Embed Size (px)
Citation preview

Untersuchungen zum Ionentransport aus demXylem in den Symplasten des Maisblattes
Dissertation zur Erlangung des Doktorgradesder Mathematisch-Naturwissenschaftlichen Fakultät
der Christian-Albrechts-Universität zu Kiel
vorgelegt von Maike Keunecke
Kiel, Dezember 1999

Referent: Prof. Dr. Ulf-Peter Hansen
Koreferent:
Tag der mündlichen Prüfung:
Zum Druck genehmigt:
Der Dekan

Vorbemerkung:
Teilergebnisse dieser Dissertation sind bereits mit Genehmigung des Dekans der Ma-thematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität zuKiel wie folgt vorab veröffentlicht bzw. eingereicht worden:
Keunecke M, Sutter JU, Sattelmacher B, Hansen UP 1997Isolation and patch clamp measurements of xylem contact cells for the study of their
role in the exchange between apoplast and symplast of leaves.Plant and Soil 196: 239-244.
Keunecke M, Hansen UP 1999Different pH-dependences of K+ channel activity in bundle sheath and mesophyll cells
of maize leaves.Planta (im Druck).
Keunecke M, Lindner B, Seydel U, Schulz A, Hansen UPBundle sheath cells in maize are the location of uptake from the xylem.Journal of Experimental Botany (Eingereicht).

I
Inhaltsverzeichnis
1. EINLEITUNG 1
1.1 ABKÜRZUNGEN UND BIOLOGISCHE BEGRIFFE 3
2 EINFÜHRUNG 5
2.1 ENTWICKLUNG UND AUFBAU EINER MAISPFLANZE 52.2 TRANSPORTVORGÄNGE IN DER WURZEL 72.3 TRANSPORTVORGÄNGE IM MAISBLATT 92.3.1 XYLEM: AUFBAU UND FUNKTION 112.3.2 PHLOEM: AUFBAU UND FUNKTION 122.4 AUFBAU EINER PFLANZENZELLE 122.4.1 PLASMODESMEN 132.5 PHOTOSYNTHESE 152.5.1 DER CALVIN-ZYKLUS 152.5.2 PHOTOSYNTHESE DER C4-PFLANZEN 16
3 BESTIMMUNG DES ÜBERTRITTSORTES DER IONEN AUS DEM XYLEM INDEN BLATTSYMPLASTEN 18
3.1 MOTIVATION 183.2 VERWENDETE PFLANZEN 213.3 ANFÜTTERUNG MIT RUBIDIUM UND ERSTELLEN DER SCHNITTE 213.4 ANALYSE MIT LAMMA 223.5 ERGEBNISSE 25
4 BIOMEMBRANEN UND MEMBRANTRANSPORT 29
4.1 PUMPEN, COTRANSPORTER UND UNIPORTER 304.1.1 PUMPEN 304.1.2 COTRANSPORTER 314.1.3 CARRIER 314.1.4 KANÄLE: STRUKTUR UND MODELLE 314.2 MEMBRANPOTENTIALE 354.3 TRANSPORTWEGE INS PHLOEM UND AUS DEM XYLEM 364.4 LÖSUNGEN 674.5 DIE PATCH-CLAMP-TECHNIK 42
5 ISOLATIONSVERFAHREN 46
5.1 ISOLATION VON XYLEMKONTAKTZELLEN BEI BOHNE 475.2 ISOLATION VON BÜNDELSCHEIDENZELLEN BEI MAIS 475.3 WHOLE-CELL-PRÄPARATION 505.4 ISOLATION VON MESOPHYLLZELLEN 515.5 VITALITÄTSTESTS 525.6 GRÖßEN VON BÜNDELSCHEIDENZELLEN UND -PROTOPLASTEN 53

II
5.7 SEALBILDUNG 56
6 EXPERIMENTELLER AUFBAU DER PATCH-CLAMP-ANLAGE 57
6.1 MECHANISCHER AUFBAU MIT MIKROSKOP 576.2 ELEKTRISCHE SCHALTUNGEN UND DATENAUFNAHME 586.2.1 WHOLE-CELL-MESSUNGEN 606.3 HERSTELLUNG DER PIPETTEN 61
7 EINZELKANALMESSUNGEN 62
7.1 SYMMETRISCHER K+-KANAL UND EINWÄRTSGLEICHRICHTER 647.2 BLOCKIERUNG DER KANÄLE DURCH WEICHGLAS 657.3 EINWÄRTSGLEICHRICHTER 657.4 SELEKTIVITÄT UND UMKEHRPOTENTIAL DES SYMMETRISCHEN K+ KANALS 667.5 DURCHLÄSSIGKEIT DER K+-KANÄLE FÜR NH4
+ 687.6 NA+-MESSUNGEN 697.7 PHYSIOLOGISCHE BEDINGUNGEN 72
8 WHOLE-CELL-MESSUNGEN AN BÜNDELSCHEIDENZELLEN 72
8.1 SPANNUNGSPROTOKOLL 748.2 VERSCHIEDENE K+-KANAL-TYPEN BEI BÜNDELSCHEIDENZELLEN 768.3 ATP-ABHÄNGIGKEIT 788.4 GIBT ES ZWEI ARTEN VON BÜNDELSCHEIDENZELLEN? 808.5 NA+-WHOLE-CELL-MESSUNGEN 858.6 RB+-EFFEKT AUF BÜNDELSCHEIDENZELLEN 878.7 PH-EFFEKT BEI BÜNDELSCHEIDENZELLEN 91
9 WHOLE-CELL-MESSUNGEN AN MESOPHYLLZELLEN 94
9.1 RB+-EFFEKTE AUF MESOPHYLLZELLEN 979.2 PH-EFFEKT BEI MESOPHYLLZELLEN 100
10 DISKUSSION 102
10.1 VERGLEICH DER EINZELKANAL- UND WHOLE-CELL-MESSUNGEN 10210.2 VERGLEICH VON BÜNDELSCHEIDEN- UND MESOPHYLLZELLEN 10410.3 WIRKUNG VON NA+ 10510.4 EINFLUß VON RB+ 10510.5 EINFLUß DES PH-WERTES 11010.6 PHYSIOLOGISCHE BEDEUTUNG DER PH-ABHÄNGIGKEITEN 11210.7 BIOPHYSIKALISCHE ASPEKTE DER NA+- UND RB+-MESSUNGEN 11410.8 VERGLEICH DER ERGEBNISSE VON WURZEL UND BLATT 11510.9 AUSBLICK 117
11 ZUSAMMENFASSUNG 119
12 ANHANG 122

III
12.1 BESCHICHTUNG DER LAMMA-GRIDS12.2 FÄRBEN MIT NEUTRALROT12.3 APHIDENTECHNIK12.4 LEACHING12.5 WURZELDRUCKKAMMER
13 LITERATUR 127
14 DANKSAGUNG 138

1
1. Einleitung
Pflanzen nehmen Nährstoffe aus dem Boden auf (z.B. Marschner, 1995). Diese müssenzum Ort der Photosynthese in die Blätter transportiert werden. Die im Leitbündel dafürverantwortlichen Gefäße heißen Xylem. Das Xylem und der Raum zwischen den Blatt-zellen bilden den Apoplasten. Während die Funktion der Zellen sehr intensiv untersuchtwird, ist die Kenntnis über die Prozesse im Apoplasten gering.
Die Einrichtung eines DFG-Schwerpunktprogrammes „Der Apoplast der höherenPflanze: Speicher-, Transport- und Reaktionsraum“ hatte das Ziel, diese Wissenslückezu schließen. Hier geht es z.B. um die Besiedelung mit Pilzen (Heyser, 1997, Hecht-Buchholz und Höflich, 1997, Schulz et al., 1998), Ablagerung (Oxalatkristalle, Türkund Fink, 1997) und insbesondere um die Rolle bei der Versorgung der Blattzellen.
Von den etwa 30 in Deutschland am Schwerpunkt beteiligten Arbeitsgruppen hattesich eine Teilgruppe als Ziel gesetzt, eine Stoffwechselbilanz für eine höhere Pflanzeaufzustellen. Hierbei studiert die AG Schurr (Heidelberg) mit der Wurzeldruckkammerden Xylemsaft hinsichtlich der Nährstoffe, die von der Wurzel bis ins Blatt transportiertwerden (Gollan et al., 1992). Die AG Heldt (Göttingen) untersucht mit Hilfe der Aphi-dentechnik (Riens et al., 1991) die Zusammensetzung des Phloemsaftes (enthält diePhotosyntheseprodukte). Zusammen mit der AG Sattelmacher analysiert die AG Heldtauch die Apoplastenlösung (Lohaus et al., 1995), während in der AG Sattelmacher zu-sätzlich der Einfluß der Beregnung (Leaching) untersucht wird (Pennewiß und Sattel-macher, 1998). Die AG Hansen hat die Aufgabe, den Transport vom Apoplasten (Xy-lem) in den Symplasten zu untersuchen (Keunecke et al., 1997, Keunecke et al.,1999a,b).
Der Apoplast in Maisblättern ist durch die undurchlässige Suberinlamelle um dieBündelscheide in zwei Kompartimente geteilt (Hattersley and Browing, 1981, Botha etal., 1982, Canny, 1995, Evert et al., 1977, 1985, 1996). Die Forschung des Transportesmuß sich daher auf zwei Bereiche der Wechselwirkungen von Apoplast und Symplastkonzentrieren: innerhalb und außerhalb der Suberinlamelle. Außerhalb der Suberinla-melle liegen die Mesophyllzellen um die Bündelscheide. Hier ist der Austausch Sym-plast-Blattapoplast von Interesse für das Projekt.
Innerhalb der Suberinlamelle mußte zunächst die Frage geklärt werden, an wel-chem Ort dieser Übergang stattfindet. Dies machte es erforderlich, den Weg der Ionenmit einem Tracer (Rubidium) zu verfolgen (Keunecke et al., 1999).
Transporteigenschaften von Membranen werden meistens durch elektrische Meß-verfahren ermittelt. Dafür müssen die betreffenden Zellen zugängig gemacht werden.Vorarbeiten von Sutter (1996) zur Isolation von Xylemkontaktzellen an Bohnen (Vicia

2
faba) bildeten dazu die Grundlage. Die Überwindung der speziellen Schwierigkeiten,die der Aufbau der Maisblätter dabei mit sich brachte, erforderte großen Aufwand.Auch von der DFG wurde dies als sehr schwierig eingestuft, so daß das Projekt zu-nächst nur auf ein Probejahr genehmigt wurde. Einen weiteren Hinweis auf die Schwie-rigkeiten gab (indirekt) eine Literaturrecherche: In den 10 Jahren (von 1989 bis Oktober1998) sind 127 Arbeiten erschienen, die sich mit Patch-Clamp-Messungen an Pflan-zenmembranen beschäftigen1 (ermittelt mit dem Literaturrechercheprogramm „win-spirs“ der CAU Kiel). Davon entfallen nur 11 auf Mesophyllzellen, die anderen befas-sen sich nicht mit Zellen, die der Versorgung des Blattes dienen (beim Mais ist diePhotosynthese zwischen den Mesophyll- und Bündelscheidenzellen aufgeteilt). Zur Zeitgibt es nur die Arbeit von Keunecke et al. (1997), die sich mit Zellen beschäftigt, dieweiter im Blattinneren liegen als die Mesophyllzellen.
Zur Untersuchung des Membrantransportes dient heutzutage die Patch-Clamp-Technik (Numberger und Draguhn, 1996): Eine Mikroelektrode wird auf die Membrangesetzt und mit einer zweiten Elektrode im Bad die durch vorgegebene Spannungenhervorgerufenen Ströme aufgezeichnet. Die Patch-Clamp Technik benötigt saubereObjekte, d.h. die Zelloberflächen müssen sehr sorgfältig von Zellwandresten befreitsein.
Es gibt zwei Möglichkeiten der Untersuchung mit der Patch-Clamp-Technik: Ein-zelkanaluntersuchungen ermöglichen die Betrachtung einzelner Kanäle, Whole-Cell-Messungen geben einen Überblick über das Verhalten der ganzen Zelle. Mit beidenVerfahren können Stromspannungskurven, Umkehrpotential (Bestimmung der Selekti-vitäten) und Blöcke durch Ionen ermittelt werden. Während in Einzelkanalmessungendie Klassifizierung der Kanäle im Vordergrund steht, kann bei Whole-Cell-Messungendas Verhalten der Zelle unter physiologischen Bedingungen betrachtet werden (Schroe-der et al., 1994, Hedrich et al., 1988, 1989).
Bündelscheiden- und Mesophyllzellen wurden mit der Patch-Clamp-Technik unter-sucht, und es ergaben sich erste Hinweise auf Besonderheiten der physiologischen Rolledes Austausches zwischen Apoplast und Symplast.
1 Verteilung der Paper: 37 Schließzellen (27 davon Vicia faba), 23 Tonoplasten, 15Wurzelzellen, 11 Mesophyllzellen, 6 Aleuronen oder Samen, 5 Reviews, 3 Koleoptilen,26 sind nicht auf eine Zellart spezifiziert, 1 Bündelscheidenzellen (Keunecke et al.,1997).

3
1.1 Abkürzungen und biologische Begriffe
Apoplast gas- und wassergefüllter toter Raum außerhalb der ZellenAssimilate Produkte der PhotosyntheseATP Adenosintriphosphat; energetisches „Kleingeld“ der Zelle;
die chemische Energie (≈ 450 meV) wird durch Abspal-tung eines Phosphatrestes frei
BSA bovine serum albumin (Serumprotein des Rindes)BS = Bündelscheidenzellen Zellschicht um das LeitbündelBTP-ATP Bis-Tris-Propan-AdenosintriphosphatCalvin-Zyklus Reihe von enzymatischen Photosynthesereaktionen, in
denen CO2 zu 3-PGA reduziert wird, während der CO2-Akzeptor RuBP wiederhergestellt wird. Für je 6 MoleküleCO2, die den Zyklus durchlaufen, wird ein Nettogewinn an2 Molekülen 3-PGA erzielt
Cotransporter Transportprotein, welches den Transport eines Molekülsgegen einen Gradienten an einen Transport mit einemGradienten koppelt
Cytoplasma viskose Grundmasse des ProtoplastenEpidermis äußerste Zellschicht um einen mehrzelligen OrganismusEGTA Ethylenglycol-bis-(β-aminoethylether)N,N,N‘,N‘-Tetra-
essigsäure (Calciumpuffer)FeEDTA Eisen-Ethylendiamin-bis-(β-aminoethylether)N,N,N‘,N‘-
TetraessigsäureHEPES N-[2-Hydroxyethyl]piperazine-N‘-[2-ethan Schwefelsäu-
re] (Puffer zum Einstellen eines pH Wertes, pH 6,8 - 8,2)i EinzelkanalstromI Ganzzellenstrom pro FlächeKanal passives Transportmolekül mit hohen TransportratenkDa kilo Dalton; ein Dalton steht für das Gewicht eines Was-
serstoffatomsKoleoptile oft als erstes Blatt bezeichnet: beim Graskeimling schüt-
zende Scheide um das Wachstumsgewebe (Spitze desBlattes)
LAMMA Laser Mikroproben MassenanalysatorLeitbündel zu Strängen zusammengefaßtes Assimilat- und Wasser-
leitungsgewebe (= vaskuläres Gewebe)Lignin komplexes polymeres Molekül, zusammengesetzt aus
Phenylpropan-Verbindungen, gehört zu den Gerüstsub-stanzen der Zellwand und führt zur Verholzung
Lumen wässriges KompartimentMembranspannung Spannung, die über der Membran einer Zelle anliegt
(Masse im Lumen)MES 2-[N-Morpholino]ethan Schwefelsäure (Puffer zum Ein-
stellen eines pH-Wertes, pH 5,5 – 6,7)

4
M = Mesophyllzellen innere Zellen des Blattes; bei Mais: die Schicht um dieBündelscheidenzellen
NADPH Nicotinamidadenindinucleotidphosphat (siehe Photosyn-these)
Nernstspannung Spannung, die aus dem Konzentrationsgradienten einesIons resultiert
PEP Phosphoenolpyruvat (siehe Photosynthese, Abschnitt 2.5)3-PGA Phosphoglycerinsäure (3-phospho-glyceric-acid, siehe
Photosynthese, Abschnitt 2.5)P = Phloem Assimilatleitungsbahnen; Transport der Produkte der
Photosynthese durch lebende ZellenPlasmodesmen röhrenartige Proteinkomplexe (Durchmesser 30 – 60 nm),
die die Cytoplasmen aneinander grenzender Zellen ver-binden
Protoplast lebender Zellkörper (ohne Zellwand)Pumpe Proteinkomplex, welcher unter Verbrauch von ATP Ionen
entgegen einem Gradienten transportiertRinde Gewebe in der Wurzel zwischen Epidermis und vaskulä-
rem GewebeRuBP Ribulose-1,5-bis-phosphate, Produkt des Calvin-ZyklussesSuberin Komplexe Mischung aus Fettsäuren; Suberinschichten
(oder –lamellen) sind für Wasser undurchlässigSymplast Kontinuum lebender Zellen, durch Plasmodesmen verbun-
denTRIS Tris(hydroxymethyl)aminoethan (Puffer zum Einstellen
eines pH Wertes, pH 10,2 – 10,6)Vakuole vom Tonoplasten (Vakuolenmembran) umgebener Raum
innerhalb der Zelle, der Speicher- und Abfallstoffe enthält;pH < 6, oft über 90% des Zellvolumens
Vaskuläres Gewebe Gewebe, welches direkt der Leitung dient oder Leitungbewirkt
VP = Vaskuläres Parenchym Teil des vaskulären Gewebes; Funktion des vaskuläremParenchyms liegt wahrscheinlich in Transport, Speiche-rung und Abgabe von Nährstoffen, insbesondere von As-similaten
X = Xylem vaskuläres Gewebe, welches Ionen und Wasser von derWurzel durch die Pflanze transportiert

5
2 Einführung
Die vorliegende Arbeit behandelt einen Teilaspekt des Ferntransportes von Nährstoffenvon der Wurzel bis in den Symplasten (Zellraum) des Blattes. Um zu verdeutlichen, wiedie Ergebnisse dieser Arbeit einzuordnen sind, soll zunächst eine Einführung in denAufbau von höheren Pflanzen gegeben werden. Abbildungen und Erläuterungen be-schränken sich auf den Mais als Beispielpflanze. Ihre Besonderheiten im Vergleich zuanderen Pflanzen bestehen darin, daß bei allen Gräsern Bündelscheidenzellen auftretenund daß insbesondere beim Mais als C4-Pflanze die Photosynthese zwischen Meso-phyll- und Bündelscheidenzellen aufgeteilt wird.
2.1 Entwicklung und Aufbau einer Maispflanze
A B
Abbildung 1. Entwicklungsstadieneiner Maispflanze. A. Junge Mais-pflanze (ca. 1 Woche alt). Aus demMaiskorn haben sich die Wurzel unddie Koleoptile entwickelt. B. Mais-pflanzen mit vier Blättern. Die Wur-zeln übertreffen die Blätter an Län-ge. Koleoptilen, Blätter und Wurzelnwerden für Transportstudien genutzt.

6
In den ersten Tagen ernährt sich der Keimling ganz durch die im Maiskorn enthaltenenSpeicherstoffe und dann erst aus dem Boden bzw. der Nährlösung. Nährlösungskulturenhaben den Vorteil, daß die Zusammensetzung der Nährstoffe genau bekannt ist. ImRahmen des Apoplastenprojektes sind diese genau vorgegeben (Abschnitt 3.2), damitdie Ergebnisse der verschiedenen Arbeitsgruppen vergleichbar sind.
Nach dem Keimen bildet sich als erstes die Wurzel und die Koleoptile (Abbildung 1A). Die Pflanze bezieht nun aus dem Boden (oder der Nährlösung) die benötigten Nähr-stoffe, kann aber noch keine Photosynthese betreiben. Bereits mit Ausbildung desKeimblattes (Mais ist eine monokotyle Pflanze, d.h. sie besitzt nur ein Keimblatt) be-ginnt die Photosynthese. Sobald das Blatt in Kontakt mit Licht kommt, ist die Differen-zierung abgeschlossen (Evert et al., 1996). Die Pflanzenteile, z.B. junge Blätter, die(noch) keine Photosynthese betreiben, bezeichnet man als „Sink“. Sie müssen die ener-giereichen Produkte von anderen Blättern oder Speichergeweben bekommen. Ausdiffe-renzierte, photosynthetisch aktive Blätter dienen als „Source“, d.h. sie sind eine Quellevon Photosyntheseprodukten (z.B. Zucker, Aminosäuren).
Der Transport der Produkte der Photosynthese läuft über das sogenannte Phloem(Abschnitt 2.3.2) Richtung der Wurzel oder Sinkblätter; Wasser und die darin gelöstenNährstoffe aus dem Boden werden über das Xylem transportiert (Abschnitt 2.3.1).
2.2 Transportvorgänge in der Wurzel
Die komplexe Anatomie der Wurzel (Abbildung 2) muß berücksichtigt werden, wennman die Aufnahmemechanismen und –wege der Nährstoffe ins Blatt verstehen will. Dasvaskuläre Gewebe (Gewebe, welches mit Flüssigkeit gefüllt ist und dem Transportdient) ist in der Wurzel im Zentralzylinder lokalisiert. Der Zentralzylinder wird von derRinde umgeben. Die Rinde und der Zentralzylinder sind durch die Endodermis vonein-ander getrennt. Die Endodermiszellen sind in ihren Radialwänden mit einem suberi-nähnlichen Stoff, dem Endodermin, inkrustiert. Dieses Endodermisband wird auch alsCasparyscher Streifen bezeichnet. Er ist für Wasser undurchlässig. Bis zum Caspary-schen Streifen gibt es zwei Möglichkeiten des radialen Transportes, apoplastisch (überdie Zwischenräume zwischen den Zellen) oder symplastisch (über Plasmodesmen, sieheAbschnitt 2.4.1). Der Casparysche Streifen ermöglicht keinen apoplastischen Transportin den Zentralzylinder. Für den weiteren Transport muß die Aufnahme von Ionen in denWurzelsymplasten erfolgen. Die Ionenabgabe in den Apoplasten des Zentralzylindersüber die Xylemparenchymzellen erfolgt über verschiedene K+-Kanäle (KORC undNORC, Wegner und Raschke, 1994, Wegner et al., 1995, Abschnitt 10.8) und Anionen-

7
kanäle (X-IRAC, inwardly rectifying anion channel, X-SLAC, slowly activating anionconductance, X-QUAC, xylem quick activating anion conductance, Raschke et al.,1998, Köhler et al., 1999, Köhler und Raschke 1999). Die Kationenkanäle werden inAbschnitt 10.8 besprochen. Ionenaufnahme und –abgabe in den Apoplasten des Zen-tralzylinders stellen wesentliche Schritte im radialen Transport von Salzen zum Xylemund somit von der Wurzel bis ins Blatt dar.
Abbildung 2. Querschnitt durch eine Maiswurzel (Marschner, 1995). A und B markierenzwei mögliche Wege von der Rinde (= Cortex) in den Zentralzylinder: symplastisch (A) oderapoplastisch (B). Der Casparysche Streifen trennt den Apoplasten in die Bereiche innerhalbund außerhalb des Zentralzylinders (= Stele). Xylem, Phloem und Xylemparenchym liegeninnerhalb.

