Embed Size (px)
Citation preview

Xylans
Xiao Liu10/22/2010
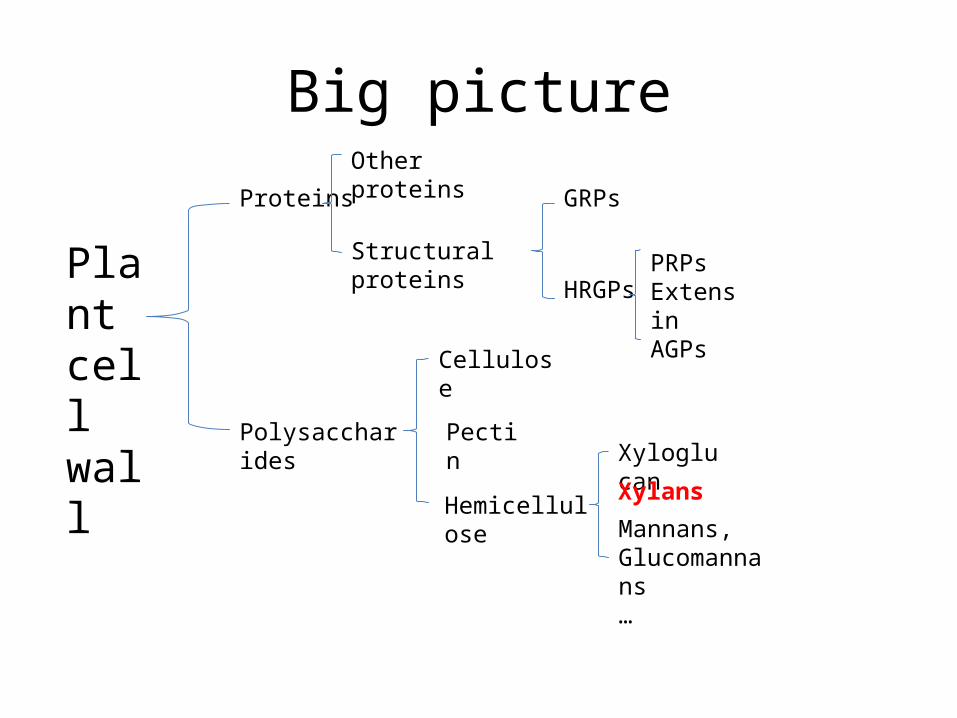
Big picture
Plant cell wall
Proteins
Polysaccharides
Other proteins
Structural proteins
GRPs
HRGPsPRPsExtensinAGPs
Cellulose
Pectin
Hemicellulose
Xyloglucan
Xylans
Mannans, Glucomannans…

How do xylans affect our well-being?
• Xylans can help reduce some diseases in humans• Xylans are important functional ingredients in
baked products.• Xylans impact brewing properties of grains.• Xylans can be converted to xylitol, a natural food
sweetener.• Xylans are major constituents in the nonnutritional
constituent of feed in monogastric animals.
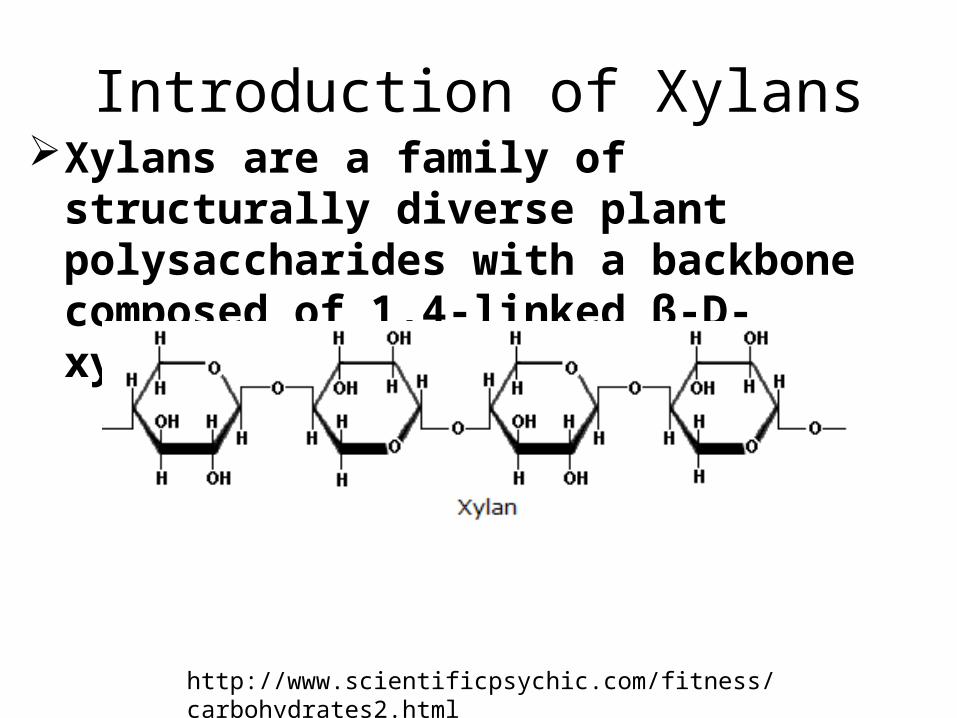
Introduction of XylansXylans are a family of structurally diverse plant
polysaccharides with a backbone composed of 1,4-linked β-D-xylosyl residues.
http://www.scientificpsychic.com/fitness/carbohydrates2.html

Major types of xylans
In almost all cases, the backbone is substituted with monosaccharide or disaccharide side chains, to varying degrees.
glucuronic acid and 4-O-methyl glucuronic acid (glucuronoxylan, GX),
arabinose (arabinoxylan, AX), a combination of acidic and neutral sugars
(glucuronoarabinoxylan, GAX).

