
I
Aus dem Institut für Medizinische Biochemie und Molekularbiologie
(Direktorin Prof. Dr. rer. nat. habil. Elke Krüger)
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
Einfluss der pharmakologischen Modulation des Angiotensin-(1-7) Rezeptors MAS auf die
Expression von TRPC-Kanälen in Osteosarkom-Zelllinien
Inaugural - Dissertation
zur
Erlangung des akademischen
Grades
Doktor der Medizin
(Dr. med.)
der
Universitätsmedizin
der
Ernst-Moritz-Arndt-Universität
Greifswald
2018
vorgelegt von: Florian Lässig
geb. am: 28.02.1990
in: Magdeburg

I
Dekan: Prof. Dr. rer. nat. Max P. Baur
1. Gutachter: Prof. Dr. rer. nat. Heinrich Brinkmeier
2. Gutachter: Prof. Dr. med. Bernhard H. Rauch
Ort, Raum: Greifswald, Seminarraum J02.15
Tag der Disputation: 19.07.2018

I
INHALTSVERZEICHNIS
1 ABKÜRZUNGSVERZEICHNIS .................................................................................................................1
2 EINLEITUNG ................................................................................................................................................3
2.1 OSTEOSARKOM ...........................................................................................................................................3
2.2 ZELLLINIEN ................................................................................................................................................7
2.3 KLASSISCHES RAS UND ALTERNATIVES RAS ........................................................................................7
2.4 TRPC-KANÄLE ....................................................................................................................................... 13
2.5 ZIELSTELLUNG ........................................................................................................................................ 16
3 MATERIAL UND METHODEN ............................................................................................................. 17
3.1 MATERIAL ............................................................................................................................................... 17
3.2 METHODEN ............................................................................................................................................. 23
4 ERGEBNISSE ............................................................................................................................................. 30
4.1 EINFLUSS VON IL1 UND AVE-0991AUF DIE MAS-MRNA-GEHALTE ........................................... 30
4.2 MRNA-EXPRESSION DER TRPC-KANÄLE ........................................................................................... 32
4.3 EINFLUSS VON IL1 UND AVE-0991 AUF DIE TRPC-KANAL MRNA-GEHALTE .......................... 34
4.4 DER EINFLUSS VON D-ALA AUF MAS- UND TRPC- MRNA-GEHALTE ............................................ 39
4.5 DER EINFLUSS VON LY294002 UND PD98059 AUF DIE MAS- UND TRPC- MRNA-GEHALTE . 44
5 DISKUSSION .............................................................................................................................................. 51
6 ZUSAMMENFASSUNG ............................................................................................................................ 58
7 LITERATURVERZEICHNIS ................................................................................................................... 59

1
1 ABKÜRZUNGSVERZEICHNIS
ACE Angiotensin-Converting-Enzyme
ADH Antidiuretische Hormon
AngI, II, III, IV Angiotensin I bis IV
APA Aminopeptidase A
APN Aminopeptidase N
AT1R/AT2R/AT4R Angiotensinrezeptor 1, 2 und 4
AVE-0991 synthetisches Ang-(1-7) Analogon
bp Basenpaare
cDNA complementary DNA, dt. komplementäre DNA
D-Ala D-Alanin7-Ang-(1-7); synthetischer Mas-Rezeptor-Antagonist
DMSO Dimethylsulfoxid
EGFR Epidermal Growth Factor Receptor
ER Endoplasmatisches Retikulum
ERK1/2 Extracellular Signal Regulated Kinase 1/2
FCS Fetal Calf Serum, dt. fetales Kälberserum
GPCR G-Protein Coupled Receptor
IL1β Interleukin 1β
IP3 Inositol-1,4,5-trisphosphat
Ly294002 synthetischer Inhibitor der PI3-Kinase
MAP-Kinasen Mitogen-Activated Protein Kinasen
MrgD Mas-related G-protein coupled receptor member D
MMNG/HOS N-Methyl-N-Nitro-N-Nitrosoguanidin/human osteosarcoma
mRNA messenger RNA
NEP Neutrale Endopeptidase
NFκB Nuclear Factor-κB
NOD2 nucleotide-binding and oligomerization domain 2
PBS Phosphate buffered saline, dt. Phosphat-gepufferte Salzlösung
PI3-Kinase Phosphatidylinositol-4,5-Bisphosphate 3-Kinase
PCR Polymerase Chain Reaction
PD98059 synthetischer Inhibitor von ERK 1/2

ABKÜRZUNGSVERZEICHNIS
2
POP Prolyl-Oligopeptidase
RAS Renin-Angiotensin-System
RAAS Renin-Angiotensin-Aldosteron-System
RT-qPCR Reverse Transcription quantitative Polymerase Chain Reaction
SMOC Second messenger-operated Channel
SOCE Store-operated Calcium Entry
TOP Thimet-Oligopeptidase
TNFα Tumor Necrosis Factor α
TRPC Transient Receptor Potential Canonical
U-2 Os U-2 osteosarcoma
v/v Volumen pro Volumen
w/v Gewicht pro Volumen
ZNS Zentrales Nervensystem

3
2 EINLEITUNG
2.1 OSTEOSARKOM
Das Osteosarkom ist ein primärer und hochmaligner Knochentumor des Kindes- und Jugendalters.
Sowohl die frühzeitige Metastasierung als auch die damit einhergehende ungünstige Prognose
machen diese Tumorentität zu einem wichtigen Teil aktueller Bestrebungen effizientere Therapien
zu entwickeln um den Heranwachsenden ein längeres und qualitativ höheres Überleben zu sichern.
In den letzten Jahrzehnten gab es glücklicherweise schon eine deutliche Steigerung der
Überlebensraten durch den Einsatz von systemischer prä- und postoperativer Polychemotherapie.
Trotzdem überlebt jeder fünfte Patient die ersten fünf Jahre nach Diagnosestellung nicht. [1;2;3]
Wie es zur Entartung der Zellen kommt, ist nicht zufriedenstellend geklärt. Jedoch treten sie
häufiger im Zusammenhang mit familiären Neoplasiesyndromen auf, bei denen
Tumorsupressorgene wie das Retinoblastom-Protein oder der „Wächter des Genoms“ p53 mutiert
sind. [4] Diese beiden Proteine sind auch in den sporadisch auftretenden Osteosarkomen
regelmäßig betroffen. Zusätzlich tragen die häufigen Alterationen des Insulin-like Growth Factor 1
Receptor-Signalweges über die ungehemmte Aktivierung von Proteinkinasen wie der
Phosphatidylinositol-4,5-Bisphosphate 3-Kinase (PI3-Kinase) oder der Extrazellulär Signal-
Regulierte Kinase 1/2 (ERK1/2) zur Progression des Osteosarkoms bei. [5;6;7]
Bei 2 bis 5 von 1.000.000 Europäern wird jedes Jahr die Diagnose Osteosarkom gestellt. Meist sind
es junge Menschen im Alter von 15 bis 25 Jahren und mit 60 % sind Jungen etwas häufiger betroffen.
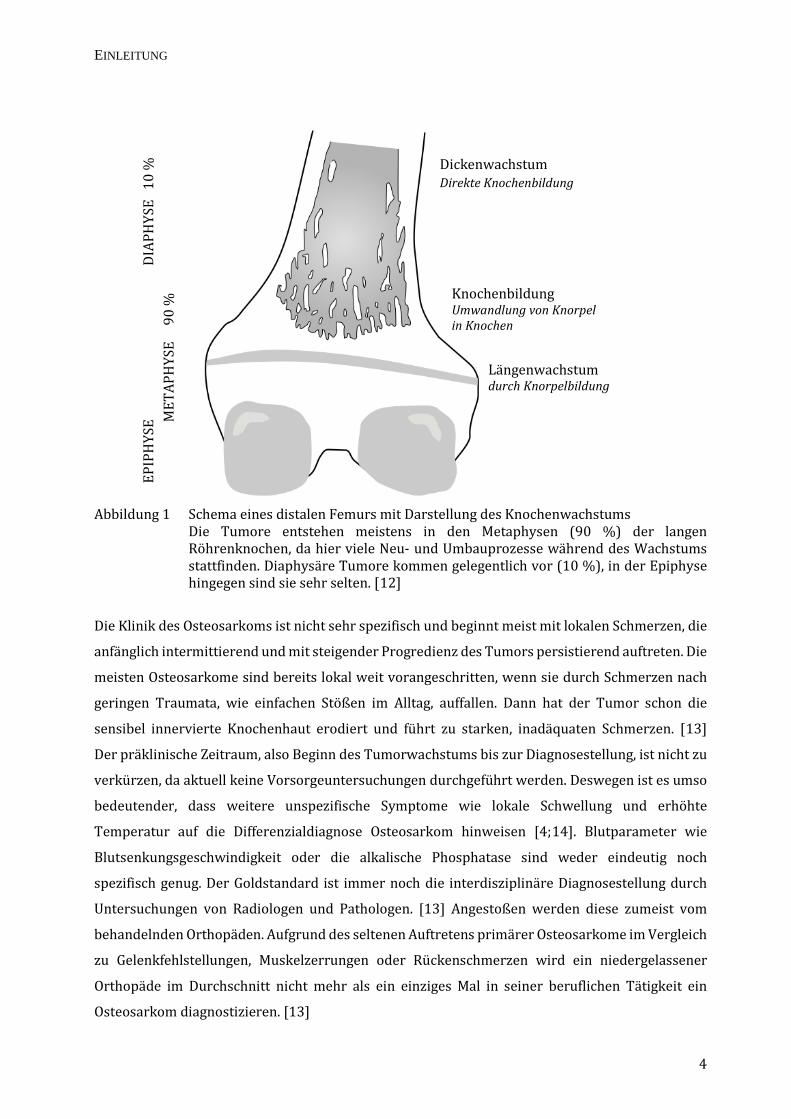
Grundsätzlich kann das Osteosarkom überall im Knochen auftreten, aber die Metaphysen – die
Bereiche des Längenwachstums der Röhrenknochen (siehe Abbildung 1) – sind am häufigsten
betroffen. Laut Statistik ist die häufigste Lokalisation das distale Femur, gefolgt von der proximalen
Tibia, dem Humerus, Ober- und Unterkiefer sowie den Beckenschaufeln. Mit circa 6 von 10 Fällen
sind die meisten Manifestationen kniegelenksnah. [8;9] Stammnahe Osteosarkome (typisch für
ältere Patienten in der sechsten Lebensdekade) sind eher sekundärer Natur wie beim
Morbus Paget – einer eventuell viral bedingten Knochengewebsvermehrung – oder durch
Strahlung induziert. [10;11]
EINLEITUNG
4
Abbildung 1 Schema eines distalen Femurs mit Darstellung des Knochenwachstums Die Tumore entstehen meistens in den Metaphysen (90 %) der langen Röhrenknochen, da hier viele Neu- und Umbauprozesse während des Wachstums stattfinden. Diaphysäre Tumore kommen gelegentlich vor (10 %), in der Epiphyse hingegen sind sie sehr selten. [12]
Die Klinik des Osteosarkoms ist nicht sehr spezifisch und beginnt meist mit lokalen Schmerzen, die
anfänglich intermittierend und mit steigender Progredienz des Tumors persistierend auftreten. Die
meisten Osteosarkome sind bereits lokal weit vorangeschritten, wenn sie durch Schmerzen nach
geringen Traumata, wie einfachen Stößen im Alltag, auffallen. Dann hat der Tumor schon die
sensibel innervierte Knochenhaut erodiert und führt zu starken, inadäquaten Schmerzen. [13]
Der präklinische Zeitraum, also Beginn des Tumorwachstums bis zur Diagnosestellung, ist nicht zu
verkürzen, da aktuell keine Vorsorgeuntersuchungen durchgeführt werden. Deswegen ist es umso
bedeutender, dass weitere unspezifische Symptome wie lokale Schwellung und erhöhte
Temperatur auf die Differenzialdiagnose Osteosarkom hinweisen [4;14]. Blutparameter wie
Blutsenkungsgeschwindigkeit oder die alkalische Phosphatase sind weder eindeutig noch
spezifisch genug. Der Goldstandard ist immer noch die interdisziplinäre Diagnosestellung durch
Untersuchungen von Radiologen und Pathologen. [13] Angestoßen werden diese zumeist vom
behandelnden Orthopäden. Aufgrund des seltenen Auftretens primärer Osteosarkome im Vergleich
zu Gelenkfehlstellungen, Muskelzerrungen oder Rückenschmerzen wird ein niedergelassener
Orthopäde im Durchschnitt nicht mehr als ein einziges Mal in seiner beruflichen Tätigkeit ein
Osteosarkom diagnostizieren. [13]
EPIP
HYS
E
D
IAPH
YSE
10
%
M
ETAP
HYS
E
90 %
Längenwachstum durch Knorpelbildung
Knochenbildung Umwandlung von Knorpel in Knochen
Dickenwachstum Direkte Knochenbildung

EINLEITUNG
5
Nach einer detaillierten Analyse von Poppe im Jahr 1978 dauerte es beginnend bei dem Symptom
Schmerz durchschnittlich mehr als 38 Wochen bis zur Einleitung der Therapie. Aufgrund der
Bemühungen von Spezialisten wurden viele Kollegen in den letzten Jahrzehnten sensibilisiert,
sodass diese Zeitspanne um knapp 80 bis 90 % verkürzt werden konnte. [13;15]
Nicht nur die Diagnostik wurde in den letzten Jahrzehnten verbessert, auch die Therapie wurde mit
Einführung der adjuvanten Chemotherapie wirksamer. Bei diesem Vorgehen werden nach der
operativen Resektion des Tumors systemische Zytostatika appliziert. Dabei war zunächst
Methotrexat in Kombination mit Bleomycin oder Cyclophosphamid die Therapie der Wahl. [16]
Nach diesem Erfolg wurde zusätzlich die neoadjuvante Therapie – also ein präoperativer Einsatz
der Medikamente – getestet und neben Methotrexat bilden Doxorubicin und Cisplatin weitere
Standard-Zytostatika, die nach aktuellem Protokoll eingesetzt werden. Bei sogenannten
„Respondern“ zeigt sich eine klinische und radiologische Verringerung des Tumorgewebes um
mindestens 90 %, sodass danach versucht wird, Extremitäten erhaltend zu resezieren. Insgesamt
wird durch die neoadjuvante und adjuvante Polychemotherapie nach kurativer Resektion ein
metastasenfreies Überleben von über 70 % erreicht. [1;2;3] Wenn das Osteosarkom erst im
metastasierten Stadium entdeckt wird, sind die Tochtergeschwülste meist hämatogen in die Lunge
gestreut, sehr selten auch in die regionalen Lymphknoten. Lungenmetastasen werden zwar
routinemäßig entfernt, bedeuten allerdings für 50-70 % der Patienten längerfristig eine infauste
Prognose. [17,18;19;20]
Mikrometastasen sind jedoch mit herkömmlichen Bildgebungsverfahren nicht darstellbar und
dementsprechend auch nicht resezierbar. Daher ist ein weiterer Therapieansatz die Modulation der
körpereigenen Tumorabwehr. Dafür erhielt Mifamurtid 2010 eine Zulassung als sogenanntes
Orphan-Drug (eine besondere Zulassung für Medikamente gegen sehr seltene Krankheiten).
Grundlage dafür ist eine Phase-III-Studie mit 678 Osteosarkompatienten. Im Vergleich zur
alleinigen Polychemotherapie mit Doxorubicin/Cisplatin/Methotrexat konnte die zusätzliche Gabe
von Mifamurtid die 6-Jahres-Überlebensrate von 70 auf 78 % signifikant erhöhen. [21]
Mifamurtid bewirkt eine Aktivierung des Immunsystems, da es bakteriellen Bestandteilen ähnelt
und der Körper über Freisetzung von Botenstoffen auf die potentiell schädliche Substanz reagiert.
Im Rahmen der dadurch gesteigerten Aktivität des Immunsystems wird auch die humorale und
zytotoxische Aktivität gegen Tumorzellen erheblich gesteigert.
Dieses Prinzip dieser Tumortherapie ist nicht neu. Es geht auf den amerikanischen Chirurgen
William Coley zurück, der im 19. Jahrhundert krankheitserregende Bakterien in Tumorgewebe
injizierte und seinen Angaben zufolge – kontrollierte klinische Studien gab es noch nicht - eine
Tumorregression erreichen konnte.
Heutzutage kennt man die Mechanismen der Tumorabwehr durch das Immunsystem sehr viel
genauer und ein wichtiger Bestandteil ist der Botenstoff Tumor Nekrose Faktor α (TNFα), der im

EINLEITUNG
6
Zuge einer Immunreaktion gebildet wird und Immunzellen, wie zum Beispiel Makrophagen,
stimuliert. Deren Aufgabe ist neben der Abwehr pathogener Mikroorganismen auch die
Demarkierung entarteter körpereigener Zellen. Über die folgende Aktivierung zellinterner
Signalkaskaden, die unter anderem Nuclear Factor-κB (NFκB) und Mitogen-Assoziierte-Protein-
(MAP)-Kinasen beinhalten, wird der Untergang entarteter Zellen induziert. [22]
Wenn im Organismus also eine Infektion mit bakteriellen Keimen simuliert wird, reagiert er
stereotyp mit der Initiation einer Entzündungsreaktion, die zu gesteigerter Kompetenz und
Effizienz des Immunsystems führen soll.
Diesen Effekt versucht man beim Einsatz von Mifamurtid zu nutzen. Als synthetisches Analogon
eines Peptidoglykan-Polymers namens Muramyldipeptid ähnelt es bakteriellen Zellwänden und
wird daher von einer Untergruppe der NOD-like-Rezeptoren, speziell NOD2, gebunden und der
NFκB-Signalweg aktiviert. Zusätzlich wird über Bindung an den NOD-like-Rezeptor 3 die Bildung
eines Inflammosomes angeregt. Beide bewirken in Synergie die Sekretion von immunreaktiven
Signalstoffen wie TNFα, Interleukin 1β oder Interleukin 6. [23;24] Dadurch wird eine systemische
Immunreaktion getriggert, die eine potentielle Bedrohung von außen abwehren soll, allerdings
werden durch die gesteigerte „Wachsamkeit“ des Immunsystems auch endogene Bedrohungen wie
zum Beispiel entartete Tumorzellen bekämpft (siehe Abbildung 2). [25]
Der anti-onkogene Nebeneffekt wurde in der oben genannten klinischen Studie gezeigt, was
allerdings auf zellulärer Ebene dazu führt, ist aktuell noch unklar und deswegen thematischer
Schwerpunkt der vorliegenden Untersuchung.
Abbildung 2 Schema der Wirkungsweise von Muramyldipeptid (Beschreibung im Text)
Abkürzungen: MPD – Muramyldipeptid, NOD2 - nucleotidebinding and oligomerization domain 2, NLRP3 – NOD-like-receptor P3, NFκB – Nuclear Factor-κB, TNFα – Tumor Necrosis Factor α, IL1β – Interleukin 1β

EINLEITUNG
7
2.2 ZELLLINIEN
Für die Untersuchungen wurden die Zelllinien MNNG/HOS und U-2 Os-Zellen gewählt. Erstere
stammt ursprünglich aus dem Tumorgewebe einer 13-jährigen Patientin. Dieser sogenannte TE-85
Klon F-5 wurde durch das chemische Kanzerogen N-Methyl-N-nitro-N-nitrosoguanidin in der
Konzentration 0,01 µg/ml transformiert und immortalisiert. [26;27] Die U-2 Os-Zellen wurden
einer 15-jährigen Patientin entnommen und in vitro kultiviert. [28]
Die beiden Osteosarkom-Zelllinien werden häufig zur Untersuchung von neuen Wirkstoffen und
deren Effekten auf Proliferation und Invasion verwandt. Zum Beispiel wurden im Jahr 2016 Studien
veröffentlicht, die den Einsatz oder die Wirkung von Kaltplasma, Vorinostat (ein Histon-
Deacytylasehemmer mit Orphan-Drug-Zulassung) oder Metformin (ein orales Antidiabetikum)
sowie Gadolinium (Kontrastmittel für MRT-Bildgebung) untersucht haben. [29;30;31;32]
2.3 KLASSISCHES RAS UND ALTERNATIVES RAS
Bislang nicht untersucht sind Expression und Funktion des alternativen Ang-(1-7)/Mas-Systems
bei den Osteosarkom-Zelllinien und dessen Einfluss auf die Tumorregression. [33]
Das alternative Ang-(1-7)/Mas-System ist eine vor relativ kurzer Zeit entdeckte alternative und oft
antagonistisch wirkende Achse des Renin-Angiotensin-Aldosteron-Systems (RAS, RAAS). Das
klassische RAS mit Angiotensin II (AngII) und Angiotensin-II-Typ-1-Rezeptor (AT1R) ist in der
Forschung schon lange etabliert [34], sodass die physiologische und pathophysiologische
Bedeutung seit geraumer Zeit Bestandteil der naturwissenschaftlichen Lehre ist. Lange bestand
Konsens darüber, dass AngII das Endprodukt einer proteolytischen Kaskade in (pulmonalen)
Blutgefäßen ist, deren Hauptaufgabe darin besteht, den Blutdruck und den Wasserhaushalt zu
regulieren. [35;36] Im Laufe der letzten Jahrzehnte wurden unabhängig davon agierende
Komponenten des RAS in bestimmten Organen (mit anderen Funktionen) entdeckt, die als lokales
RAS bezeichnet wurden. Im Gegensatz dazu wurde das humoral wirkende (und
blutdruckbeeinflussende) RAS als systemisches RAS tituliert.
Jedoch wurde auch die bis dato erforschte Geradlinigkeit des RAS durch Entdeckung von
alternativen Achsen, wie der Ang-(1-7)/Mas-Achse oder der Angiotensin IV/AT4R-Achse korrigiert
(siehe Abbildung 3) und bietet seit mittlerweile vielen Jahren einen ungemein großen Raum für
Forschungen, Neuentdeckungen und letztlich mitunter auch neuen Therapieansätzen vieler
Krankheiten. [37;38;39]
Das systemische RAS vermittelt seinen Einfluss auf Blutdruck und Wasserhaushalt über den
Protagonisten AngII, der über Bindung an AT1R Blutgefäße verengt und die Natrium-Retention
(und dadurch auch die Wasserretention) in der Niere erhöht. Außerdem steigert es über mitogene

