Embed Size (px)
Citation preview

魚 病 研 究 Fish Pathology,38(4),151-161,2003.12 2003 The Japanese Society of Fish Pathology
Virulence Properties of a Newly Identified Species
Aeromonas sp. T8 Group Isolated from
EUS-affected Fish
M.M. Rahman1,T. Somsiri2, K. Tajima1*and Y. Ezura1
1Laboratory of Microbiology, Graduate School of Fisheries Sciences, Hokkaido University, Hakodate, Hokkaido 041-8611, Japan
2Aquatic Animal Health Research Institute , Kasetsart University Campus, Jatujak, Bangkok 10900, Thailand
(Received May 30,2003)
ABSTRACT-The present study was conducted to determine the virulence and virulence proper
ties of 4 bacterial strains belonging to a newly identified species Aeromonas sp. T8 group, isolated from epizootic ulcerative syndrome (EUS)-affected fish of Southeast Asian countries. Virulence of
a representative strain T8 to fish was investigated through intramuscular injection to silver barb Barbodes gonionotus, striped snakehead Channa striatus and tilapia Oreochromis sp., where the
LD50 value was determined to be 1.5×105,3.2×105 and 2.9×105 CFU/fish, respectively.
However, all strains were found non-virulent for mice upon intraperitoneal or intravenous injections
at a concentration of 3×107 CFU/mouse. Live bacterial cells of these strains showed lipase ,
protease and hemolytic activities in agar plate assays. The hemolytic activity of the extracellular
products(ECPs)was found to be higher for fish blood cells than for mammalian blood cells. Heat
stability, EDTA and PMSF sensitivities of β-hemolytic activity of the ECPs differed from that of A.
hydrophila and A. caviae reference strains. ECP of only one strain(P2)showed cytotoxicity
against both fish(RTG-2, FHM, EPC, BF-2 and SNN-1)and human(HeLa)cell lines, while none of
the strains exhibited enterotoxicity to suckling mice. All of the strains also failed to amplify
aerolysin and cytolytic enterotoxin(AHCYTOEN)genes by PCR.
Key words:Aeromonas sp. T8, virulence factor, protease, hemolysin, cytotoxin, enterotoxin, EUS
Bacteria of the genus Aeromonas have been reported as pathogens of poikilothermic animals for more than one hundred years (Carnahan and Altwegg, 1996). Recently, several motile species are also increasingly recognized as primary, opportunistic disease agents in human (Janda, 1991; Janda and Abbott, 1996). Although several Aeromonas species have been found in association with healthy or diseased fish
(Sugita et al., 1994, 1995; Esteve, 1995; Iqbal et al., 1998) and human clinical samples (Huys et al., 1996; Valera and Esteve, 2002), the importance of recently identified species in fish and human pathology is mostly unknown. During the last decade, the taxonomy of the genus
Aeromonas has undergone major changes due to the application of extended DNA-DNA hybridization and
other molecular techniques (Kozinska et al., 2002). At
present, the genus Aeromonas comprises 16 wellrecognized genomic species (Valera and Esteve, 2002). Recently, a new species designated as Aeromonas sp. T8 has been reported (Iqbal, 1999). The first identified strain T8 (IAM 14920T, JCM 11177T) of this species possesses distinct phenotypic properties, which represents a cluster with Aeromonas caviae in the phylogenetic dendogram constructed on the basis of 16S rDNA sequences. However, it comprises discrete DNA-DNA homologies with all recognised Aeromonas genomic species (Iqbal, 1999). Recently, we identified three more strains of the new species on the basis of phenotypic properties, phylogenetic analysis based on 16S rDNA sequences and DNA-DNA hybridization studies
(Unpublished). All of these strains were isolated from epizootic ulcerative syndrome (EUS)-affected fish of Southeast Asian countries. Evaluation of the patho
genic importance of these strains for fish and mammals
*Corresponding author
E-mail:[email protected]

152 M.M. Rahman, T. Somsiri, K. Tajima and Y. Ezura
is needed.
In the recent years, various putative virulence fac
tors have been ascribed to explain the process of patho
genicity of Aeromonas spp.(Krovacek et al.,1994).
Such factors include the production of extracellular pro
tease,α-andβ-hemolysins, cytotoxin, enterotoxin, leci
thinase etc. Crude extracellular products(ECPs)of
motile Aeromonas spp. have been suggested to play a
role in virulence process(Allan and Stevenson,1981;
Rodriguez et al.,1992).
The present study was conducted to determine
the pathogenesis of the strain (s) belonging to the
Aeromonas sp. T8 group to fish and mouse, characterize
in vitro enzymatic activities of bacterial cells and ECPs
as putative virulence factors, and screen for the pres
ence of aerolysin and cytolytic enterotoxin genes by
polymerase chain reaction as susceptive virulence mark
ers.
Materials and Methods
Bacterial strains
Four bacterial strains of Aeromonas sp. T8 group
collected from Thailand(T4, T8 and T27)and the Philip
pines(P2), and some reference strains were used
(Table 1). Strains of Aeromonas sp. T8 group were iso
lated from EUS-affected fish and previously identified
through their phenotypic characteristics, phylogenetic
analysis based on 16S rDNA sequences and DNA-DNA
homology studies(Unpublished). In the present study,
reference strains Aeromonas hydrophila ATCC 7966T, A.
caviae ATCC 15468T and A. veroniibiotype sobria ATCC
9071T were included either to confirm the results or to
compare the putative virulence properties with those of
the experimental strains. All bacterial strains were rou
tinely sub-cultured on nutrient agar(NA;pH 7.2)at
25℃. The composition of NA was 1.0% polypepton
(Nihon Seiyaku, Japan),0.5% beef extract(Wako,
Japan),0.12% NaCl and 1.5% agar(Wako,Japan)dis
solved in distilled water. Stock cultures were preserved
in nutrient broth supplemented with 10% glycerin at
-80℃.
V
irulence for fish The virulence assay was conducted with a repre
sentative strain(T8)to fingerlings(average weight 10g)
of three tropical fish species, silver barb Barbodes
gonionotus, striped snakehead Channa striatus and tila
pia Oreochromis sp. The bacterial strain was cultured on
tryptic soy agar(Difco, USA)and then suspended in
sterile physiological saline(0.85% NaCl). Desired dilu
tions were adjusted by the ten-fold dilution method.