Arabinoxylan• Arabinoxylan is the predominant
hemicellulose of grasses. • L-arabinofuranose attached randomly by
1α→2 and/or 1α→3 linkages to the xylose units throughout the chain.
• Side chains containing arabinosyl, galactosyl, glucosyluronic acid, and 4-O-methyl glucosyluronic acid residues have been identified.
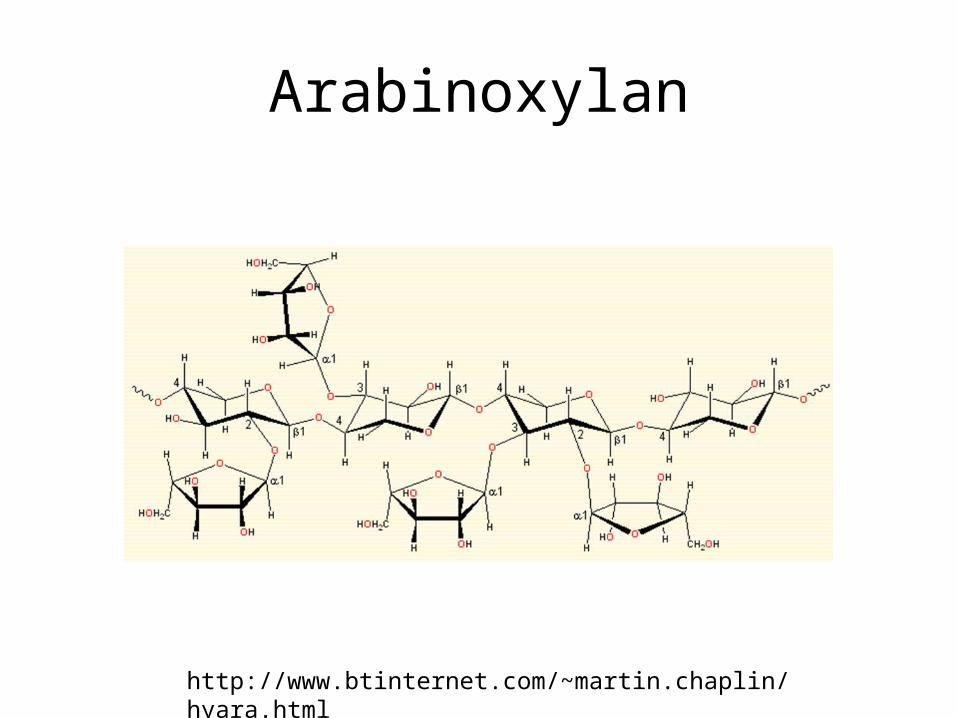
Arabinoxylan
http://www.btinternet.com/~martin.chaplin/hyara.html

Glucuronoxylan
Major components of the secondary cell walls of dicots (15%-30%):• α(1,2)-linked D-glucuronyl (GlcA) • 4-O-methyl-GlcA (MeGlcA) residues attached to
C-2 ~ every 10 Xyl residues• ~ 70% contain one O-acetyl group at C-2 or C-3.• Contain a distinct “glycosyl sequence” at the
reducing end.• Devoid of Ara units

Glucuronoxylan
William S York and Malcolm A O’Neill, Biochemical control of xylan biosynthesis – which end is up?
• Two-domain architecture: a typical polymer domain and a distinct “glycosyl sequence” domain:

Glucuronoarabinoxylan (GAX)
Ara units are added to the O-3 position of the xylosyl units of the backbone Feruloyl groups are esterified to the O-5 position of the Ara units in about every
50 units. (5’5 biphenyl bond forms cross-link between two ferulic acid residues) GlcAs are added to the O-2 position of the xylosyl units. (In all GAXs)
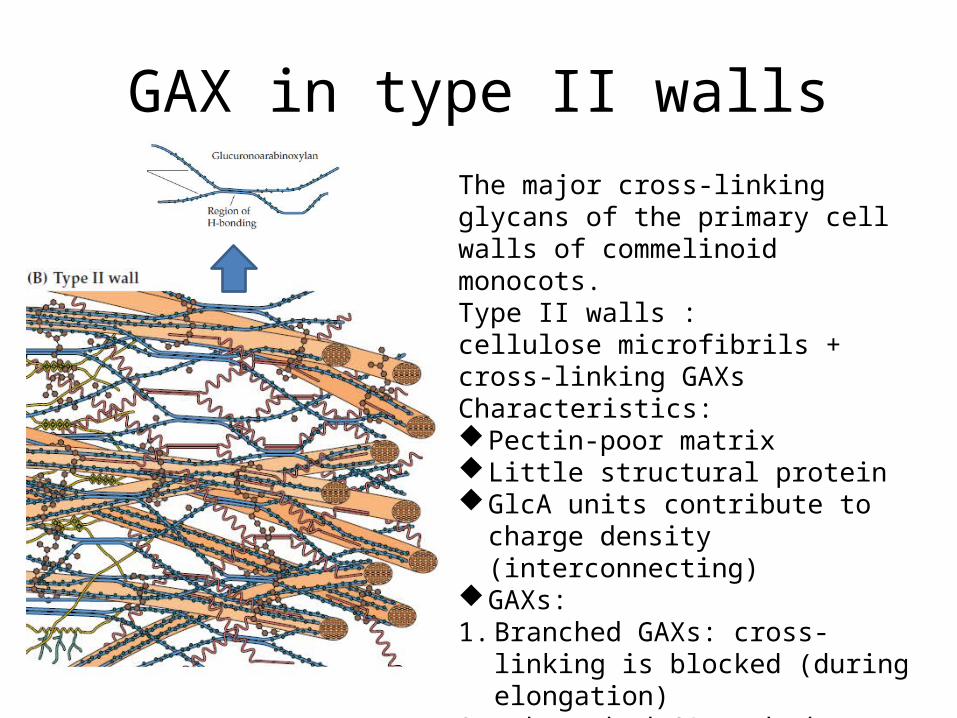
GAX in type II wallsThe major cross-linking glycans of the primary cell walls of commelinoid monocots.Type II walls :cellulose microfibrils + cross-linking GAXsCharacteristics:Pectin-poor matrixLittle structural proteinGlcA units contribute to charge density
(interconnecting)GAXs:1. Branched GAXs: cross-linking is blocked
(during elongation) 2. Unbranched GAXs: hydrogen-bond to
cellulose or to each other (after elongation)

