO) för internationella insatser i minskad antibiotikaresistens. Vi deltar också i ”European Antibiotic

14
Awareness Day” initierat av europeiska smittskyddsmyndigheten för att sprida kunskap om korrekt användning av antibiotika. Det som görs i Sverige för minskad resistens är utförlig resistens- och utskrivningsbevakning samt behandlingsrekommendationer och information till sjukvårdspersonal. Resistensövervakning sker genom odling av prover från patienter som ska behandlas med antibiotika. Data samlas sedan in och jämförs nationellt. Utskrivnings- och försäljningsövervakning sker genom att apotek dagligen skickar statistik över sålda mediciner till e-Hälsomyndigheten, vilka sammanställer data och ger en nationell statistik över läkemedelsbruket i Sverige. En bristande faktor i detta sammanhang har varit avsaknad av information om varför antibiotika har föreskrivits till personen. En viktig faktor i spridandet av resistens är framtagandet av behandlingsrekommendationer och information till både sjukvårdspersonal och allmänheten. Detta infattar information om när antibiotika bör och ej bör användas samt allmän information om risken vid uppkomst av resistenta bakterier (Folkhälsomyndigheten 2014). Det är inte bara Sverige som har en plan för att på lång sikt minska spridningen av antibiotikaresistens. 2015 publicerade WHO en ny internationell plan för att minska problemen globalt. Planen liknar i många fall den i Sverige där ökad förståelse, ökad bevakning och optimering av antibiotikaanvändning är viktiga punkter. WHO vill även arbeta med att förebygga orsakerna till infektion och sjukdom via bättre hygieniska standarder i sjukhus världen över. De uppmuntrar även vaccinationer som ett förebyggande medel mot farliga sjukdomar (WHO 2015). Ytterligare ett globalt problem är den drastiska minskningen av nya antibiotikum och vacciner de senaste åren. Detta beror till stor del av det minskade intresset för investering i nya former av läkemedel. Av de 18 största läkemedelsföretagen världen över, har 15 av dessa övergivit antibiotikafältet. De har även minskat antalet forskningsgrupper som arbetar inom fältet (Ventola 2015). Detta beror på flera olika faktorer, där de största orsakerna är ekonomiska och regulatoriska barriärer. Antibiotika är relativt billigt och behandlingstiden är ofta kort, vilket gör tillverkningen av antibiotika inte är tillräckligt lönsamt för företag. Detta om man jämför med att tillverka mediciner för kroniska sjukdomar, exempelvis diabetes och astma, vilket genererar mer pengar. Mikrobiologer och infektionsspecialister har även uppmanat till försiktighet vid användning av antibiotika på grund av resistensproblem, vilket gör att företag inte alltid börjar sälja en ny antibiotika direkt. Istället kan läkemedlet sparas för att användas vid nödsituationer, när ingen annan antibiotika fungerar. En annan orsak till minskad forskning inom antibiotikafältet är regulatoriska barriärer. För att ett företag ska få producera nya antibiotikum krävs tillstånd. Antalet tillstånd som delats ut har minskat drastiskt de senaste årtiondena på grund av rädsla för resistensproblem. Att kliniskt få testa nya antibiotikum har också blivit en svår barriär att passera på grund av ändringar i lagar om hur kliniska prövningar får utföras. Äldre metoder, där exempelvis placeboeffekter studeras anses numera vara oetiska och kan därför inte längre användas. Att testa antibiotikan blir därför en kostnadsfråga då andra testmetoder kräver fler antal prover och är därför dyrare (Ventola 2015). På grund av dessa faktorer som hindrar uppkomsten av nya läkemedel vill Världshälsoorganisationen öka intresset för investering i nya antibiotika och vacciner samt skapa en stabil ekonomisk marknad för detta (WHO 2015). Framtidsutsikter för streptomycin och antibiotika. Kan de fortsätta användas eller behövs nya metoder? Trots att flera länder och organisationer försöker minska spridningen av antibiotikaresistens globalt, är det inte säkert att detta är lösningen på de medicinska problemen. Bakterier kommer alltid att mutera och antibiotika kommer alltid att selektera ut de resistenta bakterierna, även om användningen av läkemedlen minskar. Istället behöver mänskligheten kanske upptäcka nya former av antibiotikum, som på ett säkert sätt kan behandla infektioner

