Embed Size (px)
Citation preview

UNIVERZA V LJUBLJANI
PEDAGOŠKA FAKULTETA
Maja Dacar
VEDENJSKI ODZIV JAMSKIH IN
POVRŠINSKIH VODNIH OSLIČKOV (Asellus
aquaticus) NA VODNI TOK
Diplomsko delo
Ljubljana, 2017


UNIVERZA V LJUBLJANI
PEDAGOŠKA FAKULTETA
DVOPREDMETNI UČITELJ BIOLOGIJA - GOSPODINJSTVO
Maja Dacar
VEDENJSKI ODZIV JAMSKIH IN
POVRŠINSKIH VODNIH OSLIČKOV (Asellus
aquaticus) NA VODNI TOK
Diplomsko delo
Mentor: dr. Peter Trontelj
Ljubljana, 2017


Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
III
ZAHVALA
Najprej se zahvaljujem svojemu mentorju, dr. Petru Trontlju, ki me je tekom lovljenja vodnih
osličkov, izvajanja eksperimentov ter pisanja strokovno vodil ter mi pomagal pri samem
procesu ustvarjanja diplomskega dela.
Zahvaljujem se dr. Simoni Prevorčnik, ki mi je pomagala pri določanju samcev in samic
vodnih osličkov, s čimer mi je olajšala delo pri ločevanju.
Zahvaljujem se tudi vsem osebam, ki so mi v času pisanja diplomskega dela nudili podporo,
motivacijo in pomoč.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
IV

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
V
POVZETEK
Veliko je vprašanj, kaj vpliva na nastanek novih vrst. Eno je predvsem pojav razlik znotraj
določene vrste, kjer se je del izoliral od skupine in nadaljeval svojo evolucijo v smeri
nastanka nove vrste. Ena od teh razlik se med površinskimi in jamskimi vodnimi oslički
(Asellus aquaticus) kaže kot moč oprijemanja na podlago.
Ugotavljanje teh razlik je bilo izvedeno s pomočjo metode poskusa, in sicer na jamskih
vodnih osličkih iz Planinske jame in Zelških jam ter na površinskih vodnih osličkih iz
Ljubljane (okolica Biotehniške fakultete) ter Planinskega polja. S poskusom sem ugotavljala
mejne hitrosti toka, pri katerih se vodni oslički ne morejo več obdržati na podlagi. Pri tem
sem uporabila v ta namen konstruirano napravo (PVC kanal s sanmi). Na saneh je podlaga iz
grobega papirja, na katero se postavi vodnega oslička. Sani se s pomočjo vrvi na eni strani in
elastiko na drugi napne in pri določeni dolžini izpusti. Potovanje sani vzdolž naprave simulira
vodni tok, hitrost pa določa začetna napetost elastike, ki vleče sani. Nato sem zapisala
raztezek elastike, pri katerem se je osebek izpustil od podlage. Po večkratnih ponovitvah sem
izračunala povprečne hitrosti vodnega toka ob izpustu osebkov.
Predhodno sem opravila meritve hitrosti potovanja sani brez vodnih osličkov in izrisala grafa
hitrosti v odvisnosti od raztezka za šibkejšo in močnejšo elastiko. Rezultate meritev sem
predstavila v obliki škatlastega diagrama, razlike v oprijemu med skupinami pa sem preverila
s pomočjo ne-parametričnih statističnih testov. Ti so pokazali, da se med površinskimi in
jamskimi vodnimi oslički pojavljajo pomembne razlike, znotraj ene in druge skupine pa
razlik ni.
Rezultati so pokazali, da so jamski vodni oslički veliko bolje prilagojeni na oprijemanje
podlage ob nenadnih povečanjih hitrosti vodnega toka. Te ugotovitve bodo pomagale pri
razumevanju vedenjskih razlik, ki lahko skupaj z drugimi dejavniki evolucije vplivajo na
nastajanje novih podzemnih vrst kljub neposredni bližini površinske predniške populacije.
KLJUČNE BESEDE
Asellus aquaticus, ekosistem, podlaga, vodni tok, jame, podzemeljske vode, nastanek vrst

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
VI
ABSTRACT
There are many questions regarding what influences the emergence of new species. Firstly
and above all, is the appearance of differences within a certain specie, where a certain part is
isolated from the group and continues its own evolution. One of these differences appear
between the surface- and cave-dwelling Asellus aquaticus, as the ability to hold on to their
surface.
The discovery of these differences was carried out using a method of experiment, namely on
the cave-dwelling Asellus aquaticus from Planinska jama and Zelške jame and on surface-
dwelling Asellus aquaticus from Ljubljana (near Biotehniška fakulteta) and Planinsko polje.
With the experiment I tried to determine the maximum limit of the current velocities in which
Asellus aquaticus can no longer hold on to their ground. For this purpose I had to built a
contraption (PVC tube with a »slide«) that would allow me to do the experiment. The slide is
wrapped with coarse paper, so that it represents a surface on which the organsms can hold on
to. After an Asellus aquaticus is placed on that surface, the slide is pulled by a string on one
side. On the other side, a different pull of the slide causes different tensions to the elastic,
therefore causing different velocities. The movement of the slide along the contraption
simulates the water flow, in where the initial tension of the elastic, determines the speed.
Then I measured and recorded the expansion of the elastic string, each time the specimen let
go of the surface. After many repetitions, I calculated the average speed of the water current
during the release.
Before the actual experiment, I measured the speed the slide without Asellus aquaticus and
created graphs of speed in dependence to the elongation of the weaker and stronger elastic. I
presented the results of the measurements in form of a box plot and checked the differences
in the grip between groups by using nonparametric statistical tests. Those have shown that
there are significant differences between surface- and cave-dwelling Asellus aquaticus and no
differences within the populations themselves.
The results have shown that cave-dwelling Asellus aquaticus are better adapted to gripping to
the substrate, if the intensity of the water flow suddenly increases. These findings will help
understand the behavioral differences which, together with other factors of evolution, affect
the proces of formation of new underground species, despite the close vicinity to the surface
ancestor populations.
KEY WORDS
Asellus aquaticus, ecosistem, substratum, current velocity, caves, groundwater, speciation

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
VII

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
VIII
KAZALO VSEBINE
1 UVOD .................................................................................................................................................. 1
1.1 OPIS VODNIH OSLIČKOV ........................................................................................................ 2
1.2 MODELNI ORGANIZMI ............................................................................................................ 3
1.3 EKOLOŠKA SPECIACIJA .......................................................................................................... 3
1.4 JAMSKI EKOSISTEM ................................................................................................................. 4
1.5 RAZISKOVALNI PROBLEM IN HIPOTEZE ............................................................................ 4
2 MATERIALI IN METODE ................................................................................................................. 6
2.1 ANALIZIRANI VZORCI ............................................................................................................. 6
2.1.1 Opis vzorca ............................................................................................................................ 6
2.1.2 Postopek zbiranja podatkov ................................................................................................... 6
2.2 OPIS NAPRAVE .......................................................................................................................... 7
2.3 POSTOPEK OBDELAVE PODATKOV ..................................................................................... 8
3 REZULTATI........................................................................................................................................ 9
3.1 UMERITEV ELASTIK ................................................................................................................ 9
3.2 MERITVE MEJNIH HITROSTI ................................................................................................ 10
3.3 STATISTIČNI TESTI ................................................................................................................ 12
4 DISKUSIJA ....................................................................................................................................... 13
5 LITERATURA IN VIRI .................................................................................................................... 14
6 VIRI SLIKOVNEGA GRADIVA ..................................................................................................... 15
7 PRILOGA .......................................................................................................................................... 16

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
IX
KAZALO SLIK
Slika 1: Površinski (A) in jamski (B) vodni osliček. (Ž. Fišer, 2017). ................................................... 2
Slika 2: Skica delov telesa vodnih osličkov. ........................................................................................... 2
Slika 3: Slika samca (levo) in samice (desno) jamskih vodnih osličkov (Ž. Fišer, 2017). ..................... 3
Slika 4: Fotografija pripomočka. ............................................................................................................ 7
Slika 5: Skica naprave iz ptičje perspektive. ........................................................................................... 8
Slika 6: Približana fotografija "sani". ..................................................................................................... 8
Slika 7: Graf hitrosti vodnega toka v odvisnosti od razdalje raztega sani pri šibkejši elastiki. .............. 9
Slika 8: Graf hitrosti vodnega toka v odvisnosti od razdalje raztega sani pri močnejši elastiki. ............ 9
Slika 9: Škatlasti diagram hitrosti pri izpustu pri določenem kraju in spolu populacije. ...................... 11
KAZALO PREGLEDNIC
Preglednica 1: Število osebkov glede na spol ter kraj ulova. .................................................................. 6
Preglednica 2: Statistične vrednosti posameznih skupin vodnih osličkov. ........................................... 10
Preglednica 3: Test normalne porazdelitve (Shapiro-Wilk) in p-vrednosti podatkov........................... 12
Preglednica 4: Mann-Whitney test signifikantnosti med populacijami. ............................................... 12
Preglednica 5: Meritve in izračun hitrosti "sani" pri različnih odmikih pri šibkejši elastiki. ............... 16
Preglednica 6: Meritev in izračun hitrosti "sani" pri različnih odmikih pri močnejši elastiki. ............. 17
Preglednica 7: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinske jame pri
močnejši elastiki.................................................................................................................................... 19
Preglednica 8: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinske jame pri
šibkejši elastiki. ..................................................................................................................................... 20
Preglednica 9: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Zelške jame pri
močnejši elastiki.................................................................................................................................... 20
Preglednica 10: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Zelške jame pri šibkejši
elastiki. .................................................................................................................................................. 20
Preglednica 11: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinskega polja pri
šibkejši elastiki. ..................................................................................................................................... 21
Preglednica 12: Izračun posamezne povprečne hitrosti osebkov v populaciji iz okolice Ljubljana pri
šibkejši elastiki. ..................................................................................................................................... 21

