
Kloning och expression av arsR från Ideonella dechloratans
- Cloning and expression of arsR from Ideonella dechloratans
Bartal Fimti Mikladal
Fakulteten för Hälsa, Natur- och teknikvetenskap
Biovetenskapliga programmet: Bioteknik
C-uppsats 15 hp Biokemi
Handledare: Maria Rova
Examinator/Granskare: Anna Smedja Bäcklund
Datum: 2017-12-13

2
Sammanfattning
Klorat som avfallsprodukt från pappersindustrin har lett till miljöproblem på grund av klorats
toxiska verkan på växter och alger, och har även lett till bekymmer för människohälsan där
klorat avfallet har kommit i kontakt med dricksvatten. För att åtgärda detta så har mycket
forskning gjorts på bakterier med förmågan att reducera klorat till syrgas och kloridjoner, en
anaerob process som vissa naturligt förekommande bakterier kan utföra. Med ökad kunskap
om regleringen av denna kloratreducerande process, kan dessa bakterier utgöra en effektiv
reningsprocess av pappersbrukens avloppsvatten. Ideonella dechloratans är en sådan bakterie,
den har ett genkluster som kodar för de kloratreducerande enzymerna. Nedströms för dessa
finns en arsR-sekvens som tros att koda för en transkriptionsfaktor; och ytterligare information
om denna transkriptionsfaktor kan möjligen bidra till förståelse av genuttrycket hos den
kloratreducerande funktionen. Syftet med detta arbete är att transformera expressionsceller
med förmågan att uttrycka arsR, så framtida försök kan göras för att identifiera potentiella
bindningssäten för ArsR-proteinet. arsR-sekvensen amplifierades med primrar specifika för
ändarna hos arsR, sekvensen ligerades i en pET-21a(+) vektor från Novagen och
transformerades med BL21 (DE3) expressionsceller. Med en IPTG inducering kunde ett stort
uttryck av olösligt ArsR observeras. Komplikationer med resultatet och framtida
tillvägagångsätt diskuteras.

3
Abstract
Chlorate as a waste product from the paper industry has caused environmental problems due
its toxic effect on plants and algae and is also a concern for human health where chlorate has
contaminated the tap water supplies. To address this issue, a great deal of research has been
carried out on naturally occurring bacteria that can anaerobically reduce chlorate to oxygen and
chloride ions. With additional knowledge of how this chlorate-reducing process is regulated,
these bacteria may one day provide an effective purification process of wastewater from paper
mills. Ideonella dechloratans is such a bacterium that has a gene cluster which encodes the
chlorate-reducing enzymes. Downstream of this cluster is an arsR-sequence believed to encode
a transcription factor, which could aid in the understanding of the gene expression for the
chlorate-reducing operon. The goal of this research is to transform expression cells with the
ability to express the arsR-sequence so that future trials can be made to identify any potential
binding sites for the ArsR-protein. The arsR-sequence was amplified with primers specific to
the ends of the arsR-sequence. The sequence was then ligated into a pET-21a (+) vector from
Novagen and transformed with BL21 (DE3) expression cells. By IPTG inducing these
transformants it was possible to observe a significant expression of insoluble ArsR.
Complications with the outcome and future approaches are discussed.

4
Förkortningar
bp Baspar
ChIP Chromatin immunoprecipitation
Klorat ClO3-
Klorit ClO2-
Klordioxid ClO2
SDS
PCR
Natriumdodecylsulfat
Polymeraskedjereaktion

5
SAMMANFATTNING ......................................................................................................................................... 2
ABSTRACT ........................................................................................................................................................ 3
FÖRKORTNINGAR ............................................................................................................................................ 4
INLEDNING ...................................................................................................................................................... 6
MATERIAL OCH METOD ................................................................................................................................. 11
2.1 PRIMERDESIGN ............................................................................................................................................ 11
2.2 DNA-EXTRAKTION ..................................................................................................................................... 11
2.3 KONTROLL AV PRIMRAR .............................................................................................................................. 11
2.4 PCR AMPLIFIERING ..................................................................................................................................... 12
2.5 KLYVNING AV PCR PRODUKT ..................................................................................................................... 12
2.6 VEKTORPREPARATION ................................................................................................................................ 12
2.7 KLYVNING AV VEKTOR ............................................................................................................................... 13
2.8 LIGERING .................................................................................................................................................... 13
2.9 TRANSFORMERING AV E. COLI BL21(DE3) ................................................................................................. 13
2.10 KONTROLL AV TRANSFORMATIONEN ........................................................................................................ 14
2.11 IPTG INDUCERING .................................................................................................................................... 14
2.12 CELLYSERING ............................................................................................................................................ 14
2.13 PROTEINANALYS MED SDS-PAGE ........................................................................................................... 15
RESULTAT & DISKUSSION ............................................................................................................................... 15
3.1 PRIMERDESIGN ............................................................................................................................................ 15
3.2 ARSR AMPLIFIERING .................................................................................................................................... 16
3.3 KLYVNING AV VEKTOR ............................................................................................................................... 18
3.4 LIGERING .................................................................................................................................................... 19
3.5 TRANSFORMERING ...................................................................................................................................... 20
3.6 ARSR EXPRESSION ...................................................................................................................................... 21
SLUTSATS ......................................................................................................................................................... 24
REFERENSER .................................................................................................................................................. 25
APPENDIX 1. .................................................................................................................................................. 28
APPENDIX 2. .................................................................................................................................................. 29
APPENDIX 3. .................................................................................................................................................. 30

6
Inledning
Nutidens samhälle präglas i stor utsträckning av debatter om globala miljöproblem och vilka
åtgärder som behövs, det har lett till mycket forskning för att åtgärda dessa bekymmer. Ett av
många tilltagande problem är klorat och kloritavfall i miljön runt i världen, där den primära
källan är rening av vatten med klordioxid, som bildar klorat och klorit som en biprodukt (1).
Den andra källan är industriell tillverkning där klorat används till framställning av ogräsmedel,
avlövningsmedel, behandling av läder, färgämnen, tändstickor och explosiva ämnen (2), medan
den problematiken som berör Sverige mest är användningen av klordioxid som blekningsmedel
i den stora pappersindustrin (3). Det har lett till att mycket klorat och klorit har läckt ut i miljön,
där de framför allt har ansamlat sig i grundvatten och sjöar eftersom de är vattenlösliga, samt
väldigt stabila, så håller de sig länge i det fria (4). Ansamlingen av dessa ämnen i grundvatten
har ställt till en problematik för människor där dricksvatten dras ur jorden. Även om klorat och
klorit bryts ner i stor utsträckning vid intag med hjälp av saliv och magsäckens innehåll, så
absorberas resterande mängder snabbt in i plasman där de sprider sig runt kroppen innan de
utsöndras genom urinen. Djurstudier har primärt visat att klorit påverkar kroppens erytrocyter,
medan klorat har samma effekt i viss utsträckning (5). Däremot har studier inte kunnat visa
några signifikanta cancerogena egenskaper eller reproduktiva defekter hos klorat behandlade
försöksdjur (5). Men det är tydligt att dessa ämnen har en toxisk effekt, eftersom många
kloratförgiftningar har rapporterats i samband med användningen av kloratbaserat ogräsmedel
(6). Patienterna visade en funktionsnedsättning i röda blodkropparnas förmåga att transportera
syre och även en minskning i antalet blodkroppar (7). Mängden klorat som ställer till
hälsoeffekter skiljer sig mycket i de olika studierna, beroende på försöksdjurens ras och
behandlingsmetod, därför har man också gjort mänskliga studier, där försökspersoner fick
dricka doser upp till 2,4 mg klorat, tillsammans med ytterligare försökspersoner som skulle
konsumera 36 µg per kilo kroppsvikt om dagen i 12 veckor. Inga av dessa klorathalter visade
någon signifikant effekt. Därför räknas lägsta observerade effektnivån av klorat att vara 36
µg/kg om dagen, jämfört med dödliga dosen som anses att vara 230 mg/kg. US Enviromental
Protection Agency har därför bestämt att högsta mängden klorat i dricksvatten får vara 210
µg/l (8) och för att förstå vad detta innebär, så kan man jämföra med kloratmätningar runt i
USA, där 26 av 74 vattensystem överskred rekommenderade nivån och på några platser var
halterna av klorat högre än 1000 µg/l (9). Hälsoeffekten av klorat är kanske inte särskilt
betydelsefull i de mängderna som förekommer i dricksvatten. Men det gör inte problemet
mindre, eftersom den största effekten av klorat i miljön ses i sjöar, där klorats starka oxidativa

