
1
PODŁOśA MIKROBIOLOGICZNE
podstawowe cechy podłoŜy:
• źródło pierwiastków biogennych• o odpowiednim pH, izotoniczne• klarowne• STERYLNE
poŜywka mikrobiologiczna moŜe być:
• uniwersalna• wybiórcza• róŜnicująca
• płynna• półpłynna (0.6% agar-agar)• stała (2% agar-agar)
Escherichia colii Micrococcus luteus
Wzrost mikroorganizmów na uniwersalnym podłoŜu stałym
MoŜemy odróŜnić te bakterie na uniwersalnym podłoŜu, gdyŜ jedna z nich,
Micrococcus luteus, wytwarza Ŝółty barwnik
PodłoŜa róŜnicujące – podłoŜe MacConkey’a
Escherichia colii Salmonella enteritidis
Na tym podłoŜu bakterie fermentujące laktozę (Escherichia
coli) produkuj ą kwasy organiczne, indykator pH zmienia kolor i
bakterie te rosną w postaci
czerwonychkolonii
Bakterie nie fermentujące laktozy(Salmonella enteritidis) rozkładają pepton obecny w podłoŜu bez zmiany pH i rosną w postaci
białychkolonii
hodowla bakterii
zbyt duŜobakterii dopoliczenia
liczba kolonii
9 ml bulionu
rozcieńczenie
1 ml próbki na płytki
Określanie ilości mikroorganizmów –posiew na podłoŜe stałe
Trzeba jednak pamiętać, Ŝe
• w skład mikroflory dorosłego człowieka
wchodzi ponad 400 gatunków mikroorganizmów,
z których 60% nie umiemy hodować w laboratorium
• w laboratorium umiemy hodować tylko około
5% mikroorganizmów obecnych w środowisku
Zalety bycia niewielkim
• szybsza wymiana substancji odŜywczych
i szkodliwych produktów metabolizmu• większe tempo wzrostu
• większe rozmiary populacji• większa zdolność akumulacji mutacji –
szybsza adaptacja do zmieniającego się środowiska
Istnieje jednak dolna granica wielkości komórki
około 0.2 µµµµm
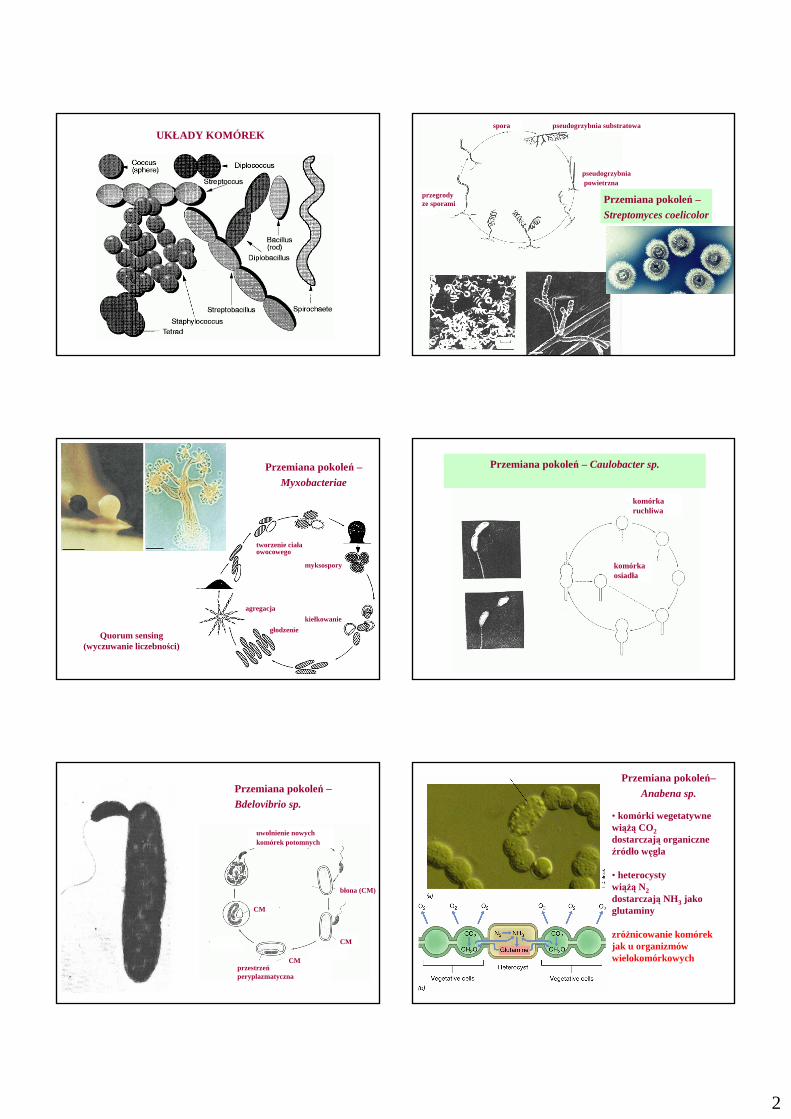
2
UKŁADY KOMÓREKpseudogrzybnia substratowa
pseudogrzybniapowietrzna
przegrody ze sporami
spora
Przemiana pokoleń –Streptomyces coelicolor
Przemiana pokoleń –Myxobacteriae
myksospory
kiełkowanie
głodzenie
agregacja
tworzenie ciałaowocowego
Quorum sensing(wyczuwanie liczebności)
Przemiana pokoleń – Caulobacter sp.
komórka ruchliwa
komórka osiadła
Przemiana pokoleń –Bdelovibrio sp.
błona (CM)
przestrzeń peryplazmatyczna
CM
uwolnienie nowych komórek potomnych
CM
CM
Przemiana pokoleń–Anabena sp.
• komórki wegetatywnewiąŜą CO2dostarczają organiczne źródło węgla
• heterocystywiąŜą N2dostarczają NH3 jakoglutaminy
zróŜnicowanie komórekjak u organizmów wielokomórkowych

3
Budowa komórki prokariotycznej
błona cytoplazmatyczna
ściana komórkowa
nukleoid
cytoplazmai rybosomy
Lipidy błonowe u bakterii
• kwasy tłuszczowe nierozgałęzione,• rzadko wielonasycone• brak steroli, są hopanoidy
• brak kwasów tłuszczowych• są alkohole poliizoalkilowe,często rozgałęzione
• czasem obecne sterole
i archae
wiązanie estrowe
wiązanie ETEROWE
hopanoid
cholesterol
monowarstwa
Lipidy u archae
dwuwarstwadwuetery glicerolu
czteroetery diglicerolu
Peptydoglikan (mureina) u bakterii
N-acetylo-glukozamina
kw.N-acetylomuraminowy
L-alanina
kw. D-glutaminowy
kw. mezo-diaminopimelinowy
D-alanina
(ββββ1,4)
Archae:pseudomureina
• wiązanie (ββββ1,3)
• brak form D aa
Tworzy rodzaj siatkiwokół komórki
Ściana komórkowa bakterii gramdodatnich
peptydoglikan
błona cytoplazmatyczna
błona cytoplazmatyczna
peptydoglikan
kwasylipotejchojowe
kwasy tejchojowebiałka związane ześcianą komórkową
Struktura i funkcje kwasów tejchojowych
• polisacharydy o negatywnym ładunku
• nadają komórce negatywny ładunek(ochrona przed toksycznymi związkami hydrofobowymi)
• wiąŜą dwuwartościowe jony(Ca2+ i Mg2+)

