
Journal of South American Earth Sciences, Vol. 6, No. 3, pp. 191-206, 1992 Printed in Great Britain
0895-9811/92 $5.00+.00 © 1993 Pergamon Press Ltd
& Earth Sciences & Resources Institute
First planktonic foraminifera from the Early Cretaceous (Albian) of the Upper Magdalena Valley, Colombia
J. BLAU, 1 L. VERGARA .2 and H. W. STOCK 1
]Institut ftir Angewandte Geowissenschaften, Fachgebiet Palllontologie, Justus-Liebig-Universitiit Giel3en, DiezstraBe 15, 6300 Giel3en, Germany; 2Ingeominas, A. A., 4865 Bogota, Colombia.
(Received July 1992," Revision Accepted March 1993)
Abstraa-.--Albian planktonic foraminifera have been found in the Caballos and "Villeta" formations at two localities in the Upper Magdalena Valley. This is the first documented record of Early Cretaceous planktonic foraminifera in Colombia. Hedbergellids and heterohelieids predominate; keeled forms are absent. The sedimentologie features and the associated microfauna indicate the onset of restricted environments from the middle Albian on.
Resumen--Se deseriben foraminfferos planct6nicos de edad Albiana de las formaciones Caballos y "Villeta" en dos localidades del Valle Superior del Magdalena. El pmsente trabajo eonstituye la primera documentaeion de foraminfferos planet6nicos del Cretticeo temprano en Colombia. La fauna est~ compuesta por hedbergellidos y heteronelieidos. Las formas quilladas estfin ausentes. Esto indica, junto con las caraeterfstieas sedimentol6gicas, condieiones paleoceanogr~ifieas restringidas a partir del Albi- ano medio.
INTRODUCTION
IN THE GEOLOGIC RECORD, the Early Cretaceous was an epoch in which the simple globigeriniform foraminiferal species that first appeared in the Jurassic, or possibly even in the Triassic, began to diversify (Lipps, 1973). Before the Aptian, planktonic foraminifers had particular charac- teristics such as low diversity or small size. They are con- sidexexl primitive, opportunist forms that (re)colonized the oceanic realm after periods of environmental stress (Caron and Homewood, 1983). Their degree of speciation and morphologic evolution is likewise low, in comparison with the morphologically highly specialized and diverse taxa of the Late Cretaceous (see, e.g., Robaszynski et al., 1984; Caron, 1985).
In Colombia, only Late Cretaceous planktonic foramin- ifera have been studied in any detail (e.g., Gandolfi, 1955; Cushman and I-Iedberg, 1941; Martinez, 1989). The only references to Early Cretaceous forms are limited to a few citations summarized below.
Early Cretaceous beds bearing foraminifera were reported in the Eastern Cordillera by Colom (1962), who identified foraminifers in thin sections as Globigerina. He described a sample replete with specimens of this genus, which he believed belonged to the middle Albian, but gave neither species identification nor arguments for the dating of the samples.
A record of the planktonic species Globigerinella escheri (Kaufmann) in middle Albian beds of the Western Cordillera of Colombia, identified by H. Duque-Caro Oageominas, Bogota), was given by Etayo et al. (1980). This taxon is a synonym of Globigerinelloides escheri (Kaufmann) (among many others) according to Masters (1977. p. 411), who gave an "Upper Albian, Turonian- Maastrichtian" range for this species. Considering Etayo's ammonite-supported age determination, the range of the species could be extended to the middle A1bian. In fact, this is the only record of a planktonic Early Cretaceous species in Colombia.
Martlnez (1990) reported Praeglobotruncana stephani and Rotalipora micheli from beds of the Hi16 Formation of the Eastern Cordillera, which also contain Albian ammo- nites, in particular Oxytropidoceras. These are also associ- ated with Rotalipora reicheli, whose biochron lies between late early Cenomanian and late Cenomanian (Stock. 1993). Therefore, the Hil6 Formation and the range of Oxytropidoceras may not be limited to the Albian.
Benthonic Early Cretaceous foraminifera were docu- mented by Petters (1954), in particular the assemblage Choffatella. Epistomina raosquensis, Orbitolina concav- ata texana, and Haplostiche texana, which he assigned to ages between earliest Barremian (?) and late Albian. Althongh some of these assemblages would appear to be
I~-G-f~a~ imalt t
Address all correspondence and reprint requests to Dr. Joachim Blau: telephone [49] (641) 702-8240; fax [49] (641) 702-4711. *Present address of Dipl.-Geol. Luis Vergara: e/o Institut fur Geowissenschaften und Lithospharenforschung, Justus-Liebig-Universit~it Gie6en, Senekenbergstrage 3, 6300 GieBen, Germany
191
192 J. BLAU, L. VERGARA, and H.W. STOCK
-vT,
"-4
e¢
z~
La PlataL~ ~ (f~ / / ~ lqeiva
Subbasin
76 °
Ibague
Bogot~
i Gira[dot Subbasin
'-4 '-4
tg (b
(o
& % 0
I ry
ov
k m
I
1: Quebradas Palrnotosa and Bambuc~i
2: Quebrada Ocal Magdalena Valley
100 I
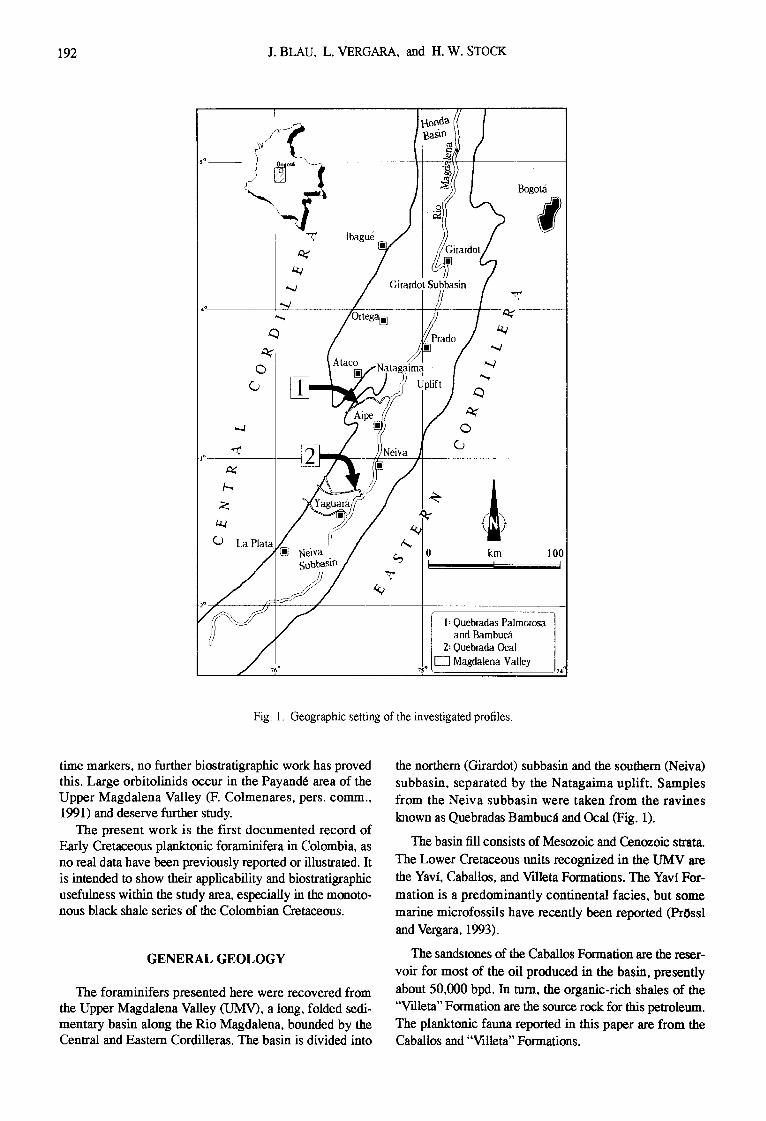
Fig. 1. Geographic setting of the investigated profiles.
time markers, no further biostratigraphic work has proved this. Large orbitolinids occur in the Payand6 area of the Upper Magdalena Valley (F. Colmenares, pers. comm., 1991) and deserve further study.
The present work is the first documented record of Early Cretaceous planktonic foraminifera in Colombia, as no real data have been previously reported or illustrated. It is intended to show their applicability and biostratigraphic usefulness within the study area, especially in the monoto- nous black shale series of the Colombian Cretaceous.
GENERAL GEOLOGY
The foraminifers presented here were recovered from the Upper Magdalena Valley (UMV), a long, folded sedi- mentary basin along the Rio Magdalena, bounded by the Central and Eastern Cordilleras. The basin is divided into
the northern (Girardot) subbasin and the southern (Neiva) subbasin, separated by the Natagaima uplift. Samples from the Neiva subbasin were taken from the ravines lalown as Quebradas Bambuc~ and Ocal (Fig. 1).
The basin fill consists of Mesozoic and Cenozoic strata. The Lower Cretaceous units recognized in the UMV are the Yavi, Caballos, and Villeta Formations. The Yavi For- mation is a predominantly continental facies, but some marine microfossils have recently been reported (PrOssl mid Vergara, 1993).
