
Biochimie - Curs 2 - 2019
Aminoacizii
Aminoacizii sunt compuși biochimici cu masă moleculară mică și importanţă
deosebită pentru organismul uman. Aceştia sunt elemente structurale de bază care intră în
componenţa proteinelor, compuşi cu proprietăți biologice remarcabile. Aminoacizii au
fost obţinuţi din proteine prin hidroliza acidă (HCl 6M) a legăturilor peptidice. De
asemenea, aminoacizii sunt precursori ai hormonilor (tiroxina) sau moleculelor cu funcţii
specializate (dopamina-neurotransmiţător). Primul aminoacid descoperit a fost asparagina
(în rădăcina şi tulpina sparanghelului; 1806). Ultimul aminoacid, treonina, a fost
descoperit în 1938. La început aminoacizii au fost denumiţi în funcţie de sursa de
provenienţă (asparagina din Asparagus, glutamatul din gluten şi tirozina din brânza -
“tyros”= brânză) sau după alte proprietăţi (glicina-“Glykos”- dulce).
Aminoacizii sunt prezenți în sânge sau alte fluide biologice atât în formă liberă
cât şi combinată. De asemenea, aminoacizii sunt precursori ai hormonilor, zaharurilor,
purinelor, pirimidinelor, porfirinelor, vitaminelor sau aminelor cu rol fiziologic.
Structura aminoacizilor
Structura aminoacizilor (ionizată) este ilustrată în figura 1. Proprietăţile chimice și
fizice ale aminoacizilor sunt dictate de grupările care sunt grefate pe atomul de carbon
din poziţia . La acest atom de carbon se disting următoarele grupări:
a) gruparea amino - grupare protonată (-NH3+) în condiţii fiziologice (pH 7);
b) gruparea carboxil - grupare deprotonată (-COO-) în condiţii fiziologice;
c) catena laterală - conferă proprietăţi specifice aminoacizilor, fiind responsabilă şi
de asimetria atomului de carbon din poziţia .
Până în prezent se cunosc 22 de aminoacizi care sunt elemente structurale de bază ale
proteinelor, dintre care numai 20 sunt criptate de către codul genetic universal. Ceilalți
doi, selenocisteina (abreviată Sec, descoperită în 1986) și pirolizina (abreviată Pyl,
descoperită în 2002, prezentă doar în componența proteinelor din bacterii sau arhee)
fuzionează cu ADN-ul de transfer care este capabil să formeze perechi de baze cu codonii
de stop ai ARN-ului mesager în timpul procesului de translație. Doar în această situație
acești doi aminoacizi pot fi încorporați în proteine. Configuraţia celor 21 de aminoacizi
existenţi în proteinele umane (excepţie face aminoacidul glicina) este L (Figura 1,
structura generală ionizată a unui aminoacid la pH neutru).
Figura 1. Configurația L- și D- a aminoacizilor
Deși în procesul de translație (copierea informației de pe ARN-ul mesager sub
formă de secvență proteică) se utilizează doar 21 de aminoacizi se cunosc peste 140 de
aminoacizi care apar în proteine. Este important de subliniat faptul că numărul
aminoacizilor care nu apar în proteine (neproteinogeni) este cu mult mai ridicat. Unii
dintre aceştia apar numai în plante sau în anumite microorganisme. În natură aminoacizii

Biochimie - Curs 2 - 2019
există şi în configuraţia D. De exemplu, D-alanina, acizii D-glutamic şi D-aspartic intră
în componenţa peptidelor din peretele bacterian. Fluidul intracelular al unor viermi
marini şi nevertebrate conţine drept componentă principală D-aminoacizii, iar în unele
crustacee marine cantitatea acestora poate depăşi 1%. De asemenea, acidul D-aspartic, D-
asparagina, acidul D-glutamic, D-glutamina, D-serina și D-alanina au fost detectați în
majoritatea plantelor, în timp ce D-prolina, D-valina, D-leucina and D-lizina doar în
unele plante (angiosperme dicotiledonate). Proteinele metabolice stabile din mamifere
conţin cantităţi însemnate de acid D-aspartic rezultat în urma procesului de racemizare:
concentraţia acidului D-aspartic în materia albă creierului atinge circa 3%, iar în proteina
bazică din măduva spinării până la 10%.
Laptele, carnea şi diversele cereale nu conţin cantităţi substanţiale de D-
aminoacizi. În decursul procesării, aminocizii din aceste alimente suferă adesea
fenomenul de racemizare. Mai mult, concentrația D-aminoacizilor din probele de lapte
(bio) creşte substanţial în decursul stocării la 4 oC. Astfel, se recomandă determinarea
conţinutului de D-alanină pentru a monitoriza eventualele contaminări ale laptelui cu
bacterii.
Concentrația aminoacizilor în sângele total este de 5-8 mg/ml. S-a constatat o
creștere a concentrației aminoacizilor în sânge în cazul afecțiunile acute ale
parenchimului hepatic, icterelelor, cirozelor, leucemiilor, nefritelor sau intoxicațiilor cu
arsen sau fosfor. Variațiile patologice ale acestora sunt corelate în multe situații cu erorile
metabolice înnăscute, încadrându-se în sindroamele cu severitate ridicată și pot fi asociate
în unele cazuri cu debilitate mintală. Hiperaminoacidimiile se produc prin inhibiția
sistemului de transport tubular (în care nivelul plasmatic al aminoacizilor rămâne
neschimbat) sau datorită unui deficit enzimatic strict localizat. O creștere a nivelului total
de aminoacizi din sânge poate fi pusă pe seama: eclapsiei (afecțiune care survine, în
general, la sfârșitul perioadei de sarcină, caracterizată prin convulsii asociate cu
hipertensiune arterială), intoleranței la fructoză, cetoacidozei, insuficiențelor renale sau
instalării sindromului Reye (care apare la copii cu vârstele între 4-12 ani cărora le-a fost
administrată aspirină; se manifestă prin meningită, encefalită, disfuncții ale ficatului).
