10.1128/JB.185.15.4508-4518.2003.
2003, 185(15):4508. DOI:J. Bacteriol. Bonnie J. Stewart and Linda L. McCarter parahaemolyticus
VibrioLateral Flagellar Gene System of
http://jb.asm.org/content/185/15/4508Updated information and services can be found at:
These include:
REFERENCEShttp://jb.asm.org/content/185/15/4508#ref-list-1This article cites 85 articles, 46 of which can be accessed free at:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from

JOURNAL OF BACTERIOLOGY, Aug. 2003, p. 4508–4518 Vol. 185, No. 150021-9193/03/$08.00�0 DOI: 10.1128/JB.185.15.4508–4518.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Lateral Flagellar Gene System of Vibrio parahaemolyticusBonnie J. Stewart and Linda L. McCarter*
Department of Microbiology, The University of Iowa, Iowa City, Iowa 52242
Received 28 January 2003/Accepted 1 May 2003
Vibrio parahaemolyticus possesses dual flagellar systems adapted for movement under different circum-stances. A single polar flagellum propels the bacterium in liquid (i.e., swimming) with a motor that is poweredby the sodium motive force. Multiple proton-driven lateral flagella enable translocation over surfaces (i.e.,swarming). The polar flagellum is produced continuously, while production of lateral flagella is induced whenthe organism is grown on surfaces. This work describes the isolation of mutants with insertions in thestructural and regulatory laf genes. A Tn5-based lux transcriptional reporter transposon was constructed andused for mutagenesis and subsequent transcriptional analysis of the laf regulon. Twenty-nine independentinsertions were distributed within 16 laf genes. DNA sequence analysis identified 38 laf genes in two loci. Amongthe mutants isolated, 11 contained surface-induced lux fusions. A hierarchy of laf gene expression wasestablished following characterization of the laf::lux transcriptional fusion strains and by mutational andprimer extension analyses of the laf regulon. The laf system is like many enteric systems in that it is aproton-driven, peritrichous flagellar system; however, laf regulation was different from the Salmonella-Es-cherichia coli paradigm. There is no apparent flhDC counterpart that encodes master regulators known tocontrol flagellar biosynthesis and swarming in many enteric bacteria. A potential �54-dependent regulator,LafK, was demonstrated to control expression of early genes, and a lateral-specific �28 factor controls lateflagellar gene expression. Another notable feature was the discovery of a gene encoding a MotY-like product,which previously had been associated only with the architecture of sodium-type polar flagellar motors.
Vibrio parahaemolyticus can differentiate into a specializedcell type, called the swarmer cell, which is adapted for rapidmovement on surfaces. Upon differentiation, numerous neworganelles—lateral flagella—are peritrichously elaborated onthe cell surface (4). Lateral flagella are encoded by one of thetwo distinct flagellar gene sets in V. parahaemolyticus. Each setcontains more than 35 structural and regulatory genes. Thepolar flagellum, which is sheathed by an extension of the cellouter membrane, is constitutively expressed (48). It is a highlyeffective propulsive organelle in liquid environments. Rotationspeed of the polar flagellum, which acts as a propeller, hasbeen measured to average 1,100 revolutions per s for closelyrelated V. alginolyticus (45, 46). The sodium motive force pow-ers this rotation, and swimming speeds achieve �60 �m per s(9). However, the polar organelle becomes less effective asviscosity increases, and lateral flagella are induced (11, 47).The proton motive force powers the peritrichous lateral fla-gella, similar to the peritrichous flagella of Escherichia coli (9).Production of numerous lateral flagella enables many marineVibrio species to move through viscous environments or oversurfaces (10). Some other bacteria possess both peritrichousand polar flagellation, including Aeromonas and Azospirillumspecies and Rhodospirillum centenum; however, there appearsto be some functional overlap of the two kinds of flagella (2,35, 55, 57, 74). Other swarming bacteria typically regulate geneexpression of a single peritrichous system to accommodate theneed for different numbers of flagella (28, 29). For example,swarming in Proteus mirabilis and Serratia liquefaciens involvesupregulation of flagellar biosynthesis through the master reg-ulatory genes flhD and flhC (22, 24).
For V. parahaemolyticus, the two flagellar systems appeardistinct, although central chemotaxis elements are shared (73).All mutants that have been isolated with defects in the polarflagellar system (fla) show no defect in swarming (13, 34, 49).Although the polar gene system and its regulation have beenstudied in V. parahaemolyticus, the lateral flagellar system (laf)has not been well characterized. This work employs transposonmutagenesis by a Tn5 derivative in combination with the sen-sitive reporter function of a promoterless luxCDABE operon toanalyze the laf gene system of V. parahaemolyticus. Until now,the genetics of dual flagellar systems in a single organism havenot been dissected.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The strains and plasmids used in thiswork are described in Table 1. V. parahaemolyticus strains were grown at either30°C or room temperature (�27°C) in heart infusion medium (HI broth) con-taining 25 g of heart infusion (Difco) and 20 g of NaCl per liter. Swarm plateswere prepared by adding 15 g of Bacto agar (Difco) to HI broth. Swimmingmedium was made with Vogel-Bonner salts medium (84) supplemented with 20 gof NaCl, 3.25 g of agar per liter, and 0.4% galactose. Translucent V. parahae-molyticus strains were used in this study. V. parahaemolyticus undergoes phasevariation between opaque and translucent colony types (51). Translucent strainsproduce less capsular polysaccharide and are more swarming proficient thanopaque strains (20). Strain LM5431 was used as the target strain for transposonmutagenesis. This strain, a derivative of the translucent swarming-proficient strainLM5395, was made spontaneously resistant to phosphonomycin (100 �g/ml) accord-ing to the method of Alper and Ames (5). Strain LM5431 is translucent, swarm andswim competent, and a prototroph. Antibiotics were used at final concentrationsof 50 �g of kanamycin per ml, 10 �g of chloramphenicol per ml, 60 �g ofphosphonomycin per ml, 30 �g of gentamicin per ml, and 10 �g of tetracyclineper ml. E. coli strains were maintained with the same antibiotic concentrations asjust described, except that 15 �g of gentamicin per ml was used. Overexpressionplasmids were induced with 1 mM isopropyl-�-D-thiogalactopyranoside (IPTG).
Genetic and molecular techniques. General molecular biology methods wereadapted from Sambrook et al. (72). Chromosomal DNA was prepared accordingto the protocol of Woo et al. (87). Some insertion and deletion mutations weremade with a � Red recombinase system (17) to introduce Camr cassettes onto
* Corresponding author. Mailing address: Department of Microbi-ology, The University of Iowa, Iowa City, IA 52242. Phone: (319)335-9721. Fax: (319) 335-7679. E-mail: [email protected].
4508
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
cosmids carrying 40-kb insertions of wild-type V. parahaemolyticus DNA (53) inE. coli. Conjugation and gene replacement methods for V. parahaemolyticus havebeen described elsewhere (77). All gene replacements were confirmed by South-ern blot analysis performed using Hybond-NX membranes (Amersham) and
32P-dCTP-labeled DNA probes (Pharmacia). RNA isolation for surface- andliquid-grown cells was performed using trizol reagent (Gibco BRL) according tothe manufacturer’s suggestions. Reverse transcription-PCR (RT-PCR) was per-formed as specified using Access RT-PCR (Promega).
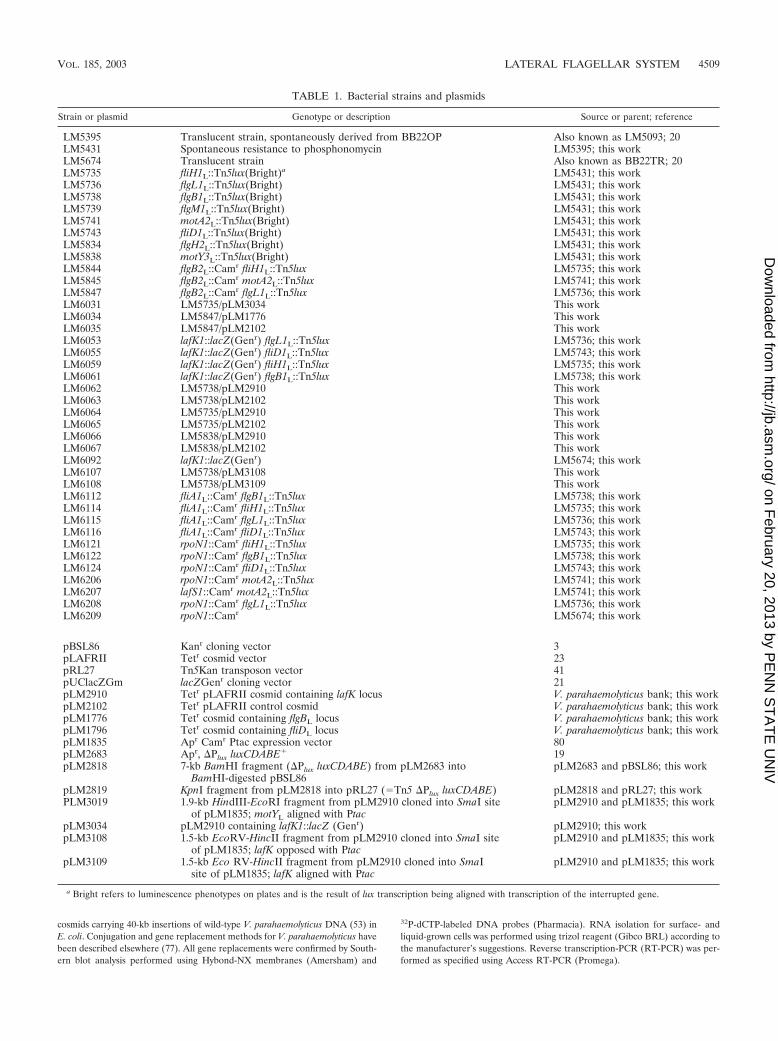
TABLE 1. Bacterial strains and plasmids
Strain or plasmid Genotype or description Source or parent; reference