The sandstones of the Caballos Formation are the reser- voir for most of the oil produced in the basin, presently about 50,000 bpd. In turn, the organic-rich shales of the "Villeta" Formation are the source rock for this petroleum. The planktonic fauna reported in this paper are from the CabaUos and "Villeta" Formations.

First Early Cretaceous planktonic foraminifera, Upper Magdaleaa Valley, Colombia 193
Caballos and "Villeta" Formations
I L,T.o.ooY I[ S~PLEN~-15~O.I ~M11̂ O~l
~ ~_~:-:=:-= ~- =-5=--__=----_ .-- (~¢~ -=-:"="=-----:-:"="
(3 ¢ ~ ~ : - = ~.-_--~-~_=.'
~_=_--.~--::-.~:~.~:
,~ . . . . . . , . . . ~ , . . ~ •
~ z • .-~./.
5-"
.II
,2 ~. C: '3 '3 '3 '= '_- C: ~
c0 250144 C) ~1~
c0
250107 0
t~
250102
<
° ~
250098 0
0 ! . t~
~ 1 ~
Quebradas Palmorosa ] and Bambuc~ /
(Upper Magdalena V.) 1
S i l i c i c l a s t i c s
~ Sandstone
Siltstone
~ Shale/Marl
Carbonates Sparite / Biosparite
~ 1 Micrite/Biomicrite
[ ~ l Biosparudite
~ Siderite
~ - i Calcareous concr.
~ Sideritic concretions
Biogenic remains ~ i Planktonic Forams
~ - - ] Benthonic Forams
~ - - ~ Ammonites
~ - - ~ Bioturbation
~ - ] Gastropods
F ~ Bivalves
Fig. 2. Lithologic column of the CabaUos and "ViUeta" Formations in Quebradas Palmorosa and BambucL
CabaUos Formation "Villeta" Formation
This formation was named by Olsson (1956, p. 307) in the region of Prado Dolores, Tolima. Its age ranges from Aptian to Albian, according to Corrigan (1967) and Bel- t r~ and Gallo (1969), who correlated the Caballos Forma- tion with the Une Formation in the Eastern Cordillera and with the HollfLn Formation in Ecuador. The Caballos For- mation consists mainly of sandstones, with carbonate intercalations, and it records the arrival of the Cretaceous sea in the UMV during the Early Cretaceous. It either unconformably overlaps pre-Cretaceous rocks (Corrigan, 1967) or lies conformably on top of the Yavt Formation where this formation is present. The CabaUos Formation has been mapped as such in the UMV by Ntiflez et al. (1984) and Ftiquen et al. (1989), and as horizon K-9 by Raasveldt and Carvajal (1957a,b).
Hettner (1892) defined the "Villetaschichtea" as slxati- graphically intermediate between the Gir6n and Guada- lupe beds of the Eastern Cordillera and originally assigned it an age of"Neoceaaian to Gault."
The name Villeta Formation has been applied in differ- eat areas of Colombia (and even Venezuela) where Creta- ceous strata of similar lithology crop out (e.g., Grosse, 1930; Stutzer, 1926). In this paper we follow Corrigan (1967, p. 232), who recommended using this name for what he described as a "well defined lithologic unit sand- wiched between the older CabaUos sands and the overly- ing Guadalupe sands." Prevalent lithologies are black shales with carbonate concretions or intercalated carbon- ate beds.
In the UMV, however, physical and temporal limits of the V'flleta Formation (sensu Corrigan, 1967) are different
194 J. BLAU, L. VERGARA, and H.W. STOCK
"Villetd' Formation
0%
I ,,T.OLOOY li S"PLE:-I FMIAoEi
0 .4 0
o ~
• 3MSI . - - .~.~¢
• 11-10-08-90 t~ 0
0
C-~..
(D
~ a..l
• 11-10-07-90 • 11-10-06-90 ID t'~ • II-10-05-90 ~ "~r'~
• 2MSI ~
• 11-10-04-90 " ~ > - : ~.~
• 11-10-03-90
• 11-10"t12-90 .4 • 11-10-01-90
• IMSI <
I l O m l
I " ~ I
¥ I U r n |
Quebrada Ocal (Upper Magdalena
Valley) SECTION ACCORDING TO GROSSER
(PERS COMM. 1991}
Siliciclastics
Sandstone
Siltstone
~ Shale/Marl
Carbonates
Massive Limestone
Irregular Bedded Lst.
~ Sandy Limestone
Biogenic Remains ~ Ammonites
[ ~ - - ~ Bivalves
[ ~ - i Planktonic Forams
V ~ i Benthonic Forams
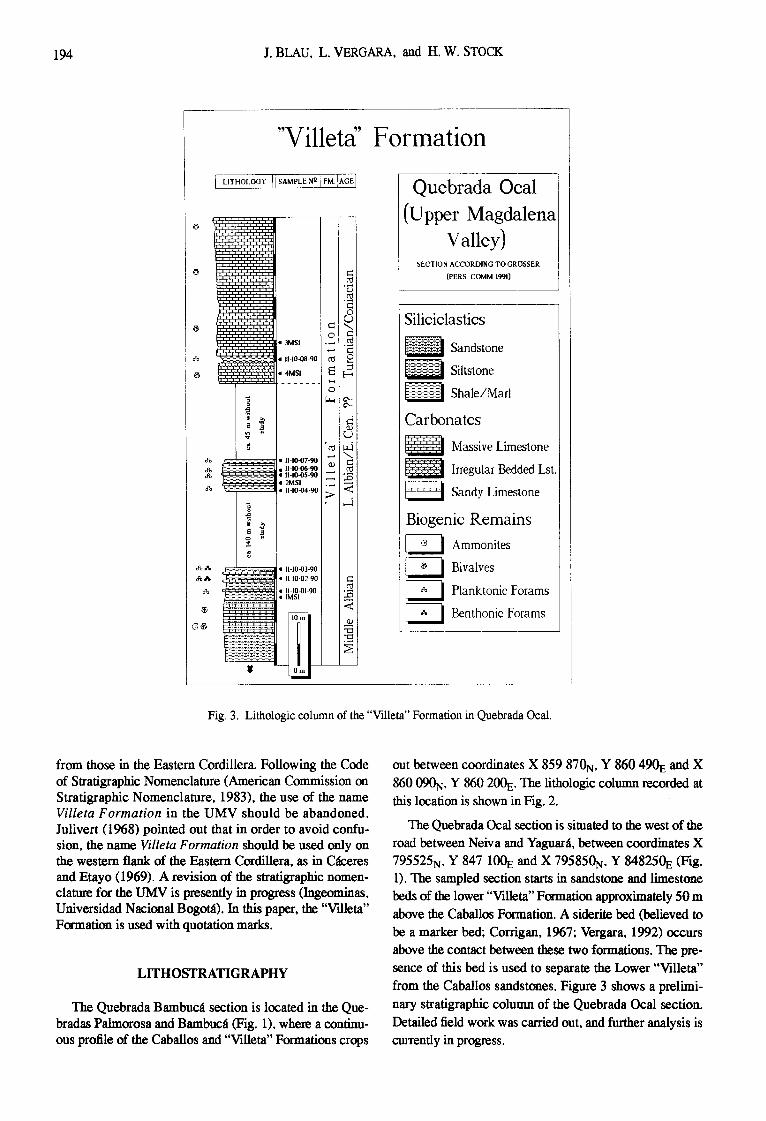
Fig. 3. Lithologic column of the "Villeta" Formation in Quebrada Ocal.
from those in the Eastern Cordillera. Following the Code of Stratigraphic Nomenclature (American Commission on Stratigraphic Nomenclature, 1983), the use of the name Villeta Formation in the UMV should be abandoned. Julivert (1968) pointed out that in order to avoid confu- sion, the name Villeta Formation should be used only on the western flank of the Eastern Cordillera, as in C~eres and Etayo (1969). A revision of the stratigraphic nomen- clature for the UMV is presently in progress (Ingeominas, Universidad Nacional Bogotti). In this paper, the "V'flleta" Formation is used with quotation marks.
LITHOSTRATIGRAPHY
The Quebrada Bambucli section is located in the Que- bradas Palmorosa and Bambuclt (Fig. 1), where a continu- ous profile of the Caballos and "Villeta" Formations crops
out between coordinates X 859 870N, Y 860 490E and X 860 090 N, Y 860 200E. The lithologic col-mn recorded at this location is shown in Fig. 2.
The Quebrada Ocal section is situated to the west of the road between Neiva and Yaguar~l, between coordinates X 795525N, Y 847 100E and X 795850N, Y 848250E (Fig. 1). The sampled section starts in sandstone and limestone beds of the lower "Villeta" F~rmation approximately 50 m above the Caballos Formation. A siderite bed (believed to be a marker bed; Corrigan, 1967; Vergara, 1992) occurs above the contact between these two formations. The pre- sence of this bed is used to separate the Lower "Villeta" from the Caballos sandstones. Figure 3 shows a prelimi- nary stratigraphic column of the Quebrada Ocal section. Detailed field work was carried out, and further analysis is cmTently in progress.

First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley, Colombia 195
Cabailos Formation BIOSTRATIGRAPHY
In the Quebrada Bambuc~l section, the CabaUos Forma- tion can be divided into five lithologic segments (from oldest to youngest):
1. Lithic, feldspathic, or quartz arenites, where sideritic concretions appear in light-colored mudstones.
2. Black shales and intercalated carbonates containing the first appearance of metazoans - - in particular, lamelli- branchs. The carbonates are biosparites and biomicrites and contain unoriented bivalve shells. The shale inter- calations of this segment have yielded the earliest Cre- taceous planktonic foraminifers.
