Embed Size (px)
Citation preview

Mycologist, Volume 19, Part 1 February 2005. ©Cambridge University Press Printed in the United Kingdom.DOI: 10.1017/S0269915XO5001035
18
Keywords: Metabolomic profiles, water availability,solid substrate fermentation.
Introduction
The pharmaceutical industry still makes use ofmicrobial secondary metabolites as a source of newdrugs for major clinical conditions. New leadcompounds are searched for in high throughput drugdiscovery programs involving the rapid screening ofthousands of samples. Microbial extracts are normallyobtained in liquid culture fermentation systems undersomewhat ‘standardised’ conditions, in terms ofculture media choice and typical fermentationparameters. In particular the use of solid materials asgrowth media for fungal species is a largely neglectedarea within the industry, which appears to favour theuse of submerged liquid fermentation (SLF) technology.This is in spite of major advances in this type oftechnology in other industries, such as fermented foodproduction, mainly in the Far East (Smits et al., 1998).Solid substrate fermentation (SSF) has also been subjectto renewed interest in a number of other areas, such as
enzyme production, solid waste management,production of animal feed and production of a range offungal secondary metabolites (Aldred, 2000).
In principle, SSF offers a very attractive method forthe culture of fungi. Solid substrates are the natural‘habitat’ for most fungal species, in contrast to bacteriawhich usually require freely available water (Griffin,1981). In fact fungi, with their filamentous growthform, are superbly adapted to a terrestrial life style,living on (and in) solid matrices, sometimes in relativelydry conditions. For some fungal species (particularlycertain basidiomycetes) SSF represents the only methodavailable for laboratory culture, and therefore providesunique opportunities for obtaining potentially usefulmetabolites (Penn, 1994). In addition, theheterogeneous conditions that occur in SSF systemsprovide a multiplicity of micro-environments andnutrient gradients which have been shown toencourage secondary metabolite production in somecases (Penn, 1994).
Very little work has been published on the effects ofthe manipulation of environmental factors andadaptation of filamentous fungi to growth and
Water availability and metabolomic profiles ofEpicoccum nigrum and Sarophorum palmicola grown insolid substrate fermentation systems
DAVID ALDRED, JULIA PENN* & NARESH MAGAN
Applied Mycology Group, Institute of BioScience and Technology, Cranfield University, Silsoe, Bedford MK45 4DT, UKand *Cubist Pharmaceuticals (UK) Ltd., 545 Ipswich Road, Slough, Berkshire, SL1 4EQ, UK. ([email protected]).
There has been interest in using environmental screening procedures for enhancing the metabolomic productionprofiles by fungi which are of pharmaceutical interest. This study examined two ecologically distinct species,Epicoccum nigrum and Sarophorum palmicola, in solid substrate fermentation systems over a range of water availability conditions (water activity, aW). Total secondary metabolite profiles were obtained by HPLC + diodearray detection after 14 days incubation on cereal-based substrates in relation to four aW treatments. Temporalstudies (24 d, E. nigrum and 18 d, S. palmicola) showed that metabolite production profiles varied markedly betweenthe two fungi. For E. nigrum, metabolite production generally increased with reduction in aW, and was optimal inthe range 0.99 – 0.97. In contrast to this, for S. palmicola, metabolite production was restricted to the highest aW
level used, 0.998, and declined to zero at 0.99 aW. Statistical analysis revealed that time, aW and their interactionswere significant in all cases (p<0.001). The potential for using ecophysiological stresses for enhancing naturalproduct discovery is discussed.

Mycologist, Volume 19, Part 1 February 2005
19
potential for modifying the production of novelpharmaceutically relevant metabolites. Part of theadaptation to restricted water availability may involvethe production of metabolites for maintaining nichedominance.
A few recent studies have demonstrated that themanipulation of water availability (water activity, aW)and nutrient status can result in increased productionof certain metabolites, notably work reported onsqualestatins, a class of potential cholesterol loweringcompounds. For example, Baxter et al. (1998), workingwith a Phoma sp. grown on glycerol-amended MEA,reported a 200 fold increase in production bydecreasing available water to about 0.98 aW (= 98%equilibrium relative humidity), and Aldred et al.(1999), working with the same organism growing on arange of starchy grain type substrates, reportedincreased squalestatin production at substrate aW levelsas low as 0.96 (= 96% equilibrium relative humidity),compared to those with freely available water (100%relative humidity). Most other studies have concernedfungal toxins and related compounds. Recently, Maganet al. (2002) reported increased production of nivalenol(NIV) and deoxynivalenol (DON) by Fusarium sppgrowing on wheat grain in the presence of somefungicidal agents at lowered aW levels.