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
X
SEZNAM KRATIC IN OKRAJŠAV
SZ …... severo-zahod
Npr. … na primer
N..........velikost vzorca
PJ …... Planinska jama
ZJ …... Zelške jame
L ……. Ljubljana
PP …... Planinsko polje
idr. ….. in drugi
n. l. ….. ni letnice
PVC .... Polivinilklorid (sintetični plastični polimer)
itd. …... in tako dalje
SLOVAR STROKOVNIH IZRAZOV
Speleobiološki laboratorij = Laboratorij namenjen gojenju in proučevanju jamskih živali
Reducirano = Odvzeto, zmanjšano ali omejeno
Morfološka struktura = Zgradba in oblika organizma
Specializirano = Usmerjena in pridobljena sposobnost na določenem področju ali
aktivnosti
Variacija = Različica, raznolikost
Ventralno = Trebušno
Dorzalno = Hrbtno
Spolni dimorfizem = Različnost v obliki, barvi ali drugi biološki lastnosti med spoloma
znotraj vrste
Adaptacija = Prilagoditev
Speciacija = Proces nastajanja novih vrst
Divergentna selekcija = Nastanek različnih populacij ali vrst iz enotne predniške
populacije ali vrste
Kompeticija = Tekmovanje med osebki za omejene vire (hrana, partner, prostor,…)
Predacija = Odnos med organizmi, kjer en napada in se prehranjuje z drugo (plenjenje)
Habitat = Življenjsko okolje organizmov
Selekcijski pritisk = Učinkovanje dejavnikov, ki povzročajo naravno selekcijo
Aklimacija = Navaditi se na iste ali enake pogoje (ista temperatura, hrana, tema…)
Signifikantno = Pomembno

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
1
1 UVOD
Ključno vlogo pri nastanku novih vrst ima naravna selekcija, vendar še vedno ne vemo veliko
o mehanizmu, ki vodi do speciacije. V mojem diplomskem delu raziskujem prav ta proces
ločitve dveh populacij, kot sta populaciji jamskih in površinskih vodnih osličkov. Zaradi
njihove enostavnosti in boljšega razumevanja biologije spadajo te živali pod modelne
organizme. Jamski vodni oslički imajo za razliko od površinskih sorodnikov številne
morfološke posebnosti, kot so reducirane oči, reduciran pigment, ki se kaže v beli
obarvanosti, podaljšanje okončin in prilagojene zaznavne strukture (Verovnik, Sket in
Trontelj, 2004). Zakaj določen organizem postane tak, veliko pove njegov ekosistem oziroma
interakcija, ki jo ima z njo. Ti sistemi na organizme, kot so vodni oslički, delujejo z
določenimi dejavniki, kot so podlaga in hitrost ali sunek vodnega toka.
V svojem diplomskem delu sem s poskusom preverjala zmožnost obstanka vodnih osličkov
na podlagi ob določenem sunku vode. Preučevala sem jamski populaciji iz Planinske jame
(območje kraja Planina) in Zelških jam (območje Rakovega Škocjana) ter površinski
populaciji iz Ljubljane (okolica Biotehniške fakultete) in Planinskega polja. Jamski populaciji
razločno kažeta morfološke spremembe povezane z življenjem v podzemeljskem okolju.
Manj znane od morfoloških značilnosti jamskih živali pa so njihove vedenjske prilagoditve.
V diplomskem delu sem se osredotočila na odziv živali na vodni tok. Pri površinskih vodnih
osličkih tok vode (reke) redko predstavlja usoden pojav, medtem ko jamske vodne osličke
lahko tok ponikalnice odnese na površje, kjer hitro propadejo. Na podlagi tega se pričakuje,
da je višja sposobnost kljubovanja vodnemu toku ena od pomembnih vedenjskih sprememb,
ki se zgodi v evoluciji podzemeljskih populacij vodnih osličkov.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
2
1.1 OPIS VODNIH OSLIČKOV
Vodni oslički ali Asellus aquaticus (Linnaeus, 1758) so enakonožni raki, ki jih najdemo po
celotni Evropi (Stahl idr., 2015). Sposobni so živeti v različnih okoliščinah. V Dinarskem
Krasu SZ Balkana je znanih 6 podvrst, med njimi tudi specializirane jamske (podzemne)
populacije. Kljub temu da se površinske in jamske populacije razlikujejo že po videzu, se za
odkrivanje genskih variacij med njima uporablja različne tehnike (Verovnik, Sket in Trontelj,
2004). Jamski vodni oslički imajo z razliko od površinskih sorodnikov številne morfološke
posebnosti, kot so reducirane oči in pigment (kar se kaže v beli obarvanosti), podaljšane
okončine in prilagojene zaznavne strukture. V naravi se ti dve populaciji ne srečujeta,
oziroma se to zgodi zelo redko, zaradi česar se populaciji težje križata. V ujetništvu se
križanje pojavi pogosteje, saj tam obstaja večja možnost razmnoževanja in proizvajanja
plodnih potomcev med jamskimi in površinskimi vodnimi oslički (Stahl idr., 2015).
Slika 1: Površinski (A) in jamski (B) vodni osliček. (Ž. Fišer, 2017).
Skupna lastnost površinskih in jamskih vodnih osličkov je sestava telesa. Osnovna delitev
telesne regije, ki so prisotne pri večini rakov, so glava, oprsje in zadek. V skupini izopodov
(raki enakonožci) se glava in oprsje združita v glavoprsje, ki je sestavljeno iz 6 členov glave
in 7 členov oprsja ter zadek, ki je sestavljen iz 6 členov. Ti segmenti so skupaj zaraščeni, zato
se jih po zgornji strani telesa ne loči med seboj. Vodni oslički so prepoznani še po njihovi
dorzo-ventralni sploščenosti (Fox, 2006).
Slika 2: Skica delov telesa vodnih osličkov.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
3
Majhnim organizmom, kot so vodni oslički, je včasih težko določiti spol. Imajo za nas dokaj
težje opazni spolni dimorfizem, vendar z natančnejšim ogledom značilnosti posameznega
vodnega oslička lahko določimo spol osebka. Značilnosti, ki ločujejo spol, so npr. širše
zgornje oprsje pri odrasli samici, manjši tretji par nožnih okončin in močnejši nožni krempelj
pri samcih, spremenjene spolne strukture pri samcih, itd. (Riehl, 2016). Za natančnejše
opazovanje se uporablja metoda opazovanja osebkov pod stereomikroskopom.
Slika 3: Slika samca (levo) in samice (desno) jamskih vodnih osličkov (Ž. Fišer, 2017).
1.2 MODELNI ORGANIZMI
Za izvajanje raziskav v biologiji in drugih področjih se uporabljajo tako imenovani modelni
organizmi. Njihova prilagoditev na določeno okolje omogoča razumevanje obnašanja in
delovanja organizmov v tem okolju.
Pod pojem modelni organizem razumemo kot ne-človeško vrsto, ki je znanstveno dobro
raziskana. Organizmi so priljubljeni predvsem zaradi dobrega poznavanja njihove biologije,
njihovih procesov, enostavnega vzdrževanje, razmnoževanja (veliko število mladičev),
raziskovanja in manipulacije, imajo kratki generacijski čas (čas med rojstvom in
reprodukcijsko sposobnostjo), so splošno razširjeni in so primerni za genske analize ter
študijo karakteristik in bolezni (Yourgenome, 2017).
Eden izmed modelnih organizmov jamskega okolja je prav vodni osliček. So odlični modeli,
na katerih se izvajajo raziskave adaptacije, prav zaradi njihovih prilagoditev na jamski svet.
Te prilagoditve so redukcija ali popolna izguba oči, depigmentacija telesa in oči, razvoj
specializiranih organov za okus in tip ter podaljševanje okončin (Baldwin in Beatty, 1941;
Culver in Pipan, 2009).
1.3 EKOLOŠKA SPECIACIJA
V današnjem času razumemo ključno vlogo naravne selekcije pri nastanku novih vrst, vendar
še vedno ostaja vprašanje o mehanizmih, ki vodijo do speciacije. V diplomskem delu
raziskujem dejavnike speciacije, ki bi lahko bili posledica prilagajanja populacij na različni
življenjski okolji. Imenovana ekološka speciacija predstavlja proces nastajanje novih vrst,