EINLEITUNG
8
Signale zellproliferative Vorgänge. Die Enzyme Renin und Angiotensin-Converting-Enzyme (ACE)
sind verantwortlich für die biologische Aktivierung von AngII aus den Vorläuferproteinen
Angiotensinogen (Renin) und Angiotensin I (ACE). Das Steroidhormon Aldosteron wird nach
Anstieg des AngII-Spiegels im Blut aus der Nebennierenrinde freigesetzt und übt synergistische
Effekte vor allem in der Niere aus. [29;30]
Ein weiterer Metabolit von Angiotensin I (AngI) und AngII ist das Heptapeptid Angiotensin-(1-7),
Ang-(1-7), welches die Hauptrolle im alternativen RAS einnimmt. Eine Reihe von Enzymen wurde
bisher beschrieben, die häufig gewebespezifisch exprimiert werden und Ang-(1-7) aus
Vorläufermolekülen bilden. Durch die Enzyme Neutrale Endopeptidase (NEP, EC3.4.24.11), Prolyl-
Oligopeptidase (POP) und Thimet-Oligopeptidase (TOP) werden von dem Dekapeptid AngI
C-terminal drei Aminosäuren abgespalten, sodass Ang-(1-7) entsteht. POP sowie TOP sind auf das
Gefäßsystem beschränkt und NEP ist lokal in der Niere aktiv. [40;41;42] Jedoch ist Letztere
aufgrund der membranständigen Lokalisation im Lumen der Nierengefäße systemisch wirksam
und für den größten Teil der Ang-(1-7)-Produktion im Kreislauf verantwortlich. [43;44]
Einige Jahre nach diesen Erkenntnissen zeigten Studien, dass die Peptidase Angiotensin-
Converting-Enzyme 2 (ACE2) sowohl von AngI als auch AngII C-terminal eine Aminosäure
abspaltet. Aus dem Oktapeptid AngII entsteht direkt Ang-(1-7). Ausgehend von dem Dekapeptid
AngI entsteht Ang-(1-7) nach einem Zwischenschritt über Angiotensin-(1-9).
Abbildung 3 Schema des Renin-Angiotensin-Systems Die klassische (schraffierte) RAS-Achse mit AngII/AT1R sowie die Alternativen Achsen mit Ang-(1-7)/Mas und AngIV/AT4R.
* AT2R zählt nicht zum klassischen RAS, da er andere Effekte als AT1R bewirkt
Angiotensinogen
AngI
AngII
Renin
ACE1
Ang-(1-9)
Ang-(1-7)
ACE1 NEP, POP TOP
ACE2
ACE2
AT1R AT2R*
Mas-R
ALTERNATIV KLASSISCH ALTERNATIV
AngIII APA AngIV APN
AT4R
ANG-(1-7)/MAS ANGII/AT1R ANGIV/AT4R
V Y I H P F V Y I H P F R D V Y I H P R D V Y I H P F R
D V Y I H P F H R D V Y I H P F H R L

EINLEITUNG
9
Da die Affinität von ACE2 signifikant höher gegenüber AngII als gegenüber seinem
Vorläufermolekül AngI ist, wird dem indirekten Weg über Ang-(1-9) nicht so viel Bedeutung
beigemessen. Der direkte Weg von AngII zu Ang-(1-7) (und damit der alternativen Achse) wird
mittlerweile in verschiedenen pathophysiologischen Situationen als wichtiger Gegenspieler des
klassischen RAS bewertet. [45]
Im systemischen RAS ist vor allem die Balance zwischen der klassischen und der alternativen RAS-
Achse von Bedeutung. ACE2-Knockout Mäuse leiden entsprechend unter schweren
kardiovaskulären Dysfunktionen. [46] Allerdings führt auch ein Übergewicht der alternativen RAS-
Achsen ebenfalls zu pathologischen Veränderungen an Herz und Nieren. [47;48;49]
In dem Mas-Protoonkogen, welches bereits 1986 entdeckt wurde, fand sich um die
Jahrtausendwende der Rezeptor, welcher für die Wirkung von Ang-(1-7) verantwortlich gemacht
wird. [50;51] Der Ang-(1-7)-Rezeptor Mas ist ein G-Protein gekoppelter Rezeptor (GPCR), der als
Mitglied einer sehr großen Familie von membranständigen Rezeptoren, genau wie seine
Homologen sieben Transmembrandomänen besitzt. Üblicherweise wird bei Stimulierung der
GPCRs ein G-Protein aktiviert, welches eine Signalkaskade in Gang setzt.
Die genauen Wirkmechanismen des Mas-Rezeptors sind allerdings weiterhin Bestandteil
intensiver Forschungen. Aktuell besteht Einigkeit darin, dass die Gabe von Ang-(1-7) nach
Aktivierung der p38 Kinase zur Freisetzung von Arachidonsäure führt. [47;51] Außerdem wird die
NO-Synthase über einen PI3-Kinase-abhängigen Signalweg aktiviert. [52;53] Obwohl Karnik et al.
vermuten, dass second messenger Moleküle wie cAMP und IP3 nicht Teil der intrazellulären
Signalkaskade sind [54;55], konnten mehrere Arbeiten eine Mas-abhängige Aktivierung der
Adenylatcyclase und einen damit verbundenen Anstieg der intrazellulären cAMP-Konzentration
nachweisen. [56;57] Des Weiteren wird kontrovers diskutiert, ob Ang-(1-7) die PI3-Kinase
ausschließlich aktiviert oder möglicherweise auch hemmt. Einige Arbeiten zeigen eine Ang-(1-7)
abhängige Aktivierung der PI3-Kinase [53;58;59] und andererseits berichten sowohl Ni et al. als
auch Zhang et al. über eine Inaktivierung der PI3-Kinase. [60;61] Eine inhibierende Wirkung von
Ang-(1-7) auf die Extrazellulär Signal Regulierte Kinase 1/2 (ERK1/2) ist ebenfalls beschrieben.
Letztgenannte Kinase ist ein wichtiger Mediator von Proliferation, Migration und
Inflammation. [61; 62;63]
Insgesamt ist der intrazelluläre Signalweg der Ang-(1-7)/Mas-Achse bis dato nicht ausreichend
aufgeklärt. Neueste Phosphoproteom-Analysen haben zwar zahlreiche nach Ang-(1-7)-Gabe
regulierte Phosphoproteine identifiziert, jedoch lassen sich weiterhin keine klaren Aussagen zu den
Bedeutungen einzelner Signalkaskaden in Bezug auf den Mas-Rezeptor ableiten. Die beteiligten
Kinasen werden Vorgängen zugeschrieben, die zum Beispiel mit den intrazellulären Mechanismen
der Karzinomentstehung, p53-Signaling, Apoptose oder Zellzykluskontrolle zusammenhängen.

EINLEITUNG
10
Außerdem haben Ang-(1-7)-abhängig regulierte Proteine Einfluss auf das VEGF- und Insulin-
Signaling. [64;65]
Die Wirkungen der Ang-(1-7)/Mas-Achse auf den Organismus sind sehr vielfältig und Ang-(1-7)
scheint insgesamt ein Gegenspieler der AngII/AT1R-Achse zu sein. [45]
Es verbindet antihypertensive, antiarrhythmische und kardioprotektive Effekte, jedoch kann es in
zu hoher Dosierung zu Arrhythmien führen. [66;67;68;69;70] In der Niere fördert das Heptapeptid
die Durchblutung und steigert die glomeruläre Filtrationsrate, sowie die Natrium- und
Wasserausscheidung. [71;72]
Ang-(1-7) wird durch ACE (und NEP) wieder abgebaut, was möglichweise zu einem gewissen Teil
zu den kardio- und nephroprotektiven Effekten von ACE-Hemmern beiträgt. [73;74]
Während das systemische RAS bereits seit Langem klinische Relevanz in der Behandlung von
Bluthochdruck und Herzinfarkt besitzt, sind die lokalen RAS – und insbesondere die alternativen
Achsen – gegenwärtig Bestandteil vieler Forschungen.
Wie beim systemischen RAS haben die klassische und die alternativen Achsen meist
antagonistische Effekte, weshalb im Folgenden hauptsächlich die Wirkungen von Ang-(1-7)/Mas
beschrieben werden. Der Mas-Rezeptor vermittelt einige sehr positive Wirkungen, die bei vielen
Krankheitsbildern in der westlichen Welt einen möglichen therapeutischen Ansatz erhoffen lassen.
So konnte gezeigt werden, dass Ang-(1-7) die Proliferation von Fibroblasten und die Produktion
von Kollagen inhibiert. Damit verringert es beispielsweise kardiales Remodeling, Leber- oder
Lungenfibrose. [75;76;77]
Mit AVE-0991 einem Ang-(1-7)-Rezeptor-Agonisten, konnte bei Mäusen und Ratten mit exogen
induzierter Arthritis eine Verringerung von Neutrophilen, proinflammatorischen Zyto- sowie
Chemokinen und der Leukozytenadhäsion in den entzündeten Gelenken gezeigt werden. [78]
Gesundheitsökonomisch relevant könnte die antiinflammatorische Wirkung des alternativen RAS
auf die Artherosklerose sein, da die Inflammation des Endothels als Hauptrisikofaktor für
kardiovaskuläre Ereignisse gilt und damit die häufigste Todesursache in Deutschland darstellt. [79]
Im Tiermodell konnte der Progress der Artherosklerose durch AVE-0991 gestoppt werden. Diese
Beobachtung liefert damit einen Ansatz zur Prophylaxe von Krankheiten wie Herzinfarkt,
peripherer arterieller Verschlusskrankheit und Apoplexie. [80]
Ein weiterer sehr interessanter Aspekt ist die Wirkung der alternativen RAS-Komponenten auf die
Glucose-Homöostase von Nagetieren. So konnte gezeigt werden, dass die Aktivierung von Mas
einen antihyperglykämischen Effekt hat, der sowohl auf einer Attenuierung der peripheren
Insulinresistenz, als auch auf der Verstärkung der Glukose-stimulierten Insulinsekretion
beruht. [56;81;82]

EINLEITUNG
11
Auch im zentralen Nervensystem (ZNS) nimmt das RAS eine wichtige Rolle ein. Jedoch wird der
klassischen Achse mit AngII weniger Bedeutung beigemessen als in anderen Geweben. Dafür haben
Angiotensin III (AngIII; Ang 2-8) und Angiotensin IV (AngIV; Ang 3-8) – Metabolite von AngII, die
durch die Aminopeptidasen A und N gebildet werden – im ZNS wichtige Aufgaben. [83;84]
In Synergie zum systemischen RAS steigert AngIII über das Antidiuretischen Hormon (ADH) aus
dem Zwischenhirn die Natrium-Retention in der Niere und den Blutdruck durch Verengung von
Arteriolen im Gefäßsystem. Außerdem werden zentral das Durstgefühl und der Salzhunger
gesteigert, damit von außen Flüssigkeit und Natrium zugeführt werden. [85] AngIV hingegen übt
andere Effekte aus. Einerseits wird der zerebrale Blutfluss verstärkt und andererseits ist es für
Gedächtnis- und Lernprozesse von Bedeutung. [86] Sehr wichtig scheint auch hier die
Ang-(1-7)/Mas-Achse zu sein, denn unter den Angiotensinen im ZNS ist Ang-(1-7) am häufigsten.
Der Hippocampus, ein obligater Teil des Gedächtnisses, zeigt dabei die höchsten Ang-(1-7) Spiegel.
Bei Mas-Knockout Mäusen wurde eine defizitäre Objektwiedererkennung festgestellt, sodass der
Mas-Rezeptor für Gedächtnis- und Lernprozesse relevant ist. [87;88;89]
Zusammenfassend lässt sich feststellen, dass das Spektrum der alternativen RAS Komponenten in
seinen Wirkungen sehr weitreichend und verschieden ist, denn die Aufzählung über Ang-(1-7)
könnte weiter über die Bedeutung in Plazenta und Fortpflanzungsorganen, sowie Beeinflussung
des Augeninnendrucks oder antiischämische Effekte im ZNS geführt werden. In Abbildung 4 sind
die verschiedenen Wirkungen des Mas-Rezeptors in unterschiedlichen Geweben
dargestellt. [90;91;92;93;94;95;96;97]

EINLEITUNG
12
Abbildung 4 Wirkungen des Mas-Rezeptors modifiziert nach Passos-Silva et al. [97] Der G-Protein gekoppelte Rezeptor hat viele verschiedene Downstream-Effektoren (grau), die er entweder inhibiert () oder aktiviert (⊕). Dadurch hat er sehr verschiedene Wirkungen in verschiedenen Geweben.
Aktuell gibt es nur wenige Daten über das lokale RAS im Knochen – dem Zielorgan der vorliegenden
Arbeit. Teile des klassischen RAS mit AT1-Rezeptor, Renin und ACE wurden auf Osteoblasten
nachgewiesen. [98;99] Durch Inhibierung dieser Achse mit Losartan (AT1-Rezeptorblocker) oder
Aliskiren (Renininhibitor) wurde unter anderem die Aktivierung des ERK 1/2 Signalweges oder die
Freisetzung von IL1β und TNFα gehemmt. [98;100] In einer Vorarbeit konnte erstmals die
Expression von Bestandteilen des alternativen RAS in Osteosarkomzellen
nachgewiesen werden. [33]
Besonderes Augenmerk liegt in dieser Arbeit auf der Wirkung des lokalen RAS in Bezug auf
Tumorwachstum. Es konnten bereits mehrfach onkogene Effekte durch die AngII/AT1Rezeptor-
Achse beispielsweise in cholangio- und hepatocellulären Karzinomzellen sowie in Nieren- und
Pankreaskarzinomzellen nachgewiesen werden. [60;63] Andererseits kann eine Stimulierung des
Mas-Rezeptors über die Inhibierung des ERK 1/2 Signalweges das Zellwachstum verlangsamen.
Anhand von Bronchial- und Mammakarzinomzellen wurde gezeigt, dass Ang-(1-7) die Proliferation
inhibiert. Das konnte sogar im Mausmodell in vivo bestätigt werden. [62;101;102;103]
Die vorliegende Arbeit liefert einen kleinen Beitrag zum Verständnis des Einflusses der alternativen
Ang-(1-7)/Mas-Achse auf das Wachstum von Osteosarkomen, vor allem in Zusammenhang mit
Mifamurtid.
Antifibrotisch Antidiabetogen Antiproliferativ Antiinflammatorisch
Mas-Rezeptor
VEGF CTGF JNK
JNK ERK COX2 HDAC1 VEGF-A ⊕ FOXO1
ERK c-Src NFκB NADP(H)- Oxidase
ERK NFkB c-Src NADP(H)-Oxidase ⊕ PI3K ⊕ PPARγ ⊕ cAMP ⊕ AS160
Myokard Lunge Leber
Bronchialkarzinom Mammakarzinom
Artherosklerose Arthritis
Pankreas Fettgewebe
β γ α

EINLEITUNG
13
2.4 TRPC-KANÄLE
G-protein-gekoppelte Rezeptoren können auf die Aktivität von TRPC-Kanälen Einfluss
nehmen..[104] Inwieweit der Ang-(1-7)-Rezeptor Mas mit der Expression/Aktivität von TRPCs
assoziierte ist und ob dies beim Osteosarkom von Relevanz ist, wurde bislang nicht untersucht.
Die TRPC Kanäle sind Teil der TRP-Superfamilie, einer Gruppe von homologen Ionenkanälen, die
mit dem TRP-Gen von Drosophila melanogaster verwandt sind. Phylogenetisch teilt man die TRP-
Superfamilie in drei Zweige ein: die TRPC-Kanäle, die TRPM-Kanäle (für Melastin) und TRPV-
Kanäle (für Vanilloid). [105]
Die TRPC Kanäle wurden zuerst als Funktionseinheiten des Store-operated Calcium Entrys (SOCE)
beschrieben. SOCE bezeichnet den Calcium Einstrom von extrazellulär nach intrazellulär über
Kanäle in der Zellmembran – zum Beispiel TRPC-Kanäle – die durch den Abfall der Calcium-
Konzentration im Inneren des Endoplasmatischen Retikulums (ER) aktiviert werden.
[106;107;108]
Nacheinander wurden sieben Kanäle entdeckt (TRPC1 bis TRPC7), jedoch stellt TRPC2 im
menschlichen Genom ein Pseudogen dar und besitzt keine Funktion mehr. Die TRPC Kanäle sind
aber nicht nur Teil des SOCE, sondern sind auch als second messenger-operated Channels (SMOC)
Bestandteil in anderen Signalwegen. Eine schematische Übersicht gibt Abbildung 5 wider. [104]
Abbildung 5 Stark vereinfachte Darstellung von SOCE und SMOC SOCE - Calcium-Depletion im ER (1) führt über Interaktionsproteine zur Aktivierung der SOCE-Kanäle (2), sodass Calcium in die Zelle strömt und über Calcium-Pumpen ins ER befördert wird (3).
SMOC – Ein Agonist bindet an den transmembranalen Rezeptor (4) und aktiviert einen second messenger (5), der einen Calcium-Kanal öffnet (6). Calcium strömt ins Zytosol und kann unter anderem wieder ins ER gepumpt werden (7).
Agonist
Rezeptor
second messenger
SMOC TRPC 3 TRPC 5 TRPC 6 TRPC 7
SOCE
Homo- Hetero-tetramere tetramere
Endoplasmatisches Retikulum Ca2+ Ca2+↓ 1
2
3
4
5
6
Ca2+ Ca2+ Ca2+
TRPC 1∩1 TRPC 4∩4
TRPC 1∩3 TRPC 4∩7 2
7

EINLEITUNG
14
Neben der Abstammung ist den Kanälen der TRP-Superfamilie der strukturelle Aufbau aus einem
Protein mit jeweils sechs Transmembrandomänen, die jeweils zu viert ein porenbildendes
Tetramer bilden, gemeinsam und innerhalb einer Subfamilie (wie z.B. TRPC) können die vier
Untereinheiten Heterotetramere bilden, siehe Abbildung 6. [109;110]
Abbildung 6 Schematischer Aufbau eines TRPC-Kanals Jeweils sechs Transmembrandomänen der vier Untereinheiten bilden einen Tetramer. Zwei Transmembrandomänen jeder Untereinheit sind für die Porenbildung verantwortlich.
Die Kanaleigenschaften der TRPC Kanäle werden weitestgehend durch deren Interaktionspartner
bestimmt. Sie haben Einfluss auf die Anzahl und Lokalisation der Kanäle sowie
Kanalöffnungseigenschaften und damit insgesamt auf die Aufgaben der Kanäle auf zellulärer oder
organischer Ebene. Aufgrund der komplexen Interaktionen der TRPC-Kanäle können die
vielfältigen Funktionen nicht vollständig beschrieben werden, daher sind nachfolgend wesentliche
Charakteristika aufgeführt.
TRPC1 ist im Gegensatz zu den anderen TRPCs am ehesten eine wichtige Komponente des SOCE. Es
wurde gezeigt, dass Calcium Einstrom nach Depletion zellinterner Speicher über TRPC1 Kanäle
fließt. [111;112] Dabei ist der molekulare Aufbau verschieden, sodass einige Calcium-selektiv sind,
andere hingegen unselektive Kationenkanäle bilden. [113;114]
TRPC1 bildet mit vielen Proteinen Komplexe, so kann TRPC1 mit TRPC4 oder TRPC3
Heterotetramere bilden, allerdings bildet TRPC1 auch Homotetramere. [109] Mit dem Inositol-
trisphosphat-Rezeptor (IP3R), HOMER, STIM1 oder Orai1 sind einige Interaktionspartner genannt,
welche die Aktivität des Kanals direkt regulieren. [115;116;117] Dabei ist der IP3-Rezeptor
besonders hervorzuheben, da es der Interaktion von IP3R und TRPC1 zur Aktivierung des SOCE
bedarf. Der IP3-Rezeptor ist ein Calcium-Kanal an der Membran des ER und führt nach der Bindung
von IP3 zur Entleerung der endoplasmatischen Calcium-Speicher, sodass die Regeneration durch
SOCE ein folgerichtiger Schritt ist.
Der Kanal scheint eine Rolle bei Flüssigkeitssekretion, Endothelzellmigration und -permeabilität,
Muskelzellfunktion und Wundheilung zu spielen, sowie Zellproliferation, Zelldifferenzierung und
antiapoptotische Effekte zu fördern. [111;112;118;119]
Das Pseudogen TRPC2 des Menschen besitzt bei Nagetieren eine Funktion im Pheromon-sensitiven
Vomeronasalorgan. Untersuchungen konnten zeigen, dass TRPC2-Knockout Mäuse ihre

EINLEITUNG
15
Artgenossen „nicht mehr riechen können“ und Störungen im Sexual- und Sozialverhalten
zeigten. [120;121]
Die Funktion des TRPC3 wird durch das Ausmaß der Expression bestimmt, wobei hohe Level zur
Bildung von Homotetrameren mit konstitutiver Aktivität führen, ohne von den Calcium-Speichern
gesteuert zu werden. Eine geringe Expression fördert die Formung von TRPC1-TRPC3-
Heterodimeren, die wiederum Calciumspeicher-abhängig sind. TRPC3 ist eine wichtige Grundlage
für die artgerechte Entwicklung des zentralen (besonders zerebellaren) Nervensystems. Die
Überexpression in transgenen Mäusen führte hingegen zu lebensverkürzender Kardiomyopathie
durch Hypertrophie. Der Nachweis erhöhter Protein-Expression bei Patienten mit
schwerer Hypertonie oder mit (idiopathischem) Bluthochdruck im Lungenkreislauf
weist auf einen pathophysiologischen Zusammenhang von TRPC3 und dem Herzkreislauf-
system hin. [122;123;124;125;126;127]
TRPC4 ist eng verwandt mit TRPC5, drei Viertel ihrer Aminosäuresequenzen gleichen einander.
Trotzdem sind ihre Funktionen divers. Ersterer zeigte im Tiermodell Bedeutung bei
Endothelzellfunktion, Kontraktilität glatter Gefäßmuskelzellen sowie der kornealen
Epithelzellproliferation. [128;129] TRPC5 scheint für neuronales Wachstum von Bedeutung zu
sein. [130;131] Über die Zuordnung zu SOCE oder SMOC gehen die Meinungen bei beiden Kanälen
auseinander. [132;133;134]
TRPC6 hat im Hinblick auf Aminosäuresequenz eine große Ähnlichkeit mit TRPC3 sowie TRPC7 und
kann Heterotrimere bilden, welche derzeit noch nicht gut untersucht sind. Anderseits konnte in
Tiermodellen die Bedeutung des Kanals für die Kontraktilität glatter Gefäßmuskelzellen gezeigt
werden und es ist gesichert, dass TRPC6 vor allem durch Diacylglycerol und nicht durch interne
Calcium-Depletion aktiviert wird. [135;136]
TRPC7 ist sowohl Bestandteil des SOCE als des SMOC und wurde in einigen Regionen des Gehirns
nachgewiesen. Es wird angenommen, dass er an der Ausbildung von neuronalen Störungen
beteiligt ist. Dafür spricht, dass bei Patienten mit bipolaren Störungen eine signifikante
Verringerung der Genexpression in B-Lymphoblasten zu finden ist. [104;135;137;138]
Zusammenfassend kann gesagt werden, dass die Familie der TRPC Kanäle in vielen Geweben
mitunter sehr unterschiedliche Funktionen ausüben, die noch viel Raum für weitere
Untersuchungen bieten.
Inwieweit Osteosarkomzellen TRPC-Kanäle exprimieren und welche funktionelle Relevanz diese
haben, war Gegenstand der vorliegenden Arbeit.