In recent years very few drug leads have beendiscovered by the methods currently in use by thepharmaceutical industry. Interest has been increasingin approaches to enhance the production of specificmetabolites and the number of metabolites. This hasincluded the use of both molecular and physiologicalscreening approaches to examine such metabolomicprofiles of interesting microorganisms. This workreports on the impact of the manipulation of wateravailability of the solid substrate on metabolomicprofiles of two unrelated fungal species, Epicoccumnigrum and Sarophorum palmicola, as an alternativeapproach for drug discovery screening systems. Thiswas achieved by conducting temporal experiments overa range of aW levels using a processed bulgar wheatmatrix. Metabolomic profiles were followed by diode-array UV HPLC analysis. Although the identity of themetabolites are not detailed because of commercialconsiderations, the findings are of relevance to drugdiscovery approaches generally.
Materials and Methods
Micro-organismsThe following fungal species were selected from theculture collection of Cubist Pharmaceuticals Limited(formerly Terragen Discovery Limited), Slough, UK:
Epicoccum nigrum Link.Sarophorum palmicola (Henn.) Seifert and Samson.
These both have well characterised metabolomicprofiles when grown on a multi-component solidmedium (Penn, unpublished data), with some ofpotential pharmaceutical relevance. They also havewidely contrasting ecological niches in nature: E.nigrum is a cosmopolitan phyllosphere dweller, while S.palmicola is limited to a partially aquatic environment,this isolate being obtained from a water-logged palmfruit in a rain forest swamp.
Solid growth media and preparationThe solid substrate used in this study was bulgar wheat(Bedford Wholefoods Ltd, Bedford, UK), a processedform of hard (durum) wheat grain. Bulgar wheat isproduced by steaming wheat under pressure until thestarch content is partially gelatinised. The grain is thendried and ‘cracked’ to produce a uniform particle size ofapprox. 3 x 3 mm.
Bulgar wheat was prepared in bulk at aW levels of0.998, 0.99, 0.980 and 0.970 by precise addition ofreverse osmosis water with reference to moisturesorption isotherms previously prepared for thesesubstrates at 25°C (Aldred, 2001). The media wereautoclaved in bulk (121°C, 40 min.) and thendispensed into 72 mm tissue culture jars withpolypropylene microporous vented lids (Sigma, Poole,UK.) and autoclaved for a second time. Jars were filled toa constant volume (40 cm3; substrate depth 15 mm).For each water availability level, sufficient jars wereprepared to destructively analyse at each samplingpoint in triplicate.
Inoculum preparation and culture conditionsFungal spores (S. palmicola) and hyphal fragments (E.nigrum) were harvested from 14 day malt extract agarplates and filtered through sterile glass wool. Theconcentration was adjusted to approx. 1.0 x 106 colonyforming units (CFU) ml-1. Jars were inoculatedaseptically with 0.5 ml of filtered inoculum.
Inoculated treatments were incubated in sealedtransparent containers. Jars of the same wateravailability were incubated together and the humiditywas controlled by using appropriate glycerol/watersolutions of the same aW as the substrates. Alltreatments were incubated at 25 ± 1°C with diurnal UVlighting.
Studies carried out:(a) Metabolomic profiles: an experiment was carried outto determine the metabolites produced by the two fungiwhen growing on bulgar wheat at the treatment aW
Mycologist, Volume 19, Part 1 February 2005
20
levels. Samples were analysed for metabolomic profilesafter 14 days incubation. Effects on growth patternsand sporulation were also noted.(b) Temporal study on optimum production of specificmetabolites of interest: a detailed temporal experimentover 24 days for E. nigrum and 18 days for S. palmicola,with sampling at days 0, 4, 7, 10, 18 and 24. For bothfungal isolates three of the candidate secondarymetabolites were selected for examination. These weredesignated as e1 (6.9 min.), e2 (7.3 min.) and e3 (8.9
min.) and s1 (4.6 min.), s2 (6.4 min.) and s3 (6.6 min.)for E. nigrum and S. palmicola respectively (retentiontimes in brackets). Statistical analysis was performed onthe results of these experiments by analysis of variance(ANOVA: GenStat, fifth edition, RothamstedExperimental Station, UK). Two factor analyses wereperformed to investigate the effects of time and aW, andtheir interactions.