15
och sjukdomar utan att generera resistens. Ett exempel på en sådan ny metod är användandet av en ny klass av antibiotika som kallas LpxC-hämmare. Dessa medel verkar inte genom att aktivt döda bakterier via hämmandet av viktiga processer i bakteriecellen, så som traditionella antibiotikum gör. Istället verkar medlet genom att öka bakteriens känslighet för fagocytos av makrofager från värdorganismens immunförsvar (Lin et al. 2012). Detta beror på att medlet hämmar ett protein i bakterier som kallas för LpxC. Proteinet är involverad i syntesen av lipider som bidrar till cellens toxicitet (Barb & Zhou 2008). De rent molekylära mekanismerna bakom detta är inte helt kända men det har visats att LpxC-hämmarens aminosyror interagerar med LpxC-protinet via specifika vätebindingar (Kurasaki et al. 2016). Experiment med LpxC-hämmare har visat att möss som infekterats med den patogena bakterien Acinetobacter baumanni hade 100% chans att överleva infektionen om de blivit behandlade med LpxC-hämmaren. Detta jämfört med obehandlade möss som hade 0% chans att överleva infektionen (Lin et al. 2012). Det nyligen upptäckta CRISPR-Cas9 är ännu ett sätt att kunna motverka antibiotikaresistens hos bakterier (Kim et al. 2016). CRISPR-Cas9 är en genmodifierande metod som isolerats från bakteriers egna immunsystem. Det kan användas för att exempelvis rätta till fel i genom samt inducera eller stänga av genuttryck (Redman et al. 2016). Experiment har visat att multiresistenta E. coli med plasmider för antibiotikaresistens kan återgå till att vara antibiotikakänsliga med hjälp av CRISPR. Detta har gjorts genom att identifiera viktiga DNA-sekvenser i bakterierna och sedan manipulera dessa med CRISPR-Cas9 för att återinföra känslighet med antibiotika i cellerna (Kim et al. 2016). Det har även spekulerats om att modifierade bakteriofager ska användas för att vända bakteriens egna CRISPR-system mot den själv. De modifierade fagerna innehåller genetiskt information som ska trigga igång bakteriens CRISPR-system till att producera enzymer som bryter ner bakteriens DNA och därmed dödar den (Reardon 2017). Förutom att vara ett substitut mot dagens antibiotika, har det även testats om CRISPR-Cas9 skulle kunna användas för att bota genetiska sjukdomar och HIV (Redman et al. 2016). Även om antibiotikaresistens är ett globalt problem som hotar människors välfärd, finns det fortfarande hopp för framtiden. Att fler och fler länder börja reglera sin användning gör att mänskligheten är på god väg att minska resistensutvecklingen. Uppkomsten av nya metoder och teknologier för att bekämpa infektioner och sjukdomar antyder även en ljusare framtid för människors hälsa. Eftersom jag har insett problematiken med resistenta bakterier hoppas jag att fler företag framöver vill investera i forskning om nya antibiotikum. Jag skulle även vilja se fortsatt forskning av de nya metoder jag har skrivit om, så att mänskligheten i framtiden kan slippa de resistensproblem som existerar idag. Med ny forskning kanske antibiotika som läkemedel kan fasas ut och ersättas med bättre och lindrigare metoder. � Tack Ett stort tack till mina medstudenter Carl Smith och Victor Malmsjö som återkopplat och gett värdefulla kommentarer på min uppsats. Jag vill även tacka min handledare Erik Holmqvist som gett goda råd under skrivprocessen. Jag är även tacksam mot min sambo Paul Wallin, som tagit sig tiden att läsa och kommentera på min text.

16
Referenser Aminov RI. 2010. A Brief History of the Antibiotic Era: Lessons Learned and Challenges for
the Future. Frontiers in Microbiology 1:134. Antonova ES, Hammer BK. 2011. Quorum-sensing autoinducer molecules produced by
members of a multispecies biofilm promote horizontal gene transfer to Vibrio cholerae. FEMS Microbiology Letters 322: 68-76.
Barb AW, Zhou P. 2008. Mechanism and Inhibition of LpxC: an essential zinc-dependent deacetylase of bacterial lipid A synthesis. Current pharmaceutical biotechnology 9: 9-15.
Barker KF. 1999. Antibiotic resistance: a current perspective. British Journal of Clinical Pharmacology 48: 109-124.
Bennett PM. 2008. Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic resistance genes in bacteria. British Journal of Pharmacology 153: S347-S357.