EINLEITUNG
16
2.5 ZIELSTELLUNG
Mit der vorliegenden Arbeit sollte (I) eine umfassende Beschreibung der TRPC-Expression in
Osteosarkomzellen unter basalen Bedingungen vorgelegt werden, (II) die Einflüsse der
Modulation der alternativen Ang-(1-7)/Mas Achse des RAS (mittels Ang-(1-7) und AVE0991) bzw.
(III) der Einfluss der Immunstimulation (IL-1β) auf die Expression von TRPC-Kanälen
systematisch untersucht werden.
Dazu wurde ein in vitro-Modell genutzt, in dem mittels Administration von Interleukin 1β die
immunmodulatorische Wirkung nachgebildet wird, die den klinisch nachgewiesenen Effekten von
Mifamurtid bei der Therapie des Osteosarkoms zugrunde liegt und welches die Analyse der
Auswirkungen auf die Proliferation von Osteosarkomzellen ermöglicht.
Dabei lag der Fokus auf der quantitativen Analyse von mRNA-Mengen für die einzelnen TRPC-
Kanäle unter den verschiedenen experimentellen Bedingungen.

17
3 MATERIAL UND METHODEN
3.1 MATERIAL
GERÄTE HERSTELLER/LIEFERANT
Agarosegelkammer Bio-Rad
Feinwaage Pt600 Sartorius
Feinwaage AC210S Analytic Sartorius
Brutschrank HeraCell
Eismaschine KF 85 Migel
Fluoreszensreader infinite F200 Tecan
Invertierer Sarstedt
Mikroskop Telaval 3 Carl Zeiss Jena
Mikrowelle Severin
Nanodrop 2000c Thermo Scientific
PCR Cycler Biometra
Reinstwasseranlage Milli-Q Synthesis A 10 Millipore
Laminarbox Thermo Scientific
Thermocycler CFX69 Real-Time-PCR Detection System Bio-Rad
Neustädter-Zählkammer Neustädt
pH-Meter Mettler Toledo
VacuGene Pump Pharmacia LKB
Wasserbad 37 °C Techne
Zellinkubator CO2 Cell Medcenter GmbH
Zentrifuge RS54 Eppendorf
Zentrifuge Zellzucht Hettich Zentrifuge

MATERIAL UND METHODEN
18
HILFSMITTEL UND VERBRAUCHSMATERIALEN
6-Well-Gewebekulturschalen Nunclon
96-Well-Platten Nunclon
BZO Seal Film Biozym
Einfrierbox Cryo-SafeTM -1°C neoLab
Falconröhrchen 15 ml Sarstedt
Falconröhrchen 50 ml BD Biosciences
Kryo-Röhrchen Sarstedt
Pasteurpipetten Brand
PCR 96-Well-Platten TW-MT-Platte Biozym
Pipettierhilfe Eppendorf
Pipettenspitzen 0,5-10 µl, 10-200 µl, 100-1000 µl Sarstedt
Reaktionsgefäße 0,2 ml, 1,5 ml, 2,0 ml Sarstedt
Zellkulturflaschen TC25, TC75, TC175 Sarstedt
Zellschaber TPP

MATERIAL UND METHODEN
19
CHEMIKALIEN HERSTELLER/LIEFERANT
Acrylan Antiseptica
AVE-0991 Sanofi Aventis
D-Alanin7-Ang-(1-7) Bachem
DMSO AppliChem
Dinatriumhydrogenphosphat Sigma
Ethanol 96 %, vergällt und unvergällt Zentralapotheke
FCS PAN Biotech
GeneRulerTM 100bp DNA Ladder Fermentas
Guanidiniumhydrochlorid Ferak
Hydrogenchlorid Sigma
Interleukin 1β PAN Biotech
Isopropanol Roth
Kaliumhydrogenphosphat Roth
L-Glutamin PAA
Ly294002 Sigma
McCoy´s 5A medium PAA
MEM with Earle´s Salts PAA
Natriumchlorid Roth
Natrium-Pyruvat Sigma
Natriumhydroxid Sigma
PBS PAA
PD98059 Sigma
Penicillin/Streptomycin PAA
RedSafeTM Intron Biotechnologies
TriFastTM peqGOLD
Trypsin Sigma

MATERIAL UND METHODEN
20
ZELLLINIEN UND KULTURMEDIEN
MNNG/HOS American Type Culture Collection
Katalognummer: CRL-1547TM
Kulturmedium MEM with Earle´s Salts
10 % FCS
Natrium Pyruvat
100 U/ml Penicillin
100 µg/ml Streptomycin
37°C, 5 % CO2
U-2 Os American Type Culture Collection
Katalognummer: HTB-96TM
Kulturmedium McCoy´s 5A medium
10 % FCS
Natrium Pyruvat
100 U/ml Penicillin
100 µg/ml Streptomycin
37°C, 5 % CO2
KITS HERSTELLER/LIEFERANT
MaximaTM First Strand cDNA Synthesis Kit Fermentas
RevertAidTM First Strand cDNA Synthesis Kit Fermentas
SensiMixTM SYBR Kit Bioline
Analytik Jena RNA Kit Analytik Jena

MATERIAL UND METHODEN
21
PRIMER
Zielgen Sequenz Amplifikatgröße (bp) Tannealing
Tubulin 528 63 °C vorwärts CAATGCCGACTGGTACTTG
rückwärts ACATCTCAACTGGCAGGAAC Mas-Rezeptor 407 63 °C
vorwärts CAATGCCGACTGGTACTTG rückwärts ACATCTCAACTGGCAGGAAC
TRPC1 177 60 °C vorwärts TTCTTGCTGGCGTGCGACAAG
rückwärts CTGACAACCGTAGTCCAAAAG TRPC3 492 60 °C
vorwärts AAGAAGGAGAACCTGGCGCG rückwärts GTCATTCTTGAACTCCTTCTC
TRPC4 126 56 °C vorwärts CTGCTTATCAAGGAGTATAGGA
rückwärts GTCTATGTTGGAGATGCTCTAT TRPC5 210 56 °C
vorwärts GTTGTAACTTGTTCTTCCTGTC rückwärts CAGTGAGGTAGAATTAGGTGAA
TRPC6 191 55 °C vorwärts CCATCGTAACATTATAGACTCC
rückwärts GTTCAATCTCTACTCCTACTAC TRPC7 194 64 °C
vorwärts CTGCTTATCAAGGAGTATAGGA rückwärts GTCTATGTTGGAGATGCTCTAT
Alle Primer wurden von der Firma Invitrogen geliefert.

MATERIAL UND METHODEN
22
PUFFER UND LÖSUNGEN
Guanidiniumhydrochlorid 0,3 mol/l Guanidiniumhydrochlorid in 96 (v/v)% unvergälltem Ethanol
PBS 10x 10 mmol/l Kaliumhydrogenphosphat
1,5 mol/l Natriumchlorid
37 mmol/l Dinatriumhydrogenphosphat
pH 7,4
Probenpuffer 10x 0,2 mol/l EDTA
25 % Saccharose
0,25 (w/v)% Bromphenolblau
TAE-Puffer 50x 2 mol/l Tris
0,95 mol/l Eisessig
50 mmol/l EDTA

MATERIAL UND METHODEN
23
3.2 METHODEN
ZELLKULTUR
Alle folgenden Vorgänge und Experimente mit lebenden Zellen wurden an der Laminarbox unter
Einhaltung steriler Bedingungen durchgeführt. Die Inkubation erfolgte in einem 37°C warmen CO2
Brutschrank mit 5 % CO2-Gehalt sowie 95 % Luftfeuchtigkeit.
AUFTAUEN
Die Zellen wurden in einem Gemisch aus 80 % Kulturmedium, 10 % fetalem Kälberserum (FCS)
und 10 % DMSO in Kryogefäßen konserviert. Das weitere Verfahren während des Auftauens sollte
schnell erfolgen, da DMSO zytotoxisch auf Zellen wirkt. Deswegen wurden jeweils 1 ml der Proben,
sobald sie flüssig waren, in ein 15 ml Falconröhrchen mit 6-7 ml Kulturmedium überführt. Danach
wurde die Zellsuspension bei 700 × g 5 Minuten lang zentrifugiert und nach Verwerfen des
Überstandes erneut in 6-7 ml Kulturmedium resuspendiert. Dieses Gemisch wurde anschließend in
eine TC25-Zellkulturflasche übertragen, nach mindestens 24 Stunden Inkubation bei 37°C im CO2-
Brutschrank passagiert und fortan in einer TC75-Zellkulturflasche kultiviert.
KULTIVIEREN UND PASSAGIEREN
Um einen möglichst geringen Stress für die Zellen zu gewährleisten, mussten alle verwendeten
Flüssigkeiten, wie Kulturmedien, PBS oder Trypsin auf 37 °C erwärmt werden.
Zuerst wurde das Kulturmedium aus den Zellkulturflaschen mittels Saugpumpe und
Pasteurpipetten entfernt, der Zellrasen mit 10 ml Dulbecco´s PBS 1x gespült und dann je nach
Flaschengröße mit 1 ml, 2 ml oder 4 ml Trypsin-EDTA versetzt. Nach 3 Minuten Inkubation bei
37°C waren die Zellen von der Oberfläche gelöst und nach Resuspension mit dem Kulturmedium
auf ein Gesamtvolumen von 10 ml wurde dies in ein 50 ml Falconröhrchen überführt. Davon
wurden 10 µl in einer Neubauer Zählkammer zur Bestimmung der Zellzahl eingesetzt, während der
Rest der Zellsuspension bei 700 × g und 4°C 5 Minuten zentrifugiert wurde. Danach wurde das
Pellet nach dem Verwerfen des Überstandes erneut im Kulturmedium resuspendiert. Zum Erhalt
der Zelllinien wurden 3x105 Zellen in eine TC75-Zellkulturflasche mit 25 ml Kulturmedium
gegeben.

MATERIAL UND METHODEN
24
EINFRIEREN
Zum Konservieren wurden Zellen möglichst niedriger Passage genommen. Die Zellen wurden wie
oben beschrieben geerntet.
Das entstandene Pellet wurde nach dem Zentrifugieren abhängig von der Zellzahl so resuspendiert,
dass 4 x 106 Zellen in je 1 ml enthalten waren. Das Einfriermedium bestand zu 80 % aus dem
Kulturmedium und enthielt 10 % FCS sowie weitere 10 % DMSO, um die Zelllysierung durch
Kristallbildung während des Gefriervorgangs zu verhindern. Die Zellen wurden in dem
Einfriermedium in Kryoröhrchen überführt und in einem Isopropanolbehälter schonend
auf – 80 °C gekühlt, bevor sie im flüssigen Stickstoff auf lange Zeit gelagert wurden.
STIMULIERUNG/INHIBIERUNG
Für die Zellversuche wurden 5x105 Zellen in jedes Well einer 6-Well-Platte eingesät und mit 3 ml
Kulturmedium versorgt. Nach 24-stündiger Inkubation im CO2 Brutschrank wurden die Zellen
durch Zugabe von Substanzen in das Kulturmedium stimuliert oder inhibiert. Je nach Fragestellung
der Experimente fand die RNA Isolierung nach 3, 6 oder 12 Stunden, meistens jedoch nach 24
Stunden statt.
Folgende Agenzien waren Bestandteil der Versuche - Interleukin 1β in den Konzentrationen
500 pg/ml Kulturmedium, sowie zur Titrierung 5 pg/ml, 50 pg/ml, 500 pg/ml und 1.000 pg/ml,
außerdem AVE-0991, der Agonist des Mas-Rezeptors, hauptsächlich in der Konzentration
von 10-8 M im Kulturmedium, zur Überprüfung der optimalen Dosis in den Konzentrationen 10-7 M
und 10-6 M.
Als Antagonist des Mas-Rezeptors wurde D-Alanin7-Ang-(1-7) (D-Ala) in den Konzentrationen
10-7, 10-6, 10-5 M eingesetzt.
Die PI3-Kinase und ERK1/2 als Bestandteil des Mas-Rezeptor Signaling pathways, wurden mit
Ly294002 einem reversiblen PI3-Kinase Inhibitor und mit PD98059 einem Inhibitor der
Phosphorylierung und damit Aktivierung von ERK1/2 untersucht. PD98059 wurde auf 50 µM und
Ly294002 auf 100 µM im Kulturmedium verdünnt; bei der Kombination beider Substanzen wurden
die gleichen Mengen verwendet.
Da die Agenzien AVE-0991, Ly294002 und PD98059 in DMSO gelöst waren, wurden die jeweiligen
Kontrollen in den Versuchen mit gleichen Volumina DMSO versetzt. Die Kontrollsubstanz für D-Ala
und Interleukin 1β war jeweils einfaches Kulturmedium.

MATERIAL UND METHODEN
25
RNA ISOLIERUNG
Die RNA-Isolierung erfolgte entweder mit dem Analytik Jena RNA Kit oder mittels der Trizol-
Methode.
Die RNA wurde laut Protokoll des Analytik Jena Kits isoliert, sodass die RNA schließlich mit
30-75 µl RNase-freiem Wasser eluiert werden konnte.
Die RNA-Isolierung mit Trizol wurde nach Absaugen des Kulturmediums mit je 1 ml Trizollösung
pro Well der 6-Well-Platte begonnen. Nach 5 Minuten Einwirkzeit bei Raumtemperatur wurde das
Gemisch aus Trizol und Zellen mit einem Zellschaber abgelöst und in ein 1,5 ml Reaktionsgefäß
überführt. Nach Zugabe von 200 µl Chloroform in jeder Probe wurde invertiert und weitere drei
Minuten auf dem Invertierer bis zur einsetzenden Phasentrennung bei Raumtemperatur inkubiert.
Anschließend wurden durch Zentrifugation bei 12.000 × g und 4°C nach 15 Minuten die wässrige,
die Interphase und die organische Phase getrennt. Die obere wässrige Phase enthielt die RNA,
wohingegen die trizolhaltige, organische Phase Proteine beinhaltete. Die genomische DNA befand
sich zwischen beiden in der Interphase.
Die RNA in der wässrigen Schicht wurde in einem neuem 1,5 ml Reaktionsgefäß mit je 0,5 ml
Isopropanol zur Fällung gebracht. Dafür wurde zuerst das Isopropanol durch Intervertieren im
Gefäß verteilt und nach 10-minütiger Inkubation bei Raumtemperatur bei 12.000 × g und 4°C 12
Minuten lang zentrifugiert.
Das RNA-Pellet wurde nach Verwerfen des Überstandes dreimal gewaschen, das heißt mit 1 ml
unvergälltem 75 % Ethanol versetzt, 3 Minuten auf dem Invertierer inkubiert und bei 7.500 × g und
4°C für 5 Minuten zentrifugiert. Nach dem letzten Waschschritt wurde der Überstand erneut
verworfen und 20 bis 30 Minuten gewartet bis alle Ethanolreste verdunstet waren. Die getrocknete
RNA wurde mit RNase-freiem Wasser versetzt, um danach über Nacht im Kühlschrank zur
vollständigen Lösung gebracht zu werden.
Bis zur weiteren Verwendung wurde die RNA bei – 20°C kurzfristig und für längere Zeiträume
bei – 80°C gelagert.
REVERSE TRANSKRIPTION VON RNA IN CDNA
Um die RNA-Mengen mit quantitativer PCR (RT-qPCR) zu untersuchen, wurde jeweils 1 µg RNA in
cDNA umgeschrieben. Dafür wurde das MaximaTM First Strand cDNA Synthesis Kit von Thermo
Fisher Scientific benutzt. Die RNA-Konzentration wurde vorher photometrisch mit dem
NanoDropTM 2000c bestimmt und auf Verunreinigungen untersucht. Das Messprinzip beruht auf
der Analyse der unterschiedlichen Absorptionsmaxima kurzwelligen Lichts von RNA und
Proteinen.

MATERIAL UND METHODEN
26
Das Reaktionsgemisch für 30 µl setzte sich aus 6 µl Reaction Buffer Mix 5x, 3 µl Maxima Enzyme
Mix, 1 µg RNA sowie nucleasefreiem Wasser ad 30 µl zusammen. Nach kurzem Vermischen der
Proben folgten 10 Minuten Inkubation bei 25°C mit anschließender 15-minütiger Inkubation
bei 50°C. Die Reaktion wurde durch 5-minütige Inkubation bei 85°C terminiert.
Das Produkt („first strand cDNA“) konnte direkt in der quantitativen PCR eingesetzt werden. Die
Lagerung fand analog zur RNA bei – 20°C beziehungsweise bei – 80°C im Falle längerer
Aufbewahrung statt.
QUANTITATIVE PCR
Die quantitative Polymerasekettenreaktion (RT-qPCR) beruht auf dem Prinzip der herkömmlichen
Polymerasekettenreaktion (PCR) mit zeitechter Quantifizierung mittels Fluoreszenzmessung.
Diese dient der Amplifikation spezifischer DNA-Abschnitte. Oligonukleotide mit circa 20 Basen
binden nach Zerfall des DNA-Doppelstranges an komplementäre Abschnitte auf der Einzelstrang-
DNA. Als sogenannte Primer binden jeweils ein Vorwärts-Primer den Plus-Strang und ein
Rückwärts-Primer den komplementären Minus-Strang und grenzen den DNA-Abschnitt ein,
welcher amplifiziert werden soll. Im nächsten Schritt synthetisieren DNA-Polymerasen ausgehend
von den Primern in 5’-3’-Richtung einen neuen Strang, analog zur Matrize des jeweiligen DNA-
Stranges. Aus den beiden Einzelsträngen werden dadurch Doppelstränge, die wieder durch Hitze
denaturiert werden und in Einzelstränge zerfallen. Der Zyklus aus Denaturierung (1),
Primerbindung (2) und DNA-Synthese (3) kann wiederholt werden, bis ausreichend Kopien des
spezifischen DNA-Abschnitts zur Quantifizierung vorliegen. Eine schematische Darstellung findet
sich in Abbildung 7.
Bei der RT-qPCR kann die Menge der DNA-Amplifikate über Fluoreszenzmessung bestimmt
werden. Das Reaktionsgemisch der PCR wird durch einen Fluoreszenzfarbstoff erweitert. Dieser
bindet interkalierend in der doppelsträngigen DNA und nach Anregung durch Licht mit der
Wellenlänge von 494 nm emittiert der Farbstoff Licht mit 521 nm. Je mehr DNA amplifiziert wird,
desto mehr Farbstoff kann gebunden werden und fluoreszieren. Da sich theoretisch die Menge der
Amplifikate je Zyklus verdoppelt, steigt die Stärke der Fluoreszenz exponentiell an. Die Anzahl der
Kopien des Zielgens zu Beginn der RT-qPCR hat damit direkten Einfluss auf die Stärke der
Fluoreszenz. Die relative Quantität des spezifischen DNA-Abschnitts wird anhand eines
Floureszenzwertes bestimmt und in Zyklen – die benötigt wurden um den Schwellenwert zu
überschreiten – als sogenannter Cq-Wert angegeben. Mit dem absoluten Cq-Wert, können
Rückschlüsse auf die relative Menge der Kopien eines Zielgens zu Beginn der RT-qPCR gezogen
werden.

MATERIAL UND METHODEN
27
Die verwendete cDNA wurde wie oben beschrieben aus der RNA synthetisiert und jeweils ein 1 µl
diente als Template für die Amplifikation. Diese wurde mit dem SensiMixTM SYBR Hi-ROXKit, den
Vorwärts- und Rückwarts-Primern in einer Konzentration von 250 nM sowie nukleasefreiem
Wasser zur Reaktion gebracht. Das Kit enthielt in zweifacher Konzentration die DNA-Polymerase,
die Bausteine der DNA als desoxy-Nukleotidtriphosphate, Adenosintriphosphat als Energieträger,
Magnesium-Ionen, eine Pufferlösung sowie den Fluoreszenzfarbstoff SYBR Green.
Je analysiertem Zielgen wurden 2 Wells einer 96-Well-Platte mit einem Endvolumen von 20 µl
pipettiert – 1 µl cDNA, 1 µl Primer, 12 µl SensiMixTM SYBR Hi-ROX, 8 µl nukleasefreies Wasser.
Die RT-qPCR wurde mit dem Thermocycler CFX96 Real Time PCR Detection System von Bio-rad
nach folgendem Protokoll durchgeführt:
(1) 10 min 95 °C Denaturierung des inhibitorischen Antikörpers der DNA-Polymerase
(2) 10 s 95 °C Denaturierung der Doppelstrang-DNA
(3) 30 s [ ]°C Primeranlagerung, Tannealing siehe Material, Primer
(4) 30 s 72 °C Elongation und Messung der Fluoreszenz
(5) 39 Mal Wiederholung der Schritte (2) bis (4)
(6) 10 s 95 °C Denaturierung der Doppelstrang-DNA
(7) 5𝑠𝑠 0,5°𝐶𝐶� 65-95 °C Ermitteln der Schmelzkurve (zur Validierung des korrekten Amplifikates und zum Auschluss von Nebenprodukten)
(8) 4 °C Ende
Die Ergebnisse wurden mit dem Programm CFX ManagerTM von Bio-rad ausgewertet. Das
verwendete Prinzip ist die ΔΔCq-Methode, welche eine Normalisierung der Werte auf ein
Housekeeping-Gen (hier: α-Tubulin) beinhaltet. Damit ist eine Aussage über die relative Anzahl der
Startkopien des Zielgens im Vergleich untereinander möglich. Der Cq Wert entspricht dem Zyklus
in dem ein „Threshold“ – der bereits erwähnte Schwellenwert – überschritten wird und
nachfolgender Formel wird eine Ratio bestimmt, welche Fehler durch Unterschiede bei der RNA-
Isolierung sowie der Effizienz in der cDNA Synthese eliminiert.
𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟 = 2−∆∆𝐶𝐶𝐶𝐶
∆∆𝐶𝐶𝐶𝐶 = ∆𝐶𝐶𝐶𝐶𝐴𝐴𝐴𝐴𝐴𝐴𝐴𝐴𝐴𝐴 − ∆𝐶𝐶𝐶𝐶𝐾𝐾𝐾𝐾𝐴𝐴𝐾𝐾𝐾𝐾𝐾𝐾𝐾𝐾𝐾𝐾𝐴𝐴
∆𝐶𝐶𝐶𝐶 = 𝐶𝐶𝐶𝐶𝑍𝑍𝑍𝑍𝐴𝐴𝐾𝐾𝐴𝐴𝐴𝐴𝐴𝐴 − 𝐶𝐶𝐶𝐶𝐻𝐻𝐾𝐾𝐻𝐻𝐻𝐻𝐴𝐴𝐻𝐻𝐴𝐴𝐴𝐴𝐻𝐻𝑍𝑍𝐴𝐴𝐴𝐴−𝐺𝐺𝐴𝐴𝐴𝐴

MATERIAL UND METHODEN
28
Abbildung 7 Schematische Darstellung der quantitativen PCR
(Schritt 4) 30 s 72 °C Elongation
(Schritt 3) 30 s Primeranlagerung
(Schritt 4)
(Schritt 2) 10 s 95 °C Denaturierung der Doppelstrang-DNA
(Schritt 3)
(Schritt 2)
(Schritt 4)
(Schritt 3)
Primer aus ca. 20 Basenpaaren
DNA-Polymerase