4
Ściana komórkowa bakterii gramujemnych
błona cytoplazmatyczna
peptydoglikanbłona zewnętrzna
błonawewnętrzna
błonazewnętrzna
LPS
poryna
lipoproteina
peptydoglikan
przestrzeńperyplazma-tyczna
Lipopolisacharyd (LPS)
lipid Ałańcuch O-specyficzny rdzeń
Błona zewnętrzna bakterii gramujemnych jest asymetryczna
• wewnętrzna warstwa składa się z fosfolipidow
• zewnętrzna warstwa zbudowana jest głównie
z lipopolisacharydu (LPS)
lipopolisacharyd
fosfolipidy
fosfolipidy
Escherichia coli (G-) i Staphylococcus aureus (G+)
zalewamy utrwalony preparat fioletemkrystalicznym na 1 minutę
dodajemy płynu Lugolana 1 minutę
odbarwiamyetanolem
dobarwiamy fuksynąprzez 30 sekund
Barwienie Grama
Zmiana kierunku ruchu – urz ęsienie peritrichalne
obrót rzęsek w leworuch komórki
obrót rzęsek w prawo„koziołkowanie”
obrót rzęsek w leworuch komórki
nowy kierunek ruchu jest przypadkowy
Chemotaksjakierunek ruchu losowy
ukierunkowany ruch• czas ruchu w kierunku atraktanta dłuŜszy• czas ruchu w kierunku przeciwnym do atraktanta krótszy
ATRAKTANT
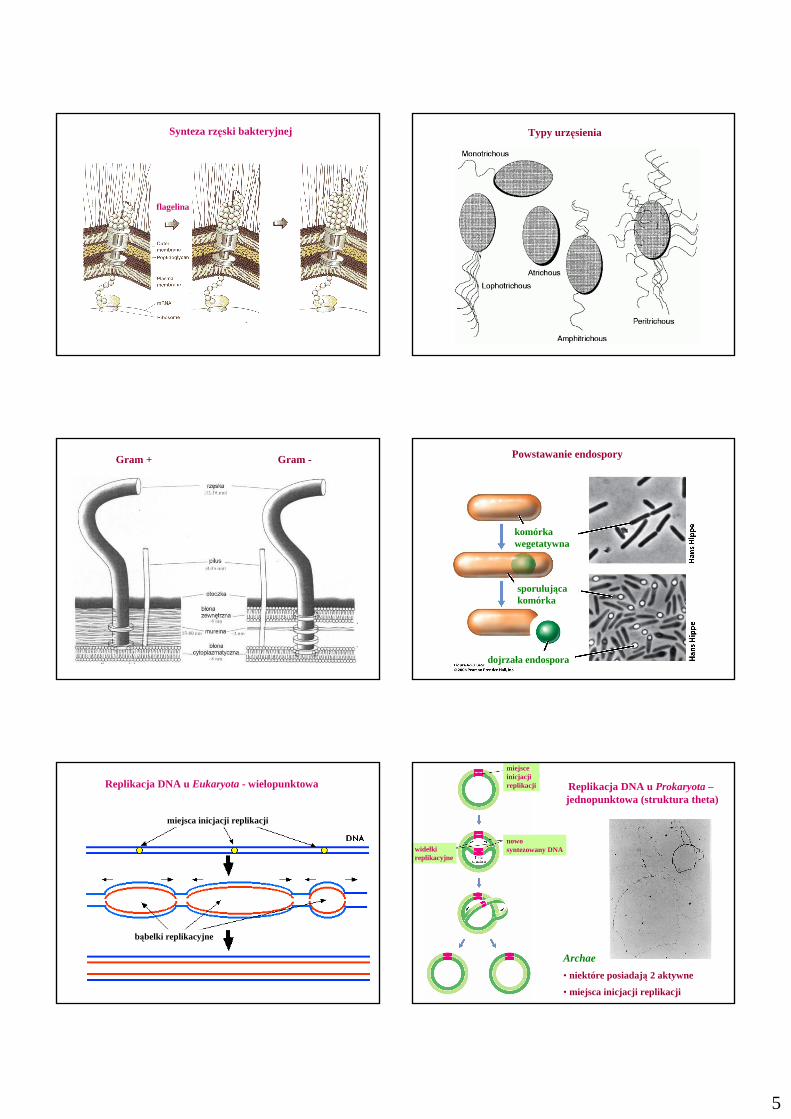
5
Synteza rzęski bakteryjnej
flagelina
Typy urzęsienia
Gram + Gram - Powstawanie endospory
komórkawegetatywna
sporulująca komórka
dojrzała endospora
Replikacja DNA u Eukaryota- wielopunktowa
miejsca inicjacji replikacji
bąbelki replikacyjne
Replikacja DNA u Prokaryota–jednopunktowa (struktura theta)
Archae
• niektóre posiadają 2 aktywne
• miejsca inicjacji replikacji
widełkireplikacyjne
nowo syntezowany DNA
miejsceinicjacjireplikacji

6
Podział komórkibakteryjnej
• z jednej komórki powstają dwie identyczne
(jedno pokolenie)
• czas podziału róŜny -
minimum 20 minut dlaE. coli
przegroda
Fazy wzrostu populacji bakteryjnej
wzrost – zwiększenie liczby komórek w populacji
stacjonarnazamieranialag
wykładnicza
czas
gęsto ść optyczna
liczba Ŝywychkomórek
• faza lag– czas potrzebny komórkom na adaptację do nowych warunków środowiska; bardzo wolny wzrost populacji
• faza wykładnicza– większość komórek w populacji dzieli się
dając początek dwóm komórkom; faza najszybszego, równomiernego wzrostu
• faza stacjonarna– wzrost spowalnia się w miarę wyczerpania ilości składników odŜywczych i nagromadzenia toksycznych
produktów przemiany materii; niektóre komórki gin ą, więc ilośćkomórek w populacji jest stała
• faza zamierania– wzrost ustaje; coraz więcej komórek
w populacji obumiera
Hodowla okresowa – fazy wzrostuFaza stacjonarna
• najczęściej występuje w środowisku
• komórki znajduj ą się w odmiennym stanie fizjologicznym
podobnym do stanu komórek rosnących pod postacią BIOFILMU(wolne tempo wzrostu, duŜa odporność na niekorzystne warunki środowiska)
FtsZ – homolog tubuliny
• homologia 10-18% na poziomie sekwencji aminokwasów• podobieństwo struktury trzeciorzędowej• polimeryzacja zaleŜna od GTP• występuje:
• u większości bakterii i archae• u eukaryota w plastydach • w mitochondriach niektórych eukaryota
MreB, Mbl, ParM – homologi aktyny
• homologia ok.15% na poziomie sekwencji aminokwasów
• podobieństwo struktury trzeciorzędowej
• polimeryzacja zaleŜna od ATP
• rola:rozdział DNA plazmidowego i chromosomowegodeterminacja kształtu komórki