7
effekt gör att den är väldigt toxisk för växter och alger även vid låga koncentrationer (10). Detta
har lett till att klorat som miljöavfall nu erkänns som ett problem som måste lösas (11).
I pappersindustrin har man övergått till att använda klordioxid i större utsträckning, istället för
klorgas, men även det skapar lite klorat som biprodukt (12). Så för att verkligen få kontroll på
problemet har man utvecklat olika metoder för att rena klorat från vatten. Det finns flera
metoder, var vissa drar nytta av omvandling genom reduktion såsom biologisk reducering,
kemisk reducering och elektrokemisk reducering. Alla metoderna har sina fördelar och
nackdelar, varav de nämnda metoderna är ofta dyra, eftersom kemikalierna kan vara
kostsamma (13). Däremot finns det andra tillvägagångssätt som använder sig av anjonväxling,
membranfiltrering och elektrodialys, dessa metoder är billigare men behöver däremot en viss
renhet på vattnet för att fungera optimalt och för att minska behovet av underhåll (14). Av
denna anledning betraktas de därför som mindre lämpliga till att användas som en extern
behandling i ett pappersbruk där mycket slam och annat förekommer i avloppet som ska
behandlas (15). Därför har man börjat att fokusera mer på biosanering eller så kallad biologisk
reducering, där bakterier genom sin respiration kan reducera klorat till kloridjoner. Själva
bakterierna är billiga och sköter sig själva, så det behövs inte mycket underhåll, förutom att
eventuellt tillsätta lite kväve och fosfor som näring, vilket det vanligen inte förekommer så
mycket av i ett pappersbruks avfallsvatten (15). Det förekommer redan användning av bakterier
i reningsverk, som genom aerob-reducering kan ta hand om olika avfallsprodukter (12). Men
eftersom de är aeroba, så behövs en ständig luftning av avloppsvattnet för att bibehålla denna
funktion. Detta gör det till en väldigt energikrävande process som ofta står för mer än 50 % av
den förbrukade elenergin hos reningsverket, dessutom producerar bakterierna mycket slam
som biprodukt i processen (13). Anaerob reducering kan ses som en lösning till detta, eftersom
den kan bilda upp till sju gånger mindre slam (13) och inte behöver den energikrävande
tillsatsen av syre. Men nackdelen är att det är mycket svårare att bibehålla en effektiv
reducering med anaeroba bakterier, eftersom de har en långsam tillväxt och är oftast ganska
känsliga för ändringar i miljön, vilket också gör att de återhämtar sig långsamt vid eventuella
störningar (12). Lyckas man att komma över dessa problem, så har man enligt dessa källor en
metod med mycket potential.
Bakterier med förmågan att respirera klorat förekommer naturligt i naturen vilket tyder på att
det måste finnas en naturlig kloratkälla. Man har hittat klorat överallt på jorden, även på ställen
där mänskligt kloratutsläpp är väldigt osannolikt (16). Det visar sig nämligen att klorat kan

8
bildas i atmosfären, för att sedan föras ner på jorden med regnet (17). Detta har gett upphov till
välutvecklade kloratreducerande bakterier. Dessa bakterier delar en gemensam funktion, det är
att växa anaerobt med klorat som elektronacceptor (18). De använder sig först av enzymet
kloratreduktas för att reducera klorat till klorit, följt av en reducering av klorit till kloridjoner
och syre, med hjälp av kloritdismutas (19).
𝐶𝑙𝑂3− + 2𝐻+
𝑘𝑙𝑜𝑟𝑎𝑡𝑟𝑒𝑑𝑢𝑘𝑡𝑎𝑠→
2𝑒− 𝐶𝑙𝑂2
− + 𝐻2𝑂 ; 𝐶𝑙𝑂2−
𝑘𝑙𝑜𝑟𝑖𝑡𝑑𝑖𝑠𝑚𝑢𝑡𝑎𝑠→
𝐶𝑙− + 𝑂2
Reaktionsformel 1.
Denna reducering sker i elektrontransportkedjan hos bakterien (20), som en del av
cellandningen. Att använda dessa bakterier till biosanering gör att det krävs en anaerob miljö.
Detta krav gör det svårare att ha en jämn och effektiv reducering av klorat, eftersom det är svårt
att hålla en vattenmiljö helt anaerob, speciellt då syre är en biprodukt av klorat-reduceringen.
Därför har kontrollen av metabolismen blivit ett intressant forskningsområde, i hopp om att en
dag kunna få cellerna att reducera klorat även i en aerobmiljö. Detta hade gjort biosanering
effektiv och billig nog till kommersiellt bruk.
Figur 1. Genkluster i Ideonella dechloratans som kodar för kloratreducerande ämnen och andra cellulära funktioner.
I 1994 upptäcktes bakterien Ideonella dechloratans. En gramnegativ, stavformad, fakultativt
anaerob bakterie med förmågan att växa anaerobt med klorat som elektronacceptor och aerobt
med syre (21). Mycket forskning har utförts på I. dechloratans för att förstå hur denna
reducering av klorat till kloridjoner utförs. Ett genkluster har sekvenserats, som kodar för
kloritdismutas (cld) och kloratreduktas (clrABDC); enzymerna som bryter ner klorat till
kloridjoner och syrgas i en tvåstegsreaktion (Reaktionsformel 1.) (22). Kloritdismutas (cld)
ligger längst uppströms relativt till de andra kodande sekvenserna i genklustret som visas i
figur 1 och skiljs åt av ett insertionselement (ISIde1) från den kloratreduktas-kodande
sekvensen (clrABDC) där clrD tros att vara ett chaperon med funktion att integrera kofaktorer
rätt vid veckningen av Clr (22). Mellan clrABDC och mobB är en gensekvens för cytokrom c,
som skulle kunna ha en elektrondonerande funktion till kloratreduktas, men studier har visat

9
att detta inte är fallet (23) (24). Hur själva uttrycket av dessa gener kontrolleras är fortfarande
oklart.
arsR-sekvensen som ligger nedströms för cyc och mobB, tros att vara en transkriptionsfaktor
av ArsR familjen. Det som är kännetecken för ArsR familjen är en homodimer med helix-turn-
helix motiv, vars funktion är att reglera de intracellulära nivåerna av olika metalljoner, för att
kunna reagera mot påfrestningar inducerade av metalltoxicitet (25). Detta görs möjligt med
hjälp av ArsR som normalt är bundet till operatorn/promotorn hos resistensoperonet. Kommer
cellen i kontakt med de metalljoner som kan leda till metalltoxicitet, så vill dessa binda sig
kovalent till ArsR, vilket leder till en allosterisk reglering av ArsR (26). Detta i sin tur
nedreglerar/inhiberar DNA-bindningsförmågan hos ArsR så att operaton/promotorn blir
tillgänglig för polymeraset, så transkriptionen av resistensoperonet kan utföras som svar till
metalltoxiciteten (25). Detta är den generella uppfattningen av ArsR, men det finns mer än
3000 ArsR liknande transkriptionsfaktorer enligt UniProt databasen, där få har egentligen
analyserats noga för deras specifika funktion. Så det upptäcks nya egenskaper inom ArsR
superfamiljen hela tiden. Till att börja med hade man enbart visat att ArsR hade en specificitet
emot metaljonerna Zn(II) och As(III), men nu inkluderas även Cd(II), Pb(II), Bi(III), Co(II)
och Ni(II) (25). Studier har också visat att den intracellulära miljön i bakterien har stor
betydelse för vilken specificitet ArsR egentligen har in vivo jämfört med in vitro observationer
(25). En studie från 2011 har till och med visat att ArsR i växtpatogenerna Xylella fastidiosa
och Agrobacterium tumefaciens genomgår en konformationsändring vid reducering och
oxidering som påverkar dess interaktion med operonet, och fungerar därför som en sorts redox
switch (27). arsR i Ideonella dechloratans som nämnt tidigare ligger nedströms för det
kloratreducerande operonet, består av 387 baspar (bp) som kodar för 128 aminosyror. Genom
att utföra ett BLAST av denna sekvens erhålls resultat som tyder på stora likheter med ArsR
superfamiljen, med flera olika aminosyror som tillsammans kan utgöra DNA-bindande säten
och även en ledtråd om tre eventuella Zn(II) bindningssäten, dock har dessa aminosyror inte
en position som är typisk för ArsR metall-bindande proteiner (28). Det finns även en annan
sekvens i I. dechloratans som har likheter med arsR familjen enligt BLAST undersökningar,
men eftersom genomet hos I. dechloratans är sekvenserat i form av kontiger, så går det inte att
tala om vilken position i genomet denna arsR-sekvens har relativt till andra kodande sekvenser,
dessutom har denna sekvens en likhet under 30 % med arsR som ligger nedströms för det
kloratreducerande operonet, så den kan inte klassificeras som en homolog.