MATERIAL UND METHODEN
29
AGAROSEGELELEKTROPHORESE
Die entstandenen PCR-Produkte wurden durch elektrophoretische Auftrennung der Nucleinsäuren
nach ihrer Größe im Agarosegel überprüft. Die negativ geladenen DNA-Moleküle wandern von dem
gemeinsamen Startpunkt Richtung positiv geladener Anode, kleine Moleküle laufen schneller durch
die Poren des Agarosegels als größere Moleküle. Bei den darzustellenden Größen zwischen 100 und
500 Basenpaaren eignete sich ein 1,5 %-iges Gel, welches mit dem Fluoreszenzfarbstoff RedSafeTM
versetzt wurde.
Nach Festwerden des Agarosegels wurden die Geltaschen nahe der Kathode mit dem Gemisch aus
10 µl PCR-Proben und 1 µl Probenpuffer beladen. Als Laufpuffer wurde TAE-Puffer verwendet und
bei einer Spannung von 100V wurden die Nucleinsäuren aufgetrennt. Durch die Interkalation des
Fluoreszenzfarbstoffes wurden die vorhandenen doppelsträngigen DNA-Banden mit UV-Licht
sichtbar gemacht. Bei spezifischer Produktbildung in der RT-qPCR sollte nur eine Bande entstehen,
die der erwarteten Sequenzlänge des PCR-Produktes (siehe Material, Primer) entspricht. Mit einer
DNA-Ladder wurde die Größe der DNA-Sequenzen bestimmt.
STATISTIK
Die Ergebnisse der RT-qPCR wurden der Software entnommen und mit GraphPad Prism zur
statistischen und graphischen Auswertung weiterverwendet. Die Parameter wurden mindestens
viermal durch unabhängige biologische Replikate erhoben. Diese Daten wurden nach Mann-
Whitney analysiert und Unterschiede in den Kontroll- bzw. Versuchsgruppen auf Signifikanz
überprüft. Abschließend wurden die Daten ebenfalls mit GraphPad Prism in Diagrammen mit
BoxPlots visualisiert.
30
4 ERGEBNISSE
4.1 EINFLUSS VON IL1β UND AVE-0991AUF DIE MAS-MRNA-GEHALTE
Wie bereits von Ender et al. gezeigt, werden die Komponenten der alternativen RAS-Achse auch in
den Osteosarkomzelllinien MNNG/HOS und U-2 Os-Zellen exprimiert. Dabei wurde außerdem ein
inhibitorischer Effekt von Interleukin 1β und Ang-(1-7) im Migrationsassay festgestellt. [33]
Der Einfluss von IL1β oder AVE-0991 auf die Mas-mRNA-Gehalte wurde nach 24-stündiger
Exposition der MNNG/HOS bzw. U-2 Os-Zellen analysiert.
Die Darstellung der Ergebnisse erfolgt in Box-Plot-Diagrammen. Die Behandlungsgruppen sind
jeweils zur Kontrollgruppe in Relation gesetzt, wodurch sich ein guter Überblick über die
Veränderungen der Transkriptmengen ergibt.
Wie in Abbildung 8 erkennbar, bewirkt die Administration der beiden Substanzen, IL1β und
AVE-0991, eine Veränderung der Mas-mRNA-Menge in beiden Zellen in unterschiedlichem
Ausmaß.
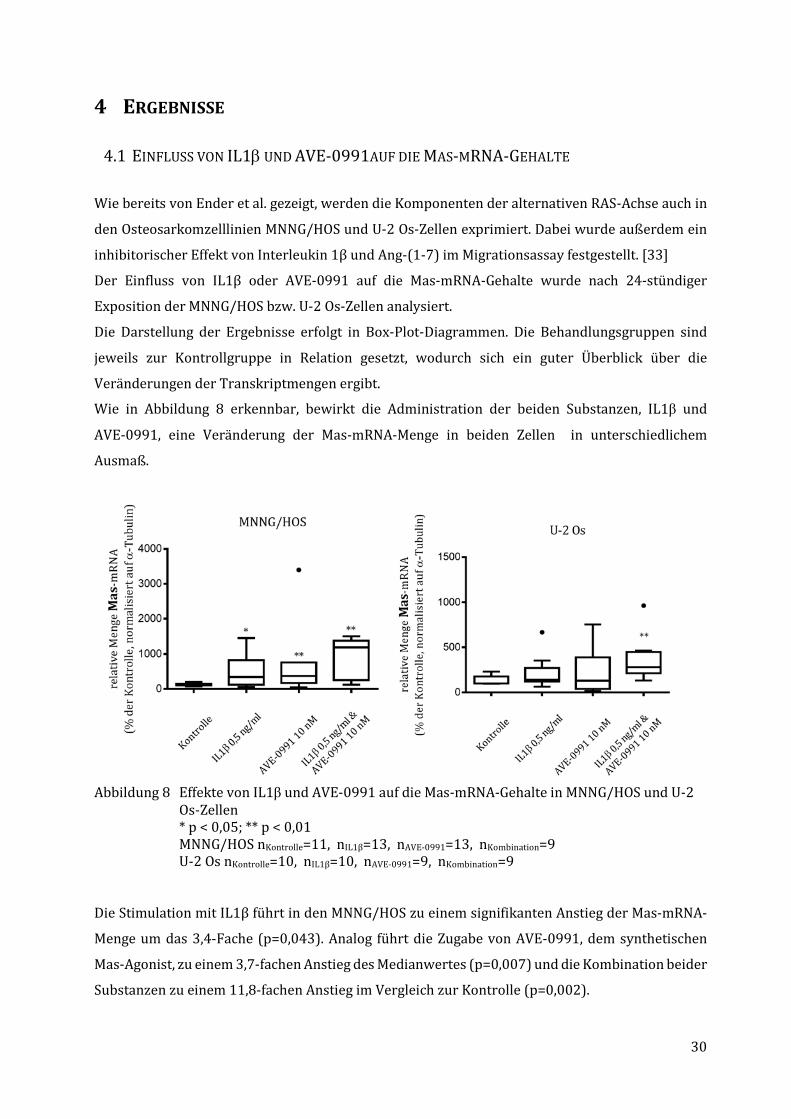
Abbildung 8 Effekte von IL1β und AVE-0991 auf die Mas-mRNA-Gehalte in MNNG/HOS und U-2 Os-Zellen * p < 0,05; ** p < 0,01 MNNG/HOS nKontrolle=11, nIL1β=13, nAVE-0991=13, nKombination=9
U-2 Os nKontrolle=10, nIL1β=10, nAVE-0991=9, nKombination=9 Die Stimulation mit IL1β führt in den MNNG/HOS zu einem signifikanten Anstieg der Mas-mRNA-
Menge um das 3,4-Fache (p=0,043). Analog führt die Zugabe von AVE-0991, dem synthetischen
Mas-Agonist, zu einem 3,7-fachen Anstieg des Medianwertes (p=0,007) und die Kombination beider
Substanzen zu einem 11,8-fachen Anstieg im Vergleich zur Kontrolle (p=0,002).

ERGEBNISSE
31
Bei den U-2 Os-Zellen sind die Effekte weniger deutlich – so ergibt die Stimulation mit IL1β zwar
einen leichten Anstieg des Medianwertes auf das 1,4-Fache des Kontrollwertes, allerdings ist hier
nur eine Tendenz erkennbar (p=0,06).
Durch die alleinige Zugabe von AVE-0991 steigt der Median der Gruppe auf das 1,3-Fache des
Kontrollwertes, jedoch ohne Signifikanz. Anders ist es beim gleichzeitigen Einsatz von AVE-0991
und IL1β. Dabei zeigt sich ein Anstieg des Medianwertes auf das 2,8-Fache des Vergleichswertes,
was statistisch gesichert ist (p=0,001).
Bei beiden Zelllinien fällt insbesondere der ähnlich stark ausgeprägte synergistische Effekt von
IL1β und AVE-0991 auf den Anstieg der Mas-mRNA-Mengen auf.
Diese auf RT-qPCR basierenden Ergebnisse zu den veränderten Mas-mRNA-Gehalten der
Osteosarkom-Zellen wurden durch die Visualisierung der PCR-Amplifikate im Agarosegel und
RedSafe-Färbung validiert. Durch die elektrophoretische Auftrennung der PCR-Produkte können
sowohl die korrekte Größe der Genabschnitte verifiziert und mögliche Nebenprodukte in Form von
zusätzlichen Banden dargestellt werden. Aus Abbildung 9 wird ersichtlich, dass die Primer für den
Mas-Rezeptor zu dem erwarteten Produkt mit 407 Basenpaaren geführt haben und unter den
Versuchsbedingungen keine Nebenprodukte amplifiziert worden sind. Das ist in völliger
Übereinstimmung mit den Ergebnissen der Schmelzpunktbestimmung, die am Ende jeder RT-qPCR
durchgeführt wurde. Ebenso bildeten die Primer für das konstitutiv exprimierte α-Tubulin nur das
erwartete Amplifikat mit 528 Basenpaaren.
Abbildung 9 Agarosegel zur Validierung der Spezifität von den humanen RT-qPCR-Primern für den
Mas-Rezeptor und α-Tubulin. Die Amplifikatgrößen von 407 bp und 528 bp entsprechen den aus der Sequenz abgeleiteten erwarteten Größen. Die cDNA-freien Kontrollen (H2O) zeigen keine Amplifikate.
mas-Rezeptor
407bp
α-Tubulin
528bp
500bp
400bp
300bp
200bp 150bp 100bp

ERGEBNISSE
32
4.2 MRNA-EXPRESSION DER TRPC-KANÄLE
Um die Grundlage für die Verknüpfung von antiproliferativen Effekten der Ang-(1-7)/Mas-Achse
und den TRPC-Kanälen zu schaffen, wurde zunächst der mRNA-Gehalt der individuellen TRPC
Kanäle unter basalen Bedingungen, das heißt ohne Zugabe von Agenzien, überprüft. Dafür wurden
die beiden Zelllinien in 6-Well-Platten eingesät, nach 24 Stunden mit neuem Kulturmedium ohne
Zusätze von Agenzien versorgt und nach weiteren 6 oder 24 Stunden für die quantitative Analyse
der TRPC-mRNAs geerntet.
Auf der mRNA-Ebene werden in beiden Zelllinien die TRPC-Kanäle 1 bis 7 exprimiert, wobei keine
deutlichen Differenzen zwischen MNNG/HOS und U-2 Os-Zellen beobachtet wurden. Die
Abbildung 10 gibt einen Überblick über die jeweiligen relativen mRNA-Mengen der Kanäle in
Relation zu dem konstitutiv exprimierten α-Tubulin.
Abbildung 10 Vergleich der mRNA-Mengen der TRPC-Kanäle in den Osteosarkomzelllinien
MNNG/HOS und U-2 Os in Relation zu der α-Tubulin-mRNA-Menge
Auffallend ist allerdings der etwas geringere mRNA-Gehalt der TRPC7-Kanäle. Wie in der Einleitung
beschrieben, sind diese Kanäle im Zentralen Nervensystem von großer Bedeutung, sodass sie in
Knochentumorzellen durchaus von geringerer Bedeutung sein könnten. Zwar wurde der Kanal
durchweg analysiert, jedoch zeigte sich in keiner der folgenden Versuchsreihen eine deutliche
Veränderung der Transkriptmengen.
Die Spezifität der TRPC-Primer bzw. der entsprechenden RT-qPCR wurde analog zu den Mas- und
α-Tubulin-Primern ebenfalls im Agarosegel verifiziert. Wie in Abbildung 11 und Abbildung 12 zu
sehen ist, erfolgte ausschließlich die spezifische Amplifikation von Produkten der erwarteten Größe
für alle TRPC-Kanäle unter den verwendeten Bedingungen.

ERGEBNISSE
33
Abbildung 11 Agarosegel zur Validierung der Spezifität der humanen RT-qPCR für TRPC1, TRPC3
und TRPC4. Die Amplifikatgrößen entsprachen mit 177 bp, 492 bp und 126 bp der erwarteten Größe. Die cDNA-freien Kontrollen (H2O) zeigen keine Amplifikate.
Abbildung 12 Agarosegel zur Validierung der Spezifität der humanen RT-qPCR für TRPC5, TRPC6
und TRPC7. Die Amplifikatgrößen entsprechen mit 210 bp, 191 bp und 194 bp den erwarteten Größen. Die cDNA-freien Kontrollen (H2O) zeigen keine Amplifikate.
500bp 400bp
300bp
200bp 150bp 100bp
TRPC5
210 bp
TRPC6
191 bp
TRPC7
194 bp
TRPC1
177 bp
TRPC3
492 bp
TRPC4
126 bp
500bp
400bp
300bp
200bp 150bp 100bp

ERGEBNISSE
34
4.3 EINFLUSS VON IL1β UND AVE-0991 AUF DIE TRPC-KANAL MRNA-GEHALTE
Wie in der Einleitung beschrieben, bewirken sowohl IL1β als auch Ang-(1-7) antiproliferative
Effekte in Osteosarkomzelllinien. Im Gegensatz dazu stehen die TRPC-Kanäle, deren Aktivität eher
mit proliferativen Effekten assoziiert sind. Daher war es von Interesse, die Expression der TRPC-
Kanäle in den beiden Osteosarkomzelllinien MNNG/HOS und U-2 Os unter dem Einfluss der beiden
antiproliferativen Substanzen zu analysieren.
Wie bei der Analyse der Mas-Rezeptor mRNA (siehe 4.1) wurden die Zellen in 6-Well-
Gewebekulturschalen eingesät, nach 24 Stunden mit frischem Kulturmedium und den jeweiligen
Agenzien –IL1β und AVE-0991 – versetzt, um wiederum 24 Stunden später die Zellernte und RNA-
Präparation vorzunehmen.
In der Abbildung 13 ist der Vergleich der mRNA-Gehalte von TRPC1 unter der jeweiligen
Stimulation dargestellt.
Abbildung 13 Effekte von IL1β und AVE-0991 auf die TRPC1-mRNA-Gehalte in MNNG/HOS und
U-2 Os Osteosarkomzellen MNNG/HOS nKontrolle=14, nIL1β=15, nAVE-0991=15, nKombination=11
U-2 Os nKontrolle=10, nIL1β=11, nAVE-0991=9, nKombination=10
Bei den MNNG/HOS ist erkennbar, dass sich der Median kaum verändert. Die Mediane variieren im
Vergleich zur Kontrolle um maximal fünf Prozent. Es zeigt sich auch keine deutliche Veränderung
bei den U-2 Os-Zellen. Der Anstieg des Medianwertes unter Kombination beider Substanzen liegt
bei dem 1,3-Fachen der Kontrolle und erreicht keine Signifikanz (p=0,07). Die alleinige Zugabe der
Agenzien hat keinen signifikanten Effekt auf den Median, wobei eine marginale Steigerung um das
1,1-Fache, bei AVE-0991 zu beobachten war (p=0,15).

ERGEBNISSE
35
Anders sieht es bei den Untersuchungen zu TRPC3 aus. In Abbildung 14 sind die Veränderungen
der mRNA-Mengen bei MNNG/HOS- und U-2 Os-Zellen unter dem Einfluss von IL-1β bzw. AVE0991
analog zu Abbildung 13 dargestellt.
Abbildung 14 Effekte von IL1β und AVE-0991 auf die TRPC3-mRNA-Gehalte in MNNG/HOS und
U-2 Os Osteosarkomzellen * p<0,05
MNNG/HOS nKontrolle=13, nIL1β=15, nAVE-0991=15, nKombination=11 U-2 Os nKontrolle=10, nIL1β=11, nAVE-0991=9, nKombination=10
Der Median der Werte ändert sich bei der Stimulation der MNNG/HOS-Zellen mit IL1β oder
AVE-0991 nur sehr marginal. Allerdings führt die Kombination beider Stoffe zu einem signifikanten
Abfall der mRNA Transkriptmengen auf 80 % der Kontrolle (p=0,02).
Auch bei den U-2 Os-Zellen ist unter Zugabe von IL1β zum Kulturmedium keine signifikante
Veränderung der mRNA-Mengen nachzuweisen. AVE-0991 hingegen führt zu einer signifikanten
Steigerung um 20 % im Vergleich zur Kontrolle (p=0,02) und im Zusammenspiel der beiden
Substanzen fällt der Median um 10 % (analog zu den MNNG/HOS), jedoch ist dies nicht signifikant.

ERGEBNISSE
36
TRPC4, als dritter Vertreter der humanen TRPC-Familie, zeigt unter Stimulation mit IL1β und/oder
AVE-0991 keine signifikanten Veränderungen im Vergleich zu den basalen mRNA-Gehalten. Die
Abbildung 15 stellt die Ergebnisse analog zu den vorherigen Abbildungen dar.
Abbildung 15 Effekte von IL1β und AVE-0991 auf die TRPC4-mRNA-Gehalte in MNNG/HOS und
U-2 Os Osteosarkomzellen MNNG/HOS nKontrolle=6, nIL1β=4, nAVE-0991=6, nKombination=7 U-2 Os-Zellen nKontrolle=5, nIL1β=5, nAVE-0991=5, nKombination=6
Es zeigt sich insbesondere bei den U-2 Os-Zellen eine größere Variabilität der TRPC4-mRNA-
Gehalte in allen drei Behandlungsgruppen im Vergleich zur Kontrolle. Ein solcher Effekt ist bei den
MNNG/HOS-Zellen lediglich unter Einfluss von IL-1β zu erkennen. Hier scheinen die mRNA-Gehalte
von TRPC4 zuzunehmen, jedoch ist der Median nahezu unverändert.

ERGEBNISSE
37
Ein ähnliches Bild ergibt sich für die Expression von TRPC5 in den untersuchten Osteosarkom-
Zelllinien. Die Abbildung 16 zeigt für beide Zelllinien die Veränderung der TRPC5-mRNA-Gehalte
unter Stimulation mit IL1β, AVE-0991 bzw. nach Administration beider Substanzen zusammen.
Abbildung 16 Effekte von IL1β und AVE-0991 auf die TRPC5-mRNA-Gehalte in MNNG/HOS und U-2 Os Osteosarkomzellen
MNNG/HOS nKontrolle=14, nIL1β=15, nAVE-0991=15, nKombination=11 U-2 Os nKontrolle=14, nIL1β=14, nAVE-0991=14, nKombination=14
In dem Diagramm der MNNG/HOS fallen die vielen als Punkte gekennzeichneten Outlier auf. Diese
verdeutlichen die Abweichung der Werte innerhalb einer Gruppe und sind Ausdruck einer hohen
Variabilität der Einzelwerte, insbesondere nach Administration der Substanzen einzeln oder in
Kombination. Im Gesamtbild ergibt sich dann für die TRPC5-Transkriptmengen keine signifikante
Veränderung bei den MNNG/HOS, obwohl die Mediane der drei Stimulationsgruppen um das 1,4-
Fache (IL1β oder AVE-0991) und das 1,3-Fache (beide in Kombination) ansteigen.
Bei den U-2 Os-Zellen führen weder IL1β noch AVE-0991 zu einer Veränderung des Medianwertes
um mehr als 5 %. Wenn beide Agenzien zusammen eingesetzt werden, sinkt zwar der Median auf
knapp 60 % des Kontrollwertes, jedoch ist auch dies aufgrund der deutlichen Streuung keine
signifikante Veränderung.
Schließlich sind in der Abbildung 17 die Effekte von IL1β und AVE-0991 auf die TRPC6-mRNA-
Gehalte von MMNG/HOS und U-2 Os-Zellen gezeigt.

ERGEBNISSE
38
Abbildung 17 Effekte von IL1β und AVE-0991 auf die TRPC6-mRNA-Gehalte in MNNG/HOS und
U-2 Os Osteosarkomzellen ** p<0,01; * p<0,05 MNNG/HOS nKontrolle=12, nIL1β=12, nAVE-0991=12, nKombination=9 U-2 Os nKontrolle=5, nIL1β=7, nAVE-0991=8, nKombination=8
Bei den MNNG/HOS-Zellen gibt es trotz deutlicher Unterschiede des Medianwertes der Kontrolle
zu den Medianen der Behandlungsgruppen keine signifikanten Veränderungen der TRPC6-mRNA-
Gehalte. Unter Stimulation mit IL1β steigt der Median auf das 1,5-Fache und der Median für
AVE-0991 sogar auf das 1,7-Fache der Kontrolle. Auch die Steigerung des Medianwertes durch
Kombination von IL1β und AVE-0991 um 30 % ist nicht signifikant.
Anders ist die Situation bei den U-2 Os-Zellen. Unter dem Einfluss von IL1β – mit oder ohne
AVE-0991 – ist eine signifikante Verringerung des Medianwertes der TRPC6-mRNA-Gehalte um
circa 40 % im Vergleich zum Kontrollwert (p=0,005; p=0,03) zu beobachten. Auch die
Administration von AVE-0991 allein führt zu einem Abfall des Medianwertes um 15 %, allerdings
ohne Signifikanz.
Zusammenfassend ist deutlich erkennbar, dass sich die Zelllinien MNNG/HOS und U-2 Os
hinsichtlich der Transkriptmengen der untersuchten TRPC Kanäle nach Zugabe von IL1β oder
AVE-0991 oder beidem wenig gleichen. Die in Punkt 5.2 angesprochene Ähnlichkeit der Zelllinien
in der basalen Expressionsstärke ist mitunter deutlichen Unterschieden gewichen. Bei den
MNNG/HOS kommt es beispielsweise unter Einfluss von IL1β und AVE-0991 zu einem signifikanten
Abfall der mRNA-Menge von TRPC3. Bei den U-2 Os-Zellen hingegen führt nur AVE-0991 zu einer
signifikanten Veränderung – und das in die entgegengesetzte Richtung. Weiterhin zeigt sich der
signifikante inhibitorische Effekt von IL1β auf die TRPC6-mRNA-Mengen nur bei den U-2 Os-Zellen.

ERGEBNISSE
39
4.4 DER EINFLUSS VON D-ALA AUF MAS- UND TRPC- MRNA-GEHALTE
In einer zweiten Versuchsreihe sollte der Effekt von D-Alanin7-Ang-(1-7) (D-Ala), einem Derivat
von Ang-(1-7) mit antagonistischer Wirkung am Mas-Rezeptor, auf die Transkriptmengen der
TRPC-Kanäle untersucht werden.
Erneut wurden dafür Zellen in die 6-Well-Gewebekulturschalen eingesät, nach 24 Stunden mit
frischem Kulturmedium und D-Ala versehen, um drei oder sechs Stunden später zum Zwecke der
RNA-Präparation geerntet zu werden.
In den Abbildungen 18 bis 22 sind die Ergebnisse aus der Analyse der mRNA-Gehalte der TRPC-
Kanäle 1 bis 7 unter Einfluss von D-Ala im Vergleich zur jeweiligen Kontrolle (ohne D-Ala)
dargestellt. Die Analyse der TRPC1-mRNA-Gehalte ergab für beide untersuchten Zelllinien keinen
Effekt nach Administration von D-Ala (Abbildung 18).
Abbildung 18 Effekte von D-Ala auf die TRPC1-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen MNNG/HOS nKontrolle=9, nD-Ala=8
U-2 Os nKontrolle=8, nD-Ala=9
Der Median der D-Ala Gruppe fällt bei den MNNG/HOS marginal um circa 8 % ab und steigt bei den
U-2 Os-Zellen um knapp 7 %. Beide Veränderungen sind statistisch gesehen nur zufällig.
ERGEBNISSE
40
Abbildung 19 Effekte D-Ala auf die TRPC3-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen MNNG/HOS nKontrolle=9, nD-Ala=8
U-2 Os nKontrolle=8, nD-Ala=9
In Analogie zu TRPC1, unterliegen auch die TRPC3-mRNA-Mengen in den MNNG/HOS-Zellen
keinen Veränderungen unter Einfluss von D-Ala. Ein leichter Anstieg der medianen TRPC3-mRNA-
Gehalte in den U-2 Os-Zellen unter D-Ala erreicht ebenfalls keine statistische Signifikanz.
Abbildung 20 Effekte D-Ala auf die TRPC4-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen MNNG/HOs nKontrolle=9, nD-Ala=8
U-2 Os nKontrolle=8, nD-Ala=9
Gleichermaßen zeigten sich auch die TRPC4-mRNA-Mengen durch die Administration von D-Ala
unbeeinflusst (Abbildung 20).