7
Wielkość genomu
Eukaryota• 3 000 kb (2000)mikrosporidia
• 12 000 (6000)Saccharomyces cerevisiae
• 3 000 000 kb (30-35 tys)człowiek
Prokaryota
• 600 kb(500)Mycoplasma sp.
• 4700 kb(4000)Escherichia coli
• 8000 kb(8000)Mezorhizobium loti
• maksymalnie 12 000 kb
ok. 40% OFR o funkcji nieznanejMycobacterium tuberculosis
zawartość GC26% 76%
Mycoplasma sp. Micrococcus sp.
Ilość róŜnych chromosomów w komórce•Vibrio sp. 2•Agrobacterium sp. 3
Borelia burgdorferii1 chromosom liniowy5 plazmidów• 3 liniowe (160-500 kb)• 2 koliste (270 i 300 kb)
Rhodobacter sphaeroideschromosom I (3 045 kb)chromosom II (914 kb)4 plazmidy po 100 kb1 plazmid 42 kb
Ilość kopii chromosomu w komórce•Escherichia coli maks. 4•Azotobacter vinelandii 4-40, maks. 80•Methanococcus sp. 3-15
Eukaryota1n, 2n do 500 w niektórych tkankach roślin
otwarte koło
superskręcone
białka
superskręconadomena
Chromosom bakteryjny –kolisty
liniowe:•Borelia sp.
•Streptomyces sp.
•Agrobacterium sp.
(1 liniowy i 1 kolisty)
Chromosom Eukaryota
u Archae
•homologi histonów H1, H3 i H4
• tetramery histonowe tworzą nukleosom
• brak „ogonków” histonowych
• występują warianty histonów
3 drogi wymiany informacji genetycznej u Prokaryota:
• transdukcja – bakteriofagi
• koniugacja – plazmidy koniugacyjne
• transformacja – bezpośrednio ze środowiska
3% genów wolno Ŝyjących bakterii pochodzi od Archae lub Eukaryota
nawet do 24% genów u bakterii hypertermofilnych pochodzi od hypertermofilnych Archae
Liczenie bakteriofagów
łysinki
wylewamy mieszankębakterii i bakteriofaga na plytk ęz poŜywką dla bakterii
na murawie rosnącychbakterii obserwujemy„łysinki”
inkubujemy w cieplarceprzez noc

8
Cykl lityczny – bakteriofag T4
cząstka transdukuj ąca
cząstka transdukuj ąca
homologiczna rekombinacja integracja DNA dogenomu biorcy
cząstka fagowa
do komórek biorcy przekazywany jest dowolny fragment genomu dawcy
transdukcja ogólna
Cykl lizogenny (infekcja) – bakteriofag λλλλ
cząstka fagowa
cykl lityczny cykl lizogennyy
integracja faga dogenomu biorcy
profag
podział komórki lizogennej
Cykl lizogenny (indukcja) –bakteriofag λλλλ
komórka lizogenna
INDUKCJA
często rzadko
• część DNA dawcy (czerwony)wydziela się wraz z DNA faga
• namnaŜanie cząsteczek transdukuj ących
• do następnych komórek przekazywane zawsze te samefragmenty DNA dawcy
transdukcja specyficzna
Koniugacja
• plazmid wbudowany w chromosom
tzw. szczepy Hfr
• przekazywany DNA chromosomu bakterii
• plazmid wolny w cytoplazmie
• przekazywany DNA plazmidu
Koniugacja
chromosom
DAWCA BIORCApilus
plazmid
• jedna nić DNA plazmidunacinana i przekazywanado biorcy
• komplementarna nić DNA dosyntezowywana zarównou biorcy jak i dawcy
• po transferze OBIE komórkizawierają cząsteczkę plazmidu
Gen C przekazywany pierwszy CW
Gen L przekazywany pierwszy CCW
Gen X przekazywany pierwszy CW
Gen C przekazywany pierwszy CCW
• plazmid wbudowanyw chromosom tzw. szczepy Hfr
• do dawcy przekazywane sągeny chromosomowe
• geny plazmidowe przechodząna końcu

9
Koniugacyjna mapa chromosomu E.coliTransformacja
• komórka dawcy DNA obumiera
• DNA uwalniany jest do środowiska
• komórka kompetentna pobiera DNAdo swego wnętrza
geny z jednej komórki są przekazywanedo drugiej bez kontaktu komórek
i bez pośrednictwa wirusów
• nie wszystkie gatunki bakterii mogą pobierać DNA ze środowiska
• stan kompetencji moŜna wywołać sztucznie w laboratorium
Transformacja - Bacillus subtilis
•kompetencja indukowanaprzez dwa feromony
9-10 aa oligopeptydpentapeptyd
•pobierana jedna nić DNA,druga degradowana
Quorum sensing – wykrywanie liczebności
peptydylaktony homoserynowe
produkcja autoinduktora
wykrywanie autoinduktora
indukcja transkrypcjiwydzielanie efektorów• enzymów, • antybiotyków, • sideroforów
Trends in Microb.(2002) 10: 365
• ruch i chemotaksja• wydzielanie enzymów degradacyjnych• wydzielanie antybiotyków• stan kompetencji, pobieranie DNA• sporulacja
Adaptacja komórek Bacillus sp.do zmiennych warunków środowiskaProkaryota Eukaryota
NIERESTRYKCYJNY RESTRYKCYJNY
stan aktywny
stan przygotowania
STANPODSTAWOWY
stan represowany
stan wyciszenia
Pro-• promotory silne• regulacja przez represję
Eu-• promotory słabe• regulacja przez aktywację(zmiana struktury chromatyny)
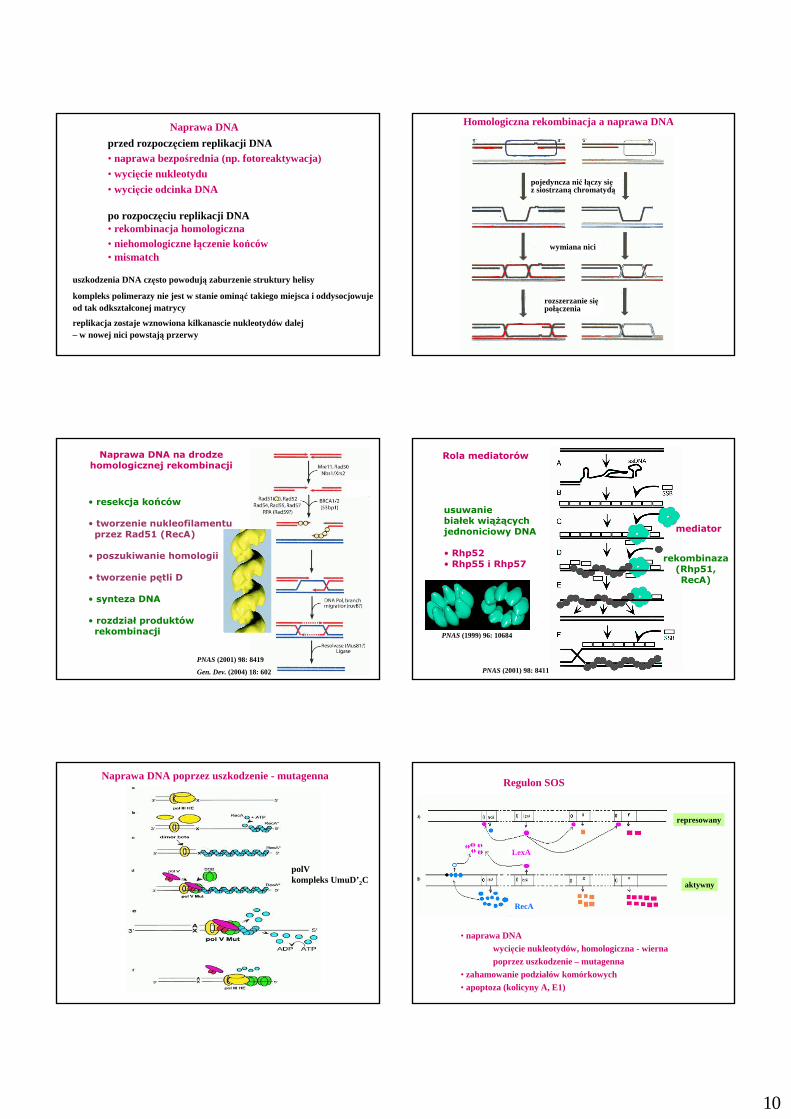
10
Naprawa DNA
przed rozpoczęciem replikacji DNA• naprawa bezpośrednia (np. fotoreaktywacja)• wycięcie nukleotydu• wycięcie odcinka DNA
po rozpoczęciu replikacji DNA• rekombinacja homologiczna• niehomologiczne łączenie końców• mismatch
uszkodzenia DNA często powodują zaburzenie struktury helisy
kompleks polimerazy nie jest w stanie ominąć takiego miejsca i oddysocjowuje od tak odkształconej matrycy
replikacja zostaje wznowiona kilkanascie nukleotydów dalej – w nowej nici powstają przerwy
Homologiczna rekombinacja a naprawa DNA
wymiana nici
pojedyncza nić łączy się z siostrzaną chromatydą
rozszerzanie się połączenia
Naprawa DNA na drodze homologicznej rekombinacji
• resekcja końców
• tworzenie nukleofilamentuprzez Rad51 (RecA)
• poszukiwanie homologii
• tworzenie pętli D
• synteza DNA
• rozdział produktów rekombinacji
Gen. Dev.(2004) 18: 602
PNAS(2001) 98: 8419
Rola mediatorów
usuwanie białek wiąŜącychjednoniciowy DNA
• Rhp52• Rhp55 i Rhp57
mediator
rekombinaza(Rhp51,RecA)
PNAS(2001) 98: 8411
PNAS(1999) 96: 10684
Naprawa DNA poprzez uszkodzenie - mutagenna
polVkompleks UmuD’2C
Regulon SOS
aktywny
represowany
LexA
RecA
• naprawa DNA
wycięcie nukleotydów, homologiczna - wiernapoprzez uszkodzenie – mutagenna
• zahamowanie podziałów komórkowych• apoptoza (kolicyny A, E1)