Blanco P, Hernando-Amado S, Reales-Calderon JA, Corona F, Lira F, Alcalde-Rico M, Bernardini A, Sanchez MB, Martinez JL. 2016. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 4: 14.
Brown L, Wolf JM, Prados-Rosales R, Casadevall A. 2015. Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nature Reviews Microbiology 13: 620-630.
Burmeister AR. 2015. Horizontal Gene Transfer. Evolution, Medicine, and Public Health 2015: 193-194.
Burian J, Ramón-García S, Sweet G, Gómez-Velasco A, Av-Gay Y, Thompson CJ. 2012. The Mycobacterial Transcriptional Regulator whiB7 Gene Links Redox Homeostasis and Intrinsic Antibiotic Resistance. The Journal of Biological Chemistry 287: 299-310.
Campbell NA, Reece JB, Urry LA, Cain ML, Wasserman SA, Minorsky PV, Jackson RB. 2014. Expression of Genes. I: Wilbur B (red.). Biology: A Global Approach, ss. 392-403. Pearson Education Limited, Kendallville.
Carter AP, Clemons WM, Brodersen DE, Morgan-Warren RJ, Wimberly BT, Ramakrishnan V. 2000. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 407: 340-348.
Center for Disease Control and Prevention (CDC). 2013. Antibiotic Resistance Threats in the Unites States, 2013. Rapport, U.S. Departement of Health and Human Services.
Chater KF. 2006. Streptomyces inside-out: a new perspective on the bacteria that provide us with antibiotics. Philosophical Transactions of the Royal Society B: Biological Sciences 361: 761-768.
Clardy J, Fischbach M, Currie C. 2009. The natural history of antibiotics. Current biology 19: R437-R441
Cooper GM. 2000. The Cell - a Molecular Approach, 2: a uppl. Sinauer Associates, Sunderland. Corey RR, Starr MP. 1957. Genetic Transformation of Streptomycin Resistance in
Xanthomonas Phaseoli. Journal of Bacteriology 74: 146-150. Dagert M, Ehrlich SD. 1979. Prolonged incubation in calcium chloride improves the
competence of Escherichia coli cells. Gene 6: 23-28. Davies J, Davies D. 2010. Origins and Evolution of Antibiotic Resistance. Microbiology and
Molecular Biology Reviews 74: 417-433. Denamur E, Matic I. 2006. Evolution of mutation rates in bacteria. Molecular Microbiology
60: 820-827. Duffy B, Holliger E, Walsh F. 2014. Streptomycin use in apple orchards did not increase
abundance of mobile resistance genes. FEMS Microbiology Letters 350: 180-189. Filloux A. 2010. A Variety of Bacterial Pili Involved in Horizontal Gene Transfer. Journal of
Bacteriology 192: 3243-3245.

17
Folkhälsomyndigheten. 2014. Svenskt arbete mot antibiotikaresistens. WWW-dokument: https://www.folkhalsomyndigheten.se/publicerat-material/publikationsarkiv/s/svenskt-arbete-mot-antibiotikaresistens/. Hämtad 2017-11-15.
Griffiths AJF, Gelbart WM, Miller JH, Lewontin RC. 1999. Modern Genetic Analysis. W.H Freeman, New York.
Griffiths AJF, Miller JH, Suzuki DT, Lewontin RC, Gelbart WM. 2000. An Introduction to Genetic Analysis, 7: e uppl. W.H Freeman, New York.
Hashimoto K. 1960. Streptomycin Resistance in Escherichia Coli Analyzed by Transduction. Genetics 45: 49-62.
Hollingshead S, Vapnek D. 1985. Nucleotide sequence analysis of a gene encoding a streptomycin/spectinomycin adenyltransferase. Plasmid 13: 17-30.
Honoré N, Cole ST. 1994. Streptomycin resistance in mycobacteria. Antimicrobial Agents and Chemotherapy 38: 238-242.
Keseler IM, Mackie A, Santos-Zavaleta A, Billington R, Bonavidez-Martínez C, Caspi R, Fulcher C, Gama-Castro S, Kothari A, Krummenacker M, Latendresse M, Muñiz-Rascado L, Ong Q, Paley S, Peralta-Gil M, Subhraveti P, Velázquez-Ramírez DA, Weaver D, Collado-Vides J, Paulsen I, Karp PD. 2017. The EcoCyc database: reflecting new knowledge about Escherichia coli K-12. Nucleic Acids Research 45: D543-D550.