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
4
kjer je oviran genski pretok med populacijama zaradi njunega ekološkega razhajanja. Ta
speciacija vsebuje komponente, kot so ekološki viri ločevanja, oblike reproduktivne izolacije
ter genetski mehanizem, ki povezujejo oboje (Rundle in Nosil, 2005).
Na organizme, kot so vodni oslički, delujejo faktorji, kot so podlaga in hitrost vodnega toka.
Podlaga je eden izmed večjih dejavnikov okolja, v katerem organizmi živijo, saj predstavlja
njihov medij. Hitrost vodnega toka pa v povezavi z drugimi fizičnimi silami ustvarja pogoje
njihovega habitata (Peeters idr., 2002). Kljub neprijaznosti pogojev, se vodni organizmi
pojavijo na številnih območjih. Culver (1971) omenja, da »izpiranje« organizmov z vodnim
tokom na dolgi rok ne zmanjšuje njihovega števila. Ti se kaj kmalu ponovno kolonizirajo ali
se vrnejo nazaj proti toku, ko ta upade. Izjema so tiste živali, ki so se poškodovala ali bila
»izprana« čez mejo oziroma oviro vrnitve.
1.4 JAMSKI EKOSISTEM
Vodne sisteme delimo na površinske ali sladkovodne ter na jamske ali podzemeljske
ekosisteme. Prvi sistem je značilen po tem, da mu kljub raznolikosti v pogojih ne
primanjkujejo hranila, biogenih elementov (fosfor, nitrati in silikon), prostora, svetlobe,
biotskih dejavnikov in podobno. Omenjeni dejavniki velikokrat niso konstantni, temveč
nihajoči (Menshutkin, Rukhovets in Filatov, 2014). Kar pa je ravno nasprotno z jamskim
sistemom, ki nam kaže številne ekološke razlike.
Jamski sistemi so prostori znotraj zemeljske skorje, ki so po velikosti lahko zelo raznoliki ter
so napolnjeni bodisi z vodo, zrakom ali celo nafto (Culver in Pipan, 2009). Divergentna
selekcija je tam mogoča zaradi številnih in pomembnih ekoloških razlik, kot so: absolutna
tema, v kateri človek ne vidi ničesar; primanjkovanje hrane; pomanjkanje predatorjev in
kompeticije; visoka vlažnost in konstantnost okolja; stalno vlažna (kamnita) podlaga, nizka
koncentracija kisika v podzemnih vodah, itd. (Trontelj, 2012; Baldwin in Beatty, 1941,
Culver in Pipan, 2009). Pomemben abiotski dejavnik je tudi temperatura, ki je bolj ali manj
konstantna. Nihanje temperature je zgolj nekaj stopinj Celzija. Ker so določeni dražljaji
odsotni ali ne tako izraziti v jamah kot na površju, je pomembna iztočnica za razmnoževanje
prav spomladna poplavitev jame (Culver in Pipan, 2009).
1.5 RAZISKOVALNI PROBLEM IN HIPOTEZE
Eden od dejavnikov za naravno selekcijo, ki jo raziskujem v diplomskem delu, je sposobnost
oprijema na podlago ob trenutnem sunku vodnega toka pri vodnih osličkih. Vodni tok je
pomemben faktor, ki vpliva na to, koliko in kateri posamezniki se bodo vzdržali na podlagi.
Izpiranje namreč največkrat pomeni poškodbo ali ovirano vrnitev na prvotno mesto (Culver,
1971). Tisti, ki so dovolj sposobni oprijema oziroma premagovanja hitrosti začetnega sunka,
bodo lahko imeli večje možnosti za preživetje. Sunek vodnega toka je bil pri tem poskusu
ključen, saj osebki pokažejo drugačno vedenje kot če bi jim postopno višali hitrost. Med
populacijami površinskih in jamskih vodnih osličkov bi se pri postopnem višanju hitrosti
vodnega toka pokazale le majhne razlike v zmožnosti oprijema, najverjetneje zaradi
prilagajanja osebkov na tok vode (Fišer, 2017). Nova odkritja o vedenjskih razlikah med
obema populacijama vodnih osličkov lahko vodijo k večjemu razumevanju mehanizma
evolucije oziroma razhoda dveh skupin ene vrste v novi vrsti.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
5
Domnevamo, da je selekcijski pritisk vodnega toka v jamah močnejši kot na površju, saj bi
izpiranje največkrat pomenilo smrt, kar je v nasprotju s površinskih živali, kjer je lahko
izpiranje pomenilo »novo priložnost«. Jamske populacije vodnih osličkov so se morale
postopoma, tekom več generacij, prilagajati na novo okolje oziroma nove pogoje. Bolj ko so
se te populacije prilagajale jamskemu okolju, bolj so se začele morfološko oddaljevati od
svojih površinskih sorodnikov. Te morfološke razlike so torej posledica naravne selekcije.
V diplomskem delu želim na podlagi tega znanja preveriti naslednjo hipotezo:
Jamske populacije vodnih osličkov so izpostavljene močnejšemu selekcijskemu pritisku
vodnega toka kot površinske, zato je verjetno, da so se nanj prilagodile z učinkovitejšim
kljubovanjem. Razlike v odzivu na vodni tok pričakujem predvsem pri nenadnih
spremembah hitrosti (poplavni val), ko se odloči, ali bo žival obstala v svojem okolju.
Postavljeno hipotezo sem preverila s poskusom, kjer sem jamske in površinske vodne osličke
na dveh različnih lokacijah testirala glede sposobnosti oprijema na podlago. Iz dobljenih
rezultatov sem izračunala povprečje hitrosti ter jih vnesla v preglednico in škatlast diagram.
Iz tega bo razvidno, če je med populacijami prisotna razlika.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
6
2 MATERIALI IN METODE
Pri svojem delu sem uporabila kvantitativno metodo za ocenjevanje sposobnosti kljubovanja
vodnemu toku. Vključila sem statistično reprezentativne vzorce živali iz več populacij.
2.1 ANALIZIRANI VZORCI
2.1.1 Opis vzorca
Vzorec je vključeval 30 osebkov vsakega spola na populacijo, izjemoma manj, če to ni bilo
izvedljivo. V vzorcu so vključeni odrasli osebki, torej taki, ki so večji od 0,5 cm. Te sem
ločila po kraju in spolu v posebne plastične posodice – skupaj 8 skupin.
Preglednica 1: Število osebkov glede na spol ter kraj ulova.
Kraj ulova Spol Število osebkov Skupaj
Ljubljana (okolica
Biotehniške fakultete)
Moški 30 47
Ženski 17
Planinsko polje Moški 5
7 Ženski 2
Zelške jame Moški 11
36 Ženski 25
Planinska jama Moški 30
60 Ženski 30
SKUPAJ 150
Kot je razvidno iz tabele, je vseh osebkov 150. Od tega je bilo 47 osebkov iz Ljubljane
(okolice Biotehniške fakultete), 7 iz Planinskega polja, 36 iz Zelških jam in 60 iz Planinske
jame. Pri večini je moški spol številčno prevladoval, z izjemo pri Zelških jamah, kjer je bilo
samic več. Spol posameznega osebka sem določila s pomočjo stereomikroskopa, pri čemer
sem iskala določene značilnosti, kot so oblika glavoprsja, velikost tretjih parov nožnih
okončin ter prisotnost/odsotnost drugih spolnih značilnosti.
2.1.2 Postopek zbiranja podatkov
Podatke za poskus sem zbirala od februarja do maja 2017. Postopek se je začel tako, da sem z
mentorjevo pomočjo ulovila nekaj primerkov vodnih osličkov v različnih območjih.
Ulovljene vodne osličke sem s pomočjo večje plastične pipete previdno prenesla v lonček, ki
je bil napolnjen z okoliško vodo. Na območjih, kjer je bilo veliko blata ter rastlinja
(površinski ekosistemi), sem vodne osličke nabralo tako, da sem po dnu drsala s tanko mrežo.
Kar se je nabralo na mreži, sem dala v kadičko, iz katere sem nato izolirala vodne osličke. Ob
ustreznem številu osebkov (ali zgolj število osebkov, ki jih je bilo mogoče uloviti), se je
nabiranje končalo. Za meritve sem vodne osličke prenesla v posebne plastične posodice
napolnjene z deklorirano vodo in napol razkrojenimi listi za hrano in oprijem. Preden sem
začela izvajati poskus, sem naredila pred-test, kjer sem s tremi različnimi papirji poskušala

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
7
ugotoviti, ali se pojavljajo razlike med površinsko in jamsko populacijo. Ta test mi je
povedal, ali se pojavljajo razlike ter kateri papir je najbolje uporabiti za nadaljnji poskus.
Merjenja sem opravila v speleobiološkem laboratoriju na Oddelku za biologijo Biotehniške
fakultete. V za to namenjenih prostorih so imeli vodni oslički po izvajanju poskusov
zagotovljene vse ustrezne pogoje za življenje (nizke temperature, tema in izolacija). Z
aklimacijo organizmov sem izključila bolj ali manj vse vplive različnih okoljskih razmer na
izide poskusov.
Za ugotavljanje mejne hitrosti toka pri kateri vodni oslički ne morejo več obdržati stika s
podlago, sem uporabila v ta namen konstruirano aparaturo. Narejena je iz PVC kanala
napolnjenega z vodo, po katerem se z različno hitrostjo gibljejo »sani«. Na saneh je podlaga
iz grobega papirja, na katero postavimo vodnega oslička. Potovanje sani simulira vodni tok,
hitrost pa določa začetna napetost elastike, ki vleče sani. Hitrost sem izmerila s pomočjo
štetja posnetkov visoko-hitrostne kamere na definirani razdalji. Meritve sem izvedla v več
ponovitvah in v končni obdelavi rezultatov upoštevala povprečne vrednosti.
Podobno sem se lotila tudi meritve obstojnosti vodnih osličkov na podlagi. Previdno sem
prenesla osebka iz posodice na sani ter ju skupaj napela do določene razdalje. Po izpustu sem
videla, ali se je osebek od podlage ločil ali ne. Pri tisti razdalji ko se je ločil, sem razdaljo tudi
zapisala. Pri tem sem izvedla vsaj 10 ponovitev. Dobljene podatke sem vnesla v tabelo in iz
tega izračunala povprečje v programu Excel (Microsoft, 2010).
2.2 OPIS NAPRAVE
Za diplomsko delo je bila za poskus narejena posebna aparatura, ki simulira sunkovit porast
hitrosti vodnega toka. Narejena je iz PVC kanala, velikosti 65 x 3cm, napolnjenega z vodo,
po katerem se z različno hitrostjo gibljejo »sani«. Oba konca kanala imata prilepljeno
cilindrično plastiko, ki je pravokotna na sam kanal. Njuna funkcija je pritrjevanja elastike na
enem koncu ter ovoj vrvice za lažje nategovanje na drugem.
Slika 4: Fotografija pripomočka.
Sani so narejene iz pokrova PVC kanala in meri 12 x 3cm. Na obeh delih ima zvrtano luknjo
za pritrditev elastike in vrvice. Na ravno površino sani je prilepljen ulito narezan trak