ERGEBNISSE
41
Abbildung 21 Effekte D-Ala auf die TRPC5-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen * p<0,05 MNNG/HOS nKontrolle=9, nD-Ala=8
U-2 Os nKontrolle=8, nD-Ala=8
Die Veränderung der TRPC5-mRNA-Menge unter Einfluss von D-Ala bei den MNNG/HOS ist in
Abbildung 21 visualisiert. Es zeigt sich ein signifikanter Abfall der Transkriptmengen auf das
0,7-Fache des Kontrollwertes (p=0,05). Bei den U-2 Os-Zellen steigt der Median der D-Ala Gruppe
um 20 % an. Jedoch ist schon anhand der Geometrie des Box-Plots zu sehen, dass die Einzelwerte
in der Gruppe stark schwanken und dadurch keine klare Tendenz erkennbar ist.

ERGEBNISSE
42
Abbildung 22 Effekte D-Ala auf die TRPC6-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen MNNG/HOS nKontrolle=9, nD-Ala=7
U-2 Os nKontrolle=8, nD-Ala=6
Wie aus Abbildung 22 ersichtlich ist, bewirkt die Administration von D-Ala keine eindeutige
Veränderung der TRPC6-Transkriptmenge in MNNG/HOS-Zellen. Der Median steigt zwar um
knapp 25 %, jedoch unterliegen die anderen Einzelwerte einer zu großen Schwankungsbreite, um
eine statistisch gesicherte Aussage zu erhalten. Anders ist es bei den U-2 Os-Zellen, wo unter
Einfluss von D-Ala der Median der TRPC6-mRNA-Gehalte auf 80 % des Kontrollwertes
abfällt (p=0,02).
TRPC6 ist der Kanal, dessen mRNA-Mengen unter Einfluss von IL1β in den U-2 Os-Zellen signifikant
vermindert wird. Außerdem zeigt sich unter IL1β-Administration ein Anstieg des mRNA-Gehaltes
des Mas-Rezeptors. Das bedeutet, dass unter gleichen in vitro-Bedingungen die Transkriptmengen
des antiproliferativen Mas-Rezeptors ansteigen, während die mRNA-Menge des TRPC6-Kanals
verringert wird.
Interessanterweise zeigt die D-Ala Versuchsreihe ein ähnliches Phänomen, indem hier die Zugabe
von D-Ala bei U-2 Os-Zellen zu einer signifikanten Verringerung der TRPC6-Transkriptmenge führt,
während es gleichzeitig zu einer signifikanten Erhöhung der Mas-mRNA-Menge kommt (p=0,02).
Dies ist in Abbildung 23 dargestellt. Sowohl bei den MNNG/HOS als auch bei den U-2 Os-Zellen
steigen die mRNA-Gehalte des Mas-Rezeptors nach Administration von D-Ala signifikant an. Bei
erstgenannten Zellen steigt der Median auf den 3,3-fachen Wert (p=0,05), bei Letzteren auf das
Dreifache des Kontrollwertes (p=0,02).

ERGEBNISSE
43
Abbildung 23 Effekte D-Ala auf die Mas-mRNA-Gehalte in MNNG/HOS und U-2 Os
Osteosarkomzellen * p<0,05 MNNG/HOS nKontrolle=9, nD-Ala=8
U-2 Os nKontrolle=6, nD-Ala=8

ERGEBNISSE
44
4.5 DER EINFLUSS VON LY294002 UND PD98059 AUF DIE MAS- UND TRPC- MRNA-GEHALTE
Die intrazelluläre Signaltransduktion des Mas-Rezeptors wird unter anderem über die Enzyme
Phosphatidylinositol-3-Kinase (PI3-Kinase) und Extrazellulär Signal Regulierte Kinase 1/2
(ERK1/2) vermittelt. Die Untersuchungsergebnisse verschiedener Arbeitsgruppen bezüglich
Aktivierung oder Inaktivierung der Kinasen gehen aktuell noch auseinander. Karnik et al. gehen in
einer aktuellen Übersichtsarbeit von einer Ang-(1-7)/Mas-abhängigen Aktivierung von
beispielweise der PI3-Kinase aus, jedoch berichten andere Arbeitsgruppen von einer Inaktivierung
der oben genannten Kinasen durch Ang-(1-7). [60;61;139]
Pharmakologische Inhibitoren für die PI3-Kinase (Ly294002) bzw. den ERK1/2-Signalweg
(PD98059) stehen zur Verfügung. Die Anwendung dieser Inhibitoren wurde in der vorliegenden
Arbeit vorgenommen, um zu untersuchen, inwieweit die Auswirkungen von Mas-Rezeptor-
Agonisten (AVE0991) oder Antagonisten (D-Ala) auf die TRPC-Expression mit der Aktivität dieser
Signalwege in Osteosarkomzellen assoziiert sind bzw. die Expression von Mas selbst beeinflussen.
Für die Versuche wurden analog zu den oben beschriebenen Versuchsreihen beide Zelllinien in
jeweils 6-Well-Gewebekulturschalen eingesät, nach 24 Stunden mit frischem Kulturmedium sowie
den Substanzen versorgt und 3 beziehungsweise 6 Stunden später geerntet. Die Ergebnisse der
RT-qPCR Untersuchung der TRPC-Kanäle und des Mas-Rezeptors sind in den Abbildungen 24 bis
29 dargestellt.
Wie aus der Abbildung 24 ersichtlich ist, bewirkt weder die Administration von Ly294002 noch
die von PD98059 Veränderungen der Transkriptmengen von TRPC1 in den Osteosarkom-Zelllinien
MNNG/HOS und U-2 Os.

ERGEBNISSE
45
Abbildung 24 Effekte von Ly294002 und PD98059 auf die TRPC1-mRNA-Gehalte in MNNG/HOS
und U-2 Os Osteosarkomzellen MNNG/HOS nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
U-2 Os nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
Bei den MNNG/HOS ist zwar ein leichter Anstieg des Medianwertes in der Ly294002-Gruppe auf
das 1,4-Fache des Kontrollwertes zu sehen, jedoch erreicht dieser Unterschied keine statistische
Signifikanz. Auch der Median für die TRPC1-Werte unter PD98059 Gabe ändert sich nicht im
Vergleich zur Kontrolle.

ERGEBNISSE
46
Die Diagramme in Abbildung 25 illustrieren die Ergebnisse der Analyse der TRPC3-mRNA-Mengen
in MNNG/HOS und U-2 Os-Zellen. In der Übersicht sind leichte Veränderungen zu erkennen.
Abbildung 25 Effekte von Ly294002 und PD98059 auf die TRPC3-mRNA-Gehalte in MNNG/HOS und U-2 Os Osteosarkomzellen
* p<0,05 MNNG/HOS nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
U-2 Os nKontrolle=9, nLy 100 µM =9, nPD 50 µM=8, nKombination=9
Die Transkriptmengen des TRPC3-Kanals zeigen sich in den MNNG/HOS-Zellen durch Ly294002
unbeeinflusst. In diesen Zellen bewirkt PD98059 einen tendenziellen Anstieg der TRPC3-mRNA-
Mengen (p=0,08). Dieser Effekt von PD98059 ist in U-2 Os-Zellen deutlicher ausgeprägt und
erreicht auch statistische Signifikanz (1,2-fache TRPC3-mRNA-Menge im Vergleich zur Kontrolle;
p=0,02). Bei den MNNG/HOS und U-2 Os-Zellen führt die Gabe von Ly294002 allein oder die
Kombination von Ly294002 mit PD98059 zu keiner eindeutigen Veränderung der TRPC3-mRNA-
Gehalte. Auch bei der in Abbildung 26 dargestellten Analyse des TRPC4-Kanals sind Veränderungen
der TRPC4-mRNA-Gehalte in der PD98059-Gruppe auffallend.

ERGEBNISSE
47
Abbildung 26 Effekte von Ly294002 und PD98059 auf die TRPC4-mRNA-Gehalte in MNNG/HOS
und U-2 Os Osteosarkomzellen ** p<0,01
MNNG/HOS nKontrolle=7, nLy 100 µM =7, nPD 50 µM=9, nKombination=6
U-2 Os nKontrolle=8, nLy 100 µM =8, nPD 50 µM=8, nKombination=9
Die MNNG/HOS zeigen unter dem Einfluss des MAP-Kinase-Inhibitors PD98059 eine signifikante
Erhöhung der TRPC4-mRNA-Gehalte auf das 2,1-Fache (p=0,008). Ein solcher Effekt deutet sich
auch bei den U-2 Os-Zellen an, erreicht aber keine Signifikanz (Abbildung 26). Die Administration
von Ly294002 allein oder in Kombination mit PD98059 bewirkt hingegen keine Veränderungen
der TRPC4-mRNA Gehalte in den beiden Osteosarkom-Zelllinien.
Ein anderes Bild ergibt sich bezüglich der transkriptionellen Regulation von TRPC5 in
Osteosarkom-Zellen. Wie aus Abbildung 27 ersichtlich ist, bewirkt die Administration des PI3-
Kinase Inhibitors Ly294002 sowohl in den MNNG/HOS als auch in den U-2 Os-Zellen einen
deutlichen Anstieg der TRPC5-mRNA-Gehalte.

ERGEBNISSE
48
Abbildung 27 Effekte von Ly294002 und PD98059 auf die TRPC5-mRNA-Gehalte in MNNG/HOS
und U-2 Os Osteosarkomzellen *** p<0,001
MNNG/HOS nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
U-2 Os nKontrolle=9, nLy 100 µM =9, nPD 50 µM=8, nKombination=9
Die Zugabe von Ly294002 hat einen Anstieg des Medianwertes in Relation zu den jeweiligen
Kontrollwerten auf das 9,6-Fache bei den MNNG/HOS und auf 5,9-Fache bei den U-2 Os Zellen zur
Folge (p=0,0004; p=0,0008). Die Administration von PD98059 führt in beiden Zelllinien zu einem
marginalen Anstieg der TRPC5-mRNA-Mengen um weniger als 10 %, ohne jegliche Signifikanz.
Deswegen liegt die Annahme nahe, dass auch die deutliche Zunahme der Transkriptmengen unter
Verwendung beider Substanzen eher in der inhibierenden Wirkung von Ly294002 auf die PI3-
Kinase begründet ist. Sowohl der Anstieg des Medianwertes nach Gabe von Ly294002 und
PD98059 auf das 8,9-Fache bei den MNNG/HOS (p=0,0004), als auch der Anstieg auf das 5,2-Fache
der Kontrollwerte bei den U-2 Os-Zellen (p=0,0004) sind statistisch signifikant.
Die in der Abbildung 28 dargestellten Effekte der verwendeten Inhibitoren auf die TRPC6-mRNA-
Mengen zeigen interessante Ergebnisse.

ERGEBNISSE
49
Abbildung 28 Effekte von Ly294002 und PD98059 auf die TRPC6-mRNA-Gehalte in MNNG/HOS
und U-2 Os Osteosarkomzellen ** p<0,01; * p<0,05
MNNG/HOS nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
U-2 Os nKontrolle=6, nLy 100 µM =6, nPD 50 µM=5, nKombination=6
Bei den MNNG/HOS führen Ly294002 und PD98059 unabhängig voneinander zu einem
signifikanten Anstieg der Mediane auf das Doppelte des Kontrollwertes (p=0,004; p=0,005).
Interessanterweise führt die Kombination beider Stoffe nicht etwa zu einem additiven oder gar
superadditiven Effekt. Der Median in der jeweils letzten Gruppe (kombinierte Gabe beider
Inhibitoren) steigt „nur“ auf das 1,9-Fache des Kontrollwertes (p=0,02) und liegt damit jeweils
unter dem Niveau der Einzelgaben der Stoffe.
Auch die U-2 Os-Zellen zeigen signifikante Veränderungen ihrer TRPC6-mRNA-Gehalte, wenn sie
mit Ly294002 und PD98059 behandelt werden. Der Median in der Ly294002-Gruppe steigt auf das
Zweieinhalbfache des Kontrollwertes (p=0,005). Der augenscheinliche Anstieg der TRPC6-mRNA-
Mengen nach Gabe von PD98059 erreicht aber lediglich tendenziellen Charakter (p=0,08).
Dahingegen ist der Median der Versuchsgruppe mit dem gleichzeitigen Einsatz der Substanzen
dreimal so groß wie der Median der Kontrollgruppe und beschreibt damit einen statistisch
gesicherten Unterschied (p=0,008). Es lässt sich zusammenfassen, dass Ly294002 und PD98059
offensichtlich unabhängig voneinander die Transkriptmengen von TRPC6 beeinflussen.
In den vorherigen Versuchsreihen zum Einfluss von IL1β und AVE-0991 sowie von D-Ala ist die
gegenläufige Veränderung der mRNA-Mengen des Mas-Rezeptors und des TRPC6 Kanals (bei den
U-2 Os-Zellen) aufgefallen. In dieser Versuchsreihe ist das Ergebnis ein anderes. Die Verwendung
von Ly294002 und PD98059 erhöht nicht nur die Transkriptmengen von TRPC6 (siehe
Abbildung 28), sondern auch – wie in Abbildung 29 erkennbar – die mRNA-Gehalte des Mas-
Rezeptors.
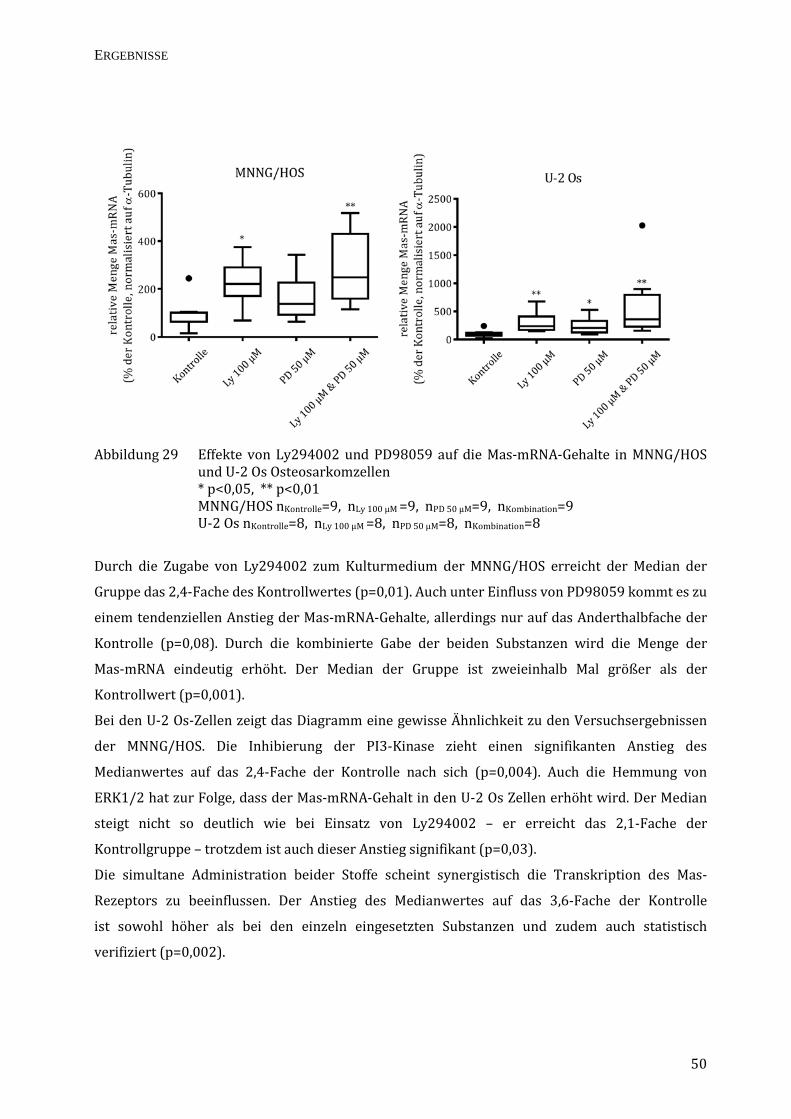
ERGEBNISSE
50
Abbildung 29 Effekte von Ly294002 und PD98059 auf die Mas-mRNA-Gehalte in MNNG/HOS
und U-2 Os Osteosarkomzellen * p<0,05, ** p<0,01
MNNG/HOS nKontrolle=9, nLy 100 µM =9, nPD 50 µM=9, nKombination=9
U-2 Os nKontrolle=8, nLy 100 µM =8, nPD 50 µM=8, nKombination=8
Durch die Zugabe von Ly294002 zum Kulturmedium der MNNG/HOS erreicht der Median der
Gruppe das 2,4-Fache des Kontrollwertes (p=0,01). Auch unter Einfluss von PD98059 kommt es zu
einem tendenziellen Anstieg der Mas-mRNA-Gehalte, allerdings nur auf das Anderthalbfache der
Kontrolle (p=0,08). Durch die kombinierte Gabe der beiden Substanzen wird die Menge der
Mas-mRNA eindeutig erhöht. Der Median der Gruppe ist zweieinhalb Mal größer als der
Kontrollwert (p=0,001).
Bei den U-2 Os-Zellen zeigt das Diagramm eine gewisse Ähnlichkeit zu den Versuchsergebnissen
der MNNG/HOS. Die Inhibierung der PI3-Kinase zieht einen signifikanten Anstieg des
Medianwertes auf das 2,4-Fache der Kontrolle nach sich (p=0,004). Auch die Hemmung von
ERK1/2 hat zur Folge, dass der Mas-mRNA-Gehalt in den U-2 Os Zellen erhöht wird. Der Median
steigt nicht so deutlich wie bei Einsatz von Ly294002 – er erreicht das 2,1-Fache der
Kontrollgruppe – trotzdem ist auch dieser Anstieg signifikant (p=0,03).
Die simultane Administration beider Stoffe scheint synergistisch die Transkription des Mas-
Rezeptors zu beeinflussen. Der Anstieg des Medianwertes auf das 3,6-Fache der Kontrolle
ist sowohl höher als bei den einzeln eingesetzten Substanzen und zudem auch statistisch
verifiziert (p=0,002).

51
5 DISKUSSION
In der vorliegenden Arbeit erfolgte eine umfassende Beschreibung der TRPC-Expression in
Osteosarkomzellen unter basalen Bedingungen. Es wurden die Einflüsse der Modulation der
alternativen Ang-(1-7)/Mas Achse des RAS (mittels Ang-(1-7) und AVE0991) bzw. der Einfluss der
Immunstimulation (IL-1β) auf die Expression von TRPC-Kanäle systematisch untersucht.
Mittels Administration von Interleukin 1β wurden die immunmodulatorische Wirkung und der
klinisch nachgewiesene Effekt von Mifamurtid nachgebildet, die der Therapie des Osteosarkoms
zugrunde liegen. [140] Spezielles Augenmerk wurde dabei auf die alternative RAS-Achse mit den
Protagonisten Ang-(1-7) und dem Rezeptor Mas gelegt – besonders die mögliche Assoziation
mit Expression/Aktivität von TRPC Kanälen und deren mitunter proliferative
Wirkungen. [119;123;129]
Dass ein Zusammenhang von Osteosarkomen und dem RAS besteht, legt die Arbeit von Wasa et al.
nahe, in welcher die antiproliferative Wirkung des Ang-II Rezeptorantagonisten Candesartan auf
Osteosarkome im Mausmodell gezeigt wurde. [141] In der Arbeit von Souza et al. konnte
nachgewiesen werden, dass die Expression des Mas-Rezeptors in Makrophagen nach Gabe von
Lipopolysacchariden (einem natürlichem Bestandteil der Bakterienwand und damit Analogon zum
Mifamurtid) gesteigert wird. [142]In unserer Voruntersuchung wurde ebenfalls die Induktion der
Mas-Rezeptorexpression durch Interleukin 1β gezeigt. [33]
Zunächst wurde eine Analyse der basalen mRNA-Gehalte der TRPCs in Osteosarkom-Zelllinien
vorgenommen und eventuelle Veränderungen nach Modulation der Ang-(1-7)/Mas-Achse
bzw. der intrazellulären Signaltransduktion bestimmt. Wie bereits einleitend beschrieben, weisen
die aktuellen Forschungen den TRPC-Kanälen eine wichtige Rolle bei Tumorentstehung und -
wachstum zu. Sie sind Mediatoren einer veränderten Calcium-Homöostase in den Tumorzellen und
können für proliferative Prozesse von großer Bedeutung sein. Diese Prozesse wurden von Hanahan
und Weinberg als „Hallmarks of Cancer“ bezeichnet – Merkmale, die für den malignen Phänotyp
eines Tumors stehen, wie zum Beispiel autonomes Wachstum durch eigene Wachstumssignale,
Angiogenese, Metastasierung und Vermeidung von Apoptose. [143;144]
Allerdings ist es wichtig festzuhalten, dass Calcium-Signaling keineswegs nur für proliferative
Zellvorgänge steht. In Podozyten führt beispielsweise ein Calcium-Einstrom über TRPC6 zu
proapoptotischen Signalen und genauso kann ein Calcium-Einstrom über TRPC1 zur Apoptose von
glatten Muskelzellen führen. [145]
In der Mehrzahl der Publikationen ist TRPC1 jedoch als Schlüsselprotein einer veränderten
Calcium-Homöostase und damit wichtiger Akteur in verschiedenen Tumorzellgeweben
beschrieben. Tajeddine et al. vermuten, dass die Expression des TRPC1-Kanals einer
autoregulativen Schleife im Sinne eines positiven Feedback-Mechanismus unterliegen könnte.