Xylan biosynthesis• Synthesized in Golgi Apparatus.• Backbone synthesis: IRX9, IRX10, IRX10L, IRX14• Synthesis of side chain primer or terminator
oligosaccharide: IRX7, IRX8, PARVUS

GX biosynthesisServeral glycosyltransferases may be involved in the initiation and elongation of the polymer backbone; other enzymes for the addition and /or modification of the side chain.
Five genes (FRA8, IRX8, IRX9, PARVUS, IRX14) in Arabidopsis have been identified to be involved in GX synthesis. They encode putative GTs that may have a role in forming reducing end.
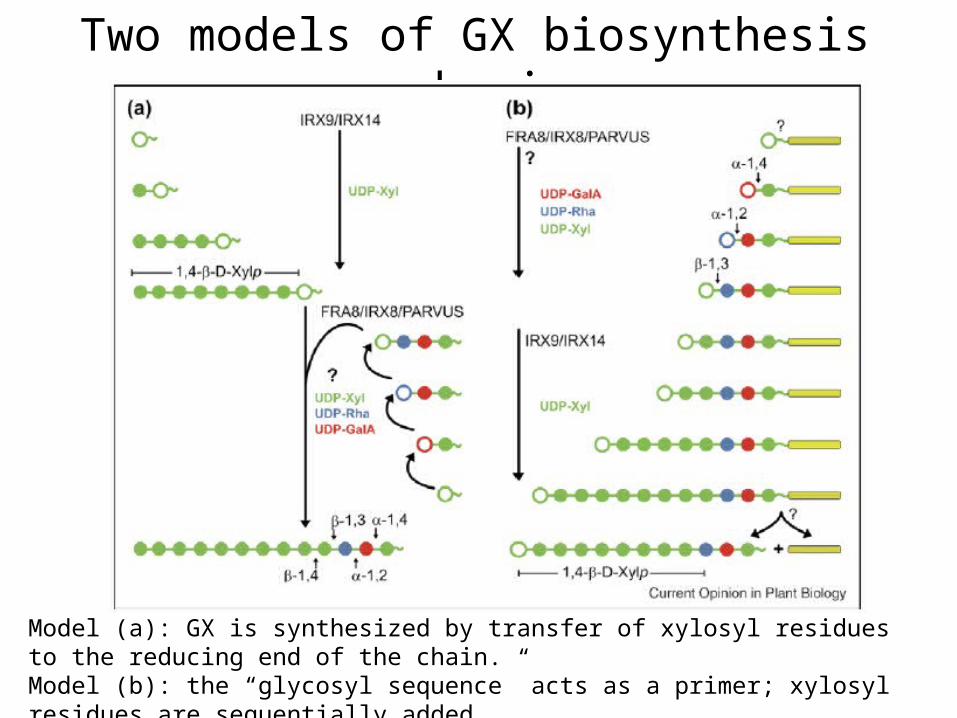
Two models of GX biosynthesis mechanism
Model (a): GX is synthesized by transfer of xylosyl residues to the reducing end of the chain.Model (b): the “glycosyl sequence” acts as a primer; xylosyl residues are sequentially added to the nonreducing end.

GAX biosynthesis• Newly synthesizsed GAX polymers have
regular structuresAfter treatment of endoxylanase III, only three kinds of oligosaccharides are released
Possible explanation of the formation of GAX fragments: two unbranched Xyl residues at the reducing end and one or two unbranced Xyl residue at the nonreducing end.
This type of regular structure is usually the result of a cooperative mechanism between enzymes:XylT, AraT, GlcAT.

AX biosynthesis
• Xylosyltransferase and arabinosyl transferase activities have been detected in microsomal fractions isolated from wheat and barley.

Why are we desired to know how Xylans are synthesized ?
• Contribution to the recalcitrance in biofuel production.
• In paper manufacture, decrease in brightness of final product
MeGlcA Hexenuronic Acid • In animal feed, loss of nutrition