Kim JS, Cho DH, Park M, Shin WJ, Ko KS, Kweon DH. 2016. CRISPR/Cas9-Mediated Re-Sensitization of Antibiotic-Resistant Escherichia coli Harboring Extended-Spectrum β-Lactamases. Journal of Microbiology and Biotechnology 26: 394-401.
Kloos DU, Strätz M, Güttler A, Steffan RJ, Timmis KN. 1994. Inducible cell lysis system for the study of natural transformation and environmental fate of DNA released by cell death. Journal of Bacteriology 176: 7352-7361.
Klug WS, Cummings MR, Spencer CA, Palladino MA. 2014. Essentials of Genetics, 8: e uppl. Pearson Education Limited, Edinburgh.
Krüger NJ, Stingl K. 2011. Two steps away from novelty – principles of bacterial DNA uptake. Molecular Microbiology 80: 860-867.
Kurasaki H, Tsuda K, Shinoyama M, Takaya N, Yamaguchi Y, Kishii R, Iwase K, Ando N, Nomura M, Kohno Y. 2016. LpxC Inhibitors: Design, Synthesis, and Biological Evaluation of Oxazolidinones as Gram-negative Antibacterial Agents. ACS Medicinal Chemistry Letters 7: 623-628.
Kuzminov A. 2011. Homologous Recombination - Experimental Systems, Analysis and Significance. EcoSal Plus, doi 10.1128/ecosalplus.7.2.6.
Lang AS, Zhaxybayeva O, Beatty TJ. 2012. Gene transfer agents: phage-like elements of genetic exchange. Nature reviews Microbiology 10: 472-482.
Leung KY, Ruschkowski SR, Finlay BB. 1992. Isolation and characterization of the aadA aminoglycoside-resistance gene from Salmonella choleraesuis. Molecular Microbiology 6: 2453-2460.
Lin L, Tan B, Pantapalangkoor P, Ho T, Baquir B, Tomaras A, Montgomery JI, Reilly U, Barbacci EG, Hujer K, Bonomo RA, Fernandez L, Hancock REW, Adams MD, French SW, Buslon VS, Spellberg B. 2012. Inhibition of LpxC Protects Mice from Resistant Acinetobacter baumannii by Modulating Inflammation and Enhancing Phagocytosis. mBio 3: e00312-12.
Martin B, Johnston C, Fichant G, Claverys JP, Polard P. 2014. Bacterial transformation: distribution, shared mechanisms and divergent control. Nature Reviews Microbiology 12: 181.
Mell JC, Redfield RJ. 2014. Natural Competence and the Evolution of DNA Uptake Specificity. Journal of Bacteriology 196: 1471-1483.
Mikheil DM, Shippy DC, Eakley NM, Okwumabua OE, Fadl AA. 2012. Deletion of gene

18
encoding methyltransferase (gidB) confers high-level antimicrobial resistance in Salmonella. The Journal of Antibiotics 65: 185-192.
Morris RP, Nguyen L, Gatfield J, Visconti K, Nguyen K, Schnappinger D, Ehrt S, Liu Y, Heifets L, Pieters J, Schoolnik G, Thompson CJ. 2005. Ancestral antibiotic resistance in Mycobacterium tuberculosis. Proceedings of the National Academy of Sciences of the United States of America 102: 12200-12205.
National Center for Biotechnology Information (NCBI). PubChem Compound Database; CID=19649. Streptomycin. WWW-dokument: https://pubchem.ncbi.nlm.nih.gov/compound/19649. Hämtad 2017-11-20.
Newcombe HB, Nyholm MH. 1950. The Inheritance of Streptomycin Resistance and Dependence in Crosses of Escherichia Coli. Genetics 35: 603-611.
Ohnuki T, Imanaka T. 1985. Self-cloning in Streptomyces griseus of an str gene cluster for streptomycin biosynthesis and streptomycin resistance. Journal of Bacteriology 164: 85-94.
Okamoto S, Nakajima C, Nishimura K, Tanaka Y, Tokuyama S, Suzuki Y, Ochi K. 2007. Loss of a conserved 7-methylguanosine modification in 16S rRNA confers low-level streptomycin resistance in bacteria. Molecular Microbiology 63: 1096-1106.