Scăderea concentrației totale a aminoacizilor din sânge poate fi datorată: unei hiperfuncții
cortico-suprarenaliane, bolii Hartnup (boală autozomală recesivă în care nivelele alaninei,
valinei, treoninei, leucinei, fenil alaninei, tirozinei, histidinei sau glicinei sunt mai
scăzute; de asemenea scăderea concentrației de triptofan pot conduce la deficiențe ale
vitaminei B3 care se manifestă prin dermatite), malnutriție, sindrom nefrotic, boala
Huntington (afecțiune neurodegenerativă rară, moștenită genetic; apar disfuncţiile motorii
tipice - mişcări spasmodice şi involuntare ale braţelor, picioarelor, toracelui şi feţei;
simptomele timpurii pot fi și pierderea motivaţiei şi o modificare a stării emoţionale
depresie), artrită reumatoidă sau febră Phlebotomus (provocată de musca de nisip
Phlebotomus рарраtasif; simptomele sunt: instalarea oboselii, pierderea în greutate, febră
urmată de un focar de leziuni ale pielii).
Fenilcetonuria (idioția fenilpiruvică) este cauzată de un defect genetic al unei
enzime care catalizează transformarea fenilalaninei în tirozină. Drept rezultat, în fluxul
sangvin se acumulează o cantitate apreciabilă de fenilalanină și produșii acesteia de
metabolizare (acid fenilpiruvic, acid fenilacetic și acid fenillactic). Concentrația ridicată
de fenilalanină inhibă o altă enzimă, fapt care va induce diminuarea sintezei melaninei
(un pigment de la nivelul pielii, părului, țesutului pigmentat care stă la baza irisului

Biochimie - Curs 2 - 2019
ochiului și peretelui de dincolo de urechea internă). Drept urmare bolnavii de
fenilcetonurie au următoarele trăsături: tenul deschis, părul blond și ochi albaștri.
Majoritatea aminoacizilor care intră în componența proteinelor se regăsesc în
urină, sub formă liberă. Conținutul zilnic de aminoacizi totali din urină este de 1-3 g.
Aminoacizii cei mai reprezentativi sunt: glicina, histidina și derivații metilați (1-metil-
histidina și 3-metil-histidina), glutamina, serina, alanina, acidul -amino-butiric. În mod
frecvent în urină se găsesc glicina, tirozina, leucina sau histidina. Zilnic se elimină
aproximativ 210 mg histidină, 140 mg glicină și 100 mg taurină. Un aport proteic
alimentar ridicat determină o creștere a cantității de histidină, treonină, lizină sau tirozină.
Hiperaciduriile renale pot fi de mai multe tipuri:
a) fiziologice: hiperacidurii globale, care apar îndeosebi la sugar și prematur;
datorate sistemelor enzimatice de transport deficitare; hiperaciduria și
histinuria ce apar în săptămâna a 5-a de sarcină;
b) patologice – ce depind de mecanismele fiziopatologice:
o aminoacidurii renale, prin tulburarea reabsorbției tubulare, cu nivel
sangvin normal;
o aminoacidurii prerenale cu nivel crescut de aminoacizi atât la nivel
sangvin cât și în urină.
Hiperaciduriile renale se caracterizează prin alterarea absorbției unui singur
aminoacid (glicocoluria) sau a unui grup de aminoacizi (cistina, lizina, arginina și
ornitina-cistinuria; acidul glutamic, acidul aspartic - aminoaciduria dicarboxilică;
metionina, tirozina, fenilalanina, acidul -hidroxibutiric - malabsorbţia metioninei).
Boala urinei cu miros de sirop de arțar (MSUD- eng. Maple Syrup Urine
Disease) este o boală cauzată de un deficit al enzimei care catalizează decarboxilarea
oxidativă a aminoacizilor cu catena ramificată (valină, leucină și izoleucină) atât în sânge
cât și în urină alături de metaboliți (cetoacizii derivați din aceștia) imprimând urinei un
miros de sirop de arțar. Deficitul în metabolizarea acestor aminoacizi induce apariția unor
leziuni nervoase, întârzierea dezvoltării mentale sau în cazul formelor severe chiar la
deces. În condițiile în care boala este depistată în primele luni de viață se impune o
alimentație săracă în valină, leucină și izoleucină, iar șansele de redresare a pacientului
sunt ridicate.
Concentrația aminoacizilor din lichidul cefalorahidian (LCR) este mai scăzută
decât cea plasmatică. LCR conține majoritatea aminoacizilor prezenți în plasmă și țesutul
nervos (cu excepția 1- și 3-metil histidinei respectiv a cisteinei). Determinarea nivelului
aminoacizilor poate furniza informații importante legate de diferențierea meningitei
tuberculoase de cea aseptică. Dacă în ambele tipuri de meningită nivelul aminoacizilor
aspartic, glutamic, GABA (acid -aminobutiric), glicinei, triptofanului sunt ridicate, în
cazul meningitei tuberculoase scade nivelul de taurină și crește concentrația fenilalaninei
din LCR.