LM5395 Translucent strain, spontaneously derived from BB22OP Also known as LM5093; 20LM5431 Spontaneous resistance to phosphonomycin LM5395; this workLM5674 Translucent strain Also known as BB22TR; 20LM5735 fliH1L::Tn5lux(Bright)a LM5431; this workLM5736 flgL1L::Tn5lux(Bright) LM5431; this workLM5738 flgB1L::Tn5lux(Bright) LM5431; this workLM5739 flgM1L::Tn5lux(Bright) LM5431; this workLM5741 motA2L::Tn5lux(Bright) LM5431; this workLM5743 fliD1L::Tn5lux(Bright) LM5431; this workLM5834 flgH2L::Tn5lux(Bright) LM5431; this workLM5838 motY3L::Tn5lux(Bright) LM5431; this workLM5844 flgB2L::Camr fliH1L::Tn5lux LM5735; this workLM5845 flgB2L::Camr motA2L::Tn5lux LM5741; this workLM5847 flgB2L::Camr flgL1L::Tn5lux LM5736; this workLM6031 LM5735/pLM3034 This workLM6034 LM5847/pLM1776 This workLM6035 LM5847/pLM2102 This workLM6053 lafK1::lacZ(Genr) flgL1L::Tn5lux LM5736; this workLM6055 lafK1::lacZ(Genr) fliD1L::Tn5lux LM5743; this workLM6059 lafK1::lacZ(Genr) fliH1L::Tn5lux LM5735; this workLM6061 lafK1::lacZ(Genr) flgB1L::Tn5lux LM5738; this workLM6062 LM5738/pLM2910 This workLM6063 LM5738/pLM2102 This workLM6064 LM5735/pLM2910 This workLM6065 LM5735/pLM2102 This workLM6066 LM5838/pLM2910 This workLM6067 LM5838/pLM2102 This workLM6092 lafK1::lacZ(Genr) LM5674; this workLM6107 LM5738/pLM3108 This workLM6108 LM5738/pLM3109 This workLM6112 fliA1L::Camr flgB1L::Tn5lux LM5738; this workLM6114 fliA1L::Camr fliH1L::Tn5lux LM5735; this workLM6115 fliA1L::Camr flgL1L::Tn5lux LM5736; this workLM6116 fliA1L::Camr fliD1L::Tn5lux LM5743; this workLM6121 rpoN1::Camr fliH1L::Tn5lux LM5735; this workLM6122 rpoN1::Camr flgB1L::Tn5lux LM5738; this workLM6124 rpoN1::Camr fliD1L::Tn5lux LM5743; this workLM6206 rpoN1::Camr motA2L::Tn5lux LM5741; this workLM6207 lafS1::Camr motA2L::Tn5lux LM5741; this workLM6208 rpoN1::Camr flgL1L::Tn5lux LM5736; this workLM6209 rpoN1::Camr LM5674; this work
pBSL86 Kanr cloning vector 3pLAFRII Tetr cosmid vector 23pRL27 Tn5Kan transposon vector 41pUClacZGm lacZGenr cloning vector 21pLM2910 Tetr pLAFRII cosmid containing lafK locus V. parahaemolyticus bank; this workpLM2102 Tetr pLAFRII control cosmid V. parahaemolyticus bank; this workpLM1776 Tetr cosmid containing flgBL locus V. parahaemolyticus bank; this workpLM1796 Tetr cosmid containing fliDL locus V. parahaemolyticus bank; this workpLM1835 Apr Camr Ptac expression vector 80pLM2683 Apr, �Plux luxCDABE� 19pLM2818 7-kb BamHI fragment (�Plux luxCDABE) from pLM2683 into
BamHI-digested pBSL86pLM2683 and pBSL86; this work
pLM2819 KpnI fragment from pLM2818 into pRL27 (�Tn5 �Plux luxCDABE) pLM2818 and pRL27; this workPLM3019 1.9-kb HindIII-EcoRI fragment from pLM2910 cloned into SmaI site
of pLM1835; motYL aligned with PtacpLM2910 and pLM1835; this work
pLM3034 pLM2910 containing lafK1::lacZ (Genr) pLM2910; this workpLM3108 1.5-kb EcoRV-HincII fragment from pLM2910 cloned into SmaI site
of pLM1835; lafK opposed with PtacpLM2910 and pLM1835; this work
pLM3109 1.5-kb Eco RV-HincII fragment from pLM2910 cloned into SmaIsite of pLM1835; lafK aligned with Ptac
pLM2910 and pLM1835; this work
a Bright refers to luminescence phenotypes on plates and is the result of lux transcription being aligned with transcription of the interrupted gene.
VOL. 185, 2003 LATERAL FLAGELLAR SYSTEM 4509
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from

Primer extension experiments were conducted as previously described (34)using RNA (�15 �g/reaction) prepared from plate- and liquid-grown LM5674.The primer extension reactions were repeated at least two times with indepen-dently isolated RNA preparations. For all laf genes examined, there were primerextension products for plate-grown but not for liquid-grown cells. As a control tonormalize the RNA preparations, polar flagellin transcription was analyzed. Forthe polar flagellin gene flaA, there were primer extension products for bothplate- and liquid-grown cells. The LafA primer extension product was so abun-dant that the reaction was diluted 1:25 prior to loading on the sequencing gel.
Mutagenesis. Plasmid pRL27 contains a hyperactive Tn5-transposase geneexpressed from the tetA promoter (41) and a Tn5-derived transposon containingthe oriR6K replication origin and a gene encoding kanamycin resistance. Tomake the transposon a luminescence reporter, a promoterless luxCDABE operon(19) from Vibrio fischeri was inserted into the transposon on pRL27. A 7-kb �Plux
cassette was cloned from pLM2683 with BamHI into vector pBSL86, thus mak-ing pLM2818. The cassette was excised from pLM2818 by using flanking KpnIsites and inserted into KpnI-digested pRL27 to yield pLM2819. Transposonmutagenesis was performed on the phosphonomycin-resistant strain LM5431.The recipient was grown overnight in medium containing phosphonomycin anddiluted to an optical density at 600 nm (OD600) of 0.01 followed by 6 h of growthwithout antibiotic. This culture was mixed with an equal volume of an exponen-tially growing E. coli strain carrying the transposon delivery plasmid pLM2819.HI plates with no NaCl were spotted with 10 25-�l drops of the mating mixtureand grown overnight at 37°C. Cells from each mating plate were suspended in 5ml of 0.3 M sucrose and diluted 10-fold to a final OD600 value of 0.2 to 0.5. Avolume of 100 �l of this mating suspension was spread onto HI medium thatcontained the phosphonomycin for counterselection of the E. coli donor andkanamycin for selection of V. parahaemolyticus transposon mutants. Approxi-mately 2,000 colonies were picked from each mating and maintained on HImedium containing kanamycin. Southern blot analysis on representative mutantsconfirmed the randomness of transposition and absence of multiple insertions.
Lateral flagellar mutant identification. Gridded arrays of mutant colonieswere transferred to the appropriate media to examine swarming and swimmingphenotypes. Colony immunoblots of the swarm� swim� mutants were used toassess lateral flagellin (LafA) production. Activity of the lux reporter gene wasmeasured by examining light production of the mutants in a luminescence imager(LAS-1000; Fujifilm). Chromosomal DNA was isolated from selected mutantsand arbitrarily primed PCR was performed by using primers designed to atransposon end and arbitrary primers ARB6 and ARB2 as described by O’Tooleand Kolter (63). The transposon-specific primers used were KANARB1 (5-GGCGATTCAGGCCTGGTATGAG-3), KANARB2 (5-GCATGCAAGCTTCAGGGTTGAG-3), LUXARB1 (5-GTGTTCTCTTCGGCGGCGCTGG-3),and LUXARB2 (5-GATCCTCGCCGTACTGCCCGC-3). PCR products weresequenced to identify the interrupted gene and were also radiolabeled to probea V. parahaemolyticus cosmid library. Subsequent DNA sequence analysis of theregion of interest was performed on the retrieved cosmid.
Surface and liquid luminescence assays. Bioluminescence on plates was mon-itored by exposing overnight colonies in a Fujifilm luminescent image analyzer(LAS-1000) for 30 s to 5 min. For time course experiments on plate-grown cells,strains were grown on plates and suspended to an OD600 of 0.05 and 50 �l wasspread onto multiple fresh HI swarm plates with antibiotics when appropriate.Plates were suspended at specified times in 5 ml of HI broth, and OD andrelative light unit (RLU) measurements were recorded. Several plates wereharvested for early time points to ensure an adequate cell number. For timecourses in liquid, overnight cultures were diluted to OD600 values of 0.10 and 30�l was inoculated into 250-ml flasks containing 15 ml of HI broth. These cultureswere incubated with aeration at room temperature with periodic sampling forOD and light readings. Luminescence was quantified by measuring 0.1-ml sam-ples for 30 s in a TD20/22 luminometer (Turner Designs). Dilutions were madeto keep all measurements (RLU) within a linear range. Luminescence is re-ported here as specific light units (SLU), which describe RLU per minute permilliliter per OD600. Each experiment was performed at least two times, withlight readings taken in triplicate.
Immunoblots. Whole-cell preparations were made by suspending 16-h plate-grown cells to an OD600 of 2.0 in 0.5 ml of Laemmli sample buffer (40). Samples(5 �l) were run on 12% acrylamide gels and blotted as previously described (13).Immunoblot probes consisted of a pool of polyclonal antibodies raised to thelateral flagellin protein LafA (antibody no. 127) at a concentration of 1:10,000(54) and to a �40 kDa V. parahaemolyticus outer membrane protein (antibodyno. 634) at a concentration of 1:5,000. Colony immunoblots were performedessentially as described by Meyer et al. (56). Anti-LafA was used at a concen-tration of 1:1,000 overnight for colony blots.
DNA sequence analysis. DNA sequence determination was performed by theDNA Core Facility of the University of Iowa. Oligonucleotides were manufac-tured by Integrated DNA Technologies, Inc. (Coralville, Iowa). The KANARB2and LUXARB2 primers were used to sequence outward from the ends of thetransposon. Sequence assembly was performed by using the Genetics ComputerGroup (GCG) software package. Searches for homology were carried out by theNational Center for Biotechnology Information using the BLAST network ser-vice (6). Sequence alignments were performed by using the CLUSTAL W pro-gram (found at http://www.ebi.ac.uk/clustalw) (83). Only the sequence for whichwe obtained double-stranded information was deposited in GenBank.
V. parahaemolyticus lateral flagellar GenBank accession numbers are U52957(lafA-motBL); U52597 (flgNL-flgEL); and AY22518 (fliML-fliJL).