Absence of branches from xylan in Arabidopsis gux mutants reveals
potential for simplification of lignocellulosic biomass
• Jennifer C. Mortimer, Godfrey P. Miles, David M. Brown, Zhinong Zhang, Marcelo P. Segura, Thilo Weimar, Xiaolan Yu, Keith A. Seffeen, Elaine Stephen, Simon R. Turner, and Paul Dupree
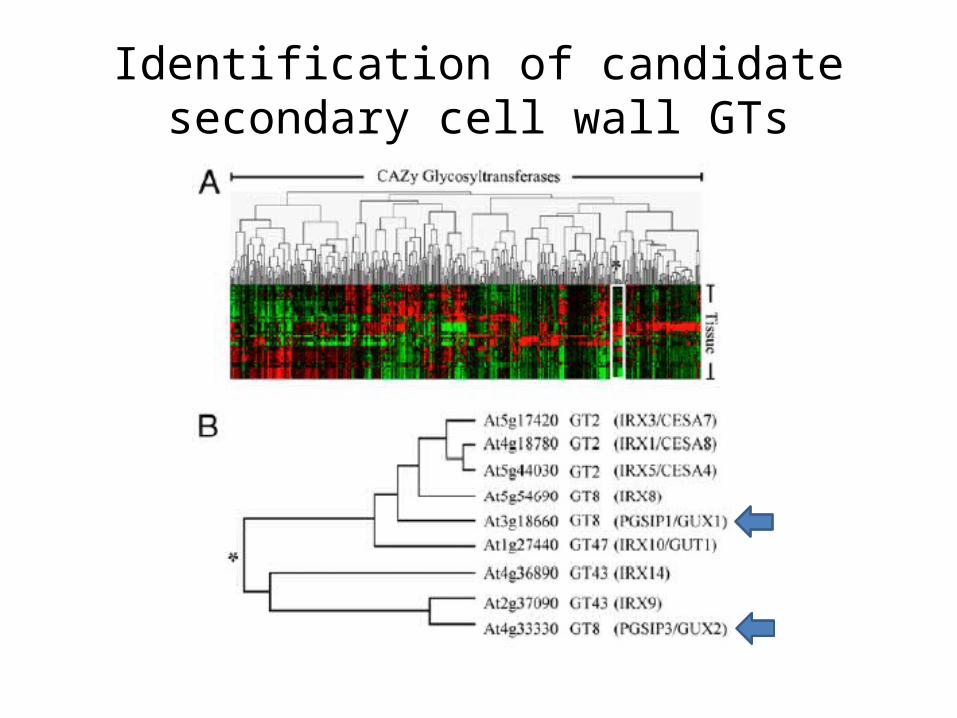
Identification of candidate secondary cell wall GTs
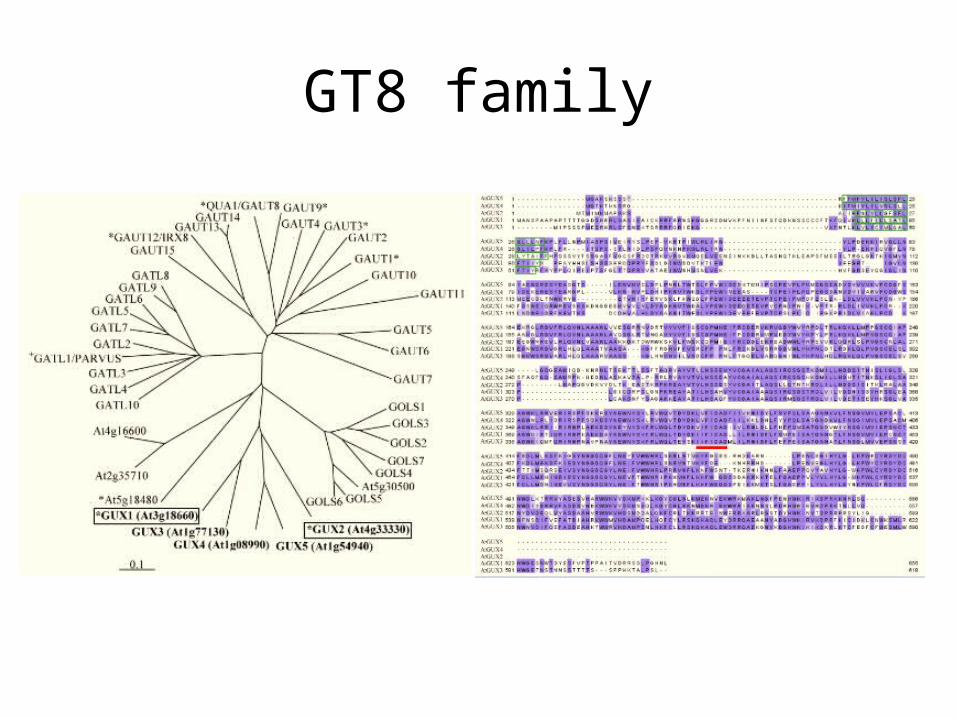
GT8 family