3. Predominartfly quartz arenites with minor intercalations of siltstones, mudstones, and stratified or concretionary siderite.
4. Another succession of black shales and stratified car- bonates, especially biomicrites and biosparites and some thin sideritic layers. Fossils found include plank- tonic as well as benthonic foraminifera, ostracods, and abundant ichnofossils.
5. Littoral deposits of sandstones and interlayered black mudstones with flasex and other typical intertidal sedi- mentary structures.
"'Villeta" Formation
In the Quebrada Bambuc~i section, the lower boundary of the "Villeta" Formation is an even, normal surface between fine-grained sandstones of the CabaUos Forma- tion and black laminated mudstones. The "V'illeta" Forma- tion represents a monotonous series of black shales and limestones. Sideritic beds containing volcanic rock flag- merits and glauconite occur in the lowest part of the for- marion. The remainder of the formation is composed of a thick series of black shales commonly intercalated with oil-bearing layers of biomicrite. Many of these have a wavy. lenticular or concretionary morphology. Thin, locally discontinuous micrite laminae commonly altexnate with the black shales. Large carbonate concretions (for- merly called "Wagon Wheels") are found in this unit.
In the part of the "Villeta" Formation studied at Que- brada Ocal, the unit begins with a series of sandstones. intercalated with sandy shales and fossiliferous lime- stones. Ammonites and oysters are abundant. This series is overlain by black shales intercalated with limestone layers tenths of a meter thick. The shales contain calcareous con- cretions. About 140 m upward in the section, 15 m of black shales were sampled. These are intercalated with thin limestcme layers containing inoceramids and dinosaur bones. Some 45 m higher, the section continues with irreg- ular bedded bluish-grey limestones with large nodular structures. These limestones are overlain by a 2-m-thick black shale intercalation, overlain in turn by massive grey fossiliferons limestones in beds a meter or more thick.
Preparation Methods
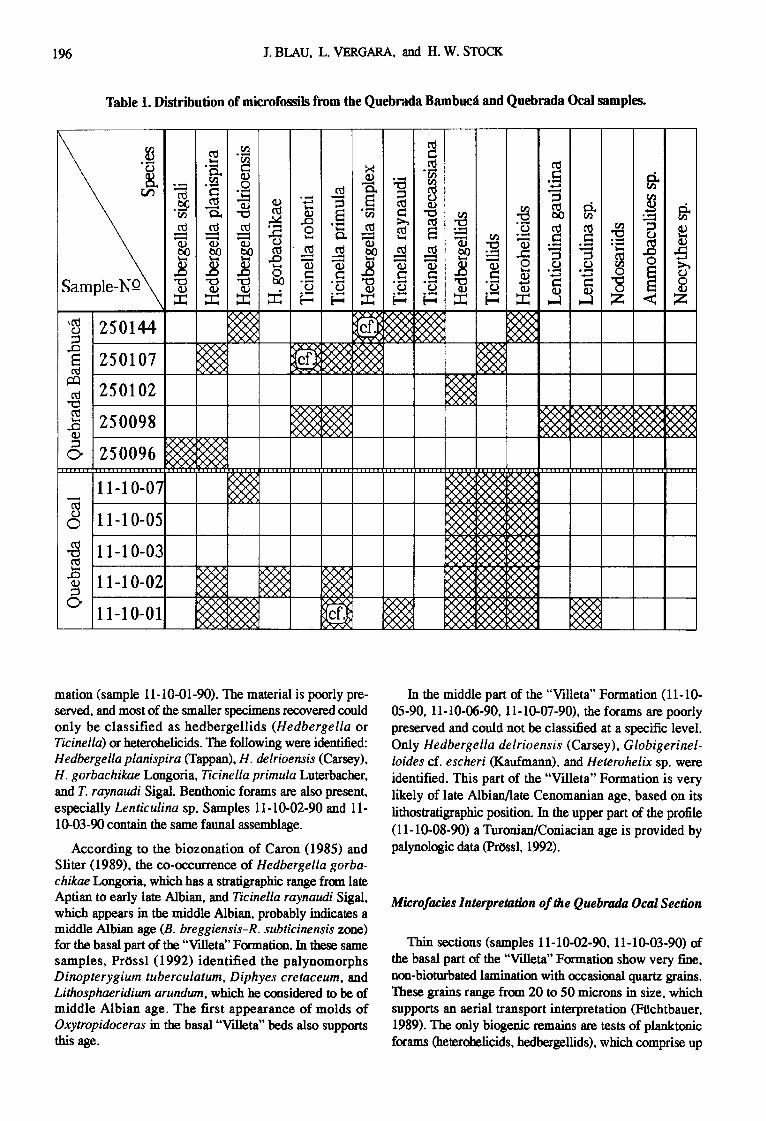
Microfossils of the Bambuc,'i section were prepared at Ingeominas using the traditional Quaternary-O method (see Lipps, 1973). Material collected from the Ocal sec- tion was treated with 1-1202 at Giel3en University. Other than these two kinds of preparation, there were no major differences in preservation of the specimens. Additionally, the foraminifers were examined in thin sections. Table 1 shows the distribution of microfossils.
Quebrada Bambucd Section
The oldest planktonic foraminifers in the Cretaceous succession are from Segment 2 (sample 250096) of the Caballos Formation. Most of the taxa can only be classi- fied as hedbergellids, but we could identify Hedbergella sigali (MouUade) (see Fig. 6.1).
According to Caron's biozonation (1985), this species has a stratigraphic range from Barremian to "middle" Aptian; however, in Sliter's zonation (1989), H. sigali becomes extinct in the middle Albian. Palynologic data (PtOssl and Vergara, 1993) from the same sample reveal an early Albian age. This supports a range of H. sigali at least into the early Albian.
The next planktonic foraminifera are from Segment 4 (sample 250098), where Ticinella cf. primula Luterbacher and T. roberti (Gandolfi) (see Fig. 6) appear in association with other indeterminable hedbergellids. Benthonic fora- minifera are also present, in particular Lenticulina gault- ina (Berthelin). The identification of the associated ostracod Neocythere sp. is worth mentioning, as it has potential stratigraphic value in Lower Cretaceous strata in Colombia (Martinez, written comm. 1991). Other asso- ciated fossils include bivalves and fish scales.
Numerous small hedbergellids are found in the lower "Villeta" Formation (sample 250102). Higher in the col- unto, sample 250107 contains an abundance of Ticinella cf. roberti (Gandolfi), T. primula Luterbacher, Hedber- gella planispira Tappan. and H. simplex (Morrow). This association allows no better stratigraphic resolution than Albian. However, in a sample (250144) higher in the col- umn, the appearance of T. madecassiana Sigal together with Heterohelix sp. indicates a late Albian age (R. ticin- ensis-R, appenninica zones). This age is valid for the uppermost part of the Bambuc~t section displayed in Fig. 2. The Heterohelix individuals are a primitive, dwarfed fauna, probably indicating restricted conditions. The dat- ing of the early and late Albian permits us to infer the position of the middle Albian. We believe this to be the age of the Caballos-"Villeta" boundary at this locality.
Quebrada Ocal Section
The first recorded planktonic foraminifera occur above the lowest sandstones and limestones of the "V'dleta" For-
SAES~6/3--G
196 J. BLAU, L. VERGARA, and H.W. STOCK
Table I. Distribution of microfossils from the Quebrada BambucA and Quebrada Ocal samples.
-: •
• . _
Sample-NQ :d [.: N N N ~ ,..a ~ Z <
250107 250102
~ 250098
~" 250096 ~ ~ 11-10-07
11-10-05
11-10-03 "~ 11-10-02
° 11-1O-Ol N N
®
mation (sample 11-10-01-90). The material is poorly pre- served, and most of the smaller specimens recovered could only be classified as hedbergellids (Hedbergella or Ticinella) or heterohelicids. The following were identified: Hedbergella planispira (Tappan), H. delrioensis (Carsey), H. gorbachikae Longoria, Ticinella primula Luterbacher, and T. raynaudi Sigal. Benthonic forams are also present, especially Lenticulina sp. Samples 11-10-02-90 and 11- 10-03-90 contain the same faunal assemblage.
According to the biozonation of Caron (1985) and Sliter (1989), the co-occurrence of Hedbergella gorba- chikae Ixragoria, which has a stratigraphic range from late Aptian to early late Albian, and Ticinella raynaudi Sigal, which appears in the middle Albian, probably indicates a middle Albian age (B. breggiensis-R, subticinensis zone) for the basal part of the "ViUeta" Formation. In these same samples, PrOssl (1992) identified the palynomorphs Dinopterygium tuberculatum, Diphyes cretaceum, and Lithosphaeridium arundum, which he considered to be of middle Albian age. The first appearance of molds of Oxytropidoceras in the basal "ViUeta" beds also supports this age.