RESULTS
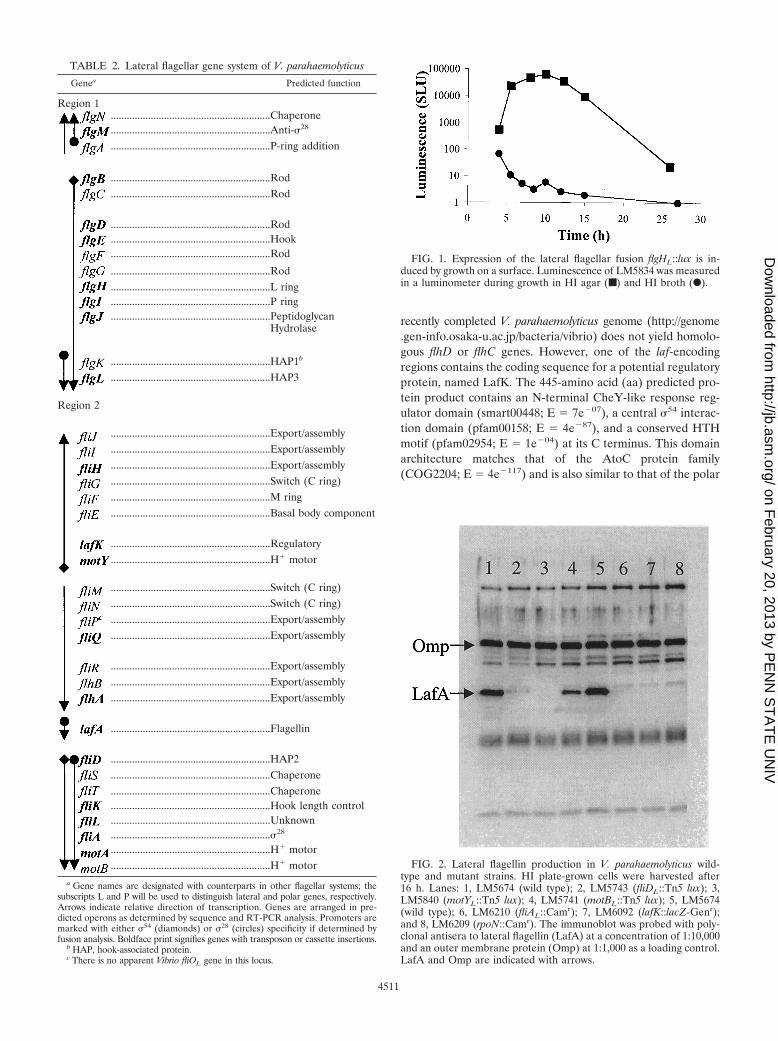
Mutagenesis and identification of lateral flagellar (laf) mu-tants. The V. parahaemolyticus strain LM5431, which is aphosphonomycin-resistant derivative of the translucent strainLM5395, was mutagenized with a Tn5-derived transposon car-rying a reporter luxCDABE operon. Approximately 12,000 mu-tants were screened for swarming defects on 1.5% agar plates.Mutants defective in swarming motility—but still capable ofswimming motility—were assayed for the production of lateralflagellin by probing colony immunoblots with an antibody spe-cific to the lateral flagellin protein, LafA. Laf� insertion mu-tants were examined for laf-dependent lux gene expression byassaying surface growth-dependent light emission. DNA se-quence analysis using either arbitrarily primed PCR fragmentsor plasmids carrying the kanamycin-resistant end of the Tn5lux and a portion of flanking chromosomal DNA was used toidentify transposon-interrupted laf genes. Twenty-nine trans-poson insertions were identified in 16 lateral flagellar genes(Table 2). All but two of these lateral flagellar genes containedat least one transcriptional fusion, which produced surface-induced light. An example of surface-induced light productionfor one of the fusion strains is shown in Fig. 1. Light expressionof flgHL, encoding a lateral flagellar L-ring component, wasincreased by at least 1,000-fold when the fusion strain wasgrown on a surface. All of the mutants with defects in thesegenes failed to produce lateral flagellin, except strains withinsertions in the lateral flagellar motor gene, motAL. The LafAproduction profiles of a subset of these mutants are shown inthe immunoblot in Fig. 2, lanes 2 to 4.
Organization of the laf loci. Nucleotide sequence and PCRanalysis were used to establish the gene order between inser-tion mutations and to identify additional potential laf genes.Table 2 lists the predicted genes and gene products in the lafsystem. A total of 38 open reading frames (ORFs) were foundin two distinct regions. DNA sequence analysis revealed littleor no intergenic spacing among large groups of ORFs andoverlap of potential coding regions. RT PCR was performed byusing RNA from surface-grown cells to confirm transcriptionallinkage between some ORFs. A PCR product contingent onthe presence of both reverse transcriptase and RNA, and sim-ilar to that amplified from chromosomal DNA, signified tran-scriptional coupling between fliRL and flhBL as well as lafKL
and fliEL (data not shown). Based on these observations (aswell as mutant analysis), at least seven operons within two lociare predicted (shown in Table 2).
A laf-specific regulator: LafK. No counterpart to the masterflagellar regulatory operon flhDC of enteric bacteria was foundin the laf-encoding regions. Moreover, BLAST analysis of the
4510 STEWART AND MCCARTER J. BACTERIOL.
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
recently completed V. parahaemolyticus genome (http://genome.gen-info.osaka-u.ac.jp/bacteria/vibrio) does not yield homolo-gous flhD or flhC genes. However, one of the laf-encodingregions contains the coding sequence for a potential regulatoryprotein, named LafK. The 445-amino acid (aa) predicted pro-tein product contains an N-terminal CheY-like response reg-ulator domain (smart00448; E � 7e�07), a central 54 interac-tion domain (pfam00158; E � 4e�87), and a conserved HTHmotif (pfam02954; E � 1e�04) at its C terminus. This domainarchitecture matches that of the AtoC protein family(COG2204; E � 4e�117) and is also similar to that of the polar
FIG. 1. Expression of the lateral flagellar fusion flgHL::lux is in-duced by growth on a surface. Luminescence of LM5834 was measuredin a luminometer during growth in HI agar (■) and HI broth (F).
FIG. 2. Lateral flagellin production in V. parahaemolyticus wild-type and mutant strains. HI plate-grown cells were harvested after16 h. Lanes: 1, LM5674 (wild type); 2, LM5743 (fliDL::Tn5 lux); 3,LM5840 (motYL::Tn5 lux); 4, LM5741 (motBL::Tn5 lux); 5, LM5674(wild type); 6, LM6210 (fliAL::Camr); 7, LM6092 (lafK::lacZ-Genr);and 8, LM6209 (rpoN::Camr). The immunoblot was probed with poly-clonal antisera to lateral flagellin (LafA) at a concentration of 1:10,000and an outer membrane protein (Omp) at 1:1,000 as a loading control.LafA and Omp are indicated with arrows.
TABLE 2. Lateral flagellar gene system of V. parahaemolyticus
Genea Predicted function
Region 1............................................................Chaperone............................................................Anti-28
............................................................P-ring addition
............................................................Rod
............................................................Rod
............................................................Rod
............................................................Hook
............................................................Rod
............................................................Rod
............................................................L ring
............................................................P ring
............................................................PeptidoglycanHydrolase
............................................................HAP1b
............................................................HAP3
Region 2
............................................................Export/assembly
............................................................Export/assembly
............................................................Export/assembly
............................................................Switch (C ring)
............................................................M ring
............................................................Basal body component
............................................................Regulatory
............................................................H� motor
............................................................Switch (C ring)
............................................................Switch (C ring)
............................................................Export/assembly
............................................................Export/assembly
............................................................Export/assembly
............................................................Export/assembly
............................................................Export/assembly
............................................................Flagellin
............................................................HAP2
............................................................Chaperone
............................................................Chaperone
............................................................Hook length control
............................................................Unknown
............................................................28
............................................................H� motor
............................................................H� motor
a Gene names are designated with counterparts in other flagellar systems; thesubscripts L and P will be used to distinguish lateral and polar genes, respectively.Arrows indicate relative direction of transcription. Genes are arranged in pre-dicted operons as determined by sequence and RT-PCR analysis. Promoters aremarked with either 54 (diamonds) or 28 (circles) specificity if determined byfusion analysis. Boldface print signifies genes with transposon or cassette insertions.
b HAP, hook-associated protein.c There is no apparent Vibrio fliOL gene in this locus.
4511
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
flagellar regulators FlaK and FlaM of V. parahaemolyticus (34,79), FlrA and FlrC in Vibrio cholerae (36), and FleQ and FleRin Pseudomonas aeruginosa (7, 71). Amino acid alignment ofLafK, other 54-interacting flagellar proteins, and NtrC fromE. coli shows that the four highly conserved residues commonlyassociated with response regulator receiver domain phosphor-ylation, corresponding to NtrC residues Asp-11, Asp-12, Asp-54, and Lys-104, are not present in LafK (58, 81). The 54
interaction domain in LafK extends from aa 126 to 344 andcontains two regions involved in ATP hydrolysis. The helix-turn-helix domain spans from aa 390 to 429. No transposoninsertions were identified in the lafK coding region; therefore,the mutant strain LM6092 was constructed bearing the lafK::lacZ(Genr) mutation. LM6092 failed to swarm or producelateral flagellin (Fig. 2, lane 7).