10
Det som gör arsR i I. dechloratans intressant i det här sambandet, är att under år 2012
publicerades en artikel där det hade forskat om en arsR-sekvens (Rv2034) i Mycobacterium
tuberculosis (29). Detta ArsR protein visade sig inte att ha någon metallreglerande funktion,
men hade däremot starka DNA-bindnings egenskaper med förmågan att inhibera sin egen
promoter genom inbindning. Med hjälp av en omfattande ChIP-sekvensering kunde även
ytterligare 614 bindningssäten identifieras. Detta tydde på att ArsR eventuellt fungerade som
en så kallad global reglerare. Det som gör det särskilt intressant är att åtta av dessa
målsekvenser var gener som blir uttryckta vid hypoxiska förhållanden. Om ArsR i I.
dechloratans delar samma egenskaper som ArsR från M. tuberculosis, så kan det vara ett stort
steg i rätt riktning för att ta reda på mekanismen bakom anaeroba reduceringen av klorat hos I.
dechloratans. Det finns anledningar till att tro att det kan vara så, för det första så tyder BLAST
undersökningen på att primära egenskapen hos ArsR i I. dechloratans är DNA-bindning och
genom att analysera genklustret som kodar för de kloratreducerande enzymerna så finns
likheter med målsekvenser hos ArsR från M. tuberculosis, i cld regionen som kodar för
kloritdismutas, så det kan vara ett potentiellt bindningssäte för ArsR hos I. dechloratans. Det
finns också saker som strider emot att ArsR i I. dechloratans kan ha funktionella likheter med
M. tuberculosis, eftersom den arsR-kodande strängen i I. dechloratans har bara en 31%
sekvens-likhet med arsR (Rv2034) i M. tuberculosis. De sekvenserade proteiner av ArsR
familjen, som delar mycket likheter med ArsR från I. dechloratans är från bakterier så som
Acidithiobacillus thiooxidans, Sulfuricella denitrificans skB26, Acidithiobacillus ferrooxidans.
Men det finns inga studier om funktionen hos ArsR i dessa bakterier. Därför finns ingen ledtråd
om att arsR i I. dechloratans delar funktionella likheter med Rv2034 och detta gör det också
svårt att göra någonting annat än spekulationer omkring rollen hos ArsR i I. dechloratans.
Syftet med detta arbetet var att extrahera genom ur Ideonella dechloratans, designa primrar för
att kunna PCR-amplifiera arsR-sekvensen med hög specificitet och med hjälp av ett
expressionssystem från Novagen skulle arsR-sekvensen sedan uttryckas. Det innebar att
arbetet inleddes med att designa lämpliga primrar till uppdraget. Den amplifierade arsR-
sekvensen skulle därefter ligeras in i en pET-21a(+) vektor från Novgen (Appendix 3), så att
en His-Tag kunde adderas till ena änden av arsR-sekvensen och samtidigt göra IPTG
inducering samt ampicillin-selektivitet möjlig. Den arsR bärande pET-21a(+) vektorn skulle
därefter införas i E. coli BL21(DE3) för att uttrycka införda arsR-sekvensen med His-Tag.
Produkten skulle sedan extraheras ur de expressionskompetenta cellerna och analyseras med
SDS-PAGE för att verifiera att ArsR hade blivit uttryckt, samt bestämma dess löslighet. Med

11
ett positivt utfall, skulle det här arbetet vara en hjälp för framtida försök med syfte att rena fram
ArsR och undersöka eventuella bindningssekvenser.
Material och metod
2.1 Primerdesign
Ett primerpar designades till att amplifiera arsR. Till nedströmsprimern adderades ett
restriktionsenzymsäte för XhoI och ett NdeI säte för uppströmsprimern, så sekvensen skulle
kunna ligeras in i en pET-21a(+) vektor från Novagen. Dessutom krävdes det att hela arsR-
sekvensen blev inkluderad i PCR-amplifieringen, så primern behövde vara komplementär till
ändarna hos arsR och samtidigt leva upp till kraven för optimitet med avseende på
smälttemperaturen på 52-58°C, längden 18-22 bp och CG fördelning 40-50 %. Det gjordes 43
olika förslag för varje primer genom att variera antalet CG på ändarna och antalet
komplementära nukleotider mot templatsekvensen (arsR) med spannet 15-25 bp. De primrar
närmast optimala kraven sorterades i från de andra, för att sedan analyseras ytterligare för deras
benägenhet att bilda hairpin struktur, self-dimer och hetero-dimer med hjälp av OligoAnalyser
3.1 (Integraded DNA Technologies®). De två primrar med mest lik smälttemperatur och
stabilast med avseende på hairpin/dimer sturkturer, såväl som att leva upp till ställda krav,
användes (se resultatdel 3.1 Primerdesign).
2.2 DNA-extraktion
Ideonella dechloratans togs från en fryst kultur och ympades i 5 ml PC-medium (se Appendix
1), för att odlas över natten i en skakinkubator vid 37°C. Därefter gjordes en extraktion av
genomiskt DNA på I. dechloratans-övernattskulturen med NucleoSpin Tissue (MACHEREY-
NAGEL) utförd enligt tillverkarens instruktioner.
2.3 Kontroll av primrar
PCR-amplifiering med två olika annealing-temperaturer utfördes på DNA-extraktet från I.
dechloratans. En med den teoretiskt mest lämpliga temperaturen 56°C och en 2°C varmare, för
att testa vilken gav ett lämpligt utbyte för de designade primrarna. Polymeraset Hoststart Fast
Ready Mix från Kapabiosystems användes med en primerkoncentration på 10 µM, och proven
tillbereds enligt tillverkarens instruktioner med 90 ng templat DNA. Amplifieringen gjordes i

12
en Veriti 96 well thermal cycler från Applied Biosystems med programmet; initial denaturering
95°C 2 min, följt av 35 cykler med 95°C 15 sek, 58°C eller 56°C i 15 sek för respektive prov,
samt 72°C 15 sek, avslutad med 72°C i 7 min. För att se om PCR-reaktionerna ovan gav något
utbyte, gjordes en agarosgel-elektrofores med TAE-buffert (Appendix 1), 1,5 % agaros och
kördes på 75 volt (V) med båda PCR-produkterna som erhölls från amplifieringen (figur 2).
2.4 PCR amplifiering
Stora mängder av arsR målsekvensen behövde amplifieras, eftersom förluster förekommer i
renings- och transformationsprocesserna. Fyra stycken 25 µl prov med 90 ng templat DNA
bereddes med Hotstart Fast Ready Mix enligt tillverkarens instruktioner. Två kontrollprover
gjordes också, ett utan primer och ett annat utan templat-DNA. PCR-programmet kördes med
inställningarna; initial denaturering 95°C 2 min, följt av 35 cykler med 95°C 15 sek, 58°C i 15
sek och 72°C 15 sek, avslutad med 72°C i 7 min. PCR produkterna renades med GeneJet PCR
Purification Kit (Fermentas). Koncentration och renhet bestämdes spektrofotometriskt genom
att mäta absorbansen vid 260 och 280 nm. PCR-produkterna analyserades sedan med agarosgel
elektrofores (TAE-buffert, 1,5 % agaros, 75 V) för att verifiera att reningsprocessen hade
lyckats.
2.5 Klyvning av PCR produkt
Ett prov tillbereddes för klyvning med 2204 ng renad PCR produkt i en volym på 40 µl, genom
att tillsätta 8 µl restriktionsenzym XhoI (10 U/µl), 4 µl NdeI (10 U/µl) från New England
Biolabs, 8 µl buffert-O och 72 µl sterilt avjonat H2O. Klyvningen utfördes i ett 37°C vattenbad
i 3¾ timmar och följdes av en 80°C inaktivering av enzymerna i 20 min. Ytterligare en rening
gjordes med GeneJet PCR Purification Kit (Fermentas) av den kluvna PCR lösningen.
2.6 Vektorpreparation
En vektorpreparation gjordes genom att transformera E. coli XL1 Blue (Stratagene) med
vektorn pET-21a(+) från Novagen, enligt Stratagenes instruktioner. Dessa transformerade
celler odlades övernatt på LB-agarplattor med ampicillin (Appendix 1). Varefter en koloni av
transformerade celler fördes över i ett 14 ml Falconrör med 5 ml LB-medium (Appendix 1)