Determinarea aminoacizilor în laboratorul clinic se efectuează prin metoda
Sorensen sau metode fotometrice (cu acid 1,2-naftachinon-4-sulfonic) sau fluorimetrice
(în prezență de tioli și o-dibenzaldehidă). În cazul separării aminoacizilor prin
cromatografia în strat subțire sau HPLC (cromatografie de lichide de înaltă performanță)
aminoacizii pot fi detectați prin reacția cu ninhidrina (colorație violet – purpura lui
Ruhemann, doar prolina conduce la un produs colorat în galben sau verde-în funcție de
pH) sau izatina (colorație albastră intensă cu prolina, roșie cu valina și roz cu alanina).

Biochimie - Curs 2 - 2019
Clasificarea aminoacizilor
Aminoacizii pot fi clasificaţi după următoarele criterii:
- proprietăţile chimice;
- utilitatea acestora în biosinteza proteinelor;
- necesitatea biosintezei acestora pentru organism;
- implicarea lor ca substrat în procesul de gluconeogeneză.
Catena laterală determină diferenţele dintre proprietăţile chimice ale
aminoacizilor
În funcţie de natura catenei laterale se disting:
a) catene laterale alifatice sau aromatice nepolare:
- aminoacizii alifatici: glicina, alanina, valina, leucina, izoleucina, metionina;
Glicina este aminoacidul cu structura cea mai simplă, având în catena laterală un
atom de hidrogen. Drept rezultat glicina este singurul aminoacid care nu este chiral (pe
atomul de carbon din poziția sunt grefați doi atomi de hidrogen) și este întâlnit de
obicei la suprafața proteinelor datorită flexibilității pe care o conferă lanțului proteic. De
asemenea, glicina este un aminoacid abundent în proteinele fibrilare. Glicina are funcție
de neurotransmițător și este intermediar metabolic în cadrul biosintezei bazelor azotate
din acizii nucleici.
Alanina, valina, leucina şi izoleucina posedă catene laterale hidrocarbonate (de la
gruparea metil din alanină la grupările butiril izomere din izoleucină şi leucină). Ultimii
trei aminoacizi constituie o sursă de energie în cazul suprasolicitării musculare. Acești
aminoacizi hidrofobi sunt localizați în special în interiorul structurii 3D a proteinelor.
Alanina este unul dintre aminoacizii care se găsesc în proteine, frecvența de
apariție în aceste macromolecule fiind depășită doar de leucină. Există o controversă
legată de faptul că leucina este un aminoacid care stimulează direct sinteza proteinelor în
mușchi. Mai mult excedentul de leucină poate fi periculos și poate induce diareea,
apariția dermatitelor, instalarea demenței și uneori decesul. Izoleucina posedă un centru
chiral suplimentar pe catena laterală.
Metionina este un aminoacid nepolar care intervine (sub formă formilată; fiind
inserat primul în lanțul proteic) în cadrul biosintezei proteinelor din bacterii, mitocondrii

Biochimie - Curs 2 - 2019
și cloroplaste. Este codat de codonul de start AUG localizat pe ARN-ul mesager.
Metionina este un intermediar în biosinteza carnitinei, taurinei sau fosfatidilcolinei.
Metionina este un precursor al S-adenozil-metioninei (SAM), o moleculă
importantă în metabolismul celular al mamiferelor. Acest compus este folosit în reacțiile
de metilare (de transfer al grupării –CH3).
- aromatici: fenilalanina, tirozina şi triptofanul;
Resturile aromatice (fenilalanina, tirozina şi triptofanul) pot fi detectate prin
spectroscopie în domeniul UV (250-300 nm). Această proprietate este folosită în special
pentru estimarea concentraţiei peptidelor sau proteinelor care conţin aceste resturi sau
pentru studiul împachetării acestor macromolecule.
Fenilalanina este un aminoacid nepolar și un precursor metabolic al tirozinei.
Boala genetică numită fenilcetonurie are drept consecință metabolizarea defectuoasă a
fenilalaninei.
Triptofanul este un aminoacid esențial care conține un rest indolic. Acest
aminoacid este precursor an serotoninei, niacinei și hormonului auxină (în plante).
Tirozina este un aminoacid neesențial care poate fi fosforilat în proteine în cadrul
proceselor de semnalizare. În celulele dopaminergice ale creierului o enzimă (tirozin
hidroxilaza) convertește tirozina la L-DOPA, un intermediar precursor al dopaminei. Mai
mult, dopamina este un precursor al norepinefrinei, epinefrinei, hormonilor tiroidieni și
melaninei.
Fluorescenţa proteinelor (polimeri care au drept unităţi structurale aminoacizii)
este în general obţinută prin excitarea aminoacizilor aromatici la lungimi de undă mai
mari de 280 nm (dat fiind faptul că fluorescenţa fenil alaninei este mai mică şi maximul
de absorbţie al acestui aminoacid este în jurul valorii de 260 nm se preferă excitarea la
valori mai mari ale absorbanţei). Proteinele absorb la 280 nm datorită triptofanului şi
tirozinei. În schimb, la lungimi de undă mai mari de 295 nm, absorbţia este atribuită în
special triptofanului. Astfel, triptofanul trebuie excitat la 295-305 nm pentru a emite
specific la lungimi de undă mai mari. Maximele de emisie ale tirozinei şi triptofanului
(aminoacizi individuali) în soluţie apoasă sunt la lungimile de undă de 303 respectiv 348
nm (triptofanul deşi puţin abundent în proteine are proprietăţi fluorescente atribuite
nucleului indolic).