11
Antybiotyki – spektrum działania
tetracykliny
penicyliny
polimyksynyny
tobramycyna
sulfonamidycefalosporyny
streptomycyna
izonazid
prątki Gram - Gram + chlamydie riketsje
Antybiotyki – mechanizm działania
ściana komórkowabłona komórkowa
hamowanie syntezyściany komórkowejpenicylina, wankomycyna
zakłócenie funkcjibłony komórkowejpolimyksyny
hamowanie syntezy białkastreptomycyna,erytromycyna,tetracyklina, chloramfenikol
hamowanie syntezykwasów nukleinowychchinolowe (replikacja)
rifamycyna (transkrypcja)
antymetabolitysulfonamidy,trimetoprim
fermentacje
oddychanie
Degradacja polimerów -zewnątrzkomórkowa
proteazy – Bacillus, Clostridium, Proteus, Pseudomonas spp.
alkaliczne, neutralne, kwaśne
specyficzne (np. typu reniny) i niespecyficzne
nukleazy
deoksyrybonukleazy –Clostridium sp., Staphylococcus aureus,hemolityczne paciorkowce
rybonukleazy –Bacillus subtilis, Penicillium citrinum(RNazaP)
lipazy
enzymy degradującepolisacharydy
Rola:• patogenność• biotechnologia
enzymy lub produkty ich działania
Fosforylacja oksydacyjna – łańcuch oddechowy
• w wyniku transportu elektronow na łańcuchu oddechowymi wyrzucaniu H+ na zewnatrz komórki powstaje potencjał w poprzek błony
• zredukowany NADH zostaje zregenerowany do NAD+
• ATP powstaje pośrednio, w wyniku działania ATP-az błonowych, wnikanie H+ do wnętrza komórki dostarcza energii
Fosforylacja oksydacyjna –róŜnice w potencjale oksydoredukcyjnym
para red-ox
fumaran bursztynian

12
rozmieszczenie elementów łańcuchaw błonie cytoplazmatycznej
mitochondria E.coli Paracoccus Micrococcus denitryficans luteus
ODDYCHANIE AZOTANOWE(dysymilacyjna redukcja azotanów)
1. denitryfikacja - Bacillus, Pseudomonas spp., Paracoccus denitrificans
glu + 4.8 NO3- + 4.8 H+ → 6 CO2 + 2.4 N2 + 8.4 H2O
Go’ = -2669 kJ(-638 kcal)
dla porównania:
glu + 6O2 → 6 CO2 + 6H2O Go’ = -2870 kJ(-686 kcal)
2. Escherichia coli, Klebsiella sp., Staphylococcus sp.
glu + 12 NO3- → 6 CO2 + 12 NO2
- + 6 H2O
Go’ = -1766 kJ(-422 kcal)
u niektórych mikroorganizmów z tej grupy zachodzi proces:
glu + 3 NO3- + 3 H+ → 6 CO2 + 3 NH3 + 3 H2O
Go’ = -1796 kJ(-429 kcal)
Fosforylacja substratowa
• ATP powstaje bezpośrednio, sprzęŜone jest z utlenianiem substratu
• powstaje zredukowany NADH
część wykorzystywana do biosyntezy
część musi być zregenerowana by odnowić pulę NAD+ do utleniania następnych cząsteczek substratu
• brak energizacji błony komórkowej
HMP EMP ED
Rozkładglukozy do pirogronianu DEKARBOKSYLACJA PIROGRONIANU
1. Eukaryota, bakterie tlenowepirogronian + CoA + NAD+ →→→→ acetylo-CoA + NADH + H+ + CO2
(kompleks dehydrogenazy pirogronianu)
2. Archebakterie, bakterie beztlenowepirogronian + CoA + 2Fd →→→→ acetylo-CoA + 2FdH + CO2 (oksydoreduktaza
pirogronianu )
U Archaebacteriae zamiast ferredoksyny (Fd) występuje koenzym F420.
3. Fermentacja mieszana pałeczek jelitowychpirogronian + CoA →→→→ acetylo-CoA + mrówczan (liaza pirogronianowa)
4. Fermentacja alkoholowa droŜdŜy i Zygomonas sp.pirogronian →→→→ acetaldehyd + CO2 (dekarboksylaza pirogronianu)
5. Bacillus sp., Enterobacteriaceae
2 cz. pirogronianu →→→→ acetomleczam + CO2 (syntaza acetomleczanu)