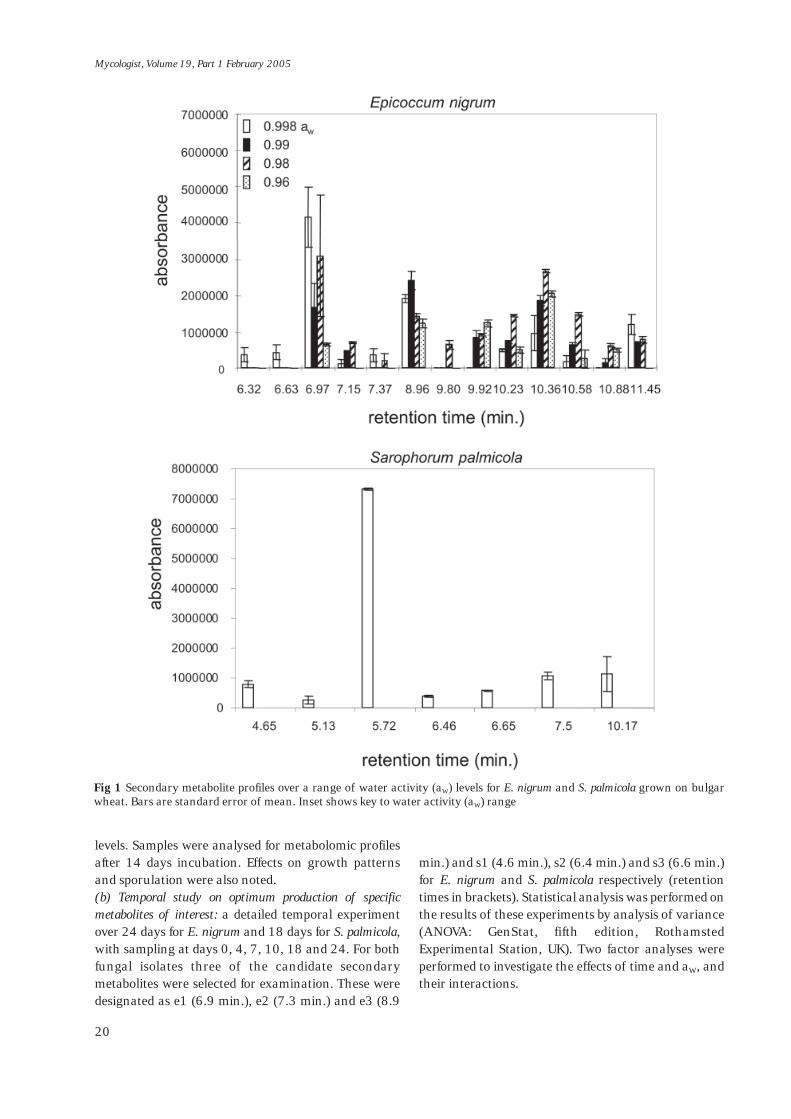
Fig 1 Secondary metabolite profiles over a range of water activity (aW) levels for E. nigrum and S. palmicola grown on bulgarwheat. Bars are standard error of mean. Inset shows key to water activity (aW) range

21
Secondary metabolite extraction and HPLC analysisDestructive sampling was carried out by coarsechopping using a sterile knife or spatula followed byfreeze-drying of the entire jar contents to eliminatewater (Edwards high vacuum freeze drier unit,Edwards, Crawley, Sussex. UK.) The freeze-driedmaterial was then dry ‘macerated’ using a foodprocessor (Kenwood, Havant, Hants. UK.) for approx.30 sec. An aliquot of 50 ml HPLC grade methanol(Prolabo, Fontenay, France) was added to each jar andplaced in a fixed frequency ultrasonicating bath (DeconUltrasonics, Hove, Sussex. UK.) for 30 min to optimisesolvent extraction. A small volume of extract was thenfiltered through a 0.45µm syringe filter (Sartorius,Epsom, Surrey, UK.). Filtrate samples were placed inHPLC vials (Chromacol, Welwyn Garden City, Herts.UK.) and analysed immediately or placed in a freezer at
–20°C pending analysis. Uninoculated samples ofbulgar wheat were also prepared in the same way forHPLC analysis to act as substrate control samples.
Samples were analysed using a Waters Alliance2960 diode-array UV system with a gradient methodand photodiode array detection (Waters 996) (Waters,Millipore, Milford, MA. USA.), Waters symmetry C8column, and dedicated Waters "millennium" software.All HPLC results were corrected against uninoculatedsubstrate control samples.
Results
Metabolite profiles for E. nigrum and S. palmicolagrowing on bulgar wheat. Figure 1 compares total metabolite profiles for E.nigrum and S. palmicola on bulgar wheat substrate. The
Fig 2 Temporal development of metabolites produced by E.nigrum grown on bulgar wheat. (a) metabolite e1, (b) metabo-lite e2, and (c) metabolite e3.