8
2.3 Transportvorgänge im Maisblatt
Im Maisblatt gibt es Leitbündel, die zu Strängen zusammengefaßten Assimilat- undWasserleitungsgewebe, verschiedener Ordnung mit unterschiedlichen Funktionen(Tabelle 1): Leitbündel erster und zweiter Ordnung haben sehr große Xylemelemente(Durchmesser: 20 - 40 µm) und dienen daher dem Ferntransport (nach Hagen-Poisseuille ist der Volumenfluß proportional zu r4). In den kleineren Leitbündeln (Leit-bündel dritter Ordnung, Durchmesser 7 - 15 µm) erfolgt die Verteilung der Nährstoffe indas angrenzende Gewebe diffusiv, d.h. die Fließgeschwindigkeit in den Xylemelemen-ten ist sehr gering (1/7 der Geschwindigkeit in den großen Xylemelementen, Tabelle 1,Canny, 1995, 1998) und ermöglicht so dem angrenzenden Gewebe wie z.B. den Bündel-scheidenzellen (Abschnitt 3.1, Abbildung 7) einen Austausch. Das bedeutet, daß hierradialer (senkrecht zu den Leitbündeln) statt axialer (in Richtung der Leitbündel) Trans-port stattfindet, z.B. passiv über Kanäle (Abschnitt 4.1.4).
Alle Leitbündel im Maisblatt (es gibt drei Ordnungen, Abschnitt 2.3) entwickeln sichähnlich, wobei die vaskulären Zellen (innerhalb der Leitbündel) ontogenetisch nicht mitden Bündelscheidenzellen zusammenhängen (Bosabalidis et al., 1994). Die Entwicklungder Leitbündel im Maisblatt ist bereits abgeschlossen, wenn es in Kontakt mit Licht tritt(Evert et al., 1996).
Tabelle 1. Eigenschaften von Xylemelementen (Canny, 1998) verschiedener Ordnung (großeLeitbündel = zweiter Ordnung, kleine Leitbündel = dritter Ordnung).
große Leitbündelgroße Gefäße
große Leitbündelkleine Gefäße
kleine Leitbün-del
kleine GefäßeDurchmesser / µm 20 – 40 7 – 15 7 – 15
Geschwindigkeitsfluß je Gefäß(willkürliche Einheit)
625 1 1
Geschwindigkeit(willkürliche Einheit)
7 1 1
Funktion Zufuhr ? Verteilung undAustausch
Austausch (Volumen / Ober-fläche = Durchmesser / 4)
1 3 3
Transportrichtung axial ? radial
9
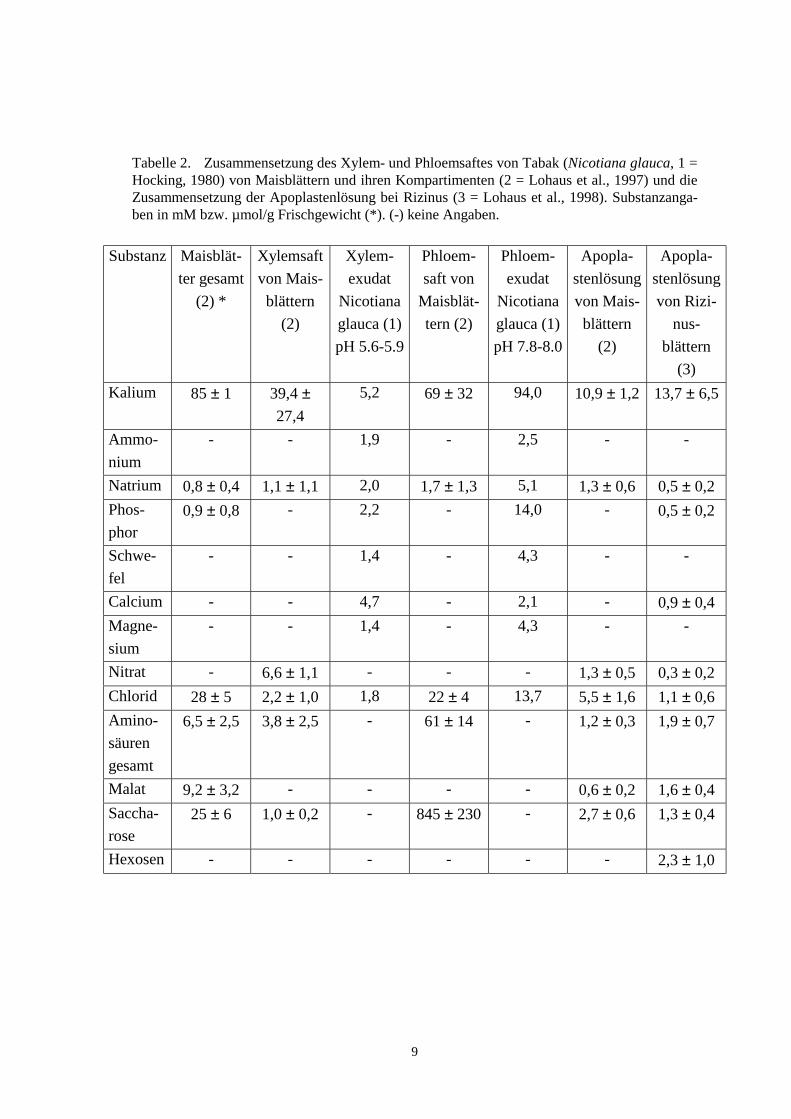
Tabelle 2. Zusammensetzung des Xylem- und Phloemsaftes von Tabak (Nicotiana glauca, 1 =Hocking, 1980) von Maisblättern und ihren Kompartimenten (2 = Lohaus et al., 1997) und dieZusammensetzung der Apoplastenlösung bei Rizinus (3 = Lohaus et al., 1998). Substanzanga-ben in mM bzw. µmol/g Frischgewicht (*). (-) keine Angaben.
Substanz Maisblät-ter gesamt
(2) *
Xylemsaftvon Mais-
blättern(2)
Xylem-exudat
Nicotianaglauca (1)pH 5.6-5.9
Phloem-saft von
Maisblät-tern (2)
Phloem-exudat
Nicotianaglauca (1)pH 7.8-8.0
Apopla-stenlösungvon Mais-
blättern(2)
Apopla-stenlösungvon Rizi-
nus-blättern
(3)Kalium 85 ± 1 39,4 ±
27,45,2 69 ± 32 94,0 10,9 ± 1,2 13,7 ± 6,5
Ammo-nium
- - 1,9 - 2,5 - -
Natrium 0,8 ± 0,4 1,1 ± 1,1 2,0 1,7 ± 1,3 5,1 1,3 ± 0,6 0,5 ± 0,2Phos-phor
0,9 ± 0,8 - 2,2 - 14,0 - 0,5 ± 0,2
Schwe-fel
- - 1,4 - 4,3 - -
Calcium - - 4,7 - 2,1 - 0,9 ± 0,4Magne-sium
- - 1,4 - 4,3 - -
Nitrat - 6,6 ± 1,1 - - - 1,3 ± 0,5 0,3 ± 0,2Chlorid 28 ± 5 2,2 ± 1,0 1,8 22 ± 4 13,7 5,5 ± 1,6 1,1 ± 0,6Amino-säurengesamt
6,5 ± 2,5 3,8 ± 2,5 - 61 ± 14 - 1,2 ± 0,3 1,9 ± 0,7
Malat 9,2 ± 3,2 - - - - 0,6 ± 0,2 1,6 ± 0,4Saccha-rose
25 ± 6 1,0 ± 0,2 - 845 ± 230 - 2,7 ± 0,6 1,3 ± 0,4
Hexosen - - - - - - 2,3 ± 1,0

10
2.3.1 Xylem: Aufbau und Funktion
Im Xylem werden das Wasser und die darin gelösten Nährstoffe aus der Wurzel bis indas Blatt transportiert. Hierbei existiert das leitende Xylem nur noch als das verholzteZellwandgerüst ehemals lebender Protoxylemzellen. Im Gegensatz dazu besteht dasPhloem (Abschnitt 2.3.2) aus lebendem Gewebe. Die Xylemelemente der verschiedenenLeitbündel haben dabei unterschiedliche Funktionen (Tabelle 1). So werden in den Leit-bündeln erster und zweiter Ordnung die Nährstoffe angeliefert und dann über die Leit-bündel dritter Ordnung im Blatt verteilt.
Pflanzen benötigen nur anorganische Nährstoffe, alle organischen Verbindungenkönnen synthetisiert werden. Dennoch finden sich kleine Mengen organischer Zuckerim Xylemsaft, in Mais z.B. bis 5 mM Zucker (Canny und McCully, 1989). Die Zusam-mensetzung der verschiedenen Xylemsäfte ist in Tabelle 2 dargestellt.
Kalium liefert den größten Anteil der Kationen. Auffallend sind die großen Schwan-kungen, insbesondere bei Mais (39,4 ± 27,4 mM). Nitrat und Chlorid stellen zusammenmit einigen Aminosäuren die Anionen. Die unterschiedliche Zusammensetzung vonXylemsaft und Apoplastenlösung (für Kalium z.B. um ca. Faktor vier) zeigt, daß inner-halb des Apoplasten Gradienten vorhanden sein müssen (Xylem macht einen Teil desApoplasten aus).
Diese Zusammensetzung hängt neben dem Nährstoffangebot auch von der Wasser-verdünnung und dem Entwicklungszustand der Pflanze (Noodén und Mauk, 1987) ab.Der pH-Wert schwankt ebenfalls, er liegt zwischen pH 5 und pH 7. Pate et al. (1964)fanden, daß bei Nitratzugabe (anstatt von Ammonium) die NO3-Konzentration mit derWeglänge abnimmt, während gleichzeitig der Gehalt an organischem Stickstoff insge-samt ansteigt.
Für die Aufnahme von Nährstoffen aus dem Xylem in den Symplasten sind zweiWege denkbar: Parenchymzellen und Bündelscheidenzellen haben direkten Kontaktzum Xylem und kommen somit hierfür in Frage. Da die Aufnahme in den Symplastenuntersucht werden soll, war es von fundamentaler Bedeutung, welche der Zellen genaudafür verantwortlich sind. In Kapitel 3 wird ausführlich auf diese Problematik einge-gangen.

11
2.3.2 Phloem: Aufbau und Funktion
Das Phloem besteht aus zwei Gefäßarten: dünn- und dickwandigen Phloemelementen.Dickwandiges Phloem ist das nicht mehr teilungsfähige Protophloem, dünnwandigesheißt Metaphloem und ist noch teilungsfähig. Beide Phloemtypen bestehen aus Siebröh-ren, den Elementen der Assimilatleitung, und Geleitzellen. Das Phloem dient zumTransport der Assimilate (Produkte der Photosynthese). Diese sind größtenteils organi-scher Natur.
Tabelle 2 stellt dies für Mais sehr anschaulich dar: alle Ionen plus Aminosäuren er-geben eine Molarität von weniger als 160 mM, dem gegenüber stehen fast 850 mMMolarität an Saccharose. Daß das Phloem zu dem lebenden Gewebe gehört (und nichtwie das Xylem zum Apoplasten), ist auch an den hohen Kaliumkonzentrationen abzule-sen (70 - 100 mM). Kalium übernimmt bei Pflanzen die dominierende Rolle, die beitierischen Zellen Natrium zukommt.
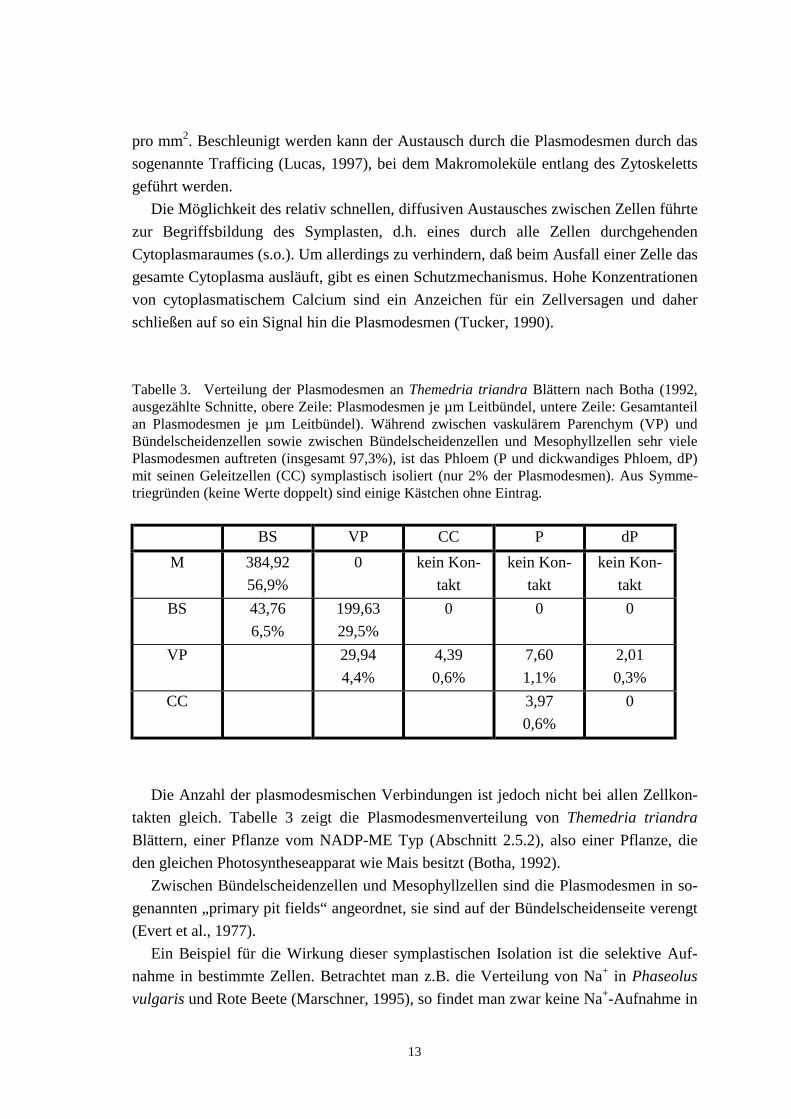
Für die Phloembeladung gibt es prinzipiell zwei Möglichkeiten: symplastische oderapoplastische. Bei der apoplastischen Beladung werden die Assimilate durch die Mem-bran über spezielle Transporter (Abschnitt 4.1) aufgenommen, während sie bei der sym-plastischen über Plasmodesmen direkt aus benachbarten Zellen in die Phloemelementegelangen. Da aber das Phloem bei Mais mitsamt den Geleitzellen symplastisch vomRest des Blattes isoliert ist (keine Verbindung über Plasmodesmen zu anderen Zellen,Abschnitt 2.4.1, Tabelle 3, Evert et al., 1993), kann die Beladung hier nur apoplastischerfolgen.
2.4 Aufbau einer Pflanzenzelle
Während die obigen Abschnitte die Grundlagen für die Einordnung der in dieser Arbeitdurchgeführten Versuche in die Funktionen des Ferntransportes beschreiben sollen, gehtes im folgenden Abschnitt um die Funktionseinheit, die diese Transportleistungen voll-bringt: die Zelle.
Abbildung 3 zeigt eine typische embryonale (A) und eine ausgewachsene (B) Pflan-zenzelle. Das Plasmalemma grenzt das von Cytoplasma gefüllte Zellinnere von derZellwand ab. Die Zelle ohne Zellwand wird als Protoplast bezeichnet. Zu anderen Zel-len besteht aber über die Plasmodesmen eine Verbindung. Die Photosynthese findet inden Chloroplasten statt, im Zellkern ist die genetische Information gespeichert. Bei derausgewachsenen Pflanze nimmt die Vakuole den Großteil des Zellvolumens ein.

12
Abbildung 3. A. Embryonale Zelle: Zellwand mit Mittellamelle und Plasmodesmen; im Cyto-plasma zwei Dictyosomen (D), glattes und rauhes Endoplasmatisches Reticulum (ER), Ribo-somen und Polysomen, verschiedene Vesikel (darunter auch Coated Vesicles, CV) und Lipid-tröpfchen (Oleosomen). Unter der Zellmembran stellenweise Mikrotubuli, längs und quer; Va-kuolen (V); im zentralen Zellkern ein Nucleolus und dichtes Chromatin; zwei Proplastiden (mitPlastoglobuli und Stärke, S) und ein Mitochondrion (mit Cistae). Die getönten Organellen ent-halten DNA. Nichtplasmatische Kompartimente sind weiß. B. Ausschnitt aus einer Gewebe-zelle mit enorm vergrößerter Vakuole, Beispiel Blattzelle. Ausgewachsene Primärwand, an denZellecken Interzellularräume (punktiert, Teil des Apoplasten); im Cytoplasma neben einemMitochondrion ein Chloroplast mit Thylakoiden, Plastoglobuli und Stärkekorn. Plasmodesmen(PD). Nach: Sitte et al. (1998).
2.4.1 Plasmodesmen
Der Transport von Zelle zu Zelle geschieht durch Plasmodesmen. Sie verbinden sowohlgleiche als auch verschiedenartige Zellen miteinander. Es handelt sich hierbei um ver-schließbare, aus Proteinen gebildete Röhren , die in ihren Ausmaßen verhältnismäßiggroß (Durchmesser von 30 – 100 nm) und daher im Elektronenmikroskop gut sichtbar(Abbildung 3) sind. Die Selektivität ist gering. So können Moleküle bis zu einer Größevon 800 Da (Durchmesser ca. 2 nm) ungehindert durch Plasmodesmen diffundieren(Sitte et al., 1998). Die Dichte der Plasmodesmen liegt gewöhnlich über einer Million
13
pro mm2. Beschleunigt werden kann der Austausch durch die Plasmodesmen durch dassogenannte Trafficing (Lucas, 1997), bei dem Makromoleküle entlang des Zytoskelettsgeführt werden.
Die Möglichkeit des relativ schnellen, diffusiven Austausches zwischen Zellen führtezur Begriffsbildung des Symplasten, d.h. eines durch alle Zellen durchgehendenCytoplasmaraumes (s.o.). Um allerdings zu verhindern, daß beim Ausfall einer Zelle dasgesamte Cytoplasma ausläuft, gibt es einen Schutzmechanismus. Hohe Konzentrationenvon cytoplasmatischem Calcium sind ein Anzeichen für ein Zellversagen und daherschließen auf so ein Signal hin die Plasmodesmen (Tucker, 1990).
Tabelle 3. Verteilung der Plasmodesmen an Themedria triandra Blättern nach Botha (1992,ausgezählte Schnitte, obere Zeile: Plasmodesmen je µm Leitbündel, untere Zeile: Gesamtanteilan Plasmodesmen je µm Leitbündel). Während zwischen vaskulärem Parenchym (VP) undBündelscheidenzellen sowie zwischen Bündelscheidenzellen und Mesophyllzellen sehr vielePlasmodesmen auftreten (insgesamt 97,3%), ist das Phloem (P und dickwandiges Phloem, dP)mit seinen Geleitzellen (CC) symplastisch isoliert (nur 2% der Plasmodesmen). Aus Symme-triegründen (keine Werte doppelt) sind einige Kästchen ohne Eintrag.
BS VP CC P dPM 384,92
56,9%0 kein Kon-
taktkein Kon-
taktkein Kon-
taktBS 43,76
6,5%199,6329,5%
0 0 0
VP 29,944,4%
4,390,6%
7,601,1%
2,010,3%
CC 3,970,6%
0
Die Anzahl der plasmodesmischen Verbindungen ist jedoch nicht bei allen Zellkon-takten gleich. Tabelle 3 zeigt die Plasmodesmenverteilung von Themedria triandraBlättern, einer Pflanze vom NADP-ME Typ (Abschnitt 2.5.2), also einer Pflanze, dieden gleichen Photosyntheseapparat wie Mais besitzt (Botha, 1992).
Zwischen Bündelscheidenzellen und Mesophyllzellen sind die Plasmodesmen in so-genannten „primary pit fields“ angeordnet, sie sind auf der Bündelscheidenseite verengt(Evert et al., 1977).
Ein Beispiel für die Wirkung dieser symplastischen Isolation ist die selektive Auf-nahme in bestimmte Zellen. Betrachtet man z.B. die Verteilung von Na+ in Phaseolusvulgaris und Rote Beete (Marschner, 1995), so findet man zwar keine Na+-Aufnahme in

14
den Blattsymplasten, aber eine selektive Na+-Aufnahme in die Xylemparenchymzellen(Rains, 1969, Drew und Läuchli, 1987).
2.5 Photosynthese
Die Unterteilung in Bündelscheidenzellen und Mesophyllzellen im Maisblatt, die indieser Arbeit getrennt untersucht werden, hängt mit ihrer speziellen Arbeitsteilung wäh-rend der Photosynthese zusammen.
Sonnenenergie in Form von Licht ist der Ursprung des Lebens. Mittels der Photo-synthese wird diese Strahlungsenergie in die Form einer chemischen Bindung umge-wandelt, die in Zellen speicherbar ist, und die für verschiedene Prozesse, z.B. chemi-sche, osmotische, elektrische oder thermische, genutzt werden kann. Gleichzeitig wirdSauerstoff freigesetzt.
Photonen werden in den Photosystemen II und I absorbiert und treiben eine Kette vonRedoxreaktionen an, die in der Thylakoidmembran lokalisiert sind. Diese Reaktions-kette wird als Elektronentransportkette bezeichnet. Als Endergebnis werden Elektronenvom Wasser auf NADPH/H+ übertragen und je zwei H+ pro Elektron ins Innere desThylakoiden (Lumen) geschafft. Der so entstandene pH-Gradient treibt eine invers be-triebene H+-Pumpe (ATPase) an, die aus ADP ATP erzeugt. Als Gesamtbilanz ergibtsich folgende Gleichung:
OH6O6OHCOH12CO6 226126h
22 ++→+ ν .
2.5.1 Der Calvin-Zyklus
Die Bindung der chemischen Energie geschieht im Calvin-Zyklus (Buschman undGrumbach, 1985, Abbildung 4, benannt nach Melvin Calvin, der dafür den Nobelpreiserhielt). Er besteht aus einem Kreisprozeß im Stroma (Thylakoidaußenraum) der Chlo-roplasten mit den folgenden drei Teilen: Karboxylierung (Fixierung von Kohlenstoff),Reduktion und Regeneration (Abbildung 4). Bei der Karboxylierung durch das EnzymRubisco binden CO2 und H2O an RuBP (ribulose-1,5-bis-phosphate), um zwei Moleküle3-PGA (3-phospho-glyceric-acid) zu formen. Da dieses erste Produkt der Photosynthese(3-PGA) 3 Kohlenstoffatome enthält, wird der Calvin-Zyklus auch C3-Zyklus genannt.Aus PGA wird unter ATP-Verbrauch in mehreren Schritten RuBP. Da dieser lineareElektronentransport den Bedarf an H+ für die ATP-Erzeugung nicht deckt, gibt es eine

15
Reihe von Hilfsprozessen: zyklischer Elektronentransport, Mehler (Abgabe von Elek-tronen an O2), Redox-Export (Backhausen et al., 1994, Krömer, 1995, Noctor und Foy-er, 1998). Insgesamt werden für die Fixierung von einem Molekül CO2 2 MoleküleNADPH/H+ und 3 Moleküle ATP benötigt.
Abbildung 4. Calvin-Zyklus und Elektronentransportkette (ohne weitere Abbildung, in denThylakoiden). In der Karboxylierung wird 3-PGA (3-phosphoglyceric acid) gebildet (welches 3C-Atome enthält und so zu dem Namen C3- Photosynthese als Synonym für Calvin-Zyklusführte). Bei der Reduktion wird die Karboxylgruppe des 3-PGA zu einer Aldehydgruppe redu-ziert (GAP = 3-phospho-glyceric-aldehyde). In weiteren Schritten wird ein C3-Körper als „Ge-winn“ abgespalten und der andere dem Zyklus wieder zugeführt.
2.5.2 Photosynthese der C4-Pflanzen
Der oben beschriebene Mechanismus der Photosynthese gilt für C3-Pflanzen. Das trifftauf alle einheimischen Pflanzen in Nordeuropa zu. Doch der Mais, der in dieser Arbeituntersucht wird, gehört zu den C4-Pflanzen. Bei ihnen ist die Gesamtphotosynthese aufzwei spezialisierte Zelltypen verteilt: Mesophyllzellen und Bündelscheidenzellen, die inscheidenartigen Lagen um die Leitbündelstränge angeordnet sind (Abschnitt 3.1, Abbil-dung 7).
Die Bündelscheidenzellen von C4-Pflanzen enthalten stärkehaltige, granafreie Chlo-roplasten, die Mesophyllzellen hingegen Chloroplasten vom Granatyp (Chloroplasten-

16
dimorphismus, Abbildung 5). Während in den Bündelscheidenzellen alle Enzyme desCalvin-Zyklusses enthalten sind, hier also der komplette Zyklus ablaufen kann, enthal-ten die Mesophyllzellen lediglich eine Reaktionskette, die als „Turbolader für CO2“ fürden Calvin-Zyklus in den Bündelscheidenzellen dient. CO2 wird in den Mesophyllzellenzuerst an PEP (Phosphoenolpyruvate) gebunden. PEP hat eine sehr hohe Affinität zuCO2, so daß die Bindung bei niedrigen CO2 Konzentrationen geschehen kann, also auchbei gering geöffneten Stomata (Schließzellen). Dadurch wird der Wasserverlust geringgehalten, und diese Pflanzen können somit auch Trockengebiete und Gebiete sehr hoherLichtintensität besiedeln. 17% der Landoberfläche sind von C4-Pflanzen besiedelt unddabei leisten sie 30% der globalen Photosynthese. C4-Pflanzen sind also durch ihre be-sonders effektive Kohlendioxidverwertung an tropische Bedingungen angepaßt, in de-nen bei hoher Lichtsättigung CO2 die Photosynthese limitiert.
Abbildung 5. Chloroplastendimorphismus in C4-Pflanzen (Gunning und Steer, 1996). Linksist die Mesophyllzelle mit einem granahaltigen, stärkefreien Chloroplasten (M) zu sehen. Diebreiten Pfeile bezeichnen die suberisierte Grenzschicht, welche von zwei Gruppen von Plasmo-desmen (P) durchbrochen wird. Rechts ist ein granafreier, stärkehaltiger Bündelscheidenchlo-roplast (BS) zu sehen. Die Chloroplasten in beiden Zellen sind nur durch eine dünne Cytoplas-maschicht gegen den die Vakuole begrenzenden Tonoplasten (T) abgegrenzt. Strich: 1µm.
Man nennt diese Pflanzen C4-Pflanzen, da in den Mesophyllzellen das PEP nachCO2 Bindung in einen C4-Körper verwandelt wird (linke Seite von Abbildung 6). Die-ser C4-Körper kann verschieden sein, je nach C4 Typ. Beim Mais ist es Malat. Das

17
Malat wird in den Bündelscheidenzellen wieder in Pyruvat zerlegt und gibt CO2 an denCalvin-Zyklus ab (rechte Seite von Abbildung 6)
Abbildung 6. Photosynthese bei C4-Pflanzen (NADP-Malatenzym-Typ). Im Cytoplasma derMesophyllzelle findet die PEP-Carboxylase (1) statt, Oxalacetat wird dann im Chloroplastenvon der Malatdehydrogenase (2) in Malat umgewandelt und in die Bündelscheidenchloropla-sten transportiert. Dort wirkt das NADP-Malatenzym (3) und auch der Calvin-Zyklus findethier statt. Das entstandene Pyruvat wird zurück in den Mesophyllchloroplasten gebracht unddurch die Pyruvat-Phosphat-Dikinase (4) und die Pyrophosphatase (5) in PEP umgewandelt.