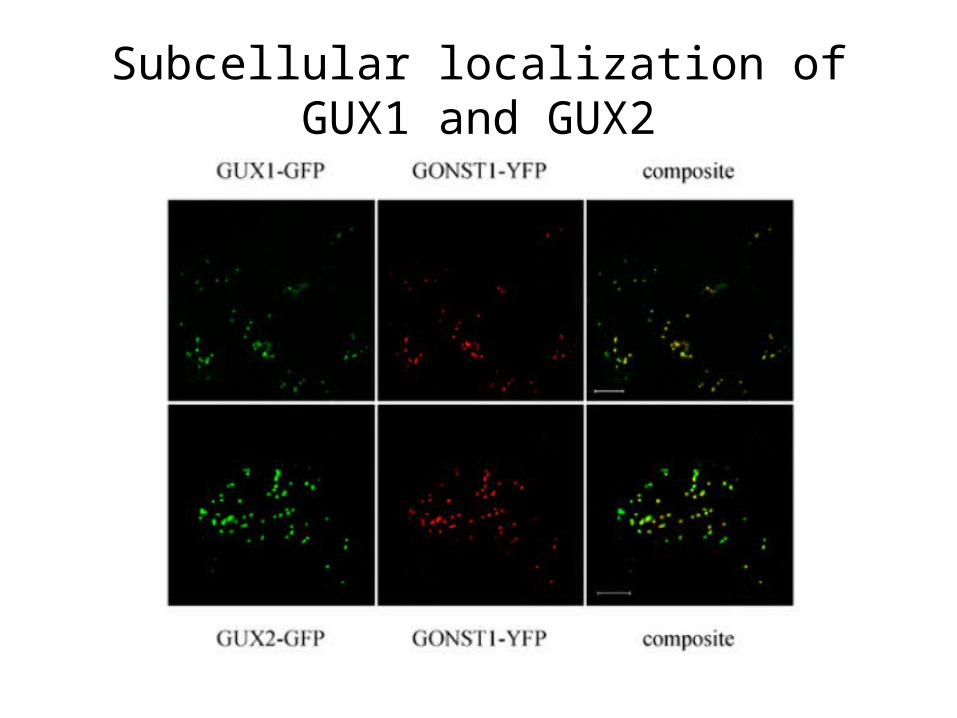
Subcellular localization of GUX1 and GUX2
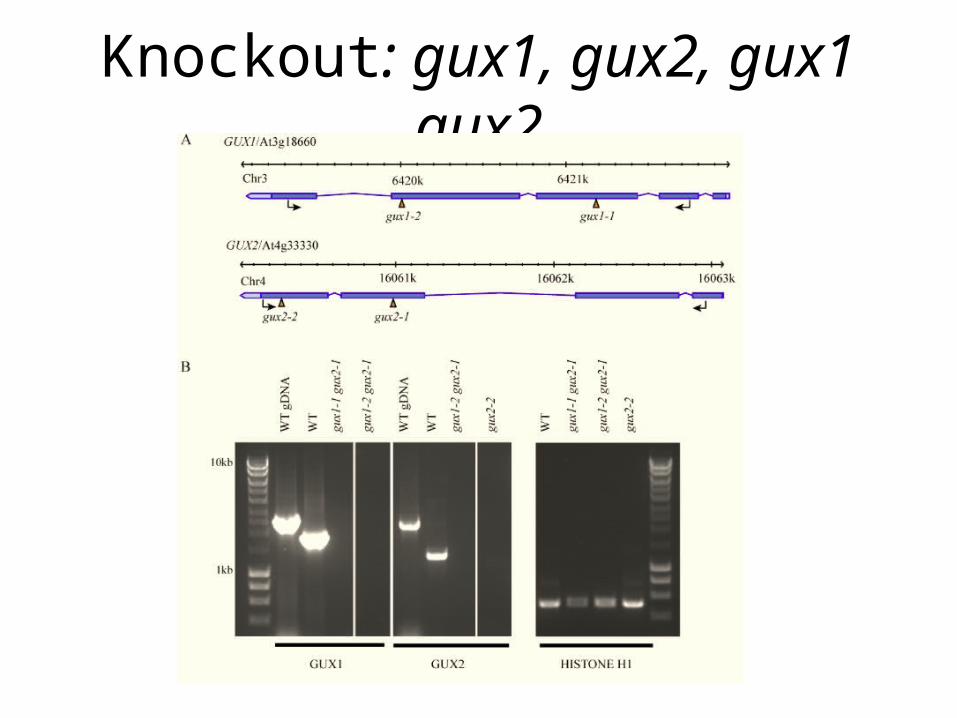
Knockout: gux1, gux2, gux1 gux2
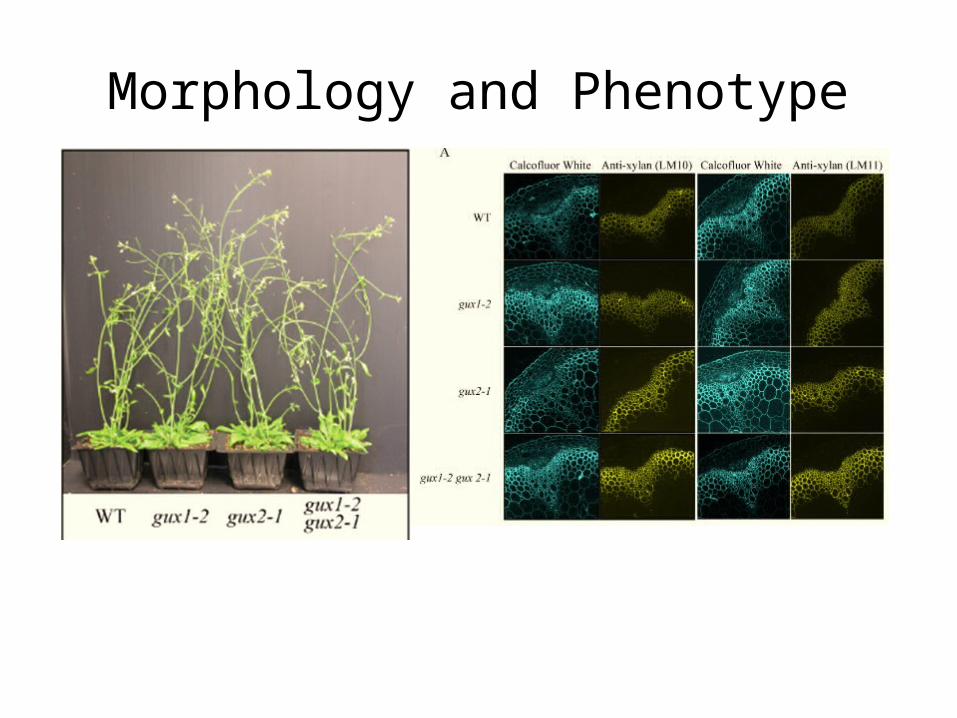
Morphology and Phenotype
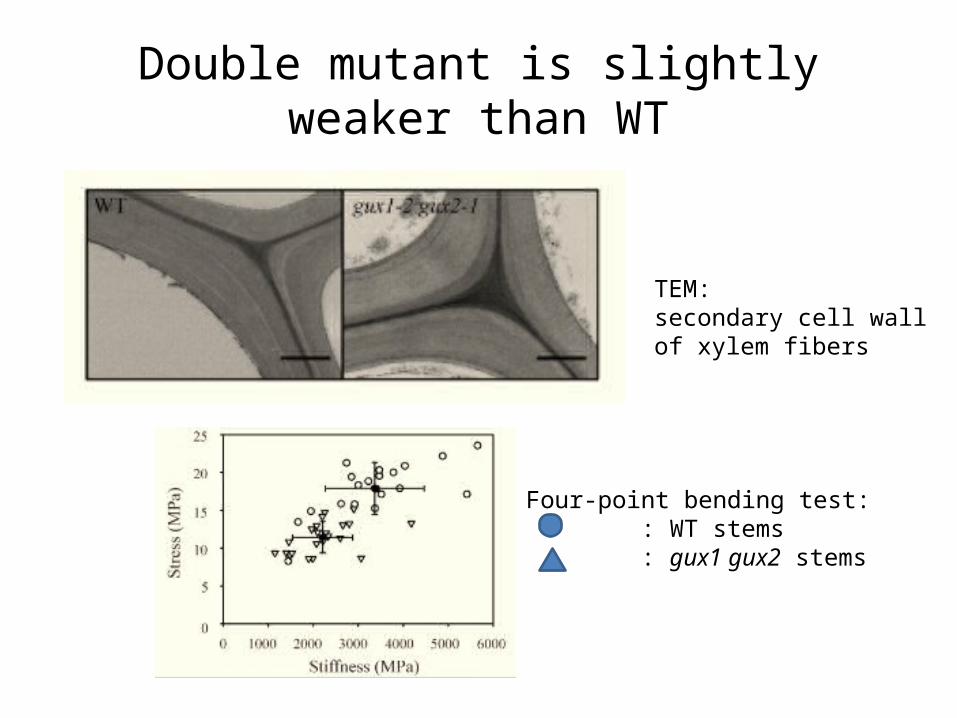
Double mutant is slightly weaker than WT
TEM:secondary cell wall of xylem fibers
Four-point bending test: : WT stems : gux1 gux2 stems
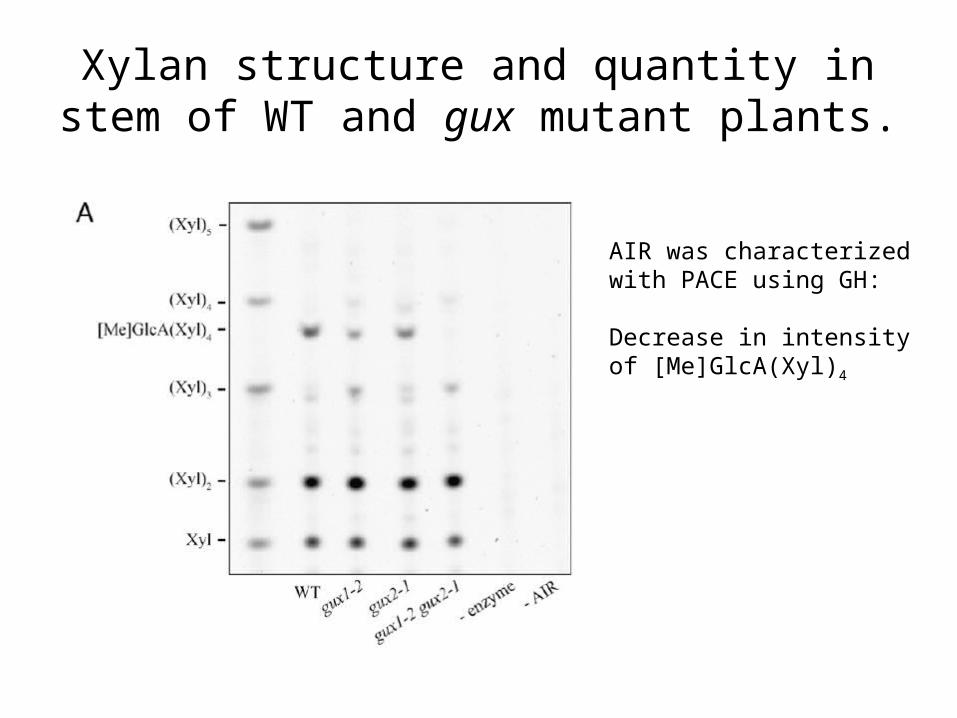