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
8
filtrirnega papirja, na katerega sem nameščala vodne osličke. Izkazalo se je, da je filtrirni
papir v vodi dovolj trpežen in zagotavlja zadostno hrapavost za oprijem.
Pri sliki 5 je narejena skica same naprave. Razvidno je, da ob potegu vrvice na eni strani
napnemo elastiko na drugi. Po izpustu vrvice se elastika povrne v prvotno stanje ter povzroči
hiter premik sani oziroma val vode, ki simulira vodni tok v naravi. Na podlagi tega, kako
močno se elastika napne, je odvisna začetna hitrost sani oziroma vodnega toka.
Slika 5: Skica naprave iz ptičje perspektive.
Slika 6: Približana fotografija "sani".
Za lažje beleženje odmika sani sem z vodoodpornim označevalcem izrisala 5 cm razmike
(modra barva) ter 1 cm razmike (rdeča barva).
2.3 POSTOPEK OBDELAVE PODATKOV
Pridobljene podatke sem vnesla v program Excel (Microsoft, 2010). Videoposnetke sem
analizirala v programu VirtualDub 1.10.4 (Lee, 2013), kjer sem odčitavala čas in razdaljo
premika »sani«. Iz meritev časa ter razdalje sem izračunala hitrost premika prvih 3─5 cm. Iz
dobljenih vrednosti osebkov sem izračunala še povprečno mejno vrednost vsakega
posameznika. Vsa povprečja sem nato vnesla v program Past (Hammer, Harper in Ryan,
2001), ki je ustvaril škatlast diagram za boljšo predstavo razlik med populacijami in spoloma.
V tem programu sem prav tako analizirala dobljene podatke z neparametričnim statističnim
testom, imenovanim Mannov-Whitneyev test. Rezultati tega so povedali signifikantnost
dobljenih hitrosti pri poskusu.
Cilindrična plastika Cilindrična plastika
Vrvica
Sani
Grob papir
Elastika
PVC kanal

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
9
3 REZULTATI
3.1 UMERITEV ELASTIK
Slika 7: Graf hitrosti vodnega toka v odvisnosti od razdalje raztega sani pri šibkejši elastiki.
Slika 8: Graf hitrosti vodnega toka v odvisnosti od razdalje raztega sani pri močnejši elastiki.
y = 1,3217x + 0,1448
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 0,1 0,2 0,3 0,4 0,5 0,6
Hitrost sani (m/s)
Razdalja (m)
y = 5,3408x + 0,0961
0
0,2
0,4
0,6
0,8
1
1,2
0 0,02 0,04 0,06 0,08 0,1 0,12 0,14 0,16 0,18
Hitrost sani (m/s)
Razdalja (m)

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
10
Pri obeh grafih hitrosti vodnega toka v odvisnosti od razdalje nategnjene sani (slika 7 in 8) se
je določil trend naraščanja premice odvisnosti. Ta nam je podala formulo, s katero sem lahko
umerila elastiko. Z določitvijo formule iz razmerja med razdaljo in začetnim sunkom (hitrost
pri prvih 3-5 cm) sem lahko pri tej razdalji izračunala vmesne oziroma povprečne hitrosti, pri
katerih so se vodni oslički izpustili. Graf, ki je kazal podatke šibkejše elastike, je imel visoka
odstopanja od premice. Največje odstopanje je bilo za okoli 0,109 m/s. Pri grafu s podatki
močnejše elastike pa je bilo odstopanje manjše, za največ 0,058 m/s.
Kljub temu da je bila ustvarjena linearna premica pri prvem grafu (slika 7), je približno
linearen odnos veljaven le do okrog raztega 25 cm. Pri večjih raztegih se linearna zveza ne
nakazujejo več, vendar se ti pri nadaljnjem računanju še vseeno upoštevajo pri formuli:
y = 1,3217x + 0,1448, ki je linearna. Drugi graf (slika 8) teh težav ni povzročal, saj je
odnos razločno linearen, s podano formulo: y = 5,3408x + 0,0961.
3.2 MERITVE MEJNIH HITROSTI
Preglednica 2: Statistične vrednosti posameznih skupin vodnih osličkov.
PJ,
samci
PJ,
samice
ZJ,
samci
ZJ,
samice
L,
samci
L,
samice
PP,
samci
PP,
samice
Povprečna
vrednost
[m/s]
0,5196 0,5899 0,5054 0,5469 0,1942 0,1739 0,1941 0,1811
Minimum
[m/s] 0,2909 0,4112 0,3431 0,3880 0,1792 0,1699 0,1792 0,1587
Maksimum
[m/s] 0,7744 1,1803 0,8011 0,6722 0,2109 0,1778 0,2082 0,2149
Standardni
odklon [m/s] 0,1151 0,1567 0,1244 0,0850 0,0133 0,0056 0,0090 0,0157
Standardna
napaka [m/s] 0,0210 0,0291 0,0375 0,0227 0,0059 0,0040 0,0017 0,0038
25 percentil
[m/s] 0,4346 0,4700 0,4012 0,5123 0,1811 0,1274 0,1858 0,1729
75 percentil
[m/s] 0,6195 0,6596 0,5466 0,6135 0,2069 0,1334 0,2026 0,1891
*PJ = Planinska jama, ZJ =Zelške jame, L = Ljubljana, PP = Planinsko polje
Iz preglednice 2 lahko razberemo številke, ki nam predstavljajo numerične vrednosti, kot so
povprečna vrednost, odklon od povprečja, standardne napake ter druge podobne vrednosti. Iz
teh vrednosti dobimo sliko škatlastega diagrama, ki nam služi za lažjo slikovno prepoznavo
razlik med skupinami.
Škatlasti diagram (slika 9) prikazuje skupek povprečnih hitrosti, ki so se pojavile pri
določenem območju in določenem spolu v populaciji. Modro obarvani kvadrati zajemajo
percentile (med 25 in 75 percentilov) oziroma območje, kjer se povprečne hitrosti
najpogosteje pojavljajo. V teh kvadratih je s črto označena povprečna vrednost vseh