The fish were injected intramuscularly just below the
anterior part of the dorsal fin with 100μL of serially
diluted concentrations of bacterial suspensions. Each
suspension was injected into 10 fish for each challenge
group, and the fish was kept separately in 10 L aquaria
at 28℃ for 4 days. No feed was given to fish during the
experiment. The clinical symptoms and mortality of arti
ficially infected fish were recorded, and the median lethal
dose(LD50)was calculated by the method of Reed and
Muench(1938).
Virulence for mouse
Virulence of all of the strains belonging to the
Aeromonas sp. T8 group was assayed in albino mice
(10-12 wk old)by intraperitoneal and intravenous injec
tions following the methods of Janda et al.(1985)and
Mateos et al.(1993). Bacterial strains were grown in
nutrient broth at 37℃ for 18 h with shaking(160
rpm). Cells were harvested by centrifugation at
4,000×g and 4℃, and suspended in sterile saline
(0.85% NaCl). The cell suspensions were adjusted to
approximately 3×108 CFU/mL and confirmed by dupli-
Table 1. Bacterial strains used in this study

Virulence properties of Aeromonas sp. T8 group153
cate plate counts. For each assay, 5 albino mice of
both sexes (average weight 30 g) were injected with 100
,ƒÊL of the bacterial cell suspension and observed for 7
days. A. hydrophila ATCC 7966T was used as a posi
tive control and sterile saline was injected into control
mice. Mice died within the experimental period were
immediately autopsied, and portions of the kidney, liver
and spleen were aseptically collected for isolation of
bacteria on NA. All surviving mice were sacrificed after
the experimental period, internal organs were visually
examined, and attempts were made to re-isolate the
bacteria from kidney, liver, spleen and cardiac
blood. Bacteria grown on the plates were tested by
physiological and biochemical characterization.
Enzymatic and hemolytic activities of live bacterial cells
Exoenzyme activities of bacterial cells were evalu
ated by spot inoculations onto NA plates supplemented
with various proteins, lipid and phospholipid as
substrates. Albumin (from bovine serum), casein, gela
tin and hemoglobin were obtained from Wako, Japan;
elastin (from bovine neck ligament) from Nakarai, Japan;
Tween-20 from Bio-Rad, UK. and egg-yolk coagulates
from Difco, USA. The composition of the supplements
for protease assay was the same as described by Shotts
et al. (1985). To determine lipase and phospholipase
activity, 1% Tween-20 and 5% egg-yolk coagulates were
supplemented into NA, respectively. All assay plates
were incubated at 25•Ž for 7 days. After incubation,
the colony size and diameter of hydrolytic halo around
the inocula were measured, and the ratios were calcu
lated as mentioned by Shotts et al. (1985).
Hemolytic activities of live bacterial cells were deter
mined against erythrocytes of both poikilothermic (com
mon carp and rainbow trout) and homoeothermic (horse,
human, rabbit and sheep) sources by the plate assay
method as described by Esteve et al. (1995). Five per
cent washed blood cells were added into NA. Spot
inoculations of the bacteria were done. Plates contain
ing fish or mammalian blood cells were incubated at
25•Ž or 37•Ž for 48 h, respectively. After incubation,
clear zones surrounding the inocula were considered a
positive result and categorized as ƒ¿ or ƒÀ as mentioned
by Janda (1991). The zone ratios were calculated as
mentioned by Esteve et al. (1995).
Preparation of ECPs
NA plates overlaid with sterile cellophane sheets
were used to collect the concentrated preparations of
ECPs as described by Esteve et al. (1995). Plates
were inoculated with 100 ƒÊL of overnight culture suspen
sion and incubated at 25•Ž for 48 h. After incubation,
ECPs on the cellophane sheet were washed off with 3
mL of phosphate buffered saline (PBS, pH 7.2) and
centrifuged. The supernatant fluids were filter-sterilized
through 0.22 ƒÊm pore-sized filters•@ (Millex-GV, Millipore,
Japan), and the aliquots were either directly used or pre
served at 4•Ž for no more than 2 days. The protein
concentration of ECPs was determined by the method of
Lowry et al. (1951), using bovine serum albumin (Wako,
Japan) as a standard.
Protease activity of ECPs
Protease activity of ECPs was determined as
described by Gudmundsdottir (1996) using azocasein
(Sigma, USA) as a substrate.
The protease activity was characterized on the
basis of the inhibition effect of different treatments on
ECPs. For this purpose, portions of ECPs were heated
at 60 and/or 80•Ž for 15 min. Other portions were
mixed with equal volume of 0.020 mol/L ethylenedi
amine-N,N,N,N-tetraacetic acid (EDTA; Dojindo, Japan)
and/or 0.050 mol/L phenyl methyl sulphonyl fluoride
(PMSF; wako, Japan) for 15 min. Then protease activ
ity was assayed as mentioned above, and the percent
age of inhibition was determined as described by
Gudmundsdottir (1996). ECPs of A. hydrophila ATCC
7966T and A. caviae ATCC 15468T were also incorpo
rated in the experiment.
Hemolytic activity of ECPs
Hemolytic activity of the ECPs (prepared as men
tioned above) against fish and mammalian erythrocytes
was assayed after Gunnlaugsdottir and Gudmundsdottir
(1997) with a little modification. Briefly, 100 ƒÊL of two
fold serial dilutions of ECPs in PBS were added to an
equal volume of 1% (v/v) blood cell suspension in micro
centrifuge tubes and incubated at 25•Ž. Fish blood
cells were incubated for 12 h, whereas mammalian
blood cells were incubated for 24 h. PBS was used
instead of ECPs as a control. The hemolytic activity
was estimated by eye observation as well as by micro
scopic observation of the titration mixture. Hemolytic
titre was expressed as the reciprocal of the highest dilu
tion that gave 50% hemolysis. Hemolytic activity of the
ECPs of A. hydrophila ATCC 7966T was also assayed at
the same time.
In order to characterize the hemolytic activity, differ
ent portions of ECPs were treated (heating at 60 and
80•Ž, and addition of EDTA and PMSF) as mentioned in
protease characterization. After the treatments,
hemolytic activity was estimated against rabbit blood
cells as mentioned above. ECPs of A. hydrophila
ATCC 7966T and A. caviae ATCC 15468T were also
included in the characterization experiment.