13
och odlades övernatt med skakning. En plasmidextraktion gjordes sedan på övernattskulturen
med E.Z.N.A. Plasmid DNA kit I (VWR) enligt tillverkarens instruktioner.
2.7 Klyvning av vektor
De extraherade plasmiderna klövs med restriktionsenzymerna XhoI (10 U/µl) och NdeI (10
U/µl) från New England Biolabs, tillsammans med två kontroller med endast XhoI eller NdeI,
för att se om enzymerna klöv vektorn som förväntat. 96 µl med 3403 ng extraherad pET-21a(+)
från Novagen klövs i en reaktionsblandning med 180 µl sterilt avjonat H2O, 20 µl buffer-O, 10
µl NdeI och 20 µl XhoI, i ett 37°C vattenbad i 4 timmar. Klyvningarna analyserades med
agarosgel elektrofores (TAE-buffert, 1 % agaros, 75 V). Varefter den kluvna vektorn
separerades ifrån de okluvna med ytterligare en agarosgel elektrofores körning, med 1711 ng
vektor på gelen. Banden med den kluvna vektorn skars ur gelen och renades från gelen med
Gel Extraction KIT (QIAGEN) enligt tillverkarens instruktioner.
2.8 Ligering
Kluven pET-21a(+)-vektor och kluven PCR-produkt innehållande arsR-sekvensen ligerades
ihop med T4 DNA ligase 5 wiess U/µl (Thermoscientific). Reaktionsblandningen bestod av
1:1 molar fördelning av vektor (95,2 ng) och insert (6,72 ng), 2 µl T4 ligas buffert, 1 µl T4
ligas och 2 µl sterilt avjonat H2O, i en total volym på 20 µl. Ligeringen utfördes vid
rumstemperatur i 1 timme och inaktiverades i ett 65ºC vattenbad i 10 min. Ligeringen
analyserades med agarosgel elektrofores (TAE-buffert, 1 % agaros, 75 V).
2.9 Transformering av E. coli BL21(DE3)
BL21 (DE3) från Agilent Technologies transformerades med pET-21a(+)-vektor innehållande
arsR eller pUC18 som kontroll, enligt tillverkarens instruktioner. Cellerna odlades vid 37ºC
övernatt på LB-agarplattor med 100 µg/ml ampicillin (Appendix 1). Varefter en renodling
gjordes på ytterligare en LB-agarplatta, samt en övernattskultur i LB-medium med 100 µg/ml
ampicillin (Appendix 1).

14
2.10 Kontroll av transformationen
En plasmidpreparation gjordes på BL21(DE3) klonerna med E.Z.N.A. Plasmid DNA kit I
(VWR), enligt tillverkarens instruktioner. Följt av en restriktionsenzyms-klyvning på de
extraherade plasmiderna enligt metoden beskriven i (2.5 Klyvning av PCR produkt). Kluvna
produkten PCR-amplifierades sedan med arsR-specifika primrar. Amplifieringen gjordes
enligt metoden beskriven i (2.4 PCR amplifiering). PCR-produkten analyserades på agarosgel
elektrofores (TAE-buffert, 1,5 % agar, 75 V).
2.11 IPTG inducering
BL21(DE3) kloner innehållande arsR-genen och kloner med pUC18 odlades övernatt i 37°C
på LB-agar med 100 µg/ml ampicillin. En koloni togs från varje odling och löstes i 10 ml LB-
medium med 50 µg/ml ampicillin för att odlas ytterligare en natt. 500 µl av övernattsodlingen
fördes över i ett nytt 50 ml Falconrör med 10 ml LB-medium utan ampicillin och ställdes i en
skakinkubator med temperaturen 37°C i 115 min. OD600 mättes på proven och 2 ml ställdes
på is för att användas som oinducerad prov. Resterande prov delades upp i tre 2 ml prov och
IPTG tillsattes till en slutkoncentration på 1 mM, varefter de inkuberades i 2 timmar vid 37°C.
2.12 Cellysering
De oinducerade och inducerade proven centrifugerades i 10 min med 5000 rpm i rotor SLA-
1500 (Sorvall™). Supernatanten dekanterades och pelleten löstes upp i 0,5 ml PBS-buffer
(Appendix 1) och 1x Protease inhibitor (Appendix 1) tillsattes. Lösningen sonikerades på is
med inställningarna: Power 4, 40 % duty och 10 pulser med Branson Sonifier 450. 3 µl av 10
U/μl DNase tillsattes efteråt, för att undvika för hög viskositet orsakad av DNA. Proven
centrifugerades återigen i 10 min på 14500 rpm i en Eppendorf minispin plus, för att skilja på
lösliga och olösliga proteiner. Supernatanten fördes över i ett nytt Eppendorfrör och användes
som löslig fas. Olösliga pelleten tvättades med 750 µl PBS, följt av en 5min centrifugering på
14500 rpm. Supernatanten kastades, varefter pelleten löstes i 1x SDS Running Buffer
(Thermoscientific) tillberedd enligt tillverkarens instruktioner, vortexades kraftigt och ställdes
i kokande vatten i 5 min. Olösliga proven sonikerades sedan ytterligare en gång med
inställningarna: Power 4, 40 % duty och 12 pulser.