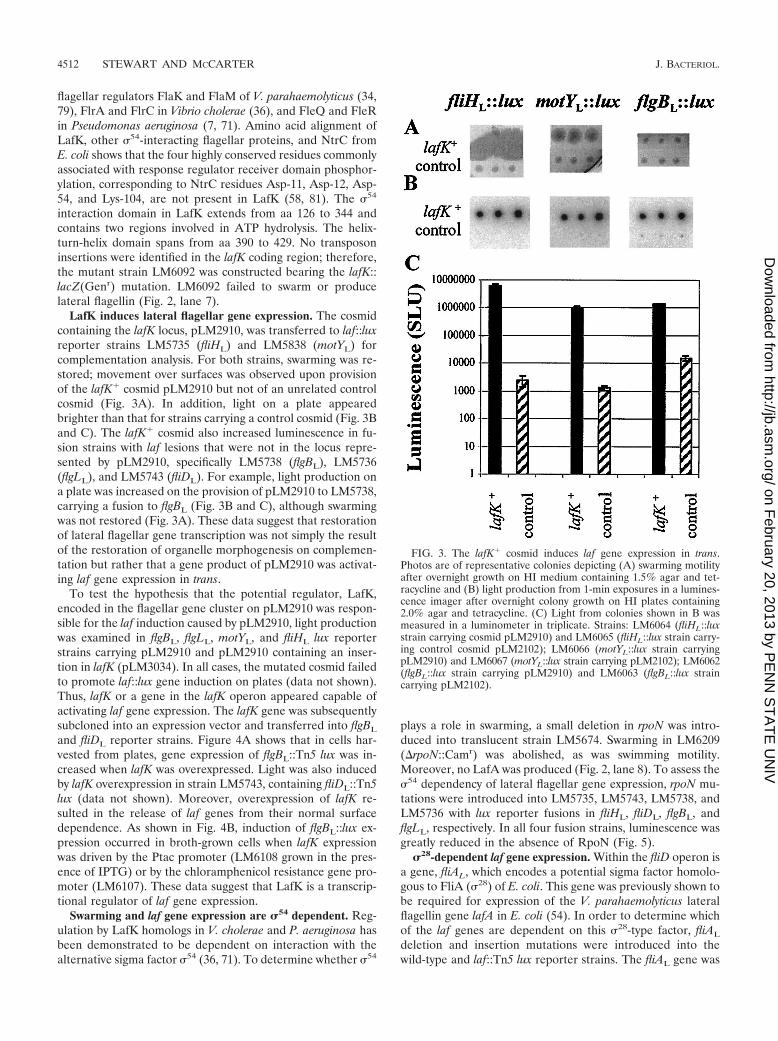
LafK induces lateral flagellar gene expression. The cosmidcontaining the lafK locus, pLM2910, was transferred to laf::luxreporter strains LM5735 (fliHL) and LM5838 (motYL) forcomplementation analysis. For both strains, swarming was re-stored; movement over surfaces was observed upon provisionof the lafK� cosmid pLM2910 but not of an unrelated controlcosmid (Fig. 3A). In addition, light on a plate appearedbrighter than that for strains carrying a control cosmid (Fig. 3Band C). The lafK� cosmid also increased luminescence in fu-sion strains with laf lesions that were not in the locus repre-sented by pLM2910, specifically LM5738 (flgBL), LM5736(flgLL), and LM5743 (fliDL). For example, light production ona plate was increased on the provision of pLM2910 to LM5738,carrying a fusion to flgBL (Fig. 3B and C), although swarmingwas not restored (Fig. 3A). These data suggest that restorationof lateral flagellar gene transcription was not simply the resultof the restoration of organelle morphogenesis on complemen-tation but rather that a gene product of pLM2910 was activat-ing laf gene expression in trans.
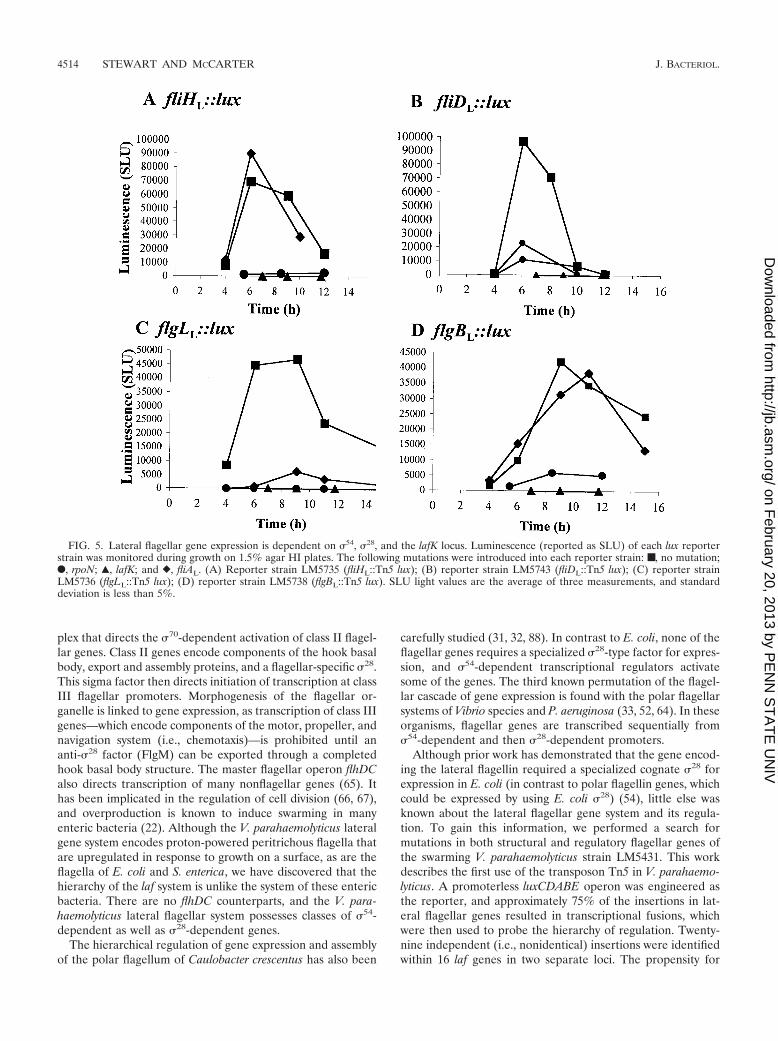
To test the hypothesis that the potential regulator, LafK,encoded in the flagellar gene cluster on pLM2910 was respon-sible for the laf induction caused by pLM2910, light productionwas examined in flgBL, flgLL, motYL, and fliHL lux reporterstrains carrying pLM2910 and pLM2910 containing an inser-tion in lafK (pLM3034). In all cases, the mutated cosmid failedto promote laf::lux gene induction on plates (data not shown).Thus, lafK or a gene in the lafK operon appeared capable ofactivating laf gene expression. The lafK gene was subsequentlysubcloned into an expression vector and transferred into flgBL
and fliDL reporter strains. Figure 4A shows that in cells har-vested from plates, gene expression of flgBL::Tn5 lux was in-creased when lafK was overexpressed. Light was also inducedby lafK overexpression in strain LM5743, containing fliDL::Tn5lux (data not shown). Moreover, overexpression of lafK re-sulted in the release of laf genes from their normal surfacedependence. As shown in Fig. 4B, induction of flgBL::lux ex-pression occurred in broth-grown cells when lafK expressionwas driven by the Ptac promoter (LM6108 grown in the pres-ence of IPTG) or by the chloramphenicol resistance gene pro-moter (LM6107). These data suggest that LafK is a transcrip-tional regulator of laf gene expression.
Swarming and laf gene expression are �54 dependent. Reg-ulation by LafK homologs in V. cholerae and P. aeruginosa hasbeen demonstrated to be dependent on interaction with thealternative sigma factor 54 (36, 71). To determine whether 54
plays a role in swarming, a small deletion in rpoN was intro-duced into translucent strain LM5674. Swarming in LM6209(�rpoN::Camr) was abolished, as was swimming motility.Moreover, no LafA was produced (Fig. 2, lane 8). To assess the54 dependency of lateral flagellar gene expression, rpoN mu-tations were introduced into LM5735, LM5743, LM5738, andLM5736 with lux reporter fusions in fliHL, fliDL, flgBL, andflgLL, respectively. In all four fusion strains, luminescence wasgreatly reduced in the absence of RpoN (Fig. 5).
�28-dependent laf gene expression. Within the fliD operon isa gene, fliAL, which encodes a potential sigma factor homolo-gous to FliA (28) of E. coli. This gene was previously shown tobe required for expression of the V. parahaemolyticus lateralflagellin gene lafA in E. coli (54). In order to determine whichof the laf genes are dependent on this 28-type factor, fliAL
deletion and insertion mutations were introduced into thewild-type and laf::Tn5 lux reporter strains. The fliAL gene was
FIG. 3. The lafK� cosmid induces laf gene expression in trans.Photos are of representative colonies depicting (A) swarming motilityafter overnight growth on HI medium containing 1.5% agar and tet-racycline and (B) light production from 1-min exposures in a lumines-cence imager after overnight colony growth on HI plates containing2.0% agar and tetracycline. (C) Light from colonies shown in B wasmeasured in a luminometer in triplicate. Strains: LM6064 (fliHL::luxstrain carrying cosmid pLM2910) and LM6065 (fliHL::lux strain carry-ing control cosmid pLM2102); LM6066 (motYL::lux strain carryingpLM2910) and LM6067 (motYL::lux strain carrying pLM2102); LM6062(flgBL::lux strain carrying pLM2910) and LM6063 (flgBL::lux straincarrying pLM2102).
4512 STEWART AND MCCARTER J. BACTERIOL.
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
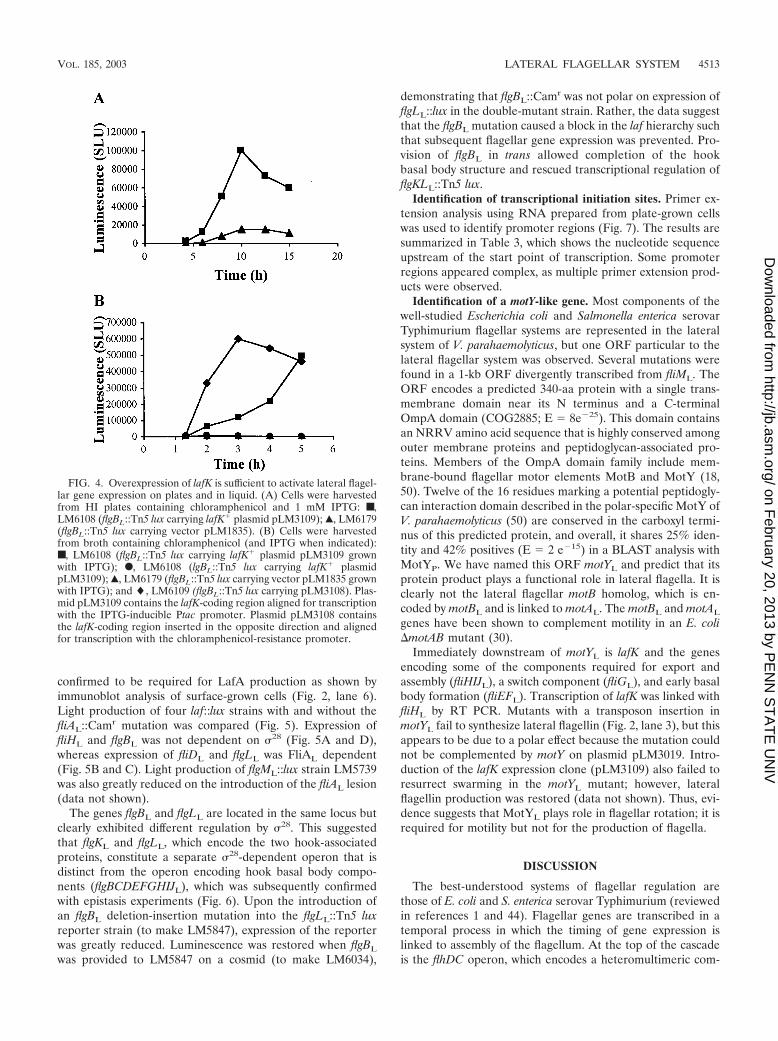
confirmed to be required for LafA production as shown byimmunoblot analysis of surface-grown cells (Fig. 2, lane 6).Light production of four laf::lux strains with and without thefliAL::Camr mutation was compared (Fig. 5). Expression offliHL and flgBL was not dependent on 28 (Fig. 5A and D),whereas expression of fliDL and flgLL was FliAL dependent(Fig. 5B and C). Light production of flgML::lux strain LM5739was also greatly reduced on the introduction of the fliAL lesion(data not shown).