Fig 3 Temporal development of metabolites produced by S.palmicola grown on bulgar wheat. (a) metabolite s1, (b)metabolite s2, and (c) metabolite s3.
Mycologist, Volume 19, Part 1 February 2005

22
most important difference between the two fungi wasthe markedly different response to lowered aW
conditions. For E. nigrum, there was a positive responseto lowered aW with increases in the levels of mostmetabolites and some other metabolites only producedunder drier treatment conditions. Qualitativeobservations of growth of this fungus in different aW
treatments on both media showed an extremelyuniform growth pattern, with slightly more diffusegrowth at 0.96 aW. The effect on S. palmicola wasexactly the opposite, with a striking decrease inmetabolite production with decreasing aW. Nometabolites were produced at <0.997 aW. This trendwas, to some extent, reflected in the observations ongrowth, with dramatically reduced stromatalproduction (conidia bearing structures) at aW levels <0.997, although some vegetative growth persisteddown to the lowest water availability level tested.
Temporal development of metabolites on bulgarwheatFigure 2 shows the temporal development of threemetabolites for E. nigrum grown on bulgar wheat over arange of aW levels. All three metabolites are produced atincreased levels under conditions of lowered wateravailability. In the case of metabolites e2 and e3,maximum production was achieved at the lowest aW
level tested (0.97). In the case of metabolite e1,maximum production was achieved at around 0.99 aW,which still represents conditions significantly lowerthan water saturation. Statistical analysis revealed thattime (in days, d), aW and their interactions weresignificant (p < 0.001).
Figure 3 shows the temporal development of threemetabolites produced by S. palmicola when grown onbulgar wheat over the same range of aW levels. In thiscase metabolites were produced only under conditionsof water saturation, and fall off steeply to zeroproduction at 0.99 aW, indicating a very narrowpermissible range of water availability for metaboliteproduction. The production patterns of metabolites s2and s3 suggest that metabolite concentrations were stillincreasing after 18 days incubation. Statistical analysisrevealed that time (d), aW and their interactions weresignificant (p < 0.001).
Discussion
This study has demonstrated that metabolomic profilescan be significantly modified by environmental stressfactors such as water availability and time in SSFsystems. The ecological background of the candidatespecies being used for screening also needs to be taken
into account for improving the chances of discoveringnew metabolites or enhance the production ofmetabolites of interest.
In a number of respects, bulgar wheat represents anideal medium for fungal SSF work. In particular, itdemonstrates a number of important attributes. Theseinclude good ‘granulometry’– a uniform particle size ofapprox. 3 mm, which gives a free flowing property overa wide range of aW levels (this facilitates handling). Thissubstrate also demonstrates good structural integrity,being able to withstand repeated autoclaving cycles. Itis also pre-processed i.e. semi-cooked with outer layersremoved, and shows abraded surface characteristics,properties which seem to facilitate hyphal colonisationand penetration. Also, from a commercial point of view,it is freely available at consistent quality.
Fungi generally show a high level of adaptation tothe particular ecological niche they occupy. Thisadaptation may be shown in a number of ways,including mode of reproduction, interactions withother organisms including other fungi, and primaryresource capture (Magan & Lacey, 1984, 1985; Cooke& Whipps, 1993; Magan et al., 2003). All of thesestrategies enhance competitive capacity and ‘fitness’helping the organism to maintain its niche position. Inthis study the use of water availability as an adaptive‘marker’ has been investigated for two dissimilar fungioriginating from very different ecological niches.
The different responses of E. nigrum and S. palmicolato aW manipulation are of particular interest. E. nigrumis an extremely cosmopolitan phyllosphere dwellerwhich can be expected to encounter water stress, in theform of drought, in its natural environment. Accordingto the classification proposed by Griffin (1981), E.nigrum was described as sensitive to changes in aW.Magan and Lacey (1984) described the lower limit forgrowth of this organism as occurring at 0.89 aW
(25°C). In this study, subtle manipulation of aW,producing mild water stress, resulted in a strongresponse from E. nigrum in terms of increasedmetabolite levels. In fact, the precise setting of wateravailability levels, resulting in carefully controlledwater stress, is a key factor in this approach. In the vastmajority of studies involving the manipulation of aW,large changes in aW have been imposed, resulting instraightforward inhibition of both growth andmetabolomic profiles.