15
2.13 Proteinanalys med SDS-PAGE
10 µl togs i från de sonikerade cellproven och blandades med 1,5 µl Reducing Agent (10x),
samt 3,5 µl LDS Sample Buffer (4x) (Thermoscientific), varefter de omedelbart värmdes till
70°C i 10 min i ett vattenbad. Proven applicerades på en 10 % Bis-Tris gel (Life Technologies)
i 1x MES-buffert (Thermoscientific), med 500 µl antioxidant blandad i överkammaren. Gelen
kördes i 35 min på 200 V. Varefter den färgades med SimplyBlue SafeStain, enligt tillverkarens
instruktioner.
Resultat & Diskussion
3.1 Primerdesign
Genom att jämföra 86 olika primrar specifika för ändarna på arsR med varierande längd på
komplementära sekvensen och CG-svans, så kunde det primerpar med bäst värden utses, med
avseende på smälttemperatur, komplementär längd, CG fördelning och förmågan att binda till
sig själva. Det utvalda primerparet som användes till att amplifiera arsR-sekvensen från I.
dechloratans visas nedan. Baspar som är rödfärgade nedan utgör restriktionsenzyms-
klyvningssäte, till vänster är CG-svans och på högersida är komplementära sekvensen till
ändarna hos arsR.
Uppströms:
5’ CGC GCC ATA TGT TCA TTA ATA TGA ATT CCG 3’
Totalt 30 bp i storlek, med 19 bp komplementära till målsekvensen. 9157,03 g/mol, 40 % CG
och smält temperaturen 66,42°C. primern hade 2 möjliga hairpin strukturer med ett ΔG och
smälttemperatur enligt tabell 1. ΔG värden överskred inte kraven (Max -3 kcal/mol). Däremot
hade uppströms primern förmågan att bilda 24 olika homo-dimer strukturer, varav sex hade för
lågt ΔG, enligt optimala kraven (Max -6 kcal/mol).
Nedströms:
5’ GCC TCG AGA GAT GCC TTC TTA GTT TT 3’
Totalt 26 bp i storlek, med 18 bp komplementära till målsekvensen. 7944,23 g/mol, 46,15%
CG och smält temperaturen 63,42°C. Nedströms primern hade en benägenhet att bilda 3 olika
hairpin strukturer med ett ΔG och smälttemperatur enligt tabell 1. Alla tre hairpin strukturerna
16
hade ett ΔG värde inom det som ansågs optimalt. 13 olika homodimer var också sannolika,
varav en överskred kraven. 25 olika hetero-dimer var möjliga, men alla hade så pass högt ΔG
att sannolikheten för att det skulle ställa till något problem var liten.
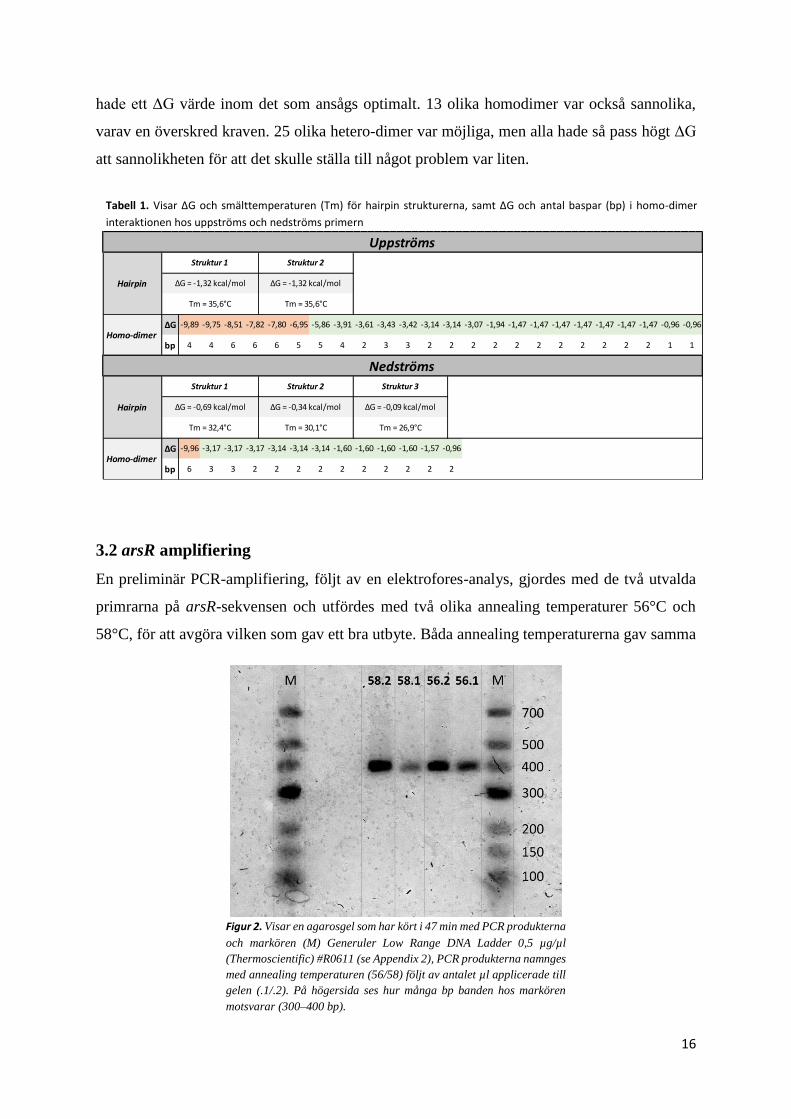
3.2 arsR amplifiering
En preliminär PCR-amplifiering, följt av en elektrofores-analys, gjordes med de två utvalda
primrarna på arsR-sekvensen och utfördes med två olika annealing temperaturer 56°C och
58°C, för att avgöra vilken som gav ett bra utbyte. Båda annealing temperaturerna gav samma
Uppströms
Nedströms
2 2 2 2 2
-0,96
bp 6 3 3 2 2 2 2 2
-3,14 -1,60 -1,60 -1,60 -1,60 -1,57
Struktur 3
ΔG = -0,09 kcal/mol
Tm = 26,9°C
∆G -9,96 -3,17 -3,17 -3,17 -3,14 -3,14
Struktur 1 Struktur 2
ΔG = -0,69 kcal/mol ΔG = -0,34 kcal/mol
Tm = 32,4°C Tm = 30,1°C
Struktur 1
ΔG = -1,32 kcal/mol
Tm = 35,6°C
Struktur 2
ΔG = -1,32 kcal/mol
Tm = 35,6°C
Hairpin
Homo-dimer
Hairpin
Homo-dimer2 2 1 12 2 2 2 2 22 3 3 2 2 2
-0,96
bp 4 4 6 6 6 5 5 4
-1,47 -1,47 -1,47 -1,47 -1,47 -0,96-3,14 -3,14 -3,07 -1,94 -1,47 -1,47-6,95 -5,86 -3,91 -3,61 -3,43 -3,42∆G -9,89 -9,75 -8,51 -7,82 -7,80
Tabell 1. Visar ΔG och smälttemperaturen (Tm) för hairpin strukturerna, samt ΔG och antal baspar (bp) i homo-dimer
interaktionen hos uppströms och nedströms primern
Figur 2. Visar en agarosgel som har kört i 47 min med PCR produkterna
och markören (M) Generuler Low Range DNA Ladder 0,5 µg/µl
(Thermoscientific) #R0611 (se Appendix 2), PCR produkterna namnges
med annealing temperaturen (56/58) följt av antalet µl applicerade till
gelen (.1/.2). På högersida ses hur många bp banden hos markören
motsvarar (300–400 bp).

17
resultat, med band som motsvarade storleken av arsR (figur 2). Annealing temperaturen på
58°C valdes att användas i fortsättningen. Den amplifierade arsR-sekvensen från I.
dechloratans analyserades på agarosgel (figur 3). PCR-produkten (#2) har ett väldigt svagt
band runt 400 bp, bredvid är PCR-produkten efter rening (#1), som visar ett tydligt och tjockt
band runt 400 bp. Analysen utfördes också med tre olika kontrollprov (K1, K2, #3), baserade
på PCR-amplifierade prover utan primrar (K1) eller DNA målsekvens (K2) och ett från en förut
lyckad amplifiering (#3). De teoretisk möjliga komplikationerna med homo-dimer och hetero-
dimer strukturer gav inga märkbara problem. Den renade PCR-produkten (#1) hade ett band
som motsvarade exakt de 400 bp som teoretiskt skulle erhållas, baserade på 381 bp arsR
längden med restriktionsenzyms klyvningssäte och GC-påhäng, som är 19 bp tillsammans.
Detta resultat förstärks också genom att kontrollproverna inte visade någon förorening eller
oväntad amplifiering. Orenade PCR-produkten (#2) hade ett mycket svagt band, men det
berodde på en misslyckad applicering.
Figur 3. Agaros-elektrofores (TAE-buffert, 1,5 % agaros, 75 V) analysen
från PCR-amplifieringen av arsR-sekvensen ur I. dechloratans. M är markör
#R0611 (se Appendix 2) med band storleken angiven med baspar på
högersida. K1 är PCR prov amplifierad utan primer och K2 utan DNA. 1# är
renad PCR produkt från arsR amplifiering och #2 är orenad. #3 är PCR
produkt av en förut lyckat amplifiering av arsR. Svaga bandet hos #2 beror
på en misslyckad applicering av prov.