18
3 Bestimmung des Übertrittsortes der Ionen ausdem Xylem in den Blattsymplasten
3.1 Motivation
Im Kapitel 2 ist der Weg der Nährstoffe von der Wurzel ins Blatt beschrieben worden.Die Messungen dieser Arbeit beschäftigen sich mit der darauf folgenden Aufnahme derNährstoffe vom Xylem in den Blattsymplasten. Bei diesem Transport vom apoplasti-schen Raum (Xylem) in den symplastischen Raum muß eine Plasmamembran überquertwerden. Hier kommt die Patch-Clamp-Technik (Abschnitt 4.5) zum Einsatz (Untersu-chung der Transporter in der Membran, Kapitel 7-9). Voraussetzung für diese Untersu-chungen ist jedoch, daß bekannt ist, welche der an das Xylem angrenzenden Zellen diezu untersuchende Transportfunktion übernehmen.
Diese Frage war noch nicht geklärt. Dies zeigte sich zum einen auf der DFG-Schwerpunkttagung des Apoplastenprojektes 1997 in Hannover, auf der es wider-sprüchliche Ansichten über den Transportweg vom Xylem in den Symplasten gab. Zumanderen führten auch die aus der Literatur bekannten Untersuchungen zu keinem voll-ständigen Bild.
Zum Verständnis der hier zu berichtenden Ergebnisse wird die bereits im Kapitel 2angesprochene Morphologie der Blattadern (Leitbündel) noch etwas detaillierter vorge-stellt. Abbildung 7 zeigt einen Schnitt durch die Leitbündel zweiter und dritter Ordnungim Maisblatt. Die dort eingezeichneten Benennungen werden bei der Beschreibung derUntersuchungen mit dem Laser-Mikroproben-Massenanalysator (LAMMA, Abschnitt3.4) benutzt.
Ein Teil der Arbeiten zum Fluß vom Xylem in den Blattsymplasten stützt sich aufFarbstoffuntersuchungen, bei denen mit dem Transpirationsstrom membranimpermea-ble Farbstoffe ins Blatt transportiert werden. Für einen direkten X-BS Übertritt (vomXylem in die Bündelscheidenzellen, Abbildung 7) sprechen Farbstoffablagerungen inder Zellwand im X-BS Bereich sowohl von Preußisch-Blau-Kristallen (Evert et al.,1985) als auch von Sulphorhodamin (Canny, 1990 a). Dies zeigt, daß der Fluß vonWasser aus dem Xylem direkt in den Apoplasten des Phloems und in die Bündelschei-denzellen geht (daher oft als „parenchymatic bundle sheath“ bezeichnet), wobei dieFarbstoffe an der Stelle des Membrandurchtrittes "hängenblieben". Dies war allerdingsnur eine Markierung des Wasserweges. Daß Ionen den gleichen Weg gehen, war einplausibler, aber nicht gesicherter Schluß.

19
Für die Zuckeraufnahme ergab sich ein anderes Bild. Daß Zucker zuerst in die Par-enchymzellen aufgenommen wird, zeigte folgender Versuch: Abgeschnittene Blattspit-zen, die für 2-8 Minuten in eine [14C] Sucrose-Lösung gegeben wurden, zeigten eineAnhäufung der radioaktiven Sucrose erst im Xylem, dann im Parenchym und dann imPhloem (Fritz et al., 1983), während in den Bündelscheidenzellen keine Ablagerungenauftraten.
Abbildung 7. Schnitt durch ein Maisblatt. Links: Leitbündel zweiter Ordnung, rechts: Leit-bündel dritter Ordnung, beide Leitbündel jeweils mit Xylem (X), Phloem (S) mit Geleitzellen(CC bei Leitbündel dritter Ordnung rechts), vaskulärem Parenchym (VP, Xylem- und Phloem-parenchym) und Bündelscheide (BS). Die Bündelscheide ist eingebettet in Mesophyll (M),wobei maximal zwei Mesophyllzellen zwischen zwei Leitbündeln liegen. Zu erkennen sindauch die Schließzellen (G, haben im Gegensatz zur Epidermis (E) Chloroplasten). Unter derSchließzelle ist besonders deutlich ein Teil des gasgefüllten Interzellularraumes (gasgefüllterApoplast) zu sehen. In den Interzellularen findet der CO2-Transport statt, denn die Diffusion inder Gasphase ist etwa 105 mal größer als in Wasser. Die weiteren „inneren Zellen“ (I) zwischenden Xylemelementen bei Leitbündeln zweiter Ordnung (Abbildung links) sind schwierig zuunterscheiden. Ihnen kommt hier keine Bedeutung zu, deshalb wurden sie zusammengefaßt.

20
Es gibt Hinweise, daß der Export von Photosyntheseprodukten aus den Bündelschei-denzellen über das Parenchym zu läuft. Anfütterungen über die Luft mit 14CO2 führtenzuerst zu einer allgemeinen Erhöhung der Radioaktivität und dann bevorzugt im Paren-chym und im Phloem.
Das Vorliegen verschiedener Wege für anorganische Nährstoffe und Photosynthese-produkte ergab sich aus Messungen an Source- und Sinkblättern (Russin et al., 1996).Die verwendete Mutante (sucrose export defective1 maize mutant) bildet keine funkti-onsfähigen Plasmodesmata zwischen Bündelscheiden- und Parenchymzellen aus.
Solange das Blatt als Sink-Blatt arbeitet, d.h. daß die Photosynthese noch nicht in derLage ist, die Eigenversorgung zu decken, bzw. einen Überschuß an Stoffwechselpro-dukten zu erzeugen, der exportiert wird, wächst es normal. Das heißt, in dieser Phase, inder hauptsächlich Nährstoffe aufgenommen werden, kommt die Unterbrechung zwi-schen Bündelscheidenzellen und Parenchymzellen nicht zum Tragen. Nach dem Sink-Source-Übergang sammelt sich Stärke in den Parenchymzellen an. Das Blatt kann kei-nen Zucker exportieren, was dazu führt, daß diese Zellen plasmolysieren. Der Normal-wuchs im Sinkzustand ist damit zu erklären, daß der Import direkt vom Xylem in dieBündelscheidenzellen geschieht, während die Stärkeanhäufung zeigt, daß der Exporthauptsächlich über die Parenchymzellen läuft. Dies bestätigten die Ergebnisse mit ra-dioaktiv markiertem CO2 (Fritz et al., 1983).
Eine unterschiedliche Rolle der Leitbündeln sowohl zweiter als auch dritter Ordnungergab sich aus Untersuchungen von Canny (1985, 1993) und Russell und Evert (1985).Während in den Leitbündeln zweiter Ordnung hauptsächlich der Ferntransport stattfin-det, übernehmen die Leitbündel dritter Ordnung die Verteilung der Nährstoffe.
Das oben beschriebene komplexe Bild und die Tatsache, daß direkte Untersuchungenüber Ionenströme bislang fehlten, machten eigene Untersuchungen notwendig. Es solltedaher versucht werden, den Weg der Ionen mit Tracern zu verfolgen. Ein viel benutzterTracer für Kalium ist Rubidium (Brindley, 1990, Cocucci und Sacchi 1993, MacRobbie,1995, Wu und Assmann, 1995). Da aber auch zwei K+-Kanäle bekannt sind, die für Rb+
nicht durchlässig ist (Hedrich et al., 1995, Bregante et al., 1997), ist in eigenen Patch-Clamp-Messungen überprüft worden, ob die K+-Kanäle in den hier interessierendenBündelscheidenzellen Rb+ durchlassen (Abschnitt 8.6).
In Zusammenarbeit mit A. Schulz (damals Botanik, Kiel, jetzt Kopenhagen) wurdenSchnitte erstellt und mit LAMMA (U. Seydel, FZ Borstel) ausgewertet.

21
3.2 Verwendete Pflanzen
Verwendet wurden Mais-Pflanzen der Sorte Helix. Die Samen quollen 6 Stunden inLeitungswasser vor, bevor sie auf mit Leitungswasser befeuchtetes Filterpapier gelegtwurden. Nach 4 Tagen wurden die Keimlinge mit den Wurzeln für eine Woche in be-lüftetes Leitungswasser gestellt. Danach wurden die Maispflanzen in Schaumstoff ein-gewickelt und jeweils vier Pflanzen in einen 5 l Eimer mit Apoplastennährlösung (0.5mM NH4NO3, 0.7 mM K2SO4, 0.1 mM KCl, 2 mM Ca(NO3)2, 0.5 mM MgSO4, 0.1 mMKH2PO4, 1 µM H3BO3, 0.5 µM MnSO4, 0.1 µM ZnSO4, 0.2 µM SnSO4, 10 nM(NH4)6MO7O24, 1µM FeEDTA) unter Normallicht in einem Gewächshaus angezogen.Der Lösungswechsel erfolgte zweimal pro Woche.
Für die LAMMA Versuche wurden etwas ältere Pflanzen verwendet (fünf bis sechsWochen alt, 7 bis 8 Blätter) als für die Patch-Clamp-Untersuchungen, da ältere sichbesser für die Cryomicrotomschnitte eigneten.
3.3 Anfütterung mit Rubidium und Erstellen der Schnitte
Das vorletzte Blatt (von 7 oder 8) wurde abgeschnitten und etwa ein Millimeter tief mitder Schnittstelle in 100 ml einer 20 mM RbCl-Lösung (niedrigere Rubidiumkonzentra-tionen führten zu keiner mit dem LAMMA nachweisbaren Beladung) gehalten. Die ge-ringe Eintauchtiefe verhinderte ein eventuelles Eindringen von Rubidium durch dieEpidermis.
Nach 22,5 Minuten wurde neben dem Leitbündel erster Ordnung 2 mm oberhalb deralten Schnittstelle ein 2 mm breites und 4 mm langes Stück herausgeschnitten und so-fort in mit flüssigem Stickstoff gekühltem Freon getaucht (dies verhindert Eiskristall-bildung). In gefrorenem Einbettmedium (Jung, über Leica, Benzheim) wurde ein 3 mmbreiter Spalt geschmolzen, die Probe hineingesteckt, sofort mit weiterem Einbettmedi-um übergossen und durch Stickstoffschnee gekühlt. Das Einbettmedium wirkt hierbeiwie ein Isolator, das heißt, es läßt keine Wärme an die Probe heran.
Das Schneiden wurde mit einem Cryomicrotom durchgeführt. Die besondereSchwierigkeit lag in der Herstellung sehr dünner Schnitte (< 10 µm), die für eine aus-reichende räumliche Auflösung mit dem Massenanalysator (LAMMA) benötigt wurden.Es gelang eine schrittweise Verfeinerung der Schneidetechnik, bis schließlich 5 µmdünne Schnitte erreicht wurden.
Die Schnitte wurden auf mit Formvar beschichtete Kupfergitter (Plano, Wetzlar) ge-legt und dann gefriergetrocknet. Der Transport von Kiel nach Borstel erfolgte in einemExsikkator mit Trockenperlen.

22
3.4 Analyse mit LAMMA
Abbildung 8. Aufbau des Laser-Mikroproben-Massenanalysators (LAMMA). Unter einemTransmissionsmikroskop (Leybold-Hereaeus, Köln) betrachtet man die auf einem mit Formvarbeschichteten Gitter aufgebrachte Probe (vergrößerter Ausschnitt). Mit dem Pilotlaser wird dieProbe anvisiert und dann mit dem Pulslaser verdampft. Die nun freien Ionen werden über einTOF (time-of-flight) Massenspektrometer analysiert mit einer Auflösung von m/∆m = 650, dasSignal wird verstärkt, mit einem Transientenrekorder (LeCroy TR 8818, Heidelberg) in einerAbtastrate von 10 ns aufgenommen, auf einem Oszilloskop angezeigt und auf dem Rechnergespeichert.
LAMMA (Abbildung 8) ist ein hauptsächlich qualitatives Verfahren zum Nachweisgeringer Mengen von Ionen. Hierbei verdampft ein Laserstrahl das Material im Zielbe-reich (NdYAG Laserpuls, 108 W/cm2, Durchmesser der verdampften Probe ca. 1 µm,Seydel et al., 1992), und ein Massenspektrometer analysiert die ausgetretenen Substan-zen. Verschiedene Schichtdicken und Matrizen (die Probe umgebende Medien) führenzu unterschiedlichen Ergebnissen. Daher war es nötig, ein Verfahren zu finden, mit demvergleichbare Meßwerte gewonnen werden konnten.
Vorversuche ergaben, daß nach 22.5 Minuten die erste Front des Rubidiums den Be-reich erreichte, in dem der Schnitt lokalisiert wurde. Bei einer Inkubation von wenigerals 20 Minuten ließ sich kein Rubidium nachweisen, nach mehr als einer halben Stundewaren alle Bereiche gesättigt, es trat eine Gleichverteilung ein; 22.5 Minuten stellte sich

23
als optimale Inkubationszeit heraus. Doch die Menge an Rubidium, die nach einer Zeitvon 22.5 Minuten durch die Leitbündel in die Blattzellen transportiert worden war, er-reichte nur geringe Werte.
Um die Streuung zu vermindern, wurde nach einer natürlichen Normierung gesucht.Hierzu bietet sich das bereits in den Zellen vorhandene Kalium an, das ebenfalls vomLAMMA angezeigt wird. Als Maß für den vorangeschrittenen Rubidiumtransport wur-de nicht die absolute Höhe des Rubidiumpeaks genommen, sondern das Verhältnis vonKalium zu Rubidium betrachtet.
Abbildung 9. LAMMA Spektrum eines BS-BS Übergangs. Die Ionenpeaks folgender Ionenkönnen zugeordnet werden: 39K+, 41K+, 85Rb+ und 87Rb+. Der 39K+-Peak zeigt Übersteuerung.
Den größten Peak im LAMMA-Spektrum ergibt 39K+(Abbildung 9). Doch leider lie-ßen sich die beiden Rubidium Peaks (85Rb+ und 87Rb+) nicht zusammen mit dem 39K+-Peak in einem Spektrum darstellen. Nach 22,5 Minuten Anfütterung mit Rubidiumlö-sung war das Verhältnis von Kaliumkonzentration zu Rubidiumkonzentration sehr groß.Das führte zu Skalierungsproblemen: war der 39K+-Peak vollständig zu erkennen, soverschwanden die Rubidiumpeaks im Rauschen.

24
Das natürliche Isotopenverhältnis von 39K+ zu 41K+ beträgt 13,7 und von 85Rb+ zu87Rb+ 2,6. Daher wurde das Verhältnis der Peaks von 41K+ zu 85Rb+ betrachtet und dasVerhältnis von 85Rb+ und 87Rb+ verfolgt, um zu kontrollieren, ob auch tatsächlich Rubi-dium ausgewertet wurde und nicht etwa organische Bruchstücke zufällig in den Rubidi-umpeak fielen.
Trotz dieser Normierung mit simultan gemessenen Peaks zeigte sich, daß dieSchwankungen von Leitbündel zu Leitbündel beträchtlich waren. Eine Ursache wird inunterschiedlichen morphologischen Gegebenheiten vermutet. Die unterschiedlichenLeitbündel (Abbildung 7, zweiter und dritter Ordnung) haben verschiedene Funktionen,wie oben erwähnt. Je näher nun das untersuchte Leitbündel dritter Ordnung an einemLeitbündel zweiter Ordnung lag, desto eher kam Rubidium dort an, denn alle Leitbündelsind durch transversale Leitbündel miteinander verknüpft (Abstand der Transversalen:1,19 mm, Abstand der Leitbündel dritter Ordnung: 0.08 mm (Evert et al., 1978); zwi-schen zwei Leitbündeln zweiter Ordnung liegen etwa 10 Leitbündel dritter Ordnung).
Um dieser Variabilität zu begegnen, wurde eine zweite Normierung eingeführt. DieFragestellung dieser Untersuchungen ist nicht, wieviel Rubidium ankommt, sondern woes zuerst ankommt. Daher wurden die 85Rb+/41K+ Verhältnisse aller Leitbündel auf dasWichtigste normiert: Dies ist der Übergang vom Xylem zur Bündelscheide (X-BS inAbbildung 7).
Um festzustellen, ob die Abweichung der Verhältnisse vom Übergang X-BS signifi-kant abweichen, wurde der Student-t-test durchgeführt. Ausgegangen wird von derNullhypothese ( )BSXx −< , wobei x der Mittelwert des zu untersuchenden Über-
gangs und BSX − = 1 (durch die Normierung) ist. Die Nullhypothese ist mit einerWahrscheinlichkeit geringer α anzunehmen, wenn
( )
σ−−< αz
nBSXx gilt
(ebenso für die Nullhypothese ( )BSXx −> die Bedingung ( )
σ+−> αz
nBSXx ),
hierbei ist αz die Student-t-Verteilung mit n Freiheitsgraden (n = Anzahl der Messun-gen). Ihre Werte können dem Bronstein (Tabelle 1.1.2.11, Seite 22) entnommen wer-den; für α = 5% ergibt sich αz = 1.645. Die Ergebnisse dieser Untersuchung sind inTabelle 4 und Tabelle 5 dargestellt.

25
3.5 Ergebnisse
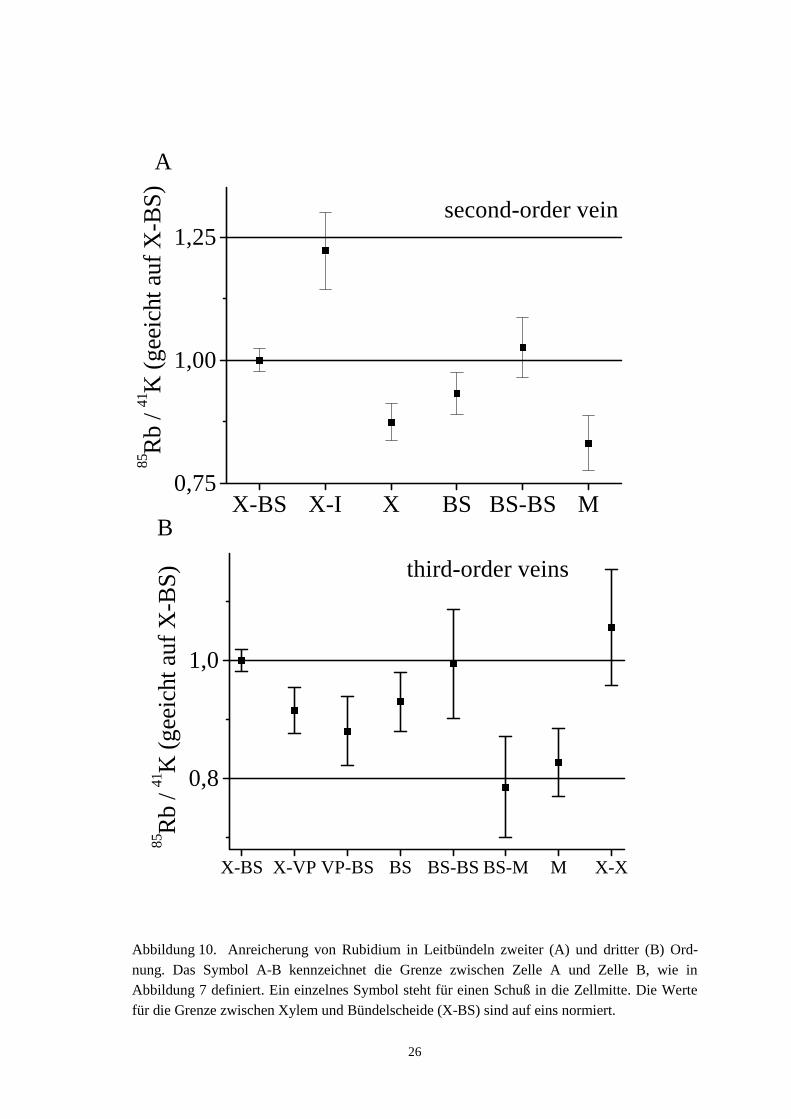
Abbildung 10 zeigt die normierten 85Rb+/41K+-Verhältnisse für Leitbündel zweiter unddritter Ordnung. Die einzelnen Punkte sind Mittelwerte aus 19 bzw. 42 Leitbündelnzweiter bzw. dritter Ordnung, bei denen jeweils 5 – 13 Schüsse (im Mittel 8, Leitbündelzweiter Ordnung), bzw. 2 – 9 Schüsse (im Mittel 5, Leitbündel dritter Ordnung) je Be-reich aufgenommen wurden. Die Bezeichnungen sind in Abbildung 7 definiert und inder Legende von Abbildung 10 für die Grenzflächen erklärt. Wie oben erwähnt, sindalle Daten zusätzlich auf eins für den X-BS Bereich normiert. Die Normierung auf die85Rb+/41K+-Verhältnisse löst auch folgendes Problem: Wenn die Zellen geschnittenwerden, ist nicht abzusehen, was mit dem Zellinhalt geschieht. Es könnte passieren, daßder mittlere Bereich verloren geht, oder daß er sich an den Rand zurückzieht, oder aberdaß alles unverändert bleibt. Hier zahlt es sich aus, daß durch die Quotientenbildung85Rb+/41K+ ein Ratio-Verfahren eingeführt wird. Da Aufkonzentrierungen an der Zell-wand sich auf K+ und Rb+ gleich auswirken, verändern sie das Verhältnis nicht.
Die Verhältnisse in den Leitbündeln zweiter Ordnung zeigt Abbildung 10 A. In Hin-blick auf die Wirksamkeit der Normierung des Ratio-Verfahrens ist wichtig, daß dieLAMMA-Schüsse in die Wandbereiche X-BS und BS-BS gleiche Verhältnisse ergeben.Rb+ hat sich also gleichmäßig im Cytosol der Bündelscheidenzellen verteilt, und derSchuß in den Wandbereich erfaßt hauptsächlich die anhaftende Cytosolschicht. Deretwas niedrigere BS-Wert enthält wahrscheinlich Vakuolenanteile, in denen die Rb+-Anreicherung noch nicht so weit fortgeschritten war. Die Mesophyllzellen liegen alsletzte im Flußweg, und somit ist eine erniedrigte Rb+ Konzentration (M-Wert, auchTabelle 4) zu erwarten.
Größere Werte für den Xyleminhalt (X-Wert in Abbildung 10 A) konnten nur seltenausgemacht werden. Xyleminhalt ist auch nur zu erwarten, wenn die Xylemzelle nochnicht vollständig ausdifferenziert ist (A. Schulz, Kopenhagen, persönliche Mitteilung),d.h. wenn sie noch Organelle und Cytoplasma enthält. Arbeitet sie als „Transportzelle“,so enthält sie nur noch Lösung, keine festeren Bestandteile, und die kann in den großenXylemgefäßen zweiter Ordnung bei der Präparation nicht erhalten bleiben. Es ist daherzu erwarten, daß dieser Wert unter dem durchschnittlichen Xylem-Bündelscheiden-Übergangswert liegt.
26
Abbildung 10. Anreicherung von Rubidium in Leitbündeln zweiter (A) und dritter (B) Ord-nung. Das Symbol A-B kennzeichnet die Grenze zwischen Zelle A und Zelle B, wie inAbbildung 7 definiert. Ein einzelnes Symbol steht für einen Schuß in die Zellmitte. Die Wertefür die Grenze zwischen Xylem und Bündelscheide (X-BS) sind auf eins normiert.
X-BS X-I X BS BS-BS M0,75
1,00
1,25second-order vein
A85
Rb
/ 41K
(gee
icht
auf
X-B
S)
X-BS X-VP VP-BS BS BS-BS BS-M M X-X
0,8
1,0
third-order veins
B
85R
b / 41
K (g
eeic
ht a
uf X
-BS)

27
Vor diesem Hintergrund, der sehr gut den Erwartungen entspricht, ist die im X-I-Wert liegende Aussage signifikant (Tabelle 4): Der Übergang X-I vom Xylem der Leit-bündel zweiter Ordnung zu weiter innen liegenden Zellen (I in Abbildung 10) hat einhöheres 85Rb+/41K+-Verhältnis als der Übergang zur Bündelscheide. Dies zeigt, daß derTransportweg vom Xylem nicht direkt zur Bündelscheide führt. Stattdessen wird Rb+
erst in innen liegende Zellen (u. a. Parenchymzellen) aufgenommen und geht dannsymplastisch in die Bündelscheidenzellen.
Bei den Leitbündeln dritter Ordnung (Abbildung 10 B, Tabelle 5) ergibt sich ein an-deres Bild: Wieder zeigen die gleichen Werte bei BS-BS und X-BS eine gleichmäßigeRb+-Verteilung im Cytosol der Bündelscheidenzellen an. An der abnehmenden FolgeBS-BS, BS-M, M sieht man, daß die Messung genau in dem Zeitbereich stattfindet, indem die Rb+-Front sich ausbreitet und somit der Weg des Rb+ noch nachzuvollziehenist. Wichtig ist jetzt, daß, im Gegensatz zu Abbildung 10 A, die Werte für VP-BS undX-VP tiefer als 1.0 liegen (allerdings beinhaltet Tabelle 5, daß für X-VP die Irrtums-wahrscheinlichkeit über 5% liegt, daß dieser Wert unter X-BS liegt, wahrscheinlichwegen eines hohen X-Anteils in X-VP), genau wie die Werte für BS-M, BS und M.Damit ist gezeigt, daß bei den Venen dritter Ordnung das Rb+ als Tracer für K+ direktvom Xylem in die Bündelscheidenzellen aufgenommen wird und nicht wie inAbbildung 10 A erst den Umweg über innen liegende Zellen (Parenchymzellen) neh-men muß.
Die X-X Wände, die in Abbildung 10 A fehlen, haben natürlich noch höhere Werteals die X-BS Wände. Auch dieses Ergebnis bestätigt, daß die 85Rb+/41K+-Verhältnisseein Verfolgen des Weges der Kationenaufnahme zulassen. Daß die B- und M-Werte soniedrig liegen, hängt wahrscheinlich wieder mit der Vermischung mit schwächer ange-reicherten Vakuoleninhalten zusammen.
Zusammenfassend läßt sich feststellen, daß in Leitbündeln dritter Ordnung derTransport vom Xylem direkt in die Bündelscheide geht. Das bedeutet, daß hier derApoplast-Symplast-Übergang lokalisiert ist, und damit die Patch-Clamp-Messungen anBündelscheidenzellen richtig lokalisiert sind.