Xylan structure and quantity in stem of WT and gux mutant plants.
AIR was characterized with PACE using GH:
Decrease in intensity of [Me]GlcA(Xyl)4
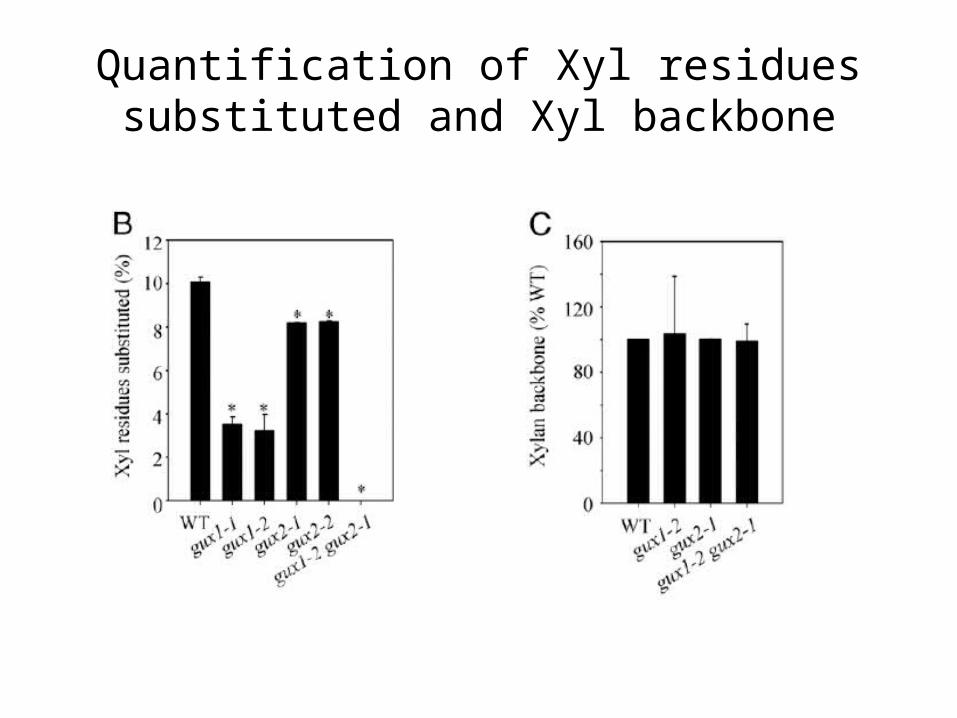
Quantification of Xyl residues substituted and Xyl backbone
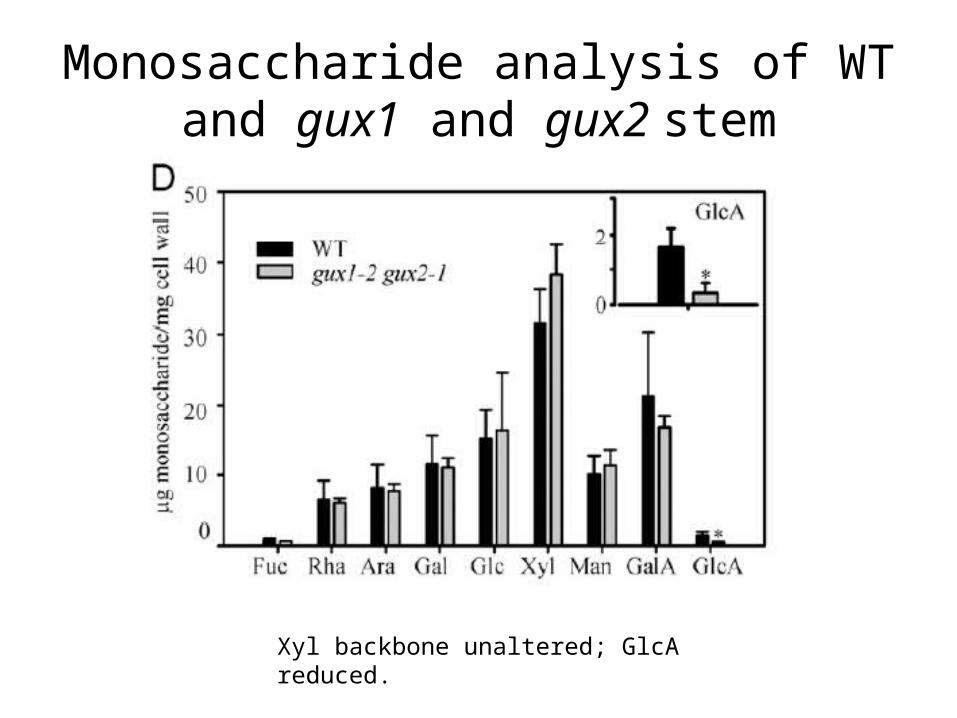
Monosaccharide analysis of WT and gux1 and gux2 stem
Xyl backbone unaltered; GlcA reduced.
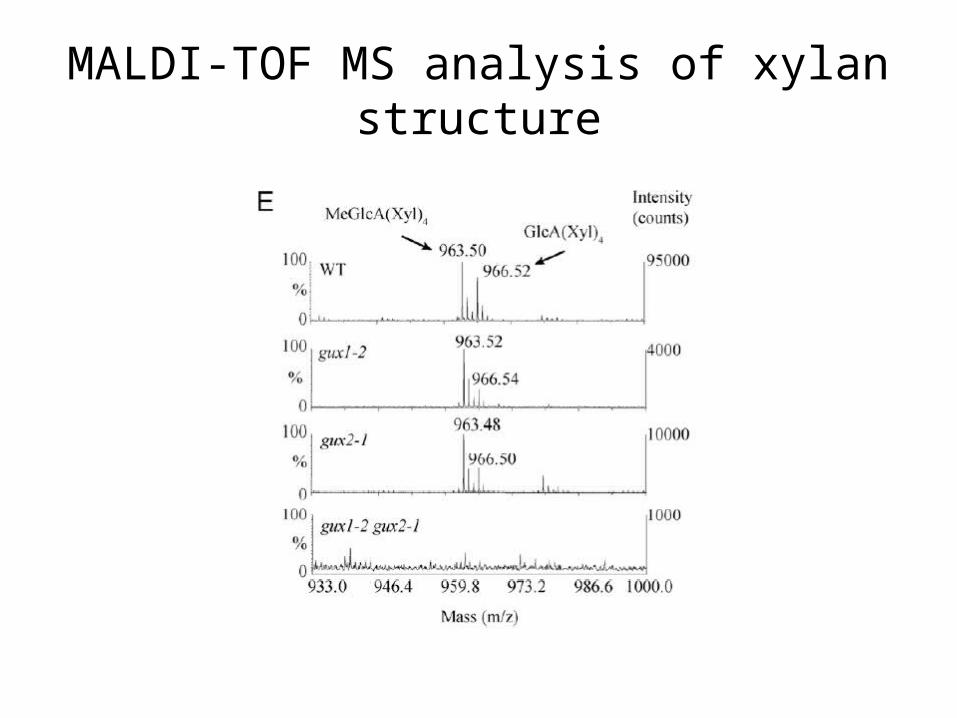
MALDI-TOF MS analysis of xylan structure

