
Hydrobiologia 186/187: 239-245, 1989.C. Ricci, T. W. Snell and C. E. King (eds), Rotifer Symposium V. 239© 1989 Kluwer Academic Publishers. Printed in Belgium.
Tasmania revisited: rotifer communities and habitat heterogeneity
R.J. Shiel', W. Koste2 & L.W. Tan3
'Murray-Darling Freshwater Research Centre, P.O. Box 921 Albury, N.S. W. 2640, Australia; 2LudwigBrill Strasse 5, Quakenbrack D-4570, Federal Republic of Germany; 3 Western Mining Corporation, P.O.Box 409 Unley, S.A. 5061, Australia
Key words: rotifers, Tasmania, dune lakes, endemism
Abstract
The results of four field surveys for Rotifera in Tasmania are summarized. Most new species and recordsin a 1987 survey were from acid waters (pH < 4.0) of dune lakes on the west coast (42 ° S). Marked intra-and interhabitat differences in rotifer communities of lakes and ponds were demonstrated by clusteranalysis and related to habitat heterogeneity.
Introduction
Our first survey of Tasmanian natural lakes on thecentral plateau (1980) provided 62 new rotiferrecords (one n. sp., Aspelta tilba). In 1984 thesurvey was extended to impoundments and arange of smaller ponds and stock dams, with 62further taxa (including n. spp. Lecane tasmanien-sis, Lepadella tyleri and Testudinella unicornuta)added to the known Rotifera (Koste & Shiel,1987a). In 1985, a third survey (100 sites, includ-ing all sites samples earlier), added 63 taxa(including Brachionus lyratus tasmaniensis, Cepha-lodella lindamaya, Lepadella tana and Testudinellamucronata tasmaniensis) (Koste & Shiel, 1986).
To provide seasonal information from the same100 localities, a fourth survey was made in spring1987, with the addition of eight new sites in aseries of dune lakes on the west coast of theisland. We had not investigated dune lakes previ-ously; work on the mainland (e.g. Timms, 1973)suggested that dune lakes were not productive ofRotifera. Indeed, in a review of these habitats,Timms (1986: 425) noted that rotifers seem to be
excluded by acidity; 'several taxa, including...rotifers... are rare or absent in acidic dune lakes'.Our attention was directed to these lakes by DrP.A. Tyler at the University of Tasmania, whohad sampled them for phytoplankton.
The 1987 survey resulted in 159 identified taxa,59 of these first records for the island (with Lecaneherzigi, Notommata tyleri, Trichotria buchneri andT. pseudocurta new) (Koste et al., 1988). Most ofthe new records came from the western plateauarea (Fig. 1). Some 250 taxa of Rotifera are nowknown from the island, and it is likely that only afraction of the rotifer fauna has been encountered.We summarize here the results of the four surveysto date, and discuss some of the peculiarities ofthe Tasmanian limnetic and littoral rotifer com-munities relative to those of the nearby mainland.
Sampling sites
All sites described in the earlier reports wereresampled in 1987. Habitats ranged from shallowroadside pools, ponds, turbid stock dams, to large
240
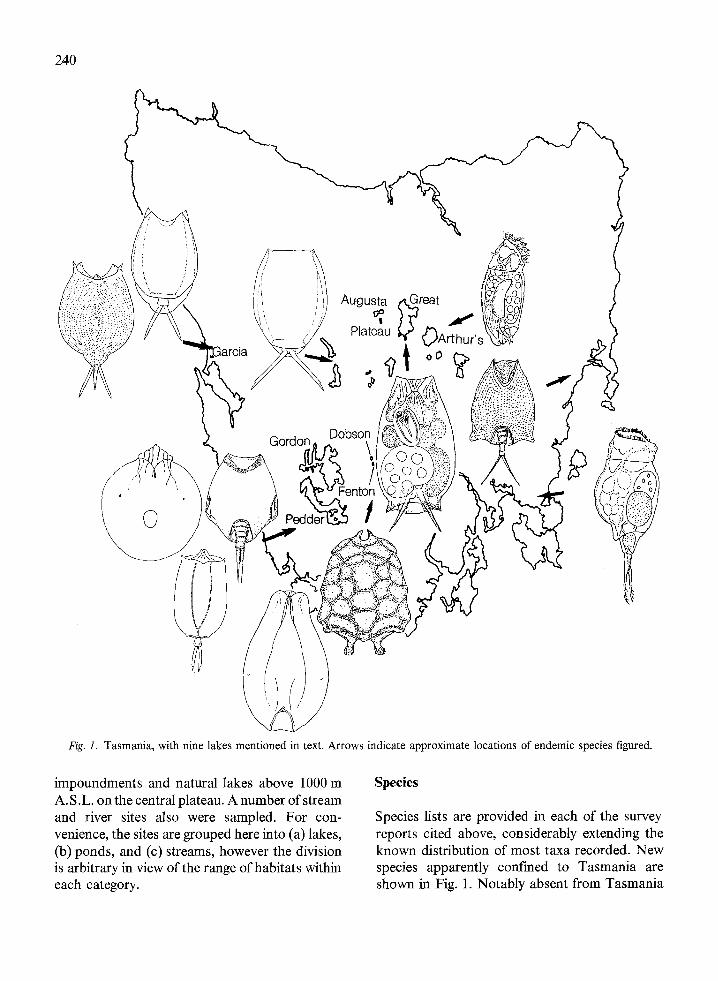
Fig. 1. Tasmania, with nine lakes mentioned in text. Arrows indicate approximate locations of endemic species figured.