The genes flgBL and flgLL are located in the same locus butclearly exhibited different regulation by 28. This suggestedthat flgKL and flgLL, which encode the two hook-associatedproteins, constitute a separate 28-dependent operon that isdistinct from the operon encoding hook basal body compo-nents (flgBCDEFGHIJL), which was subsequently confirmedwith epistasis experiments (Fig. 6). Upon the introduction ofan flgBL deletion-insertion mutation into the flgLL::Tn5 luxreporter strain (to make LM5847), expression of the reporterwas greatly reduced. Luminescence was restored when flgBL
was provided to LM5847 on a cosmid (to make LM6034),
demonstrating that flgBL::Camr was not polar on expression offlgLL::lux in the double-mutant strain. Rather, the data suggestthat the flgBL mutation caused a block in the laf hierarchy suchthat subsequent flagellar gene expression was prevented. Pro-vision of flgBL in trans allowed completion of the hookbasal body structure and rescued transcriptional regulation offlgKLL::Tn5 lux.
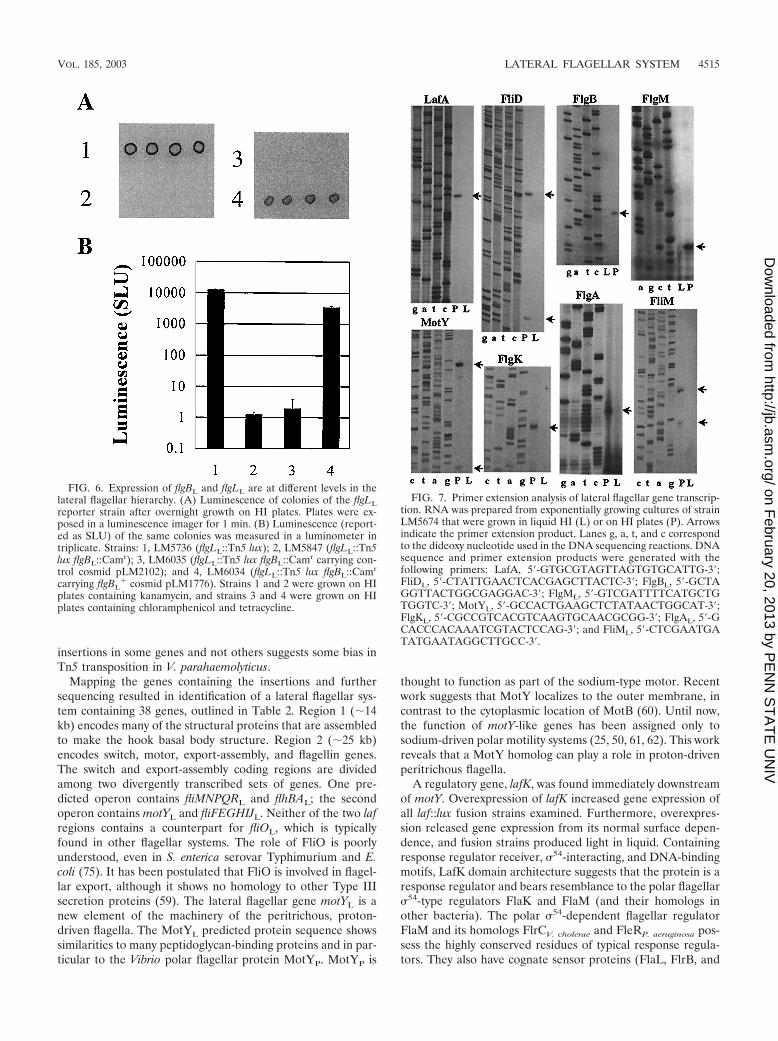
Identification of transcriptional initiation sites. Primer ex-tension analysis using RNA prepared from plate-grown cellswas used to identify promoter regions (Fig. 7). The results aresummarized in Table 3, which shows the nucleotide sequenceupstream of the start point of transcription. Some promoterregions appeared complex, as multiple primer extension prod-ucts were observed.
Identification of a motY-like gene. Most components of thewell-studied Escherichia coli and Salmonella enterica serovarTyphimurium flagellar systems are represented in the lateralsystem of V. parahaemolyticus, but one ORF particular to thelateral flagellar system was observed. Several mutations werefound in a 1-kb ORF divergently transcribed from fliML. TheORF encodes a predicted 340-aa protein with a single trans-membrane domain near its N terminus and a C-terminalOmpA domain (COG2885; E � 8e�25). This domain containsan NRRV amino acid sequence that is highly conserved amongouter membrane proteins and peptidoglycan-associated pro-teins. Members of the OmpA domain family include mem-brane-bound flagellar motor elements MotB and MotY (18,50). Twelve of the 16 residues marking a potential peptidogly-can interaction domain described in the polar-specific MotY ofV. parahaemolyticus (50) are conserved in the carboxyl termi-nus of this predicted protein, and overall, it shares 25% iden-tity and 42% positives (E � 2 e�15) in a BLAST analysis withMotYP. We have named this ORF motYL and predict that itsprotein product plays a functional role in lateral flagella. It isclearly not the lateral flagellar motB homolog, which is en-coded by motBL and is linked to motAL. The motBL and motAL
genes have been shown to complement motility in an E. coli�motAB mutant (30).
Immediately downstream of motYL is lafK and the genesencoding some of the components required for export andassembly (fliHIJL), a switch component (fliGL), and early basalbody formation (fliEFL). Transcription of lafK was linked withfliHL by RT PCR. Mutants with a transposon insertion inmotYL fail to synthesize lateral flagellin (Fig. 2, lane 3), but thisappears to be due to a polar effect because the mutation couldnot be complemented by motY on plasmid pLM3019. Intro-duction of the lafK expression clone (pLM3109) also failed toresurrect swarming in the motYL mutant; however, lateralflagellin production was restored (data not shown). Thus, evi-dence suggests that MotYL plays role in flagellar rotation; it isrequired for motility but not for the production of flagella.
DISCUSSION
The best-understood systems of flagellar regulation arethose of E. coli and S. enterica serovar Typhimurium (reviewedin references 1 and 44). Flagellar genes are transcribed in atemporal process in which the timing of gene expression islinked to assembly of the flagellum. At the top of the cascadeis the flhDC operon, which encodes a heteromultimeric com-
FIG. 4. Overexpression of lafK is sufficient to activate lateral flagel-lar gene expression on plates and in liquid. (A) Cells were harvestedfrom HI plates containing chloramphenicol and 1 mM IPTG: ■,LM6108 (flgBL::Tn5 lux carrying lafK� plasmid pLM3109); Œ, LM6179(flgBL::Tn5 lux carrying vector pLM1835). (B) Cells were harvestedfrom broth containing chloramphenicol (and IPTG when indicated):■, LM6108 (flgB L::Tn5 lux carrying lafK� plasmid pLM3109 grownwith IPTG); F, LM6108 (lgBL::Tn5 lux carrying lafK� plasmidpLM3109); Œ, LM6179 (flgBL::Tn5 lux carrying vector pLM1835 grownwith IPTG); and �, LM6109 (flgBL::Tn5 lux carrying pLM3108). Plas-mid pLM3109 contains the lafK-coding region aligned for transcriptionwith the IPTG-inducible Ptac promoter. Plasmid pLM3108 containsthe lafK-coding region inserted in the opposite direction and alignedfor transcription with the chloramphenicol-resistance promoter.
VOL. 185, 2003 LATERAL FLAGELLAR SYSTEM 4513
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
plex that directs the 70-dependent activation of class II flagel-lar genes. Class II genes encode components of the hook basalbody, export and assembly proteins, and a flagellar-specific 28.This sigma factor then directs initiation of transcription at classIII flagellar promoters. Morphogenesis of the flagellar or-ganelle is linked to gene expression, as transcription of class IIIgenes—which encode components of the motor, propeller, andnavigation system (i.e., chemotaxis)—is prohibited until ananti-28 factor (FlgM) can be exported through a completedhook basal body structure. The master flagellar operon flhDCalso directs transcription of many nonflagellar genes (65). Ithas been implicated in the regulation of cell division (66, 67),and overproduction is known to induce swarming in manyenteric bacteria (22). Although the V. parahaemolyticus lateralgene system encodes proton-powered peritrichous flagella thatare upregulated in response to growth on a surface, as are theflagella of E. coli and S. enterica, we have discovered that thehierarchy of the laf system is unlike the system of these entericbacteria. There are no flhDC counterparts, and the V. para-haemolyticus lateral flagellar system possesses classes of 54-dependent as well as 28-dependent genes.
The hierarchical regulation of gene expression and assemblyof the polar flagellum of Caulobacter crescentus has also been
carefully studied (31, 32, 88). In contrast to E. coli, none of theflagellar genes requires a specialized 28-type factor for expres-sion, and 54-dependent transcriptional regulators activatesome of the genes. The third known permutation of the flagel-lar cascade of gene expression is found with the polar flagellarsystems of Vibrio species and P. aeruginosa (33, 52, 64). In theseorganisms, flagellar genes are transcribed sequentially from54-dependent and then 28-dependent promoters.