- heterociclici: prolina (singurul aminoacid care are o grupare amino secundară),
triptofanul și histidina (vezi catene laterale încărcate pozitiv)

Biochimie - Curs 2 - 2019
Prolina este cel mai puțin flexibil aminoacid dintre prezenți în proteine. Prezența
prolinei afectează structura secundară (desface elementele secundare de tip helix și β
pliat). Hidroxilarea prolinelor din colagen (din oase, tendoane sau ligamente) imprimă o
stabilitate conformațională acestei proteine.
b) catene laterale neîncărcate (serina, treonina, asparagina, glutamina, cisteina,
selenocisteina). Grupările -OH din catena laterală a aminoacizilor (serina,
treonina – aminoacizi alifatici polari, tirozina – aminoacid aromatic – grupare
fenolică) pot fi implicate în legături de hidrogen.
Serina este un precursor al multor compuși celulari: purinelor, pirimidinelor,
sfingolipidelor, folatului și a unor aminoacizilor (glicină, cisteină și triptofan). Gruparea
hidroxilică a serinei din proteine este țintă pentru fosforilare prin intermediul kinazelor.
Treonin kinazele sunt parte integrantă a sistemelor de semnalizare celulară.
Serina face parte din triada catalitică a serin proteazelor (tripsină, chimotripsină).
În proteine, atât pe treonina cât și pe serină, pot fi grefate resturi de zaharuri
(reacția de glicozilare la gruparea –OH are loc în aparatul Golgi).
Pe catena laterală mai pot fi prezente şi alte grupări: -SH (cisteina), -SeH (seleno-
cisteina), tioeterică (metionina) sau amidică (glutamina, asparagina).
Cisteina este singurul aminoacid care are o grupare tiolică în catena laterală. Este
neesențial pentru adulți dar esențial pentru bebeluși. Grupările tiolice din două molecule
de cisteină poate fi oxidate la una disulfidică. În organismul uman cisteina poate fi
biosintetizată din metionină. În plus, acest aminoacid este o componentă a proteinelor,
peptidelor (glutation). De asemenea, poate regla activitatea unor enzime (de exemplu din
cadrul biosintezei melaninei).

Biochimie - Curs 2 - 2019
Selenocisteina este un aminoacid codat de tripletul UGA (care este şi tripletul de
oprire în sinteza proteică). Acest aminoacid este mult mai reactiv decât cisteina.
Înlocuirea atomului de S cu Se în unele enzime (catalizatori biochimici care sunt
implicaţi în unele reacţii redox) determină o creştere semnificativă (cu peste 100 de ori) a
vitezei reacţiei catalizate de acestea.
Asparagina este un aminoacid neesențial. Gruparea amidică din catena laterală
imprimă un caracter polar acestui aminoacid. Asparagina este implicată în formarea
acrilamidei în cazul alimentelor gătite la temperaturi ridicate (când reacționează cu
grupările carbonilice). De asemenea, asparagina este convertită în acid malic în cadrul
ciclului acidului citric.
Glutamina care nu este în mod normal esențial, dar în anumite situații poate fi
necesar pentru atleții care depun eforturi intensive sau pentru persoanele care sunt
afectate de boli gastro-intestinale. Glutamina este cel mai abundent aminoacid din sânge
(respectiv mușchii scheletici) și unul dintre puținii care poate traversa bariera hemato-
encefalică. Acest aminoacid susține creșterea și multiplicarea celulelor sistemului
imunitar. În reticulul endoplasmatic acest aminoacid poate fi glicozilat (la gruparea
amidică din catena laterală).
c) catene laterale încărcate:
- pozitiv (lizina, arginina, histidina-imidazolul are un pKa de aproximativ 6 în
consecinţă heterociclul din catena laterală este deprotonat, iar nucleul imidazolic încărcat
negativ, la valori ale pH-ului mai mari decât 7-8);

Biochimie - Curs 2 - 2019
Lizina asistă absorbția calciului din tractul gastro-intestinal promovând astfel
biosinteza colagenului. Histamina și carnozina (substanță ce anihilează radicalii liberi)
sunt biosintetizate pornind de la acest aminoacid. Lizina este un aminoacid esențial care
poate fi modificat postranslațional (după biosinteza proteică) pentru a forma acetilizină,
hidroxilizină și metillizină. De asemenea, acest aminoacid poate fi ubichitinat (legat de o
proteină de dimensiuni relativ mici numită ubichitină), biotinilat (legat de o vitamină),
carboxilat sau cuplat cu alte proteine (“sumoilare”, “pupilare”).
Arginina este un aminoacid care poate fi esențial sau neesențial. Copii născuți
prematur nu pot sintetiza arginina. În plus, traumele chirurgicale, septicemiile și arsurile
induc o nevoie crescută de arginină. Acest aminoacid conține o grupare complexă numită
guanidinică a cărei constantă pKa este de peste 12, fapt care îi conferă o încărcare
pozitivă la pH-ul celular. Arginina induce dilatarea vaselor de sânge (reduce presiunea
sangvină) și producerea de monoxid de azot (NO). Acest aminoacid joacă un rol
important în balanța azotului.
- negativ (grupările carboxilice aflate pe catena laterală a resturilor aspartat şi
glutamat au valori ale pKa-ului de 3,9 şi 4,3; sunt foarte polare – încărcate negativ în
condiţii fiziologice). Aceste grupări sunt implicate în interacţii electrostatice cu grupări
încărcate pozitiv, punţi de hidrogen etc.