The effect of water stress can be viewed as a similarresponse to that produced by nutrient limitation. In thecase of nutrient stress a well understood mechanismoperates which regulates the production of certainsecondary metabolites within particular limitingnutritional conditions. This mechanism operates at the
Mycologist, Volume 19, Part 1 February 2005

Mycologist, Volume 19, Part 1 February 2005
23
level of gene expression and effectively holds theproducer genes in a non-active state when nutrients arein excess. Expression of genes is therefore effectivelycoupled to a specific deterioration in environmentalconditions. In the case of E. nigrum water stress seemsto bring about a similar response, although there is noevidence at present for a specific mechanismcontrolling gene function. It can be assumed that thesecondary metabolites produced by these types ofmechanisms heighten the competitive advantage ofproducer organisms under natural conditions, and areof particular importance as environmental conditionsbecome hostile.
In contrast, S. palmicola, which was isolated from anessentially aquatic habitat (water-logged palm fruitfrom rain forest swamp), demonstrated inhibition ofboth growth and secondary metabolite production inthe same range of aW levels. This isolate of S. palmicolawas probably not adapted to respond to lowered wateravailability conditions, so that the stress imposed wasboth severe and inappropriate, with respect to itsnatural ‘lifestyle’. It is noteworthy that this organismalso showed growth limitation at lowered aW.
Caporale (1994) has described a similarecophysiological approach to the study of secondarymetabolism which he has termed ‘chemical ecology’.Chemical ecology is one approach to modern drugdiscovery which is centred on natural products, andseeks to understand the biochemical and physiologicalbasis of ecological interactions between organisms.This approach is in marked contrast to the normalmethods used in laboratory culture of micro-organisms, where rich media and optimal growthconditions have been employed, with typically littleattention paid to the natural lifestyle and relevantinteractions of the organisms. In fact as early as 1987,Tribe questioned the relevance of the majority of mediaemployed by mycologists, in particular the use ofnutritionally rich materials. A chemical ecology typeapproach to microbial drug discovery should thereforepay particular attention to the natural lifestyle andrelevant stresses experienced by candidate organisms.Most ‘standard’ conditions used in traditional
laboratory culture would be rejected in favour of morerelevant biotic and abiotic conditions. This approachwould seem to hold particular promise for future drugdiscovery programs.
ReferencesAldred, D., Magan, N. & Lane, B. S. (1999). Influence of water
activity and nutrients on growth and production ofsqualestatin S1 by a Phoma sp. Journal of AppliedMicrobiology 87: 842-848.
Aldred, D. (2000). The development of a solid phase fermentation system for the production ofpharmaceutically useful secondary metabolites in fungi.PhD Thesis Cranfield Biotechnology Centre, CranfieldUniversity, UK.
Baxter, C. J, Magan, N., Lane, B. & Wildman, H. G. (1998).Influence of water activity and temperature on in vitrogrowth of surface cultures of a Phoma sp. and production ofthe pharmaceutical metabolites, squalestatins S1 and S2.Applied Microbiology and Biotechnology 49: 328-332.
Caporale, L. H. (1994). Chemical ecology: A view from thepharmaceutical industry. Proceedings of National Acadamyof Science 92: 75-82.
Cooke, R. & Whipps, J. (1993). Ecophysiology of Fungi.Blackwell, Oxford.
Griffin, D. M. (1981). Water and Microbial Stress. In Advancesin Microbial Ecology vol.5, pp. 91-136. Edited by M.Alexander. New York: Plenum.
Magan, N. & Lacey, J. (1984). Effects of water activity, temperature and substrate on interactions between fieldand storage fungi. Transactions of the British MycologicalSociety 82: 83-93.
Magan, N. & Lacey, J. (1985). Interaction between field andstorage fungi on wheat grain. Transactions of the BritishMycological Society 85: 29-37.
Magan, N., Hope, R., Colleate, A. & Baxter, E. S. (2002).Relationship between growth and mycotoxin production byFusarium spp., biocides and environment. European Journalof Plant Pathology 108: 685-690
Magan, N., Hope, R., Cairns, V. & Aldred, D. (2003). Post-harvest fungal ecology: Impact of fungal growth and mycotoxin accumulation in stored grain. European Journalof Plant Pathology 109: 723-730.
Penn, J. (1994). Not shaken or stirred? Alternatives to theusual cocktails for fungal fermentations. In DiscoveringDrugs from Nature. Conference Documentation, 8,9December 1994. London: IBC Technical Services.
Smits, J. P., Sonsbeek, H. M., Rinzema, A. & Tramper, J.(1998). Solid state fermentation–a mini review. Agro-food-Industial Hi-Technology 9: 29-36.
Tribe, H. T. (1987). Do we think about our culture media?Mycologist 21: 69-71.



























![Systems Metabolomic Lecture[1]](https://img.dokumen.tips/doc/110x75/546af5e0b4af9f486b8b45b1/systems-metabolomic-lecture1.jpg)