28
Tabelle 4. Statistische Auswertung für die Leitbündel zweiter Ordnung (Abbildung 7 links).Berechnung der Student-t-Verteilung für α = 0.05 und somit zα = 1.645. Jeder in der Spalte„Anzahl“ auftauchende Wert ist ein Mittelwert aus 3 bis 12 Werten. Nur der X-I Wert ist größerals der normierte X-BS Wert.
Übergang Mittelwert σ = sd Anzahl 1 ± σ/√n*zα ErgebnisX-BS 1 0,453 151X-I 1,222 0,407 73 1,078 >X 0,874 0,355 88 0,938 <
BS-BS 1,025 0,327 29 1,100 ≈BS 0,932 0,376 78 0,930 ≤M 0,831 0,332 35 0,908 <
Tabelle 5. Statistische Auswertung für die Leitbündel dritter Ordnung (Abbildung 7 rechts).Berechnung der Student-t-Verteilung für α = 0.05 und somit zα = 1.645 (bzw. zα = 1,895 fürBS-M; da n < 30, ist dieser Wert größer). Jeder in der Spalte „Anzahl“ auftauchende Wert istein Mittelwert aus 3 bis 12 Werten. X-X und BS-BS sind dem X-BS Wert gleich, während diedas Vaskuläre Parenchym betreffenden Werte kleiner sind (X-VP und VP-BS).
Übergang Mittelwert σ = sd Anzahl 1 ± σ/√n*zα ErgebnisX-BS 1 0,589 205X-X 1,055 0,434 31 1,128 =
X-VP 0,915 0,444 139 0,937 <VP-BS 0,880 0,430 53 0,903 <BS-BS 0,994 0,734 63 0,855 =BS-M 0,785 0,226 7 0,859 <
BS 0,930 0,466 88 0,918 ≤M 0,827 0,500 74 0,904 <

29
4 Biomembranen und Membrantransport
Membranen grenzen Zellen und die in ihnen enthaltenen Organellen (das sind Funkti-onseinheiten, in oder an denen bestimmte Reaktionsketten ablaufen, Schnepf, 1977)gegen ihre Umgebung ab. Die Pflanzenzelle selbst ist vom Plasmalemma begrenzt undzusätzlich umgeben von einer starren Zellwand. Zu den Organellen gehören u.a. dieVakuolen (enthalten Speicher- und Abfallstoffe), Chloroplasten (Organelle der Photo-synthese), Mitochondrien (Organelle der Zellatmung) und der Zellkern (enthält die Er-binformation), die alle in das Cytoplasma (viskose Grundmasse der Zelle) eingebettetsind (siehe auch Abschnitt 2.4).
Transporter in der Membran (Proteine, die die Membran überspannen) sorgen dafür,daß nur bestimmte Stoffe die Membran passieren können, sie sind somit semipermeabel.Die Zusammensetzung innerhalb der Membran wird auf diese Art kontrolliert, so daßdie lebensnotwendigen Prozesse ablaufen können, auch wenn sich die äußere Umge-bung verändert. Die im folgenden zu beschreibenden Patch-Clamp-Messungen beschäf-tigen sich mit dem Transport durch die Plasmamembran. Deshalb folgen hier einigeGrundlagen.
Abbildung 11. Schema einer biologischen Membran (Singer und Nicolson, 1972). Nach dem„Fluid-Mosaic-Modell“ wird die zähflüssige Membran von integralem Membranproteinen durch-setzt, die mit ihren hydrophoben Domänen in einen Doppelfilm aus Strukturlipiden integriert sind.Einige dieser Proteine fungieren als Ionenkanäle. Die inneren Enden der Phospholipide, die Phos-pholipidschwänze, sind frei beweglich und ebenfalls hydrophob. Stereolipide sind in den unpola-ren Bereich der Lipidbilayer eingelagert, sie dienen zur Stabilisierung der Membran.

30
4.1 Pumpen, Cotransporter und Uniporter
Viele verschiedene Proteine sind in die Lipiddoppelschicht der Membran eingebettet(Abbildung 11), unter anderem auch die Transportmoleküle, die für den selektivenTransport bestimmter Ionen oder Moleküle durch die Membran verantwortlich sind.Neben den Transmembranproteinen gibt es auch rein periphere Membranproteine, diejedoch keinen Anteil am Membrantransport haben (Abbildung 11).
Ordnet man die Transportmoleküle nach ihren Energiequellen, so unterteilen sie sichin drei Klassen: Pumpen, Cotransporter und Uniporter (Maathuis und Sanders, 1992).
Abbildung 12. Schema der Transporter in einer Membran. Die Pumpe transportiert H+ entgegendem pH Gradienten aus der Zelle in den Apoplasten. Durch den Kanal (Uniporter) diffundierenK+ Ionen in die Zelle (von der negativen Membranspannung (Abschnitt 4.2) getragen). Der Trans-port von NO3
- wird im Cotransporter an den Import von 2 H+ gekoppelt (Symport).
4.1.1 Pumpen
Pumpen sind Enzyme, die in jeder Membran jeder lebenden Zelle enthalten sind. Siewandeln chemische Energie in Ionenbewegung um und umgekehrt. Meist wird aus derUmwandlung von ATP in ADP und H2PO4
- die benötigte Energie (etwa 50 kJ/mol =520 meV je Umsetzung, Karlson et al., 1994) gewonnen:
MgATP + H2O ↔ MgADP + H2PO4-
Diese Reaktion wird von der ATP-Phosphohydrolase, abgekürzt ATPase, katalysiert.Die freiwerdende Energie kann auch zum gekoppelten Transport verwendet werden,z.B. zum Export von Ca2+ oder Na+ aus dem Cytosol in die Vakuole oder aus der Zelleheraus unter gleichzeitigem Import von H+ (Ca2+/H+ ATPase ebenso Na+/K+ ATPase).

31
4.1.2 Cotransporter
Cotransporter bewegen Ionen oder ungeladene Moleküle gegen ihren elektroosmoti-schen Gradienten durch die Membran, indem sie deren Fluß an einen Transport einesanderen Ions oder Moleküls in Richtung des Gradienten koppeln. Erfolgt der Transportbeider Ionen oder Moleküle in gleicher Richtung, so spricht man von Symport, bei ent-gegengesetzter Richtung von Antiport (Abbildung 12, Sanders, 1988).
4.1.3 Carrier
Es gibt zwei Arten von Uniportern: Kanäle und Carrier. Beides sind passive Transpor-ter, bei denen sich Ionen in Richtung eines elektrochemischen Gradienten bewegen.
Carrier sind kugelförmige hydrophobe Moleküle, die sich frei in der Membran bewe-gen können. Sie nehmen Ionen auf der einen Seite der Membran auf und durchquerenmit ihnen die Membran, d.h. sie nehmen die Ionen „huckepack“ von einer Seite auf dieandere mit. Auf diese Art erreichen sie Transportraten von 103-104 Molekülen oder Io-nen pro Sekunde (Läuger, 1980). Heute setzt sich die Vorstellung immer mehr durch,daß die meisten Transporter Kanäle sind (Galli et al., 1995, Conejero, 1997 ). WirklicheCarrier sind beispielsweise die von Pilzen eingesetzten Kampfstoffe wie Valinomycin(Benz und Läuger, 1976) oder Nigericin. In Pflanzen gibt es H+-Carrier in der Thyla-koidmembran (Plastoquinon) und in den Mitochondrien. Allerdings werden insbesonde-re Sucrosetransporter häufig ebenfalls als Carrier bezeichnet. Daß diese nicht mit obigerDefinition eines Carriers zu vereinbaren sind, sieht man unter anderem daran, daß soge-nannte Sucrose-Carrier aus 12 Helices bestehen (Truernit et al., 1999), die die gesamteMembran überspannen.
4.1.4 Kanäle: Struktur und Modelle
Kanäle können als Poren betrachtet werden, die die Lipiddoppelschicht überbrücken(Abbildung 11, Abbildung 13). Etwa 107 Ionen pro Sekunde können einen Kanal durch-queren. Der Kanal läßt die Ionen dabei nicht in kontinuierlichem Strom auf die andereSeite der Membran, sondern er öffnet und schließt sich. Die Patch-Clamp-Methode (Ab-schnitt 4.5) ermöglicht eine direkte Beobachtung des Öffnens und Schließens eines ein-zelnen Kanals.

32
Mit Hilfe biochemischer Methoden ist es möglich, die Sequenz eines Makromoleküls(z.B. eines Kanals) zu bestimmen. Allerdings ist die Sekundär- und Tertiärstruktur, alsodie räumliche Anordnung der Kette, und die Funktion der einzelnen Aminosäuren damitnoch nicht geklärt. Mutagenese, das ist der Austausch einzelner Aminosäuren in derKette, ermöglicht die funktionelle Zuordnung. Gelingt es, die Kanäle in eine Kristall-struktur zu bringen, die hinreichend gut streut (je größer der Kristall, desto besser ist i.A. die Auflösung), so läßt sich die dreidimensionale Struktur ermitteln.
A B
Abbildung 13. Das Kanalmodell des KscA aus Streptomyces lividans nach Doyle et al. (1998).A. Das Selektivitätsfilter (1,2 nm lang) mit zwei eingezeichneten K+ Ionen an der Außenseite(oben). In der Mitte liegt eine Hohlkugel mit einem Durchmesser von 1 nm. Die Verbindungzur Innenseite (unten) vermittelt ein etwas breiterer Kanal. Die beiden kleinen schrägstehendenHelices stützen die Struktur ab. Die Ionen müssen ihre Hydrathülle abstreifen, um in den Kanalzu gelangen. Negative Ladungen an ihrer Innenseite führen zur Erhöhung der Aufenthaltswahr-scheinlichkeit von Ionen in der Hohlkugel vor dem Selektivitätsfilter. B. Anblick des Kanalsvon oben. Die viermal zwei Helices bilden das Gerüst des Kanals. Im Gegensatz zu den meistenK+ Kanälen ist dieser einfache Bakterienkanal nur aus zwei alpha-Helices je Untereinheit auf-gebaut und nicht aus sechs.
Bekannt sind bislang der Aufbau des einfach gebauten Gramicidinkanals (er enthältkeine Ladungen im Inneren), des Porinkanals (Kreusch und Schulz, 1994), und einesKaliumkanals (aus Pilzen, Abbildung 13, Doyle et al., 1998), eines Eisen transportie-renden Kanals (Ferguson et al., 1998) und eines mechanosensitiven Kanals (Chang etal., 1998).
Den großen Durchbruch lieferte die Darstellung eines K+-Kanals (Doyle et al., 1998).Der KscA aus Streptomyces lividans hat nur je zwei Helices in den vier Untereinheitenund dies vereinfachte die Interpretation der Röntgenstrahlbeugungsbilder. Abbildung 13

33
zeigt den Aufbau dieses Kanals. Die Struktur ermöglichte das Verständnis des Selekti-vitätsmechanismus. Das Selektivitätsfilter (12 AA lang) enthält eine hochkonservierteAminosäurensequenz. Diese Aminosäuren kleiden mit ihren Carbonylgruppen den Ka-nal derart aus, daß sie die Wasserhülle des Ions ersetzen. Da der Kanal aber im Gegen-satz zum Wasser starr ist, gelingt dieser Ersatz nur, wenn der Ionenradius genau stimmt.Das erklärt extrem hohe Selektivität bei Reaktionsraten von 107 bis 108 Ionen/s. Vordem Selektivitätsfilter liegt eine größere wassergefüllte Höhle. Zwei alpha-Helices ra-gen bis zur Höhle in den Kanal hinein und erhöhen mit ihren negativen Ladungen dieAufenthaltswahrscheinlichkeit vor dem Selektivitätsfilter. Eine weitere Besonderheit istdie hydrophobe Auskleidung des inneren breiteren Kanals, die eine Wechselwirkung desIons mit der Wand verhindert und damit die Durchtrittsgeschwindigkeit erhöht.
Die Auskleidung des Selektivitätsfilters geschieht durch die „P-loops“ der vier Un-tereinheiten. Sie liegen beim Shakertyp-Kanal zwischen den Helices S5 und S6 (Hegin-botham et al., 1994). Der P-loop ist nahezu identisch für alle Kaliumkanäle, aber ver-schieden für Kanäle, die andere Ionen durchlassen (Durell et al., 1998). Die Helix S4 hatdie Rolle eines Spannungsfühlers, der sich bei aktiviertem Schalten nach außen bewegt(Larsson et al., 1996, Yang et al., 1996). Die Spannungsabhängigkeit des Öffnens liegtan der Wirkung auf 13 Ladungen (das sind mehr als 3 Ladungen pro Untereinheit,Schoppa et al., 1992, Sigg et al., 1994, Aggarwal und MacKinnon, 1996, Seoh et al.,1996 ). Die Bewegung kann zum Öffnen und Schließen des Kanals führen. Ein weitererMechanismus des Kanalschaltens hängt mit den vier Untereinheiten am N-Terminus(dem Ende der Aminosäurekette, das in H3N endet) zusammen, die den Eingang zumKanal verschließen können („Ball and Chain“, Antz et al., 1997).
Mit Hilfe solch genauer Daten ist es nun denkbar, eine exakte Molekular-Dynamische (MD) Beschreibung zu finden. Allerdings ist das Ausrechnen bislang prak-tisch unmöglich: Um Aussagen über den Weg der Ionen machen zu können, müßte manmindestens zehn Durchgänge durch den Kanal berechnen. Bei einer Leitfähigkeit von 50pS und einer Spannung von 100 mV ergibt das einen Zeitraum von 0,3 µs. Beim Porinzum Beispiel müßte man dann 1020 Aminosäurereste, 300 Phosphatidylmoleküle,12992 Wassermoleküle und 27 Natriumatome betrachten. Die für 1 ps Realzeit benö-tigte Rechenzeit beträgt zwei Stunden, und das ergibt hochgerechnet etwa 70 Jahre fürdie 0,3 µs (Levitt, 1999).
Dies ist jedoch nicht das einzige Problem. Soll die Leitfähigkeit auch nur auf einenFaktor 7 genau berechnet werden, muß dazu die Wechselwirkungsenergie zwischen Ionund Kanal bis auf etwa 2kT genau bestimmt sein und das ist weniger als die Hälfte derEnergie einer einzigen Wasserstoffbindung.
Zur Modellierung sind daher Vereinfachungen nötig. Betrachtet man nicht alle Mole-küle sondern nur die an den Rändern, so ist von der Computerzeit eine Berechnung herbald denkbar. Simulationen über sehr kurze Zeiträume (ps) werden schon gemacht (Su-

34
enaga et al., 1998, Sansom et al., 1998) und sind nützlich, um Informationen über lokalepotentielle Energie und über Diffusionskoeffizienten von Ionen als Funktion der Positi-on im Kanal zu gewinnen.
Bei Brownian Dynamics (BD) werden die Wassermoleküle durch ein Kontinuum er-setzt, die Proteinstruktur bleibt fest. Für jedes Ion wird in diskreten Zeitschritten eineDifferentialgleichung gelöst. Dies ist die beste zur Zeit mögliche Beschreibung bei be-kannter Kanalstruktur.
In der nächstweitergehenden Näherung wird das Feld für jedes Ion durch ein mittle-res Feld ersetzt, welches mit der Poissongleichung und zufälligen thermischen Bewe-gungen berechnet wird. Dieses Modell wird als Poisson-Nernst-Planck-Modell (PNP)bezeichnet. Die meisten Berechnungen zum PNP Modell sind in einer Dimensiondurchgeführt worden, es gibt aber auch dreidimensionale Berechnungen (Kurnikova etal., 1999).
Da bei den meisten Kanälen die Struktur noch nicht bekannt ist, ist es nicht nötig,kompliziertere Modelle als das PNP zu benutzen, zumal sich viele Experimente damitfitten lassen (Nonner und Eisenberg, 1998, Nonner et al., 1998, Nonner et al., 1999).
Auch das PNP Modell läßt sich weiter vereinfachen: Nimmt man an, daß die Ionen inspezifischen Regionen im Kanal lokalisiert sind (nach Doyle et al., 1998, enthält dasSelektivitätsfilter zwei Ionen im Abstand von 7.5 AA), kann man die Kinetik durchSprünge zwischen diesen Bereichen und den Bereichen und der Außenlösung beschrei-ben. Diese Barrierenmodelle oder Reaktionsratenmodelle (RR) sind sinnvoll, wenn esdarum geht, den Kanal zu parametrisieren, und man weiß, daß starke Ion-Ion-Wechselwirkungen wichtig sind. Allerdings können die Ratenkonstanten nicht unbe-dingt wahren Energiebarrieren zugeordnet werden. Zur Beschreibung einer linearenStromspannungskurve, wie sie oft im Experiment gefunden wird, müssen häufig mehre-re Energiebarrieren eingeführt werden, die keine Beziehung zu wahren physikalischenEnergiebarrieren haben.
Experimente wie der anomale Molfraktionseffekt (AMFE, bei einer festen Ge-samtionenkonzentration durchläuft der Einzelkanalstrom mit der Variation des Ionen-verhältnisses ein Minimum, d.h. die reinen Lösungen haben einen größeren Strom alsdie Mischlösung) zeigen die Grenzen, aber auch die Vorteile der Modelle. So führte derAMFE dazu, daß in den meisten RR Modellen davon ausgegangen wird, daß mehrereIonen gleichzeitig im Kanal sind, die nicht aneinander vorbei können (Hille undSchwarz, 1978) und deren Wechselwirkung untereinander zu dem Minimum führt.Solch ein „single filing“ wurde nun auch experimentell von Doyle et al. (1998) gefun-den.
Im PNP Modell wird solch ein AMFE für Na+ und Ca2+ damit erklärt, daß im KanalElektroneutralität herrschen muß. Bei sehr geringer Ca2+-Konzentration wird diese Auf-gabe von Na+ übernommen, steigt die Ca2+-Konzentration, so verdrängt Calcium Natri-

35
um, da es als zweiwertiges Ion überlegen ist. Der Anstieg nach dem Minimum wird al-lerdings nicht erklärt.
Neuere Ergebnisse zeigen aber, daß gewisse Grundfragen vor dem Entwerfen vonModellen erst geklärt werden sollten. Für den K+-Kalium in Chara wurde gezeigt (Fa-rokhi et al., 1999), daß der AMFE ein Gating-Phänomen ist und kein Phänomen desPermeationsmechanismus. Die Verminderung der scheinbaren Einzelkanalleitfähigkeitliegt an einer früher nicht entdeckten Modulation des Gating-Verhaltens bei konstanterEinzelkanalleitfähigkeit.
4.2 Membranpotentiale
Von großem Interesse für die Funktion des Transportes an den Xylemkontaktzellen ist dieGröße des Membranpotentials. Die Spannung an der Membran setzt sich aus mehrerenKomponenten zusammen. Konzentrationsunterschiede zwischen Cytosol und externerLösung führen auf das Nernstpotential. Für Kalium ist dies:
i
o
]K[]K[ln
zFRTE =
mit F = Faraday-Konstante; [K]o,i = Kaliumkonzentration außerhalb und innerhalb derZelle; R = Gaskonstante; T = Raumtemperatur; z = Ladung des Ions, für Kalium: z = 1.
Für die Spannungsentstehung bei Lösungen aus mehreren Ionen gibt es noch keinefundierte Theorie. Aus Tradition wird die eigentlich nur für die Diffusion im Kontinuumgeltende Goldmann Gleichung (1943) verwendet, die phänomenologische Durchlässig-keiten (P) berücksichtigt:
∑∑∑∑
∑∑∑∑
++
=
++
=
onAimK
inAomK
onAimK
inAomK
]A[P]K[P]A[P]K[P
log*mV58
]A[P]K[P]A[P]K[P
lnzFRTE
m
m
m
m
Pumpen transportieren aktiv Ladung (z.B. H+, Na+, Ca2+) entgegen dem Konzentrati-onsgradienten aus der Zelle in die Zellwand. Dieser Strom positiver Ladung führt zu einerErhöhung der negativen Membranspannung (Cytosol gegen Lumen, Bad). Im allgemeinenist das Cytosol zwischen 100 und 250 mV negativer als seine Umgebung und zum Teilauch 30 mV negativer als die Vakuole (Sze, 1985).
Bei den kleinen Zellen höherer Pflanzen ist das Membranpotential aufgrund derunvermeidlichen Einstichverletzungen schwierig zu bestimmen: Amtmann et al. (1997)zeigen z.B. eine weitgefächerte Verteilung von -140 mV bis +30 mV mit zwei Peaks beiHaferwurzel-Protoplasten an, und auch Skerrett und Tyerman (1994) berichten von solcheiner Verteilung.

36
4.3 Transportwege ins Phloem und aus dem Xylem
Abbildung 14. Transportwege aus dem Apoplasten ins Phloem (Sieve Tube) nach Marschner(1995).
Mais ist ein apoplastischer Belader, d.h., die Assimilate werden außerhalb der Zellen bisin das Leitbündel transportiert und dann erst in den Symplasten (Phloem) aufgenommen(Abschnitt 2.4.1). Für diese Aufnahme aus dem Apoplasten ins Phloem spielen ver-schiedene Transporter eine Rolle. Abbildung 14 zeigt, daß die H+-ATPase Membran-spannung und pH-Gradient über der Membran aufbaut. Zucker und Aminosäuren wer-den über H+- Cotransporter, und K+ durch K+-Kanäle aus dem Apoplasten ins Phloemaufgenommen.
Auch für die Beladung der Zellen mit den im Xylem transportierten Nährstoffenspielen verschiedene Transporter eine Rolle. In der Plasmamembran gibt es ATPasen,die die pH-Gradienten (von Apoplast zu Cytosol) aufrechterhalten. Kationen (Cat+)werden wieder über Kanäle transportiert, Aminosäuren (As-) und Anionen (An-) überH+-Cotransporter. Ähnliche Transporter bewirken die Anreicherung von Substanzen inder Vakuole, die als Speicher- und Abfallkompartiment dient.

37
Abbildung 15. Beladung der an das Xylem angrenzenden Zellen nach Marschner (1995). Cat+
= Kationen, An- = Anionen, As- = Aminosäuren.
4.4 Lösungen
Im Rahmen des Apoplastenprojektes wurde angestrebt, die Daten aus den einzelnenBereichen so zu erheben, daß sie am Ende zusammenpassen. Für diese Arbeit bedeutetedies, neben der Festlegung auf die Maispflanze, die Lösungen möglichst physiologischzusammenzusetzen. Einen Anhaltspunkt dazu lieferte Tabelle 2 (Abschnitt 2.3), aber amAnfang der Arbeit lagen dazu kaum Daten vor. Zudem mußte ich die Lösungen sowählen, daß sich die Protoplasten darin „wohl fühlten“ (also eine lange Lebenserwar-tung hatten; besonders wichtig hierbei pH-Wert und Osmolarität), sich aber auch soge-nannte Gigaseals (gute Verbindung zwischen Meßpipette und Zelle, Abschnitt 4.5, Ab-bildung 17) bilden konnten. Dies führte insbesondere bei den Einzelkanalmessungendazu, sich mich mehr an der Patch-Literatur zu orientieren. Einen Überblick geben Ta-belle 6 für die Pipettenlösungen und Tabelle 7 für die Badlösungen. Die Kaliumkon-zentrationen (als KCl oder KGlutamat) für alle Pipettenlösungen bei Whole-Cell-Messungen liegen um 100 mM, der physiologischen Konzentration im Cytosol. Calcium(0 – 2 mM) wird mit EGTA oder EDTA gepuffert. MgCl2 liegt zwischen 1 und 2 mM.Der pH-Wert im Cytosol beträgt etwa 7.2. Eingestellt mit Mes und Tris oder HEPES

38
sind so groß auch die meisten (11 von 13) pH-Werte der Pipettenlösungen. Bei denBadlösungen sind die Werte für Kalium (1 – 100 mM) und Calcium (1 – 40 mM) weitergestreut. Calcium muß in ausreichender Konzentration im Bad vorhanden sein, um diefeste Verbindung zwischen Glas und Membran zu ermöglichen. Kaliumkonzentrationenkönnen auch im Apoplasten schwanken: In Tabelle 2 sind Ionenkonzentrationen vonXylemsaft und Apoplastenlösung von Maisblättern angegeben. Die beiden differierenum bis zu Faktor vier, obwohl das Xylem einen Teil des Apoplasten ausmacht. Kon-zentrationen über 50 mM K+ im Bad sind dennoch physiologisch nicht zu erwarten. Au-ßerhalb der Zellen ist der pH-Wert leicht sauer, daran angepaßt sind auch die Badlösun-gen (pH-Werte zwischen 5,1 und 6,0 bis auf eine Ausnahme). Sehr stark sowohl für dieBadlösungen als auch für die Pipettenlösungen variiert die Osmolarität (200 – 720mOsm). Die Osmolarität ist denn auch experimentell am schwierigsten einzustellen. Imallgemeinen liegt die Osmolarität der Pipettenlösung etwas über der der Badlösung (diessoll zur besseren Sealbildung beim Patchen, Abschnitt 4.5, führen).
Die verwendeten Lösungen finden sich in Tabelle 8. Alle Lösungen wurden gefiltertund in 1,5 ml Reaktionsgefäßen (Eppendorf, Hamburg) eingefroren, um eine längereHaltbarkeit zu erreichen. Direkt vor den Versuchen wurden sie aufgetaut.
Bei Einzelkanalmessungen wurden zumeist höher konzentrierte Lösungen verwendet,da die Ströme i.A. mit zunehmender Konzentration steigen. Da die gemessenen Zellennicht nur sehr kleine Einzelkanalströme hatten, sondern zudem noch schnell schalteten,ließen sich nur unter solch unphysiologischen Bedingungen Daten gewinnen.