impoundments and natural lakes above 1000 mA. S.L. on the central plateau. A number of streamand river sites also were sampled. For con-venience, the sites are grouped here into (a) lakes,(b) ponds, and (c) streams, however the divisionis arbitrary in view of the range of habitats withineach category.
Species
Species lists are provided in each of the surveyreports cited above, considerably extending theknown distribution of most taxa recorded. Newspecies apparently confined to Tasmania areshown in Fig. 1. Notably absent from Tasmania

241
are Brachionus species known from the mainland(28 taxa) (Koste & Shiel, 1987b). OnlyB. angularis was common in Tasmanian waters,with occasional occurrences of B. calyciflorus,B. quadridentatus and a local variant of B. lyratus(Fig. 1). The most abundant taxa in limneticassemblages were species of Keratella, with nineidentified and four (K. australis, K. cochlearis,K. procurva and K. slacki) widely distributed.Also abundant were species of Lecane (28 spp.),with four taxa common in the limnetic regions ofponds and stock dams (L. bulla, L.flexilis,L. hamata and L. lunaris), and an endemic(L. tasmaniensis) occurring widely in acid watersin the southwest. Trichocerca species (26) werewidespread, but only T. similis abundant.
Of 250 taxa recorded, 116 (48%) were foundonly once (or twice from the same locality in dif-ferent surveys). Such patchiness is generally inaccord with Australian mainland surveys (Shiel &Koste, 1986) and with studies reported elsewhere(Dumont, 1983). Approx. 20 % (49 spp.) have notbeen reported from the mainland; in most casesthese are acidophile species known previouslyfrom tropical regions of South America or SouthEast Asia, e.g. Testudinella ahlstromi, Trichocercabraziliensis (Koste, 1978). About half the recordedtaxa were of limited distribution (< 20 % of locali-ties). Only 10 species (4 %) occurred in > 20 % ofsamples (n = 267): K. slacki (38%), T. similis(35 %), K. cochlearis (32 %), K. australis/L. flexilis(30%), L. lunaris (28%), L. bulla (23%),B. angularis/Polyarthra vulgaris (21 %) and K. pro-curva (20 %).
Comparison of species distribution data fromthe Tasmanian series with those from a moreintensive sampling effort reported by Hillman(1986) from the R. Murray catchment on themainland suggests that the Tasmanian rotiferfauna is more diverse (Table 1), i.e. the R. Murraydata represent > 1500 samples from a300000 km2 area, the Tasmanian data 267samples from a 68000 km2 area. More speciesoccur in Tasmanian standing waters, with amarked decrease in streams possibly reflecting thesteeper gradients and greater turbulence atTasmanian sites. R. Murray gradients are gentler,
Table 1. Summary of rotifer recorded from Tasmanian andAustralian mainland habitats. Lakes includes artificialimpoundments; Ponds includes turbid stock dams.(R. Murray data after Hillman, 1986).
No. of species
Tasmania R. Murray(240) (229)
1. Lakes 148 1262. Ponds 189 174*3. Streams 50 734. Lakes and ponds 59 855. Lakes and streams 3 456. Ponds and streams 10 547. Lakes. ponds and streams 41 39
* Billabongs
the flows slower, and a complex rotifer planktonis maintained.
A simple plot of species numbers againstsampling frequency (Fig. 2) suggests that morerotifer species await discovery in Tasmania. Typi-cally, such plots produce an asymptote asrepeated surveys produce fewer unknown taxa.Our sample series has not reached an asymptote,although first records have declined (90 %, 47 %,32%, 28% for respective surveys). Given that wehave collected from only 100 of > 4000 availablesites, it is conceivable that more rotifer taxa ulti-mately will be recorded from Tasmania than pres-ently are known from the mainland (about 600species).