Cytotoxicity of ECPs
Both poikilothermic and homoeothermic cell lines
were used to evaluate the cytotoxic effect of the ECPs of
the Aeromonas sp. T8 group. The fish cell lines
included RTG-2, EPC, BF-2, FHM and SNN-1. HeLa
was used as mammalian cell line. The cytotoxicity

154 M.M. Rahman, T. Somsiri, K. Tajima and Y. Ezura
assay was carried out with a slight modification of the
method described by Gunnlaugsdottir and Gudmundsdottir
(1997). Briefly, confluent monolayers of the cells were
grown in flat-bottomed 96-well tissue culture plates in
minimal essential medium(MEM)supplemented with
10% fetal bovine serum and 1% penicillin-streptomycin
except for the SNN-1 cell line. SNN-1 cell line was
grown in L-15 medium. The reaction system was pre
pared by removing 100 mL of medium from each well
and adding serial two fold dilutions of 100μL of ECPs
(ECP content in the first dilution was adjusted to 450μg
protein)in PBS (pH 7.2). PBS was used as a
control. The fish cells were incubated at 20℃ for 2 d,
and the mammalian cells were incubated at 37℃ in a
5%CO2 atmosphere for 1 d. The cytotoxic activity was
examined under an inverted microscope. Toxicity titres
were expressed as the reciprocal of the highest dilution
of ECPs that caused complete or partial destruction of
cell monolayers. Heat stability of the cytotoxicity was
investigated by heating the ECPs at 80℃ for 15
min. ECPs collected from A. hydrophila ATCC 7966T
was used as a positive control.
Enterotoxicity of ECPs
Enterotoxic activity of bacterial culture supernatant
was evaluated by the suckling mouse test using three
types of culture media to screen whether the growth
medium had any effect on the assay. These media
included double-strength nutrient broth, double-strength
brain heart infusion broth(Difco, USA)and double
-strength glucose-free tryptic soy broth(Difco, USA). A
5 mL portion of each growth medium in an Erlenmeyer
flask was inoculated with bacteria and incubated at 37
℃ with shaking(175 rpm)for 24 h. Cell-free supernatant
was prepared by centrifugation at 10,000×g for 15 min at
4℃ and subsequent filtration through a 0.22μm pore
sized filter(Millex-GV, Millipore, Japan). Culture super
natant solution(100μL)containing sterile 0.02%(w/v)
trypan blue was administered orally into the stomachs of
2-4 day-old infant mice with a fine silicon tube con
nected with a pipette tip attached to a micropipette.
Three mice were used per challenge group. The infant
mice were then individually placed on a layer of tissue
paper in a petridish and left at 30℃ for 3-4 h. The ani
mals were then sacrificed, and entire intestinal tracts
were removed and weighed. The remaining body
weight was also measured. The ratio of intestinal
weight to remaining body weight was calculated, and
enterotoxicity was considered positive if the ratio
exceeded 0.08. Supernatant from A. hydrophila ATCC
7966T and sterile growth media were used as positive
and negative controls. Supernatant from A. caviae
ATCC 15468T was also included.
PCR for detection of virulence genes
PCR was conducted in order to detect aerolysin and
cytolytic enterotoxin(AHCYTOEN)genes in the strains
of Aeromonas sp. T8 group. Bacterial cells were grown
in nutrient broth, and genomic DNA was extracted
by using the Wizard Genomic DNA Purification Kit
(Promega, USA)following the manufacturer's instruc
tions. The aerolysin gene was amplified using forward
primer 5'-GC(A/T)GA(A/G)CCC(A/G)TCTATCC(A/T)G-
3'and reverse primer 5'-TTTCTCCGGTAAGACCATTG-
3'(Santos et al.,1999). The primer combination
AHCF1(5'-GAGAAGGTGACCACCAAGAACA-3')and
AHCR1(5'-AACTGACATCGGCCTTGAACTC-3')were
used to amplify the AHCYTOEN gene as described by
Kingombe et al.(1999).
A final volume of 50 mL PCR mixture contained 1.25
μL of each 0.2 mM deoxyribonucleotide tri-phosphate,5
μL of 25 mM MgCl2,100-300 ng genomic DNA,5μL of
10×PCR buffer,2.5μL of a 20μM solution of each prim
ers,0.3μL of Taq DNA polymerase(Promega SUSA)at
5U/μL and double-distilled sterile water. The PCR am
plification was performed with a Gene Amp 9700 PCR
system(PE Applied Bio Systems). The aerolysin gene
was amplified following the protocol described by
Gonzalez-Serrano et al.(2002), and the AHCYTOEN
gene was amplified following the temperature profile de
scribed by Rahman et al.(2002). The PCR amplicons
were separated by electrophoresis in a 1.5% agarose
gel and visualised after ethidium bromidestaining. DNA
templates from A. hydrophila ATCC 7966T and A. veron
ii biotype sobria ATCC 9071T were used as positive con
trols, whereas the template from A. caviae ATCC 15468T
was used as a negative control.
Results
Virulence for fish
Strain T8 showed a high degree of virulence for a
ll fish species. The LD50 of the strain for silver barb,
striped snakehead and tilapia was 1.5×105,3.2×105
and 2.9×105 CFU/fish, respectively(Table 2). In all of
the challenged fish, swelling and inflammation of the tis
sue surrounding the injection site were observed within
12 h after injection. Hemorrhagic and necrotic lesions
developed in the muscle adjacent to the injection site
within 24 h. Lesions became more pronounced at the
2nd and 3rd day after inoculation. The internal clinical
symptoms that occurred after the injection are
summarised in Table 2. Bacteria were re-isolated from
the moribund or dead fish and confirmed by the slide
agglutination test against anti-T8 serum.
Virulence for mouse
Neither mortality nor clinical disease symptom was observed in any mice challenged with strains T4, T8,
T27 and P2. The internal organs of the mice, sacrificed
after the experimental period, seemed apparently nor
mal, and no expected bacteria were re-isolated from any

Virulence properties of Aeromonas sp. T8 group 155
Table 2. Virulence and clinical symptoms expressed by the fish challenged with Aeromonas sp. strain T8
of the mice. When mice were challenged with A.
hydrophila ATCC 7966T, 20% mortality was observed.