DISKUSSION
52
TRPC1 stimuliert dabei den Wachstumshormonrezeptor Epidermal Growth Factor Receptor
(EGFR), welcher wiederum über IP3 die Freisetzung von Calcium aus dem ER fördert, was (im
Rahmen des SOCE) zu Calcium-Einstrom über TRPC1 führt. Da der EGF-Rezeptor auch die PI3-
Kinase und ERK aktiviert, könnte dieser sich selbst unterhaltende Kreislauf als Korrelat für das
unkontrollierte Wachstum von Tumorzellen stehen. [146;147;148]
In der vorliegenden Arbeit konnte weder durch Aktivierung oder Inhibierung des Mas-Rezeptors
noch durch Inhibierung der Aktivität der Proteinkinasen PI3-Kinase beziehungsweise ERK1/2 ein
Einfluss auf die Expression von TRPC1 festgestellt werden. Schlussfolgernd lässt sich feststellen,
dass der oben genannte Effekt von Mas und Interleukin1β nicht über eine Veränderung der mRNA-
Mengen des TRPC1 Kanals vermittelt wird.
Im Gegensatz zu seinem Pendent TRPC1 ist die Rolle von TRPC3 laut Literatur nicht nur als
tumorigen einzuordnen. So korreliert beispielsweise die Expression von TRPC3 in
Bronchialkarzinomen mit einer insgesamt verbesserten 5-Jahres-Überlebensrate sowie einem
längeren krankheitsfreien Überleben. [149] Bei einigen gynäkologischen Tumoren hingegen
scheint eine erhöhte Expression von TRPC3 eher proliferativ zu wirken. In Mammakarzinomzellen
wurde beispielsweise durch die Gabe von Arachidonsäure die Inhibition von TRPC3 und damit die
Reduktion von in vitro Zellmigration erreicht. [150] In Ovarial-Karzinomen fördern TRPC3-Kanäle
ebenfalls das Tumorwachstum und nach deren Herabregulation zeigte sich eine signifikant
verminderte Proliferation der ovarialen Tumorzellen SKOV3. [151]
Eine mögliche Ursache für die offenbar diskrepanten pro-onkogenen bzw. anti-onkogenen
Wirkungen ist zum Teil über die unterschiedliche Aktivierung von TRPC3 im Vergleich zu TRPC1
erklärlich. Wie einleitend beschrieben, ist TRPC1 wichtiger Bestandteil des store-operated-
Calcium-Einstroms (SOCE) und im Gegensatz dazu wird TRPC3 eher store-independent aktiviert
und ist damit von einer Ligandenaktivierung abhängig. [152] Aufgrund des spezifischeren
Mechanismus der Aktivierung kann auch die Funktion hinsichtlich der Beeinflussung von
Proliferation bzw. Antiproliferation differieren. So kann man zumindest die Vermutung aufstellen,
dass die unterschiedlichen molekularen Sequenzen der Tumorigenese der jeweiligen
Tumorentitäten einen unterschiedlichen Effekt der TRPC3-Bindung nach sich ziehen.
In der vorliegenden Arbeit zeigte sich ein positiver Effekt des Mas-Agonisten AVE-0991 auf die
Transkriptmengen des TRPC3-Kanals bei U-2 Os-Zellen. Es zeigte sich hingegen keine signifikante
Veränderung bei den MNNG/HOS-Zellen und keine Veränderung der mRNA-Mengen nach
Interleukin 1β-Gabe, sodass wahrscheinlich auch TRPC3 nicht als Vermittler der antiproliferativen
Effekte von Interleukin 1β und Mas fungiert. Allerdings ist ein interessanter Aspekt, dass sowohl
AVE-0991 als auch der ERK-Inhibitor PD98059 eine signifikante Steigerung der TRPC3-mRNA-
Mengen bewirken. Dies steht im Gegensatz zu den Ergebnissen von Yang et al., bei denen die
Inhibierung des TRPC3-Expression in den ovarialen Tumorzellen SKOV3 zu einer Reduktion der

DISKUSSION
53
Proliferation via Suppression des EGF-Rezeptor-induzierten Calcium-Einstroms mit konsekutiver
Dephosphorylierung von Cdc2 und CaMKIIα führt. [153] Vorstellbar wäre ein Mechanismus, bei
dem die Gabe von AVE-0991 direkt oder indirekt die Inhibierung der ERK zur Folge hat und
daraufhin möglicherweise autoregulativ die TRPC3 Expression erhöht wird, um im Sinne eines
Escape-Mechanismus das Tumorwachstum aufrecht zu erhalten.
Die Transkription von TRPC4 zeigte sich fast unbeeinflusst unter den hier vorgenommenen
Zellkulturbedingungen. Einzig durch die Gabe von PD98059 wurde bei den MNNG/HOS die TRPC4-
mRNA-Menge erhöht. Da zur pathophysiologischen Bedeutung von TRPC4 weitaus weniger
Forschungsdaten existieren als zu den anderen Mitgliedern der TRPC-Familie, fällt es schwer, diese
Veränderung einzuordnen. Carson et al. untersuchten die Wirkung von Englerin A (ein Wirkstoff
aus der Katzenminze) auf das Wachstum von renalen Tumorzellen. Dabei konnten sie zeigen, dass
Englerin A über die Aktivierung von TRPC4 Kanälen das Tumorwachstum reduziert. Durch den
Calcium-Einstrom wurden in den renalen Tumorzellen -498 Nekrose- und Apoptoseneigung unter
Inhibierung der ERK-Kinase beobachtet. [154] Analog dazu zeigte sich auch in den MNNG/HOS eine
Verbindung zwischen der Inhibierung von ERK (durch PD98059) und TRPC4. Jedoch sind Ursache
und Wirkung nicht in allen Konstellationen in Übereinstimmung zu bringen. So wird im oben
genannten Experiment ein extrazelluläres Signal in wachstumshemmende Effekte transduziert,
deren Bestandteil unter anderem die Inhibierung der ERK ist. Dahingegen unterbindet die Gabe des
ERK-Inhibitors PD98059 direkt die proliferative Funktion von ERK. Insgesamt scheint der
antiproliferative Effekt, den IL1β und Mas ausüben nicht über den TRPC4-Kanal zu wirken.
TRPC5 ist wie in der Einleitung erwähnt bis jetzt vorwiegend im Zusammenhang mit neuronalem
Wachstum beschrieben. [130;131] Dass eine Veränderung der TRPC5-Genexpression in
Zusammenhang mit Interleukin 1β beziehungsweise Mas steht, ist nach aktuellem Wissenstand
nicht zu erwarten. Diese Sicht wird durch die Ergebnisse der vorliegenden Arbeit – auf
Transkriptebene - weitgehend bestätigt. Einen signifikanten Einfluss auf die TRPC5-mRNA-Mengen
hatte jedoch die Gabe des Mas-Antagonisten (D-Ala) bei den MNNG/HOS. D-Ala führte zu einer
Verminderung der TRPC5-mRNA-Mengen. Deutlich eindrücklicher war jedoch die Auswirkung der
Inhibition der PI3-Kinase mittels Ly294002 auf die TRPC5-mRNA-Mengen. Es zeigte sich in beiden
Zelllinien ein fünf- bis zehnfacher Anstieg der mRNA-Transkriptmengen. Auch wenn dies sicherlich
nicht Erklärung für die in der Einleitung formulierten antiproliferativen Effekte ist (da AVE-0991
und/oder IL1β keinen Einfluss haben), so steht es doch im Zusammenhang mit den Ergebnissen
von Bezzerides et al. Eine Aktivierung von EGFR mit konsekutiver Aktivierung der PI3-Kinase führt
in kultivierten Hippocampus-Neuronen zu einer schnellen Translokation von TRPC5 Vesikeln mit
Insertion in die Plasmamembran. Dies wiederum supprimiert das Wachstum von
Neuritenfortsätzen (growth cones) und Axonen. Durch die Gabe von Ly294002 kann diese Insertion
von voll funktionsfähigen TRPC5 Kanälen in die Plasmamembran unterbunden werden. [155]

DISKUSSION
54
Nun zeigte sich in unseren Untersuchungen, dass durch Ly294002 die Transkription von TRPC5 um
ein Vielfaches gesteigert wird. Möglicherweise wird die Expression von TRPC5 durch einen
negativen Feedback-Mechanismus über die PI3-Kinase reguliert. Eine aktivierte PI3-Kinase könnte
zu einer Translokation von TRPC5-Kanälen in die Plasmamembran führen und solange die TRPC5
Kanäle in die Plasmamembran inkorporiert werden, stagniert die Expression derselben auf einem
basalen Niveau. Sollte allerdings die Aktivität der PI3-Kinase herabgesetzt werden und dadurch
weniger TRPC5-Kanäle die Plasmamembran erreichen, führen kompensatorische feedback-
Schleifen zu einer vermehrten Expression von TRPC5-Kanälen. Dass der Mechanismus der TRPC5
Translokation in die Plasmamembran nicht nur für neuronale Zellen zutreffend ist, konnte bereits
in HEK-Zellen (embryonale Nierenzellen) nachgewiesen werden. Daher ist es möglich, dass dieses
Prinzip der Aktivierung auch bei Osteosarkomzellen wirksam ist.
Ein in vielerlei Hinsicht interessanter Vertreter der TRPC-Familie ist TRPC6. Dieser Kanal ist
Gegenstand intensiver Forschung und seine pathophysiologische Bedeutung wird stetig mit neuen
Publikationen belegt bzw. erweitert.
Eine verstärkte Expression von TRPC6 wurde in Prostata-, Magen-, Leberkarzinom-Zellen und
Glioblastomzellen nachgewiesen. [156;157;158;159] Die pharmakologische Suppression durch
spezifische Antagonisten oder der genetische Knockout von TRPC6 führen zu einer Suppression der
Proliferation und zum Zellzyklusarrest in den Tumorzellen. Der Kanal wird dabei entweder über
second messenger aktiviert – wie zum Beispiel durch Diacylglycerol nach der Aktivierung des AT1-
Rezeptors [152;160] – oder durch Depletion der internen Calcium-Speicher aktiviert. [161] Folge
ist in beiden Fällen der Calcium-Einstrom in die Zelle mit Erhöhung der intrazellulären
Calciumkonzentration. In verschiedenen Arbeiten wurden die daraus resultierenden Effekte
aufgezeigt. Über die Aktivierung von Calcium-abhängigen Kinasen werden unter anderem die PI3-
Kinase oder die MAP-Kinase ERK1/2 phosphoryliert und damit ein proliferativer beziehungsweise
anti-apoptotischer Weg gefördert. [162;163] Die bereits mehrfach erwähnte Veränderung der
Calciumhomöostase – ein „Hallmark of Cancer“ – ist eine wichtige Grundlage der malignen
Transformation. [143] Folge sind unter anderem ein unkontrollierter Zellzyklus und eine
eingeschränkte Differenzierung. Aufgrund der Induktion der TRPC6 Expression in vielen
Tumorzellen und der deutlichen Effekte, die eine Suppression von TRPC6 verursacht, kommt
diesem Kanal eine besondere Bedeutung bei Entstehung und Progression des malignen Phänotyps
zu. In der Arbeit von van der Vlag et al. wurde die Existenz eines positiven Feedback-Mechanismus
für die Expression von TRPC6 Kanälen beschrieben. [164] Nach Aktivierung des TRPC6-Kanals
durch Diacylglycerol (oder mechanische Stimuli [160]) folgte ein Calcium-Einstrom der
nachfolgend Calmodulin aktivierte und anschließend zur Phosphorylierung von NFAT (=Nuclear
Factor of Activated T-Cells) führte. Die Translokation von NFAT in den Nucleus bewirkte eine

DISKUSSION
55
vermehrte Transkription von TRPC6. Diese Autoinduktion könnte eine Ursache für das maligne
Wachstum von bestimmten Tumorzellen sein.
In den von uns untersuchten Osteosarkomzellen sind Expression bzw. pathophysiologische
Relevanz von TRPC6-Kanälen noch nicht explizit analysiert worden. Durch die vorliegenden
Ergebnisse wird die Expression der TRPC6-Kanäle in Osteosarkomzellen bewiesen. Des Weiteren
zeigt sich eine Verminderung der TRPC6-Transkriptionsmengen unter Einfluss von Interleukin 1β
in den U-2 Os-Zellen. Dies könnte den antiproliferativen Effekt von Interleukin 1β durch
Reduzierung des TRPC6 getragenen Calciumeinstroms und daraufhin verminderten
calciumabhängigen proliferativen Signalen erklären. Nach Gabe des Mas-Agonisten AVE-0991
zeigte sich weder bei den U-2 Os- noch bei den MNNG/HOS-Zellen eine Veränderung der TRPC6
mRNA-Menge. Wenn die antiproliferativen Effekte der Ang-(1-7)/Mas-Achse über die Expression
eines onkogenen TRPC6 wirken würden, wäre ein Abfall der mRNA-Transkription zu erwarten
gewesen. Dies konnte jedoch nur bei der Gabe von D-Ala, einem Mas-Antagonisten beobachtet
werden. Eine Einordnung der Ergebnisse in die aktuellen Schemata zum Mas-Signaling fällt schwer.
Möglicherweise wird die Wirkung von D-Ala nicht über Mas, sondern einem weiteren Rezeptor
vermittelt. Ein möglicher Kandidat ist der Mas-related-GPCR Member D (MrgD), dessen Rolle für
die TRPC-Expression/Aktivität in weiterführenden Untersuchungen geklärt werden sollte. [165]
Alternativ kann auch eine pleiotrope Rezeptorkinetik von Mas (sogenanntes biased signaling
[166]) angenommen werden. Die Signalkaskaden, die durch Mas-Rezeptor-Liganden adressiert
werden sind – wie in Tirupula et al. beschrieben – komplex. [167] Demnach wird dem Mas-
Rezeptor eine konstitutive Aktivität zugeschrieben, die mehrere downstream-Effektoren wie
Phospholipase C, Adenylatcyclase und Raf umfasst. Die Bindung von Agonisten wie beispielsweise
Ang-(1-7) hat allerdings kaum Einfluss auf die konstitutive Aktivität, sondern führt zu einem
schnellen Calcium-Anstieg in den untersuchten Zellen. Andere synthetische nicht-peptidische Mas-
Agonisten bewirken ein davon abweichendes Aktivierungsmuster, sodass kein allgemein gültiges
Bild des Mas-Signalings existiert. Speziell die widersprüchlichen Arbeiten bezüglich der
Aktivierung bzw. Inhibierung der PI3-Kinase durch Mas lassen großen Spielraum für die
Einordnung der aktuell erhobenen Ergebnisse in das Mas-Signaling bezüglich der Wirkungen des
PI3-Kinase-Inhibitors Ly294002. Geht man davon aus, dass der Mas-Rezeptor wie in der Arbeit von
Ni et al. zu einer Inhibierung der PI3-Kinase führt [60], würde der nachgewiesene Anstieg der
mRNA-Mengen des Mas-Rezeptors nach Gabe von Ly294002 dem Anstieg nach Gabe von AVE-0991
ähneln und damit möglicherweise einer Form der Autostimulation entsprechen. Allerdings ist die
Aktivierung der PI3-Kinase durch Ang-(1-7)/Mas durch Arbeiten belegt [53;58;59], sodass die
Wirkung von Ly294002 auch der (partiellen) Wirkung von D-Ala entsprechen könnte, welches
ebenfalls zum Anstieg der mRNA-Mengen des Mas-Rezeptors führt.

DISKUSSION
56
Auch die Inhibierung der MAP-Kinase ERK1/2 durch PD98059 führt zu einer Zunahme des mRNA-
Gehaltes – analog zu AVE-0991, welches über Bindung an den Mas-Rezeptor ebenfalls zu einer
Hemmung der ERK1/2 führt. Betrachtet man allein die Veränderungen der Mas-mRNA-Mengen
sind diese in Einklang mit einem autostimulativen Feedback-Mechanismus. Da dies aber jede
posttranskriptionale Modulation beispielsweise auf Proteinebene oder Aktivierungsstatus
unbeachtet lässt, ist dies sehr spekulativ.
Im nächsten Teil der Arbeit wurde die Wirkung der Kinase-Inhibitoren Ly294002 und PD98059 auf
die mRNA-Mengen von TRPC6 untersucht. Wenn TRPC6 in den Osteosarkomzellen, analog zu den
oben genannten Karzinomzellen, einen promalignen Effekt hat, könnte nach Inhibierung einer
sogenannten „Anti-Apoptose-Kinase“ (PI3-Kinase) [162] und einer mitogenen Kinase (ERK1/2)
eigentlich eine Verminderung der TRPC6-Aktivität und konsekutiv möglichweise auch der
Expression zu erwarten sein. In beiden Zelllinien wird unter Inhibition der PI3-Kinase verstärkt
TRPC6-mRNA gebildet. Zusätzlich zeigt sich bei den MNNG/HOS eine signifikante Steigerung der
TRPC6 mRNA-Transkriptmengen nach Hemmung der ERK. Möglicherweise wird durch die Gabe
von Ly294002 und PD98059 die Fokussierung auf die von Nijenhuis et al. beschriebene
Selbstinduktion bewirkt. [168] Jedoch wäre dann ein additiver Effekt von Ly294002 und PD98059
zu erwarten, der sich nicht einstellte. Eine andere Erklärung ist die Steigerung der Expression im
Sinne eines Escape-Phänomens bei dem die Tumorzelle gegenregulative Mechanismen aktiviert,
um die antiproliferativen Effekte von Ly294002 und PD98059 zu kompensieren. Letztlich handelt
es sich auch nur um eine Annahme, dass TRPC6 auch in den Osteosarkomzellen onkogene Effekte
ausübt. Wie bereits einleitend erwähnt wurde, ist der Calcium-Einstrom zwar wichtig für eine
Tumorprogression, jedoch werden auch pro-apoptotische Signale durch Calcium vermittelt. [145]
Zhang et al. zeigten, dass Podozyten nach Gabe von Angiotensin II via Induktion von TRPC6 in die
Apoptose übergehen. [163] Demgegenüber stehen die Ergebnisse von Dryer et al., die in Maus-
Podozyten eine Aktivierung des Zellzyklus via STAT3 (= Signal Transducer and Activator of
Transcription 3) und NFAT durch AT1-Rezeptor abhängige TRPC6-Aktivierung nachweisen
konnten. [160] Diese proliferativen Effekte des AT1-Rezeptors in Verbindung mit TRPC6 konnte
indirekt durch Wasa et al. bestätigt werden. Dabei wurde ein tumorsuppressiver Effekt von
Sartanen (einem pharmakologischen AT1-Rezeptorblocker) auf Osteosarkome bei Mäusen
nachgewiesen. [141] Allerdings mit der Einschränkung, dass diese Effekte nur in vivo und nicht in
vitro zu beobachten sind. Dies ist ein Hinweis darauf, dass die antiproliferativen Effekte von
Ang-(1-7)/Mas (als Gegenspieler des klassischen RAS AT1-Rezeptors) möglicherweise nicht nur in
den Osteosarkomzellen selbst ausgeübt werden.
Dementsprechend sind die vorliegenden Ergebnisse wahrscheinlich nicht das laborchemische
Korrelat für die klinisch relevante Wirkung von Mifamurtid oder die publizierten Effekte der
Ang-(1-7)/Mas-Achse. Die – möglicherweise – proliferativen Effekte der TRPC6-Kanäle im

DISKUSSION
57
Osteosarkom sind möglicherweise ebenfalls nicht in den Osteosarkomzellen, sondern in dem
umgebenden Tumorgewebe zu finden. Die TRPC6-abhängig induzierte Angiogenese durch
Endothelzellen wurde bereits mehrfach beschrieben [169;149] und müsste gegebenenfalls im
Zusammenhang mit Interleukin 1β und Ang-(1-7)/Mas genauer untersucht werden. Außerdem
könnten sich die Effekte von Mas und Interleukin 1β auch in der VEGF-Expression zeigen. Dieser
Wachstumsfaktor der vaskulären Endothelzellen ist ein in vielerlei Hinsicht interessanter
Beteiligter bei dem Tumorwachstum von Osteosarkomzellen. Daft et al. thematisierten eine
autokrine Stimulation der Osteosarkomzellen durch VEGF. [169] Außerdem ist ein erhöhter VEGF
Spiegel im Blutplasma mit einer schlechteren Prognose beim Osteosarkom assoziiert, so treten
häufiger Rezidive oder Fernmetastasen auf. Gerade bei prognostisch schlechterem Verlauf sind die
Ansprechraten von Mifamurtid besser (was die Verbesserung von Lebensqualität und
Verlängerung der Lebensjahre betrifft). [170] Da VEGF nicht nur autokrin, sondern auch parakrin
wirkt, ist die Wirkung von VEGF auf die TRPC6-induzierte Angiogenese in Endothelzellen sehr
interessant. Ohne die tumortypische Angiogenese würden die Tumorzellen unter anderem in ihrer
Nährstoffversorgung beschränkt und damit weniger aggressiv wachsen können. Es bleibt weiterhin
herauszufinden, an welchem Punkt die tumorsuppressiven Effekte von Interleukin 1β und
Ang-(1-7)/Mas hauptsächlich entstehen. Die gezeigten Wirkungen von Interleukin 1β auf die
Genexpression von TRPC6 liefern zwar einen Anhalt für einen möglichen Effekt, entsprechen jedoch
wahrscheinlich nicht der hauptsächlichen Wirkung.

58
6 ZUSAMMENFASSUNG
Vordergründiges Ziel der Arbeit war eine Grundlagenforschung über die Expression der TRPC-
Kanäle in Osteosarkomzellen unter basalen und stimulierten Bedingungen durchzuführen. Als
thematisches Setting wurde zum einen die bisher unklare Wirkung des erfolgversprechenden
Medikaments Mifamurtid gewählt sowie zum anderen ein möglicher Zusammenhang mit der
alternativen Ang-(1-7)/Mas-Achse gesucht.
Die vorliegende Arbeit hat einerseits die Expression der TRPC-Kanäle in Osteosarkomzellen
nachgewiesen und andererseits interessante Schnittpunkte zu der alternativen
ACE2/Ang-(1-7)/Mas-Achse des RAS gezeigt. So wird in Zukunft möglicherweise ein
Zusammenhang zwischen der Mas-Aktivität und den TRPC-Kanälen – insbesondere von TRPC6 –
substantiviert werden können. Die massive Induktion der Transkription von TRPC5 unter
Inhibierung der PI3-Kinase ist eine interessante Beobachtung, die Ansätze für weiterführende
Untersuchungen eröffnet. Inwieweit die pharmakologische Modulation von Mas- (oder MrgD-)
Rezeptoren eine therapeutisch relevante Option beim Osteosarkom darstellt und inwieweit die
Modulation der TRPC-Kanäle dazu beitragen kann, ist mit dieser Arbeit nicht geklärt. Die
vorgelegten Daten zeigen jedoch eindeutig, dass der Mas-Antagonist D-Ala die Expression von
zumindest TRPC 5, 6 und 3 sowie von Mas selbst in Osteosarkom-Zellen beeinflusst; Effekte die
auch durch die Modulation von PI3K und MAP-Kinase Signalwegen erreicht werden können.