13
Znaczenia słowa FERMENTACJA
1. KaŜdy proces, tlenowy lub beztlenowy, w którym na duŜą skalę wykorzystuje się hodowlę mikroorganizmów.
2. KaŜdy proces biologiczny zachodzący pod nieobecność tlenu.
3. Psucie się poŜywienia.
4. Produkcja napojów alkoholowych.
5. UŜywanie organicznego substratu jako dawcy i akceptora elektronów.
6. UŜywanie organicznego substratu jako czynnika redukującego i tego samego, częściowo rozłoŜonego organicznego substratu jako czynnika utleniającego (akceptora elektronów).
7. Wzrost zaleŜny od fosforylacji na poziomie substratu.
Homofermentacja mlekowa
FERMENTACJA MLEKOWA
homofermentacja
glukoza →→→→ 2 cz. mleczanu
(Lactobacillus lactis, L.delbrueckii, L. bulgaricus, L.casei,Streptococcus faecalis, S.cremoris, S.lactis, Pediococcus damnosus)
heterofermentacja
glukoza →→→→ mleczan + etanol + CO2
(Lactobacillus brevis, L.fermentum, Leuconostoc mesenteroides,
L.dextranicum)
szlak bifidum
2 glukoza →→→→ 3 cz. octanu + 2 cz. mleczanu
(Bifidobacterium bifidum)
Udział mikroorganizmów w przetwarzaniu mleka
kefir Lactococcus lactis, Lactobacillus bulgaricus, Saccharomyces spp.
jogurt Streptococcus thermophilus, L.bulgaricus
masło Lactococcus diacetylactis, Leuconostoc cremoris
sery
Roquefort L.lactis, L.cremoris, Penicillium roqueforti
Camembert L.lactis, L.cremoris, Penicillium camamberti,
Brevibacterium linens
Gouda L.lactis, L.diacetylactis, L.cremoris
Ementaler S. thermophilus, L.lactis, Propionibacterium shermani
L.helveticus, P.freudenreichii
Fermentacja alkoholowa
dekarboksylaza pirogronian/aldehyd octowyrzadka u bakterii
ENTEROBACTERIACEAE
Escherichia coli komensal przewodu pokarmowego ssaków Salmonella sp.Shigella sp. patogeny człowieka
Enterobacter sp. występują w glebie i wodzieSerratia sp.Proteus sp.Erwinia sp. patogeny roślin
Yersinia sp. patogeny człowieka
Vibrio sp. występują w wodzie, niektóre patogenneAeromonas sp.Photobacterium sp.

14
Fermentacja mieszana -produkty kwa śne
Fermentacja mieszana -produkty oboj ętne
IMViC
+ + - - - - + +
Fermentacja masłowa
ATP
ATP
CO2
CO2
H2
etanol
aceton
butanol
octan
maślan
Fermentacja masłowa Toksyczność tlenu
wynika z obecności w komórce :
enzymów flawinowych
oksydaz (np. oksydaza NADPH)barwników uczulających na światło (np. chlorofil, cytochromy)
funkcj ę ochronna pełnią:
dysmutazy ponadtlenkowe O2- + 2H+ →→→→ H2O2
katalazy 2H2O2 →→→→ 2H2O + O2
karotenoidy
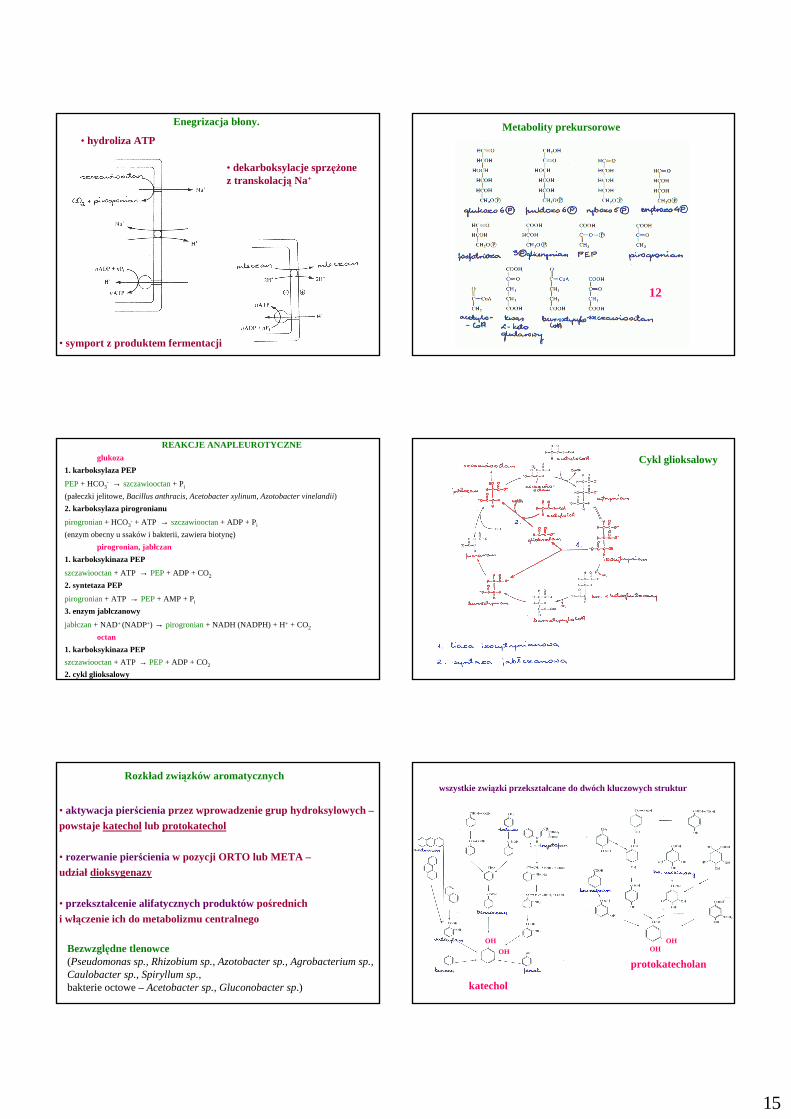
15
Enegrizacja błony.
• dekarboksylacje sprzęŜone z transkolacją Na+
• symport z produktem fermentacji
• hydroliza ATPMetabolity prekursorowe
12
REAKCJE ANAPLEUROTYCZNEglukoza
1. karboksylaza PEP
PEP+ HCO3- → szczawiooctan+ Pi
(pałeczki jelitowe,Bacillus anthracis, Acetobacter xylinum, Azotobacter vinelandii)
2. karboksylaza pirogronianu
pirogronian+ HCO3- + ATP → szczawiooctan+ ADP + Pi
(enzym obecny u ssaków i bakterii, zawiera biotynę)
pirogronian, jabłczan
1. karboksykinaza PEP
szczawiooctan+ ATP → PEP+ ADP + CO2
2. syntetaza PEP
pirogronian+ ATP → PEP+ AMP + Pi
3. enzym jabłczanowy
jabłczan+ NAD+ (NADP+) → pirogronian+ NADH (NADPH) + H+ + CO2
octan
1. karboksykinaza PEP
szczawiooctan+ ATP → PEP+ ADP + CO2
2. cykl glioksalowy
Cykl glioksalowy
Rozkład związków aromatycznych
• aktywacja pierścienia przez wprowadzenie grup hydroksylowych –
powstaje katechollub protokatechol
• rozerwanie pierścieniaw pozycji ORTO lub META –udział dioksygenazy
• przekształcenie alifatycznych produktówpośrednich
i włączenie ich do metabolizmu centralnego
Bezwzględne tlenowce(Pseudomonas sp., Rhizobium sp., Azotobacter sp., Agrobacterium sp.,Caulobacter sp., Spiryllum sp.,bakterie octowe –Acetobacter sp., Gluconobacter sp.)
wszystkie związki przekształcane do dwóch kluczowych struktur
protokatecholan
katechol
OH
OH
OHOH