Although prior work has demonstrated that the gene encod-ing the lateral flagellin required a specialized cognate 28 forexpression in E. coli (in contrast to polar flagellin genes, whichcould be expressed by using E. coli 28) (54), little else wasknown about the lateral flagellar gene system and its regula-tion. To gain this information, we performed a search formutations in both structural and regulatory flagellar genes ofthe swarming V. parahaemolyticus strain LM5431. This workdescribes the first use of the transposon Tn5 in V. parahaemo-lyticus. A promoterless luxCDABE operon was engineered asthe reporter, and approximately 75% of the insertions in lat-eral flagellar genes resulted in transcriptional fusions, whichwere then used to probe the hierarchy of regulation. Twenty-nine independent (i.e., nonidentical) insertions were identifiedwithin 16 laf genes in two separate loci. The propensity for
FIG. 5. Lateral flagellar gene expression is dependent on 54, 28, and the lafK locus. Luminescence (reported as SLU) of each lux reporterstrain was monitored during growth on 1.5% agar HI plates. The following mutations were introduced into each reporter strain: ■, no mutation;F, rpoN; Œ, lafK; and }, fliAL. (A) Reporter strain LM5735 (fliHL::Tn5 lux); (B) reporter strain LM5743 (fliDL::Tn5 lux); (C) reporter strainLM5736 (flgLL::Tn5 lux); (D) reporter strain LM5738 (flgBL::Tn5 lux). SLU light values are the average of three measurements, and standarddeviation is less than 5%.
4514 STEWART AND MCCARTER J. BACTERIOL.
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
insertions in some genes and not others suggests some bias inTn5 transposition in V. parahaemolyticus.
Mapping the genes containing the insertions and furthersequencing resulted in identification of a lateral flagellar sys-tem containing 38 genes, outlined in Table 2. Region 1 (�14kb) encodes many of the structural proteins that are assembledto make the hook basal body structure. Region 2 (�25 kb)encodes switch, motor, export-assembly, and flagellin genes.The switch and export-assembly coding regions are dividedamong two divergently transcribed sets of genes. One pre-dicted operon contains fliMNPQRL and flhBAL; the secondoperon contains motYL and fliFEGHIJL. Neither of the two lafregions contains a counterpart for fliOL, which is typicallyfound in other flagellar systems. The role of FliO is poorlyunderstood, even in S. enterica serovar Typhimurium and E.coli (75). It has been postulated that FliO is involved in flagel-lar export, although it shows no homology to other Type IIIsecretion proteins (59). The lateral flagellar gene motYL is anew element of the machinery of the peritrichous, proton-driven flagella. The MotYL predicted protein sequence showssimilarities to many peptidoglycan-binding proteins and in par-ticular to the Vibrio polar flagellar protein MotYP. MotYP is
thought to function as part of the sodium-type motor. Recentwork suggests that MotY localizes to the outer membrane, incontrast to the cytoplasmic location of MotB (60). Until now,the function of motY-like genes has been assigned only tosodium-driven polar motility systems (25, 50, 61, 62). This workreveals that a MotY homolog can play a role in proton-drivenperitrichous flagella.
A regulatory gene, lafK, was found immediately downstreamof motY. Overexpression of lafK increased gene expression ofall laf::lux fusion strains examined. Furthermore, overexpres-sion released gene expression from its normal surface depen-dence, and fusion strains produced light in liquid. Containingresponse regulator receiver, 54-interacting, and DNA-bindingmotifs, LafK domain architecture suggests that the protein is aresponse regulator and bears resemblance to the polar flagellar54-type regulators FlaK and FlaM (and their homologs inother bacteria). The polar 54-dependent flagellar regulatorFlaM and its homologs FlrCV. cholerae and FleRP. aeruginosa pos-sess the highly conserved residues of typical response regula-tors. They also have cognate sensor proteins (FlaL, FlrB, and
FIG. 6. Expression of flgBL and flgLL are at different levels in thelateral flagellar hierarchy. (A) Luminescence of colonies of the flgLLreporter strain after overnight growth on HI plates. Plates were ex-posed in a luminescence imager for 1 min. (B) Luminescence (report-ed as SLU) of the same colonies was measured in a luminometer intriplicate. Strains: 1, LM5736 (flgLL::Tn5 lux); 2, LM5847 (flgLL::Tn5lux flgBL::Camr); 3, LM6035 (flgLL::Tn5 lux flgBL::Camr carrying con-trol cosmid pLM2102); and 4, LM6034 (flgLL::Tn5 lux flgBL::Camr
carrying flgBL� cosmid pLM1776). Strains 1 and 2 were grown on HI
plates containing kanamycin, and strains 3 and 4 were grown on HIplates containing chloramphenicol and tetracycline.
FIG. 7. Primer extension analysis of lateral flagellar gene transcrip-tion. RNA was prepared from exponentially growing cultures of strainLM5674 that were grown in liquid HI (L) or on HI plates (P). Arrowsindicate the primer extension product. Lanes g, a, t, and c correspondto the dideoxy nucleotide used in the DNA sequencing reactions. DNAsequence and primer extension products were generated with thefollowing primers: LafA, 5-GTGCGTAGTTAGTGTGCATTG-3;FliDL, 5-CTATTGAACTCACGAGCTTACTC-3; FlgBL, 5-GCTAGGTTACTGGCGAGGAC-3; FlgML, 5-GTCGATTTTCATGCTGTGGTC-3; MotYL, 5-GCCACTGAAGCTCTATAACTGGCAT-3;FlgKL, 5-CGCCGTCACGTCAAGTGCAACGCGG-3; FlgAL, 5-GCACCCACAAATCGTACTCCAG-3; and FliML, 5-CTCGAATGATATGAATAGGCTTGCC-3.
VOL. 185, 2003 LATERAL FLAGELLAR SYSTEM 4515
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from

FleS, respectively). In addition, phosphorylation has beendemonstrated to be necessary for FlrC-mediated regulation inV. cholerae (15). In contrast, LafK does not possess a cognatesensory protein, nor does it contain the signature highly con-served residues found in the receiver domain of response reg-ulators. In particular, it lacks the site of phosphorylation cor-responding to Asp54 of NtrC. In these respects, LafK is mostsimilar to the polar flagellar regulators FlaKV. parahaemolyticus,FlrAV. cholerae, and FleQP. aeruginosa, which also possess noknown sensor partners and are not believed to be phosphory-lated. Evidence suggests that the activity of nonphosphorylatedFleQ is modified posttranslationally by the binding of the an-tiactivator protein FleN in P. aeruginosa (16). How lafK isregulated, and/or LafK activity modulated, remains to be de-termined.
Swarming and swimming motility were entirely dependenton 54, whereas FliAL (lateral 28) and LafK appear to beswarming-specific regulators, as only swarming was abolished,not swimming, when mutations in the genes were introducedinto the wild-type strain. To dissect the regulatory cascade, luxreporter fusions were employed in combination with mutationsin rpoN, fliAL, and lafK. Upon the introduction of an rpoNlesion, luminescence was greatly reduced in all fusion strainsexamined. Expression of fliHL, motYL, and flgBL was not af-fected by a mutation in fliAL. Expression of fliDL, flgML, andflgLL showed significant FliAL dependence. Primer extensionresults, which identified the start points of transcription andlocalized the promoter regions, were generally consistent withthe fusion analysis. The regions preceding the transcriptioninitiation sites for lafA, fliDL, flgML, and flgKL contained con-served nucleotides that could represent a 28-dependent con-sensus promoter. The promoter region for fliDL appeared tohave two transcriptional starts, a major promoter (i.e., strongprimer extension band) containing sequences resembling the
28-dependent consensus, and a weaker promoter containingsequences that might constitute a 54-type promoter. Such anarrangement would be consistent with the need to have somemiddle gene expression of the fliDL operon because it encodes28 (FliAL). No Tn5 lux fusions were isolated in the potentialoperon defined by fliMNPQRflhBAL, and for this reason theplacement of the operon in the hierarchy is uncertain. Primerextension analysis identified one major and one minor productoriginating upstream of fliML; however, the upstream se-quences do not clearly resemble other promoter regions. Thiswas also true for the major product of motYL. Taken together,these results suggest the backbone of the potential hierarchydiagrammed in Fig. 8. However, it should be emphasized thatfuture work is required to confirm and establish the details ofthe circuitry.
Many bacteria have invested heavily in flagellar systems withrespect to numbers of genes and amount of protein and energyexpended for organelle biogenesis and propulsion. As a result,flagellar systems are carefully regulated, both transcriptionallyand posttranscriptionally, by a number of environmental con-ditions and global regulators (1, 8, 27, 37, 38, 68, 69, 76, 82).Many of these global regulators act to modulate the level ofexpression of the master flagellar operon flhDC, e.g., CAP,OmpR, H-NS, CsrA, and SdiA (12, 39, 42, 78, 85, 86). V.parahaemolyticus is remarkable in that it possesses two distinctflagellar systems and at times, both flagella are simultaneouslyassembled. On contact with surfaces, the polarly flagellatedswimmer cell differentiates to an elongated swarmer cell andproduces hundreds of lateral flagella. The molecular mecha-nism for these cells’ surface recognition and gene expressionreprogramming is not known, although surface sensing islinked to polar flagellar performance and iron starvation (48).Recent work has identified a GGDEF-EAL motif containingprotein that modulates laf gene expression (14). Since the V.
TABLE 3. Lateral flagellar promoter regionsa
Geneb Nucleotide sequencec Distance to start codon
E. coli 54 TGGCA N6-7 TGC(A/t) N8-10 �1V. para 54 TGGC N7 TTGC N11-13 �1flgB ATTTATGGCACGTATCTTGTTTTGTATCTTGGCGT N27 ATGfliD2 GGCGGAACACATAAGGTGGAAAATCTACTTCCT N129 GTGmotY2 TCACCAGGTTCCTCTATCTGGTGACTAACGT N123 ATGflgMd AATTTAATGTTCACCACATCCTTGTCGCCTATTGGATTTAG N23 ATGlafA AAATTATCATTAATGGCAAGCGACCGCCTCTAGTTAGTG N54 ATGfliD1 AATTAAATATTTTCCGCTTTTTTAGTCGCCTTGTCTTATAG N188 GTGflgKd AGTTAAACAAGTGCTTGGAGAAGCCGGTATGACGACCGCT N36 ATGE. coli 28 TAAA N15 GCCGATAAV. para CTAAAG N14 GCCGTTAA28
PfliM1 GTTAAAACTAAAACATCATGAAAAACTCTGTGTAGACTACCGAA N57 ATGfliM2 TGAAAAACTCTGTGTAGACTACCGAATATTAAGTTAGAAGAACT N29 ATGmotY1 CATGCGAAGCTTTTAATAAAAATTAAAAAGGGTTAGTTTCATTTA N207 ATGflgA AAGAGTGGAAACCAAACTTCCGGTTTACGGAAAGCAAA N39 CTGE. coli 70 TTGACA N16-19 TATAAT N5-9
a For comparison, consensus promoters are provided: E. coli 70 (26), E. coli 54 (70), E. coli 28 (43), V. parahaemolyticus polar 28 (34), and V. parahaemolyticus54 (34).
b For the cases in which multiple primer extension products were observed, the major product is designated with the subscript 1 and the minor is designated withthe subscript 2.
c Boldface underlined nucleotides identify the start points of transcription (�1) as determined by primer extension analysis. Underlined sequences indicate potentialconsensus sequences.
d Transcription of flgML and flgKL may also originate from upstream promoters because multiple faint ladders of long abortive primer extension products wereobserved near the top of the sequencing gels.