59
7 LITERATURVERZEICHNIS
1 Bieling P., Rehan N., Winkler P., Helmke K., Maas R., Fuchs N., Bielack S., Heise U., Jurgens H., Treuner J., Romanowski R., Exner U., Kotz R., Winkler K.. (1996) Tumor size and prognosis inaggressively treated osteosarcoma. J Clin Oncol 14: 848
2 Bielack S., Flege S., Kempf-Bielack B. (2000) Behandlungskonzept des Osteosarkoms. Onkologe 6: 747
3 Flege S., Bielack S. (2004) Ziele und Ergebnisse der COSS-Studien. Handchir Mikrochir Plast Chir 36: 282
4 Bielack S., Zoubek A., Kotz R. (2006) Osteosarkom in "Kompendium internistische Onkologie: Standards in Diagnostik und Therapie, Teil 2: Therapiekonzepte maligner Tumoren" Springer, Berlin, Heidelberg, New York, Barcelona, Hongkong, London, Mailand, Paris, Tokio
5 Burrow S., Andrulis I.L., Pollak M., Bell R.S. (1998) Expression of insulin-like growth factor receptor, IGF-1, and IGF-2 in primary and metastatic osteosarcoma. J Surg Oncol 69(1): 21-7.
6 MacEwen E.G., Pastor J., Kutzke J., Tsan R., Kurzman I.D., Thamm D.H., Wilson M., Radinsky R. (2004) IGF-1 receptor contributes to the malignant phenotype in human and canine osteosarcoma. J Cell Biochem 1;92(1): 77-91.
7 Jentzsch T., Robl B., Husmann M., Bode-Lesniewska B., Fuchs B. (2014) Worse prognosis of osteosarcoma patients expressing IGF-1 on a tissue microarray. Anticancer Res 34(8) :3881-9.
8 Dahlin D.C. (1978) Bone tumors. Thomas, Springfield
9 Schajowicz F. (1981, 1994) Tumors and tumorlike lesions of bone and joints. Springer, Berlin, Heidelberg, New York, Tokyo
10 Adler C. P. (1998) Knochenkrankheiten: Diagnostik makroskopischer, histologischer und radiologischer Strukturveränderungen des Skeletts. Springer, Berlin, Heidelberg, New York, Barcelona, Hongkong, London, Mailand, Paris, Tokio
11 Thomas C. (1996) Spezielle Pathologie. Schattauer, Stuttgart, New York
12 Unni K.K. (1982) Case report 214. Skeletal Radiol 9: 129
13 Freyschmidt J., Ostertag H., Jundt G., (2010) Knochentumoren mit Kiefertumoren, Klinik Radiologie Pathologie. Springer, Dordrecht, Heidelberg, London, New York: 161-183
14 Schumacher K. (2000) Therapie maligner Tumoren, Integration konventioneller und komplementärer Therapie, Manual für Klinik und Praxis. Schattauer, Stuttgart
15 Poppe H. (1978) Aussagewert, Grenzen und Schwierigkeiten der Röntgendiagnostik und der Computertomographie bei primär und potentiell malignen Geschwulsten des Skeletts. Z Orthop 116: 460
16 Rosen G., Marcove R.C., Huvos A.G., Caparros B.I., Lane J.M., Nirenberg A., Cacavio A., Groshen S. (1983) Primary osteogenic sarcoma: eight-year experience with adjuvant chemotherapy. J Cancer Res Clin Oncol 106 [Abstract]: 55.

LITERATURVERZEICHNIS
60
17 Goorin A. M., Delorey M. J., Lack E. E., Gelber R. D., Price K., Cassady J. R., Levey R., Tapper D., Jaffe N., Link M. (1984). Prognostic significance of complete surgical resection of pulmonary metastases in patients with osteogenic sarcoma: analysis of 32 patients. J Clin Oncol, 2, 425.
18 Kager L., Zoubek A., Pötschger U., Kastner U., Flege S., Kempf-Bielack B., Branscheid D., Kotz R., Salzer-Kuntschik M., Winkelmann W., Jundt G., Kabisch H., Reichardt P., Jürgens H., Gadner H., Bielack S.S.; (2003) Primary metastatic osteosarcoma: presentation and outcome of patients treated on neoadjuvant Cooperative Osteosarcoma Study Group protocols. J Clin Oncol 21:2011–2018.
19 Kempf-Bielack B., Bielack S. S., Jürgens H., Branscheid D., Berdel W. E., Exner G. U., Göbel U., Helmke K., Jundt G., Kabisch H., Kevric M., Klingebiel T., Kotz R., Maas R., Schwarz R., Semik M., Treuner J., Zoubek A., Winkler K. (2005) Osteosarcoma relapse after combined modality therapy: an analysis of unselected patients in the Cooperative Osteosarcoma Study Group (COSS). J Clin Oncol 23:559–568
20 Bielack S. S., Kempf-Bielack B., Delling G., Exner G. U., Flege S., Helmke K., Kotz R., Salzer-Kuntschik M., Werner M., Winkelmann W., Zoubek A., Jürgens H., Winkler K. (2002) Prognostic factors in high-grade osteosarcoma of the extremities or trunk: an analysis of 1,702 patients treated on neoadjuvant cooperative osteosarcoma study group protocols. J Clin Oncol 20:776–790.
21 Meyers P. A., Schwartz C. L., Krailo M. D., Healey J. H., Bernstein M. L., Betcher D., Ferguson W. S., Gebhardt M. C., Goorin A. M., Harris M., Kleinerman E., Link M.P., Nadel H., Nieder M., Siegal G. P., Weiner M. A., Wells R. J., Womer R. B., Grier H. E. (2008) Osteosarcoma: the addition of muramyl tripeptide to chemotherapy improves overall survival – a report from the Children’s Oncology Group. J Clin Oncol 26:633–638
22 Balkwill F. (2009) Tumour necrosis factor and cancer. Nat Rev Cancer 9: 361–371
23 Meyers P.A. (2009) Muramyl tripeptide (mifamurtide) for the treatment of osteosarcoma. Expert Rev Anticancer Ther 9: 1035–1049
24 Geddes K., Magalhaes J.G., Girardin S.E. (2009) Unleashing the therapeutic potential of NOD-like receptors. Nat Rev Drug Discov 8: 465–479
25 Kager L., Pötschger U., Bielack S. (2010) Review of mifamurtide in the treatment of patients with osteosarcoma. Ther Clin Risk Manag 6: 279-286
26 McAllister R.M., Filbert J.E., Nicolson M.O., Rongey R.W., Gardner M.B., Gilden R.V., Huebner R.J. (1971) Transformation and productive infection of human osteosarcoma cells by a feline sarcoma virus. Nat New Biol 230(17): 279-282.
27 Rhim J.S., Park D.K., Arnstein P., Huebner R.J., Weisburger E.K., Nelson-Rees W.A. (1975) Transformation of human cells in culture by N-methyl-N_-nitro-N-nitrosoguanidine. Nature 256: 751–753
28 Ponten J., Saksela E. (1967) Two established in vitro cell lines from human mesenchymal tumours. Int J Cancer 2: 434-447
29 Gümbel D., Gelbrich N., Weiss M., Napp M., Daeschlein G., Sckell A., Ender S.A., Kramer A., Burchardt M., Ekkernkamp A., Stope M.B.. (2016) New Treatment Options for Osteosarcoma - Inactivation of Osteosarcoma Cells by Cold Atmospheric Plasma. Anticancer Res 36(11): 5915-5922
30 Pettke A., Hotfilder M., Clemens D., Klco-Brosius S., Schaefer C., Potratz J., Dirksen U. (2016) Suberanilohydroxamic acid (vorinostat) synergistically enhances the cytotoxicity of doxorubicin and cisplatin in osteosarcoma cell lines. Anticancer Drugs 27(10):1001-10.

LITERATURVERZEICHNIS
61
31 Ko Y., Choi A., Lee M., Lee J.A. (2016) Metformin displays in vitro and in vivo antitumor effect against osteosarcoma. Korean J Pediatr 59(9): 374-380
32 Tsai Y.F., Huang C.W., Chiang J.H., Tsai F.J., Hsu Y.M., Lu C.C., Hsiao C.Y., Yang J.S. (2016) Gadolinium chloride elicits apoptosis in human osteosarcoma U-2 OS cells through extrinsic signaling, intrinsic pathway and endoplasmic reticulum stress. Oncol Rep 36(6): 3421-3426
33 Ender S., Dallmer A., Lässig F., Lendeckel U., Wolke C. (2014) Expression and function of the ACE2/angiotensin(1-7)/Mas axis in osteosarcoma cell lines U-2 OS and MNNG-HOS. Mol Med Rep 10(2): 804-10
34 Wright J.W., Yamamoto, B.J., Harding J.W. (2008) Angiotensin receptor subtype mediated physiologies and behaviors: New discoveries and clinical targets. Prog Neurobiol 84(2): 157–181
35 Klinke R., Pape, H.-C., Kurtz, A., Silbernagl S. (2009) Physiologie. Thieme, Stuttgart, New York, 6. Auflage: 42-43, 375-376
36 Behrends J., Bischofsberger J., Deutzmann R., Kurtz A., Wagner C., Hoth M., Rettig J. (2009) Duale Reihe Physiologie. Thieme, Stuttgart: 319-321
37 Donoghue M., Hsieh F., Baronas E., Godbout K., Gosselin M., Stagliano N., Donovan M., Woolf B., Robison K., Jeyaseelan R., Breitbart R. E., Acton S. (2000) A novel angiotensinconverting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ Res 87: E1—9
38 Tipnis S. R., Hooper N. M., Hyde R., Karran E., Christie G., Turner A. J. (2000) A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem 275: 33238—43
39 Santos R.A.S., Ferreira A.J., Simoes E., Silva A.C. (2008) Recent advances in the angiotensin-converting enzyme 2-angiotensin(1-7)-Mas axis. Exp Physiol 93:519–527.
40 Chappell M.C., Tallant E.A., Brosnihan K.B., Ferrario C.M. (1994) Conversion of angiotensin I to angiotensin-(1-7) by thimet oligopeptidase (EC 3.4.24.15) in vascular smooth muscle cells.J Vasc Med Biol 5: 129–137
41 Santos R.A.S., Brosnihan K.B., Jacobsen D.W., DiCorleto P.E., Ferrario C.M. (1992) Production of angiotensin-(1-7) by human vascular endothelium. Hypertension 19(Suppl II): 56-61
42 Turner A.J. (2003) Exploring the structure and function of zinc metallopeptidases: old enzymes and new discoveries. Biochem Soc Trans 31: 723–727
43 Campbell D.J., Anastasopoulos F., Duncan A.M., James G.M., Kladis A., Briscoe T.A. (1998) Effects of neutral endopeptidase inhibition and combined angiotensin converting enzyme and neutral endopeptidase inhibition on angiotensin and bradykinin peptides in rats. J Pharmacol Exp Ther 287: 567–577
44 Iyer S.N., Ferrario C.M., Chappell M.C. (1998) Angiotensin-(1-7) contributes to the antihypertensive effects of blockade of the renin–angiotensin system. Hypertension 31: 356–361
45 Rice G.I., Thomas D.A., Grant P.J., Turner A.J., Hooper N.M. (2004) Evaluation of angiotensin-converting enzyme (ACE), its homologue ACE2 and neprilysin in angiotensin peptide metabolism. Biochem J 383: 45–51
46 Crackower M.A., Sarao R., Oudit G.Y., Yagil C., Kozieradzki I., Scanga S.E., Oliveira-dos-Santo A.J., da Costa J., Zhang L., Pei Y., Scholey J., Bray M.R., Ferrario C.M., Backx P.H., Manoukian A.S.,

LITERATURVERZEICHNIS
62
Chappell M.C., Yagil Y., Penninger J.M. (2002) Angiotensinconvertingenzyme 2 is an essential regulator of heart function. Nature 417: 822–828
47 Zimpelmann J., Burns K.D. (2009) Angiotensin-(1-7) activates growth-stimulatory pathways in human mesangial cells. Am J Physiol Renal Physiol 296(2): F337–F346.
48 Velkoska E., Dean R.G., Griggs K., Burchill L., Burrell L.M. (2011) Angiotensin-(1-7) infusion is associated with increased blood pressure and adverse cardiac remodelling in rats with subtotal nephrectomy. Clin Sci (Lond) 120(8): 335–345
49 Shao Y., He M., Zhou L., Yao T., Huang Y., Lu L.M. (2008) Chronic angiotensin (1-7) injection accelerates STZ-induced diabetic renal injury. Acta Pharmacol Sin 29(7): 829–837
50 Young D., Waitches G., Birchmeier C., Fasano O., Wigler M. (1986) Isolation and characterization of a new cellular oncogene encoding a protein with multiple potential transmembrane domains. Cell 6;45(5): 711-9.
51 Santos R.A.S., Simoes e Silva A.C., Maric C., Silva D.M., Machado R.P., de Bul I., Heringer-Walther S., Pinheiro S.V., Lopes M.T., Bader M., Mendes E.P., Lemos V.S., Campagnole-.-Santos M.J., Schultheiss H.-P., Speth R., Walther T. (2003) Angiotensin-.-(1-.-7) is an endogenous ligand for the G protein-.-coupled receptor Mas. Proc Natl Acad Sci USA 100: 8258–8263
52 Dias-Peixoto M.F., Santos R.A.S., Gomes E. R., Alves M.N., Almeida P.W., Greco L., Rosa M., Fauler B., Bader M., Alenina N., Guatimosim S. (2008) Molecular mechanisms involved in the angiotensin-(1–7)/mas signaling pathway in cardiomyocytes, Hypertension 52(3): 542–548
53 Yuan L., Lu C.L., Wang Y., Li Y., Li X.Y. (2014) Ang (1-7) protects islet endothelial cells from palmitate-induced apoptosis by AKT, eNOS, p38 MAPK, and JNK pathways. J Diabetes Res. 2014;2014:391476.
54 Karnik S.S., Unal H., Kemp J.R., Tirupula K.C., Eguchi S., Vanderheyden P.M., Thomas W.G. (2015) International Union of Basic and Clinical Pharmacology. XCIX. Angiotensin Receptors: Interpreters of Pathophysiological Angiotensinergic Stimuli. Pharmacol Rev. 2015 Oct;67(4):754-819.
55 Singh KD, Karnik SS. (2016 ) Angiotensin Receptors: Structure, Function, Signaling and Clinical Applications. J Cell Signal. 2016 Jun;1(2). pii: 111.
56 Sahr A., Wolke C., Maczewsky J., Krippeit-Drews P., Tetzner A., Drews G., Venz S., Gürtler S., van den Brandt .J, Berg S., Döring P., Dombrowski F., Walther T., Lendeckel U. (2016) The Angiotensin-(1-7)/Mas Axis Improves Pancreatic β-Cell Function in Vitro and in Vivo. Endocrinology. 2016 Dec;157(12):4677-4690.
57 Meinert C., Kohse F., Böhme I., Gembardt F., Tetzner A., Wieland T., Greenberg B., Walther T. (2016) Further intracellular proteins and signaling pathways regulated by angiotensin-(1-7) in human endothelial cells. Data Brief. 2016 Dec 8;10:354-363.
58 Sampaio W.O., Souza dos Santos R.A., Faria-Silva R., da Mata Machado L.T., Schiffrin E.L., Touyz R.M. (2007) Angiotensin-(1-7) through receptor Mas mediates endothelial nitric oxide synthase activation via Akt-dependent pathways. Hypertension. 2007 Jan; 49(1):185-92.
59 Giani J.F., Gironacci M.M., Muñoz M.C., Peña C., Turyn D., Dominici F.P. (2007) Angiotensin-(1 7) stimulates the phosphorylation of JAK2, IRS-1 and Akt in rat heart in vivo: role of the AT1 and Mas receptors. Am J Physiol Heart Circ Physiol. 2007 Aug;293(2):H1154-63

LITERATURVERZEICHNIS
63
60 Ni L., Feng Y., Wan H., Ma Q., Fan L., Qian Y., Li Q., Xiang Y., Gao B. (2012) Angiotensin-(1-7) inhibits the migration and invasion of A549 human lung adenocarcinoma cells through inactivation of the PI3K/Akt and MAPK signaling pathways. Oncol Rep. 2012 Mar;27(3):783-90.
61 Zhang F., Ren X., Zhao M., Zhou B., Han Y. (2016) Angiotensin-(1-7) abrogates angiotensin II-induced proliferation, migration and inflammation in VSMCs through inactivation of ROS-mediated PI3K/Akt and MAPK/ERK signaling pathways. Sci Rep. 2016 Sep 30;6:34621
62 Tallant E. A., Ferrario C. M., Gallagher P. E. (2005) Angiotensin- (1–7) inhibits growth of cardiac myocytes through activation of the mas receptor. Am J Physiol 289(4): 1560-1566
63 Haznedaroglu IC, Malkan UY. (2016) Local bone marrow renin-angiotensin system in the genesis of leukemia and other malignancies. Eur Rev Med Pharmacol Sci. 2016 Oct;20(19):4089-4111.
64 Verano-Braga T., Schwämmle V., Sylvester M., Passos-Silva D.G., Peluso A.A., Etelvino G.M., Santos R.A., Roepstorff P. (2012) Time-resolved quantitative phosphoproteomics: new insights into Angiotensin-(1-7) signaling networks in human endothelial cells.J Proteome Res.;11(6):3370-81.
65 Meinert C., Kohse F., Böhme I., Gembardt F., Tetzner A., Wieland T., Greenberg B., Walther T. (2016) Further intracellular proteins and signaling pathways regulated by angiotensin-(1-7) in human endothelial cells.Data Brief. 2016 Dec 8;10:354-363.
66 Oliveira M.A., Fortes Z.B., Santos R.A.S., Kosla M.C., De C.M. (1999) Synergistic effect of angiotensin-(1-7) on bradykinin arteriolar dilation in vivo. Peptides 20:1195–1201
67 Iyer S.N., Chappell M.C., Averill D.B., Diz D.I., Ferrario C.M. (1998) Vasodepressor actions of angiotensin-(1-7) unmasked during combined treatment with lisinopril and losartan. Hypertension 31: 699–705
68 De Mello W.C. (2004) Angiotensin (1-7) re-establishes impulse conduction in cardiac muscle during ischaemia-reperfusion. The role of the sodium pump. J Renin Angiotensin Aldosterone Syst 5:2 03–208
69 Grobe J.L., Mecca A.P., Lingis M., Shenoy V., Bolton T.A., Machado J.M., Speth R.C., Raizada M.K., Katovich M.J. (2007) Prevention of angiotensin II-induced cardiac remodeling by angiotensin-(1-7). Am J Physiol 292: 736–742
70 Ferreira A.J.,Oliveira T.L., Castro M.C., Almeida A.P., Castro C.H., Caliari M.V., Gava E., Kitten G.T., Santos R.A.S. (2007) Isoproterenol induced impairment of heart function and remodeling are attenuated by the nonpeptide angiotensin-(1-7) analogue AVE-0991 Life Sci 81: 916–923
71 Ren Y., Garvin J.L., Carretero O.A. (2002) Vasodilator action of angiotensin-(1-7) on isolated rabbit afferent arterioles. Hypertension 39: 799–802
72 DelliPizzi A., Hilchey S.D., Bell-Quilley C.P. (1994) Natriuretic action of angiotensin (1-7). Br J Pharmacol 111: 1–3
73 Ferrario C.M., Smith R.D., Brosnihan K.B., Chappell M.C., Campese V.M., Vesterqvist O., Liao W., Ruddy M.C., Grim C.E. (2002) Effects of omapatrilat on the renin angiotensin system in salt sensitive hypertension. Am J Hyperten 15: 557–564
74 Luque M., Martin P., Martell N., Fernandez C., Brosnihan K.B., Ferrario C.M. (1996) Effects of captopril related to increased levels of prostacyclin and angiotensin-(1-7) in essential hypertension. J Hypertens 14: 799–805

LITERATURVERZEICHNIS
64
75 Wang, K., Hu, X., Du, C., Tu, S., Zhang, F. and Xie, X. (2012) Angiotensin-(1–7) suppresses the number and function of the circulating fibrocytes by upregulating endothelial nitric oxide synthase expression. Mol Cell Biochem 365, 19–27
76 Lubel, J. S., Herath, C. B., Tchongue, J., Grace, J., Jia, Z.,Spencer, K., Casley, D., Crowley, P., Sievert, W., Burrell, L. M., Angus, P. W. (2009) Angiotensin-(1–7), an alternative metabolite of the renin-angiotensin system, is up-regulated in human liver disease and has antifibrotic activity in the bile-duct-ligated rat. Clin Sci 117, 375–386
77 Shenoy, V., Ferreira, A. J., Qi, Y., Fraga-Silva, R. A., Diez-Freire, C., Dooies, A., Jun, J. Y., Sriramula, S., Mariappan, N., Pourang, D., Venugopal C. S., Francis J., Reudelhuber T., Santos R. A., Patel J. M., Raizada M. K., Katovich M. J. (2010) The angiotensin-converting enzyme 2/angiogenesis-(1–7)/Mas axis confers cardiopulmonary protection against lung fibrosis and pulmonary hypertension. Am J Respir Crit Care Med 182, 1065–1072
78 da Silveira, K. D., Coelho, F. M., Vieira, A. T., Sachs, D., Barroso, L. C., Costa, V. V., Bretas, T. L., Bader, M., de Sousa, L. P., Silva T. A., dos Santos R. A., Simões e Silva A. C., Teixeira M. M. (2010) Anti-inflammatory effects of the activation of the angiotensin-(1–7) receptor, MAS, in experimental models of arthritis. J Immunol 185, 5569–5576
79 Statistisches Bundesamt, (2012) Todesursachen in Deutschland. Fachserie 12, Reihe 4, 3
80 Tesanovic, S., Vinh, A., Gaspari, T. A., Casley, D., Widdop, R. E. (2010) Vasoprotective and atheroprotective effects of angiotensin (1–7) in apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 30, 1606–1613
81 Giani, J. F., Mayer, M. A., Munoz, M. C., Silberman, E. A., Hocht, C., Taira, C. A., Gironacci, M. M., Turyn, D., Dominici, F. P. (2009) Chronic infusion of angiotensin-(1–7) improves insulin resistance and hypertension induced by a high-fructose diet in rats. Am J Physiol Endocrinol Metab 296, E262–E271
82 Santos, S. H., Fernandes, L. R., Mario, E. G., Ferreira, A. V., Porto, L. C., Alvarez-Leite, J. I., Botion, L. M., Bader, M., Alenina, N., Santos, R. A.S. (2008) Mas deficiency in FVB/N mice produces marked changes in lipid and glycemic metabolism. Diabetes 57, 340–347
83 Ardaillou R. (1997) Active fragments of angiotensin II: enzymatic pathways of synthesis and biological effects. Curr Opin Nephrol Hypertens 6(1):28–34.
84 Ardaillou R, Chansel D. (1997) Synthesis and effects of active fragments of angiotensin II. Kidney Int 52(6):1458–1468.
85 Zini S., Fournie-Zaluski M.C., Chauvel E., Roques B.P., Corvol P., Llorens-Cortes C. (1996) Identification of metabolic pathways of brain angiotensin II and III using specific aminopeptidase inhibitors: predominant role of angiotensin III in the control of vasopressin release. Proc Natl Acad Sci 93(21): 11968-73
86 Braszko J.J., Walesiuk A., Wielgat P. (2006) Cognitive effects attributed to angiotensin II may result from its conversion to angiotensin IV. J Renin Angiotensin Aldosterone Syst. (3): 168-74
87 Becker, L. K., Etelvino, G. M., Walther, T., Santos, R. A.S., Campagnole-Santos, M. J. (2007) Immunofluorescence localization of the receptor Mas in cardiovascular-related areas of the rat brain. Am J Physiol Heart Circ Physiol 293, H1416–H1424
88 Metzger, R., Bader, M., Ludwig, T., Berberich, C., Bunnemann, B., Ganten, D. (1995) Expression of the mouse and rat mas proto-oncogene in the brain and peripheral tissues. FEBS Lett 357, 27–32