Acidul aspartic este un aminoacid neesențial
(produs prin transaminarea oxaloacetatului).
Acidul glutamic este un aminoacid neesențial
(produs prin transaminarea cetoglutaratului)
care are rol și de neurotransmițător.
Ambii aminoacizi constituie o sursă
energetică rapidă.
Nou-născuții preferă gustul de acid glutamic,
aminoacid care se află în concentrație mare
în laptele matern.
Aminoacizii cu catene laterale alifatice sau aromatice nepolare cu excepţia glicinei sunt
hidrofobi, iar cei care posedă catene laterale încărcate sunt hidrofili. Aminoacizii polari
care au catene laterale neîncărcate au încărcări intermediare (atât hidrofobi cât şi
hidrofili).
În funcţie de aminoacizii care intră în compoziţia proteinelor se disting:
a) aminoacizi proteinogeni-aminoacizii care intră în compoziţia proteinelor (vezi
exemplele de mai sus);
b) aminoacizi neproteinogeni-aminoacizii care nu intră în compoziţia proteinelor:
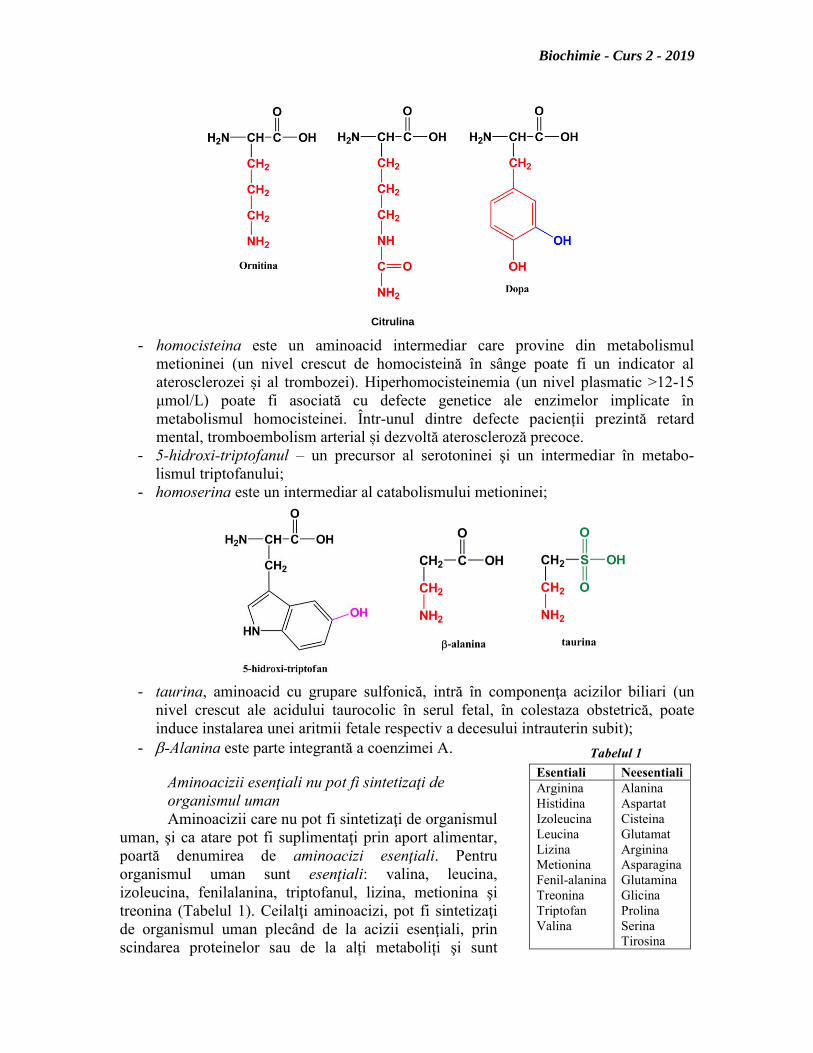
- ornitina (un precursor al citrulinei în mitocondrii) şi citrulina (un precursor al
argininei în citozol; în această reacție se produc și oxizi de azot – molecule semnal
în sistemul cardiovascular, în neuroni sau în sistemul imunitar) apar în decursul
ciclului ureei. Sinteza și eliminarea ureei este esențială pentru eliminarea
metaboliților toxici cu azot (concentrația mare de amoniac poate conduce la
encefalopatie hepatică; alcaloza poate induce apariția unor boli la nivelul unor
organe) din organism;
Biochimie - Curs 2 - 2019
Esentiali Neesentiali
Arginina
Histidina
Izoleucina
Leucina
Lizina
Metionina
Fenil-alanina
Treonina
Triptofan
Valina
Alanina
Aspartat
Cisteina
Glutamat
Arginina
Asparagina
Glutamina
Glicina
Prolina
Serina
Tirosina
Citrulina
- homocisteina este un aminoacid intermediar care provine din metabolismul
metioninei (un nivel crescut de homocisteină în sânge poate fi un indicator al
aterosclerozei și al trombozei). Hiperhomocisteinemia (un nivel plasmatic >12-15
μmol/L) poate fi asociată cu defecte genetice ale enzimelor implicate în
metabolismul homocisteinei. Într-unul dintre defecte pacienții prezintă retard
mental, tromboembolism arterial și dezvoltă ateroscleroză precoce.