The kidney, liver, spleen and other internal organs of the
infected mice became pale, and slight proliferation of
bacteria was found in the organs of infected
mouse. The internal organs of the surviving mice
seemed normal, and no bacterium was re-isolated from
the internal organs.
Enzymatic and hemolytic activities of bacterial cells
Exoenzymatic activities of bacterial cells based on
the ratio of the hydrolytic halo zone diameter to colony
diameter are illustrated in Table 3. All strains of the
Aeromonas sp. T8 group showed moderate to strong
hydrolysis activity against casein, gelatin, hemoglobin
and lipid. These strains showed weak hydrolytic activ
ity against albumin, and no activity against elastin and
phospholipid. Exoenzyme activities exhibited by A.
hydrophila ATCC 7966T and A. caviae ATCC 15468T
were very similar to those of the strains of Aeromonas
sp. T8 group except for hydrolysis against albumin,
which was negative in both strains.
All strains of the Aeromonas sp. T8 group exhibited
hemolytic activity against all of the erythrocytes except
sheep erythrocytes, which was susceptible only to strain
P2 (Fig. 1). Among the strains, Thailand strains (T4, T8
and T27) showed a clear hydrolytic zone in blood agar
plates, hence categorized as ƒÀ-hemolytic. On the other
hand, the Philippines strain (P2) showed an opaque,
incomplete type of hemolysis in blood agar plates, and
therefore was considered as ƒ¿-hemolytic strain. A.
hydrophila ATCC 7966T showed ƒÀ-hemolytic activity for
all of the erythrocytes tested.
Protease activity of ECPs The protease activities of ECPs of the Aeromonas
sp. T8 group and the reference strains are summarized in Table 4. Among the experimental strains, strain P2 had the highest activity (2865 units/mg protein) and strain T4 had the lowest activity (282 units/mg
protein). Strains T8 and T27 produced 461 and 447 protease units/mg protein, respectively. The protease activities of A. hydrophila ATCC 7966T and A. caviae ATCC 15468T were 325 and 171 units/mg, respectively.
The protease activity of all experimental and reference strains was almost completely inhibited by heating
Table 3. Protease, lipase and phospholipase activities of bacterial cells at 25℃

156 M. M. Rahman, T. Somsiri, K. Tajima and Y. Ezura
at 80•Ž for 15 min (Table 4). When ECPs were heated
at 60•Ž for 15 min, protease activities of A. hydrophila
ATCC 7966T and A. caviae ATCC 15468T were inhibited
92 and 80%, respectively, but the activities of the
Aeromonas sp. T8 group remained stable. Protease
activities of the strains T4, T8, and T27 were inhibited
38, 43 and 44%, respectively, by EDTA treatment and
87, 88 and 88%, respectively, by PMSF treatment.
Both EDTA and PMSF inhibited the activity of strain P2
at 19 and 24%, respectively. PMSF inhibited 91% of
the protease activity of A. hydrophila ATCC 7966T but
EDTA had little effect (9%). A. caviae ATCC 15468T
protease possessed similar characters exhibited by theThailand strains (T4, T8 and T27).
Hemolytic activity of ECPsThe hemolytic activities of bacterial ECPs are sum-
marized in Fig. 2. Although hemolytic activities of ECPsvaried among the strains of the Aeromonas sp. T8 groupagainst different blood cells, generally higher activity wasobserved against fish blood cells compared to mamma-lian blood cells. On the other hand, hemolytic activity of
Fig. 1. Hemolytic activity of bacterial cells on blood agar plates, prepared by 5% blood cells from different fish and mammaliansources. Hemolytic activity was expressed as zone ratio (ratio of hemolytic zone diameter to colony diameter) . Strains T4,T8, T27 and P2 belong to the Aeromonas sp. T8 group and ATCC 7966T is the type strain of A. hydrophila .
Table 4. Protease activity of ECPs
*
% inhibition: (difference between the protease activity of untreated ECPs and treated
ECPs •€ activity of untreated ECPs)•~ 100
Abbreviations: EDTA: Ethylenediamine-N,N,N,N-tetraacetic acid
PMSF: Phenyl methyl sulphonyl fluoride
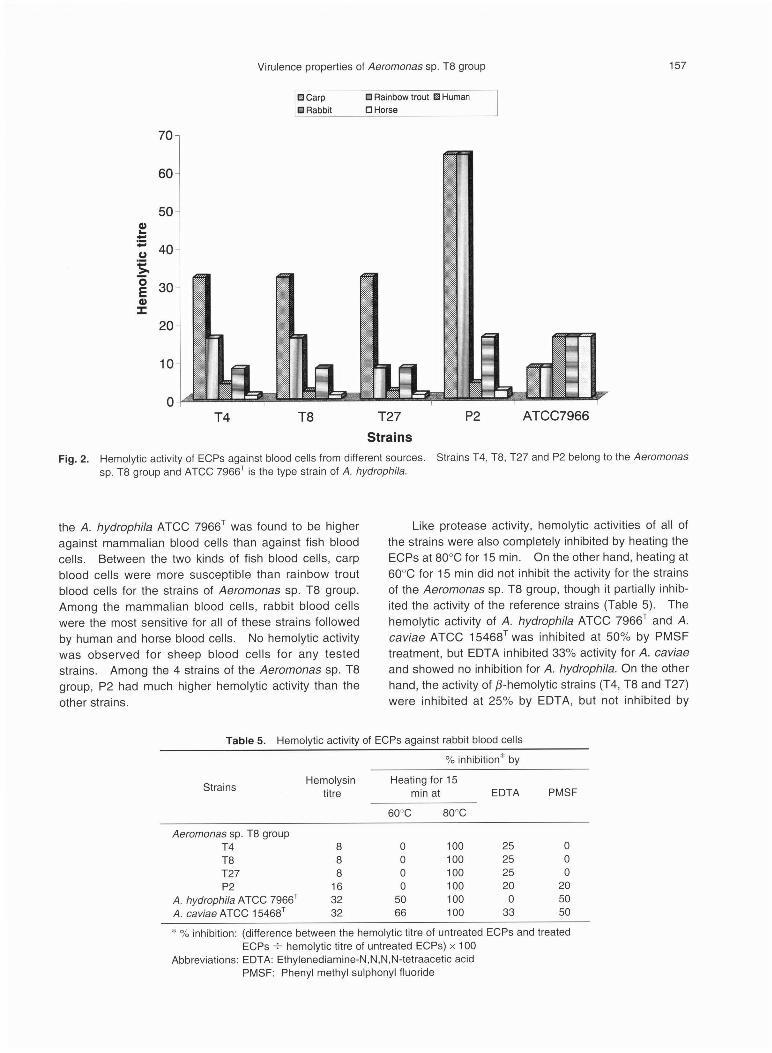
Virulence properties of Aeromonas sp. T8 group 157
the A. hydrophila ATCC 7966T was found to be higher
against mammalian blood cells than against fish blood
cells. Between the two kinds of fish blood cells, carp
blood cells were more susceptible than rainbow trout
blood cells for the strains of Aeromonas sp. T8 group.