In the middle part of the "Villeta" Formation (11-10- 05-90, 11-10-06-90, I1-10-07-90), the forams are poorly preserved and could not be classified at a specific level. Only Hedbergella delrioensis (Carsey), Globigerinel- loides cf. escheri (Kaufmann), and Heterohelix sp. were identified. This part of the "Villeta" Formation is very likely of late Albian/late Cenomanian age, based on its lithostratigraphic position. In the upper part of the profile (11-10-08-90) a Turonian/Coniacian age is provided by palynologic data (PrOssl, 1992).
Microfacies Interpretation of the Quebrada Ocal Section
Thin sections (samples 11-10-02-90. 11-10-03-90) of the basal part of the "Villeta" Formation show very fine. non-bioturbated lamination with occasional quartz grains. These grains range from 20 to 50 microns in size, which supports an aerial transport interpretation (Ftlchtbauer. 1989). The only biogenic remains are tests of planktonic forams (heterohelicids, hedbergellids), which comprise up

First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley, Colombia 197
to 10% of the sediment. Nevertheless, washed samples also yielded some rare benthonic forams (see Fig. 4.12).
The planktonic forams axe very small, ranging from 60 to 120 microns (see, e.g., Figs. 7.7 and 7.8). Many of the tests are filled with clay minerals or framboidal pyrite (cf Figs. 7.4 and 7.6), which is also scattered throughout the sediment.
The small size of the planktonic forams might be due to shallow marine conditions (Berger, 1971) and/or to a stressed ecologic environment in the water column (Caron and Homewood, 1983). However, the presence of ben- thonic forams suggests periods of better paleoxygenation.
Thin sections from the middle part of the profile (11- 10-04-90 through 11-10-07-90) also show a very fine lamination without biomrbation. Quartz is absent, whereas planktonic forams (hedbergellids, heterohelicids) are occasionally rock-forming. These forams are approxi- mately twice as large as those from the basal part of the profile (see Figs. 7.1, 7.2, 7.3). The chambers are filled with sparry calcite. One thin section (see Fig. 7.9) shows a cyclic alternation of layers consisting exclusively of forams and organic-rich laminated layers with scattered forams. Layers rich in formas range from 350 to 700 microns in thickness. The intercalated layers are approxi- mately 1500 microns thick and contain approximately 30- 40% forams. Probably the pure foramlnlferal layers reflect blooms in the water column in a poorly supplied terri- genous basin. These might be induced by periodic changes in local hydrodynamics affecting paleoecologic factors such as nutrient supply (Lipps, 1979). An additional indi- cator for those effects are phosphate laminae in the lower "V'tlleta" Formation of Quebrada Bambuc~i.
DISCUSSION
column. This is also supported by the presence of dwarfed microfauna, as mentioned above.
Blooms of planktonic foraminifera and metazoans, as well as the presence of terrigenous quartz, indicate both marine and terrigenous influx for the basin. The forams can provide additional clues to the origin of the Creta- coons black shales in the study area. They reveal quiet sedimentation over a long period of time. This fact can hardly be compared to the sharply bounded events of the Cretaceous (e.g., Jenkyns. 1980, 1991; StOhr, 1993). Thus. we suggest that black shale was deposited here during the Early Cretaceous because the basin had a restricted circu- lation and was poorly oxygenated. Tribovillard et al. (1991) presented similar factors giving rise to a regional "anoxic event" for the Upper Cretaceous La Luna Forma- tion (Venezuela). Further work on this question is in progress.
Future research utilizing better preserved foraminifers can lead to evolutionary conclusions, in particular to the early appearance of the genus Heterohelix. According to Caron (1985) and Nederbragt (1991), this taxon is thought to appear in the late Albian with H. moremani (Cushman). Our specimens could represent unknown ancestors.
SYSTEMATIC DESCRIPTIONS
Order FORAMINIFERA Eichwald 1830
Suborder GLOBIGERININA Delage & Herouard 1896
Superfamily ROTALIPORACEA Sigal 1958
Family ROTALIPORIDAE Sigal 1958
Subfamily TICINELLINAE Longoria 1974
The planktonic foraminifera presented here constitute a reference fox future biostratigraphic and exploratory work. Although biostratigraphic zones could not be established fox the sections studied, an age determination was possible where no macropaleontologic data were available. Our foram-supported stratigraphy is in agreement with the palynologic results of Pr6ssl (1992) in the Quebrada Ocal section. The proposed age fox the strata of the Caballos and "Villeta" Formations dealt with herein seems to be applicable for the Upper Magdalena Valley.
The foram-bearing segments (nos. 2 and 4) from the Caballos Formation obviously represent episodes of deeper water conditions than during deposition of the sandstone segments (nos. 1, 3, 5). For an interpretation of these facies in terms of sea-level changes, see Vergara (1992).
During the middle Albian, a deepening of the basin. deduced from lithologic changes, made it possible for planktonic forams to proliferate. However, these deposits do not represent deep-water environments (cf., e.g., Hart. 1980; Hart and Bailey, 1979). The relatively low variety and high number of species in our samples may reflect stressed paleoceanographic conditions at the water
Genus Ticinella Reichel 1950
* 1966
1985
Ticinella madecassiana Sigal 1966
Fig. 6.6a-c
Ticinella madecassiana n.sp. - - Sigal: p. 197; pl. 3, figs. 7a-b (Holotype).
Ticinella madecassiana Sigal - - Caron: p. 76; figs. 36/4-5.
Description
Spiral side: frequently 6, sometimes 5 or 7 chambers in the last whorl; chambers globular, increasing gradually in size as they are added; sutures radial and depressed; out- line lobulate.
Umbilical side: chambers ovoid, the first chambers sometimes subtrapezoidal; sutures radial and depressed; umbilicus very narrow; primary aperture extra-umbilical, nearly peripheral; supplementary apertures not visible.
Lateral view: low trochospiral.
198 J. BLAU, L. VERGARA, and H.W. STOCK

First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley, Colombia 199
* 1963
1974
1985
Ecinella primula Luterbacher 1963 Figs. 5.3a-c, 5.11a-c (cf.). 6.8a-c
Ticinella primula n.sp. - - Renz, Luterbacher & Schneider: p. 1085; text-fig. 4 (Holotype).
Ticinella primula Luterbacher - - Longoria: pp. 96-98; pl. 25. figs. 1-6; pl. 26, figs. 12- 14.
Ticinella primula Luterbacher - - Caron: p. 79; figs. 36/6-7.
Description
Spiral side: 7-8 chambers in the last whorl; chambers ovoid to subtrapezoidal in shape, increasing gradually in size as they are added; sutures radial and depressed; out- line slightly lobulate.
Umbilical side: chambers ovoid to subtrapezoidal in shape; sutures radial and depressed; umbilicus moderately wide; primary aperture extra-umbilical, nearly peripheral bordered by long portici which extend into the umbilicus; supplementary apertures umbilical.
Lateral view: very low trochospiral, pseudo-plani- spiral.
Remarks
The specimen shown in Fig. 5.11a-c is described here as Ticinella cf. primula. It shows the typical characteristics of T. primula but exhibits only 6 chambers in the last whorl.
* 1966
TicineUa raynaudi Sigal 1966 Figs. 4.2a-c, 4.4a-c, 5.12a-c
Ticinella raynaudi n.sp. - - Sigal: p. 200; pl. 6, figs. l(a-b) - 3(a-b).