Palchevskiy V, Finkel SE. 2006. Escherichia coli Competence Gene Homologs Are Essential for Competitive Fitness and the Use of DNA as a Nutrient. Journal of Bacteriology 188: 3902-3910.
Perry D, Slade HD. 1962. Transformation of Streptococci to Streptomycin Resistance. Journal of Bacteriology 83: 443-449.
Pezzella C, Ricci A, DiGiannatale E, Luzzi I, Carattoli A. 2004. Tetracycline and Streptomycin Resistance Genes, Transposons, and Plasmids in Salmonella enterica Isolates from Animals in Italy. Antimicrobial Agents and Chemotherapy 48: 903-908.
Reardon S. 2017. Modified viruses deliver death to antibiotic-resistant bacteria. Nature News 546: 586.
Redfield RJ. 1991. sxy-1, a Haemophilus influenzae mutation causing greatly enhanced spontaneous competence. Journal of Bacteriology 173: 5612-5618.
Redman M, King A, Watson C, King D. 2016. What is CRISPR/Cas9? Archives of Disease in Childhood. Education and Practice Edition 101: 213-215.
Ruiz P, Rodríguez-Cano F, Zerolo FJ, Casal M. 2002. Investigation of the in vitro activity of streptomycin against Mycobacterium tuberculosis. Microbial Drug Resistance 8: 147-149.
Scherer A, Vogt HR, Vilei EM, Frey J, Perreten V. 2013. Enhanced antibiotic multi-resistance in nasal and faecal bacteria after agricultural use of streptomycin: Increase in resistance after streptomycin use. Environmental Microbiology 15: 297-304.
Sharma D, Cukras AR, Rogers EJ, Southworth DR, Green R. 2007. Mutational analysis of S12 protein and implications for the accuracy of decoding by the ribosome. Journal of molecular biology 374: 1065-1076.
Slager J, Kjos M, Attaiech L, Veening JW. 2014. Antibiotic-Induced Replication Stress Triggers Bacterial Competence by Increasing Gene Dosage near the Origin. Cell 157: 395-406.
Sparling PF. 1966. Genetic Transformation of Neisseria gonorrhoeae to Streptomycin Resistance. Journal of Bacteriology 92: 1364-1371.
Spies FS, Ribeiro AW, Ramos DF, Ribeiro MO, Martin A, Palomino JC, Rossetti MLR, da Silva PEA, Zaha A. 2011. Streptomycin Resistance and Lineage-Specific Polymorphisms in Mycobacterium tuberculosis gidB Gene. Journal of Clinical Microbiology 49: 2625-2630.
Springer B, Kidas YG, Prammananan T, Ellrott K, Böttger EC, Sander P. 2001. Mechanisms of Streptomycin Resistance: Selection of Mutations in the 16S rRNA Gene Conferring Resistance. Antimicrobial Agents and Chemotherapy 45: 2877-2884.

19
Stewart GJ, Carlson CA. 1986. The Biology of Natural Transformation. Annual Review of Microbiology 40: 211-231.
Sunde M, Norström M. 2005. The genetic background for streptomycin resistance in Escherichia coli influences the distribution of MICs. Journal of Antimicrobial Chemotherapy 56: 87-90.
Thompson PR, Hughes DW, Wright GD. 1996. Mechanism of aminoglycoside 3'-phosphotransferase type IIIa: His188 is not a phosphate-accepting residue. Chemistry & Biology 3: 747-755.
van Hoek AHAM, Mevius D, Guerra B, Mullany P, Roberts AP, Aarts HJM. 2011. Acquired Antibiotic Resistance Genes: An Overview. Frontiers in Microbiology 2: 203.
Ventola CL. 2015. The Antibiotic Resistance Crisis. Pharmacy and Therapeutics 40: 277-283. Watanabe T, Fukasawa T. 1961. Episome-Mediated Transfer of Drug Resistance in
Enterobacteriaceae. Journal of Bacteriology 81: 669-678. World Health Organisation (WHO). Antimicrobial resistance. WWW-dokument 2017-11:
http://www.who.int/mediacentre/factsheets/fs194/en/. Hämtad 2017-11-20. World Health Organisation (WHO). 2015. Global Action Plan on Antimicrobial Resistance.
Rapport, World Health Organisation.