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
11
povprečnih hitrosti ene skupine (eno območje in en spol). Iz kvadratov pa izhajajo tudi
navpične črte, ki zajemajo vse vrednosti, ki odstopajo od večinskega deleža.
Slika 9: Škatlasti diagram hitrosti pri izpustu pri določenem kraju in spolu populacije.
Znotraj ene in druge ekološke skupine so povprečne vrednosti hitrosti dokaj podobne. Pri
jamski populaciji se je povprečje vrtelo med 0,5054 m/s in 0,5899 m/s, pri površinskih pa
med 0,1739 m/s in 0,1942 m/s. Standardni odklon pri jamskih vodnih osličkih je 0,1151 m/s
(samci) in 0,1567 m/s (samice) iz Planinske jame ter 0,1244 m/s (samci) in 0,0850 m/s
(samice) iz Zelške jame. Standardni odklon pri površinskih vodnih osličkih pa je 0,0133 m/s
(samci) in 0,0056 m/s (samice) iz Ljubljane ter 0,0090 m/s (samci) in 0,0157 m/s (samice) iz
Planinskega polja. Ti podatki nam povedo, da je odklon od povprečja pri jamskih populacijah
veliko večji kot pri površinskih populacijah. Jamski vodni oslički imajo večji razpon
zmožnosti oprijemanja na podlago ob vodnem sunku.
Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
12
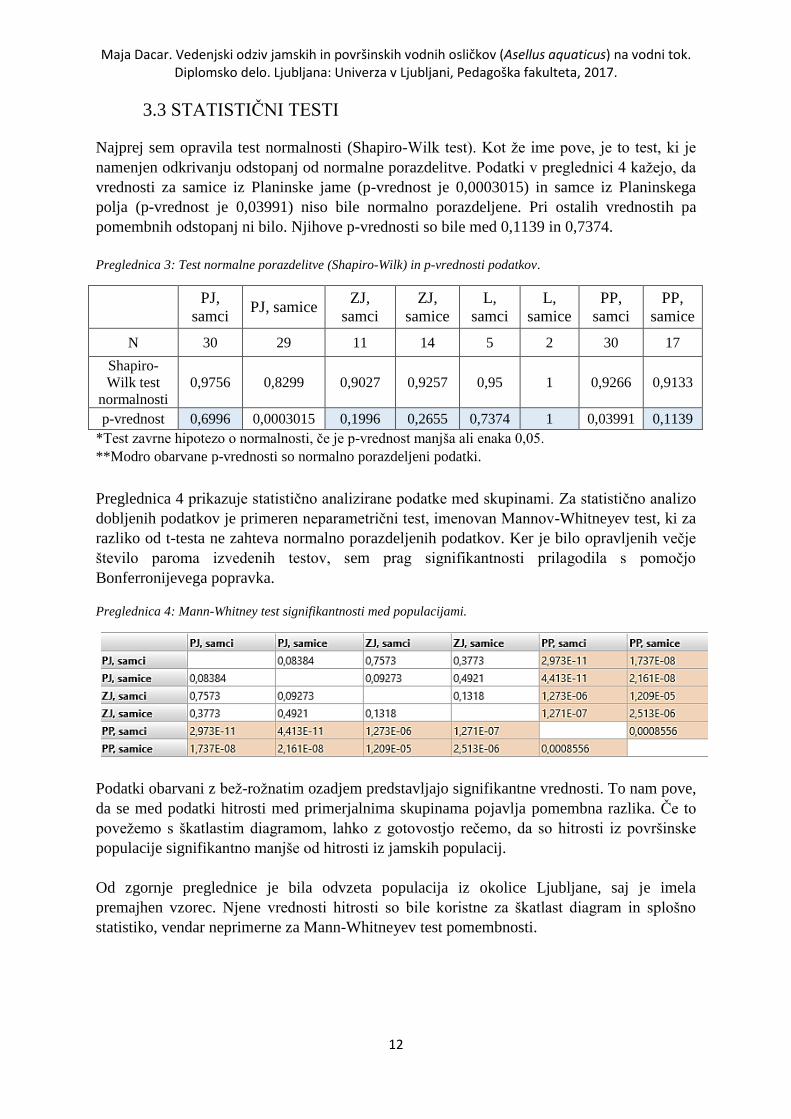
3.3 STATISTIČNI TESTI
Najprej sem opravila test normalnosti (Shapiro-Wilk test). Kot že ime pove, je to test, ki je
namenjen odkrivanju odstopanj od normalne porazdelitve. Podatki v preglednici 4 kažejo, da
vrednosti za samice iz Planinske jame (p-vrednost je 0,0003015) in samce iz Planinskega
polja (p-vrednost je 0,03991) niso bile normalno porazdeljene. Pri ostalih vrednostih pa
pomembnih odstopanj ni bilo. Njihove p-vrednosti so bile med 0,1139 in 0,7374.
Preglednica 3: Test normalne porazdelitve (Shapiro-Wilk) in p-vrednosti podatkov.
PJ,
samci PJ, samice
ZJ,
samci
ZJ,
samice
L,
samci
L,
samice
PP,
samci
PP,
samice
N 30 29 11 14 5 2 30 17
Shapiro-
Wilk test
normalnosti
0,9756 0,8299 0,9027 0,9257 0,95 1 0,9266 0,9133
p-vrednost 0,6996 0,0003015 0,1996 0,2655 0,7374 1 0,03991 0,1139
*Test zavrne hipotezo o normalnosti, če je p-vrednost manjša ali enaka 0,05.
**Modro obarvane p-vrednosti so normalno porazdeljeni podatki.
Preglednica 4 prikazuje statistično analizirane podatke med skupinami. Za statistično analizo
dobljenih podatkov je primeren neparametrični test, imenovan Mannov-Whitneyev test, ki za
razliko od t-testa ne zahteva normalno porazdeljenih podatkov. Ker je bilo opravljenih večje
število paroma izvedenih testov, sem prag signifikantnosti prilagodila s pomočjo
Bonferronijevega popravka.
Preglednica 4: Mann-Whitney test signifikantnosti med populacijami.
Podatki obarvani z bež-rožnatim ozadjem predstavljajo signifikantne vrednosti. To nam pove,
da se med podatki hitrosti med primerjalnima skupinama pojavlja pomembna razlika. Če to
povežemo s škatlastim diagramom, lahko z gotovostjo rečemo, da so hitrosti iz površinske
populacije signifikantno manjše od hitrosti iz jamskih populacij.
Od zgornje preglednice je bila odvzeta populacija iz okolice Ljubljane, saj je imela
premajhen vzorec. Njene vrednosti hitrosti so bile koristne za škatlast diagram in splošno
statistiko, vendar neprimerne za Mann-Whitneyev test pomembnosti.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
13
4 DISKUSIJA
Osrednji cilj, ki sem si ga zastavila v diplomskem delu, je bilo iskanje morebitnih razlik med
površinskimi in jamskimi vodnimi oslički v odzivu na vodni sunek. Obenem sem preverjala
tudi morebitne razlike, ki bi se lahko pojavile med samci in samicami. Jamski organizmi so
se veliko bolje odrezali pri oprijemanju na podlago, zato sem predvidevala, da so bolje
prilagojeni na ta selekcijski pritisk. Moja predvidevanja so potrdili rezultati, ki sem jih
pridobila s pomočjo poskusov. Iz vseh podatkov in analiz sem tako potrdila svojo hipotezo.
Razlike v odzivu na vodni tok so signifikantno prisotne in so pomembni dejavniki pri
določanju, ali bo žival obstala v svojem okolju ali ne.
To nam pove, da so jamski vodni oslički izpostavljeni močnejšemu selekcijskemu pritisku in
so nanj veliko bolje prilagojeni. Če povežem znanje z mojim poskusom, je razvidno, da je
omenjeni selekcijski pritisk eden od dejavnikov, ki prilagaja organizme na njihovo okolje.
Sedaj je razumljivo, zakaj so večje okončine in telo, izboljšani senzorični organi itd., uspešne
prilagoditve za jamske organizme.
Svojo raziskavo bi lahko izboljšala tako, da bi uporabila močnejšo elastiko za jamske
predstavnike in šibkejšo elastiko za površinske. Izkazalo se je namreč, da je kar nekaj
jamskih vodnih osličkov »premagalo« največji možni razteg šibkejše elastike. Ta je bolje
služila pri površinskih vodnih osličkih oziroma pri nizkih hitrostih, saj nam je ta nudila
natančnejše meritve kot močnejša elastika. Uporaba različnih elastik pri posameznih
skupinah bi olajšalo zapise ter preglednost pri analizi rezultatov. Sam princip uporabe PVC
kanala in sani se je izkazal za dokaj korektnega, vendar se je zaradi njegove višine in
velikosti večkrat pripetilo, da je veliko vode pljusknilo ven. Kljub temu se mi zdi, da je tak
način eksperimentiranja koristen in uporaben. Pri nekaterih populacijah je bilo število
osebkov malo in so predstavljali manjši reprezentativen vzorec. V tem primeru bi bilo
pametno uloviti osebke v času, ko ti najbolje uspevajo. Izkazalo se je, da je začetek februarja
za lovljenje vodnih osličkov še prezgodaj.
Pridobljene ugotovitve nam bodo pomagale razumeti ostale evolucijske procese, ki se
pojavljajo skupaj z adaptacijo v določeni skupini organizmov.

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
14
5 LITERATURA IN VIRI
Baldwin Ernest in Beatty R. A. (1941). The Pigmentation of Cavernicolous Animals. Journal
of Experimental Biology 18. Založba: The Company of Biologists Ltd. Str. 136-143
Culver David C. in Pipan Tanja (2009). The Biology of Caves and Other Subterranean
Habitats, založnik Oxford University Press: New York . str. 143-273
Culver David C. (1971). Analysis of Simple Cave Communities: III. Control of Abundance.
The American Midland Naturalist, Vol. 85, št. 1. Založnik: The University of Notre Dame,
str. 177
Fišer Ž. (2017). Doktorska disertacija: Evolucija reproduktivne izolacije ob prilagajanju na
podzemeljsko okolje. Ljubljana: Univerza v Ljubljani: Biotehniška fakulteta. Str. 118
Fox Richard (2006). Invertebrate Anatomy OnLine. Pridobljeno preko:
<http://lanwebs.lander.edu/faculty/rsfox/invertebrates/caecidotea.html> (12.7.2017)
Menshutkin V. V., Rukhovets L. A. in Filatov N. N. (2014). Ecosystem modeling of
freshwater lakes (review): 2. Models of freshwater lake’s ecosystem. Water resources, vol.
41, številka 1, str. 32
Peeters E.T.H.M., Camu J.M., Beijer J.A.J., Scheffer M. in Gardeniers J.J.P. (2002).
Response of the waterlouse Asellus aquaticus to multiple stressors: effects of current velocity
and mineral substratum. Journal of Aquatic Ecosystem Stress and Recovery, št. 9, založba:
Kluwer Academic Publishers, Nizozemska, str. 193–194
Riehl Torben (2016). ResearchGate. Pridobljeno preko
<https://www.researchgate.net/post/What_are_reliable_sexual_characteristics_of_the_freshw
ater_isopod_Asellus_aquaticus> (9.7.2017)
Rundle Howard D. in Nosil Patrik (2005). Ecological speciation. Ecology Letters, št. 8.
Založba: Blackwell Publishing Ltd. Str. 336–352
Stahl Bethany A., Gross Joshua B., Speiser Daniel I., Oakley Todd H., Patel Nipam H.,
Gould Douglas B. in Protas Meredith E. (2015). A Transcriptomic Analysis of Cave, Surface,
and Hybrid Isopod Crustaceans of the Species Asellus aquaticus. PLoS ONE 10(10).
Urednik: Sylvie Rétaux, Francija. Str. 1-2
Trontelj P. (2012). Natural Selection. V: Encyclopedia of Caves. 2nd ed. Uredila: White W.
B. in Culver D. C. (eds.). Založba: Academic Press, London. Str. 543-549
Yourgenome (2017). What are model organisms? Pridobljeno preko:
<http://www.yourgenome.org/facts/what-are-model-organisms> (12.7.2017)
Verovnik Rudi, Sket B. in Trontelj Peter (2004). Phylogeography of subterranean and surface
populations of water lice Asellus aquaticus (Crustacea: Isopoda). Molecular Ecology 13.
Založnik: Blackwell Publishing Ltd. Str. 1519

