Embed Size (px)
Citation preview

VIA DE LAS PENTOSAS
FOSFATO

La Vía de las Pentosas Fosfato


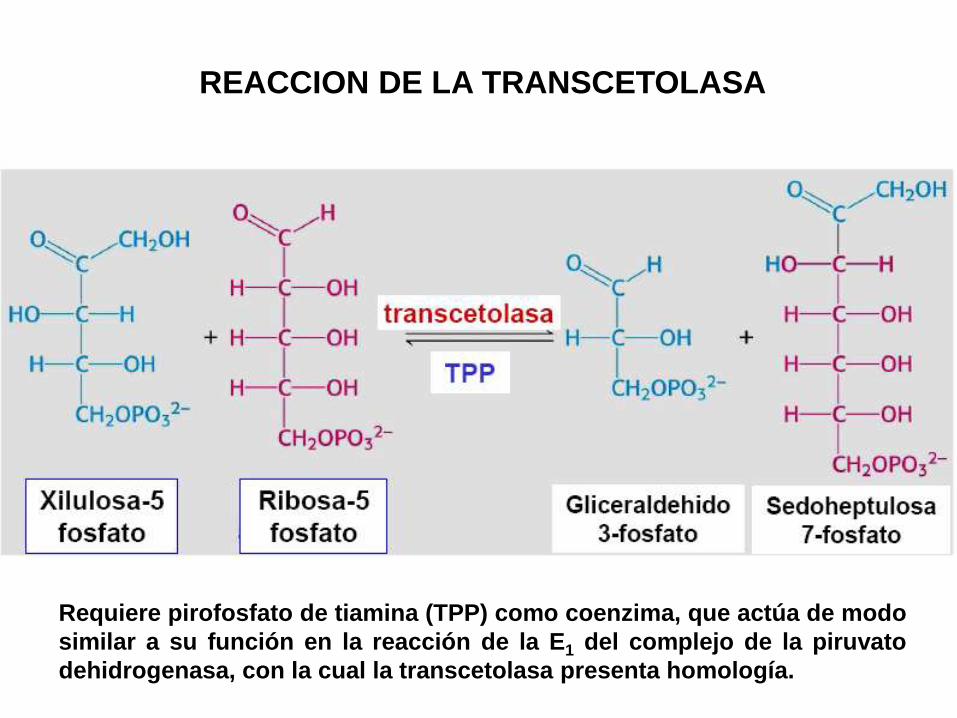
REACCION DE LA TRANSCETOLASA
Requiere pirofosfato de tiamina (TPP) como coenzima, que actúa de modo
similar a su función en la reacción de la E1 del complejo de la piruvato
dehidrogenasa, con la cual la transcetolasa presenta homología.

REACCION DE LA TRANSALDOLASA
No requiere coenzima. La reacción transcurre con formación de una base
de Schiff entre el carbonilo de la cetosa sustrato y un e-NH2 de lisina en la
transaldolasa, que tiene homología con la aldolasa de la glucólisis.

SEGUNDA REACCION DE LA TRANSCETOLASA


ulosa
sphate
ose
sphate

LA VIA DE LAS PENTOSAS FOSFATO EN
TRYPANOSOMA CRUZI.
Las siete enzimas de la vía están presentes en los
cuatro estadíos del ciclo biológico del parásito.
Los genes que codifican a la 6-fosfogluconolactonasa,
la 6-fosfogluconato dehidrogenasa, la ribosa 5-fosfato
isomerasa, la transaldolasa y la transcetolasa están presentes
como copia única por genoma haploide, en el clon CL Brener.
La ribulosa 5-fosfato epimerasa está codificada por dos
genes diferentes, y la glucosa 6-fosfato dehidrogenasa por un
número elevado de genes similares.
Las siete enzimas de la vía fueron clonadas y
expresadas como proteínas recombinantes activas en
Escherichia coli.