18
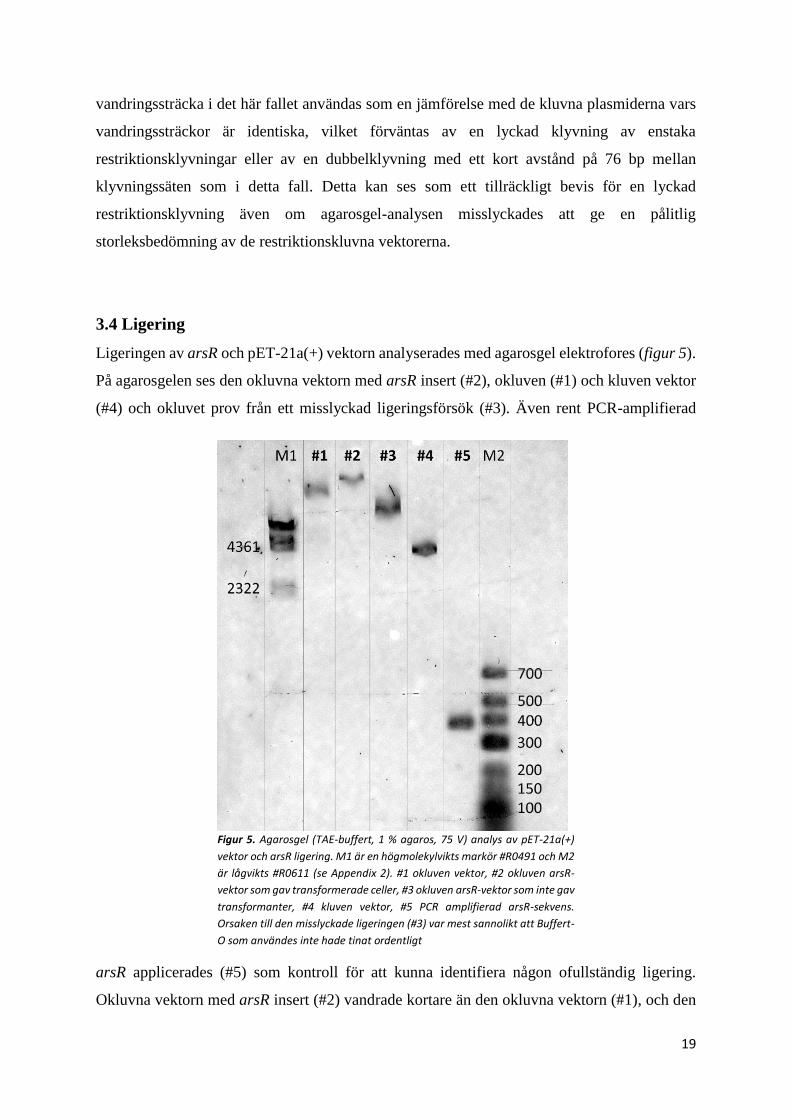
3.3 Klyvning av vektor
En preliminär restriktionsklyvning av vektorn utfördes, för att verifiera att restriktions-
enzymerna fungerade (figur 4). De utfördes genom att göra tre klyvningar, två med varsitt
restriktionsenzym (#1, #2) och ett tredje prov med både NdeI och XhoI (#3). En kontroll i form
av okluven vektor (#4) användes också. Alla klyvningarna (#1,#2,#3) gav ett distinkt band
under 4361 bp markör bandet och kontrollprovet med okluvna vektorn (#4) hade ett tjockt band
större än 9416 bp och två svaga band mellan 4361–9416 bp. Denna 4361 bp storleken på de
kluvna proven avviker väldigt mycket från den riktiga plasmidstorleken som är 5443 bp. En
orsak till dessa stora avvikelser är att markören inte blev värmebehandlad innan applicering,
enligt labprotokollet. Detta kan leda till att ändarna på det linjära λ-DNA:t, som motsvarar 4361
och 23130 bp binder till varandra och cirkulariseras. Med denna komplikation i åtanke, så blir
avvikelsen från den riktiga plasmidstorleken 663 bp istället för 1082 bp. Allt smear som
markören (M) visar, beror på att det applicerades för hög koncentration markör (5000 ng), detta
gör det också mycket svårare att bedöma placeringen hos markörbanden större än 2027 bp.
Bortsett från detta så kan okluvna Plasmiden (4#) observeras att ha vandrat mycket kortare än
de kluvna (#1, #2, #3), vilket kan tyda på att det är en nickad plasmid. Oavsett det, så kan dess
Figur 4. Agaorsgel (TAE-buffert, 1 % agaros, 75 V) analys av pET-21a(+) vektorklyvning
innan ligering. M är markör #R0491 (se Appendix 2). #1 vektor kluven med Ndel, #2 XhoI
kluven, #3 kluven med både NdeI och XhoI. #4 är okluven vektor. Sifforna till höger visar
molekylvikten som markörbanden motsvarar och höger om dessa illustreras
molekylvikten efter korrigeringen på grund av λ-DNA cirkulariseringen.
19
vandringssträcka i det här fallet användas som en jämförelse med de kluvna plasmiderna vars
vandringssträckor är identiska, vilket förväntas av en lyckad klyvning av enstaka
restriktionsklyvningar eller av en dubbelklyvning med ett kort avstånd på 76 bp mellan
klyvningssäten som i detta fall. Detta kan ses som ett tillräckligt bevis för en lyckad
restriktionsklyvning även om agarosgel-analysen misslyckades att ge en pålitlig
storleksbedömning av de restriktionskluvna vektorerna.
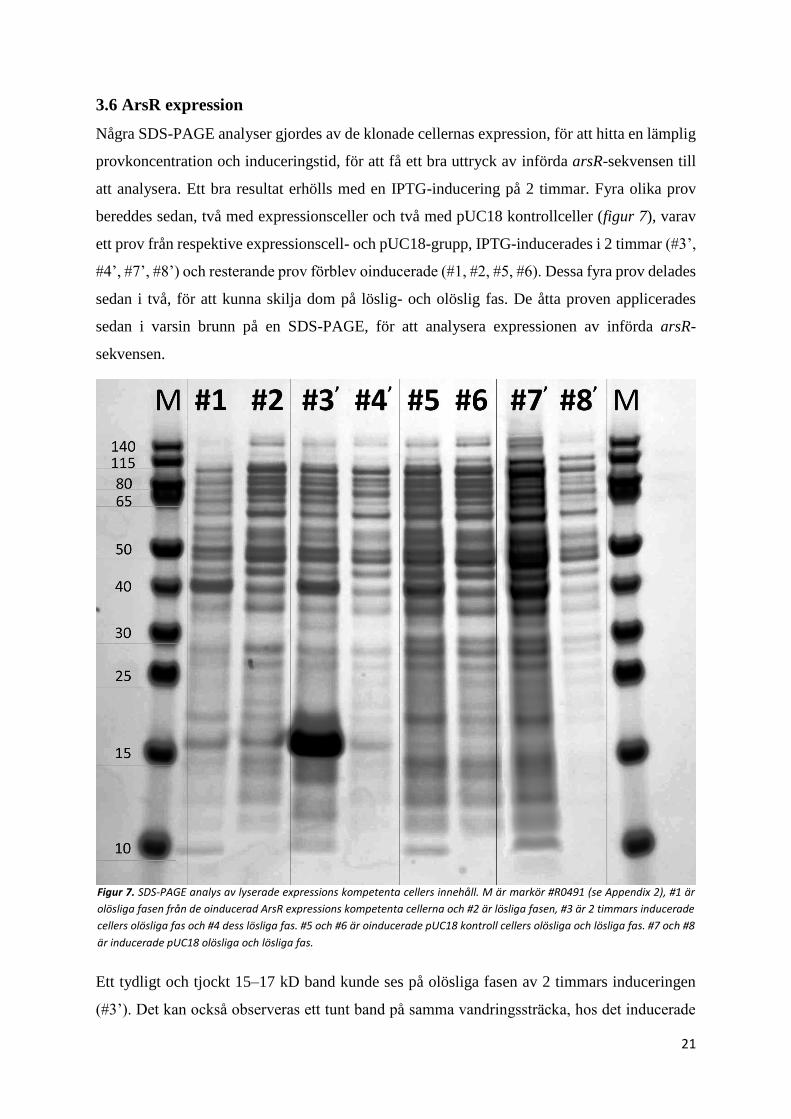
3.4 Ligering
Ligeringen av arsR och pET-21a(+) vektorn analyserades med agarosgel elektrofores (figur 5).
På agarosgelen ses den okluvna vektorn med arsR insert (#2), okluven (#1) och kluven vektor
(#4) och okluvet prov från ett misslyckad ligeringsförsök (#3). Även rent PCR-amplifierad
arsR applicerades (#5) som kontroll för att kunna identifiera någon ofullständig ligering.
Okluvna vektorn med arsR insert (#2) vandrade kortare än den okluvna vektorn (#1), och den
Figur 5. Agarosgel (TAE-buffert, 1 % agaros, 75 V) analys av pET-21a(+)
vektor och arsR ligering. M1 är en högmolekylvikts markör #R0491 och M2
är lågvikts #R0611 (se Appendix 2). #1 okluven vektor, #2 okluven arsR-
vektor som gav transformerade celler, #3 okluven arsR-vektor som inte gav
transformanter, #4 kluven vektor, #5 PCR amplifierad arsR-sekvens. Orsaken till den misslyckade ligeringen (#3) var mest sannolikt att Buffert-
O som användes inte hade tinat ordentligt

20
kluvna vektorn (#4) vandrade längre än alla andra fragment, och med hjälp av arsR kontrollen
(#5) kunde någon form av ligering bekräftas eftersom det inte observerades några fria arsR
ligeringsfragment. Att okluvna vektorn med arsR insert gav ett band som hade vandrat kortare
än den okluvna vektorn, innebär att den förmodligen hade en högre molekylvikt än
ursprungsplasmiden, vilket den borde ha om den har tagit upp arsR.
3.5 Transformering
Tre transformerade kolonier erhölls från det andra ligeringsförsöket. Två av kolonierna
analyserades på agarosgel efter utförd plasmidpreparation kluven med restriktionsenzymerna
NdeI och XhoI som är specifika för införda arsR-sekvensen. Ett distinkt band för varje
klyvning erhölls med en molekylvikt på 400 bp (figur 6), vilket stämmer överens med storleken
på införda arsR-sekvensen som är tillsammans 400 bp, bestående av 381 bp kodande sekvens
och 19 bp restriktionsenzyms klyvningssäte och GC-påhäng. Detta tyder på att de
transformerade cellerna bär på den införda arsR-sekvensen.
Figur 6. Agaorsgel (TAE-buffert, 1,5 % agar, 75 V) analys
av två transformerade cellers kolonier, genom att klyva
införda arsR-sekvensen ut med NdeI och XhoI. M är
lågvikts markör #R0611 (se Appendix 2)., #1 extraherad
arsR från koloni 1 och #2 är från koloni 2.
21
3.6 ArsR expression
Några SDS-PAGE analyser gjordes av de klonade cellernas expression, för att hitta en lämplig
provkoncentration och induceringstid, för att få ett bra uttryck av införda arsR-sekvensen till
att analysera. Ett bra resultat erhölls med en IPTG-inducering på 2 timmar. Fyra olika prov
bereddes sedan, två med expressionsceller och två med pUC18 kontrollceller (figur 7), varav
ett prov från respektive expressionscell- och pUC18-grupp, IPTG-inducerades i 2 timmar (#3’,
#4’, #7’, #8’) och resterande prov förblev oinducerade (#1, #2, #5, #6). Dessa fyra prov delades
sedan i två, för att kunna skilja dom på löslig- och olöslig fas. De åtta proven applicerades
sedan i varsin brunn på en SDS-PAGE, för att analysera expressionen av införda arsR-
sekvensen.
Ett tydligt och tjockt 15–17 kD band kunde ses på olösliga fasen av 2 timmars induceringen
(#3’). Det kan också observeras ett tunt band på samma vandringssträcka, hos det inducerade
Figur 7. SDS-PAGE analys av lyserade expressions kompetenta cellers innehåll. M är markör #R0491 (se Appendix 2), #1 är
olösliga fasen från de oinducerad ArsR expressions kompetenta cellerna och #2 är lösliga fasen, #3 är 2 timmars inducerade
cellers olösliga fas och #4 dess lösliga fas. #5 och #6 är oinducerade pUC18 kontroll cellers olösliga och lösliga fas. #7 och #8
är inducerade pUC18 olösliga och lösliga fas.