GuxT activity in WT and gux1 gux2
GuxT activity is strongly reduced in the double mutant
The activity of XylT was unaffected in the double mutant

GuxT activity in WT and gux1 gux2
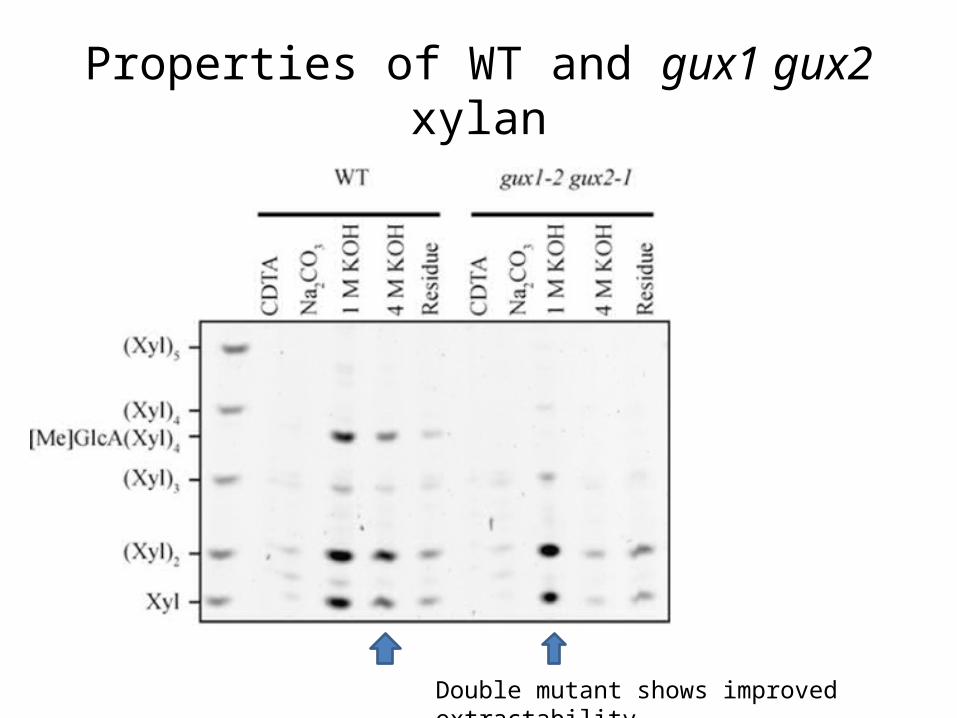
Properties of WT and gux1 gux2 xylan
Double mutant shows improved extractability
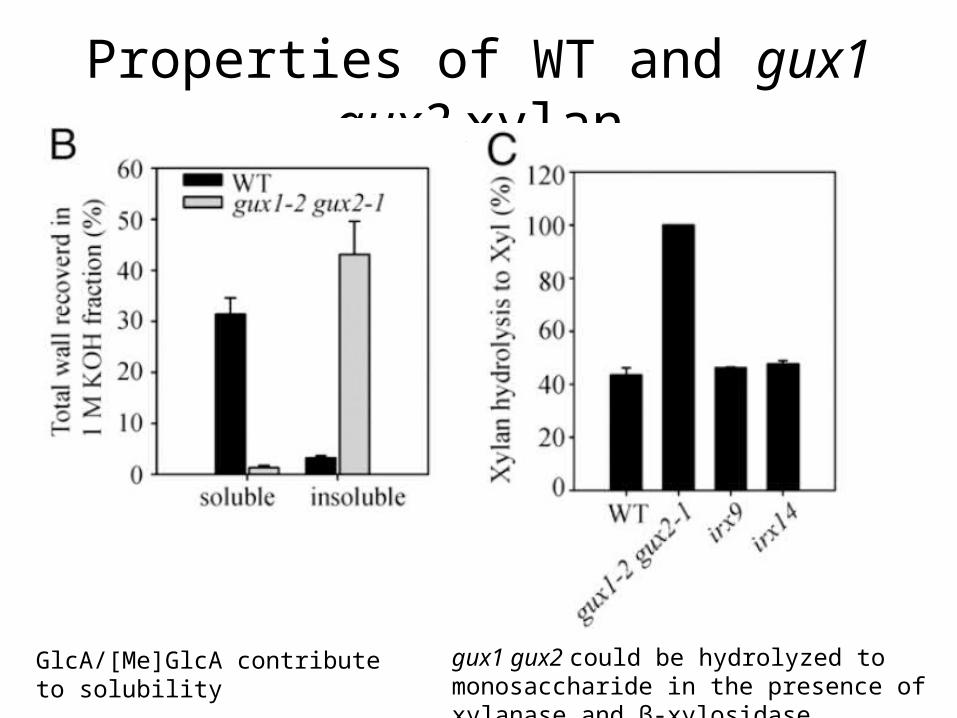
Properties of WT and gux1 gux2 xylan
GlcA/[Me]GlcA contribute to solubility gux1 gux2 could be hydrolyzed to monosaccharide in the presence of xylanase and β-xylosidase