Al menos tres enzimas de esta vía pueden llegar a
convertirse en blancos adecuados para la quimioterapia de la
Enfermedad de Chagas: la glucosa 6-fosfato dehidrogenasa,
por su papel en la protección contra el stress oxidativo; la 6-
fosfogluconato dehidrogenasa, por el efecto letal de su
inhibición en otras células y sus diferencias cinéticas con la
enzima de mamífero; y la ribosa 5-fosfato isomerasa, por su
condición de exclusiva del parásito, al ser de Tipo B, ausente
en los mamíferos.
Estas características justifican la búsqueda de
inhibidores selectivos, que puedan servir como base para el
desarrollo de nuevas drogas antiparasitarias.

La enzima es inducida en trypomastigotes metacíclicos de T. cruzi por el
stress oxidativo causado por peróxido de hidrógeno. Por el contrario, en el
caso de los epimastigotes, que normalmente no están expuestos al stress
oxidativo, no se observó inducción, y en cambio se encontró inactivación
parcial.
La glucosa 6-fosfato dehidrogenasa de T. cruzi
Estudios de otros autores indican que algunos inhibidores de la G6PDH son
capaces de inhibir el crecimiento de T. cruzi y de T. brucei.

Hay dos tipos no homólogos de Rpi: A y B.
La Rpi A está presente en eucariotas superiores, hongos, y
muchas bacterias y Archaeas. La Rpi B está presente casi
exclusivamente en procariotas, con la excepción de algunos
eucariotas inferiores, incluyendo algunos Protozoarios. Algunas
bacterias, como Escherichia coli, tienen ambos tipos.
El genoma de T. cruzi presenta dos ORFs que codifican
proteínas homólogas a la Rpi B y ninguna homóloga a la Rpi A.
Determinaciones de actividad y Western blots mostraron que la
enzima se expresa en los cuatro estadíos principales del parásito.
Dado que se ha demostrado que la actividad enzimática es
esencial en levadura y en E. coli, y no hay homólogos de la RpiB en
los mamíferos, esta enzima podría ser un buen blanco para
quimioterapia. Estudios recientes de otros autores sugieren que la
enzima sería esencial tanto en Leishmania infantum como en T.
brucei.
La Ribosa-5-P Isomerasa (Rpi) de T.cruzi.

Estructura de la TcRpiB con R5P
Resolución 1.8Å
A.L. Stern, A. Naworyta, J.J. Cazzulo, S.L. Mowbray. Structures of type B ribose 5-phosphate
isomerase from Trypanosoma cruzi shed light on the determinants of sugar specificity in the
structural family. FEBS J. , 2011; 278, 793 – 808.

FOTOSINTESIS
EL CICLO DE CALVIN
Las reacciones de la fase luminosa de la fotosíntesis, que
estudiaremos conjuntamente con la cadena de transporte de
electrones y la fosforilación oxidativa, para resaltar sus semejanzas y
diferencias, llevan a la producción de ATP y NADPH. Las reacciones
de la fase oscura de la fotosíntesis, que veremos ahora, utilizan ese
ATP y NADPH para la reducción del CO2 a hexosa, y varias de las
enzimas e intermediarios son comunes con la vía de las pentosas
fosfato.

ESQUEMA DEL CLOROPLASTO

ESQUEMA SIMPLIFICADO DE LAS REACCIONES LUMINOSAS DE LA
FOTOSÍNTESIS

Las
reacciones
oscuras de la
fotosíntesis:
El Ciclo de
Calvin.
Podemos considerar que tiene 3
etapas:
1) la fijación del CO2 sobre la
ribulosa 1,5 bis-fosfato para
dar 2 moléculas de 3-
fosfoglicerato.
2) la reducción del 3-
fosfoglicerato para dar
hexosas.
3) la regeneración de la ribulosa
1,5 bis-fosfato para continuar
fijando CO2.