22
provets lösliga fas (#4’), samt i respektive oinducerade prov (#1, #2,), men inte hos pUC18
kontrollen (#5, #6, #7’, #8’). Detta tyder på att en viss expression förekommer naturligt utan
inducering, vilket verifierar att expressionscellerna innehåller införda plasmiden, för att den
bör ha en basal expression av T7 RNA polymerase även utan inducering (30). Sannolikheten
är stor att det tjocka 15-17 kD olösliga bandet från 2 timmars induceringen (#3’) är en produkt
av den införda arsR-sekvensen, eftersom den förväntade molekylvikten hos ArsR med His-Tag
är 15,580 kD och chansen att någon annan sekvens skulle ha ett så stort uttryck vid inducering
och ha samma molekylvikt är liten, och kan även delvis uteslutas eftersom den inducerade
pUC18 kontrollen inte gav samma resultat, vilket innebär att det tjocka bandet av intresse är
en produkt av modifieringen av BL21(DE3) med expressionsvektorn pET-21a(+) med införd
arsR-sekvens.
Stora mängden ArsR som ackumulerades med en IPTG inducering på 2 timmar, kan göra
protein reningen av ArsR mycket lättare, eftersom den utgör en så stor procentdel av hela
proteinmängden. Trots det, så är det enbart en fördel så länge inga kvantitativa komplikationer
förekommer, såsom att proteinerna bildar aggregat vilket försvårar reningsprocessen eller att
stora mängder har någon toxisk effekt på expressionsceller. Men med pET-21a(+) vektorn, så
är det möjligt att kontrollera expressionsförmågan ifall det utgör något problem. Det enkla
sättet är det som redan är gjort; att använda vektorn i en DE3 lysogen, eftersom vektorn har en
sekvens som kodar för lac-repressor (lacI) nedströms för T7-promotern och målsekvensen, så
att en expression av målsekvensen också leder till produktionen av lacI, vilket i sin tur minskar
uttrycket av LacUV5 promotorn i värdcellen samt blockerar T7-promotern i vektorn, och på
sätt minskar det både produktionen av T7 RNA-polymeras från värdcellen samt inhiberar i viss
omfattning uttrycket av målsekvensen arsR. Om komplikationerna kvarstår, så kan
expressionen justeras ytterligare genom att införa plasmider med möjligheten att uttrycka T7-
lysozyme (30). Däremot är den största problematiken med bandet som motsvarar ArsR att det
befinner sig i olösliga fasen. Detta utgör en utmaning eftersom separationsprocessen kräver att
proteinet befinner sig i lösliga fasen, och för att få ett olösligt protein lösligt används ett
tillvägagångsätt som består mycket av trial-and-error. I det oinducerade uttrycket av ArsR kan
som tur är en viss mängd ArsR observeras inte bara i olösliga fasen, men samma mängd finns
också i lösliga fasen. Detta innebär att ArsR trivs i löslig form till en viss mängd, därför kan
det vara lämpligt med ett tillvägagångssätt där justeringar görs i odlingen av
expressionscellerna, så hastigheten på arsR-uttrycket minskas. T.ex. reducera temperaturen på
övernattskulturen av expressionscellerna från 37°C till 15–20°C, näringsfattigare

23
odlingsmedium, eventuellt introducera T7-lysozymeuttryckande plasmider eller i värsta fall
byta värdcell. Ett annat tillvägagångsätt som kan kombineras med ovan nämnda är att tillsätta
ämnen som neutraliserar de intramolekylära krafter som gör ArsR olösligt, såsom
reduktionsmedlen 2-mercaptoetanol βME eller dithiothreitol (DTT), kaotropiskamedel urea
eller thiourea eller helt enkelt justera pH och salthalten.
Lyckas man att få en lämpligt mängd ArsR vattenlöslig, då är nästa steg att separera ArsR från
resten av cellproteinerna, detta kan uppnås genom att utnyttja den His-Tag som har adderats på
ena änden av ArsR i detta arbete och med hjälp av affinitetskromatografi som har hög affinitet
för histidin-repeats, kan ArsR selektivt sorteras ifrån de flesta cellproteinerna, för att sedan
elueras ut med imidazole. Om ArsR inte behöver vara i en mycket ren form, så kan en enkel
affinitetskromatografi separation vara tillräcklig, annars kan den användes som preliminär
grovrening efterföljt av andra reningsmetoder för att uppnå högst möjliga renhet. His-Tag
separationen med affinitetskromatografi kräver fortfarande en del optimering för att uppnå en
relativt effektiv rening och ställer korta His-Taggen till några problem i efterföljande analyser,
så kan en proteolytisk klyvning göras för att ta bort den.
Med ArsR i relativt ren form, så är målet att undersöka vilka DNA-sekvenser i I. dechloratans
har förmågan att interagera med ArsR och vilka andra egenskaper ArsR har. Det kan inledas
med att först göra ett försök på specifika sekvenser i I. dechloratans som har likheter med
målsekvenser hos ArsR i M. tuberculosis, som exempel cld-sekvensen i det kloratreducerande
operonet. Om det är en målsekvens för ArsR eller inte, så kan det fortfarande vara värt att göra
en ChIP-undersökning, för att hitta vilka målsekvenser ArsR har i I. dechloratans, eftersom det
kan bidrag till mer information om stora ArsR-superfamiljen. Identifieras en målsekvens för
ArsR är nästa steg att också undersöka om Zn(II), As(III), Cd(II), Pb(II), Bi(III), Co(II) och
Ni(II) har någon påverkan på dess DNA-bindningsförmågan och om ArsR från I. dechloratans
agerar som en homodimer.

24
Slutsats
De BL21(DE3) expressionscellerna som transformerades hade förmågan att uttrycka införda
arsR-sekvensen med ett stort utbyte vid 2 timmars IPTG-inducering. Sonikeringen av
inducerade BL21(DE3) gjorde det möjligt att hitta proteinet uttryckt av införda arsR-sekvens
och kan därför anses som en lämplig metod att extrahera ArsR ur BL21(DE3). Stora mängder
av ArsR observerades i olösliga fasen av det sonikerade extraktet. Detta innebär att det krävs
ett mer tillämpat tillvägagångsätt för att isolera och karaktärisera ArsR i framtida försök. Men
med hjälp av expressionssystemet pET-21a(+) från Novagen som användes, så finns det många
sätt att försöka åtgärda just detta problem. Metoderna som användes för att göra de ArsR
expressions kompetenta celler hade acceptabla materialförluster, varav största
materialförlusten förekom vid transformeringen av BL21(DE3) med arsR-vektorn, där endast
tre transformerade kolonier erhölls.
Resultaten tyder på att expressionen av ArsR har lyckats, och att själva baspar kompositionen
av arsR-sekvensen identifierad i genomet hos Ideonella dechloratans stämmer i en viss mån,
eftersom det designade primerparet som var specifikt för ändarna hos arsR-sekvensen gav
upphov till en produkt med den förväntade storleken 381 bp, däremot blev ingen sekvensering
utförd på den amplifierade arsR-produkten och därför kan den exakta baspar sekvensen inte
verifieras.