16
ORTO
META
dioksygenaza
O2 O2
O2 O2
Rozkład węglowodorów
• metan –metanotrofy
• do C8 –niewiele bakterii (Nocardia sp., Mycobacterium sp.)specjalna struktura ściany komórkowej
• C10-C18 –wiele mikroorganizmów (Pseudomonas sp., Nocardia sp., Mycobacterium sp., Corynebacrium sp., droŜdŜe np. Candida sp., Torulopsis sp.)
oksydacja subterminalna
oksydacja terminalna
O2 O2
O2
monooksygenaza metylotrofy
CH4 + NADH + H+ + O2 →→→→ CH3OH + NAD+ + H2O monooksygenaza
CH3OH + PQQ →→→→ CH2O + PQQH2
dehydrogenaza
CH2O + NAD+ + H2O →→→→ HCOOH + NADH + H+
HCOOH + NAD+ →→→→ CO2 + NADH + H+
u droŜdŜy brak PQQ
CH3OH + O2 →→→→ CH2O + H2O2
oksydaza
2 H2O2 →→→→ 2 H2O + O2
katalaza
CH2O punktem wyjścia do biosyntezy
PQQ
3 drogi asymilacji aldehydu mrówkowego (CH2O)
• cykl serynowy CH2O + CO2 →→→→ acetyloCoA
3 drogi asymilacji aldehydu mrówkowego (CH2O)
• cykl rybulozomonofosforanu →→→→ fruktozo-6-fosforan
• cykl ksylulozomonofosforanu →→→→ dihydroksyaceton
tylko u droŜdŜy

17
Niepełne utlenianie substratu – bakterie octowe
Acetobacter sp.Gluconobacter sp.
Niepełne utlenianie-produkcja acetoiny i butandiolu
Bacillus spp.
Oddychanie beztlenowe
mrówczan + fumaran →→→→ bursztynian + CO2
H2 + fumaran →→→→ bursztynian
oddychanie azotanowe lub fumaranowe
brak akceptora elektronów – uwalnianie H2
H2dawcą energii
5H2 + 2NO3- + 2H+ →→→→ N2 + 6H2O
Oddychanie beztlenowe – dysymilacyjna redukcja siarczanów
oddychanie biosynteza (asymilacjyjna redukcja siarczanów)
3 grupy mikroorganizmów
• niepełne utlenianie substratuDesulfovibrio sp.(gram-), Desulfotomaculum sp.(gram+)
• pełne utlenianie substratuzmodyfikowany CKTK Desulfobacter sp. (gram-)
oksydatywnej dehydrogenazy CODesulfobacterium sp.(gram-), Desulfotomaculum sp. (gram+)
Arechaeglobus sp.(Archae)
jako źródło węgla mogą słuŜyć:cukry aminokwasyalkohole kw. organicznezw. aromatyczne węglowodory (C12-C20)
tlen włączany do cząsteczki substratu pochodzi z H2O -hydroksylazy nie będące oksydazami
Rozmieszczenie elementów łańcucha oddechowego w błonie
niepełne utlenienie substratu
Desulfovibrio sp.
mleczan+ SO42- + 8H+ →→→→ octan + CO2 + H2S
mleczan pirogronian
octan

18
Pełne utlenianie substratu – zmodyfikowany CKTK
Desulfobacter sp.
• dehydrogenaza kw. αααα-ketoglutarowego zaleŜna od Fd a nie NAD+
• dehydrogenaza jabłczanu niezaleŜna od NAD+,związana z błoną
• liaza a nie syntaza cytrynianu, CoA przenoszone na octan z bursztynylo-CoA, powstaje ATP
Pełne utlenianie substratu – oksydatywna dehydrogenaza CO
Desulfobacterium sp. Arechaeglobus sp.(Archae) Desulfotomaculun sp. (inne koenzymy)
Bakterie homoacetogenne – oddychanie węglanowe
Clostridium aceticum, Acetobacterium woodi
CO2 CO2
fruktoza →→→→ 3 cz. octanu
4H2 +2CO2 →→→→ octan- +2H2O + H+
redukcyjna dehydrogenaza CO
H2dawcą energii
CO2 akceptorem elektronów
CO2 jako końcowy akceptor elektronów:oddychanie węglanowe
dwie grupy organizmów:
acetogeny- redukcja CO2 do octanu
dawcy elektronów:• głównie H2• cukry, kwasy organiczne, aa, alkohole
metanogeny-redukcja CO2 do metanu
formylo-
metylo-
CO2
metylo-
acetylo-OCTAN
Szlak redukcyjnej dehydrogenazy CO
4H2 +2CO2 →→→→ octan- + 2H2O + H+
Bakterie metanogennne (Archae)
4H2 + CO2 →→→→ CH4 + 2H2O
Methanobacterium thermoautotrophicum
CO2 akceptorem elektronów –oddychanie węglanoweH2 dawcą energii
CH3COOH →→→→ CH4 + CO2
Methanosarcina sp. Methanothrix sp.
3CH3OH →→→→ 3CH4 + CO2 + 2H2O
4(CH3)3N + 6H2O →→→→ 9CH4 + 3CO2 + 4NH3
Methanolobus sp., Methanococcoides sp.
Methanosarcina barkeri- modelowy organizm
ograniczona ilość substratów dla metanogenezy