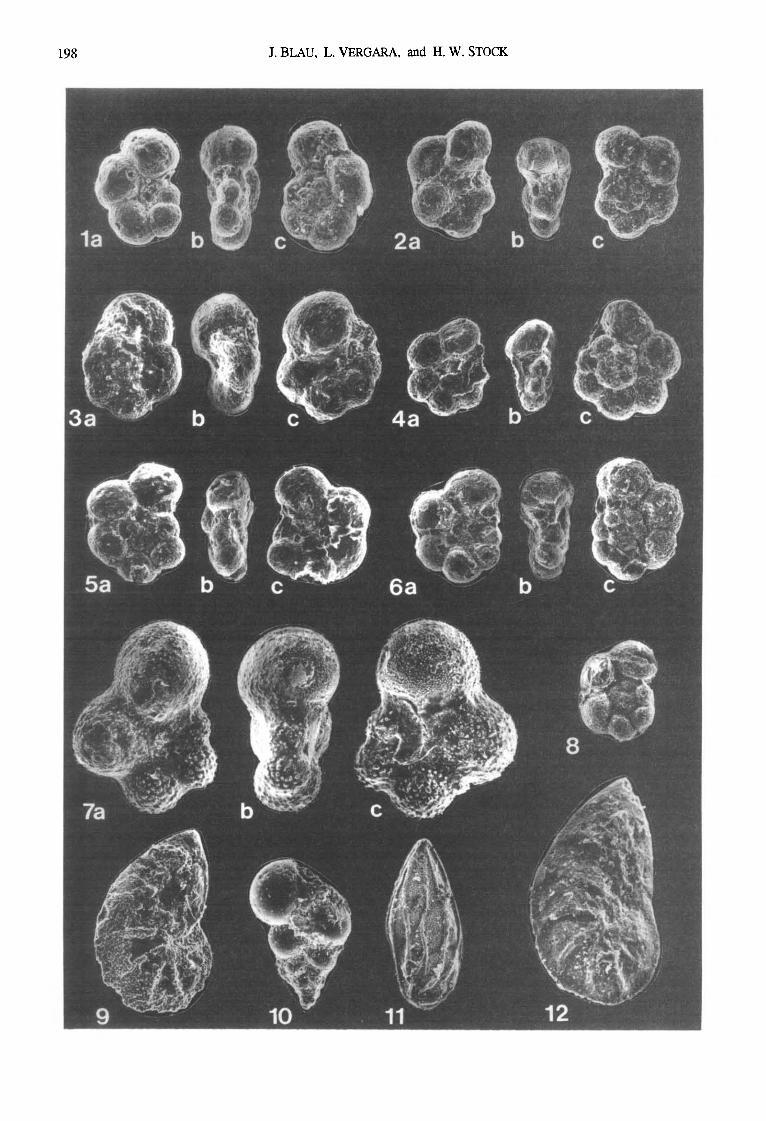
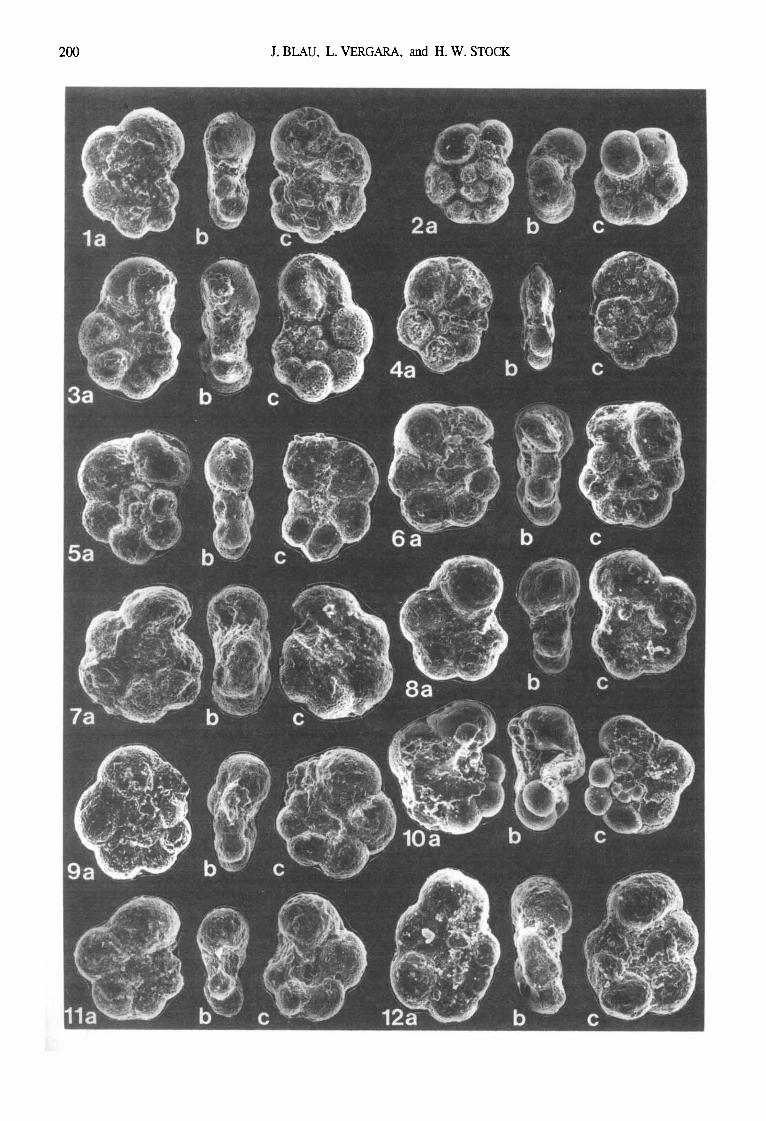
Fig, 4. Foraminifera from the Quebrada Ocal section (sample numbers in brackets; 8 mm = 50 microns):
la-c: Hedbergella sp. [11-10-01-90]
2a-c: Ticinella raynaudi Sigal [11-10-01-90]
3a-c: Hebergella delrioensis (Carsey) [11-10-01-90]
4a-c: Ticinella raynaudi Sigal [11-10-01-90]
5a-c: Hedbergella planispira (Tappan) [11-10-01-90]
6a-c: Hedbergella planispira (Tappan) [11-10-02-90]
7a-c: Globigerinelloides of, escheri (Kaufmann) [ 11-10-07- 90]
8: Hedbergella or Ticinella sp. [11-10-02-90]
9: Lenticulina sp. [11-10-01-90]
10: Sp. ind. exHeterohelicidae [11-10-05-90]
11: Gen. et sp, ind. ex Lagenina [ 11-10-02-90]
12: Lenticulina sp. [11-10-01-90]
1985 Ticinella raynaudi Sigal - - Carom p. 79; figs. 36/10-12.
Descr~aon
Spiral side: 6-8 chambers in the last whorl; chambers globular, the later chambers are either radially elongated or digitiform; chambers increasing gradually in size as they are added; sutures radial and depressed; outline lobu- late.
Umbilical side: chambers globular, the last ones becoming radially elongated or digitiform; the last cham- ber sometimes extends into the umbilicus; sutures radial and depressed; umbilicus narrow; primary aperture extra- umbilical, nearly peripheral; supplementary apertures umbilical (Fig. 4.2a).
Lateral view: low trochospiral.
Ticinella of. roberti (Gandolfi 1942) Figs. 6.5a-c
Remarks
The specimen illustrated shows close relationships to Ticinella roberti, but the trochospire is too low and possesses fewer chambers than Gandolfi's (1942) holo- type.
FamUyHEDBERGELLIDAELoeb~ch&Tappan1961 Subfam~yHEDBERGELLINAEIx~bfich&
Tappan1961
Genus Hedbergella Br6nnimann & Brown 1958
1926
1936
1974
1979
1980
1980
1980
Hedbergella delrioensis (Carsey 1926) Figs. 4.3a-c and 5.2a-c
Globigerina cretacea D'Orbigny var. del- rioenis n.var. - - Carsey: p. 43 (without figs.).
Globigerina cretacea D'Orb. - - Brotzen: 169; pl. 13, figs. la-c.
Hedbergella delrioensis (Carsey) - - Longo- ria: p. 54; pl. 10, figs. 1-3 (Neotype); pl. 10, figs. 1-12.
Hedbergella delrioensis (Carsey 1926) - - Robaszynski & Carom p. 123; pl. 22, figs. 1, 2; pl. 23, figs. 1-2.
Hedbergella delrioensis (Carsey 1926) - - Weiss: p. 110; pl. 1, figs. 1-3.
Hedbergella delrioensis (Carsey 1926) - - Peryt: p. 54; pl. 10, figs. la-c.
Hedbergella portsdownensis (Williams & Mitchell 1948) - - Peryt: p. 55; pl. 9. figs. la- c.
200 J. BLAU, L. VERGARA, and H. W. STOCK

First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley. Colombia 201
1984
1985
1993
Hedbergella delrioensis (Carsey 1926) - - Weidich: p. 81; pl. 3, figs. 1-4.
Hedbergella delrioensis (Carsey) - - Carom p. 57; figs. 25/6-7.
Hedbergella delrioensis (Carsey 1926) - - Stock: pp. 54-56; pl. 4, fig. la-e.
Description
Spiral side: 5-6 chambers in the last whorl; chambers globular, increasing gradually in size as they are added; suture.s radial and depressed; outline slightly lobulate.
Umbilical side: chambers globular; sutures radial and depressed; umbi l icus very narrow; aperture extra- umbilical, nearly peripheral.
Lateral view: low to slightly high trochospiral; the last chambers are displaced towards the umbilical side.
Remarks
The great intraspecific variability of Hedbergella del- rioensis is already well known. According to Weidich (1984) and Stock (1993), all variations from low trocho- spiral forms to higher trochospiral "portsdownensis" forms can be found in mid-Cretaceous sediments.
Hedbergella gorbachikae Longoria 1974
* 1974 Hedbergella gorbachikae Longoria, n.sp. - - Longoria: p. 56; pl. 15, figs. 11-13 (Holo- type), 1-10, 14-16 (Paratypes).