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
15
6 VIRI SLIKOVNEGA GRADIVA
Slika 1: Površinski (A) in jamski (B) Vodni osliček. Avtor: Žiga Fišer, 2017
Slika 2: Skica delov telesa vodnih osličkov. Avtorica: Maja Dacar, 2017
Slika 3: Fotografija samca in samice. Avtor: Žiga Fišer, 2017
Slika 4: Fotografija pripomočka. Avtorica: Maja Dacar, 2017
Slika 5: Skica naprave iz ptičje perspektive. Avtorica: Maja Dacar, 2017
Slika 6: Približana fotografija »sani«. Avtorica: Maja Dacar, 2017

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
16
7 PRILOGA
PRILOGA A
Preglednica 5: Meritve in izračun hitrosti "sani" pri različnih odmikih pri šibkejši elastiki.
1 cm 1 cm 1cm 1cm 1cm 1cm 1cm 1cm 1cm 1cm
s (cm) 1 1 1 1 1 1 1 1 1 1
t (št. Sličic -1) 12 10 19 20 23 18 21 20 24 19
t (s) 0,1 0,083333 0,158333 0,166667 0,191667 0,15 0,175 0,166667 0,2 0,158333
v (cm/s) 10 12 6,315789 6 5,217391 6,666667 5,714286 6 5 6,315789
v(m/s) 0,1 0,12 0,063158 0,06 0,052174 0,066667 0,057143 0,06 0,05 0,063158 0,069229923 Povprečna hitrost (m/s)
v (km/h) 0,36 0,432 0,227368 0,216 0,187826 0,24 0,205714 0,216 0,18 0,227368 0,249227721 Povprečna hitrost (km/h)
2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm
s (cm) 2 2 2 2 2 2 2 2 2 2
t (št. Sličic -1) 23 23 21 21 20 22 23 19 23 20
t (s) 0,191667 0,191667 0,175 0,175 0,166667 0,183333 0,191667 0,158333 0,191667 0,166667
v (cm/s) 10,43478 10,43478 11,42857 11,42857 12 10,90909 10,43478 12,63158 10,43478 12
v(m/s) 0,104348 0,104348 0,114286 0,114286 0,12 0,109091 0,104348 0,126316 0,104348 0,12 0,112136943 Povprečna hitrost (m/s)
v (km/h) 0,375652 0,375652 0,411429 0,411429 0,432 0,392727 0,375652 0,454737 0,375652 0,432 0,403692995 Povprečna hitrost (km/h)
3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm
s (cm) 3 3 3 3 3 3 3 3 3 3
t (št. Sličic -1) 25 23 25 24 24 25 24 23 23 25
t (s) 0,208333 0,191667 0,208333 0,2 0,2 0,208333 0,2 0,191667 0,191667 0,208333
v (cm/s) 14,4 15,65217 14,4 15 15 14,4 15 15,65217 15,65217 14,4
v(m/s) 0,144 0,156522 0,144 0,15 0,15 0,144 0,15 0,156522 0,156522 0,144 0,149556522 Povprečna hitrost (m/s)
v (km/h) 0,5184 0,563478 0,5184 0,54 0,54 0,5184 0,54 0,563478 0,563478 0,5184 0,538403478 Povprečna hitrost (km/h)
4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm
s (cm) 4 4 4 4 4 4 4 4 4 4
t (št. Sličic -1) 27 28 32 26 30 26 29 28 30 30
t (s) 0,225 0,233333 0,266667 0,216667 0,25 0,216667 0,241667 0,233333 0,25 0,25
v (cm/s) 17,77778 17,14286 15 18,46154 16 18,46154 16,55172 17,14286 16 16
v(m/s) 0,177778 0,171429 0,15 0,184615 0,16 0,184615 0,165517 0,171429 0,16 0,16 0,168538293 Povprečna hitrost (m/s)
v (km/h) 0,64 0,617143 0,54 0,664615 0,576 0,664615 0,595862 0,617143 0,576 0,576 0,606737855 Povprečna hitrost (km/h)
5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm
s (cm) 5 5 5 5 5 5 5 5 5 5
t (št. Sličic -1) 31 29 31 27 30 27 28 27 30 29
t (s) 0,258333 0,241667 0,258333 0,225 0,25 0,225 0,233333 0,225 0,25 0,241667
v (cm/s) 19,35484 20,68966 19,35484 22,22222 20 22,22222 21,42857 22,22222 20 20,68966
v(m/s) 0,193548 0,206897 0,193548 0,222222 0,2 0,222222 0,214286 0,222222 0,2 0,206897 0,208184226 Povprečna hitrost (m/s)
v (km/h) 0,696774 0,744828 0,696774 0,8 0,72 0,8 0,771429 0,8 0,72 0,744828 0,749463213 Povprečna hitrost (km/h)
6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm
s (cm) 4 3 3,5 4 3,5 3 4 4 3,5 4
t (št. Sličic -1) 16 12 15 18 15 13 17 17 14 18
t (s) 0,133333 0,1 0,125 0,15 0,125 0,108333 0,141667 0,141667 0,116667 0,15
v (cm/s) 30 30 28 26,66667 28 27,69231 28,23529 28,23529 30 26,66667
v(m/s) 0,3 0,3 0,28 0,266667 0,28 0,276923 0,282353 0,282353 0,3 0,266667 0,255803922 Povprečna hitrost (m/s)
v (km/h) 1,08 1,08 1,008 0,96 1,008 0,996923 1,016471 1,016471 1,08 0,96 0,920894118 Povprečna hitrost (km/h)
15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm
s (cm) 5 3,5 4 5 3 4 4 5 4 5
t (št. Sličic -1) 24 10 12 15 9 12 9 12 10 11
t (s) 0,2 0,083333 0,1 0,125 0,075 0,1 0,075 0,1 0,083333 0,091667
v (cm/s) 25 42 40 40 40 40 53,33333 50 48 54,54545
v(m/s) 0,25 0,42 0,4 0,4 0,4 0,4 0,533333 0,5 0,48 0,545455 0,432878788 Povprečna hitrost (m/s)
v (km/h) 0,9 1,512 1,44 1,44 1,44 1,44 1,92 1,8 1,728 1,963636 1,558363636 Povprečna hitrost (km/h)
20 cm 20 cm 20 cm 20 cm 20 cm 20 cm 20 cm 20 cm 20 cm 20 cm
s (cm) 3 4 4 4 5 4 3 5 4 5
t (št. Sličic -1) 8 9 10 7 9 11 8 12 10 12
t (s) 0,066667 0,075 0,083333 0,058333 0,075 0,091667 0,066667 0,1 0,083333 0,1
v (cm/s) 45 53,33333 48 68,57143 66,66667 43,63636 45 50 48 50
v(m/s) 0,45 0,533333 0,48 0,685714 0,666667 0,436364 0,45 0,5 0,48 0,5 0,518207792 Povprečna hitrost (m/s)
v (km/h) 1,62 1,92 1,728 2,468571 2,4 1,570909 1,62 1,8 1,728 1,8 1,865548052 Povprečna hitrost (km/h)