19
Metanogeneza z CO2
formylo-
metyleno-
metylo-
METAN
CO2
Metanogeneza z octanu
METAN
OCTAN
biosynteza
Typy troficzne – źródła energii
Typy troficzne – źródła węgla
CO2 – autotrofy zwiazki organiczne - heterotrofy
CHEMOLITOTROFY
• bakterie wodorowe
H2 + 0.5O2 →→→→ H2O Nocardia, Alcaligenes, Pseudomonas spp.
5H2 + 2NO3- + 2H+ →→→→ N2 + 6H2O Paracoccus denitryficans
4H2 + CO2 →→→→ CH4 + 2H2O Methanobacterium sp.
4H2 +2CO2 →→→→ octan- + 2H2O + H+ Clostridium aceticum, Acetobacterium woodi
• bakterie siarkowe
S2- + 2O2 →→→→ SO42- Beggiatoa, Thiothrix spp.
S0 + 1.5O2 + H2O →→→→ SO42- + 2H+ Thiobacillus, Sulfolobus spp.
S2O32- + 2O2 + H2O →→→→ 2SO4
2- + 2H+ Thiobacillus thiooxidans
5S2O32- + 8NO3
- + H2O →→→→ 10SO42- + 2H+ +4N2 Thiobacillus denitryficans
• bakterie Ŝelazowe
2Fe2+ + 2H+ + 0.5O2 →→→→ 2Fe3+ + H2O Gallionella sp., Thiobacillus ferroxidans
• bakterie nitryfikacyjne
utleniające amoniak
NH4+ + 1.5O2 →→→→ NO2
- + H2O + 2H+ Nitrosomonas, Nitrosospira, Nitrosococcus spp.
utleniająceazotyny
NO2- + 0.5O2 →→→→ NO3
- Nitrobacter, Nitrospira, Nitrococcus spp.
Odwrotny transport elektronów
siła redukcyjna ATPodwrotny transport
bakterie nitryfikacyjne
potencjał dawcy elektronów nie jest dostatecznie ujemny by zapewnić redukcję NAD+ (wyjątek H2)
Figure 20.27b
Odwrotny transport elektronów –bakterie siarkowe
wiele z nich to autotrofy – bardzo wolny wzrost
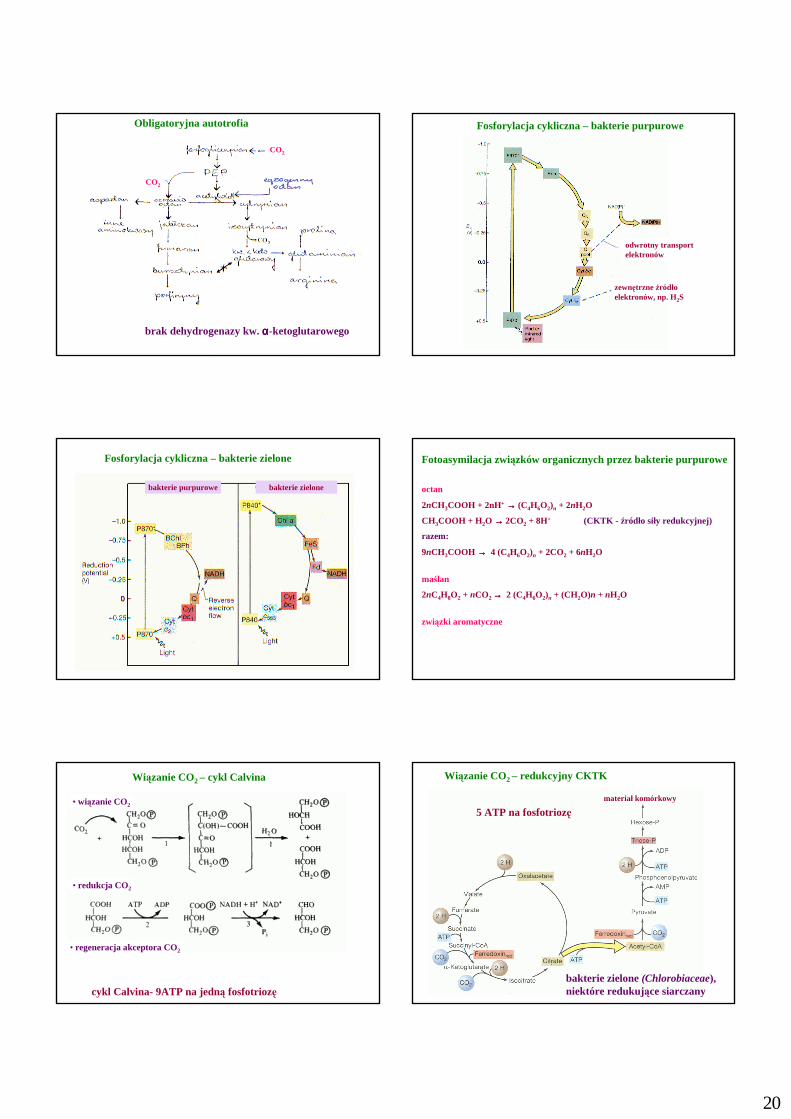
20
CO2
CO2
Obligatoryjna autotrofia
brak dehydrogenazy kw. αααα-ketoglutarowego
Fosforylacja cykliczna – bakterie purpurowe
odwrotny transportelektronów
zewnętrzne źródło elektronów, np. H2S
Fosforylacja cykliczna – bakterie zielone
bakterie purpurowe bakterie zielone
Fotoasymilacja związków organicznych przez bakterie purpurowe
octan
2nCH3COOH + 2nH+ →→→→ (C4H6O2)n + 2nH2O
CH3COOH + H2O →→→→ 2CO2 + 8H+ (CKTK - źródło siły redukcyjnej)
razem:
9nCH3COOH →→→→ 4 (C4H6O2)n + 2CO2 + 6nH2O
maślan
2nC4H8O2 + nCO2 →→→→ 2 (C4H6O2)n + (CH2O)n + nH2O
związki aromatyczne
Wiązanie CO2 – cykl Calvina
cykl Calvina- 9ATP na jedną fosfotriozę
• wiązanie CO2
• redukcja CO2
• regeneracja akceptora CO2
Wiązanie CO2 – redukcyjny CKTK
5 ATP na fosfotriozęmateriał komórkowy
bakterie zielone(Chlorobiaceae),niektóre redukujące siarczany

21
Wiązanie CO2 – szlak redukcyjnej dehydrogenazy CO
metanogeny, acetogeny, niektóre redukujące siarczany3 ATP na fosfotriozę
formylo-
metylo-
CO2
metylo-
acetylo-CoA
fosfotrioza3ATP
Dodatkowe karboksylacje
1. karboksylaza PEP
PEP + CO2 → szczawiooctan+ Pi
(pałeczki jelitowe,Bacillus sp.)
2. karboksylaza pirogronianu
pirogronian + CO2 + ATP → szczawiooctan+ ADP + Pi
(Pseudomonas sp., Bacillus sp.)
3. cykl glioksalowy
4. acetylo-CoA + CO2 + FdH2 → pirogronian+ Fd + CoA
UmoŜliwiaj ą wzrost na octanie, u niektórych (np. Clostridium sp.)
aŜ 30% węgla komórkowego pochodzi z CO2
polimery(polisacharydy,bia łka)
monomery(cukry, aminokwasy)
octan
CH4+CO2
propionian, ma ślan,bursztynian, alkohole
H2+CO2 octan
H2+CO2
octan
fermentacja I
fermentacja II
metanogeneza
acetogeneza
hydroliza
Fermentacje II rzędu
1. 3 mleczan →→→→ 2 kw. propionowy + octan + CO2(Propionibacterium sp., Clostridium sp.)
2. etanol + H2O →→→→ octan + 2H2Go’ = +9.6 kJ
3. kw. masłowy+ 2H2O →→→→ 2octan + 2H2Go’ = +48.1 kJ
(Syntrophomonas wolfei)
4. kw. propionowy + 3H2O →→→→ octan + + CO2 + 3H2Go’ = +76.1 kJ
(Syntrophobacter wolfinii)
syntropiamiędzygatunkowe przekazywanie wodoru
przełyk
odbyt
Układ pokarmowy
Enterococcus faecalis, pałeczki jelitoweBacteroides sp.Bifidobacterium sp.Peptococcus sp.Peptostreptococcus sp.Ruminococcus sp.Closrtidium sp.Streptococcus sp.Staphylococcus sp.Lactobacillus sp.
Lactobacillus sp.Enterococcus sp.
Główne grupy mikroorganizmów1010 – 1011 kom./g
układ moczowyEscherichia coli, Proteus mirabilis– patogeny oportunistyczne(zmiana pH, spadek oporności)
układ płciowyniskie pH utrzymywane przezLactobacillus acidophilusinne organizmy to:Escherichia coli,Streptococcus sp. droŜdŜaki:Candidai Torulopsis spp.

22
OBECNOŚĆ patogenów
ADHEZJA
WZROST i KOLONIZACJAlokalna
TOKSYCZNO ŚĆefekt lokalny lub systemowy
INWAZYJNO ŚĆwzrost lokalny lub
w miejscach oddalonych
USZKODZENIE TKANEKCHOROBA
Czynniki adhezyjne:
glycocalyx enterotoksyczna Escherichia coli(ETEC)Streptococcus mutans
białka adhezyjne Streptococcus pyogenes białkoMNeiseria gonorrhoeae białko Opa
kwas lipotejchojowy Streptococcus pyogenes
fimbrie (pili) Neiseria gonorrhoeae enterotoksyczna Escherichia coli(ETEC)
Wnikanie do wnętrza komórki
• namnaŜanie się, wzrost, wywoływanie choroby ściśle związane
z inwazją cytoplazmy komórek gospodarzariketsie, chlamydie– pasoŜyty wewnątrzkomórkowe
• ucieczka przed układem immunologicznym, moŜliwość rozprzestrzeniania się po całym organizmie
Mycobacterium tuberculosis, Neisseria gonorrhoeae,
Shigella sp., Salmonella sp., patogenne szczepyEscherichia coli
Neutrofil – fagocytoza komórek bakteryjnych
lizosomy
bakterie
fagosom
fagolizosom
adhezja
wydzie-lanie
wchła-nianie
trawienie
pseudo-podium
cytoplazma
błonacytoplazmatyczna
1. repelenty, toksyny –brak chemotaksji
2. otoczki, śluzy, biofilm –brak adhezji i pochłaniania
3. blokowanie fuzji lizosomów z fagosomem
4. wydzielanie katalazy
5. nieprzepuszczalne osłony
komórkowe
6. blokowanie odpowiedzi makrofagów na czynniki stymulujące
7. utrata zdolności do prezentacji antygenów
8. ucieczka patogenu z fagosomu
Unikanie fagocytozy Inwazja przez wymuszoną fagocytozę – Shigella sp.
komórka M
makrofag
komórkanabłonkajelita
aktyna-ruch