39
Tabelle 6. Pipettenlösungen bei Pflanzen. Alle Angaben, soweit nicht anders vermerkt, inmM. G = Schließzellen, M = Mesophyllzellen, P = Protoplasten, S = Suspensionszellen, W =Wurzel.
KCl oderKglu (g)
CaCl2oder Ca2+
frei (f)MgCl2 HEPES
MgATPoderK2ATP(K) oderBTP-ATP(B)
EGTAoderBAPTA(B)
pHOsm mitSorbitoloderMannitol(M)
Zellenwhole-cell(wc) odersinglechannel (sc)
Autoren
100 g Kein 2 10 4 K 2 7,2 720 Mais W Pwc
Roberts undTester, 1995
100 Kein 2 10 4 K 2 7,22 ~720 RoggenW Pwc
White undLemtiri-Chlieh, 1995
100 g Kein 2 10 5 B 10 7,0 ~600 GetreideS
Ketchum undPoole, 1991
100 2 - - - 4 5,8 ~200 AsclepiastuberosaP, wc, sc
Schauf undWilson, 1987
100 0,2 f 2 4 - - 7,5 ~390 Eremos-phera
Linz undKöhler, 1993
100 0,1f 2 4 - 1 7,5 ~390 Tabak De Boer et al.,1994
100 0,1 2 5 - 1 B 7,2 450 M W Psc,wc
Vogelzangund Prins,1992
4 0,001 f 2 - - 1 7,2 210 M P, sc Elzenga et al.,1991
120 < 0,001f
2 - 2 - 7,2 530 M GerstenW Pwc
Wegner et al.,1995
100 1 1 - 2 ? 5,7 500 Gersten Swc
Amtmann etal., 1997
100 g Kein 2 20 4 2 7,2 520 Vicia fabaG wc
Ilan et al.,1995
100 150 nMf
1 ? 2 -? 7,5 600 Mais M Karley(email)

40
Tabelle 7. Badlösungen bei Pflanzen. Alle Angaben, soweit nicht anders vermerkt, in mM. G= Schließzellen, M = Mesophyllzellen, P = Protoplasten, S = Suspensionszellen, sc = Einzelka-nal, W = Wurzel, wc = Whole-Cell.
KCloderKglu (g)
CaCl2oderCa2+ frei(f)
Mg-Cl2
HE-PESoderMes(M)
TEAglu
NaClpH
Osm. mitSorbitoloder Man-nitol (M)
Zellen whole-cell (wc)oder singlechannel (sc)
Autoren
10 10 - 10 M - - 6,0 700 Mais W Pwc
Roberts undTester, 1995
100 10 - 10 M - - 5,5 636 Roggen WPwc
White undLemtiri-Chlieh,1995
1 g 1 2 - 9 - 6,25 ~580 M Getreide S Ketchum, andPoole, 1991
100 g 2 - - - 100 5,8 ? Asclepiastuberosa P,wc, sc
Schauf undWilson, 1987
100 40 2 4 M - - 5,1 ~280 Eremos-phera
Linz und Köh-ler, 1993
10 –100
1 – 2 1 - 2 - - - 5,1 –5,7
? M Tabak De Boer et al.,1994
10 2 2 5 M - - 6,0 400 M W P sc,wc Vogelzang undPrins, 1992
10citrate
5 2 - - - 5,5 210 M P, sc Elzenga et al.,1991
30 1 2 10 - - 5,8 500 M Gersten WP wc
Wegner et al.,1994
100 1 1 - - - 7,5 ? Gersten Swc
Amtmann et al.,1997
10 –50 g
1 2 10 M - - 5,5 480 M Vicia fabaG wc
Ilan et al., 1995
- 1 - ? - 100 5,7 600 Mais M Karley (email)

41
Tabelle 8. Verwendete Lösungen (alle Angaben in mM). Um Verwechslungen zu vermeiden,wurden die Eppendorf Caps mit den gefilterten Lösungen vor dem Einfrieren farblich makiert.Diese Farbbezeichnungen werden im Folgenden als Abkürzungen für die Lösungen verwendet.sc in der linken Spalte steht für die Lösungen, die bei Einzelkanalmessungen (Single-Channel)verwendet wurden, wc für Whole-Cell.
BadlösungenFarbe KCl MgCl2 CaCl2 Sorb. sonstiges pH Osmgelb sc 2 2 5 370 - 5,8 460
orange sc 50 2 5 280 - 5,8 280rot wc 10 2 5 350 - 5,8 450
+ (rot) wc 10 2 5 350 - 6,15 450rot-braun wc 10 2 5 350 - 6,5 450
braun wc 10 2 5 350 - 7,2 450schwarz wc 10 2 5 350 - 7,9 450R (braun) wc - 2 5 350 10 RbCl 7,2 450+ (braun) wc 10 2 5 350 - 7,2 550N (grün) sc - 2 5 290 100 NaCl 7,2 500K (grün) sc 100 2 5 290 - 7,2 5002 (grün) sc 200 2 5 40 - 7,2 450
O (braun) wc - 2 5 350 10 KNO3 7,2 4501 (rot) wc 1 2 5 400 - 6,15 450
1 (orange) wc 0,5K2SO4
2MgSO4
5CaSO4
400 - 6,15 450
PippettenlösungenFarbe KCl MgCl2 CaCl2 Sorb. sonstiges pH Osmgrün sc,
wc100 2 5,6 150 10 EG-
TA7,2 470
blau wc 100 2 5,6 150 10 EG-TA2
MgATP
7,2 470
+ (blau) wc 100 2 5,6 150 10 EG-TA2
MgATP
7,2 470
2 (blau) wc 100 2 5,6 50 10 EG-TA + 2MgATP
7,2 470
ohne (En-zym-lösung)
sc,wc
- - 5 400 1,5% Cellula-se, 0,5% Ma-
cerozyme,2,0% Pectina-
se
6,5

42
4.5 Die Patch-Clamp-Technik
Bei der klassischen Mikroelektrodentechnik von Umrath (1932) wurde, um elektrischeEigenschaften von Membranen zu messen, mit einer Elektrode die Zelle oder Vakuoleangestochen, während sich eine Referenzelektrode in der Badlösung befand (Einstichelek-trodentechnik). Der Nachteil dieser klassischen Technik liegt darin, daß nur relativ großeObjekte damit angestochen werden können.
Neher und Sakman entwickelten diese Technik 1976 entscheidend weiter. Sie legten andie Pipette einen Unterdruck an, sobald die Pipette die Membran berührte. Auf diese Artwird die Verbindung zwischen Glas und Membran sehr hochohmig und der Widerstandgeht in den Gigaohmbereich, es bildet sich das sogenannte Gigaseal (Abbildung 17 A –D). Diese Verbindung ist so fest, daß es nun möglich ist, Ströme durch einzelne Kanäle inder Membran zu beobachten, insbesondere deren Öffnen und Schließen, dasEinzelkanalschalten.
Abbildung 16. Die Patch-Clamp Technik. In einer Petrischale liegt das Ausgangsobjekt für alleverschiedenen Patchtechniken in einer Salzlösung. Eine Patchelektrode wird auf die Membrangesetzt, so daß sich ein sehr hoher Widerstand bildet, während die andere (Erd-) Elektrode dasBad auf Masse- oder Sollwertpotential legt. Durch einen Regelkreis zwischen Patch- und Erde-lektrode können Spannungen angelegt werde, und dann die Ströme durch das „gepatchte“ Mem-branstück gemessen werden.
Verschiedene Gigaseal-Techniken sind möglich. Wird, wie in Abbildung 17 A, nur diePipette auf die Membran gesetzt, spricht man von der On-Cell Konfiguration. Ihr Vorteilliegt darin, daß innerhalb der Zelle (oder des Vesikels) physiologische Bedingungenvorliegen. Damit ist aber auch die Zusammensetzung der Lösung unbekannt und ebenso

43
das Potential. Zudem kann die Eintauchtiefe nicht nachträglich verringert werden,wodurch es zu einem relativ hohen Rauschen (Keunecke, 1995) kommt.
Bewegt man die Pipette ruckartig von der Zelle weg, so haftet nur noch das kleineZellstück an der Pipette, man bekommt einen Excised-Patch (Abbildung 17 C) mit derInnenseite der Zelle zur Badlösung hin (Inside-Out). Nun liegt auf beiden Seiten derMembran eine definierte Lösung vor, und auch das Potential läßt sich direkt ermitteln.
Legt man in der On-Cell-Konfiguration einen Unterdruck (mit dem Mund) oder einenSpannungspuls an, so reißt das Membranstück im Pipettenmund, und man erreicht dieWhole-Cell-Konfiguration. Die Pipette ist mit dem Zellinneren verbunden. Der Austauschder Lösungen von Zelle und Pipette geht recht schnell, und da die Pipette zumeist ein sehrviel größeres Lösungsvolumen beinhaltet als die Zelle, liegt nach kurzer Zeit diePipettenlösung in der Zelle vor. Hat man nicht alle Substanzen in die Pipettenlösunggemischt, die der Kanal zur Aktivierung benötigt, ist ein Auswascheffekt beobachtbar:Anfangs ist eine Aktivierung der Kanäle zu beobachten, aber nach einigen Minutenverschwindet diese Antwort zunehmend (Abschnitt 8.3).
Abbildung 17. Die vier Gigaseal-Techniken werden danach unterschieden, ob der Patch amPipettenmund zerstört wird oder nicht, und ob die Zelle intakt bleibt oder ein isoliertes Zellstückherausgerissen wird. Es ergeben sich damit die vier Konfigurationen (Hamill et al., 1981): On-Cell oder Cell-Attached (A), ganze Zelle (B, Whole-Cell), Excised-Patch mit Innenseite außen (C,Inside-Out) oder Excised-Patch mit Außenseite außen (D, Outside-Out).
Von der Whole-Cell-Konfiguration kann man durch ruckartiges Ziehen wieder in ei-ne Excised-Patch-Konfiguration (Abbildung 17 D) gelangen. Hierbei ist nun, im Gegen-satz zu Abbildung 17 C, die Außenseite der Zelle dem Bad zugewandt (Outside-Out-Konfiguration).

44
Abbildung 18 zeigt den Vorgang einer Sealbildung bei der Whole-Cell-Konfiguration. Sie läuft unter Beobachtung der Stromantwort ab, die durch eine Recht-eckspannung (Abbildung 18 A) erzeugt wird. Ist die Pipette ins Bad eingetaucht, fließtein sehr hoher Strom. Er nimmt ab bei Annäherung an die Zelle. Hat sich ein Seal gebil-det (Abbildung 18 C), sieht man einen sehr kleinen stationären Strom, der sich ausMembranstrom und Leckstrom durch das Seal zusammensetzt. Die Peaks sind Umlade-ströme für die Kapazität, die jetzt hauptsächlich von der Pipette bestimmt werden. Nachdem Whole-Cell-Durchbruch (Abbildung 18 D) wird der stationäre Strom sehr klein,weil jetzt die Membranfläche der ganzen Zelle die Leitfähigkeit bestimmt. Die Peaksdienen der Umladung der Zellmembrankapazität der Gesamtzelle. Mit Hilfe eines Kom-pensationsmechanismus (negative Kapazität; Sakmann und Neher, 1985) kann jetzt derkapazitive Sprung kompensiert werden. Die Peaks sollen dann verschwinden (Abbil-dung 18 E). Jetzt können die Messungen beginnen. Die Kapazität ist dabei ein Maß fürdie Zellgröße (i.A. haben Membranen eine Kapazität von 1 µF/cm2), die Kenntnis derZellgröße ist Voraussetzung für eine Eichung auf Strom pro Fläche.

45
A
B
C
D
E
Abbildung 18. A. Eingangssignal (Rechteckspannung) B. Pipette im Bad, bei Annäherung andie Zelle nimmt der Widerstand leicht zu. C. On-Cell-Konfiguration. Bei hoher Verstärkungläßt sich aus dem Offset der Sealwiderstand berechnen. D. Whole-Cell-Durchbruch. E. Nachder Kapazitätskompensation (aus: Numberger und Draguhn, 1996).

46
5 Isolationsverfahren
Der erste schwierige Schritt zum Studium des Transportes zwischen Apoplast und Sym-plast ist die Isolierung von Xylemkontaktzellen (Bündelscheidenzellen), damit sie fürPatch-Clamp-Untersuchungen zugänglich werden. Dies können Xylemparenchymzellenoder anliegende Bündelscheidenzellen sein. Nach den Untersuchungen mit LAMMA inKapitel 3 sind die Bündelscheidenzellen die Zellen, die Ionen aus dem Xylem aufneh-men.
Für die Zellen im Blatt gab es bisher kein Isolationsverfahren, doch erfolgreiche Iso-lierungen anderer pflanzlicher Zellen im Transportweg Wurzelblatt ergaben Orientie-rungshilfen bei der Entwicklung eines Verfahrens für die hier interessierenden Blattzel-len.
Die meisten Patch-Clamp-Messungen an Blättern wurden bislang an Schließzellendurchgeführt (z.B. Assmann et al., 1985), da diese leicht zu isolieren sind: Die Epider-mis vieler Pflanzen läßt sich abstreifen, besonders leicht z.B. bei Vicia faba. Diese gibtman dann in eine geeignete Enzymlösung. Die Enzyme bauen zuerst die Epidermiszel-len ab, bevor die Zellwände der Schließzellen so weit verdaut werden, daß dann dieSchließzellprotoplasten aus ihnen schlüpfen können (meist mit Hilfe von Plasmolyse,d.h. osmotischem Schock).
Für Gerstenwurzeln ist von Wegner (Wegner und Raschke, 1994) ein Isolationsver-fahren für Xylemparenchymzellen entwickelt worden. Hier werden die Hauptgersten-wurzeln (nicht die Seitenwurzeln) 1 cm weit abgeschnitten und dann die folgenden 2 cmabgestrippt und in 1 mM CaCl2 auf Eis gesammelt. Mit Haushaltspapier getrocknet,wurden die Stränge klein geschnitten und in Enzymlösung gegeben (0.02% PectolyaseY 23 (Seishin Pharmaceutical, Tokyo, Jaspan), 2% Cellulase Onozuka R 10 (YakultHonsha), 1 mM CaCl2, 10mM Na-Ascorbat, 2% BSA, 500 mOsm (Mannitol), pH 5,5mit H2SO4) und dort bei 22 – 25°C 2 Stunden im Wasserbad geschüttelt. Anschließendwurden die Protoplasten mit 50 ml einer 1 mM CaCl2 + 500 mM Mannitol Lösungdurch ein 200 µm Netz (Züricher Beuteltuchfabrik, Rütlikon, Schweiz) gespült, um dieProtoplasten von den Zellwandtrümmern zu trennen. In der Zentrifuge bei 100 g wurdendie Protoplasten 10 min pelletriert (da sie schwerer als die Lösung sind, sinken sie zuBoden), durch ein 80 µm und 23 µm Netz gespült und erneut bei 100 g pelletriert. Daßdie Xylemkontaktzellen zu einem ganz bestimmten Zeitpunkt abgebaut werden (unddadurch von anderen Zellen unterschieden werden können), wurde von Wegner undRaschke (1994) an Hand von Schnitten und Färbeversuchen nachgewiesen.

47
5.1 Isolation von Xylemkontaktzellen bei Bohne
Ein Verfahren zur Isolation von Xylemkontaktzellen von Bohnenblättern (Vicia faba)entwickelte Sutter (1996). Patchbare Xylemkontaktzellen wurden auf folgende Weise ge-wonnen: An ein gerade voll ausgewachsenes Blatt wurde in einer mit Leitungswasser ge-füllten 10 ml Spritze ein Unterdruck angelegt, der Luft durch die Stomata herausperlenließ. Bei Normaldruck füllte sich dann der vorher gasgefüllte apoplastische Raum. DieseProzedur wurde dreimal wiederholt. Wider Erwarten lieferte eine Infiltration mit der En-zymlösung keine besseren Ergebnisse (obwohl dann die Lösung schneller im Blatt ist undnicht erst hinein diffundieren muß), hingegen war sie wesentlich teurer (höherer Ver-brauch) und kam daher nicht in Betracht.
Danach wurde mit einer Pinzette die untere Epidermis vollständig abgezogen und einetwa 3 cm2 großer Bereich aus dem Blatt geschnitten. Er wurde mit der Unterseite nachoben in eine 3 cm-Petrischale mit der folgenden Enzymlösung gelegt: 1.5% Zellulase,0.5% Macerozyme (Yakult Honsha), 2.0% Pectinase (Sigma), 0.5% BSA (Sigma), 5 mMCaCl2, 0.5 M Sorbitol, pH 6.5 (gepuffert mit Tris / Mes). Nach drei Stunden wurde dieobere Epidermis mit einer Pinzette entfernt, was mitunter etwas mühsam war. Anschlie-ßend verweilten die Blattstücke weitere 30 Minuten in der Enzymlösung, bevor sie in dieBadlösung kamen. Die Mesophyllzellen wurden ausgewaschen, indem sie mit einer Pa-steurpipette vorsichtig eingesogen und wieder in die Badlösung gelassen wurden. Hierbeigingen zwar auch einige Xylemkontaktzellen verloren, es ist jedoch eine sehr geringeAusbeute für die Patch-Clamp-Messungen ausreichend. Die Xylemstränge mit anhängen-den Xylemkontaktzellen wurden dann in die Meßlösung gegeben.
5.2 Isolation von Bündelscheidenzellen bei Mais
Die Notwendigkeit, an Mais zu arbeiten, ergab sich aus einem Standardisierungsversuchin der Teilgruppe des Apoplastenprojektes. Man einigte sich auf die Maispflanze dainsbesondere die Aphidentechnik (Abschnitt 12.3, Anhang), die von der ArbeitsgruppeHeldt/Lohaus angewendet wird, an Vicia faba nicht, an Mais aber sehr gut funktioniert.
Für die Präparation von Xylemkontaktzellen von Mais waren einige Veränderungengegenüber der oben beschriebenen Isolation bei Vicia faba nötig. Insgesamt ist das Blattvon Mais stabiler, dies liegt vor allem an der Suberinschicht um die Bündelscheide(Eastman et al., 1986, Evert et al., 1977, 1996), aber auch an der Lignifizierung, die mitdem Blattalter zunimmt (Lopez et al., 1993).

48
A B
Abbildung 19. Xylemstränge von Mais mit parenchymatischen Bündelscheidenzellen bereitzum Patchen. Durch die Isolationsprozedur sind alle Mesophyllzellen ausgewaschen. A. VierLeitbündelstränge mit angrenzenden Bündelscheidenzellen. B. Vergrößerter Ausschnitt. DieLeitbündel sind untereinander über transversale Leitbündel verbunden. Diese Leitbündel beste-hen nur aus einem Xylem- und einem Phloemelement.
Ausgehend von der Bohnenpräparation (Abschnitt 5.1) wurde die Isolationsmethodefür die parenchymatischen Bündelscheidenzellen modifiziert.
Es wurden mittlere Blattabschnitte (Blattlänge etwa 20-25 cm) des vorletzten Blattes(meist das 4. von 5) von 4 Wochen alten Maispflanzen verwendet. Anders als bei Boh-nenblättern, reicht ein Unterdruck nicht aus, um die Luft aus den Interzellularen durchdie Spaltöffnungen aus dem Blatt perlen zu lassen. Bei Mais ist das Verhältnis vonapoplastischem Gas- zu Wasserraum viel kleiner als bei Vicia faba (Lohaus et al.,1999). Mit einem in einer 50 ml Spritze angelegter Überdruck gelang es, die Luft ausden Interzellularen zu pressen. Bei langsamem Nachlassen des Überdruckes lief dasBlatt dann mit Wasser voll. Durch dreimaliges Wiederholen dieses Vorganges ließ sichdie Luft vollständig aus dem Blattgewebe entfernen. Für die Präparation wurde, wie beiKeunecke et al. (1997) beschrieben, nun die untere Epidermis abgezogen. Nur ein be-stimmtes Blattalter ist für die Präparation geeignet (optimal sind drei bis vier Wochenalte Pflanzen, je nach Jahreszeit).
Danach wurde das Blattstück in Längsrichtung angeritzt. Nach 6 Stunden in Enzym-lösung (1.5% Zellulase, 0.5% Macerozyme (Yakult Honsha), 2.0% Pectinase (SigmaChemie), 0.5% BSA (Sigma Chemie), 5 mM CaCl2, 0.5 M Sorbitol, pH 6.5, gepuffertmit Tris/Mes) wurden die Fasern mit Präpariernadeln auseinandergezogen, um Einzel-stränge zu separieren. Nach einer weiteren Stunde in der Enzymlösung wurde das Blatthochosmolarer Lösung (KCl + Sorbitol, 700 mOsm) ausgesetzt. Es verblieben die Xy-lemkontaktzellen an den Xylemelementen, das Mesophyll war vollständig enzymatischabgebaut (Abbildung 19 A). Auch andere Verunreinigungen traten praktisch nicht auf

49
(Abbildung 19 A und B). Die Zellwand war nun so locker, daß Teile des Protoplastenvorquellen oder daß die Zellwand von der Patchelektrode zur Seite geschoben werdenkonnte.
Übrig blieben an größeren Strukturen nur noch Xylemnetze mit anhängenden Xy-lemkontaktzellen. Der Nachteil hierbei war, daß sich, zum einen durch die lange Inku-bation in der Enzymlösung, zum anderen durch die hochosmolare Lösung, anschließendeine starke Plasmolyse der verbleibenden Zellen ergab (Abbildung 20).
Abbildung 20. Plasmolysierte Zellen. Wenn sich Zellen zu lange in hochosmolarer Lösungbefinden, kommt es zur Verkleinerung des Protoplasten (Plasmolyse), die zum Zelltod führenkann.
In einer anschließenden Modifikation des Verfahrens konnte die Inkubationszeitdurch Veränderung der Inkubationstemperatur entscheidend verkürzt werden. Die be-nutzten Enzyme arbeiten am besten bei etwa 40°C, es wurde daher die Inkubation für 2hbei 38°C in einem Wasserbad durchgeführt. Die verwendete Temperaturregelung (IKA-Combimag RCT, Eydam, Kiel) war aber leider zu schlecht, häufig stieg die Temperaturauf über 40°C. Dabei ist eine Veränderung der Membran zu befürchten. Die Messungenergaben zwar keine Unterschiede (Einzelkanalströme veränderten sich nicht im Ver-gleich zu vorigen Ergebnissen), aber es wurde doch bevorzugt, zu niedrigeren, unbe-denklichen Temperaturen überzugehen. Eine konstante Temperatur, die nicht über 37°Canstieg, und zugleich für eine gute Durchmischung von Enzymlösung und Blattstücksorgte, wurde erreicht, indem die Probe in der Hosentasche aufbewahrt wurde. Zusätz-lich wurde keine hochosmolare Lösung mehr verwendet, da diese die Zellen belastet.Insgesamt konnte die Inkubationszeit so von ehemals 7 Stunden auf 135 min + 15 min

50
(bei Raumtemperatur) reduziert und die Plasmolyse verhindert werden (Verwendungvon Lösungen mit nur 550 mOsm).
In diesem Zustand war die verbleibende Zellwand so aufgelockert, daß eine Elektro-de hindurch geführt werden konnte und sealte. Die Patch-Clamp-Untersuchungen wur-den dann nicht, wie eigentlich üblich, an isolierten Protoplasten, sondern an noch beste-henden Gewebeverbänden von Xylemelementen und anhängenden Kontaktzellen vor-genommen. Allerdings gelangen auf diese Weise nur Excised-Patch-Messungen.
Diese Art der Präparation, bei der die zu untersuchenden Bündelscheidenzellen nocham Xylem anhaften, hatte einen weiteren Vorteil: Eine Kennzeichnung der Zellen durchAnfärbung, die sich zudem in Vorversuchen als unmöglich erwies, war überflüssig.
5.3 Whole-Cell-Präparation
Eine verbreitete Methode, Whole-Cell-Präparationen (Abbildung 17) herzustellen, be-ruht darauf, daß die Membran im Patch zerstört wird und damit das Zellinnere mit demPipetteninneren elektrisch verbunden wird. Diese Methode erschien anfangs als beson-ders geeignet, da mit obiger Präparationstechnik ein Seal an der Zelle im Zellwandko-kon hergestellt werden konnte.
Es traten aber folgende, für die Xylemkontaktzellen spezifische Probleme auf: Fürdas Durchdringen des lignifizierten Zellwandrestes mußte die Pipette einen kleinenDurchmesser (Elektrodenwiderstand 10 bis 20 MΩ) haben, um zur Sealbildung zurPlasmamembran vordringen. Es gelang bei diesen Pipetten zwar, durch Anlegen einesSpannungspulses, einen Durchbruch zu erzeugen, doch die so entstandene Whole-Cell-Präparation ist nur von sehr kurzer Dauer. Die Durchbruchstelle schloß sich schnellwieder.
Für einen dauerhaften Durchbruch waren größere Pipettenöffnungen erforderlich,d.h. Widerstände um 2,5 MΩ (Sakman und Neher, 1995). Diese Pipetten sealten abernicht bei dem vorliegenden Mais-Präparat. Offenbar vermochten nur die dünne Pipet-tenspitze beim Vordringen zum Plasmalemma lignifizierte Teile zur Seite zu schieben.Bei den großen Pipetten blieben wahrscheinlich einige „Balken“ vor dem Pipettenmundliegen und verhinderten den Kontakt mit dem Plasmalemma.
Ein Erfolg ergab sich, als die Präparationstechnik in Zusammenarbeit mit BarbaraKöhler (AG Raschke, Göttingen, jetzt Wye, Großbritannien) folgendermaßen verändertwurde: Da die im Abschnitt 5.2 beschriebene Präparation Xylemkontaktzellen ohneVerunreinigungen durch Mesophyllzellen lieferte, konnten die Protoplasten durch Spü-len mit der Badlösung (s.u.) gewonnen werden.
Im einzelnen wurde die Isolation jetzt folgendermaßen durchgeführt: Nach 2 StundenInkubation bei 33°C wurden die Blattstücke an Leitbündeln mit Präpariernadeln ausein-

51
andergezogen und noch etwa 15 Minuten bei Zimmertemperatur stehen gelassen. An-schließend wurden sie erst mit einer Lösung (alle Angaben in mM) aus 10 KCl, 2MgCl2, 5 CaCl2, pH 7.2 (Tris/Mes), 550 mOsm (Sorbitol) gespült und durch ein Netz(100 µm) mit der gleichen Lösung, aber geringerer Osmolarität (450 mOsm) gespült.Die Zellen waren so vollständig von Zellwandresten befreit.
Diese Protoplasten hatten folgende Vorteile:(i) Eine Färbung der Zellen durch Farbstoffe zur Identifikation war unnötig.(ii) Die Protoplasten ermöglichten Whole-Cell-Messungen.(iii) Der Sealprozeß lief schneller ab (meist unter einer Minute) und funktioniert auchmit Hartglas, welches rauschärmer als das bisher verwendete Weichglas war.
5.4 Isolation von Mesophyllzellen
Da die Bündelscheide von einer kompletten Suberinschicht umgeben ist, kann man da-von ausgehen, daß der apoplastische Raum in zwei Kompartimente unterteilt ist: vorund hinter der Bündelscheide. Diese Hypothese wird von zahlreichen Autoren unter-stützt (Evert et al., 1977, 1985, 1996, Hattersley and Browing, 1981, Botha et al. 1982,Canny, 1995).
Aufgrund der Verschiedenartigkeit des xylem- und mesophyllseitigen Apoplasten istanzunehmen, daß die angrenzenden Zellen auch verschiedene Transportaufgaben haben(dies bestätigt sich bei den unter Abschnitt 8.7 und 9.2 berichteten pH-Messungen). Ausdiesen Überlegungen folgte der Bedarf an Vergleichsmessungen an Mesophyllzellen.Somit mußte ein Verfahren zur Isolation der Mesophyllzellen entwickelt werden.
Für Mesophyllzellen wurde die Isolationsmethode für Bündelscheidenprotoplastenfolgendermaßen verändert: Die mittleren Blattabschnitte von vier Wochen alten Pflan-zen wurden nicht für 120 min, sondern nur für 70 Minuten in Enzymlösung inkubiert,die auch für die Isolierung der Bündelscheidenzellen benutzt wurde (Abschnitt 5.2).Anschließend wurden die Blattstücke sofort in die hochosmolare Lösung (wie bei denBS) gegeben und nach 10 Minuten in die Lösung gleicher Zusammensetzung aber nie-derer Osmolarität überführt. Dort wurden die Leitbündelstränge mit Präpariernadelnauseinandergezogen.