4516 STEWART AND MCCARTER J. BACTERIOL.
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from

parahaemolyticus lateral system does not possess the FlhDCregulators, how the global coordination of gene expressionconverges to regulate the lateral flagellar hierarchy will bemost interesting to dissect.
ACKNOWLEDGMENTS
We thank Yun-Kyeong Kim and Debbie Noack for their excellentmolecular biology support and Sandford Jaques for critical discussion.We are indebted to Rachel Larsen and Bill Metcalf for their kindprovision of pRL27 and Barry Wanner for � Red.
This work was supported by the National Science Foundation re-search grant MCB0077327 to L.L.M. and the National Institutes ofHealth training grant T32 AI07511 to B.J.S.
REFERENCES
1. Aldridge, P., and K. T. Hughes. 2002. Regulation of flagellar assembly. Curr.Opin. Microbiol. 5:160–165.
2. Alexandre, G., R. Rohr, and R. Bally. 1999. A phase variant of Azospirillumlipoferum lacks a polar flagellum and constitutively expresses mechanosens-ing lateral flagella. Appl. Environ. Microbiol. 65:4701–4704.
3. Alexeyev, M. F. 1995. Three kanamycin resistance gene cassettes with dif-ferent polylinkers. BioTechniques 18:52–56.
4. Allen, R. D., and P. Baumann. 1971. Structure and arrangement of flagellain species of the genus Beneckea and Photobacterium fischeri. J. Bacteriol.107:295–302.
5. Alper, M. D., and B. N. Ames. 1978. Transport of antibiotics and metaboliteanalogs by systems under cyclic AMP control: positive selection of Salmo-nella typhimurium cya and crp mutants. J. Bacteriol. 133:149–157.
6. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller,and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generationof protein database search programs. Nucleic Acids Res. 25:3389–3402.
7. Arora, S. K., B. W. Ritchings, E. C. Almira, S. Lory, and R. Ramphal. 1997.A transcriptional activator, FleQ, regulates mucin adhesion and flagellargene expression in Pseudomonas aeruginosa in a cascade manner. J. Bacte-riol. 179:5574–5581.
8. Arricau, N., D. Hermant, H. Waxin, C. Ecobichon, P. S. Duffey, and M. Y.Popoff. 1998. The RcsB-RcsC regulatory system of Salmonella typhi differ-entially modulates the expression of invasion proteins, flagellin and Vi an-tigen in response to osmolarity. Mol. Microbiol. 29:835–850.
9. Atsumi, T., L. McCarter, and Y. Imae. 1992. Polar and lateral flagellarmotors of marine Vibrio are driven by different ion-motive forces. Nature355:182–184.
10. Baumann, P., and L. Baumann. 1977. Biology of the marine enterobacteria:genera Beneckea and Photobacterium. Annu. Rev. Microbiol. 31:39–61.
11. Belas, R., M. Simon, and M. Silverman. 1986. Regulation of lateral flagellagene transcription in Vibrio parahaemolyticus. J. Bacteriol. 167:210–218.
12. Bertin, P., E. Terao, E. H. Lee, P. Lejeune, C. Colson, A. Danchin, and E.Collatz. 1994. The H-NS protein is involved in the biogenesis of flagella inEscherichia coli. J. Bacteriol. 176:5537–5540.
13. Boles, B. R., and L. L. McCarter. 2000. Insertional inactivation of genesencoding components of the sodium-type flagellar motor and switch of Vibrioparahaemolyticus. J. Bacteriol. 182:1035–1045.
14. Boles, B. R., and L. L. McCarter. 2002. Vibrio parahaemolyticus scrABC, anovel operon affecting swarming and capsular polysaccharide regulation. J.Bacteriol. 184:5946–5954.
15. Correa, N. E., C. M. Lauriano, R. McGee, and K. E. Klose. 2000. Phosphor-ylation of the flagellar regulatory protein FlrC is necessary for Vibrio choleraemotility and enhanced colonization. Mol. Microbiol. 35:743–755.
16. Dasgupta, N., and R. Ramphal. 2001. Interaction of the antiactivator FleNwith the transcriptional activator FleQ regulates flagellar number in Pseudo-monas aeruginosa. J. Bacteriol. 183:6636–6644.
17. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of chromo-somal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad.Sci. USA 97:6640–6645.
18. deMot, R., and J. Vanderleyden. 1994. The C-terminal sequence conserva-tion between OmpA-related outer membrane proteins and MotB suggests acommon function in both gram-postitive and gram-negative bacteria, possi-bly in the interaction of these domains with peptidoglycan. Mol. Microbiol.12:333–334.
19. Engebrecht, J., M. Simon, and M. Silverman. 1985. Measuring gene expres-sion with light. Science 227:1345–1347.
20. Enos-Berlage, J. L., and L. L. McCarter. 2000. Relation of capsular poly-saccharide production and colonial cell organization to colony morphologyin Vibrio parahaemolyticus. J. Bacteriol. 182:5513–5520.
21. Ferrandez, A., A. C. Hawkins, D. T. Summerfield, and C. S. Harwood. 2002.Cluster II che genes from Pseudomonas aeruginosa are required for anoptimal chemotactic response. J. Bacteriol. 184:4374–4383.
22. Fraser, G. M., and C. Hughes. 1999. Swarming motility. Curr. Opin. Micro-biol. 2:630–635.
23. Friedman, A. M., S. R. Long, S. E. Brown, W. J. Buikema, and F. M. Ausubel.1982. Construction of a broad host range cosmid cloning vector and its usein the genetic analysis of Rhizobium mutants. Gene 18:289–296.
24. Givskov, M., L. Eberl, G. Christiansen, M. J. Bendik, and S. Molin. 1995.Induction of phospholipase and flagellar synthesis in Serratia liquefaciens iscontrolled by expression of the master operon flhD. Mol. Microbiol. 15:445–454.
25. Gosink, K. K., and C. C. Hase. 2000. Requirements for conversion of theNa�-driven flagellar motor of Vibrio cholerae to the H�-driven motor ofEscherichia coli. J. Bacteriol. 182:4234–4240.
26. Gross, C. A., C. Chan, A. Dombroski, T. Gruber, M. Sharp, J. Tupy, and B.Young. 1998. The functional and regulatory roles of sigma factors in tran-scription. Cold Spring Harbor Symp. Quant. Biol. 63:141–155.
27. Han, Y. W., M. A. Uhl, S. J. Han, and W. Shi. 1999. Expression of bvgAS ofBordetella pertussis represses flagellar biosynthesis of Escherichia coli. Arch.Microbiol. 171:127–130.
28. Harshey, R. M. 1994. Bees aren’t the only ones: swarming in gram-negativebacteria. Mol. Microbiol. 13:389–394.
29. Harshey, R. M., and T. Matsuyama. 1994. Dimorphic transition in Esche-richia coli and Salmonella typhimurium: surface-induced differentiation intohyperflagellate swarmer cells. Proc. Natl. Acad. Sci. USA 91:8631–8635.
30. Jaques, S., Y. K. Kim, and L. L. McCarter. 1999. Mutations conferringresistance to phenamil and amiloride, inhibitors of sodium-driven motility ofVibrio parahaemolyticus. Proc. Natl. Acad. Sci. USA 96:5740–5745.
31. Jenal, U. 2000. Signal transduction mechanisms in Caulobacter crescentusdevelopment and cell cycle control. FEMS Microbiol. Rev. 24:177–191.
32. Jenal, U., and C. Stephens. 2002. The Caulobacter cell cycle: timing, spatialorganization and checkpoints. Curr. Opin. Microbiol. 5:558–563.
33. Joyt, J., N. Dasgupta, and R. Ramphal. 2002. FleQ, the major flagellar generegulator in Pseudomonas aeruginosa, binds to enhancer sites located eitherupstream or atypically downstream of the RpoN binding site. J. Bacteriol.184:5251–5260.
34. Kim, Y.-K., and L. L. McCarter. 2000. Analysis of the polar flagellar genesystem of Vibrio parahaemolyticus. J. Bacteriol. 182:3693–3704.
35. Kirov, S. M., B. C. Tassell, A. B. T. Semmler, L. A. O’Donovan, A. A. Rabaan,and J. G. Shaw. 2002. Lateral flagella and swarming motility in Aeromonasspecies. J. Bacteriol. 184:547–555.
36. Klose, K. E., and J. J. Mekalanos. 1998. Distinct roles of an alternative sigmafactor during both free-swimming and colonizing phases of the Vibrio chol-erae pathogenic cycle. Mol. Microbiol. 28:501–520.
FIG. 8. Model for the lateral flagellar hierarchy of lateral flagellar gene expression. This scheme, based on lux reporter fusion and primerextension analyses, represents the simplest model for temporal regulation of laf genes. Details of the regulatory cascade remain to be elucidated.No lux fusions were obtained in the fliM operon, so it has not been placed in the hierarchy.
VOL. 185, 2003 LATERAL FLAGELLAR SYSTEM 4517
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from

37. Ko, M., and C. Park. 2000. H-NS-dependent regulation of flagellar synthesisis mediated by a LysR family protein. J. Bacteriol. 182:4670–4672.
38. Ko, M., and C. Park. 2000. Two novel flagellar components and H-NS areinvolved in the motor function of Escherichia coli. J. Mol. Biol. 303:371–382.
39. Kutsukake, K. 1997. Autogenous and global control of the flagellar masteroperon, flhD, in Salmonella typhimurium. Mol. Gen. Genet. 254:440–448.
40. Laemmli, U. K. 1970. Cleavage of structural proteins during assembly of thehead of bacteriophage T4. Nature (London) 227:680–685.
41. Larsen, R. A., M. M. Wilson, A. M. Guss, and W. W. Metcalf. 2002. Geneticanalysis of pigment biosynthesis in Xanthobacter autotrophicus Py2 using anew, highly efficient transposon mutagenesis system that is functional in awide variety of bacteria. Arch. Microbiol. 178:193–201.