1985 Hedbergella gorbachikae Longor ia Carou: p. 59; figs. 25/8-9.
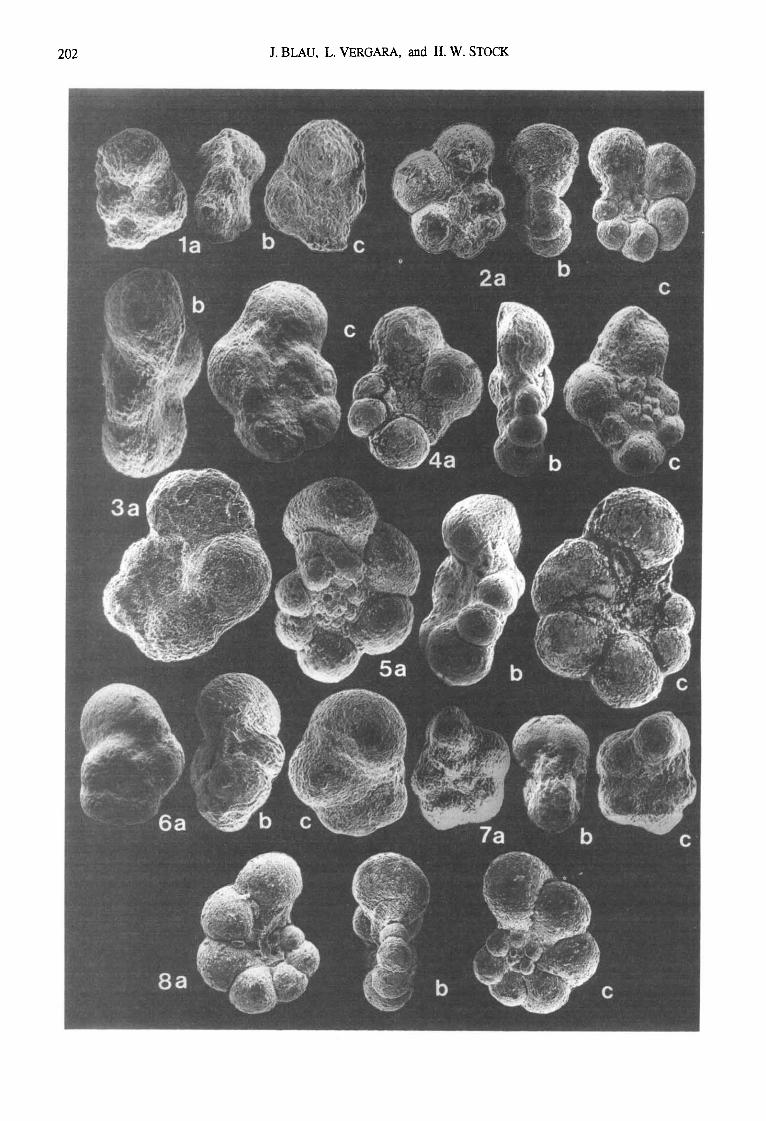
Fig. 5. Planktonic foraminifera from the Quebrada Ocal s~ction (sample numbers in brackets; 8 mm = 50 microns):
1a-c:
2a-c:
3a-c:
4a-e:
5a-c:
6a-c:
7a-c:
8a-c:
9a-c:
lOa-c:
lla-c:
12a-c:
Hedbergella planispira (Tappan) [11-10-01-90]
Hedbergella delrioensis (Carsey) [ 11-10-07-90]
Ticinella primula Luterbacher [ 11-10-02-90]
Hedbergella or Ticinella sp. [11-10-01-90]
Hedbergella or Ticinella sp. [11-10-01-90]
Hedbergella or Ticinella sp. [11-10-01-90]
Hedbergella or Ticineila sp. [11-10-01-90]
Hedbergella sp. [11-10-01-90]
Hedbergella or Ticinella sp. [11-10-01-90]
Hedbergella or Ticinella sp. [11-10-03-90]
Ticinella of. primula Luterbaeher [ 11-10-01-90]
Ticinella raynaudi Sigal [11-10-01-90]
Description
Spiral side: 5 chambers in the last whorl; chambers ovoid to globular, increasing gradually in size as they are added; sutures radial and depressed; outline more or less lobulate.
Umbilical side: chambers ovoid, sometimes subtri- angular; sutures radial and depressed; umbilicus very nar- row, completely or partly covered by the last chamber, which protrudes into the umbilical area; primary aperture extra-umbilical, nearly peripheral.
Lateral view: low trochospiral; piano-convex, spiral side flattened, umbilical side convex.
Remark
The specimen was ruined during SEM preparation and is thus not illustrated here.
* 1940
pars 1972
1979
1980
1984
1985
Hedbergella planispira (Tappan 1940) Figs. 4.5a-c, 4.6a-c, 5.1a-c, 6.2a-c
Globigerina planispira n.sp. - - Tappan: p. 122; pl. 19, figs. 12a-c (Holotype).
Hedbergella planispira (Tappan 1940) - - Hanzlikova: p. 101; pl. 26, figs. 1-2; non pl. 25. fig. 15.
Hedbergella planispira (Tappan 1940) - - Robaszynski & Caron: p. 139; pl. 27, figs. 1- 3; pl. 28, figs. 1-4.
Hedbergella planispira (Tappan, 1940) Peryt: p. 54; pl. 10, figs. 5,6.
Hedbergella planispira (Tappan 1940) - - Weidich: p. 81; pl. 3. figs. 5-8.
Hedbergella planispira (Tappan) - - Carom p. 59; figs. 25/23-24.
Description
Spiral side: 6-8 chambers in the last whorl; chambers globular with a smooth surface, increasing gradually in size as they are added; sutures radial and depressed; out- line slightly lobulate.
Umbilical side: chambers globular with a smooth sur- face; sutures radial and depressed; umbilicus wide; aper- ture extra-umbilical, nearly peripheral.
Lateral view: very low trochospiral; pseudo-plani- spiral.
* 1966
Hedbergella sigali Moullade 1966 Fig. 6.1
Hedbergella (Hedbergella) sigali n.sp. - - Moullade: p. 87; pl. 7, figs. 20-23, 24-25 (Holotype).
202 J. BLAU, L. VERGARA, and H.W. STOCK
First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley, Colombia 203
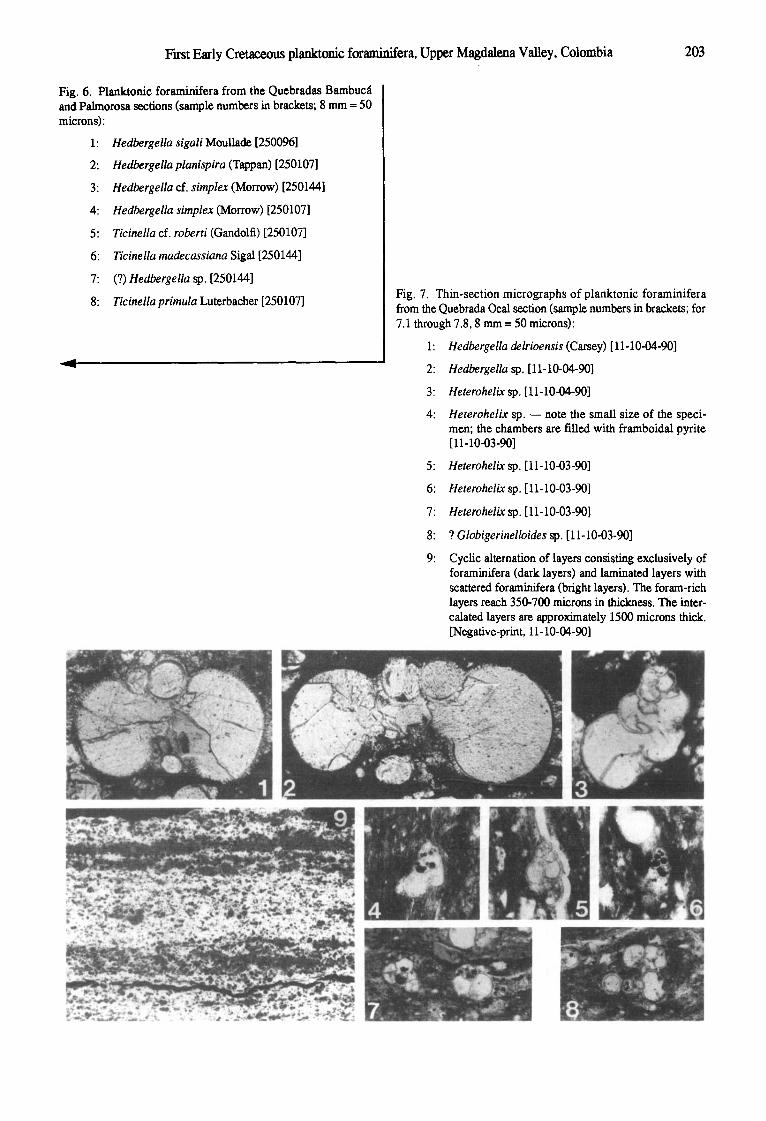
Fig. 6. Planktonic foraminifera from the Quebradas Bambuc~l and Palmorosa sections (sample numbers in brackets; 8 mm = 50 microns):
1:
2:
3:
4:
5:
6:
7:
8:
Hedbergella sigali Moullad~ [250096]
Hedbergella planispira (Tappan) [250107]
Hedbergella of. simplex (Morrow) [250144]
Hedbergella simplex (Morrow) [250107]
Ticinella of. roberti (Gandolfi) [250107]
Ticinella madecassiana Sigal [250144]
(?) Hedbergella sp. [250144]
Ticinella primula Luterbaeher [250107] Fig. 7. Thin-section micrographs of planktonic foraminifera from the Quebrada Ocal section (sample numbers in brackets; for 7.1 through 7.8, 8 mm= 50 microns):
1: Hedbergella delrioensis (Carsey) [11-10-04-90]
2: Hedbergella sp, [11-10-04-90]
3: Heterohelix sp. [11-10-04-90]
4: Heterohelix sp. ~ note the small size of the speci- men; the chambers are filled with framboidal pyrite [11-10-03-90]
5: Heterohelix sp. [11-10-03-90]
6: Heterohelix sp. [11-10-03-90]
7: Heterohelix sp. [11-10-03-90]
8: ? Globigerinelloides sp. [11-10-03-90]
9: CycLic alternation of layers consisting exclusively of foraminifera (dark layers) and laminated layers with scattered foraminifera (bright layers). The foram-rich layers reach 350-700 microns in thickness. The inter- calated layers are approximately 1500 microns thick. [Negative-print, 11-10-04-90]
4
8

204 J. BLAU, L. VEROARA, and H.W. STOCK
1974
1985
Hedbergella sigali Moullade 1966 - - Lon- goria: p. 68; pl. 21, figs. 6-8; pl. 22, figs. 1- 13.