Melvin Calvin descubríó el
producto primario de la
incorporación del CO2
utilizando 14CO2. Iluminando
un cultivo de algas en
presencia de este compuesto
radioactivo por 5 seg. observó
que el 14CO2 se incorporó al 3-
fosfoglicerato. Pero después
de 60 seg, la radioactividad
estuvo presente en muchos
compuestos, que son los
intermedarios del Ciclo de
Calvin.

La reacción requiere Mg2+. Además debe formarse un carbamato por reacción de otra molécula de CO2 con un
grupo e-NH2 de lisina, catalizada por una rubisco activasa. Si el catión se une al O2, da una reacción de oxigenasa,
que produce 3-fosfoglicerato y fosfoglicolato. 2 moléculas de este se procesan para reciclar 3 C; el cuarto se
pierde como CO2; como se consume O2, el proceso se denomina fotorrespiración.

La carboxidismutasa: La cadena L contiene un centro catalítico y un centro
regulador. La cadena S estimula la acción catalítica de L.


Transcetolización


Ciclo de Calvin

BALANCE DEL CICLO DE CALVIN
Para sintetizar una hexosa, se requieren seis vueltas del ciclo de
Calvin, porque en cada una de ellas se reduce un átomo de C. Se
gastan 12 moléculas de ATP para fosforilar 12 moléculas de 3-
fosfoglicerato a 1,3-bisfosfoglicerato, y 12 moléculas de NADPH
para reducirlas a gliceraldehído 3-fosfato. Otras 6 moléculas de
ATP se gastan para regenerar la ribulosa 1,5-bisfosfato. Por lo
tanto, se consumen 3 moléculas de ATP y 2 de NADPH para la
conversión del CO2 hasta el nivel de hexosa

REGULACIÓN DEL CICLO DE CALVIN
La actividad de la rubisco aumenta notablemente con la luz.
La formación del carbamato es favorecida por el pH alcalino
y la alta concentración de Mg2+ en el estroma, debidos al
bombeo de protones desde el estroma al espacio tilacoidal y
a la liberación del Mg2+ desde este último para compensar la
entrada de protones.
Además la rubisco, la gliceraldehido 3-fosfato DH (GADPDH),
la sedoheptulosa 1,7-bisfosfatasa y la ribulosa 5-fosfato
quinasa (R5PK) están reguladas por la reducción de puentes
disulfuro a SH libres, que las activa, producida por la
tioredoxina reducida, a su vez regenerada por la ferredoxina
reducida, proveniente de las reacciones luminosas. Además,
la GADPDH y la R5PK en la oscuridad se asocian a la
proteína CP12 que las mantiene inactivas, y el NADPH las
libera, activándolas.

La síntesis de la sacarosa
en el citosol y del almidón
en el cloroplasto.

LA VIA C4 DE LAS PLANTAS TROPICALES
(Via de Hatch y Slack)
La actividad de oxigenasa de la rubisco
aumenta mas rápidamente que la de
carboxilasa con la temperatura. Plantas
como la caña de azúcar evitan el exceso de
fotorrespiración aumentando la
concentración de CO2 en los cloroplastos de
las células de la vaina del haz. Esto se lleva
a cabo fijándolo primero en las células del
mesófilo a través de la carboxilación del
PEP por la PEP carboxilasa. Se obtiene
oxaloacetato que se reduce a malato, el cual
es transportado a través de plasmodesmos
a la célula de la vaina del haz, donde se
decarboxila por una enzima málica NADP-
dependiente, liberando el CO2, que puede
alcanzar en los cloroplastos una
concentración 20 veces mayor que en la
célula del mesófilo. Esto reduce al mínimo
la pérdida de energía por la fotorrespiración,
pero implica un gasto mayor de ATP, que
resulta de 30 por molécula de hexosa.






























![[6]Gluconeogénesis y via de las pentosas fosfato](https://img.dokumen.tips/doc/110x75/5572021e4979599169a2fe2d/6gluconeogenesis-y-via-de-las-pentosas-fosfato.jpg)