- 5-hidroxi-triptofanul – un precursor al serotoninei şi un intermediar în metabo-
lismul triptofanului;
- homoserina este un intermediar al catabolismului metioninei;
- taurina, aminoacid cu grupare sulfonică, intră în componenţa acizilor biliari (un
nivel crescut ale acidului taurocolic în serul fetal, în colestaza obstetrică, poate
induce instalarea unei aritmii fetale respectiv a decesului intrauterin subit);
- -Alanina este parte integrantă a coenzimei A.
Aminoacizii esenţiali nu pot fi sintetizaţi de
organismul uman
Aminoacizii care nu pot fi sintetizaţi de organismul
uman, şi ca atare pot fi suplimentaţi prin aport alimentar,
poartă denumirea de aminoacizi esenţiali. Pentru
organismul uman sunt esenţiali: valina, leucina,
izoleucina, fenilalanina, triptofanul, lizina, metionina şi
treonina (Tabelul 1). Ceilalţi aminoacizi, pot fi sintetizaţi
de organismul uman plecând de la acizii esenţiali, prin
scindarea proteinelor sau de la alți metaboliți şi sunt
Tabelul 1

Biochimie - Curs 2 - 2019
denumiţi neesenţiali. De exemplu, alanina este sintetizată în organism plecând de la
piruvat și din acest motiv este considerat un aminoacid neesențial.
Aminoacizii esenţiali se găsesc atât în proteinele de origine vegetală cât şi în cele
de origine animală. Compoziţia în aminoacizi diferă de la o proteină la alta. Clasificarea
în aminoacizi esenţiali şi neesenţiali este relativă. Aminoacizii esențiali pot varia de la un
organism la altul, pot fi determinați de vârsta etc. Aminoacizii care sunt în mod normal
neesențiali pot fi necesari în anumite situații. Indivizii care nu sintetizează cantități
suficiente de arginină, cisteină, glutamină, prolină, selenocisteină, serină și tirozină,
datorită unor maladii, au nevoie de suplimente alimentare care conțin acești aminoacizi.
Din scheletul atomilor de carbon al unor aminoacizi poate fi sintetizată glucoza
Scheletul hidrocarbonat al unor aminoacizi poate fi utilizat la sinteza glucozei.
Aceşti aminoacizi (cu excepţia leucinei şi lizinei) se numesc aminoacizi glucogeni, iar
restul aminoacizi cetogeni deoarece sunt transformaţi în corpii cetonici (excretați în
urină).
Proprietăţile acido-bazice ale aminoacizilor
Aminoacizii sunt amfoliţi
Aminoacizii pot ceda sau accepta protoni şi ca atare pot să se comporte asemeni
acizilor şi bazelor (amfolit).
Gruparea amino poate fi protonată:
Valoarea constantei de aciditate, pKa, a acestei reacţii este de 9-10,5 (în funcţie de catena
laterală a aminoacidului). În majoritatea condiţiilor fiziologice de pH gruparea amino este
protonată, dar gruparea carboxil din aminoacizi
este deprotonată:
Valoarea constantei de aciditate, pKa, pentru reacţia de deprotonare a grupării carboxil
este de 1,7-2,4. Acest fapt dictează o încărcare negativă a acestei grupări la valori
fiziologice ale pH-ului.
Proprietăţile acido-bazice ale grupărilor din catena laterală
De asemenea, grupările disociabile din catena laterală a aminoacizilor au valori
specifice ale pKa-ului şi pot contribui la proprietatea de amfolit a acestora. O importanţă
deosebită o are nucleul imidazolic din catena laterală a histidinei (cu un pKa = 6). Din
acest motiv unele resturi de histidină din enzime iau parte la diverse reacţii (fixarea
oxigenului în hemoglobină, proteazele posedă 3 resturi catalitice în situsul activ – cisteină
sau serină, histidină și acid glutamic). Mai mult, biologia moleculară modernă permite
producerea artificială a unor proteine care posedă la unul din capetele lanțului proteic o
secvență de 6-10 histidine (6-10 nuclee imidazolice), proprietate care permite izolarea
prin cromatografie de afinitate a acestora (prin interacțiune cu o matrice pe care sunt
grefați/complexați ionii metalici: Ni2+, Co2+ sau Zn2+).