52
Die Zellwand war im Gegensatz zu Bündelscheidenzellen viel stärker zerstört wor-den, da Mesophyllzellen keine Suberinlamelle besitzen, so daß die Mesophyllzellen sichdazwischen herauslösten. Die verbleibenden Xylemstränge mit anhängenden Bündel-scheidenzellen dagegen widerstanden noch der Zersetzung und konnten mit einer Pin-zette samt anhängenden Mesophyllzellwandresten entfernt werden.
Nach 3 bis 5 Minuten setzen sich die Mesophyllzellen auf dem Glasboden der Patch-Meßkammer ab. Die verbliebenen darüber schwimmenden Trümmer konnten ausgespültwerden, ohne daß die festgesetzten Protoplasten mitgerissen wurden.
Auch dieses Verfahren erwies sich als sehr schonend, denn eine Untersuchung derGrößenverteilung der Mesophyllzellen in situ und der Protoplasten (wie in Abbildung23 für die Bündelscheidenzellen) zeigte, daß diese nicht zerfallen (Mittelwert bei 14.8 ±5.6 pl).
5.5 Vitalitätstests
A
B
Abbildung 21. Veranschaulichung der Undurchlässigkeit der Suberinlamelle für Neutralrot.Nicht isolierte Xylemkontaktzellen sprechen im Gegensatz zu den Mesophyllzellen nicht aufeine Behandlung mit Neutralrot an (A). Erst als Protoplasten färben sie ihre Vakuole rot (B).

53
Die Besonderheit des Präparates wurde deutlich, als mit Neutralrot Vitalitätstests durch-geführt werden sollten. In der Präparation der Abbildung 19, wie sie zu Einzelkanalmes-sungen verwendet wurde, ließen sie sich nicht anfärben. Normalerweise nimmt man dieNicht-Färbbarkeit als Anzeichen, daß die Zelle tot ist: Die Rotfärbung geschieht im sau-ren Milieu der Vakuolen (Anhang, Abschnitt 12.2). In einer toten Zelle tritt sie nichtauf, weil die H+-Pumpen am Tonoplasten (ähnlich Abbildung 12) die Vakuole nichtmehr ansäuern können. Doch dies traf hier nicht zu. Eine Anfärbung von Präparaten, beidenen nur die Zellwand abgezogen wurde, ergab eine sehr schöne Anfärbung der Meso-phyllzellen (Abbildung 21), aber nicht der Bündelscheidenzellen. Die Suberinschichtum die Bündelscheide verhindert offensichtlich den Zutritt des Farbstoffes. Erst nachder vollständigen Isolation (die tote Zellen ja nicht zum Leben erwecken kann) zeigtenBündelscheidenzellen eine kräftige Rotfärbung ihrer Vakuolen.
5.6 Größen von Bündelscheidenzellen und -protoplasten
Bei dem Verfahren zur Isolation von Xylemparenchymzellen aus Gerstenwurzeln zer-fielen die Protoplasten (Wegner und Raschke, 1994) in durchschnittlich 6 Protoplasten.Ein Zeichen, daß die in Abschnitt 5.3 und 5.4 vorgestellte Isolationsmethode sehr scho-nend war, besteht darin, daß hierbei ein solcher Zerfall nicht auftrat.
Die Anzahl an Bündelscheidenzellen von Leitbündeln zweiter und dritter Ordnungund deren Zellgrößen wurden bestimmt (Abbildung 22). Das Auszählen unter dem Mi-kroskop ergab, daß Leitbündel zweiter Ordnung meist sechs etwa gleich große Bündel-scheidenzellen besitzen (Abbildung 22 D), die in Kontakt zum Xylem sind. Bei Leit-bündeln dritter Ordnung findet man 4 bis 9 Bündelscheidenzellen (Abbildung 22 B).Betrachtet man bei ihnen die Leitbündel mit 5 Bündelscheidenzellen (diese sind am häu-figsten zu finden, Abbildung 22 B), ergibt sich eine Verteilung mit zwei ausgeprägtenMaxima (Abbildung 22 C). Das oder die Xylemelemente (im Mittel sind es 1,4 Xyle-melemente pro Leitbündel dritter Ordnung) liegen an zumindest einer von diesen großenZellen.
Es ist nicht möglich gewesen, die am Xylem liegenden Bündelscheidenzellen zumarkieren. Daher kann bei Patch-Clamp-Untersuchungen nur die Größenverteilung allerBündelscheidenzellen gemittelt werden, und nicht getrennt nach BS mit und ohne Xy-lemkontakt oder nach Leitbündeln mit 5 Bündelscheidenzellen.

54
A
Abbildung 22. Größenverteilung aus visueller Bestimmung. In den Leitbündeln zweiter unddritter Ordnung wurden die Zellen gemäß A gezählt. B. Anzahl der Bündelscheidenzellen inLeitbündeln dritter Ordnung. C. Größenverteilung der Bündelscheidenzellen von Leitbündelndritter Ordnung, die genau 5 Bündelscheidenzellen besaßen und D. die Größenverteilung beider zweiter Ordnung. F = Fläche der Zellen in Schnittebene.
0 2 4 6 8 100
10
20
30
40
B
Mittelwert:
5.32 +/- 0.86
Anzahl der Bündelscheidenzellen
Leitbündel 3. Ordnung
N
1 2 3 4 50
200
400
600
800C
LB 3
Bündelscheidenzelle
F (µ
m2 )
1 2 3 4 5 60
200
400
600
800D
LB 2
Bündelscheidenzelle
F (µ
m2 )

55
Der Mittelwert des Volumens der Bündelscheidenzellen wurde vor der Isolationspro-zedur bestimmt und betrug (21,1±1,7) pl. Die Länge wurde dabei aus Ansichten wieAbbildung 19 B bestimmt, die Breite und Höhe nach Schnitten (Abbildung 22 A). DieProtoplasten (30,8 ±3,2 µm Durchmesser) hatten ein Volumen von 15,3±5,4 pl. Dies istetwas geringer als das Volumen der (nicht isolierten) Zellen, da besonders große Pro-toplasten nicht zum Patchen geeignet sind, und nur diese gemessen wurden. Protopla-sten und Bündelscheidenzellen zeigen die gleiche Verteilungsfunktion. Zum Vergleichist auch die Größenverteilung der zum Patchen verwendeten Protoplasten in Abbildung23 mit eingezeichnet.
Abbildung 23. Vergleich der Verteilungsfunktionen der Volumen von Bündelscheidenzellen(n = 448), Bündelscheidenprotoplasten (n = 954) und der Größe der gepatchten Protoplasten (n= 211). Die sehr gute Übereinstimmung der drei Verteilungen zeigt insbesondere auch die Gra-phik oben rechts, in der die Verteilungen auf 1 normiert sind.
0 20 40 600
20
40
60
Bündelscheidenprotoplasten Bündelscheidenzellen gepatchte Protoplasten
n
V (pl)
0 20 40 600,00
0,04
0,08
n (%
)
V (pl)

56
5.7 Sealbildung
An den mit Polylysine fixierten Bohnen-Protoplasten ließ sich die Pipette relativ festandrücken, so daß schon so ein Widerstand von 20 bis 30 MΩ erreicht werden konnte.Während nach der Erfahrung mit anderen Objekten der Sealwiderstand innerhalb vonwenigstens 30 Sekunden einen Wert von etwa 1 GW erreichen mußte, bedurfte es jetzteines längeren Spiels von Unterdruckbehandlungen, bis nach ca. 30 Minuten ein Seal-widerstand im Bereich um 2 GW auftrat.
Bei den Maisbündelscheidenzellen hingegen trat das Seal bei den Einzelkanalmessu-gen schneller auf. Die Maisprotoplasten hafteten bereits ohne Fixierung am Glasbodenund daher bildeten sich auch leichter Gigaseals zwischen der Protoplastenmembran undder Pipette aus Glas aus. Gigaseals bildeten sich häufig innerhalb von weniger 30 Se-kunden.
Ein gutes Seal ließ sich bei Einzelkanalmessungen nur mit Pipetten aus Weichglaserhalten, doch erst als eine innere Beschichtung mit Sigmacote den Austritt toxischerIonen verhinderte, konnte Kanalaktivität zuverlässig beobachtet werden.

57
6 Experimenteller Aufbau der Patch-Clamp-Anlage
6.1 Mechanischer Aufbau mit Mikroskop
Abbildung 24. Patch-Clamp-Aufbau zur Aufnahme von Ionenkanaldaten. Im Faraday-Käfig istnur Gleichstrom erlaubt, daher befinden sich alle Stromversorgungen außerhalb des Käfigs. In derTüröffnung steht ein mobiles Klimagerät (Pinguino, DeLonghi, Treviso, Italien), um dieRaumtemperatur zwischen 19°C und 20°C zu halten. Alle Netzgeräte wurden aus den Gerätenausgebaut und befinden sich außerhalb der Meßkammer, damit kein Netzbrummen eingefangenwird.
Der Versuchsaufbau der Patch-Clamp-Anlage, (Abbildung 24) steht in einem Faraday-Käfig, da die Messung mit hochohmigen Mikroelektroden sehr empfindlich gegenStörstrahlung ist. Alle Geräte innerhalb des Käfigs werden daher mit Gleichstrombetrieben.
Der Patch-Aufbau konzentriert sich um ein invertiertes Mikroskop, mit dem dasAndocken der Patchpipette an das Vesikel beobachtet wird. Das Mikroskop (Zeiss,

58
Jena) ist auf einem luftgelagerten Tisch aufmontiert. Dieser besteht aus vierdruckgesteuerten Shock Absorbern (4 Bar) mit Eigenfrequenzen von 2 Hz vertikal und 4Hz horizontal. Er fängt Störungen von außen wie Schritte und Schließen von Türen auf.
Mikromanipulatoren ermöglichen die Positionierung sowohl der Patch-Pipette(rechts) als auch des Bades (links). Für die Positionierung der Pipette werden mehrereManipulatoren eingesetzt. Ein grober 3-D Manipulator trägt eine schräg gestellteSchiene, an deren unteres Ende ein hydraulischer Mikromanipulator für dieFeinpositionierung montiert ist. Die Schiene kann mit einem in der Werkstatt des IEAPshergestellten Schlittenantriebs verkürzt oder verlängert werden. Dadurch sindBewegungen senkrecht zur Zelloberfläche möglich. Dies ist nützlich, um eine Pipettevon der Zelle zu entfernen und in gleicher Richtung eine neue zurückzubringen. Derhydraulische Manipulator trägt die Vorstufe des Patchverstärkers, an den über einenBNC-artigen Stecker (ohne Bajonett) der Pipettenhalter angeschlossen wird. Derhydraulische Manipulator (Narishige, Nikon, Japan) ermöglicht die sehr feinkontrollierbare Bewegung beim Andocken der Pipettenspitze an die Plasmamembrandes Protoplasten mit Hilfe eines Joysticks. Der Joystick steht auf einem Holzrahmenneben dem luftgefederten Tisch, um Störungen durch die manuelle Betätigung zuvermeiden. Die Veränderungen des Sealwiderstandes wurden mit Hilfe des Oszilloskopin der Meßkammer verfolgt.
Die normalerweise verwendete Petrischale wurde durch eine Kammer mitSpülmöglichkeit ersetzt, um einen Lösungswechsel zu ermöglichen. Sie kann mit Hilfedes rechten 2D-Manipulators in die richtige Position gebracht werden.
Um unter gleichen Bedingungen zu messen, ist es wesentlich, die Temperaturkonstant zu halten. Dazu wurde ein mobiles Klimagerät (DeLonghi, Treviso, Italien) indie Türöffnung gestellt (Abluft wird aus dem Raum geleitet). So waren auch imHochsommer Messungen möglich.
6.2 Elektrische Schaltungen und Datenaufnahme
Der elektrische Aufbau ist in Abbildung 25 gezeigt. Der Protoplast, dessen elektrischeEigenschaften untersucht werden sollen, liegt in der in Abschnitt 6.1 besprochenenSpülkammer. Die Badelektrode (E) ist eine nicht polarisierbare Ag/AgCl-Elektrode(chlorierter Silberdraht) mit einer Agar-KCl-Brücke (10 mM KCl). Die Patchelektrode(P) besteht aus der oben bereits erwähnten Pipette, die mit einer ca. 150 mM Salzlösungals Leiter gefüllt ist, und einer eingetauchten dünnen Ag/AgCl-Elektrode. Der Elektro-denhalter sitzt direkt auf der BNC-Buchse des Eingangs des Patch-Verstärkers.
Für die Messung steht ein integrierter Patch-Clamp Verstärker (Firma DAGAN 3900A mit Whole-Cell-Expander, USA) zur Verfügung. Im Voltage Clamp Mode wird eine

59
Spannung über der Zellmembran angelegt und der Strom durch die Membran gemessen.Im Gegensatz zu anderen Geräten (z.B. EPC 7, List, Darmstadt) geschieht beim Dagandas Anlegen der Spannung dadurch, daß die Sollspannung an die Badelektrode (E, weilsie normalerweise als Erdelektrode fungiert) gelegt wird und die Pipettenspitze (P) aufPseudo-Null gehalten wird. Die Messung des Stromes und das Halten der Pipettenspitzeauf Pseudo-Null ist Aufgabe der auf dem Manipulator montierten Eingangsstufe desDagans. Sie ist im wesentlichen ein als Stromspannungswandler geschalteterOperationsverstärker.
Abbildung 25. Blockschaltbild des Meßaufbaus. In der Mitte ist der Hauptstrang für dieRegistrierung des Membranstromes. Im oberen Ast wird die softwaremäßig kontrollierteSollspannung für die Zellmembran eingeben. Der untere Ast beschreibt die Eingabe vonSollspannungen aus Funktionsgeneratoren. Dies erfordert dann eine externe Synchronisation derComputer Aufzeichnung. Das eingezeichnete Dreieckssignal wird für die Ermittlung desUmkehrpotentials (Abschnitt 7.4) benötigt.
Die Empfindlichkeit liegt bei allen Patch-Verstärkern im pA Bereich. EineBesonderheit des Stromspannungskonverters im Dagan ist die Möglichkeit, einenKondensator als Rückkopplungselement anstelle eines Gigaohm-Widerstandes zuwählen. Der Kondensator muß zwar von Zeit zu Zeit entladen werden, doch dies störtweniger als das Rauschen Ohmscher Widerstände. Die Bandbreite beträgt 250 kHz.
Als Anti-Aliasing-Filter für die Signale aus dem Dagan dient ein 4-poliger Bessel-Tiefpaß. Sein Ausgangssignal wird über einen Differenzverstärker an den Computerweitergeleitet, um Erdschleifen zu vermeiden (Albertsen, 1994), und von einemDalanco-Board (Modell 250 Dalanco, Spry, USA) (Langmann, 1990) auf die 1 Giga-Byte-Festplatte eines PC486 geschrieben. Der DSP (Digital Signal Processor) auf

60
diesem Board ermöglicht eine Abtastrate von 250 kHz. Das RAM des Signalprozessorsist geteilt: Im Wechsel wird eine Hälfte von dem DSP beschrieben, während die andereHälfte auf die Festplatte gelesen wird. Dadurch ist ein kontinuierlicher Datenstrom aufdie Festplatte möglich (Albertsen, 1992).
Das Programm zur Datenaufnahme (Albertsen, 1992, 1994) besteht aus zwei Teilen:eines läuft auf dem PC, eines auf dem DSP. Das PC-Programm „Sample” fragt dieEinstellungen des DAGAN ab (gain, Filterfrequenz) sowie die Anzahl der Datenpunkte,den Namen der Datenreihe und die Abtastrate; es entsteht ein Versuchsprotokoll.Außerdem räumt das Programm der folgenden Datenaufnahme absolute Priorität ein,d.h. wenn „Sample” läuft, kann kein Programm im Hintergrund weiterlaufen. Mit einemexternen Schalter wird das DSP Programm und somit die Datenaufnahme gestartet. Einezweifarbige LED zeigt hierbei an, ob die Messung läuft oder ob die Datenaufnahmegestartet werden kann.
6.2.1 Whole-Cell-Messungen
Für Whole-Cell-Messungen wird eine Erweiterung des DAGAN benötigt (DAGANWhole-Cell-Expander): Das Gerät muß die Möglichkeit bieten, neben derPipettenkapazität auch die Zellkapazität und den Serienwiderstand elektronisch zukompensieren (Abschnitt 4.5).
Ein Standardverfahren der Elektrophysiologie ist die Aufnahme von Strom-spannungskurven und die Untersuchung des Zeitverhaltens des Stromes bei Sprüngenauf verschiedene Spannungswerte. Zu unterscheiden ist zwischen dem instantanen unddem zeitabhängigen Anteil der Stromantwort.
Es ist also nötig, ein Spannungsprotokoll vorzugeben (Abbildung 36 A). ImAllgemeinen besteht es aus einer Phase mit einer negativen Spannung nahe demUmkehrpotential des dominierenden Transporters (meistens EK) als sogenanntesHaltepotential (sozusagen als „Erholung“ für die Zelle), gefolgt von einer kurzenAktivierungs- oder Deaktivierungsphase und dann dem Sprung auf die jeweils zuuntersuchende Spannung.
Das Spannungsprotokoll wird dazu als File eingelesen (Dauer der Spannung,Spannungswert) und mit dem Programm Capstint (Albertsen, 1992) aktiviert, d.h. vomDalanco-Board in Spannungswerte umgesetzt und über einen DA-Wandler über denDAGAN an die Elektrode (E) weitergegeben.
Während bei Einzelkanalaufnahmen sehr häufig biophysikalische Fragen imVordergrund stehen, die eine hohe Zeitauflösung mit geringem Rauschen erfordern,(Keunecke, 1995, Farokhi, et al., 1999), steht bei Whole-Cell-Aufnahmen das Verhaltendes Gesamtstromes im Vordergrund, in dem sich die schnellen Prozesse weggemittelt

61
haben. Abtastraten liegen daher zwischen 0,5 und 5 kHz. Die im Rahmen dieser Arbeitdurchgeführten Messungen wurden mit 2 kHz aufgezeichnet und mit 1 kHz gefiltert.
6.3 Herstellung der Pipetten
Ausgangsmaterial sind ca. 10 cm lange Stücke von Glasröhrchen mit 1,0 mmInnendurchmesser und 1,5 mm Außendurchmesser (Einzelkanalmessungen: GarnerGlass, Kalifornien, USA, Whole-Cell-Messungen: Kimax, Kimble Products, USA).Bevor aus ihnen Pipetten gezogen werden, werden die Glasröhrchen von innen mitSigmacote (Sigma-Chemie, Deisenhofen) beschichtet: Unter dem Abzug wird mit einerSpritze ein Tropfen Sigmacote in jedes Röhrchen gegeben, so daß es unten wiederherausläuft. Die Reste, die sich an den Innenwänden befinden, trocknen in 2 Tagenunter dem Abzug. Auf eine äußere Beschichtung mit Sylgard (Dow Corning, USA)wurde verzichtet, da sich Sigmacote als effektiver erwiesen hat (Keunecke, 1995,Hansen et al., 1997).
Im Pipettenziehgerät (L/M-3P-A, List, Darmstadt) werden die Pipetten aus denGlasröhrchen gezogen. Die Glasröhrchen werden dazu oben und unten eingeklemmt,wobei sich der mittlere Teil in einer Glühwendel befindet. Die Wahl der Ströme, diedurch die Glühwendel fließen (um 20 A), beeinflußt die Form und Größe der Spitzeentscheidend. Das Ziehen geschieht in zwei Stufen: In der ersten wird das Glas nuretwas verdünnt, in der zweiten Stufe wird das Glas so weit erhitzt, daß sich die Pipettenbilden (das Glas reißt ab). Die zweite Einstellung bestimmt im Wesentlichen denPipettendurchmesser (und somit den Pipettenwiderstand). ElektronenmikroskopischeAufnahmen von Plieth (1995) ergaben einen Spitzendurchmesser von ca. 1 µm.
Anschließend wurden die Pipetten poliert. Dazu wird eine mit Glas beschichteteGlühwendel für ca. 3 Sekunden in etwa 30 µm Abstand an die Pipettenspitze gehalten.Die Spitze schmilzt hierbei leicht an und wird glatter, so daß eine Sealbildung leichtermöglich ist.

62
7 Einzelkanalmessungen
Abbildung 26. Zeitreihe einer Maisbündelscheidenzelle (0.23 s). Gemessen wurde im Excised-Patch (Stück Membran aus dem Protoplasten herausgerissen, Abschnitt 4.5) bei -140 mV. Lö-sungen: 100 mM KCl + 5 mM CaCl2 auf beiden Seiten der Membran (Pipette und Bad).
In Einzelkanalmessungen ist es möglich, den Strom von Ionen durch einzelne Kanäle zubestimmen (Abbildung 26) und deren Schließverhalten zu untersuchen. Im Excised-Patch, bei dem nur ein Stück Membran vor der Pipette ist (ohne Verbindung zum Pro-toplasten, Abschnitt 4.5), kontrolliert man zudem beide Lösungen (in der Pipette und imBad). Unter diesen Bedingungen lassen sich die Kanäle gut klassifizieren, also z.B. dasIon herausfinden, das den Kanal hauptsächlich permeiert (Umkehrpotential, Abschnitt7.4).
Sind die Einzelkanalströme sehr klein und das Schalten sehr schnell, wie bei denBündelscheidenzellen aus Abbildung 26, lassen sich die Ströme allerdings nur sehrschwer (oder gar nicht) bestimmen. Man muß daher mit unphysiologisch hohen Salz-konzentrationen arbeiten (200 mM KCl). Für die biophysikalische Klassifizierung(Wechselwirkung verschiedener Ionen auf Strom und auf Schaltverhalten) stellt das keinHindernis dar, im Gegenteil, einige Effekte treten erst unter sehr hohen Konzentrationenauf (z.B. nimmt der Strom nicht unbegrenzt mit der K+-Konzentration zu, Gambale etal., 1996). Solche Effekte sind wichtig, wenn man die Arbeitsweise eines Kanals verste-hen will.