LITERATURVERZEICHNIS
65
89 Lazaroni, T. L., Raslan, A. C., Fontes, W. R., de Oliveira, M. L., Bader, M., Alenina, N., Moraes, M. F., Dos Santos, R. A.S., Pereira, G. S. (2012) Angiotensin-(1–7)/Mas axis integrity is required for the expression of object recognition memory. Neurobiol Learn Mem 97, 113–123
90 Merrill D.C, Karoly M., Chen K., Ferrario C.M., Brosnihan K.B. (2002) Angiotensin-(1-7) in normal and preeclamptic pregnancy. Endocrine 18: 239–245
91 Valdes G., Neves L.A., Anton L., Corthorn J., Chacon C., Germain A.M., Merrill D.C., Ferrario C.M., Sarao R., Penninger J., Brosnihan K.B. (2006) Distribution of angiotensin-(1-7) and ACE2 in human placentas of normal and pathological pregnancies. Placenta 27: 200–207
92 Vaajanen, A., Vapaatalo, H., Kautiainen, H., Oksala, O. (2008) Angiotensin (1–7) reduces intraocular pressure in the normotensive rabbit eye. Invest Ophthalmol Visual Sci 49, 2557–2562
93 Mecca, A. P., Regenhardt, R. W., O’Connor, T. E., Joseph, J. P., Raizada, M. K., Katovich, M. J., Sumners, C. (2011) Cerebroprotection by angiotensin-(1–7) in endothelin-1-induced ischaemic stroke. Exp Physiol 96, 1084–1096
94 Borges, E. L., Cabral, B. M., Braga, A. A., Neves, M. J., Santos, R. A.S., Rogana, E. (2002) Effect of angiotensin-(1–7) on jejunal absorption of water in rats. Peptides 23, 51–56
95 Durik, M., Seva Pessoa, B., Roks, A. J. (2012) The renin-angiotensin system, bone marrow and progenitor cells. Clin Sci 123, 205–223
96 Baykan, H., Gunay, G. K., Ozyazgan, I., Soyuer, I. (2012) The effect of angiotensin (1–7) on survival of random pattern skin flaps with nicotine-induced ischemia in rats. Ann Plast Surg 68, 88–93
97 Passos-Silva D.G, Verano-Braga T., Santos R.A.S. (2013) Angiotensin-(1–7): beyond the cardio-renal actions Clinical Science 124, 443–456
98 Zhang Y., Kou J., Wang C., Wang K. (2014) Role of reactive oxygen species in angiotensin II: induced receptor activator of nuclear factor-κB ligand expression in mouse osteoblastic cells. Mol Cell Biochem. 2014 Nov;396(1-2):249-55.
99 Querques F., Cantilena B., Cozzolino C., Esposito M.T., Passaro F., Parisi S., Lombardo B., Russo T., Pastore L. (2015) Angiotensin receptor I stimulates osteoprogenitor proliferation through TGFβ-mediated signaling. J Cell Physiol. 2015 Jul;230(7):1466-74
100 Araújo A.A., Lopes de Souza G., Souza T.O., de Castro Brito G.A., Sabóia Aragão K., Xavier de Medeiros C.A., Lourenço Y., do Socorro Costa Feitosa Alves M., Fernandes de Araújo R. Jr. (2013) Olmesartan decreases IL-1β and TNF-α levels; downregulates MMP-2, MMP-9, COX-2, and RANKL; and upregulates OPG in experimental periodontitis. Naunyn Schmiedebergs Arch Pharmacol. 2013 Oct;386(10):875-84.
101 Gallagher, P. E., Tallant, E. A. (2004) Inhibition of human lung cancer cell growth by angiotensin-(1–7). Carcinogenesis 25, 2045–2052
102 Pantoja D.S., Tallant A.E., Gallagher P.E. (2004) Inhibition of human breast cancer cell growth by angiotensin-(1-7). Proc Am Assoc Cancer Res 45
103 Menon J., Soto-Pantoja D.R., Callahan M.F., Cline J.M., Ferrario C.M., Tallant E.A., Gallagher P.E. (2007) Angiotensin-(1-7) inhibits growth of human lung adenocarcinoma xenografts in nude mice through a reduction in cyclooxygenase-2. Cancer Res 67:2809–2815

LITERATURVERZEICHNIS
66
104 Putney J.W. (2005) Physiological mechanisms of TRPC activation. Pflugers Arch 451: 29–34
105 D.E. Clapham, D.E., Runnels L.W.,Strübing C. (2002) The TRP ion channel family, Nat Rev, Neurosci 2 387– 396.
106 Zhu X., Jiang M., Peyton M., Boulay G., Hurst R., Stefani E., Birnbaumer L. (1996) Trp, a novel mammalian gene family essential for agonist-activated capacitative Ca2+ entry. Cell 85: 661–671
107 Philipp S., Cavalie A., Freichel M., Wissenbach U., Zimmer S., Trost C., Marquart A., Murakami M., Flockerzi V. (1996) A mammalian capacitative calcium entry channel homologous to Drosophila TRP and TRPL. EMBO J 15: 6166–6171
108 Zitt C., Zobel A., Obukhov A.G., Harteneck C., Kalkbrenner F., Lueckhoff A., Schultz G. (1996) Cloning and functional expression of a human Ca2+-permeable cation channel activated by calcium store depletion. Neuron 16: 1189–1196
109 Hofmann T., Schaefer M., Schultz G., Gudermann T. (2002) Subunit composition of mammalian transient receptor potential channels in living cells. Proc Natl Acad Sci USA 99: 7461–7466
110 Cheng, W., Yang, F., Takanishi, C.L., Zheng, J. (2007) Thermosensitive TRPV channelsubunits coassemble into heteromeric channels with intermediate conductanceand gating properties. J Gen Phys 129, 191–207.
111 Ambudkar I.S. (2007) TRPC1: a core component of storeoperated calcium channels. Biochem Soc Trans 35: 96–100
112 Beech D.J. (2005) TRPC1: store-operated channel and more. Pflugers Arch 451: 53–60
113 Brueggemann L.I., Markun D.R., Henderson K.K., Cribbs L.L., Byron K.L. (2006) Pharmacological and electrophysiological characterization of store-operated currents and capacitative Ca (2+) entry in vascular smooth muscle cells. J Pharmacol Exp Ther 317: 488–499
114 Liu X., Bandyopadhyay B.C., Singh B.B., Groschner K., Ambudkar I.S. (2005) Molecular analysis of a store-operated and 2-acetyl-snglycerol-sensitive non-selective cation channel. Heteromeric assembly of TRPC1-TRPC3. J Biol Chem 280: 21600–21606
115 Lopez J.J., Salido G.M., Pariente J.A., Rosado J.A. (2006) Interaction of STIM1 with endogenously expressed human canonical TRP1 upon depletion of intracellular Ca2+ stores. J Biol Chem 281: 28254–28264
116 Ong H.L., Cheng K.T., Liu X., Bandyopadhyay B.C., Paria B.C., Soboloff J., Pani B., Gwack Y., Srikanth S., Singh B.B., Gill D., Ambudkar I.S. (2007) Dynamic assembly of TRPC1/STIM1/Orai1 ternary complex is involved in store-operated calcium influx: evidence for similarities in SOC and CRAC channel components. J Biol Chem 282(12): 9105–9116
117 Yuan J.P., Kiselyov K., Shin D.M., Chen J., Shcheynikov N., Kang S.H., Dehoff M.H., Schwarz M.K., Seeburg P.H., Muallem S., Worley P.F. (2003) Homer binds TRPC family channels and is required for gating of TRPC1 by IP3 receptors. Cell 114: 777–789
118 Bollimuntha S., Singh B.B., Shavali S., Sharma S.K., Ebadi M. (2005b) TRPC1-mediated inhibition of 1-methyl-4-phenylpyridinium-ion neurotoxicity in human SH-SY5Y neuroblastomacells. J Biol Chem 280: 2132–2140

LITERATURVERZEICHNIS
67
119 Rao J.N., Platoshyn O., Golovina V.A., Liu L., Zou T., Marasa B.S., Turner D.J., Yuan J.X., Wang J.Y. (2006) TRPC1 functions as a store-operated Ca2+ channel in intestinal epithelial cells and regulates early mucosal restitution after wounding. Am J Physiol Gastrointest Liver Physiol 290: G782–G792
120 Liman E.R., Corey D.P., Dulac C. (1999) TRP2: a candidate transduction channel for mammalian pheromone sensory signalling. Proc Natl Acad Sci USA 96: 5791–5796
121 Zufall F. (2005) The TRPC2 ion channel and pheromone sensingin the accessory olfactory system. Naunyn Schmiedebergs Arch Pharmacol 371: 245–250
122 Vazquez G., Wedel B.J., Trebak M., St. John Bird G., Putney J.W. Jr. (2003) Expression level of the canonical transient receptor potential 3 (TRPC3) channel determines its mechanism of activation. J Biol Chem 278: 21649–21654
123 Li, Y., Jia, Y.C., Cui, K., Li, N., Zheng, Z.Y., Wang, Y.Z., Yuan, X.B. (2005) Essential role of TRPC channels in the guidance of nerve growth cones by brain-derived neurotrophic factor. Nature 434, 894–898
124 Hartmann, J., Dragicevic, E., Adelsberger, H., Henning, H.A., Sumser, M., Abramowitz,J., Blum, R., Dietrich, A., Freichel, M., Flockerzi, V., Birnbaumer, L., Konnerth, A. (2008) TRPC3 channels are required for synaptic transmission and motor coordination. Neuron 59, 392–398.
125 Nakayama, H., Wilkin, B.J., Bodi, I., Molkentin, J.D. (2006) Calcineurin-dependent cardiomyopathy is activated by TRPC in the adult mouse heart. FASEB Journal 20, 1660–1670.
126 Thilo, F., Loddenkemper, C., Berg, E., Zidek, W., Tepel, M., (2009) Increased TRPC3 expression in vascular endothelium of patients with malignant hypertension. Modern Pathology 22, 426–430.
127 Yu, Y., Fantozzi, I., Remillard, C.V., Landsberg, J.W., Kunichika, N., Platoshyn, O., Tigno, D.D., Thistlethwaite, P.A., Rubin, L.J., Yuan, J.X., 2004. Enhanced expression of transient receptor potential channels in idiopathic pulmonary arterial hypertension. Proc Natl Acad Sci USA 101, 13861–13866.
128 Freichel M., Suh S.H., Pfeifer A., Schweig U., Trost C., Weibgerber P., Biel M., Philipp S., Freise D., Droogmans G., Hofmann F., Flockerzi V., Nilius B. (2001) Lack of an endothelial storeoperated Ca2+ current impairs agonist-dependent vasorelaxation in TRP4−/− mice. Nat Cell Biol 3: 121–127
129 Tiruppathi C., Freichel M., Vogel S.M., Paria B.C., Mehta D., Yang H., Mergler S., Sun X., Wang Z., Lu L., Bonanno J.A., Pleyer U., Reinach P.S. (2005) TRPC4 knockdown suppresses epidermal growth factor-induced store-operated channel activation and growth in human corneal epithelial cells. J Biol Chem 280: 32230–32237
130 Greka A., Navarro B., Oancea E., Duggan A., Clapham D.E. (2003) TRPC5 is a regulator of hippocampal neurite length and growth cone morphology. Nat Neurosci 6: 837–845
131 Hui H., McHugh D., Hannan M., Zeng F., Xu S.Z., Khan S.U., Levenson R., Beech D.J., Weiss J.L. (2006) Calcium-sensing mechanism in TRPC5 channels contributing to retardation of neurite outgrowth. J Physiol 572: 165–172
132 Parekh A.B., Putney J.W. Jr. (2005) Store-operated calcium channels. Physiol Rev 85:757–810
133 Plant T.D., Schaefer M. (2005) Receptor-operated cation channels formed by TRPC4 and TRPC5. Naunyn Schmiedebergs Arch Pharmacol 371:266–276

LITERATURVERZEICHNIS
68
134 Xu S.Z., Boulay G., Flemming R., Beech D.J. (2006) E3-targeted anti-TRPC5 antibody inhibits store-operated calcium entry in freshly isolated pial arterioles. Am J Physiol Heart Circ Physiol 291: H2653–H2659
135 Dietrich A., Kalwa H., Rost B.R., Gudermann T. (2005) The diacylgylcerol-sensitive TRPC3/6/7 subfamily of cation channels: functional characterization and physiological relevance. Pflugers Arch 451: 72–80
136 Dietrich A., Mederos Y.S.M., Gollasch M., Gross V., Storch U., Dubrovska G., Obst M., Yildirim E, Salanova B., Kalwa H., Essin K., Pinkenburg O., Luft F.C., Gudermann T., Birnbaumer L. (2005) Increased vascular smooth muscle contractility in TRPC6−/− mice. Mol Cell Biol 25: 6980–6989
137 Nagamine K., Kudoh J., Minoshima S., Kawasaki K., Asakawa S., Ito F., Shimizu N. (1998) Molecular cloning of a novel putative Ca2+ channel protein (TRPC7) highly expressed in brain. Genomics 54: 124–131
138 Yoon I.S., Li P.P., Siu K.P., Kennedy J.L., Macciardi F., Cooke R.G., Parikh S.V., Warsh J.J. (2001) Altered TRPC7 gene expression in bipolar-I disorder. Biol Psychiatry 50: 620–626
139 Karnik S.S., Singh K.D., Tirupula K., Unal H. (2017) Significance of angiotensin 1-7 coupling with MAS1 receptor and other GPCRs to the renin-angiotensin system: IUPHAR Review 22. Br J Pharmacol. 2017 May;174(9):737-753
140 Meyers P.A., Schwartz C.L., Krailo M.D., Healey J.H., Bernstein M.L., Betcher D., Ferguson W.S., Gebhardt M.C., Goorin A.M., Harris M., Kleinerman E., Link M.P., Nadel H., Nieder M., Siegal G.P., Weiner M.A., Wells R.J., Womer R.B., Grier H.E. (2008) Children's Oncology Group. Osteosarcoma: the addition of muramyl tripeptide to chemotherapy improves overall survival--a report from the Children's Oncology Group. J Clin Oncol. 2008 Feb 1;26(4):633-8.
141 Wasa J., Sugiura H., Kozawa E., Kohyama K., Yamada K., Taguchi O. (2011) The tumor suppressive effect of angiotensin II type 1 receptor antagonist in a murine osteosarcoma model. Anticancer Res. 2011 Jan;31(1):123-7
142 Souza L.L., Costa-Neto C.M. (2012) Angiotensin-(1-7) decreases LPS-induced inflammatory response in macrophages. J Cell Physiol. 2012 May;227(5):2117-22.
143 Stewart T.A., Yapa K.T., Monteith G.R. (2015) Altered calcium signaling in cancer cells. Biochim Biophys Acta. 2015 Oct;1848(10 Pt B):2502-11
144 Hanahan D, Weinberg RA. (2000) The hallmarks of cancer. Cell. 2000 Jan 7;100(1):57-70.
145 Kondratskyi A., Kondratska K., Skryma R., Prevarskaya N. (2015) Ion channels in the regulation of apoptosis. Biochim Biophys Acta. 2015 Oct;1848(10 Pt B):2532-46.
146 Tajeddine N., Gailly P. (2012) TRPC1 protein channel is major regulator of epidermal growth factor receptor signaling. J Biol Chem. 2012 May 11;287(20):16146-57.
147 Pigozzi D., Ducret T., Tajeddine N., Gala J.L., Tombal B., Gailly P. (2006) Calcium store contents control the expression of TRPC1, TRPC3 and TRPV6 proteins in LNCaP prostate cancer cell line. Cell Calcium. 2006 May;39(5):401-15.
148 El Hiani Y., Ahidouch A., Lehen'kyi V., Hague F., Gouilleux F., Mentaverri R., Kamel S., Lassoued K., Brûlé G., Ouadid-Ahidouch H. (2009) Extracellular signal-regulated kinases 1 and 2 and TRPC1 channels are required for calcium-sensing receptor-stimulated MCF-7 breast cancer cell proliferation.Cell Physiol Biochem. 2009;23(4-6):335-46.

LITERATURVERZEICHNIS
69
149 Chen J., Luan Y., Yu R., Zhang Z., Zhang J., Wang W. (2014) Transient receptor potential (TRP) channels, promising potential diagnostic and therapeutic tools for cancer. Biosci Trends. 2014 Feb;8(1):1-10
150 Zhang H, Zhou L, Shi W, Song N, Yu K, Gu Y. (2012) A mechanism underlying the effects of polyunsaturated fatty acids on breast cancer. Int J Mol Med. 2012 Sep;30(3):487-94.
151 Yang S.L., Cao Q., Zhou K.C., Feng Y.J., Wang Y.Z. (2009) Transient receptor potential channel C3 contributes to the progression of human ovarian cancer. Oncogene. 2009 Mar 12;28(10):1320-8.
152 Cheng K.T., Ong H.L., Liu X., Ambudkar I.S. (2013) Contribution and regulation of TRPC channels in store-operated Ca2+ entry. Curr Top Membr. 2013;71:149-79.
153 Yang SL, Cao Q, Zhou KC, Feng YJ, Wang YZ. (2009) Transient receptor potential channel C3 contributes to the progression of human ovarian cancer. Oncogene. 2009 Mar 12;28(10):1320-8.
154 Carson C., Raman P., Tullai J., Xu L., Henault M., Thomas E., Yeola S., Lao J., McPate M., Verkuyl J.M., Marsh G., Sarber J., Amaral A., Bailey S., Lubicka D., Pham H., Miranda N., Ding J., Tang H.M., Ju H., Tranter P., Ji N., Krastel P., Jain R.K., Schumacher A.M., Loureiro J.J., George E., Berellini G., Ross N.T., Bushell S.M., Erdemli G., Solomon J.M. (2015) Englerin A Agonizes the TRPC4/C5 Cation Channels to Inhibit Tumor Cell Line Proliferation. PLoS One. 2015 Jun 22;10(6):e0127498.
155 Bezzerides V.J., Ramsey I.S., Kotecha S., Greka A., Clapham D.E. (2004) Rapid vesicular translocation and insertion of TRP channels. Nat Cell Biol. 2004 Aug;6(8):709-20. Epub 2004 Jul 18.
156 Yue D., Wang Y., Xiao J.Y., Wang P., Ren C.S. (2009) Expression of TRPC6 in benign and malignant human prostate tissues. Asian J Androl. 2009 Sep;11(5):541-7.
157 Cai R., Ding X., Zhou K., Shi Y., Ge R., Ren G., Jin Y., Wang Y. (2009) Blockade of TRPC6 channels induced G2/M phase arrest and suppressed growth in human gastric cancer cells. Int J Cancer. 2009 Nov 15;125(10):2281-7.
158 El Boustany C., Bidaux G., Enfissi A., Delcourt P., Prevarskaya N., Capiod T. (2008) Capacitative calcium entry and transient receptor potential canonical 6 expression control human hepatoma cell proliferation. Hepatology. 2008 Jun;47(6):2068-77.
159 Ding X, He Z, Zhou K, Cheng J, Yao H, Lu D, Cai R, Jin Y, Dong B, Xu Y, Wang Y. (2010) Essential role of TRPC6 channels in G2/M phase transition and development of human glioma. J Natl Cancer Inst. 2010 Jul 21;102(14):1052-68
160 Abkhezr M., Dryer S.E. (2014) Angiotensin II and canonical transient receptor potential-6 activation stimulate release of a signal transducer and activator of transcription 3-activating factor from mouse podocytes. Mol Pharmacol. 2014 Aug;86(2):150-8
161 Cai R., Ding X., Zhou K., Shi Y., Ge R., Ren G., Jin Y., Wang Y. (2009) Blockade of TRPC6 channels induced G2/M phase arrest and suppressed growth in human gastric cancer cells. Int J Cancer. 2009 Nov 15;125(10):2281-7.
162 Song R., Tian K., Wang W., Wang L. (2015) P53 suppresses cell proliferation, metastasis, and angiogenesis of osteosarcoma through inhibition of the PI3K/AKT/mTOR pathway. Int J Surg. 2015 Aug;20:80-7
163 Zhang H., Ding J., Fan Q., Liu S. (2009) TRPC6 up-regulation in Ang II-induced podocyte apoptosis might result from ERK activation and NF-kappaB translocation. Exp Biol Med (Maywood). 2009 Sep;234(9):1029-36.

LITERATURVERZEICHNIS
70
164 Nijenhuis T., Sloan A.J., Hoenderop J.G., Flesche J., van Goor H., Kistler A.D., Bakker M., Bindels R.J., de Boer R.A., Möller C.C., Hamming I., Navis G., Wetzels J.F., Berden J.H., Reiser J., Faul C., van der Vlag J. (2011) Angiotensin II contributes to podocyte injury by increasing TRPC6 expression via an NFAT-mediated positive feedback signaling pathway. Am J Pathol. 2011 Oct;179(4):1719-32.
165 Lautner R.Q., Villela D.C., Fraga-Silva R.A., Silva N., Verano-Braga T., Costa-Fraga F., Jankowski J., Jankowski V., Sousa F., Alzamora A., Soares E., Barbosa C., Kjeldsen F., Oliveira A., Braga J., Savergnini S., Maia G., Peluso A.B., Passos-Silva D., Ferreira A., Alves F., Martins A., Raizada M., Paula R., Motta-Santos D., Klempin F., Pimenta A., Alenina N., Sinisterra R., Bader M., Campagnole-Santos M.J., Santos R.A. (2013) Discovery and characterization of alamandine: a novel component of the renin-angiotensin system. Circ Res. 2013 Apr 12;112(8):1104-11.
166 Bader M., Alenina N., Andrade-Navarro M.A., Santos R.A. (2014) MAS and its related G protein-coupled receptors, Mrgprs. Pharmacol Rev. 2014 Oct;66(4):1080-105.
167 Tirupula K.C., Desnoyer R., Speth R.C., Karnik S.S. (2014) Atypical signaling and functional desensitization response of MAS receptor to peptide ligands. PLoS One. 2014 Jul 28;9(7):e103520.
168 Nijenhuis T., Sloan A.J., Hoenderop J.G., Flesche J., van Goor H., Kistler A.D., Bakker M., Bindels R.J., de Boer R.A., Möller C.C., Hamming I., Navis G., Wetzels J.F., Berden J.H., Reiser J., Faul C., van der Vlag J. (2011) Angiotensin II contributes to podocyte injury by increasing TRPC6 expression via an NFAT-mediated positive feedback signaling pathway. Am J Pathol. 2011 Oct;179(4):1719-32.
169 Daft P.G., Yang Y., Napierala D., Zayzafoon M. (2015) The growth and aggressive behavior of human osteosarcoma is regulated by a CaMKII-controlled autocrine VEGF signaling mechanism. PLoS One. 2015 Apr 10;10(4):e0121568.
170 Song H.J., Lee J.A., Han E., Lee E.K. (2015) Lifetime effectiveness of mifamurtide addition to chemotherapy in nonmetastatic and metastatic osteosarcoma: a Markov process model analysis. Tumour Biol. 2015 Sep;36(9):6773-9.
Recommended

