25
Referenser
1. Formation of Chlorite and Chlorate Dioxide with Han River Water. Yoon-jin Lee, Hea-tae Kim, Un-
gi Lee. 2004, Korean J, Chem. Eng., pp. 647-653.
2. EPA. Reregistration Eligibility Decision for inorganic Chlorates. s.l. : Enviromental Protection
Agency (EPA), 2006. Case No. 4049.
3. Mechanism of Chlorate Formation During Bleaching of Kraft Pulp with Chlorine Dioxide. Y. NI, G.J.
Kubes and A.R.P. Van Heiningen. 1, s.l. : Journal of Pulp and Paper Science, 1993, Vol. 19.
4. A Review of Chlorine Dioxide in Drinking Water Treatment. Aieta, E. and J.D.Berg. s.l. : Journal of
American Water Works Association (AWWA) 78(6):62-72, 1986.
5. Chlorine Dioxide, Chlorate and Chlorite in Drinking-water. WHO. s.l. : WHO, 2015.
6. (NAS), National Academy of Science. Drinking Water and Health Vol. 7. Washington DC. : National
Academy Press, 1987.
7. —. Drinking Water and Health Vol. 4. Washington, DC : National Academy Press, 1980.
8. Chlorite and Chlorate in Drinking-water, Background document for development of WHO
Guidelines for Drinking-water Quality. WHO, World Health Organisation. 1986, WHO/SDE/WSH.
9. The Potential Regulatory Implications of Chlorate. Alfredo, Katherine, et al. March 2014,
American Water Works Association(AWWA).
10. Effect of pulpmill chlorate on baltic sea algae. A. Rosemarin, K.-J. Lehtinen, M. Notini, J.
Mattson. 85, 1994, environmental pollution, pp. 2-13.
11. A Review of Chlorate- and Perchlorate-Respiring Microorganisms. Logan, Bruce E. s.l. :
Bioremediation Journal 2:69-79, 1998.
12. Persson, P.O. Kompendium i miljöskydd. D. 2, Miljöskyddsteknik : strategier och teknik för ett
hållbart miljöskydd (7 uppl.). s.l. : Stockholm : Institutionen för kemiteknik, Tekniska högskolan,
2005.
13. Jansson, Erik. Extern reningsanläggning för skogsindustriellt avlopp. Karlstads Universitet :
Avdelningen för energi-, miljö- och byggteknik, 2011.
14. Ahn, Se Chang. Removal of perchlorate in ammunition wastewater by zero-valent iron and
perchlorate respiring bacteria. UNIVERSITY OF DELAWARE : Proquest, Umi Dissertation Publishing,
2008.
15. Simulering av COD-reduktionen i en aktivslamanläggning vid en sulfatmassafabrik. Moraeus,
Peter. 2004, Institutionen för informationsteknologi, avdelningen för systemteknik, Uppsala
universitet.

26
16. Simultaneous removal of perchlorate and energetic compounds in munitions wastewater by zero-
valent iron and perchlorate-respiring bacteria. Ahn S. C, Hubbard B, Cha D, Kim B. J. 49, 2014,
Journal of Environmental Science and Health, Vol. Part A, pp. 575-583.
17. The Origin of Naturally Occurring Perchlorate: The Role of Atmospheric Processes. Purnendu K.
Dasgupta, P. Kalyani Martinelango, W. Andrew Jackson, Todd A. Anderson, Kang Tian, Richard W.
Tock, Srinath Rajagopalan. 39, 2005, Environmental Science & technology, pp. 1569-1575.
18. Chlorate reduction capacity and characterisation of chlorate reducing bacteria communities in
sediments of the rio Cruces wetland in southern Chile. Schwarz AO, Urrutia H, Vidal JM, Pérez N. 46,
2012, WATER RESEARCH, pp. 3283-3292.
19. Purification and characterization of (per)chlorate reductase from the chlorate-respiring strain GR-
1. Kengen SW, Rikken GB, Hagen WR, van Ginkel CG, Stams AJ. 181, 1999, Journal of Bacteriology,
pp. 6706-6711.
20. Microbial metabolism of oxochlorates: A bioenergetic perspective. Nilsson T, Rova M, Smedja
Bäcklund A. 2013, Biochimica et Biophysica Acta (BBA), pp. 189-197.
21. Ideonella dechloratans gen. nov., sp.nov., a new bacterium capable of growing anaerobically with
chlorate as an electron acceptor. Malmqvist Å, Welander T, Moore E, Ternström A, Molin G &
Stenström I-M. 17, 1994, System Application Microbiolology, pp. 58-64.
22. A gene cluster for chlorate metabolism in Ideonella dechloratans. Danielsson Thorell H, Stenklo
K, Karlsson J & Nilsson T. 69, 2003, Applied and Environmental Microbiology, pp. 5585-5592.
23. Periplasmic c cytochromes and chlorate reduction in Ideonella dechloratans. Bäcklund AS, Bohlin
J, Gustavsson N & Nilsson T. 75, 2009, Applied and Environmental Microbiology, pp. 2439-2445.
24. Characterization of a cytochrome c gene located at the gene cluster for chlorate respiration in
Ideonella dechloratans. Bohlin J, Bäcklund AS, Gustavsson N, Wahlberg S & Nilsson T. 165, 2010,
Microbiological Research, pp. 450-457.
25. The SmtB/ArsR family of metalloregulatory transcriptional repressors: structural insights into
prokaryotic metal resistance. Laura S. Busenlehner, Mario A. Pennella, David P. Giedroc. s.l. : FEMS
Microbiology Reviews, 2003, FEMS Microbiology Reviews, Vol. 27, pp. 131-143.
26. Allosteric Inhibition of a Zinc-Sensing Transcriptional Repressor: Insights into the Arsenic
Repressor(ArsR) Family. Gregory C. Campanello, Zhen Ma, Nicholas E. Grossoehme, Alfredo J.
Guerra, Brian P. Ward, Richard D. DiMarchi Yuzhen Ye, Charles E. Dann III and David P. Giedroc.
2013, Journal of Molecular Biology, Vol. 425, pp. 1143-1157.
27. Plant Pathogenic Bacteria Utilize Biofilm Growth-associated Repressor (BigR), a Novel Winged-
helix Redox Switch, to Control Hydrogen Sulfide Detoxification under Hypoxia. Beatriz G. Guimarães,
Rosicler L. Barbosa, Adriana S. Soprano, Bruna M. Campos, Tiago A. de Souza. 29, 2011, The Jounral
of Biological Chemistry, Vol. 286, pp. 26148-2657.
28. The SmtB/ArsR family of metalloregulatory transcriptional repressors: structural insights into
prokaryotic metal resistance. Laura S. Busenlehner, Mario A. Pennella, David P. Giedroc.

27
Department of Biochemistry and Biophysics, Center for Advanced Biomolecular Research, 2128
TAMU, Texas ApM University, College Station, : Federation of European Microbiological Societies
(FEMS), 2003, Vol. 27.
29. Chun-hui, Min Yang, Zheng-Guo He*. Characterization of a Novel ArsR-like Regulator Encoded by
Rv2034 in Mycobacterium tuberculosis. PLoS ONE. e36255, 2012, Vol. 7, 4.
30. Novagen. pET System Manual. User Protocol TB055 Rev. C 0611JN. 11th Edition.
31. Cloning, characterisation, and expression of a novel gene encoding chlorite dismutase from
Ideonella dechloratans. Thorell HD, Karlsson J, Portelius E, Nilsson T.
32. Purification and characterization of a soluble cytochrome c capable of delivering electrons to
chlorate reductase in Ideonella dechloratans. T, Bäcklund AS & Nilsson. 321, 2011, FEMS
Microbiology Letters, pp. 115-120.

28
Appendix 1.
PC-medium (1000ml)
- 5g Trypton
- 2,5g Jästextrakt
- 1000ml DH2O
- pH 7 (justerad med HCl eller NaOH)
LB-medium (250ml)
- 2,5g Trypton
- 1,25g Jästextrakt
- 2,5g NaCl
- 250ml DH2O
- pH 7,0 (justerad med HCl eller NaOH)
- evt. 0,01 wt% ampicilin
50x TAE buffert (1000ml)
- 242g Tris acetate
- 100ml 0,5 M EDTA
- 57,1ml Glacial ättiksyra
- Fylls till 1 liter med DH20
- pH 8,3
TE-buffert
- 10mM Tris
- 1 mM EDTA
- pH 8 (justerad med HCl eller NaOH)
1x PBS
- 8,0g NaCl
- 0,2g KCl
- 14,4g Na2HPO4
- 2,4g KH2PO4
- 1 liter DH2O
- pH 7,4 (justerad med HCl eller NaOH)
25x Protease inhibitor
- 1 tablett upplöses i 2ml sterilt avjonat H2O

29
Appendix 2.
Generuler Low Range DNA Ladder 0,5µg/µl (Thermoscientific) #R0611
Lambda-HindIII Marker (Thermoscientific) #R0491
30
Appendix 3.
NOVAGEN pET-21a (+) Vector
Recommended

