Among the mammalian blood cells, rabbit blood cells
were the most sensitive for all of these strains followed
by human and horse blood cells. No hemolytic activity
was observed for sheep blood cells for any tested
strains. Among the 4 strains of the Aeromonas sp. T8
group, P2 had much higher hemolytic activity than the
other strains.
Like protease activity, hemolytic activities of all of
the strains were also completely inhibited by heating the
ECPs at 80•Ž for 15 min. On the other hand, heating at
60•Ž for 15 min did not inhibit the activity for the strains
of the Aeromonas sp. T8 group, though it partially inhib-
ited the activity of the reference strains (Table 5). The
hemolytic activity of A. hydrophila ATCC 7966T and A.
caviae ATCC 15468T was inhibited at 50% by PMSF
treatment, but EDTA inhibited 33% activity for A. caviae
and showed no inhibition for A. hydrophila. On the other
hand, the activity of ƒÀ-hemolytic strains (T4, T8 and T27)
were inhibited at 25% by EDTA, but not inhibited by
Fig. 2. Hemolytic activity of ECPs against blood cells from different sources. Strains T4, T8, T27 and P2 belong to the Aeromonas
sp. T8 group and ATCC 7966T is the type strain of A. hydrophila.
Table 5. Hemolytic activity of ECPs against rabbit blood cells
* %
inhibition: (difference between the hemolytic titre of untreated ECPs and treated
ECPs •€ hemolytic titre of untreated ECPs) •~ 100
Abbreviations: EDTA: Ethylenediamine-N,N,N,N-tetraacetic acid
PMSF: Phenyl methyl sulphonyl fluoride

158 M. M. Rahman, T. Somsiri, K. Tajima and Y. Ezura
PMSF treatment.
Cytotoxi city of ECPs
Among the strains of the Aeromonas sp. T8 group,
only P2 showed evidence of cytotoxicity for all of the
tested cell lines. The cytotoxicity titres of the ECPs of
strain P2 for HeLa, BF-2, SNN-1, EPC, FHM, and RTG-
2 cell lines were 4, 8, 8, 16, 64 and 64, respectively.In
general, the degenerative changes manifested by the
ECPs were vacuolization, shrinking, dendritic elongation
of the cells, cell detachment and final monolayer
destruction. The cytotoxicity of the ECPs heated at
80℃ was completely inhibited for all tested cell
lines. A. hydrophila ATCC 7966T, which was used as a
positive control for the assay, showed cytotoxic effect against all the tested cell lines. The degenerative
changes were almost similar to that observed for the strain P2 against all tested cell lines.
Enterotoxicity of ECPs
No enterotoxic activity was observed in any of the samples from the strains belonging to the Aeromonas
sp. T8 group. No fluid accumulated in the intestinal tract, and diarrhoea was not observed. The culture supernatant of A. hydrophila ATCC 7966T cultured in any
of the media, gave positive response for enterotoxic assay: fluid accumulated in the intestinal tract, and diar
rhoea occurred in the infant mice. The culture supernatant of A. caviae ATCC 15468T exhibited enterotoxicity
(both fluid accumulation and diarrhoea) only when double strength glucose-free TSB was used as a growth medium.
Detection of virulence genes by PCR
No strains belonging to the Aeromonas sp. T8 group amplified the targeted aerolysin or AHCYTOEN genes by PCR amplification. Amplified PCR products were observed in A. hydrophila ATCC 7966T for both genes.
A. veronii biotype sobria ATCC 9071T was positive for AHCYTOEN but negative for aerolysin gene, whereas A. caviae ATCC 15468T was negative for both genes.
Discussion
EUS is a severe disease of freshwater and some brackishwater fish that has extended rapidly in South and Southeast Asia over the past 25 years (Supranee
and Roberts, 1999). Although the aetiology of EUS is considered to be complex, recently, a fungus
Aphanomyces invadans has been reported as a primary causative agent and bacteria including Aeromonas spp.
are considered to be a secondary invader of the disease
(Supranee and Roberts, 1999). However, Aeromonas spp. is frequently isolated from EUS-affected fish
(Subasinghe et al., 1990; Roberts et al., 1990; lqbal et al, 1998). Some reports suggested that Aeromonas
spp. might contribute to the pathogenesis of the disease
(Costa and Wejeyaratne, 1989). Since several species
of Aeromonas are virulent for fish, and possess severa
l virulent properties, the role of Aeromonas spp. when
exist in EUS-affected fish is still unclear. The
Aeromonas sp. T8 is a newly identified species and all
strains of the group were isolated from EUS-affected
fish. Thus, whether the Aeromonas sp. T8 group has
any significance in EUS is a new research interest.
Moreover, as a newly identified microorganism, determi
nation of the pathological importance of the Aeromonas
sp. T8 group for fish and other animal, and screening of
the putative virulence properties are also essential.
This article describes the pathological importance of the
newly identified Aeromonas sp. T8 group and its putative
virulence properties.