300-
o200
100
1980 1984 1985 19871980 1984 1985 1987
Fig. 2. Cumulative species numbers per sampling survey,Tasmania 1980-1987.

242
Community composition
Using Sokal's (1961) distance measure to com-pare species assemblages (number of taxa andproportion of each) across all sites in all years, themost striking feature of the rotifer assemblageswas the marked dissimilarity in species com-position between habitats, even within the samehabitat category. The whole data set is too largeto reproduce here, however two smaller subsetsare used below as examples.
(a) LakesIn summary, over the four surveys, the naturallakes on the central plateau had the most diverse
rotifer assemblages, although not necessarily themost species. The rotifer assemblages (1987survey) of ten sites (mean no.spp. site- ' = 10.4;mean diversity (H') = 2.31) are compared byaverage linkage cluster analysis in Fig. 3a. Rela-tive proximity of the lakes is shown in Fig. 1.
Although communities of the ten lakes wereranked of low similarity (0-22% taxa shared) byJaccard coefficient run concurrently with Sokal'sniche distance, clustering grouped lakes in closeproximity, e.g. Great Lake and Arthur's Lake,both dominated by Polyarthra vulgaris, and shar-ing Gastropus minor and Filinia longiseta from 16and 10 species, respectively. Similarly, Plateau(16 spp.) and Augusta (11 spp.) were both domi-
Sokal's niche distance
Fig. 3. Average linkage dendrograms (Sokal's niche distance) for (a) ten lake sites and (b) ten ponds/stock dams (indicated bysite #). 1987 survey.

243
nated by Synchaeta oblonga and shared Euchlanismeneta, Lepadella patella and Lecane flexilis.
Generally, the widely distributed taxa (K. coch-learis, P. vulgaris/dolichoptera, F. pejleri/longisetaand T. similis) comprised the shared species; theremaining taxa were specific to each lake. No twolakes had the same combination of temperature,pH and conductivity (within the range 7-14 C,pH 4.0-7.7, 9.0-33.0 t S cm - 1), suggesting otherdifferences in chemistry which may account inpart for the disparity of resident rotifer assem-blages by their influences on the algal and bac-terial communities.
Notably, a net tow from Lake Garcia, a dunelake first sampled in the 1987 survey (Fig. 1)(17.0 C, pH 4.3, 98.3 S cm- ) had 35 rotiferspecies, including 8 new records (3 n. spp., withone (Trichocerca) yet to be described) and thehighest diversity (H' = 4.37) yet recorded fromTasmania. The assemblage was a mixture of lim-netic and littoral taxa indicative of shalloweutrophic waters, including K. procurva (domi-nant), 5 spp. of Lecane and 8 of Trichocerca.
Seasonal changes in species compositionwithin each of the lakes were reflected in lowsimilarities between surveys, with only 1-3 of thecommon species present on successive visits. Inthe larger lakes, e.g. L. Pedder, there wereappreciable community differences at oppositeends of the lake on the same day. In L. Pedder inthe 1987 series (cf. Fig. 3a), community domi-nants at the eastern end of the lake wereK. cochlearis/Synchaeta pectinata/Conochilus dos-suarius, while at the western end, 12 km away,dominants were K. australis/F. pejleri/C. dossua-rius with only 20% of taxa shared.
(b) PondsSmaller standing waters commonly had 8-20tychoplanktonic species, with dominants one ormore species of Keratella (the most common inponds and stock dams was K. slacki, followed byK. australis > K. cochlearis > K. procurva >K. javana, with incidental occurrences of K. qua-drata, K. valga, K. hispida and K. tropica). Threesites had six co-occurring Keratella species. Other
abundant pond rotifers were species of Polyarthra,Lecane, Filinia, Trichocerca and Synchaeta.
As in the lake communities, there was littlesimilarity in inter-site rotifer species compositionon each survey. Fig. 3b shows clusters producedfor ten unnamed pond sites (mean no. spp.site-' = 17.3; mean H' = 3.25) selected fromwidely separated areas of the island i.e. 1977,1992, 1993 are north coast, 2004 and 2014 eastcoast, 2025 near L. Pedder, 2050 near Garcia andso on. Most obviously, tighter clustering suggeststhat the pond rotifer communities are more uni-form than those of the lakes, i.e. more species areshared. Again, the analysis grouped sites inclosest geographical proximity, e.g. 1992/1993and 2004/2014, with outliers (1977, 2032, 2045)the most isolated sites.