63
7.1 Symmetrischer K+-Kanal und Einwärtsgleichrichter
Bei den Einzelkanalmessungen an Bündelscheidenzellen traten zwei Typen von K+-Kanälen auf, die durch ihre Umkehrspannungen charakterisiert wurden (Keunecke et al.,1997):- ein Kanal mit symmetrischer Stromspannungskurve (SC, Abbildung 27); bei den mei-sten Messungen (ca. 92%) wurde dieser Typ gefunden;- ein Einwärtsgleichrichter (IR), der im negativen Spannungsbereich mit dem SC iden-tisch schien, aber bei positiven Spannungen entweder gar nicht öffnete (z.B. +60 mVoder +80 mV in Abbildung 27), oder öffnete, aber einen wesentlich kleineren Stromzeigte, als der SC. Dies wird auch deutlich in der Offenwahrscheinlichkeit (Abbildung28).
Abbildung 27. Spannungsabhängigkeit des symmetrischen Kanals (SC) und des Einwärts-gleichrichters (IR). Bei positiven Spannungen schaltet der Einwärtsgleichrichter mitunter garnicht auf (60 und 80 mV). Bad und Pipette: 200 mM KCl + 5 mM CaCl2.
Beide Kanaltypen spielen ebenfalls in anderen Objekten eine Rolle. So ergeben derIR und der spannungs-insensitive K+-Kanal den gesamten K+-Strom für das Wachstumvon Keimlingen (Gassmann und Schroeder, 1994, White und Lemtiri-Chlieh, 1995).
-100 0 100
-2
-1
0
1
2
IR
SC
i (pA
)
U (mV)

64
Für den Ferntransport der Pflanze ist interessant, daß Wegner und Raschke (1994)in den Parenchymzellen der Wurzel überwiegend Auswärtsgleichrichter fanden. Im Sin-ne eines K+-Stromes von der Wurzel über das Xylem in den Blattsymplasten war zuerwarten, daß es im Blatt in Zellen mit Xylemkontakt einen Einwärtsgleichrichter gab.
7.2 Blockierung der Kanäle durch Weichglas
Erste Messungen an Bündelscheidenzellen ergaben, wenn überhaupt, dann nurkurzzeitiges Schalten. Nach kurzer Zeit sank die durchschnittliche Zahl der schaltendenKanäle beträchtlich. Nach Copello et al. (1991) kann dies auf zweiwertige Ionen zu-rückgeführt werden, die aus der Pipette austreten. Dieser Austritt konnte vermiedenwerden, indem die Pipette vor dem Ziehen von innen mit Sigmacote (Sigma) beschich-tet wurde (Abschnitt 6.3). So stieg die durchschnittliche Anzahl von offenen Kanälen ineinem Patch von 3,4 ± 0,3 ohne Beschichtung auf 8,8 ± 0,4 an. Die Beschichtung mitSigmacote hatte sich zudem als rauschreduzierend erwiesen (Keunecke, 1995).
Borosilikatglas, ein Hartglas, welches weniger Rauschen erzeugt (Fairley and Wal-ker, 1989), konnte dann für die vollständig isolierten Protoplasten verwendet werden.
7.3 Einwärtsgleichrichter
Der Einwärtsgleichrichter trat bei KCl-Konzentrationen von 10, 100 und 200 mMauf. Er hatte ein deutlich asymmetrisches Schaltverhalten: Bei positiven Spannungenwar häufig gar kein Öffnen zu erkennen.
Die Asymmetrie der Offenwahrscheinlichkeit ist in Mehrkanalanalysen am leichte-sten durch die Anzahl maximal gleichzeitig offener Kanäle zu zeigen. In Abbildung 28sieht man, daß bei der 100 mV Kurve (IR+) maximal nur bis zu 43% der Kanäle gleich-zeitig öffnen. Im Mittel waren es 14% bis 29%. Bei negativer Spannung, also in Durch-laßrichtung, waren hingegen bei -100 mV im Mittel 57% der Kanäle (Kurve IR-) offen.
Im Gegensatz dazu zeigt der symmetrische K+-Kanal ein sehr symmetrisches Schalt-verhalten. Die Offenwahrscheinlichkeiten für positive (+100 mV, Kurve SC+ inAbbildung 28) und negative (-100 mV, Kurve SC-) Spannung liegen übereinander.

65
Der Einwärtsgleichrichter trat sehr viel seltener auf als der symmetrische (bei etwa8% der Messungen).
Abbildung 28. Offenwahrscheinlichkeit je Stromniveau des symmetrischen Kanals (SC) unddes Einwärtsgleichrichters (IR) bei - 100 mV (-) und bei + 100 mV (+). Auch bezüglich derOffenwahrscheinlichkeit verhält sich der IR für negative Spannungen wie der SC.
7.4 Selektivität und Umkehrpotential des symmetrischen K+
Kanals
Um zu klären, welches Ion den Strom durch den Kanal trägt, muß unter asymmetrischenBedingungen gemessen werden. Hier wurden Messungen zum Umkehrpotential durch-geführt, indem Stromspannungskurven bei variierenden luminalen und cytosolischenK+-Konzentrationen aufgenommen wurden, während jeweils die eine Seite auf 200 mMfestgehalten wurde.
Besonders gut kann das Umkehrpotential bestimmt werden, indem eine Dreiecks-spannung angelegt wird (Kunert, 1998, Abbildung 29). Beim Umkehrpotential ist keinSchalten zu erkennen und somit die Varianz minimal (Kunert, Abbildung 30). DiesesVerfahren hat den Vorteil, daß selbst bei sehr stark verrauschten Daten noch das Um-kehrpotential erkannt werden kann.
0 20 40 60 80 1000,0
0,2
0,4
0,6
0,8
SC -
SC +IR -
IR +
p
% channels open

66
Abbildung 29. Aufnahme von IU-Kurven mit Spannungsrampen. Das Programm erkennt mitHilfe des TTL-Signals die Vorzeichenwechsel der Dreiecksspannung und kann so die Strom-werte den Spannungen zuordnen.
Abbildung 30. Fit (Polynom 8. Ordnung) der Stromvarianz der abfallenden Flanken (abneh-mende Spannung) einer Zeitreihe (100 mM KCl + 5 mM CaCl2, beidseitig, Bündelscheiden-zellen). Frequenz der Dreiecksspannung 0.5 Hz, Samplefrequenz 50 kHz. Das Minimum derVarianz lag bei –35 mV.
Es ist zu erkennen, daß der Kanal nicht sehr K+-selektiv ist. Aus der Abweichungvon EK vom Nernstpotential (Diagonale in Abbildung 31) läßt sich die Durchlässigkeitvon K+ im Vergleich zu Cl- bestimmen, sie liegt bei PK : PCl = 3.4 : 1 (berechnet mit derGoldman-Gleichung, Abschnitt 4.2). Ca2+-Konzentrationen wurden bei den Messungennicht verändert und lagen zudem symmetrisch vor, Ca2+ kam daher nicht für die Ver-schiebung des Umkehrpotentials in Frage.
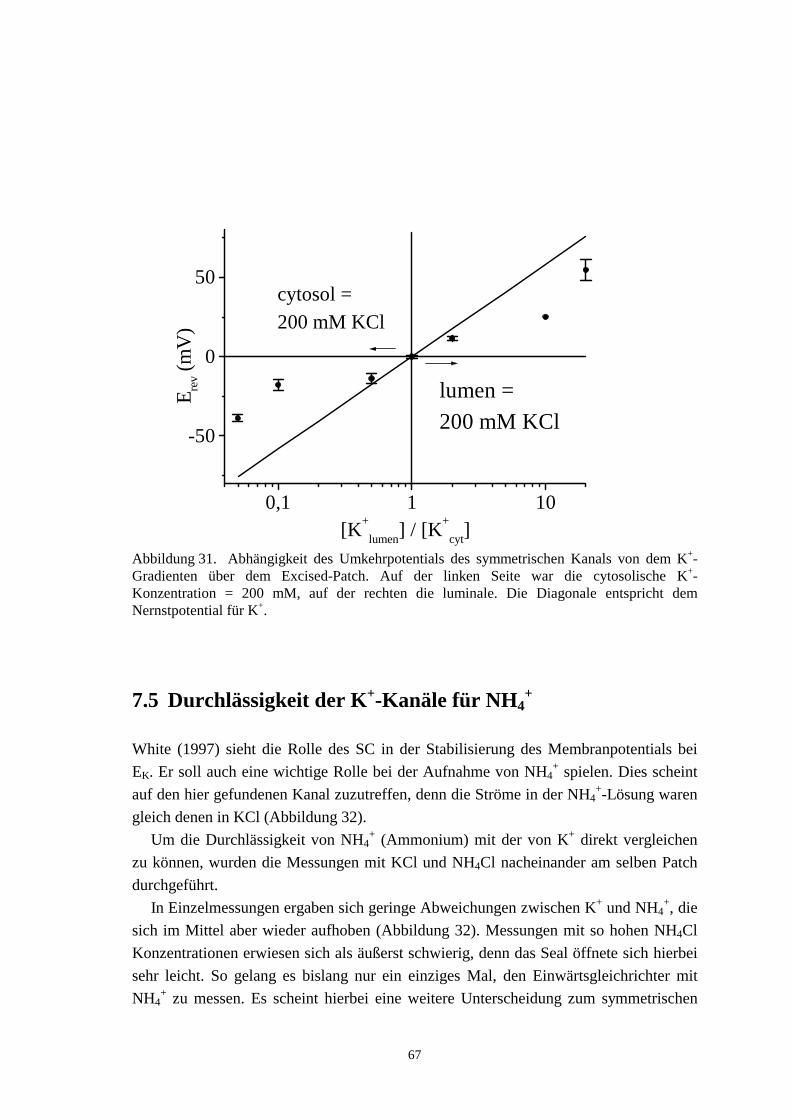
67
Abbildung 31. Abhängigkeit des Umkehrpotentials des symmetrischen Kanals von dem K+-Gradienten über dem Excised-Patch. Auf der linken Seite war die cytosolische K+-Konzentration = 200 mM, auf der rechten die luminale. Die Diagonale entspricht demNernstpotential für K+.
7.5 Durchlässigkeit der K+-Kanäle für NH4+
White (1997) sieht die Rolle des SC in der Stabilisierung des Membranpotentials beiEK. Er soll auch eine wichtige Rolle bei der Aufnahme von NH4
+ spielen. Dies scheintauf den hier gefundenen Kanal zuzutreffen, denn die Ströme in der NH4
+-Lösung warengleich denen in KCl (Abbildung 32).
Um die Durchlässigkeit von NH4+ (Ammonium) mit der von K+ direkt vergleichen
zu können, wurden die Messungen mit KCl und NH4Cl nacheinander am selben Patchdurchgeführt.
In Einzelmessungen ergaben sich geringe Abweichungen zwischen K+ und NH4+, die
sich im Mittel aber wieder aufhoben (Abbildung 32). Messungen mit so hohen NH4ClKonzentrationen erwiesen sich als äußerst schwierig, denn das Seal öffnete sich hierbeisehr leicht. So gelang es bislang nur ein einziges Mal, den Einwärtsgleichrichter mitNH4
+ zu messen. Es scheint hierbei eine weitere Unterscheidung zum symmetrischen
0,1 1 10
-50
0
50cytosol = 200 mM KCl
lumen = 200 mM KCl
E rev (
mV
)
[K+lumen] / [K
+cyt]

68
Kanal zu geben, denn die Durchlässigkeit für NH4+ ist deutlich verschieden von der für
K+.
Abbildung 32. A. Die Durchlässigkeit von NH4+ beim symmetrischen K+-Kanal. Sie ist genau-
so groß wie die von K+. B. Die Durchlässigkeit von NH4+ beim einwärtsgleichrichtenden K+-
Kanal. Da die Einzelkanalströme recht klein sind, wurde bei hohen Konzentrationen gemessen.
7.6 Na+-Messungen
Für den Transport zwischen Apoplast und Symplast ist wichtig, wie die Transportakti-vität durch das apoplastische Milieu beeinflußt wird. Im Hinblick auf Salzstreßuntersu-chungen, wie sie z.T. in der Arbeitsgruppe Heldt durchgeführt werden, ist deshalb dieBlockierung durch Na+ untersucht worden. Bei 100 mM KCl + 100 mM NaCl in derBadlösung ist im negativen Spannungsbereich der Kurvenverlauf praktisch gleich. Impositiven Bereich bleibt die Kurve mit NaCl in der Lösung deutlich unterhalb der Kurvemit symmetrischen Lösungen. NaCl wirkte stets, wenn der Strom Na+-Ionen aus derBad- oder Pipettenlösung in die K+-Kanäle schwemmt. Die Umkehrspannung bleibt inGegenwart von NaCl bei EK, dem Nernstpotential für K+. Na+ permeiert den K+-Kanalalso nicht.
-100 -50 0 50 100-2
-1
0
1
2
lumen : 200 KCl + 5 CaCl2cytosol: 5 CaCl2 +
200 KCl 200 NH4Cl
i (pA
)
U (mV)
-200 -100 0 100
-3
-2
-1
0
1
2cyt = 2 KCl + 2 CaCl2 +200 NH4Cllumen = 200 KCl
+ 2 CaCl2 + 2 NH4Cl
cyt = 200 KCl + 2 CaCl2 + 2 NH4Cli (
pA)
U (mV)

69
A B
Abbildung 33. A. Stromspannungskurve unter Einfluß von Na+. Es tritt eine kleine Verringe-rung des Auswärtsstromes auf. B. Betrachtet man die Anzahl der offenen Kanäle im Patch, soist hier ebenfalls ein Na+-Block ersichtlich: Bei reiner Kaliumlösung sind im Schnitt deutlichmehr Kanäle offen als in gemischter Lösung. Bad: 200 KCl bzw. 100 KCl + 100 NaCl jeweils+ 5 CaCl2. Pipette: 100 KCl + 5 CaCl2.
Messungen, bei denen K+ vollständig durch Na+ (200 mM) ersetzt wurde, zeigten,daß Na+ den symmetrischen K+-Kanal passieren kann. Die Leitfähigkeit ist nur um 10 -40% geringer als die von K+. Wie schon beim Umkehrpotential in KCl-Lösung inAbbildung 31, zeigt sich auch hier, daß der symmetrische Kanal nicht sehr spezifischist. Diese blockierende Wirkung von Na+ ist allgemein bekannt (Thiel und Blatt, 1991,Kourie, 1996a,b, Dietrich et al., 1998).
Auch in den Offenwahrscheinlichkeiten ist der Unterschied zwischen reiner Kali-umlösung und reiner Natriumlösung nicht sehr groß. Die Unspezifität gegenüber Kali-um fanden Wegner und Raschke (1994) auch bei einem K+-Kanal in Wurzelxylempar-enchymzellen, bei dem sogenannten NORC (non selective outward rectifying channel).
-100 -50 0 50 100
-4
-2
0
2
4
6200 K
100 K + 100 Na
i (pA
)
V (mV)0 2 4 6 8 10
0,0
0,1
0,2
0,3
0,4
0,5
0,6
Na+
K+
p
level

70
Abbildung 34. Auftreten einer erhöhten Leitfähigkeit durch Na+. Stromspannungskurve in 200mM NaCl + 2 mM KCl + 5 mM CaCl2 (Na) und Stromspannungskurve in 200 mM KCl + 5mM CaCl2 (K) sowie 200 mM KCl + 2 mM NaCl + 5 mM CaCl2 in der Pipette. Die Messungvon C. Möller (1998).
Abbildung 34 zeigt einen anderen, für den Salzstreß möglicherweise wichtigen Ef-fekt. Bei Abwesenheit von K+ auf der cytosolischen Seite (Bad) zeigt der IR eine Na+-Leitfähigkeit. Diese in reiner Natriumlösung beobachtete Leitfähigkeit fällt im negativenSpannungsbereich mit der für K+ zusammen. Bei positiven Spannungen hingegen ist beiNa+ der Gleichrichtereffekt nicht zu beobachten. Es treten sehr hohe Ströme auch impositiven Bereich auf. In diesem Bereich werden die Na+-Ionen durch den Strom vomBad in die Membran getragen, im negativen Bereich trägt der Strom die K+-Ionen ausder Pipette in die Membran. Somit ist nicht erstaunlich, daß der K+/Na+ Unterschied impositiven Bereich auftritt.
Die hier beobachtete Erhöhung der Leitfähigkeit in Gegenwart von Na+ wurde auchvon Amtmann et al. (1997) in Gerstenwurzelsuspensionszellen und Tyerman et al. (1997)in Weizencortexprotoplasten beobachtet. Na+-Leitung durch Shaker-Typ Kanäle bei K+
Abwesenheit ist jetzt auch in tierischen Zellen beobachtet worden (Starkus et al., 1997,1998, Ogielska und Aldrich, 1998).
-100 0 100 200-4
-3
-2
-1
0
1
2 K+Na+
spiky
Bad: 5 CaCl2 +200 NaCl + 2 KCl oder 200 KClPipette: 200 KCl + 2 NaCl + 5 CaCl2
i (pA
)
V (mV)

71
7.7 Physiologische Bedingungen
Abbildung 35. Messung unter physiologischen K+-Konzentrationen (100 mM KCl im Bad und10 mM KCl in der Pipette). Für den Bereich 0 > U > - 30 mV waren die Einzelkanalströme füreine Auswertung zu gering.
Patchen gelingt meistens nur unter unphysiologisch hohen Salzkonzentrationen. Diesist für biophysikalische Untersuchungen, bei denen die auftretenden Transporter ersteinmal klassifiziert und charakterisiert werden sollen, ein sinnvoller Anfang. Im Rah-men des Apoplastenprojektes geht es jedoch um die physiologische Rolle des Trans-portes zwischen Apoplast und Symplast und dafür sollten die Experimente unter denwirklich auftretenden Salzkonzentrationen stattfinden.
Abbildung 35 zeigt, daß unter physiologischen K+-Bedingungen gemessen werdenkann. Die apoplastische K+-Konzentration liegt im Bereich um 10 mM (Sattelmacher etal., 1996, Felle und Peters, 1996, Lohaus et al., 1996), die im Cytosol um 100 mM. ImExcised-Patch sind leider immer unphysiologische Calciumbedingungen nötig, damitder Patch stabil bleibt. Gemessen wird im Bereich von 1 bis 20 mM Ca2+ (Laver undFairley-Grenot, 1994). Mit der Whole-Cell Technik (Kapitel 8 und 9) sind Messungenbis zu Calciumkonzentrationen von 10 nM möglich (Skerrett und Tyerman, 1994).
-50 0 50-2
-1
0
1
2
5 CaCl2 +cytosol = 100 KCl lumen = 10 KCl
i (pA
)
V (mV)

72
8 Whole-Cell-Messungen an Bündelscheiden-zellen
Whole-Cell-Messungen ergeben den Summenstrom über alle Transporter in derMembran der ganzen Zelle. Der Nachteil dieser Methode ist, ebenso wie bei Messungenmit Einstichelektroden, daß die Zuordnung zu einem bestimmten Transporter nur dannmöglich ist, wenn dieser das elektrische Verhalten der Membran dominiert (Fisahn etal., 1986). Als Vorteile kann man werten, daß man einen Überblick über das Verhalteneiner ganzen Zelle erhält und daß die Messungen, wie bei klassischenEinstichelektroden (Umrath, 1932), unter nahezu physiologischen Bedingungenstattfinden. Die Forderung, daß Ca2+ im millimolaren Bereich in der Badlösungvorliegen muß, bedeutet im Gegensatz zum Inside-Out Patch nicht unbedingt eineunphysiologische Bedingung, da Zellen im Außenmedium hohe Ca2+-Konzentrationenmögen. Im Gegenteil fallen viele Funktionen aus (z. B. die Protonenpumpe, Spanswicket al., 1967), wenn die Außenlösung zu wenig Ca2+ enthält. Die Angaben über die freieCa2+-Konzentration schwanken allerdings sehr stark von <10 µM bis 1 mM (Plieth etal., 1999c). Welche Konzentration an der Plasmamembran wirklich herrschen, ist unklarund hängt vom Modell der Zellwand ab. Nach dem immer noch gültigen Bild vonDainty und Hope (1959) gehen wassergefüllte Poren durch die Zellwand bis an dasPlasmalemma. In ihnen (water-free space) können höhere Konzentrationen an Ca2+
vorliegen als im Bereich der Festladungen (donnan-free space).Für Whole-Cell-Messungen müssen die Pipetten eine größere Öffnung als bei den
Einzelkanalmessungen haben, damit sich die durch Saugen erzeugte Öffnung in derMembran (Abschnitt 4.5) nicht wieder verschließt. Die daraus folgendenSchwierigkeiten bei der Präparation der Bündelscheidenzellen und ihre Lösung sind im5. Kapitel beschrieben.

73
8.1 Spannungsprotokoll
Whole-Cell-Messungen wurden mit folgendem Meßprotokoll aufgenommen:Eine Meßreihe besteht aus 15 Blöcken, die jeweils aus 3 Zeitabschnitten bestanden.- 10 s lang - 60 mV Haltepotential für alle Blöcke- 2 s lang Hyperpolarisation auf - 140 mV für alle Blöcke- 15 s lang mit konstanter, aber von Block zu Block stufenweise veränderter
Spannung.Die Spannungen im 15 s Intervall stiegen in 20 mV Schritten
von - 60 mV auf - 140 mV,dann von - 40 mV auf + 40 mV,dann von - 160 bis – 200 mVund dann von + 60 mV bis von + 80 mV.
Abbildung 36 A zeigt das verwendete Spannungsprotokoll. Der Übersicht halber sinddie Spannungsblöcke ( - 60 mV, - 140 mV, Testpotential) nicht hintereinander, sondernüberlagert dargestellt.
Spannungen über + 40 mV und unter - 140 mV stellen eine starke Belastung für dieZelle dar, daher wurden diese Bereiche immer ganz zum Schluß gemessen. So konntezumindest ein breiter Bereich beidseitig des Umkehrpotentials ausgemessen werden,bevor die Zelle eventuell zerstört wurde.
Als Haltepotential wurde – 60 mV angelegt, da dies beim Nernstpotential für K+ liegt(innen etwa 100 mM K+ und außen 10 mM) und somit keine Belastung für die Zelledarstellte. Um die Kanäle in der Zelle zu deaktivieren, folgte ferner vor dem Testpoten-tial eine stark negative Spannung (- 140 mV).

74
Abbildung 36. Zeitreihen des MB-1 (B) und des MB-2 (C) gewonnen mit dem Spannungs-protokoll aus A. Der Übersicht halber sind die Spannungsblöcke ( - 60 mV, - 140 mV, Testpo-tential) überlagert dargestellt. Die Antworten des MB-2 zeigen eine zeitabhängige Aktivierung,während der MB-1 nur einen instantanen Stromfluß besitzt. Dieser ist zudem so gering, daßsich Einzelkanalschalten erkennen läßt (hier nicht auflösbar aufgrund der Zeitskala). Lösungen„braun“ und „blau“ aus Tabelle 8.
0 1 0 2 0
A
t / s
+ 8 0 m V
- 2 0 0 m V- 1 4 0 m V
- 6 0 m V

75
8.2 Verschiedene K+-Kanal-Typen bei Bündelscheidenzellen
Unter physiologischen Kaliumkonzentrationen (10 mM außen und 100 mM innen, Lö-sung "braun" in Tabelle 8) traten bei den Bündelscheidenzellen in den Whole-Cell-Messungen zwei Typen von Antworten (Abbildung 36) auf, die wahrscheinlich (Ab-schnitt 8.4 mit Tabelle 9) zwei verschiedenen Typen von Bündelscheidenzellen zuge-ordnet werden können:- Typ 1 zeigte keine Zeitabhängigkeit des Aktivierungszustands nach einem Span-
nungssprung, so daß er den vollen Wert des Stromes instantan lieferte. Er wurdeMB-1 oder auch MB-KI1 (Mais Bündelscheidenzellen-K+ instantaner Typ 1) ge-nannt.
- Typ 2 zeigte ein spannungs- und zeitabhängiges Aktivierungsverhalten. Abbildung36 C zeigt, daß er die Stromspannungskurve eines Auswärtsgleichrichters erzeugt.Er wurde MB-2 (Mais Bündelscheidenzellen Typ 2) genannt.Typ 2 besteht aus zwei Komponenten (Abbildung 37). Diese beiden Komponenten,die instantane und die zeitabhängige, erhielten die Namen MB-KI2 (Mais Bündel-scheidenzellen- K+ instantaner Typ 2) und MB-AOR (Mais Bündelscheidenzellen-aktivierter Auswärtsgleichrichter, „Outward-Rectifier“).
Die Skalierung in Abbildung 36, aber insbesondere die Stromspannungskurven inAbbildung 37 zeigen, daß der Strom des MB-1 deutlich unter dem des MB-2 liegt. Beidem instantanen Zelltyp (MB-1) ist zudem häufig schnelles Einzelkanalschalten zu er-kennen (Abbildung 38).