The representative strain T8 showed a high degree
of virulence for silver barb, striped snakehead and tilapia
with median lethal doses of 1.5×105, 3.2×105 and 2.9×
105 CFU/fish, respectively. Hemorrhagic and necrotic lesions in the external tissue were observed in the artifi
cially infected fish. The lesions seemed apparently similar to the lesions usually observed in EUS-affected
fish. On the other hand, all strains of the Aeromonas
sp. T8 group failed to bring about any disease signs in the challenged mice, which suggests that these strains
are non-pathogenic for mammals. Mateos et al (1993) found a group of A. hydrophila isolates collected from fish farm water that were virulent for rainbow trout Oncorhynchus mykiss but did not cause any disease
signs in mice injected with the isolates. They assumed that the prolonged presence of these strains in aquatic environments might lead to loss or decrease in the viru
lence of those strains for homoeothermic animals. However, it is important to note that motile aeromonads
have long been considered as opportunistic pathogens. They have an affinity to cause disease in immune-com-
promised hosts. The present study is the first attempt on pathogenesis of the newly identified species the
Aeromonas sp. T8 group. More studies are needed to determine the pathological potency of the species for homoeothermic animals.
The exoenzyme activities of the live bacterial cells evaluated by the plate assay method showed that the strains of the Aeromonas sp. T8 group and reference
strains were positive for lipase and protease activity. For protease activity, all strains showed hydrolysis of
casein, gelatin and hemoglobin, but none of them hydrolyzed elastin. The extracellular proteases are considered to play an important rule in the pathogenesis of
Aeromonas spp. (Thune et al., 1982b; Tajima et al., 1983; Kanai and Wakabayashi, 1984). Although the effect of individual protease to virulence is unknown,
caseinase and elastase activities are frequently reported to co-relate with the virulence of Aeromonas spp. (Thune et al., 1982a; Esteve et al., 1995; Cascon et al.,

Virulence properties of Aeromonas sp. T8 group 159
2000). We assume that caseinase of the experimenta
l strains play an important role in the virulence of
fish. Other proteases like gelatinase and albuminase
probably provide nutrients for the growth of bacteria in
the host body by degrading host tissue protein. When
the representative strain T8 was injected to tropical fish,
necrotic lesion was observed in the muscles adjacent to
the injection site. We guess that extracellular protease
was responsible for tissue necrosis, Thune et al.
(1982a) reported that concentrated culture filtrate of A.
hydrophila that possessed proteolytic activity was lethal
and produced dermonecrotic symptom to channel catfish
lctalurus punctatus. This report supports our suspicion
. In this study, all strains of the Aeromonas sp. T8
group exhibited hemolytic activity against a wide range
of erythrocytes. The hemolytic titers determined using
different fish and mammalian blood cells were found to
vary among the strains. However, the ECPs of all
strains of the Aeromonas sp. T8 group showed higher
hemolytic activity to fish blood cells compared to mam
malian blood cells. Since the strain T8 was virulent for
different fish but all strains were non-virulent for a mam
mal, it can be presumed that there is a positive correla
tion between the virulence for fish and hemolytic activity
against fish erythrocytes. Esteve et al. (1995) also
found that fish virulent strains of the species A.
hydrophila and A. jandaei secreted hemolysins against
fish erythrocytes and concluded that hemolysins against
fish erythrocytes might play an important role in the
spread of the disease in eels. Moreover, we assume
that hemolysin was also responsible for hemorrhages
and necrosis observed in the fish artificially infected by
Aeromonas sp. strain T8. Allan and Stevenson (1981)
reported that ECP of protease-deficient but hemolysin
positive strain of A. hydrophila produced hemorrhages in
internal organs, excess fluid in the body cavity and lesion
in external muscles upon injection challenge to speckled
trout Salvelinus fontinalis. This report indicates the role
of hemolysin in the infection of fish and also justifies our
assumption.
The protease and hemolytic activities of experimen
tal and reference strains were completely inactivated by
heating the ECPs at 80℃ for 15 min, indicating that pro
tease and hemolysin of the strains are thermo
labile. Protease activity of the strains T4, T8, and T27
was highly inhibited by PMSF treatment and moderately
inhibited by EDTA treatment, indicating that these strains
produce serine protease as the major protease compo
nent and also produce some metalloprotease.
Protease activity of strain P2 was inhibited by both
PMSF and EDTA, indicating the presence of both serine
and metalloprotease, though the inhibition rate was
lower than the three Thailand strains. PMSF highly
inhibited the protease activity of A. hydrophila ATCC
7966T, but EDTA had little effect, suggesting that serine
protease is the major protease of the strain, which also
produces a small amount of metalloprotease. Protease of A. caviae ATCC 15468T possessed similar characters
exhibited by the Thailand strains (T4, T8 and T27). The
inhibition patterns of the hemolytic activities of A
. hydrophila ATCC 7966T and A. caviae ATCC 15468T were found very similar to the protease activity of the
strains. However, the activity of β-hemolytic strains
(T4, T8 and T27) was partially (25%) inhibited by
EDTA, but no inhibition was detected by the PMSF
treatment. Thus, we assume, these strains probably
possess a novel hemolysin enzyme, which is different
from the hemolysins of A. hydrophila and A. caviae.
Although extracellular products of aeromonads are
often found to possess cytotoxicity against different cell
lines, there is probably little or no correlation with
virulence. Santos et al. (1988) reported that ECPs
obtained from both pathogenic and nonpathogenic A
. hydrophila strains displayed cytoxicity to different ce
ll lines, whereas some virulent A. sobria and A. caviae
strains did not produce any cytotoxic effect. Paniagua et al
. (1990) also reported the lack of correlation
between cytotoxic activity and virulence of Aeromonas
spp. In the present study, only strain P2 showed cyto
toxicity against all cell lines tested. Strain P2 also
exhibited high protease activity compared to other
strains of the Aeromonas sp. T8 group, which probably
enabled the strain to possess cytotoxic activity.
However, involvement of the protease in cytotoxicity is
yet to be known.
Clinical and environmental strains of motile
Aeromonas spp. are also known to produce extracellular
enterotoxin, an important virulence factor suspected to
be related with gastrointestinal diseases of humans. In
the present study, we used three types of growth media
for preparation of culture supernatant to examine
whether growth medium influences the enterotoxic
activity, because Namdari and Bottene (1990) reported
that A. caviae produced cytotoxin and enterotoxin only in
selected media such as glucose-free tryptic soy broth.