The most diverse pond rotifer assemblage(H' = 3.92) and greatest number of species (32)occurred in a small black-water humic roadsidepool in the dune lakes area (site 2050, near L.Garcia, Fig. 1). This pool had the lowest pH (3.1)of the eight sites in the dune series (pH range3.1-4.3). Dominant taxa were: K. javana, Lecanetasmaniensis, Synchaeta oblonga, Cephalodellamucronata and Trichotria buchneri. The highspecies diversity at this site accounts for theanomalous grouping with 2028 in Fig. 3b, whichwas not geographically close, but which also wasdominated by the acidophile K. javana, andshared other acidophile taxa (incl. L. flexilis andL. tasmaniensis) of 15 spp. present.
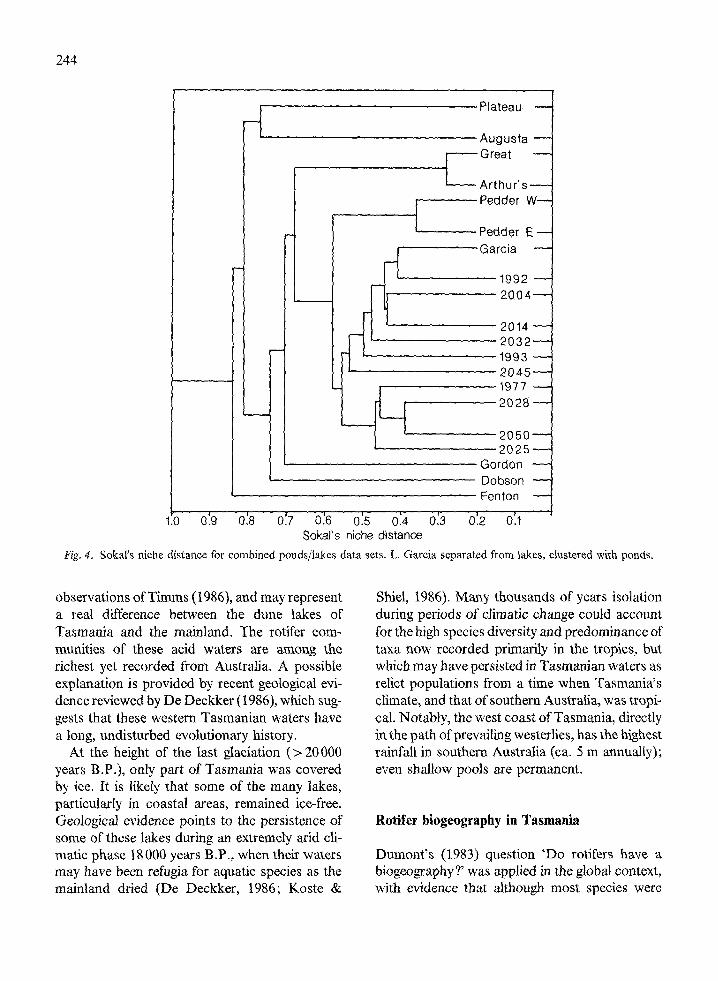
The ten pond data set was run against the tenlake set to compare the respective assemblages.The dendrogram derived (Fig. 4) maintains theseparation of all lakes except Garcia, which clus-tered with the pond assemblage, sharing five ofthe widely distributed taxa with site 1992, andsupported our impression of a mixed limnetic/littoral assemblage from the shallow margin ofthis lake.
The dune lakes
The unexpectedly high species richness of thedune lake series is clearly in contrast with the
09 0:8 0O7 0:6 0.5 0:4Sokal's niche distance
0.3 02 0:1
Fig. 4. Sokal's niche distance for combined ponds/lakes data sets. L. Garcia separated from lakes, clustered with ponds.
observations of Timms (1986), and may representa real difference between the dune lakes ofTasmania and the mainland. The rotifer com-munities of these acid waters are among therichest yet recorded from Australia. A possibleexplanation is provided by recent geological evi-dence reviewed by De Deckker (1986), which sug-gests that these western Tasmanian waters havea long, undisturbed evolutionary history.
At the height of the last glaciation (>20000years B.P.), only part of Tasmania was coveredby ice. It is likely that some of the many lakes,particularly in coastal areas, remained ice-free.Geological evidence points to the persistence ofsome of these lakes during an extremely arid cli-matic phase 18 000 years B.P., when their watersmay have been refugia for aquatic species as themainland dried (De Deckker, 1986; Koste &
Shiel, 1986). Many thousands of years isolationduring periods of climatic change could accountfor the high species diversity and predominance oftaxa now recorded primarily in the tropics, butwhich may have persisted in Tasmanian waters asrelict populations from a time when Tasmania'sclimate, and that of southern Australia, was tropi-cal. Notably, the west coast of Tasmania, directlyin the path of prevailing westerlies, has the highestrainfall in southern Australia (ca. 5 m annually);even shallow pools are permanent.