42. Lehnen, D., C. Blumer, T. Polen, B. Wackwitz, V. F. Wendisch, and G.Unden. 2002. LrhA as a new transcriptional key regulator of flagella, motilityand chemotaxis genes in Escherichia coli. Mol. Microbiol. 45:521–532.
43. Liu, X., and P. Matsumura. 1996. Differential regulation of multiple over-lapping promoters in flagellar class II operons in Escherichia coli. Mol.Microbiol. 21:613–620.
44. Macnab, R. M. 1996. Flagella and motility, p. 123–145. In F. C. Neidhardt,et al. (ed.), Escherichia coli and Salmonella. American Society for Microbi-ology Press, Washington, D.C.
45. Magariyama, Y., S. Sugiyama, K. Muramoto, I. Kawagishi, Y. Imae, and S.Kudo. 1995. Simultaneous measurement of bacterial flagellar rotation rateand swimming speed. Biophys. J. 69:2154–2162.
46. Magariyama, Y., S. Sugiyama, K. Muramoto, Y. Maekawa, I. Kawagishi, Y.Imae, and S. Kudo. 1994. Very fast flagellar rotation. Nature 371:752.
47. McCarter, L., M. Hilmen, and M. Silverman. 1988. Flagellar dynamometercontrols swarmer cell differentiation of Vibrio parahaemolyticus. Cell 54:345–351.
48. McCarter, L., and M. Silverman. 1990. Surface-induced swarmer cell differ-entiation of Vibrio parahaemolyticus. Mol. Microbiol. 4:1057–1062.
49. McCarter, L. L. 1995. Genetic and molecular characterization of the polarflagellum of Vibrio parahaemolyticus. J. Bacteriol. 177:1595–1609.
50. McCarter, L. L. 1994. MotY, a component of the sodium-type flagellarmotor. J. Bacteriol. 176:4219–4225.
51. McCarter, L. L. 1998. OpaR, a homolog of Vibrio harveyi LuxR, controlsopacity of Vibrio parahaemolyticus. J. Bacteriol. 180:3166–3173.
52. McCarter, L. L. 2001. Polar flagellar motility of the Vibrionaceae. Microbiol.Mol. Biol. Rev. 65:445–462.
53. McCarter, L. L., and M. Silverman. 1987. Phosphate regulation of geneexpression in Vibrio parahaemolyticus. J. Bacteriol. 169:3441–3449.
54. McCarter, L. L., and M. E. Wright. 1993. Identification of genes encodingcomponents of the swarmer cell flagellar motor and propeller and a sigmafactor controlling differentiation of Vibrio parahaemolyticus. J. Bacteriol.175:3361–3371.
55. McClain, J., D. R. Rollo, B. G. Rushing, and C. E. Bauer. 2002. Rhodospi-rillum centenum utilizes separate motor and switch components to controllateral and polar flagellum rotation. J. Bacteriol. 184:2429–2438.
56. Meyer, T. F., N. Mlawer, and M. So. 1982. Pilus expression in Neisseriagonorrhoeae involves chromosomal rearrangement. Cell 30:45–52.
57. Moens, S., M. Schloter, and J. Vanderleyden. 1996. Expression of the struc-tural gene, laf1, encoding the flagellin of the lateral flagella in Azospirillumbrasilense Sp7. J. Bacteriol. 178:5017–5019.
58. Morett, E., and L. Segovia. 1993. The 54 bacterial enhancer-binding proteinfamily: mechanism of action and phylogenetic relationship of their functionaldomains. J. Bacteriol. 175:6067–6074.
59. Ohnishi, K., F. Fan, G. J. Schoenhals, M. Kihara, and R. M. MacNab. 1997.The FliO, FliP, FliQ, and FliR proteins of Salmonella typhimurium: putativecomponents for flagellar assembly. J. Bacteriol. 179:6092–6099.
60. Okabe, M., T. Yakushi, M. Kojima, and M. Homma. 2002. MotX and MotY,specific components of the sodium-driven flagellar motor, colocalize to theouter membrane in Vibrio alginolyticus. Mol. Microbiol. 46:125–134.
61. Okunishi, I., I. Kawagishi, and M. Homma. 1996. Cloning and character-ization of motY, a gene coding for a component of the sodium-driven flagel-lar motor in Vibrio alginolyticus. J. Bacteriol. 178:2409–2415.
62. Ormonde, P., P. Horstedt, R. O’Toole, and D. L. Milton. 2000. Role ofmotility in adherence to and invasion of a fish cell line by Vibrio anguillarum.J. Bacteriol. 182:2326–2328.
63. O’Toole, G. A., and R. Kolter. 1998. The initiation of biofilm formation inPseudomonas fluorescens WCS365 proceeds via multiple, convergent signal-ing pathways: a genetic analysis. Mol. Microbiol. 28:449–461.
64. Prouty, M. G., N. E. Correa, and K. Klose. 2001. The novel 54 and 28-dependent flagellar gene transcription hierarchy of Vibrio cholerae. Mol.Microbiol. 39:1595–1609.
65. Pruss, B. M., X. Liu, W. Hendrickson, and P. Matsumura. 2001. FlhD/FlhC-regulated promoters analyzed by gene array and lacZ gene fusions. FEMSMicrobiol. Lett. 197:91–97.
66. Pruss, B. M., D. Markovic, and P. Matsumura. 1997. The Escherichia coliflagellar transcriptional activator flhD regulates cell division through induc-tion of the acid response gene cadA. J. Bacteriol. 179:3818–3821.
67. Pruss, B. M., and P. Matsumura. 1996. A regulator of the flagellar regulonof Escherichia coli, flhD, also affects cell division. J. Bacteriol. 178:668–674.
68. Pruss, B. M., and A. J. Wolfe. 1994. Regulation of acetyl phosphate synthesisand degradation, and the control of flagellar expression in Escherichia coli.Mol. Microbiol. 12:973–984.
69. Rashid, M. H., N. N. Rao, and A. Kornberg. 2000. Inorganic polyphosphateis required for motility of bacterial pathogens. J. Bacteriol. 182:225–227.
70. Reitzer, L., and B. L. Schneider. 2001. Metabolic context and possible phys-iological themes of 54-dependent genes in Escherichia coli. Microbiol. Mol.Biol. Rev. 65:422–444.
71. Ritchings, B. W., E. C. Almira, S. Lory, and R. Ramphal. 1995. Cloning andphenotypic characterization of fleS and fleR, new response regulators ofPseudomonas aeruginosa which regulate motility and adhesion to mucin.Infect. Immun. 63:4868–4876.
72. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
73. Sar, N., L. McCarter, M. Simon, and M. Silverman. 1990. Chemotacticcontrol of the two flagellar systems of Vibrio parahaemolyticus. J. Bacteriol.172:334–341.
74. Scheludko, A. V., E. I. Katsy, N. A. Ostudin, O. K. Gringauz, and V. I.Panasenko. 1998. Novel classes of Azospirillum brasilense mutants with de-fects in the assembly and functioning of polar and lateral flagella. Mol. Gen.Mikrobiol. Virusol. 4:33–37.
75. Schoenhals, G. J., M. Kihara, and R. M. MacNab. 1998. Translation of theflagellar gene fliO of Salmonella typhimurium from putative tandem starts. J.Bacteriol. 180:2936–2942.
76. Shi, W., Y. Zhou, J. Wild, J. Adler, and C. A. Gross. 1992. DnaK, DnaJ, andGrpE are required for flagellum synthesis in Escherichia coli. J. Bacteriol.174:6256–6263.
77. Silverman, M., R. Showalter, and L. L. McCarter. 1991. Genetic analysis inVibrio. Methods Enzymol. 204:515–536.
78. Soutourina, O., A. Kolb, E. Krin, C. Laurent-Winter, S. Rimsky, A. Danchin,and P. Bertin. 1999. Multiple control of flagellum biosynthesis in Escherichiacoli: role of H-NS protein and the cyclic AMP-catabolite activator proteincomplex in transcription of the flhDC master operon. J. Bacteriol. 181:7500–7508.
79. Stewart, B. J., and L. L. McCarter. 1996. Vibrio parahaemolyticus FlaJ, ahomologue of FliS, is required for production of a flagellin. Mol. Microbiol.20:137–149.
80. Stewart, B. S., J. Enos-Berlage, and L. L. McCarter. 1997. The lonS generegulates swarmer cell differentiation of Vibrio parahaemolyticus. J. Bacte-riol. 179:107–114.
81. Stock, J. B., A. M. Stock, and J. M. Mottonen. 1990. Signal transduction inbacteria. Nature (London) 344:395–400.
82. Takeda, S., Y. Fujisawa, M. Matsubara, H. Aiba, and T. Mizuno. 2001. Anovel feature of the multistep phosphorelay in Escherichia coli: a revisedmodel of the RcsC3YojN3RcsB signalling pathway implicated in capsularsynthesis and swarming behaviour. Mol. Microbiol. 40:440–450.
83. Thompson, J. D., D. G. Higgins, and T. J. Gibson. 1994. CLUSTAL W:improving the sensitivity to progressive multiple sequence alignment throughsequence weighting, position-specific gap penalties and weight matrix choice.Nucleic Acids Res. 22:4673–4680.
84. Vogel, H. J., and D. M. Bonner. 1956. Acetylornithinase of Escherichia coli:partial purification and some properties. J. Biol. Chem. 218:97–102.
85. Wei, B. L., A. M. Brun-Zinkernagel, J. W. Simecka, B. M. Pruss, P. Babitzke,and T. Romeo. 2001. Positive regulation of motility and flhDC expression bythe RNA-binding protein CsrA of Escherichia coli. Mol. Microbiol. 40:245–256.
86. Wei, Y., J. M. Lee, D. R. Smulski, and R. A. LaRossa. 2001. Global impactof sdiA amplification revealed by comprehensive gene expression profiling ofEscherichia coli. J. Bacteriol. 183:2265–2272.
87. Woo, T., A. Cheng, and J. Ling. 1992. An application of a simple method forthe preparation of bacterial DNA. BioTechniques 13:696–697.
88. Wu, J., and A. Newton. 1997. Regulation of the Caulobacter flagellar genehierarchy; not just for motility. Mol. Microbiol. 24:233–239.
4518 STEWART AND MCCARTER J. BACTERIOL.
on February 20, 2013 by P
EN
N S
TA
TE
UN
IVhttp://jb.asm
.org/D
ownloaded from
Recommended