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
17
Preglednica 6: Meritev in izračun hitrosti "sani" pri različnih odmikih pri močnejši elastiki.
25 cm 25 cm 25 cm 25 cm 25 cm 25 cm 25 cm 25 cm 25 cm 25 cm
s (cm) 4 4 3,5 4,5 4 3 3 4,5 4 5
t (št. Sličic -1) 7 8 7 8 9 9 6 9 9 10
t (s) 0,058333 0,066667 0,058333 0,066667 0,075 0,075 0,05 0,075 0,075 0,083333
v (cm/s) 68,57143 60 60 67,5 53,33333 40 60 60 53,33333 60
v(m/s) 0,685714 0,6 0,6 0,675 0,533333 0,4 0,6 0,6 0,533333 0,6 0,582738095 Povprečna hitrost (m/s)
v (km/h) 2,468571 2,16 2,16 2,43 1,92 1,44 2,16 2,16 1,92 2,16 2,097857143 Povprečna hitrost (km/h)
30 cm 30 cm 30 cm 30 cm 30 cm 30 cm 30 cm 30 cm 30 cm 30 cm
s (cm) 4 4 5 3 4 4,5 4 5 4 3
t (št. Sličic -1) 9 8 11 7 8 9 8 10 7 6
t (s) 0,075 0,066667 0,091667 0,058333 0,066667 0,075 0,066667 0,083333 0,058333 0,05
v (cm/s) 53,33333 60 54,54545 51,42857 60 60 60 60 68,57143 60
v(m/s) 0,533333 0,6 0,545455 0,514286 0,6 0,6 0,6 0,6 0,685714 0,6 0,587878788 Povprečna hitrost (m/s)
v (km/h) 1,92 2,16 1,963636 1,851429 2,16 2,16 2,16 2,16 2,468571 2,16 2,116363636 Povprečna hitrost (km/h)
35 cm 35 cm 35 cm 35 cm 35 cm 35 cm 35 cm 35 cm 35 cm 35 cm
s (cm) 3,5 3,5 3 5 3,5 3,5 4 3,5 4 5
t (št. Sličic -1) 8 6 6 10 8 6 8 7 8 10
t (s) 0,066667 0,05 0,05 0,083333 0,066667 0,05 0,066667 0,058333 0,066667 0,083333
v (cm/s) 52,5 70 60 60 52,5 70 60 60 60 60
v(m/s) 0,525 0,7 0,6 0,6 0,525 0,7 0,6 0,6 0,6 0,6 0,605 Povprečna hitrost (m/s)
v (km/h) 1,89 2,52 2,16 2,16 1,89 2,52 2,16 2,16 2,16 2,16 2,178 Povprečna hitrost (km/h)
40 cm 40 cm 40 cm 40 cm 40 cm 40 cm 40 cm 40 cm 40 cm 40 cm
s (cm) 4 3 4,5 3 4 3 5 4 4 4
t (št. Sličic -1) 9 7 10 5 8 6 10 7 7 7
t (s) 0,075 0,058333 0,083333 0,041667 0,066667 0,05 0,083333 0,058333 0,058333 0,058333
v (cm/s) 53,33333 51,42857 54 72 60 60 60 68,57143 68,57143 68,57143
v(m/s) 0,533333 0,514286 0,54 0,72 0,6 0,6 0,6 0,685714 0,685714 0,685714 0,61647619 Povprečna hitrost (m/s)
v (km/h) 1,92 1,851429 1,944 2,592 2,16 2,16 2,16 2,468571 2,468571 2,468571 2,219314286 Povprečna hitrost (km/h)
45 cm 45 cm 45 cm 45 cm 45 cm 45 cm 45 cm 45 cm 45 cm 45 cm
s (cm) 3 3 4,5 4 4 3 4,5 3 5 3,5
t (št. Sličic -1) 6 6 7 8 7 6 8 8 10 6
t (s) 0,05 0,05 0,058333 0,066667 0,058333 0,05 0,066667 0,066667 0,083333 0,05
v (cm/s) 60 60 77,14286 60 68,57143 60 67,5 45 60 70
v(m/s) 0,6 0,6 0,771429 0,6 0,685714 0,6 0,675 0,45 0,6 0,7 0,628214286 Povprečna hitrost (m/s)
v (km/h) 2,16 2,16 2,777143 2,16 2,468571 2,16 2,43 1,62 2,16 2,52 2,261571429 Povprečna hitrost (km/h)
2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm 2 cm
s (cm) 2 2 2 2,5 2 2 2 2 1,5 2
t (št. Sličic -1) 16 16 18 20 14 18 15 19 11 22
t (s) 0,133333 0,133333 0,15 0,166667 0,116667 0,15 0,125 0,158333 0,091667 0,183333
v (cm/s) 15 15 13,33333 15 17,14286 13,33333 16 12,63158 16,36364 10,90909
v(m/s) 0,15 0,15 0,133333 0,15 0,171429 0,133333 0,16 0,126316 0,163636 0,109091 0,14471383 Povprečna hitrost (m/s)
v (km/h) 0,54 0,54 0,48 0,54 0,617143 0,48 0,576 0,454737 0,589091 0,392727 0,520969788 Povprečna hitrost (km/h)
3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm 3 cm
s (cm) 3 3 3 3 3 3 3 3,5 3 3
t (št. Sličic -1) 11 13 14 15 13 14 12 14 12 12
t (s) 0,091667 0,108333 0,116667 0,125 0,108333 0,116667 0,1 0,116667 0,1 0,1
v (cm/s) 32,72727 27,69231 25,71429 24 27,69231 25,71429 30 30 30 30
v(m/s) 0,327273 0,276923 0,257143 0,24 0,276923 0,257143 0,3 0,3 0,3 0,3 0,28354046 Povprečna hitrost (m/s)
v (km/h) 1,178182 0,996923 0,925714 0,864 0,996923 0,925714 1,08 1,08 1,08 1,08 1,020745654 Povprečna hitrost (km/h)
4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm 4 cm
s (cm) 4 4 4,5 4 4 4 3,75 4 4 4
t (št. Sličic -1) 15 13 18 13 12 14 12 13 14 13
t (s) 0,125 0,108333 0,15 0,108333 0,1 0,116667 0,1 0,108333 0,116667 0,108333
v (cm/s) 32 36,92308 30 36,92308 40 34,28571 37,5 36,92308 34,28571 36,92308
v(m/s) 0,32 0,369231 0,3 0,369231 0,4 0,342857 0,375 0,369231 0,342857 0,369231 0,355763736 Povprečna hitrost (m/s)
v (km/h) 1,152 1,329231 1,08 1,329231 1,44 1,234286 1,35 1,329231 1,234286 1,329231 1,280749451 Povprečna hitrost (km/h)
5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm 5 cm
s (cm) 5 5 5 5 5 5 5 5 4,5 5
t (št. Sličic -1) 17 14 15 20 18 15 15 18 16 15
t (s) 0,141667 0,116667 0,125 0,166667 0,15 0,125 0,125 0,15 0,133333 0,125
v (cm/s) 35,29412 42,85714 40 30 33,33333 40 40 33,33333 33,75 40
v(m/s) 0,352941 0,428571 0,4 0,3 0,333333 0,4 0,4 0,333333 0,3375 0,4 0,368567927 Povprečna hitrost (m/s)
v (km/h) 1,270588 1,542857 1,44 1,08 1,2 1,44 1,44 1,2 1,215 1,44 1,326844538 Povprečna hitrost (km/h)

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
18
6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm 6 cm
s (cm) 4 5 4 5 5 5 4 4,5 4 5
t (št. Sličic -1) 12 14 12 15 18 16 13 15 12 16
t (s) 0,1 0,116667 0,1 0,125 0,15 0,133333 0,108333 0,125 0,1 0,133333
v (cm/s) 40 42,85714 40 40 33,33333 37,5 36,92308 36 40 37,5
v(m/s) 0,4 0,428571 0,4 0,4 0,333333 0,375 0,369231 0,36 0,4 0,375 0,384113553 Povprečna hitrost (m/s)
v (km/h) 1,44 1,542857 1,44 1,44 1,2 1,35 1,329231 1,296 1,44 1,35 1,382808791 Povprečna hitrost (km/h)
7 cm 7 cm 7 cm 7 cm 7 cm 7 cm 7 cm 7 cm 7 cm 7 cm
s (cm) 4 5 4 5 5 4 6 5 6 4
t (št. Sličic -1) 12 16 11 13 14 10 14 12 19 9
t (s) 0,1 0,133333 0,091667 0,108333 0,116667 0,083333 0,116667 0,1 0,158333 0,075
v (cm/s) 40 37,5 43,63636 46,15385 42,85714 48 51,42857 50 37,89474 53,33333
v(m/s) 0,4 0,375 0,436364 0,461538 0,428571 0,48 0,514286 0,5 0,378947 0,533333 0,450803994 Povprečna hitrost (m/s)
v (km/h) 1,44 1,35 1,570909 1,661538 1,542857 1,728 1,851429 1,8 1,364211 1,92 1,622894379 Povprečna hitrost (km/h)
8 cm 8 cm 8 cm 8 cm 8 cm 8 cm 8 cm 8 cm 8 cm 8 cm
s (cm) 5 4 4 5 5 5 4 5 5 5
t (št. Sličic -1) 10 9 9 10 12 16 13 9 10 11
t (s) 0,083333 0,075 0,075 0,083333 0,1 0,133333 0,108333 0,075 0,083333 0,091667
v (cm/s) 60 53,33333 53,33333 60 50 37,5 36,92308 66,66667 60 54,54545
v(m/s) 0,6 0,533333 0,533333 0,6 0,5 0,375 0,369231 0,666667 0,6 0,545455 0,532301865 Povprečna hitrost (m/s)
v (km/h) 2,16 1,92 1,92 2,16 1,8 1,35 1,329231 2,4 2,16 1,963636 1,916286713 Povprečna hitrost (km/h)
9 cm 9 cm 9 cm 9 cm 9 cm 9 cm 9 cm 9 cm 9 cm 9 cm
s (cm) 3 3 2,5 2,5 3 2 3 3 4 3
t (št. Sličic -1) 6 5 6 5 6 5 6 5 7 7
t (s) 0,05 0,041667 0,05 0,041667 0,05 0,041667 0,05 0,041667 0,058333 0,058333
v (cm/s) 60 72 50 60 60 48 60 72 68,57143 51,42857
v(m/s) 0,6 0,72 0,5 0,6 0,6 0,48 0,6 0,72 0,685714 0,514286 0,602 Povprečna hitrost (m/s)
v (km/h) 2,16 2,592 1,8 2,16 2,16 1,728 2,16 2,592 2,468571 1,851429 2,1672 Povprečna hitrost (km/h)
10 cm 10 cm 10 cm 10 cm 10 cm 10 cm 10 cm 10 cm 10 cm 10 cm
s (cm) 2,5 3 2,5 2 3 3 2 3,5 3 3
t (št. Sličic -1) 6 5 5 4 5 5 3 8 7 6
t (s) 0,05 0,041667 0,041667 0,033333 0,041667 0,041667 0,025 0,066667 0,058333 0,05
v (cm/s) 50 72 60 60 72 72 80 52,5 51,42857 60
v(m/s) 0,5 0,72 0,6 0,6 0,72 0,72 0,8 0,525 0,514286 0,6 0,629928571 Povprečna hitrost (m/s)
v (km/h) 1,8 2,592 2,16 2,16 2,592 2,592 2,88 1,89 1,851429 2,16 2,267742857 Povprečna hitrost (km/h)
11 cm 11 cm 11 cm 11 cm 11 cm 11 cm 11 cm 11 cm 11 cm 11 cm
s (cm) 4 2 2,5 2,5 3,5 2,5 3,5 3 2,5 3
t (št. Sličic -1) 8 4 5 3 4 6 6 6 5 4
t (s) 0,066667 0,033333 0,041667 0,025 0,033333 0,05 0,05 0,05 0,041667 0,033333
v (cm/s) 60 60 60 100 105 50 70 60 60 90
v(m/s) 0,6 0,6 0,6 1 1,05 0,5 0,7 0,6 0,6 0,9 0,715 Povprečna hitrost (m/s)
v (km/h) 2,16 2,16 2,16 3,6 3,78 1,8 2,52 2,16 2,16 3,24 2,574 Povprečna hitrost (km/h)
12 cm 12 cm 12 cm 12 cm 12 cm 12 cm 12 cm 12 cm 12 cm 12 cm
s (cm) 2 3 2,5 3 2 2 2 3,75 3 3
t (št. Sličic -1) 7 4 3 4 4 3 3 5 5 8
t (s) 0,058333 0,033333 0,025 0,033333 0,033333 0,025 0,025 0,041667 0,041667 0,066667
v (cm/s) 34,28571 90 100 90 60 80 80 90 72 45
v(m/s) 0,342857 0,9 1 0,9 0,6 0,8 0,8 0,9 0,72 0,45 0,741285714 Povprečna hitrost (m/s)
v (km/h) 1,234286 3,24 3,6 3,24 2,16 2,88 2,88 3,24 2,592 1,62 2,668628571 Povprečna hitrost (km/h)
13 cm 13 cm 13 cm 13 cm 13 cm 13 cm 13 cm 13 cm 13 cm 13 cm
s (cm) 2,5 2,25 3 2 3 1,25 3,5 3 3 2
t (št. Sličic -1) 3 3 5 4 5 3 4 5 4 3
t (s) 0,025 0,025 0,041667 0,033333 0,041667 0,025 0,033333 0,041667 0,033333 0,025
v (cm/s) 100 90 72 60 72 50 105 72 90 80
v(m/s) 1 0,9 0,72 0,6 0,72 0,5 1,05 0,72 0,9 0,8 0,791 Povprečna hitrost (m/s)
v (km/h) 3,6 3,24 2,592 2,16 2,592 1,8 3,78 2,592 3,24 2,88 2,8476 Povprečna hitrost (km/h)
14 cm 14 cm 14 cm 14 cm 14 cm 14 cm 14 cm 14 cm 14 cm 14 cm
s (cm) 3 3,5 3 3 2,5 3,75 2 2,25 2,5 2
t (št. Sličic -1) 6 4 4 4 4 5 3 5 3 3
t (s) 0,05 0,033333 0,033333 0,033333 0,033333 0,041667 0,025 0,041667 0,025 0,025
v (cm/s) 60 105 90 90 75 90 80 54 100 80
v(m/s) 0,6 1,05 0,9 0,9 0,75 0,9 0,8 0,54 1 0,8 0,824 Povprečna hitrost (m/s)
v (km/h) 2,16 3,78 3,24 3,24 2,7 3,24 2,88 1,944 3,6 2,88 2,9664 Povprečna hitrost (km/h)