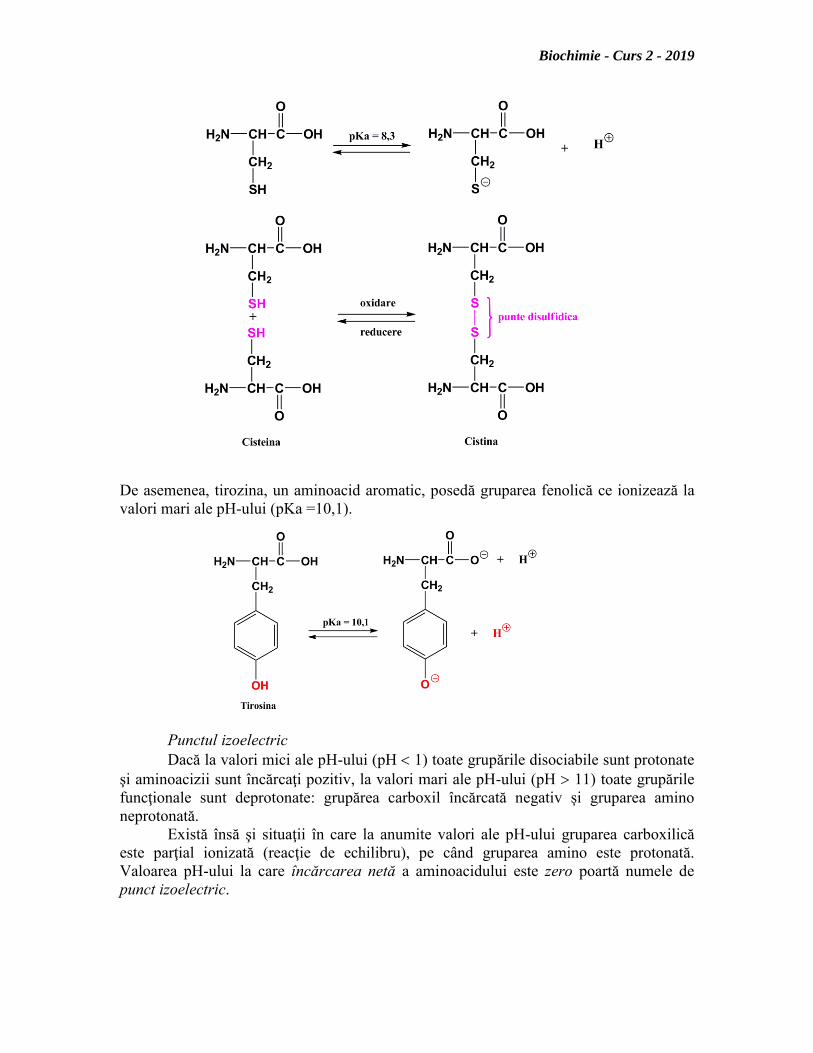
Cisteina ionizează la pH bazic (pKa = 8,3) sau poate fi oxidată la cistină, acest
fapt conferă resturilor de cisteină un rol deosebit în împachetarea tridimensională a
proteinelor.
Biochimie - Curs 2 - 2019
De asemenea, tirozina, un aminoacid aromatic, posedă gruparea fenolică ce ionizează la
valori mari ale pH-ului (pKa =10,1).
Punctul izoelectric
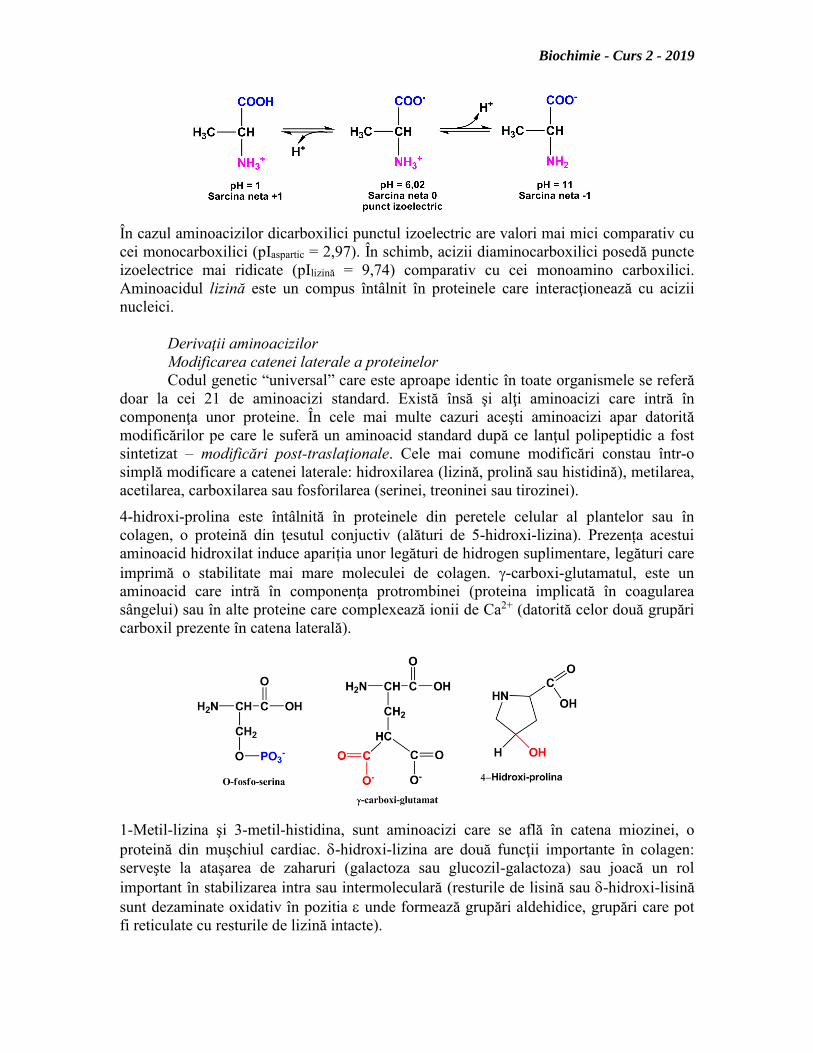
Dacă la valori mici ale pH-ului (pH 1) toate grupările disociabile sunt protonate
şi aminoacizii sunt încărcaţi pozitiv, la valori mari ale pH-ului (pH 11) toate grupările
funcţionale sunt deprotonate: grupărea carboxil încărcată negativ și gruparea amino
neprotonată.
Există însă şi situaţii în care la anumite valori ale pH-ului gruparea carboxilică
este parţial ionizată (reacţie de echilibru), pe când gruparea amino este protonată.
Valoarea pH-ului la care încărcarea netă a aminoacidului este zero poartă numele de
punct izoelectric.