Conclusions
• Identification of two Golgi-localized putative glycosyltransferases, GUX1 and GUX2, that are required for the addition of both glucuronic acid and 4-O-methylglucuronic acid branches to xylan in Arabidopsis stem cell wall.
• gux1 gux2 double mutants show loss of xylan glucuronyltansferase activity and lack almost all detectable xylan substitution, but no change in xylan backbone quantity.

• The xylan in gux1 gux2 shows improved extractibility
• Xylan chain extension and substitution are not obligatorily coupled during synthesis.
• Fermentable sugar release from lignocellulose can be increased by reducing xylan branching.
• Alterations in crop xylan structure could be a feasible goal for the bioprocessing industry.

Reference
• Plant Cell Walls, Peter Albersheim, etc, • Biochemistry & Molecular Biology of Plants, B. Buchanan, W.
Gruissem, R. Jones,Eds.• Biochemical control of xylan biosynthesis- which end is up?
William S York, etc.• Xylan biosynthesis: News from the grass. Ahmed Faik.



![7.5: PROTEINS Proteins Function Structure. Function 7.5.4: State four functions of proteins, giving a named example of each. [Obj. 1] Proteins are the](https://img.dokumen.tips/doc/110x75/56649e425503460f94b34519/75-proteins-proteins-function-structure-function-754-state-four-functions.jpg)

