Hedbergella sigali Moullade - - Caron: p.59; figs. 25/21-22.
Description Spiral side: 4-5 chambers in the last whorl; chambers
globular, increasing gradually in size as they are added; sutures radial and depressed; outline lobulate.
Umbilical side: chambers globular; sutures radial and depressed; umbilicus narrow; aperture extra-umbilical. nearly peripheral.
Lateral view: moderately high trochospiral.
the last chamber is mostly significantly elongated; cham- bers increasing abruptly in size as they are added; sutures radial and depressed; outline strongly lobulate.
Umbilical side: chamber shape very variable (see above); sutures radial and depressed; umbilicus very nar- row; aperture extra-umbilical, nearly peripheral.
Lateral view: moderately to distinctly high trocho- spiral.
Remarks The high degree of variability of Hedbergella simplex
has led to the creation of a number of new species and one genus (Clavihedbergella), most of which were placed in the synonymy of H. simplex in the micropaleontological literature in recent years.
pars
1984
1985
1985
1993
Hedbergella simplex (Morrow 1934) Fig. 6.4a-c
1934 Hastigerinella simplex n.sp. - - Morrow: pp. 198-199; text-fig. 6a-b (Holotype).
1961 Clavihedbergella simplex (Morrow). - - Loe- blich & Tappan: p. 279; pl. 3, figs. 11-14.
1961 Hedbergella amabilis n.sp. - - Loeblich & Tappan: p. 274; pl. 3, fig. 1 (Holotype), 2, 4 (Paratypes), 6-10;, non pl. 3, figs. 3, 5.
1970 Hedbergella flandrini nov.sp. - - Porthault: p. 64; pl. 10, fig. 1 (Holotype) , 2, 3 (Paratypes).
1972 ClavihedbergeUa simplex (Morrow) - - Barr: p. 14; pl. 2, figs. 2a-b.
1972 Hedbergella amabilis I_x~blich & Tappan - - Barr: p. 13; pl. 2, figs. 3a-c.
1979 Hedbegella simplex (Morrow 1934) - - Robaszynski & Caron: p. 145; pl. 29, figs. 1- 3; pl. 30, figs. 1, 2.
1979 Hedbergellaflandrini Porthault 1970 - - Robaszynski & Caron: p. 129; pl. 24, figs. 1, 2; pl. 25, figs. 1-3.
1980 Hedbergella simplicissima (Magne & Sigal 1953) - - Peryt: p. 56; pl. 10, figs. 7, 8.
Hedbergella simplex (Morrow 1934) - - Weidich: p. 82; pl. 3, figs. 9-13.
Hedbergella simplex (Morrow) - - Caron: p. 59; figs. 25/15-16.
Hedbergellaflandrini Porthault - - Caron: p. 57; figs. 25/12-14.
Hedbergella simplex (Morrow 1934) - - Stock: p. 56; pl. 4, figs. 4a-c.
Description Spiral side: 4-6 chambers in the last whorl; the cham-
ber shape is very variable: globular to radially elongated;
Superfamily PLANOMALINACEA Bolli, Loeblich & Tappan 1957
Family GLOBIGERINF+I,I OIDIDAE Longoria 1974 Subfamily GLOBIGERINELLOIDINAE Longoria 1974
Genus GlobigerbzeUoides Cushman & Ten Dam 1948
Globigerinelloides cf. escheri (Kaufmann) Fig. 4.7a-c
Description Planispiral; 5 chambers in the last whorl; chambers
globular, increasing rapidly in size as they are added; sur- face of the first chambers covered by pustules; sutures radial and depressed; outline strongly lobulate; aperture peripheral.
Remarks
A single specimen recovered from presumed late Albian strata in the Quebrada Ocal. It closely resembles Globigerinelloides escheri (Kaufmann), previously reported from the Western Cordillera by Etayo et al. (1980; see Introduction), and in northwestern Colombia by Gandolfi (1955) as Globotruncana beldingi beldingi. (For a taxonomic discussion, see Masters, 1977.)
Acknowledgements--W. Blind and F. Stibane (Giegen) critically read the manuscript. Field work with G. Renzoni (Ingeominas) and revision of some Foraminifera identifications by H. P. Luterbacher (TUbingen) are acknowledged. J. Gr6sser and M. Senff provided field data from the Que- brada Ocal section. Valuable suggestions were provided by the reviewers, Dr. E. A. Balcells-Baldwin (Houston, TX) and Dr. D. W. Engelhardt (Columbia, SC), with additional corrections by D. Tanner (Giegen). M. Schorge and G. Appel (Giel~en) are thanked for photographic work. The SF.M photographs of Fig. 6 were taken at the Gieflen University Strahlen- zentrum by Dr. D611. This paper is a contribution to DFG Project "Fazielle, palaogeographische und palaoklimatologische Untersuch- ungen an Mikrofannen aus der Kreide Kolumbiens" (B1 66/11-2), as part of the DFG working group "Globale und regionale Steuerungsprozesse biogener Sedimentation." Biostratigraphy from the Aipe section is part of L. V.'s dissertation at the University of GieBen (DAAD grant 322/ 509002/1).

First Early Cretaceous planktonic foraminifera, Upper Magdalena Valley, Colombia 205
REFERENCES
American Commission on Stratigraphic Nomenclature, 1983. Code of stratigraphic nomenclature. Bulletin of the American Association of Petroleum Geologists 67 (5), 841-875.
Burr, F. T., 1972. Cretaceous biostratigraphy and planktonic foraminifera of Libya. Micropaleontology 18 (1), 1-46.
Beltr/m, N., and Gallo, J., 1968, The geology of the Neiva sub-basin, Upper Magdalena Basin (southern portion). In: Geological Field Trips, Colombia: 1958-1978, pp. 253-276. Colombian Society of Petroleum Geologists and Geophysicists, BogotA, Colombia, 545 p.
Berger, W. H., 1971. Planktonic foraminifera: Sediment production in an oceanic front. Journal o fF oraminiferal Research 1 (3), 95-118.
Brotzen, F., 1936. Foraminiferen aus dem Schwedischen untersten Senon von Eriksdal in Schonen. Sveriges Geologiska Undersokning, C396, Arsbok 30, 234 p.
Btlrsl, H., 1961. Geologfa de los alrededores de Ortega, Tolima. Univer- sidad Industrial de Santander, Bucaramanga, Bolet[n Geol6gico 8, 21-38.
Bllrgl, H., and Dummit, 3"., 1954. E1 Cret,'iceo Superior de la regi6n de Girardot. lnstituto Geol6gico Nacional, BogoM, Bolettn Geol6gico 2 (1), 23-48.
C~ceres, C., and Etayo-Serna, E, 1969. Bosquejo Geol6gico de la Regi6n del Tequendama: Op~sculo Gufa Excursi6n, I Congreso Colombiano de Geolo~a, Universidad Nacional de Colombia, BogotA, Colombia, 24p.
Caron, M., 1985. Cretaceous planktic foraminifera. In: Plankton Stratig- raphy (edited by H. M. bolli, J. B. Saunders, and K. Pesch-Nielsen), pp. 17-86. Cambridge University Press, Cambridge, UK, 1032 p.
Caron, M., and Homewood, P., 1983. Evolution of early planktic fora- minifers. Marine Micropaleontology 7 (1981/83), 453-462.]
Carsey, D. O., 1926. Foraminifera of the Cretaceous of central Texas. University of Texas Bulletin 2612, 1-56.
Colom, G., 1962. Micropaleontologfa del CretAceo al NW de Bogot,'l (Colombia). Universidad Industrial de Santander. Bucaramanga, Boleffn de Geologfa 10, 69-75.
Corrigan, H., 1967. The geology of the Upper Magdalena Basin (north- ern portion). In: Geological Field Trips, Colombia." 1958-1978, pp. 221-252. Colombian Society of Petroleum Geologists and Geophysi- cists, BogotA, Colombia, 545 p.
Cushman, J., and Hedberg, H., 1941. Upper Cretaceous Foraminifera from Santander del Norte, Colombia, S.A. Contributions from the Cushman Laboratory for Foraminiferal Reasearch 17 (23/4), 79-108.
Etayo, F., 1979. Zonation of the Cretaceous of central Colombia by ammonites, lngeominas. Publicaciones Geol6gicas Especiales 14, 1- 235.
Etayo, E, Gonz,'tlez, H., and Alvarez, E., 1980. Mid-Albian ammonites from northern Western Cordillera, Colombia, S.A. Geologia Noran- dina 2, 25-30.
F0chtbauer, H. (ed.), 1989. Sediment-Petrologie I1: Sedimente und Sedi- rnentgesteine (4th Edition). Schweizerbart, Stuttgart, Germany, 1141 p.
Fdquen, J., Rodrfguez, G., and Cassio, U., 1989. Mapa Geol6gico Pre- liminar, Plancha 302 (Aipe), Esc: 1:100 000. Ingeominas, BogotA, Colombia.