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
19
PRILOGA B
Preglednica 7: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinske jame pri močnejši
elastiki.
15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm 15 cm
s (cm) 2 2 4,5 2 3 2,5 2,5 2 3 2,5
t (št. Sličic -1) 4 3 5 3 4 3 3 3 4 4
t (s) 0,033333 0,025 0,041667 0,025 0,033333 0,025 0,025 0,025 0,033333 0,033333
v (cm/s) 60 80 108 80 90 100 100 80 90 75
v(m/s) 0,6 0,8 1,08 0,8 0,9 1 1 0,8 0,9 0,75 0,863 Povprečna hitrost (m/s)
v (km/h) 2,16 2,88 3,888 2,88 3,24 3,6 3,6 2,88 3,24 2,7 3,1068 Povprečna hitrost (km/h)
16 cm 16 cm 16 cm 16 cm 16 cm 16 cm 16 cm 16 cm 16 cm 16 cm
s (cm) 2 2 3 2,9 2 2,5 2,75 2,5 3,75 3
t (št. Sličic -1) 4 4 4 3 3 5 3 2 3 4
t (s) 0,033333 0,033333 0,033333 0,025 0,025 0,041667 0,025 0,016667 0,025 0,033333
v (cm/s) 60 60 90 116 80 60 110 150 150 90
v(m/s) 0,6 0,6 0,9 1,16 0,8 0,6 1,1 1,5 1,5 0,9 0,966 Povprečna hitrost (m/s)
v (km/h) 2,16 2,16 3,24 4,176 2,88 2,16 3,96 5,4 5,4 3,24 3,4776 Povprečna hitrost (km/h)
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,077 0,5073416 0,099 0,6248392
0,083 0,5393864 0,113 0,6996104
0,075 0,49666 0,104 0,6515432
0,053 0,3791624 0,203 1,1802824
0,107 0,6675656 0,107 0,6675656
0,052 0,3738216 0,108 0,6729064
0,076 0,5020008 0,122 0,7476776
0,107 0,6675656 0,139 0,8384712
0,097 0,6141576 0,0895 0,5741016
0,076 0,5020008 0,059 0,4112072
0,112 0,6942696 0,069 0,4646152
0,127 0,7743816 0,0615 0,4245592
0,094 0,5981352 0,064 0,4379112
0,055 0,389844 0,0715 0,4779672
0,103 0,6462024 0,0915 0,5847832
0,107 0,6675656 0,0625 0,4299
0,068 0,4592744 0,0705 0,4726264
0,067 0,4539336 0,1105 0,6862584
0,055 0,389844 0,073 0,4859784
0,058 0,4058664
0,087 0,5607496 Povprečje
0,101 0,6355208 0,0897 0,57516976
0,083 0,5393864
Povprečje
0,083913 0,54426255

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
20
Preglednica 8: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinske jame pri šibkejši
elastiki.
Preglednica 9: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Zelške jame pri močnejši elastiki.
Preglednica 10: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Zelške jame pri šibkejši elastiki.
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,275 0,45614675 0,271 0,5029807
0,376 0,57049592 0,327 0,5769959
0,256 0,43463552 0,291 0,5294147
0,129 0,29084993 0,224 0,4408608
0,299 0,48331883 0,244 0,4672948
0,256 0,43463552 0,361 0,6219337
0,273 0,45388241 0,37 0,633829
0,355 0,6140035
Povprečje 0,341 0,5954997
0,26629 0,446285549 0,339 0,5928563
Povprečje
0,3123 0,55756691
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,078 0,5126824 0,082 0,5340456
0,069 0,4646152 0,101 0,6355208
0,132 0,8010856 0,101 0,6355208
0,083 0,5393864
Povprečje 0,083 0,5393864
0,093 0,5927944 0,0955 0,6061464
0,094 0,5981352
0,083 0,5393864
Povprečje
0,090313 0,57844367
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,256 0,4831552 0,184 0,3879928
0,179 0,3813843 0,23 0,448791
0,15 0,343055 0,302 0,5439534
0,351 0,6087167 0,323 0,5717091
0,194 0,4012098 0,196 0,4038532
0,278 0,5122326 0,399 0,6721583
0,304 0,5465968
0,272 0,5043024 Povprečje
0,27225 0,504632825
Povprečje
0,248 0,4725816

Maja Dacar. Vedenjski odziv jamskih in površinskih vodnih osličkov (Asellus aquaticus) na vodni tok. Diplomsko delo. Ljubljana: Univerza v Ljubljani, Pedagoška fakulteta, 2017.
21
Preglednica 11: Izračun posamezne povprečne hitrosti osebkov v populaciji iz Planinskega polja pri šibkejši
elastiki.
Preglednica 12: Izračun posamezne povprečne hitrosti osebkov v populaciji iz okolice Ljubljana pri šibkejši
elastiki.
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,044 0,2029548 0,025 0,1778425
0,05 0,210885 0,019 0,1699123
0,026 0,1791642
0,029 0,1831293 Povprečje
0,038 0,1950246 0,022 0,1738774
Povprečje
0,0374 0,19423158
Moški Ženski
Razdalja osebka (m) Hitrost (m/s) Razdalja osebka (m) Hitrost (m/s)
0,042 0,2003114 0,029 0,1831293
0,04 0,197668 0,022 0,1738774
0,0315 0,18643355 0,0325 0,18775525
0,048 0,2082416 0,0215 0,17321655
0,041 0,1989897 0,012 0,1606604
0,045 0,2042765 0,021 0,1725557
0,028 0,1818076 0,0105 0,15867785
0,041 0,1989897 0,035 0,1910595
0,031 0,1857727 0,0275 0,18114675
0,0435 0,20229395 0,052 0,2135284
0,031 0,1857727 0,022 0,1738774
0,0385 0,19568545 0,014 0,1633038
0,026 0,1791642 0,0235 0,17585995
0,031 0,1857727 0,028 0,1818076
0,036 0,1923812 0,0345 0,19039865
0,047 0,2069199 0,029 0,1831293
0,0445 0,20361565 0,053 0,2148501
0,0325 0,18775525
0,048 0,2082416 Povprečje
0,038 0,1950246 0,027471 0,181108421
0,045 0,2042765
0,0295 0,18379015
0,045 0,2042765
0,0305 0,18511185
0,0375 0,19436375
0,0355 0,19172035
0,031 0,1857727
0,0425 0,20097225
0,0285 0,18246845
0,0295 0,18379015
Povprečje
0,037267 0,194055794
