23
Dopełniacz
liza chemotaksja opsonizacja
bakterie bakteriefagocyt
komplement
IgA toksyny neutralizacja fagocytoza
IgG bakterie między opsonizacja fagocytoza komórkami
IgM bakterie w kompleksy liza iplazmie immunologiczne fagocytoza
fagocyt opsonizacja wiązanie
dopełniacz
dopełniaczi przeciwciała
przeciwciała
Systemy sekrecyjne dla toksyn bakteryjnych
toksyna Bor. pertusis(PT),
Legionella sp., Helicobacter sp. IV
białka Yop Yersinia sp.
białka Ipa Shigella sp.III
pullulanaza Klebsiella oxytoca
elastaza, fosfolipaza Ps. aeruginosaII
αααα-hemolizyna E.colicyklaza adenylowa Bor. pertusis
proteaza Ps. aeruginosa
I
IgA proteaza N.gonorrhoeae
hemaglutynina Bor. pertusisautotransportery
przykładowe toksynytyp systemu
sekrecyjnego
Autotransportery
peryplazma
cytoplazma
peryplazma
Typ I

24
Typ II
pili typu IV
Typ IIIkomórka eukariotyczna
rzęska
do komórki eukariotycznej
Typ IV
Curr Op Cell Biol (2000) 12:420
Podział toksyn bakteryjnych
endotoksyny
• składniki LPS, głównie lipid A• bardzo podobne, działają w taki sam sposób, objawy ogólne (gorączka, wymioty, biegunka)
• ciepłostałe
• mało toksyczne LD50 300 mikrogramów
egzotoksyny
• białka, wydzielane do środowiska przez bakterie G+ i G-• bardzo róŜnorodna grupa, specyficzne działanie (niepyrogenne)
• ciepłochwiejne (są wyjątki np. toksyna S.aureus)
• bardzo toksyczne LD50 25 pikogramów (botulina)
Podział toksyn bakteryjnych
neurotoksyny
zakłócają normalne przekazywanie impulsów nerwowychClostridium tetani, Clostridium botulinum
cytotoksynyzabijają komórki gospodarzaCorynebacterium diphteriae– hamuje elongację białka
Streptococcus pyogenes– uszkadza błonę lizosomów
enterotoksyny
stymulują komórki układu pokarmowego do wydzielania wodyi elektrolitów do światła jelita
Vibrio cholerae, Escherichia coli,Shigella dysenteriae(hamuje syntezę białka w kom. nabłonka jelita)
Bakterie chorobotwórcze
Staphylococcus aureus
• obecny na skórze i w nosogardzieli
• powoduje zatrucia pokarmowe (50% szczepów produkuje enterotoksynę)• choroby związane z produkcją ropy (zapalenia skóry, wrzody)
• produkuje kolagenazę oraz cytotoksyny (hemolizyna, leukocydyna)
Diplococcus pneumoniae
• typowa flora górnych dróg oddechowych
• powoduje zapalenie płuc, opon mózgowych, ucha środkowego• 30% śmiertelność, duŜy naciek limfocytów, cytokin,
zapalenie systemowe, uszkodzenie tkanek gospodarza• oporność warunkowana przez przeciwciała przeciwko
cukrowej otoczce – duŜa zmienność patogenu

25
Streptococcus pyogenes
• rzadko występuje u zdrowych ludzi• powoduje ropne zapalenie gardła oraz zapalenie skóry
• produkuje cytotoksyny (ββββ-hemoliza) oraz enzymy (nukleazy,lipazy, proteazy)
• białka powierzchniowe A i M to tzw. superantygeny• liczne powikłania (zapalenie kłębuszków nerkowych,
reumatyczne zapalenie mięśnia sercowego) spowodowane osadzaniem się w tkankach kompleksów immunologicznych
Corynebacterium difteriae
• powoduje infekcje gardła, nekrozę komórek epitelialnych,
toksyna ma efekt systemowy (serce, układ nerwoway, nerki)• produkuje toksynę dyfterytu (hamuje syntezę białka)
Corynebacterium difteriae – transformacja lizogenna
W.J.H. Kunicki – Goldfingerśycie Bakterii PWN 1998
Helicobacter pylori
• bardzo często występuje u zdrowych ludzi• powoduje powstawanie wrzodów oraz nowotworów Ŝołądka
• czynniki wirulencji: system sekrecyjny IV oraz cytotoksynę indukuj ącą procesy zapalne i apoptotyczne w komórkach
Bacillus anthracis
• powoduje wąglik, tzw. zoonozę, chorobę odzwierzęcą
• powoduje skórne i płucne infekcje, obrzęki, nekroza tkanek,wysoka śmiertelność
• czynniki wirulecji: otoczka uniemoŜliwiaj ąca fagocytozę, wytwarzanie egzotoksyn (zaburzenie równowagi jonowej,
zakłócenie przekazywania sygnałów w obrębie układuimmunologicznego)
Porównanie cech Bacteria, Archaeai Eukarya
taktaknieczynniki transkrypcyjne
TATATATA-10 i -35promotory
nietaktakoperony
trzy (12-14 podj. kaŜda)
kilka (8-12 podj. kaŜda)
jedna (4podj)polimeraza RNA
taktaknieintrony
MetMetf-Metinicjacyjny tRNA
80S70S70Srybosomy
estryeteryestrylipidy błonowe
róŜny składróŜny składmureinaściana komórkowa
taktakniebiałka histonowe
nietaktakDNA w postaci CCC
nietaktakkomórka prokariotyczna
EukaryaArchaeaBacteriacecha
Porównanie cechBacteria, Archaeai Eukarya
nietaktakwzrost powyŜej 80oC
nietaktakchemolitotrofia
nietaktakwiązanie azotu
tak (chloroplasty)
nietakfotosynteza (chlorofil)
nietaktakdysymilacyjna redukcja siarczanów
nietaktakdenitryfikacja
nienietaknitryfikacja
nietaktakredukcja So do H2S
nietakniemetanogeneza
EukaryaArchaeaBacteriacecha
Nature (1998) 392: 38
The hydrogen hypothesis for the first eukaryote
William Martin, Miklos Muller
Recommended











![WLS 2013 małe grucz. ślin.Przew. pokarm. [tryb zgodności] · qBłona surowicza, czyli otrzewna trzewna Żołądek. Dwunastnica Jelito krJelito cienkie ęte Błona śluzowa Kosmki](https://img.dokumen.tips/doc/110x75/5c76cf0409d3f2ff328c45be/wls-2013-male-grucz-slinprzew-pokarm-tryb-zgodnosci-qblona-surowicza.jpg)