However, the present strains grown in any media did not
show enterotoxicity, although A. hydrophila ATCC 7966T
produced enterotoxin irrespective of the growth media
and A. caviae ATCC 15468T only in a selected medium
. Aerolysin is the well-characterized ECP responsible
for β-hemolysis exhibited by Aeromonas spp. It is con
sidered an important marker for virulence of motile
Aeromonas spp. Chakraborty et al. (1987) presented
genetic evidence that the structural gene aerA (pro
aerolysin) is found in all members of this genus. Using
a primer set, Santos et al. (1999), Gonzalez-Serrano et al
. (2002) and Gonzalez-Rodriguez et al. (2002)
detected the gene in several Aeromonas spp. strains
recovered from freshwater fish, cold-smoked fish and
diarrhoea of human. The cytolytic enterotoxin gene
(AHCYTOEN) in A hydrophila has been reported as a
multivirulence gene involved in lethality (in mice),

160 M. M. Rahman, T. Somsiri, K. Tajima and Y. Ezura
hemolysis, cytotoxicity and enterotoxicity, which are the
established virulence properties of Aeromonas spp.
(Chopra et al., 1993). Various strains of aeromonads from several geno-species viz., A. hydrophila, A.
bestiarum, A. salmonicida,A. caviae, A. eucrenophila,A.
sobria, A. veronii biotype sobria, A. veronii biotype
veronii, A. encheleia and A. torta were found to hold the
AHCYTOEN gene by investigation with the primer com
bination strategy of Kingombe et al. (1999). Rahman et
al. (2002) also detected the gene in several strains of A.
veronii biotype sobria isolated mostly from EUS-affected
fish and also from environmental and human clinical
samples. In the present study, we did not detect either
gene in the strains of the Aeromonas sp. T8 group. We suspect that the strains of the Aeromonas sp. T8 group
do not possess either gene.
In summary, the newly identified Aeromonas sp. T8
group was detected as a potential pathogen for tropical fish. Extracellular proteases, especially caseinase and
hemolysins are suspected to play a role in the virulence
of the species to fish. However, the role of specific
enzyme (s) is further object to be studied. Moreover,
the role of the Aeromonas sp. T8 group, when exists in
EUS-affected fish is also further object to be studied.
Acknowledgements
The authors would like to express their sincere thanks to Prof. M. Yoshimizu, Hokkaido University,
Japan for providing cell lines and valuable suggestions
for the study. The first author also acknowledges the
Japanese Ministry of Science, Education, Culture and
Sports for his scholarship support.
References
Allan, B. J. and R. M. W. Stevenson (1981): Extracellular virulence factors of Aeromonas hydrophila in fish. Can. J.
Microbiol., 27, 1114-1122.Cascon, A., J. Yugueros, A. Temprano, M. Sanchez, C.
Hernenz, J. M. Luengo and G. Naharro (2000): A major secreted elastase is essential for pathogenicity of Aeromonas
hydrophila. Infect. Immun., 68, 3233-3241.Carnahan, A. M. and M. Altwegg (1996): Taxonomy. In:"The
genus Aeromonas" (ed. by B. Austin, M. Altwegg, P. J. Gosling, and S. Joseph). John Wiley & Sons Ltd., Chichester, pp.1-38.Chakraborty, T., B. Huhle, H. Hof, H. Bergbauer and W. Goebe
l (1987): Marker exchange mutagenesis of the aerolysin determinant in Aeromonas hydrophila demonstrates the role of aerolysin in A. hydrophila-associated systemic infections. Infect. Immun., 55, 2274-2280.Chopra, A. K., C. W. Houston, J. W. Peterson and G. F. Jin
(1993): Cloning, expression and sequence analysis of a cytolytic enterotoxin gene from Aeromonas hydrophila. Can. J. Microbiol., 39, 513-523.Costa, H. H. and M. J. S. Wejeyaratne (1989): Epidemiology of
the epizootic ulcerative syndrome occurring for the first time among fish in Srilanka. J. Appl. Ichthyol., 1, 48-52.
Esteve, C. (1995): Numerical taxonomy of Aeromonadaceae
and Vibrionaceae associated with reared fish and surrounding fresh and brackish water. Syst. Appl. Microbiol.,
18,391-402.Esteve, C., C. Amaro, E. Garay, Y. Santos and A. E. Toranzo
(1995): Pathogenicity of live bacteria and extracellular products of motile Aeromonas isolated from eels. J. Appl.
Bacteriol., 78, 555-562.Gonzalez-Rodriguez, M. N., J. A. Santos, A. Otero and M. L. G.
Lopez (2002): PCR detection of potentially pathogenic aeromonads in raw and cold-smoked freshwater fish. J.
Appl. Microbiol., 93, 675-680.Gonzalez-Serrano, C. J., J. A. Santos, M. L. G. Lopez and A.
Otero (2002): Virulence markers in Aeromonas hydrophila and A. veronii biover sobria isolated from freshwater fish
and from a diarrhoea case. J. Appl. Microbiol, 93, 414-419.
Gudmundsdottir, B. K. (1996): Comparison of extracellular proteases produced by Aeromonas salmonicida strains, isolated from various fish species. J. Appl. Bacteriol, 80,
105-113.Gunnlaugsdottir, B. and B. K. Gudmundsdottir (1997): Pathoge
nicity of atypical Aeromonas salmonicida in Atlantic salmon
compared with protease production. J. Appl. Bacteriol., 83, 542-551.Huys, G., R. Coopman, P. Janssen and K. Kersters (1996):
High resolution genotypic analysis of the genus Aeromonas by AFLP fingerprinting. Int. J. Syst. Bacteriol., 46, 572-
580.Iqbal, M. M. (1999): Taxonomical and pathological studies on
motile Aeromonas species isolated from fish with epizootic ulcerative syndrome in Southeast Asian countries. Mem.
Fac. Fish. Hokkaido Univ., 46, 103-154.