20
[Uppkomst och spridning av streptomycinresistens hos bakterier]: etisk bilaga Sofia Nordin Självständigt arbete i biologi 2017 Bör antibiotika användas på boskapsdjur? Antibiotika har de senaste årtiondena varit aktuell i flera debatter eftersom användning av dessa läkemedel har visats orsaka resistens hos bakterier. Detta skapar problem inom sjukvården, eftersom infektioner och sjukdomar blir svårare att behandla. Trots att resistensproblemen är välkända sker det fortfarande överanvändning och missbruk av antibiotika i flera länder, inte minst inom boskapsskötsel. Jag vill därför diskutera de argument som finns i debatten om användning av antibiotika på boskapsdjur. Pro-argument Det största pro-argumentet för användning av antibiotika på boskapsdjur är att kontinuerlig behandling av djuren håller dem friska. Antibiotikabehandlingar är ett billigt sätt för en djurägare att försäkra djurens hälsa eftersom sjuka boskapsdjur utgör en större kostnad. Dessutom lever behandlade djur oftast längre, vilket gör att de kan producera mer produkter (Phillips et al. 2004). Ett annat positivt argument är att antibiotika gör att djuren ökar i vikt i förhållande till den mängd föda de behöver. Detta gör det billigare att föda upp djuren då deras mat är den största utgiften för ägaren (Marshall & Levy 2011). Produkter från djur som behandlats med antibiotika anses även vara mer säkra eftersom skadliga bakterier som ofta finns i djuren, dör vid behandlingen. Om man förbjuder användandet av antibiotika på djur sker en ökning av kostnader för djurägare samt en ökning av patogena bakterier i djurmiljöer. I de länder där förbud införts har en ökning av antibiotika utskriva av veterinär skett. Detta eftersom läkemedlen behövs för att behandla sjuka djur. (Hao et al. 2014). Kontra-argument Det finns även kontra-argument inom denna debatt. Ett av dessa är att användning av antibiotika på djur medför att fler bakterier i djurens egna tarmflora blir resistenta. De resistenta bakterierna i djuret kan sedan överföras till människor via konsumtion av kött, ägg eller mjölk från djuret. Detta kan i sin tur orsaka allvarliga hälsorisker hos konsumenter (Ventola 2015). Ett annat kontra-argument som är orelaterat till resistensproblem är förekomsten av antibiotika i behandlade djurs exkrement. Avföring från djur används ofta för att producera biogas och det har visats att antibiotika påverkar biogasproduktionen negativt. Ett förbud på antibiotikaanvändning på djur har visats medföra en drastisk minskning av antalet resistenta bakterier. Ett förbud mot antibiotikan enrofloxacin i USA resulterade i en minskning av antalet infektioner i människor orsakade av resistenta bakterier (Hao et al. 2014). Diskussion Att komma fram till en slutsats i frågan om antibiotika bör användas på boskapsdjur är svårt eftersom båda sidor av debatten framför bra och faktabaserade argument. Pro-argumenten är visserligen fler, men kontra-argumenten väger i min åsikt tungt eftersom jag i min uppsats har diskuterat problemen med resistenta bakterier. Förmodligen behövs långtidseffekter av användningen av antibiotika på boskapsdjur studeras ytterligare för att en slutsats ska kunna konstateras.

21
Forskningsetik I min uppsats har jag primärt använt mig av artiklar från etablerade vetenskapliga tidskrifter. Jag har även använt mig av publicerade böcker med vetenskaplig grund. De källor jag har använt mig av anser jag vara högst trovärdiga. Jag har i min uppsats varit noggrann med att skilja på vad som är fakta samt vad som är mina respektive andras åsikter. Referenser Hao H, Cheng G, Iqbal Z, Ai X, Hussain HI, Huang L, Dai M, Wang Y, Liu Z, Yuan Z. 2014.
Benefits and risks of antimicrobial use in food-producing animals. Frontiers in Microbiology 5: 288.
Marshall BM, Levy SB. 2011. Food Animals and Antimicrobials: Impacts on Human Health. Clinical Microbiology Reviews 24: 718-733.
Phillips I, Casewell M, Cox T, De Groot B, Friis C, Jones R, Nightingale C, Preston R, Waddell J. 2004. Does the use of antibiotics in food animals pose a risk to human health? A critical review of published data. Journal of Antimicrobial Chemotherapy 53: 28-52.
Ventola CL. 2015. The Antibiotic Resistance Crisis. Pharmacy and Therapeutics 40: 277-283.
Recommended

