Biochimie - Curs 2 - 2019
În cazul aminoacizilor dicarboxilici punctul izoelectric are valori mai mici comparativ cu
cei monocarboxilici (pIaspartic = 2,97). În schimb, acizii diaminocarboxilici posedă puncte
izoelectrice mai ridicate (pIlizină = 9,74) comparativ cu cei monoamino carboxilici.
Aminoacidul lizină este un compus întâlnit în proteinele care interacţionează cu acizii
nucleici.
Derivaţii aminoacizilor
Modificarea catenei laterale a proteinelor
Codul genetic “universal” care este aproape identic în toate organismele se referă
doar la cei 21 de aminoacizi standard. Există însă şi alţi aminoacizi care intră în
componenţa unor proteine. În cele mai multe cazuri aceşti aminoacizi apar datorită
modificărilor pe care le suferă un aminoacid standard după ce lanţul polipeptidic a fost
sintetizat – modificări post-traslaţionale. Cele mai comune modificări constau într-o
simplă modificare a catenei laterale: hidroxilarea (lizină, prolină sau histidină), metilarea,
acetilarea, carboxilarea sau fosforilarea (serinei, treoninei sau tirozinei).
4-hidroxi-prolina este întâlnită în proteinele din peretele celular al plantelor sau în
colagen, o proteină din ţesutul conjuctiv (alături de 5-hidroxi-lizina). Prezența acestui
aminoacid hidroxilat induce apariția unor legături de hidrogen suplimentare, legături care
imprimă o stabilitate mai mare moleculei de colagen. -carboxi-glutamatul, este un
aminoacid care intră în componenţa protrombinei (proteina implicată în coagularea
sângelui) sau în alte proteine care complexează ionii de Ca2+ (datorită celor două grupări
carboxil prezente în catena laterală).
1-Metil-lizina şi 3-metil-histidina, sunt aminoacizi care se află în catena miozinei, o
proteină din muşchiul cardiac. -hidroxi-lizina are două funcţii importante în colagen:
serveşte la ataşarea de zaharuri (galactoza sau glucozil-galactoza) sau joacă un rol
important în stabilizarea intra sau intermoleculară (resturile de lisină sau -hidroxi-lisină
sunt dezaminate oxidativ în pozitia unde formează grupări aldehidice, grupări care pot
fi reticulate cu resturile de lizină intacte).

Biochimie - Curs 2 - 2019
De asemenea, pe catena laterală a aminoacizilor din componenţa proteinelor pot fi ataşate
grupări voluminoase (lipide, carbohidraţi – în cazul glicoproteinelor acestea pot avea
resturile de zahar atașate de catena laterală a asparaginei, treoninei sau serinei). Grupările
N- sau C- terminale din lanţurile polipeptidice/proteice pot fi modificate chimic. Aceste
modificări sunt importante pentru funcţia proteinelor.
Aminoacizii şi derivaţii biologic activi ai acestora
Cei 20 de aminoacizi standard suferă adesea transformări chimice. Aceste
transformări au drept rezultat obţinerea altor aminoacizi sau a unor compuşi înrudiţi şi
sunt parte integrantă a transformărilor care au loc în sinteza celulară sau în procesele de
degradare. De asemenea, aminocizii pot fi utilizaţi la: transportul azotului sub forma
grupelor amino, sursa de energie (în urma oxidării) sau la comunicarea dintre celule.
Alte roluri ale aminoacizilor şi derivaţilor acestora:
glicina, acidul -amino-butiric (AGAB, un produs rezultat prin decarboxilarea
acidului glutamic), dopamina (un compus rezultat prin decarboxilarea hidroxi-
tirozinei) sunt compuşi eliberaţi de celula nervoasă cu scopul de a modifica
comportamentul celulelor vecine (neurotransmiţători);
- antagoniştii dopaminei (medicamente care interferă cu dopamina la legarea
acestui compus la receptorii aferenţi) sunt utilizaţi pentru tratarea schizofreniei şi
a unor boli mentale similare. O persoană cu schizofrenie poate posedă un sistem
dopaminic hiperactiv. Din acest motiv folosirea antagoniştilor dopaminei pot ajuta
la reglarea acestui sistem prin reducerea activităţii dopaminei;
- cocaina şi alte medicamente în exces pot afecta funcţiile dopaminei (prin
stimularea sau blocarea receptorilor dopaminei).
histamina (produsul de decarboxilare al histidinei) este un potenţial mediator local
pentru reacţiile alergice, poate fi implicată în mecanismul de apărare împotriva
substanţelor exogene (sistemul imunitar) sau are funcţii de reglare a tractului
gastro-intestinal (controlează nivelul de acid gastric), a sistemului nervos central
cât şi a ritmului somn/trezire.
tiroxina (un alt derivat la tirozinei) este un hormon tiroidian (conţine iod) care
reduce presiunea sangvină, pulsul, temperatura sau greutatea vertebratelor.
Tiroxina se obține în urma reacției de iodurare a tirozinei urmată de condensarea
oxidativă a două molecule de diiodo-tirozină.

Biochimie - Curs 2 - 2019
Triiodo-tironina este un derivat cu funcții și structură asemănătoare tiroxinei
(hormon care posedă grefați doar trei atomi de iod în structură). Cei doi derivați
se leagă de proteina numită tiroglobulină. Acest complex poate fi scindat, iar
derivații pot interacționa cu proteinele plasmatice și astfel transportați la celulele
periferice, unde se leagă de ADN-ul celulei inducând o creștere a ratei
metabolismului zaharurilor și proteinelor.
Compuşi derivaţi de la aminoacizi care
posedă proprietăţi biologice
Recommended

