Gandolfi, R., 1942. Richerche micropaleontologiche e stratigraphaphiche sulla Scaglia e sul Flysch eretaeici dei dintorni di Balerna (Canton Tieino). Rivista ltaliana Paleontologia 4, 1-160.
Gandolfi, R., 1955. The genus Globotruncana in northeastern Colombia. Bulletin of American Paleontology 36 (155), 7-112.
Grosse, E., 1930. Acerea de la litologia del sur tie Colombia, 1. Com- pilaci6n de Estudios Geol6gicos Oficiales en Colombia 3, 31-137.
Hanzlikova, E., 1972. Carpathian Upper Cretaceous Foraminiferidae of Moravia (Turonian-Maastrichtian). Rozpravy Ustredniho Ustavu GeologiclMho 39, 1-160.
Hart, M. B., 1980. A water depth model for the evolution of the plank- tonic Foraminiferidae. Nature 286 (5770), 252-254.
Hart, M. B., and Bailey, H. W., 1979. The distribution of planktonic Fora- miniferidae in the mid-Cretaceous of NW Europe: Aspekte der Kreide Europas. lUGS Series A6, 527-542.
Heltner, A., 1892. Die Kordillere van BogoM [Erganzungsheft 104 zu Petermanns Mitteilungen ]. Justus Pertha Verlag, Gotha, Germany, 131 p.
Jenkyns, H. C., 1980. Cretaceous anoxic events: From continents to oceans. Journal of the Geological Society of London 137, 171-188.
Jenkyns, H, C., 1991. Impact of Cretaceous sea level rise and anoxic events on the Mesozoic carbonate platform of Yugoslavia. Bulletin of the American Association of Petroleum Geologists 75 (6), 1007- 1017.
Julivert, M., 1968. Lexique Stratigraphique International. Vol. 5: AmErique Latine -- Fasicule 4a, Colombie. Centre National de la Recherche Scientifique, Paris, France, 605 p.
Lipps, J., 1973. Micrnfossila. In: Encyclopedia of Microscopy and Micro- technique (edited by P. Gray), pp. 308-312. Van No, wand, New York, NY, USA, x + 638 p.
Loeblich, A. R., and Tappan, H., 1961. Cretaceous planktonic foramini- fera: Part 1, Cenomanian. Micropaleontology 7 (3), 257-304.
Longoria, J. E, 1974. Stratigraphic, morphologie and taxonomic studies of Aptian planktonic foraminifera. Revista Espa?lola de Micropaleon- tologfa: N~mero Extraordinario, 1-107.
Martlnez, J. I., 1989. Foraminiferal biostratigraphy and paleoenviron- ments of the Maastrichtian Colon mudstones of northern South America. Micropaleontology, 35 (2), 97-113.
Martfnez, J. I., 1990. Estratigrafia de la Plancha 227 (Villeta). Unpub- lished report, Ingeominas, Bogotfi, Colombia, 120 p.
Masters, B. A., 1977. Mesozoic planktonic foraminifera. In: Oceanic Micropaleontology, Vol. I (edited by A. T. S. Ramsay), pp. 301-731. Academic Press, New York, NY, USA, 837 p.
Mojica, J., and Macfa, C., 198 I. Caracteristicas estratigr,'ifieas y edad de la Formaci6n Yavf, Mesozoico, de la regi6n entre Prado y Dolores. Tolima, Colombia. Geologia Colombiana 12, 7-32.
Morrow, A. L., 1934. Foraminifera and Ostracoda from Upper Creta- ceous of Kansas. Journal of Paleontology 8 (8), 186-205.
Moullade, M.. 1966. Etude stratigraphique et micropaMontologique du Cr6tac6 inferieur de la 'Fosse vocontien. ' Document de la Lubo- ratoire Gdologique, Facult6 des Sciences Lyon 15, 1-369.
Nederbragt, A. I.. 1991. Late Cretaceous biostratigraphy and develop- ment of Heterohelicidae (planktic foraminifera). Micropaleontology 37 (4), 329-372.
Nthlcz, A., Mosquera, D., and Vesga, C. J., 1984. Mapa Geoldgico Pre- liminar: Plancha 263. Ortega; Scale 1.100000. Ingeominas, Ibagu6, Colombia.
Olsson, A. A., 1956. Colombia, In: Handbook of South American Geo- logy: An Explanation of the Geologic Map of South America (edited by W. E Jenks). Geological S~x:iety of America, Bulletin 65, xviii + 377 p.
Peryt. D., 1980. Planktic Foraminifera Zonation of the Upper Creta- ceous in the Middle Vistula Rivet" Valley, Poland. Palaeontologia Polonica 41,101 p.
Pessagno, E. A., Jr., 1967. tipper Cretaceous planktonic foraminifera from the western Gulf Coastal Plain. Palaeontographica Americana 5, 245-445.
Petters, V., 1954. Typical foraminili~ral horizons in the Lower Cretaoeous of Colombia, S.A. Contributions from the Cushman Foundation for Foraminiferal Research 5, 128-137.
Porthault, B., 1970. Etude de la microfaune. In: Le Senonien lnferieur de Pt~get-Theniers (Alpes Maritimes) et Sa Microfaune (edited by P. Donze, B. Porthault, G. Thomel, and O. de Villoutreys). G~obios 3 (2), 41-106.
Prt~ssl. K. E, 1992. Preliminary results of palynological investigations on the Cretaceous of Colombia, South America. Review of Paleobotany and Palynology 71,255-268.
Pr~ssl, K. E, and Vergara, L., 1993. The Yavf Formation (Lower Creta- ceous), Upper Magdalena Valley: An integrated sedimentological and

206 J. BLAU, L. VERGARA, and H. W. STOCK
palynological study. Neues Jahrbuch far Geologie und Palaontologie 188 (in press).
Raasveldt, H. C., and Carvajal, J. M., 1957a. Mapa Oeol6gico de la Rep~blica de Colombia: Sheet M-8, Ataco, Scale 1:200 000. Servicio Geol6gico Naeional, BogotA, Colombia.
Raasveldt, H. C., and Carvajal, J. M., 1957b. Mapa Geol6gico de la Repablica de Colombia: Sheet L-9, Girardot, Scale 1:200 000. Servi- cio Geol6gico National, BogotA, Colombia.
Renz, O., Luterbacher, H., and Schneider, A., 1963. Stratigraphisch- palltontologisehe Untersuchtmgen im Albien und Cenomanien des Neuburger Jura. Eclogae Geologicae Helvetiae 56, 1073-1116.
Robaszynski, F., and Caron, M., 1979. Atlas de Foraminif~res Planc- toniques du Crdtac~ Moyen (Mer Bor#ale et T~thys). Cahiers de Micropal6.ontologique I (1979), 185 pp.
Robaszynski, E, Caron, M., and the European Working Group on Plank- tonic Foraminifera, 1984. Arias of Late Cretaceous globotruncanids. Revue de Micropal$ontologie 26 (3-4), 145-305.
Sigal, J., 1966. Contribution a une monographie des Rosalines: 1, Le genre Ticinella Reiehel, souches des Rotalipores. Eclogae Geolo- gicae Helvetiae 59, 187-217.
Sliter, R., 1989. Biostratigraphic zonation for Cretaceous planktonic Foraminifera examined in thin section. Journal of Foraminiferal Research 19 (1), 1-19.
Stock, H. W., 1993. Stratigraphie, Sedimentologie und Palaogeographie der Oberkreide in den NE Dolomiten (Italien). Dissertation, Justus- Liebig-Universitat Gieflen, Germany. 115 p.
StOhr, D., 1993. Die Ammoniten der Kreide yon La Stua (Norditalien ). Dissertation, Justus-Liebig-Universitat Gieflen, Germany (in prep.).
Stutzer, O., 1926. Contdbuci6n a la Geologfa de ha Cordillera Oriental, re#ones cerca a Bogot~i. Compilaci6n de Estudios Geol6gicos Off- ciales en Colombia, 141- t83.
Tappan, H., 1940. Foraminifera from the Graysen Formation of northern Texas. Journal of Paleontology 14 (2), 93-126.
Tribovilhard, N.-P., Stephan, J. E, Manivit, H., Reyre, Y., Cotillon, P., and Jaut~e, E., 1991. Cretaceous black shales of Venezuelan Andes: Pre- liminary results on stratigraphy and paleoenvironmental interpreta- tions. Palaeogeography, Palaeoclimatology, Palaeoecology 81 (3/4), 313-321.
Vergara Streinesberger, L., 1992. Lower Cretaceous stratigraphic sequences in the Quebrada Bambuc~i (Aipe), Upper Magdalena Valley, Colombia. Gieflener Geologische Schriflen 48, 183-200.
Weidich, K. F., 1984. Feinstratigraphie. Taxonomie Planktonischer Foraminiferen und PalOkologie der Foraminiferengesamtfauna der Kalkalpinen Tieferen Oberkreide. Bayerische Akadademie der Wis- senschaften, Mathematiscb-Naturwissenschaftliche Klasse, Neue Folge 162, 151 p.
Weiss, W., 1980. Feinstratigraphie des Cenoman und des Turon mit Planktonischen Foraminiferen: Ein Korrelationsversuch Boreal- Nordtethys. Dissertation, Universitlit Ttlbingen, Germany, 174 p.
Recommended

