Rotifer biogeography in Tasmania
Dumont's (1983) question 'Do rotifers have abiogeography?' was applied in the global context,with evidence that although most species were
244
Plateau -
C--- -Augusta -
Great
Arthur's -Pedder W-
Pedder E-Garcia -
1992 -200 4-
20142032--
- 1993 -2045-1977 2028 -
2050-2025 -
Gordon -Dobson -IFenton
FI
I
I
''1.U

245
cosmopolitan or widely distributed, some wereremarkably restricted. Our Tasmanian surveysmust be regarded as preliminary, however it isapparent that Tasmanian rotifers have their ownbiogeography.
If an arc is drawn on Fig. 1 approximately fromGreat Lake southwest through Aspelta tilba, eastof Lakes Dobson and Fenton to B. lyratustasmaniensis, the eastern margin of the centralplateau is demarcated. We have named this arc'Tyler's Line'. It separates two broad rotiferassemblages: west of Tyler's Line, predominantlyacidophiles, contains 11 of the 13 endemic taxaand most of the species which have not beenrecorded from the mainland. Waters are dark-black humic acid type, low in electrolytes, andpresumably differ markedly in characteristicssuch as nutrients and phytoplankton, and rotifercomposition from those east of the line. To theeast, which is a rainshadow area, waters are lesshumic, some are alkaline, and electrolytes arehigher, including some saline and hypersalinewaters on the east coast, i.e. they reflect geologicaland climatic differences between the two areas.
The heterogeneity of rotifer assemblages acrossthe island appears to be a response to local, evenmicrohabitat, variations in their requirements.
Acknowledgements
In 1987 survey was funded by the AustralianBiological Resources Study. We thank Peter Tylerfor help with field work, and for hospitality anduse of facilities at Hobart. Initial MS preparation
was at the Botany Department, University ofAdelaide; thanks to the then Chairman, GeorgeGanf, for access to facilities. Particular thanks toTerry Hillman, MDFRC, for cluster analysis, andto Kim Jenkins for graphics.
References
De Deckker, P., 1986. What happened to the Australianaquatic biota 18 000 years ago? In P. De Deckker & W. D.Williams (eds) Limnology in Australia. CSIRO & Dr W.Junk BV, Melbourne and Dordrecht: 487-496.
Dumont, H. J., 1983. Biogeography ofrotifers. Hydrobiologia104: 19-30.
Hillman, T. J., 1986. Billabongs. In P. De Deckker & W. D.Williams (eds) Limnology in Australia. CSIRO & Dr W.Junk BV, Melbourne and Dordrecht: 457-470.
Koste, W., 1978. Rotatoria. Die Radertiere Mitteleuropas.Bestimmungswerk begriindet von Max Voigt. 2 vols.Borntraeger, Stuttgart.
Koste, W. & R. J. Shiel, 1986. New Rotifera (Aschelminthes)from Tasmania. Trans. r. Soc. S. Aust. 110: 93-109.
Koste, W. & R. J. Shiel, 1987a. Tasmanian Rotifera: affinitieswith the Australian fauna. Hydrobiologia 147: 31-43.
Koste, W. & R. J. Shiel, 1987b. Rotifera from Australianinland waters. II. Epiphanidae and Brachionidae (Rotifera:Monogononta). Inv. Taxon. 1: 949-1021.
Koste, W., R. J. Shiel & L.W. Tan, 1988. New rotifers(Rotifera) from Tasmania. Trans. r. Soc. S. Aust. 112 (inpress).
Shiel, R. J. & W. Koste, 1986. Australian Rotifera: ecologyand biogeography. In P. De Deckker & W. D. Williams(eds) Limnology in Australia. CSIRO & Dr W. Junk BV,Melbourne and Dordrecht: 142-150.
Sokal, R. R., 1961. Distance as a measure of taxonomicsimilarity. Syst. Zool. 10: 71-79.
Timms, B. V., 1973. A limnological survey of the freshwatercoastal lakes of east Gippsland, Victoria. Aust. J. mar.Freshwat. Res. 24: 1-20.
Timms, B. V., 1986. The coastal dune lakes of easternAustralia. In P. De Deckker & W. D. Williams (eds)Limnology in Australia. CSIRO & Dr W. Junk BV,Melbourne and Dordrecht: 421-432.
Recommended

