Iqbal, M. M., K. Tajima, T. Sawabe, K. Nakano and Y. Ezura (1998): Phenotypic and genotypic identification of motile aeromonads isolated from fish with epizootic ulcerative
syndrome (EUS) in Southeast Asian countries. Fish Pathol., 33, 255-263.Janda, J. M. (1991): Recent advances in the study of taxonomy,
pathogenicity and infectious syndromes associated with the genus Aeromonas. Clin. Microbiol. Rev., 4, 397-410.Janda, J. M. and S. L. Abbott (1996): Human Pathogens. In:
"The genus Aeromonas" (ed. by B. Austin, M. Altwegg, P. J. Gosling, and S. Joseph). John Wiley & Sons Ltd., Chichester, pp.151-170.Janda, J. M., R. B. Clark and R. Brenden (1985): Virulence of
Aeromonas species as assessed through mouse lethality studies. Curr. Microbiol., 12, 163-168.Kanai, K. and H. Wakabayashi (1984): Purification and some
properties of protease from Aeromonas hydrophila. Bull. Jpn. Soc. Sci. Fish., 50, 1367-1374.Kingombe, C. I. B., G. Husy, M. Tonolla, M. J. Albert, J. Swings,
R. Peduzzi and T. Jemmi (1999): PCR detection, characterization and distribution of virulence genes in Aeromonas
spp. Appl. Environ. Microbiol., 65, 175-179.Krovacek, K., V. Pasquale, S. B. Baloda, V. Soprano, M. Conte
and S. Dumontet (1994): Comparison of putative virulence factors in Aeromonas hydrophila strains isolated from the
marine environment and human diarrheal cases in southern Italy. Appl. Environ. Microbiol., 60, 1379-1382.
Kozinska, A., M. J. Figueras, M. R. Chacon and L. Soler (2002):
Phenotypic characteristics and pathogenicity of Aeromonas genomospecies isolated from common carp (Cyprinus
carpio L.). J. Appl. Microbiol., 93, 1034-1041.Lowry, O. H., N. J. Rosebrough, A. L Farr and R. J. Randall
(1951): Protein measurement with the folin phenol reagent. J. Biol. Chem., 193, 265-275.Mateos, D., J. Anguita, G. Naharro and C. Paniagua(1993):

Virulence properties of Aeromonas sp. T8 group 161
Influence of growth temperature on production of extracellular virulence factors and pathogenicity of environmental
and human strains of Aeromonas hydrophila. J. Appl. Bacteriol., 74, 111-118.
Namdari, H. and E. J. Bottone (1990): Cytotoxin and enterotoxin
production as factors delineating enteropathogenicity of Aeromonas caviae. J. Clin. Microbiol., 28, 1796-1798.Paniagua, C., O. Rivero, J. Anguita and G. Naharro (1990)
: Pathogenicity factors and virulence for rainbow trout
(Salmo gairdneri) of motile Aeromonas spp. isolated from a river. J. Clin. Microbiol., 28, 350-355.Rahman, M., P. C. Navarro,I. Kuhn, G. Huys, J. Swings and R. Mollby (2002): Identification and characterization of patho
genic Aeromonas veronii biovar sobria associated with epizootic ulcerative syndrome in fish in Bangladesh. Appl.
Environ. Microbiol., 68, 650-655.Reed, L. J. and H. Muench (1938): A simple method of estimat
ing fifty percent endpoints. Am. J. Hyg., 27, 493-497.Roberts, R. J., G. N. Frerichs and S. D. Millar (1990): Epizootic ulcerative syndrome-the current position, In "Diseases in Asian aquaculture" (ed. By M. Shariff, R. P. Subasinghe and J. R. Arthur). Asian Fisheries society, Manila, Philip
pines, pp. 431-436.Rodriguez, L. A., A. E. Ellis and T. P. Nieto (1992): Purification and characterization of an extracellular metallo protease, serine protease and hemolysin of Aeromonas hydrophila
strain B32: all are lethal for fish. Microb. Pathog., 13, 17-24.
Santos, Y., A. E. Toranzo, J. L. Barja, T. P. Nieto and T. S. Villa (1988): Virulence properties and enterotoxin production of Aeromonas strains isolated from fish. Infect. Immun., 56,
3285-3293.Santos, J. A., C. J. Gonzalez, A. Otero and M. L. G. Lopez (1999): Hemolytic activity and siderophore production in different Aeromonas species isolated from fish. Appl.
Environ. Microbiol., 65, 5612-5614.
Shotts, E. B., T. C. Hsu and W. D. Waltman (1985): Extracellular proteolytic activity of Aeromonas hydrophila complex
. Fish Pathol., 20, 37-44.Subasinghe, R. P., L. P. Jayasinghe, K. S. W. Balasuriya and M. Kulathilake (1990): Preliminary investigations into the
bacterial and fungal pathogens associated with the ulcerative fish disease syndrome in Sri Lanka, In "The second
Asian fisheries forum" (ed. by R. Hirano and I. Hanyu), Asian Fisheries Society, Manila, Philippines, pp. 655-657.Sugita, H., T. Nakamura, K. Tanaka and Y. Deguchi (1994)
: Identification of Aeromonas species isolated from freshwater fish with microplate hybridization. Appl. Environ.
Microbiol., 60, 3036-3038.Sugita, H., K. Tanaka, M. Yoshinami and Y. Deguchi (1995):
Distribution of Aeromonas species in intestinal tracts of river fish. Appl. Environ. Microbiol., 61, 4128-4130.Supranee, C. and R. J. Roberts (1999): Pathology and histopa
thology of epizootic ulcerative syndrome. Aquatic Animal
Health Research Institute, Bangkok, Thailand, 33 p.Tajima, K., T. Takahashi, Y. Ezura and T. Kimura (1983): Stud
ies on the virulent factors produced by Aeromonas salmonicida, a causative agent of furunculosis in salmonidae-l, Purification of the extracellular protease of Aeromonas salmonicida Ar-4 (EFDL). Bull. Fac. Fish. Hokkaido. Univ., 34, 104-110.Thune, R. L., T. E. Graham, L. M. Riddle and R. L. Amborski
(1982a): Extracellular products and endotoxin from Aeromonas hydrophila: effects on age 0 channel catfish
. Tran. Am. Fish. Soc., 3, 404-408.Thune, R. L., T. E. Graham, L. M. Riddle and R. L. Amborski
(1982b): Extracellular proteases from Aeromonas hydrophila: partial purification and effects on age 0 channel
catfish. Tran. Am. Fish. Soc., 3, 749-754.Valera, L. and C. Esteve (2002): Phenotypic study by numerica
l taxonomy of strains belong to the genus Aeromonas. J. Appl. Microbiol., 93, 77-95.





