76
Abbildung 37. Stromspannungskennlinie der instantanen Ströme des MB-KI2 und des MB-1.Beim MB-1 sind Einzelkanalereignisse sichtbar. Haltepotential = -60 mV, angelegte Spannung- 200 bis + 80 mV in 20 mV Schritten. Lösungen „braun“ und „blau“ aus Tabelle 8 (pH 7.2). n= 11 bzw. 15 (MB-1).
Abbildung 38. Vergrößerter Ausschnitt aus einer Whole-Cell-Antwort des MB-1. Einzelkanal-schalten mit einem Strom von –3,3 pA (Abstände zwischen den Linien) bei einer Spannung von-140 mV ist zu erkennen.
-200 -150 -100 -50 0 50-6
-3
0
3
6 MB-KI2
MB-1
I (µA
cm
-2)
V (mV)

77
8.3 ATP-Abhängigkeit
Abbildung 39. Auswascheffekt beim MB-2: Obere Kurve: Stromspannungskurve direkt nachder Sealbildung aufgenommen. Untere Kurve: Stromspannungskurve 3 min später. Die Mes-sungen fanden ohne ATP in der Pipette statt (Lösungen „rot“ und „grün“ aus Tabelle 8).
Bei Whole-Cell-Präparaten traten anfangs sogenannte Run-Down-Effekte auf: Direktnach der Herstellung der Whole-Cell-Konfiguration wurden große Auswärtsströme ge-messen (Abbildung 39). Nach ca. 3 Minuten jedoch verminderten sich diese Strömedrastisch, und es ergab sich die Stromspannungskurve mit deutlich reduziertem Aus-wärtsstrom in Abbildung 39. Solche „Run-Downs“ sind aus der Literatur bekannt (Ka-mouchi et al., 1997, Robson und Hunter, 1997). Ein Run-Down kann verhindert werden,wenn der Austausch des Zellinneren mit dem Pipetteninhalt unterbunden wird. Dieskann dadurch geschehen, daß die Integrität der Zellmembran erhalten bleibt (Boyumund Guidotti, 1997). Hierbei wird der elektrische Zugriff zum Zellinneren nicht durcheinen mechanischen Durchbruch erreicht, sondern durch den Einsatz des AntibiotikumsNigericin, ein H+-K+ Austauscher (Titievsky et al., 1996). Dieser bewirkt eine niede-rohmige Verbindung zwischen Pipette und Zellinnerem (Horn und Marty, 1988, Sala etal., 1991). Der Run-Down-Effekt ist also auf eine Auswaschung zurückzuführen.
-200 -100 0 100
0
200
400
3 min lateri / p
A
V / mV
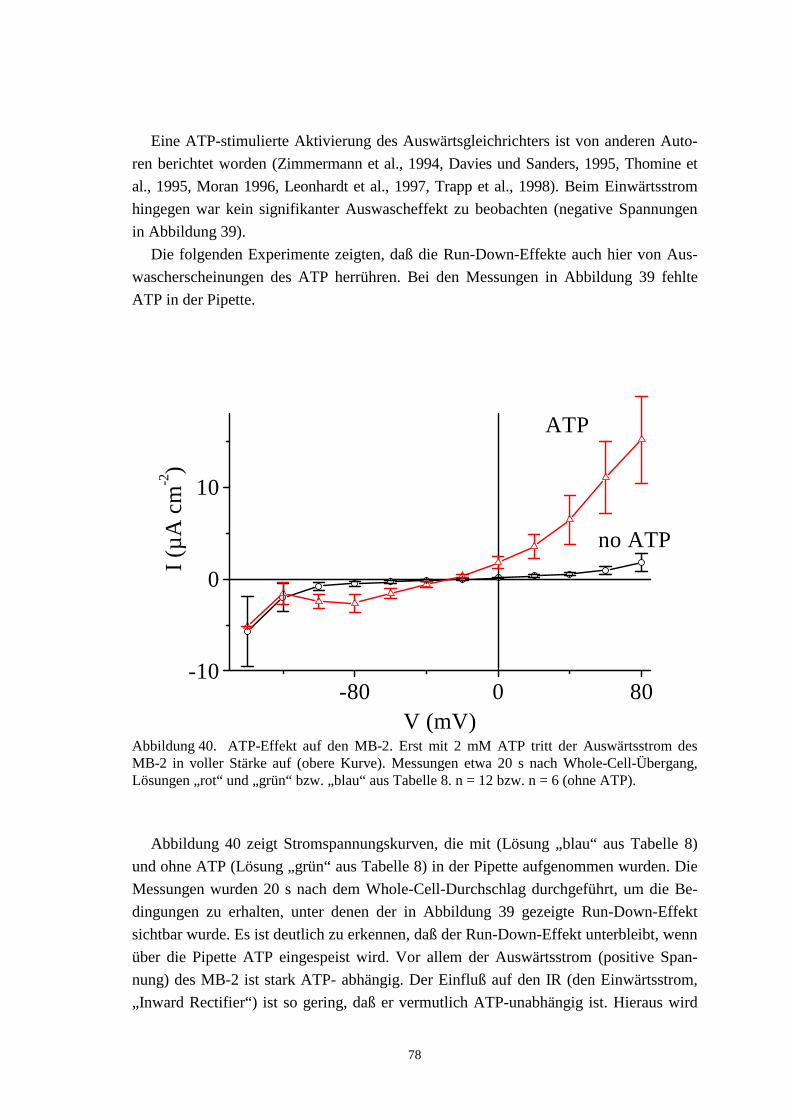
78
Eine ATP-stimulierte Aktivierung des Auswärtsgleichrichters ist von anderen Auto-ren berichtet worden (Zimmermann et al., 1994, Davies und Sanders, 1995, Thomine etal., 1995, Moran 1996, Leonhardt et al., 1997, Trapp et al., 1998). Beim Einwärtsstromhingegen war kein signifikanter Auswascheffekt zu beobachten (negative Spannungenin Abbildung 39).
Die folgenden Experimente zeigten, daß die Run-Down-Effekte auch hier von Aus-wascherscheinungen des ATP herrühren. Bei den Messungen in Abbildung 39 fehlteATP in der Pipette.
Abbildung 40. ATP-Effekt auf den MB-2. Erst mit 2 mM ATP tritt der Auswärtsstrom desMB-2 in voller Stärke auf (obere Kurve). Messungen etwa 20 s nach Whole-Cell-Übergang,Lösungen „rot“ und „grün“ bzw. „blau“ aus Tabelle 8. n = 12 bzw. n = 6 (ohne ATP).
Abbildung 40 zeigt Stromspannungskurven, die mit (Lösung „blau“ aus Tabelle 8)und ohne ATP (Lösung „grün“ aus Tabelle 8) in der Pipette aufgenommen wurden. DieMessungen wurden 20 s nach dem Whole-Cell-Durchschlag durchgeführt, um die Be-dingungen zu erhalten, unter denen der in Abbildung 39 gezeigte Run-Down-Effektsichtbar wurde. Es ist deutlich zu erkennen, daß der Run-Down-Effekt unterbleibt, wennüber die Pipette ATP eingespeist wird. Vor allem der Auswärtsstrom (positive Span-nung) des MB-2 ist stark ATP- abhängig. Der Einfluß auf den IR (den Einwärtsstrom,„Inward Rectifier“) ist so gering, daß er vermutlich ATP-unabhängig ist. Hieraus wird
-80 0 80-10
0
10
no ATP
ATP
I (µA
cm
-2)
V (mV)

79
geschlossen, daß die Verminderung der Auswärtsströme auf eine Auswaschung desATPs und damit eine Deaktivierung des MB-2 zurückzuführen war.
Der Auswascheffekt (Abbildung 39 und Abbildung 40) erklärt, warum bei bisherigenEinzelkanalmessungen (Kapitel 7) kein Auswärtsgleichrichter gefunden wurde. Die da-maligen Messungen fanden ohne ATP-Zugabe statt.
8.4 Gibt es zwei Arten von Bündelscheidenzellen?
Die beiden Typen MB-2 und MB-1 traten in Einzelkanalmessungen stets getrennt (dasheißt in verschiedenen Präparaten) auf. Dieses Verhalten konnte in Whole-Cell-Messungen wiedergefunden werden. Da die Verunreinigungen durch andere Zelltypenrecht sicher auszuschließen sind (Abschnitt 5.2), kann man davon ausgehen, daß sichdie bei der Präparation an den Leitbündelsträngen haftenbleibenden Bündelscheiden-zellen funktionell unterscheiden.
Aus Patch-Untersuchungen sind Unterschiede bei auf den ersten Blick gleichen Zel-len bekannt. In Gerstenwurzeln fanden Wegner et al. (1995) einen Unterschied bei ver-schiedenen Xylemparenchymzellen. Hier konnte ein Zusammenhang mit der Zellher-kunft (Entfernung von der Wurzelspitze) hergestellt werden (Köhler et al., in Vorberei-tung). Doch da die in dieser Arbeit behandelten Messungen im mittleren Blattabschnittstattfanden und dieser Bereich bislang keine Veranlassung gibt, weiter unterteilt zu wer-den, kommen großräumige Unterschiede nicht in Frage. Die Differenzierung des Blattesist vollständig abgeschlossen. Der Übergang von Sink zu Source setzt ein, sobald dasBlatt dem Licht ausgesetzt wird (Evert et al., 1996), während in der Wurzel unter-schiedlich zusammengesetzte Bereiche koexistieren (Schreiber und Hartmann, 1998). Esist also eine Differenzierung in kleinem Maßstab zu suchen.
Einen Hinweis auf eine Differenzierung im Kleinen gibt folgendes: Bei Betrachtungeines Leitbündels (Abbildung 7) fällt auf, daß nicht alle Bündelscheidenzellen Kontaktzum Xylem haben. Daher tauchte nun die Frage auf, ob diese Zellen verschiedeneTransportfunktionen wahrnehmen. Leider geht bei den bisherigen Präparationsverfahrendie Kenntnis verloren, ob die bearbeiteten Protoplasten von Bündelscheidenzellen mitund ohne direkten Xylemkontakt stammen.
Es wurde deshalb versucht, über eine Statistik obige Hypothese zu testen. Dazu wur-den zum einen die Zellen mit und ohne Kontakt in Schnitten von Blättern ausgezählt,die vom Blattalter und Blattabschnitt denen entsprachen, die sonst für Patch-Messungenverwandt wurden, zum anderen die Messungen mit dem MB-2 und dem MB-1.

80
Tabelle 9 zeigt das Ergebnis der Auszählung. (46±2)% der Bündelscheidenzellen ha-ben direkten Kontakt zum Xylem. Bei (43±3)% der elektrophysiologischen Untersu-chungen trat der MB-2 auf.
Tabelle 9. Vergleich der Wahrscheinlichkeiten des Auftretens von morphologischen undelektrophysiologischen Merkmalen. In 43% der Messungen trat der zeitabhängig aktivierteStrom auf. Dies stimmt überein mit dem Anteil von 46% der Bündelscheidenzellen, die in di-rektem Kontakt zum Xylem stehen. Mean = durchschnittliche Anzahl, sd = Stan-dardabweichung, se = Standardfehler der Abweichung, n = Anzahl der gemessenen Leitbündelbzw. der durchgeführten Patch-Clamp Messungen, X = Xylemelemente je Leitbündel, BS =Bündelscheidenzellen je Leitbündel, BSC = Bündelscheidenzellen je Leitbündel, die im direk-ten Kontakt zum Xylem stehen.
Mean sd se nControl BS 5,49 1,08 0,14 64
BSC 2,38 0,55 0,07 64X 1,41 0,50 0,06 64
BSC/BS 0,46 0,13 0,02 64Patch-Clamp MB-2/
(MB-1+MB-2)0,43 0,08 0,03 145
Wie in Abbildung 22 (Kapitel 5) zu sehen ist, haben Leitbündel mit 5 Bündelschei-denzellen eine bestimmte Größenverteilung: die erste und dritte Bündelscheidenzellesind groß, die anderen im Vergleich dazu klein. Nach obiger Auszählung trat die Frageauf, ob die Größenverteilung mit dem Xylemkontakt in Zusammenhang zu bringen ist.
Abbildung 41 zeigt, daß es in der Tat einen Unterschied in der Verteilung gibt. Zwargibt es in allen Zellgrößen Bündelscheidenzellen mit und ohne Kontakt, aber kleinereBündelscheidenzellen scheinen meist keinen Kontakt zu haben.
Die Größe der gepatchten Bündelscheidenprotoplasten (Abbildung 44) wurde aufzwei Arten bestimmt:
1. anhand der Kapazität (Umrechnung in Zellgröße über 1 µF/cm2 Zellfläche)2. über eine Skala im Okular.

81
Abbildung 41. Verteilung der Größen (gleitendes Mittel über 3) von Bündelscheidenzellen mitund ohne Zellkontakt, ausgezählt anhand von Schnitten (Oberfläche als Ellipsoid berechnet).Kleinere Zellen haben zumeist keinen Kontakt.
Abbildung 42. Zellkapazitäten von gepatchten Bündelscheidenprotoplasten (A) und Meso-phyllprotoplasten (B) aufgetragen über der Protoplastenoberfläche (Radius optisch bestimmt).Für beide Zelltypen ist die Kapazität pro Fläche kleiner als 1 µF/cm2 (Linie Cm = Ogem, Bündel-scheidenprotoplasten: 0,699 ± 0,016 µF/cm2, n = 141, Mesophyllprotoplasten: 0,678 ± 0,017µF/cm2, n = 105).
0 10 20 30 400
10
20
30
40
50
A
C m = O gem
Cm (p
F)
Ogem (100 µm2)0 10 20 30 40 50
0
10
20
30
40
B
C m = O ge
m
Ogem / 100 µm2
C m /
pF
0 2000 4000 6000 80000
10
20
30
40
ohne Kontakt
mit Kontakt
n
Oberfläche / µm2
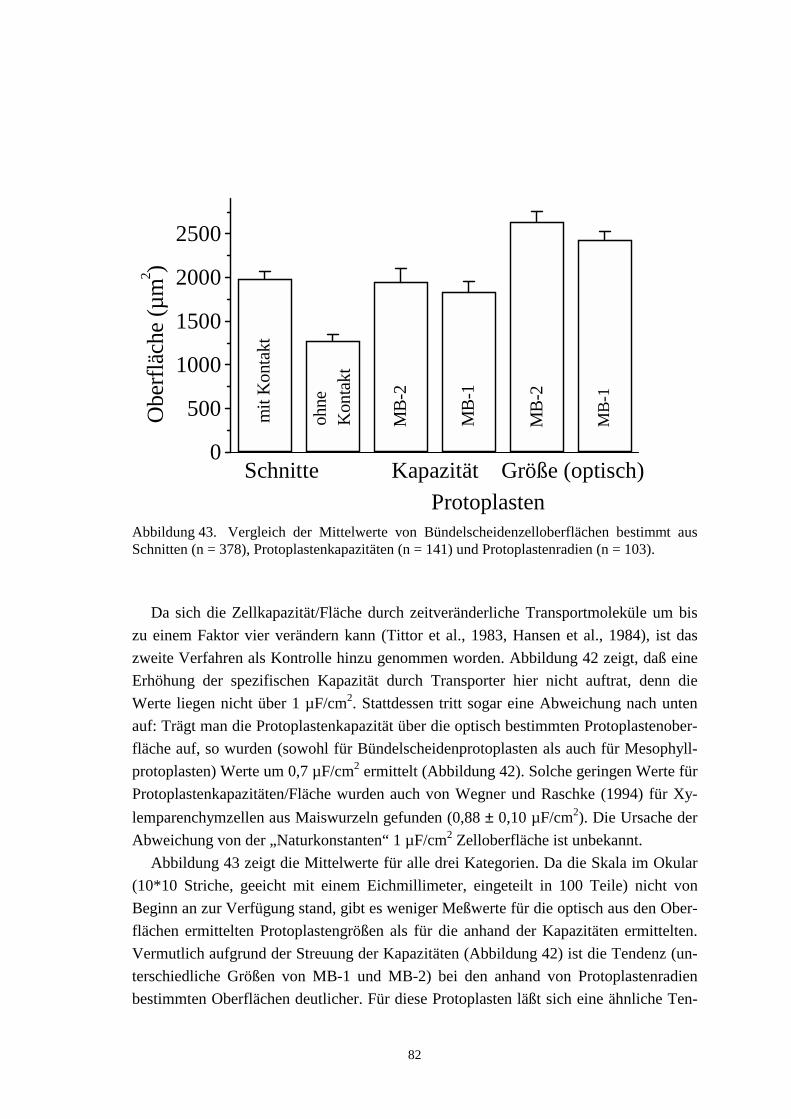
82
Abbildung 43. Vergleich der Mittelwerte von Bündelscheidenzelloberflächen bestimmt ausSchnitten (n = 378), Protoplastenkapazitäten (n = 141) und Protoplastenradien (n = 103).
Da sich die Zellkapazität/Fläche durch zeitveränderliche Transportmoleküle um biszu einem Faktor vier verändern kann (Tittor et al., 1983, Hansen et al., 1984), ist daszweite Verfahren als Kontrolle hinzu genommen worden. Abbildung 42 zeigt, daß eineErhöhung der spezifischen Kapazität durch Transporter hier nicht auftrat, denn dieWerte liegen nicht über 1 µF/cm2. Stattdessen tritt sogar eine Abweichung nach untenauf: Trägt man die Protoplastenkapazität über die optisch bestimmten Protoplastenober-fläche auf, so wurden (sowohl für Bündelscheidenprotoplasten als auch für Mesophyll-protoplasten) Werte um 0,7 µF/cm2 ermittelt (Abbildung 42). Solche geringen Werte fürProtoplastenkapazitäten/Fläche wurden auch von Wegner und Raschke (1994) für Xy-lemparenchymzellen aus Maiswurzeln gefunden (0,88 ± 0,10 µF/cm2). Die Ursache derAbweichung von der „Naturkonstanten“ 1 µF/cm2 Zelloberfläche ist unbekannt.
Abbildung 43 zeigt die Mittelwerte für alle drei Kategorien. Da die Skala im Okular(10*10 Striche, geeicht mit einem Eichmillimeter, eingeteilt in 100 Teile) nicht vonBeginn an zur Verfügung stand, gibt es weniger Meßwerte für die optisch aus den Ober-flächen ermittelten Protoplastengrößen als für die anhand der Kapazitäten ermittelten.Vermutlich aufgrund der Streuung der Kapazitäten (Abbildung 42) ist die Tendenz (un-terschiedliche Größen von MB-1 und MB-2) bei den anhand von Protoplastenradienbestimmten Oberflächen deutlicher. Für diese Protoplasten läßt sich eine ähnliche Ten-
0
500
1000
1500
2000
2500
MB-
1
MB-
1
ohne
K
onta
kt
MB-
2
MB-
2
mit
Kon
takt
Schnitte Kapazität Größe (optisch) Protoplasten
Obe
rfläc
he (µ
m2 )

83
denz in der Verteilung (kleinere Protoplasten ohne Xylemkontakt) ablesen wie für dieBündelscheidenzellen (Abbildung 44). Insgesamt sind leider noch zu wenig Meßwertevorhanden, um eine signifikante Aussage zu erhalten, die Tendenz läßt sich aber erken-nen.
Beide Ergebnisse (Tabelle 9 und Abbildung 43) unterstützen die Hypothese, daß derMB-2 in Bündelscheidenzellen mit direktem Kontakt auftritt. Die dort vermutete höhereTransportaktivität verlangt die höhere Leitfähigkeit dieses Kanals. Allerdings müßtenfür einen endgültigen Beweis überzeugendere Experimente als diese Auszählung durch-geführt werden.
Trotz der zur Zeit noch unsicheren experimentellen Unterstützung für die Hypotheseder Zuordnung von MB-1 und MB-2 zu Bündelscheidenzellen ohne und mit Kontaktzum Xylem, ist Tabelle 9 als Ausgangsbasis für eine sinnvolle Arbeitshypothese zu be-trachten. Die Gutachter von PLANTA hatten keine Probleme, diese Hypothese zu ak-zeptieren (Keunecke und Hansen, 1999).
Abbildung 44. Verteilung der Größen von Bündelscheidenprotoplasten mit und ohne zeitakti-viertes Verhalten (nach der Hypothese also MB-2 mit und MB-1 ohne Xylemkontakt) ermitteltmit Hilfe des Okularmaßstabs im Mikroskop (n = Anzahl, gleitendes Mittel über 5).
0 2000 4000 60000
2
4
6
8
MB-2
MB-1
n
Ogem(µm2)

84
8.5 Na+-Whole-Cell-Messungen
Bei den Einzelkanalmessungen zeigte sich, daß Na+ vielfältig auf die Kanäle wirkenkann. Gefunden wurde sowohl eine Blockierung (Verringerung der Offenwahrschein-lichkeit) beim SC (Keunecke et al., 1997) in K+/Na+ Mischlösungen als auch eine höhe-re Leitfähigkeit in reiner Na+-Lösung beim IR (Abschnitt 7.6).
Beide Effekte sind für Pflanzen im Salzstress wichtig. Bei guter K+-Versorgung senktNa+ die K+-Leitfähigkeit, bei K+ Mangel hingegen kann Na+ durch den K+-Kanal treten.Dies mag zu einer besonders bedrohlichen Situation führen, wenn Salzstress mit K+-Mangel einhergeht.
Diese Einzelkanalmessungen fanden allerdings unter unphysiologisch hohen Ionen-konzentrationen statt. Whole-Cell-Messungen ermöglichen eher den Einsatz physiolo-gisch sinnvoller Lösungen. In den in Abbildung 45 gezeigten Whole-Cell-Messungenwurden im Bad Ionenkonzentrationen im 10 mM-Bereich benutzt: Dies entspricht denVerhältnissen, die von der AG Sattelmacher im Apoplasten gefunden wurden (Penne-wiß et al., 1997). 10 mM KCl als Badlösung (mit 2 mM ATP in der Pipette) ergaben dieauch in den anderen Abbildungen gezeigte Stromspannungskurve (Abbildung 37): Beibeiden Typen (MB-1 und MB-2) veränderte sich der Einwärtsstrom bei Anwesenheitvon 10 mM Na+ nicht (negative Spannung). Der Auswärtsstrom wurde reduziert, beimMB-2 war der Effekt sehr viel stärker. Das Umkehrpotential (Spannung für I = 0) ver-ändert sich nicht, ein Zeichen, daß Na+ beim Na+-Block für K+ den Kanal nicht permei-ert. Die beobachtete Stromreduzierung kann deshalb nur auf Gatingverhalten zurückge-führt werden. Bei Anwesenheit von Na+ erhöht sich die Verweildauer in den geschlos-senen Zuständen relativ zu der in den offenen Zuständen (Na+-Block). Na+ wirkt hierbeivon der Badseite auf den Auswärtsstrom, d.h. entgegen der Flußrichtung. Dies sprichtdafür, daß auf der Außenseite eine Bindestelle für Na+ ist.

85
Abbildung 45. Na+-Effekt auf den MB-1 (A) und den MB-2 (B). Bei Zugabe von Na+ zurBadlösung zeigte sich ein Effekt auf den Auswärtsstrom. Die Reduktion der Leitfähigkeit desMB-2 ist stärker als beim MB-1. Lösungen „blau“ und „rot“ (MB-1: n = 10, MB-2: n= 8) ausTabelle 8 bzw. 10 mM NaCl + 10 mM KCl + 5 mM CaCl2 im Bad (MB-1: n = 6, MB-2: n = 3).
-150 -100 -50 0 50
-2
0
2
4
A
MB-1
+Na+
K+
I (µA
cm
-2)
V (mV)
-100 -50 0 50-8
0
8
16
B
MB-2
+Na+
K+
I (µA
cm
-2)
V (mV)

86
8.6 Rb+-Effekt auf Bündelscheidenzellen
Die obige Hypothese, daß die instantanen Anteile von MB-1 und MB-2 zu verschiede-nen Kanaltypen gehören, wird durch die folgenden Messungen mit Rb+ bestätigt. Beiden Untersuchungen zum Weg der Ionen vom Xylem in den Symplasten war Rb+ alsTracer für K+ genommen worden. Obwohl dies eine weit verbreitete Praxis ist(MacRobbie, 1995), entfällt nicht die Notwendigkeit des Nachweises, daß Rb+ tatsäch-lich durch die K+-Kanäle aufgenommen wird, zumal Hedrich et al. (1995) einen Kanalgefunden haben, der Rb+ nicht durchtreten läßt.
Whole-Cell-Messungen von Bündelscheidenprotoplasten in Lösung „R(braun)“ (mitRb+) oder Lösung „braun“ (mit K+) im Bad und Lösung „blau“ (mit 2 mM MgATP, alleaus Tabelle 8) in der Pipette ergaben die Stromspannungskurven in Abbildung 46. DieAufnahmen wurden mit dem in Abbildung 36 A beschriebenen Protokoll durchgeführt.
Die Resultate zeigen die Vielfalt im Verhalten der verschiedenen Zelltypen. InAbbildung 46 A ist in der Außenlösung nur mit Rb+ das Umkehrpotential positiv. Diesist eine Verlagerung des Umkehrpotentials (von –61 ± 7 mV auf +21 ± 6 mV) desWhole-Cell-Stromes in Richtung des Nernstpotentials für Rb+. ERb liegt sehr weit imPositiven, da in der Zelle bis auf geringe Konzentrationen durch Anreicherung aufgrunddes Einstromes kein Rb+ vorhanden ist. Alternativ könnte die Verschiebung auch auf ECl
zurückzuführen sein. Messungen mit 100 mM RbCl im Bad zeigen aber eine weitereVerschiebung zu positiveren Potentialen. Dies kann nur auf Rb+ zurückzuführen sein, daChlorid auf beiden Seiten gleich vorliegt (Nernstpotential: ECl
- = 0 mV).Ganz anders verhält es sich mit dem MB-2 (Abbildung 46 B): hier bleibt das Um-
kehrpotential nahezu unverändert (-36 ± 3 mV in Kaliumlösung, -31 ± 3 mV in Rubidi-umlösung), d.h. die beiden Ionenarten haben etwa die gleichen Einzelkanalleitfähigkei-ten (Abschnitt 4.2, Goldman Gleichung). Allerdings ist der Whole-Cell-Strom in Ge-genwart von Rb+ größer. Das bedeutet, daß Rb+ das Verhältnis von Offen- zu Geschlos-senzeiten erhöht.
Spaltet man den MB-2 in den instantanen und den zeitabhängigen Strom auf, so siehtman, daß der instantane für die bessere Whole-Cell-Leitfähigkeit von Rb+ durch denMB-2 verantwortlich ist (Abbildung 47): Die Whole-Cell-Leitfähigkeit von Rb+ und K+
durch den MB-AOR ist identisch.
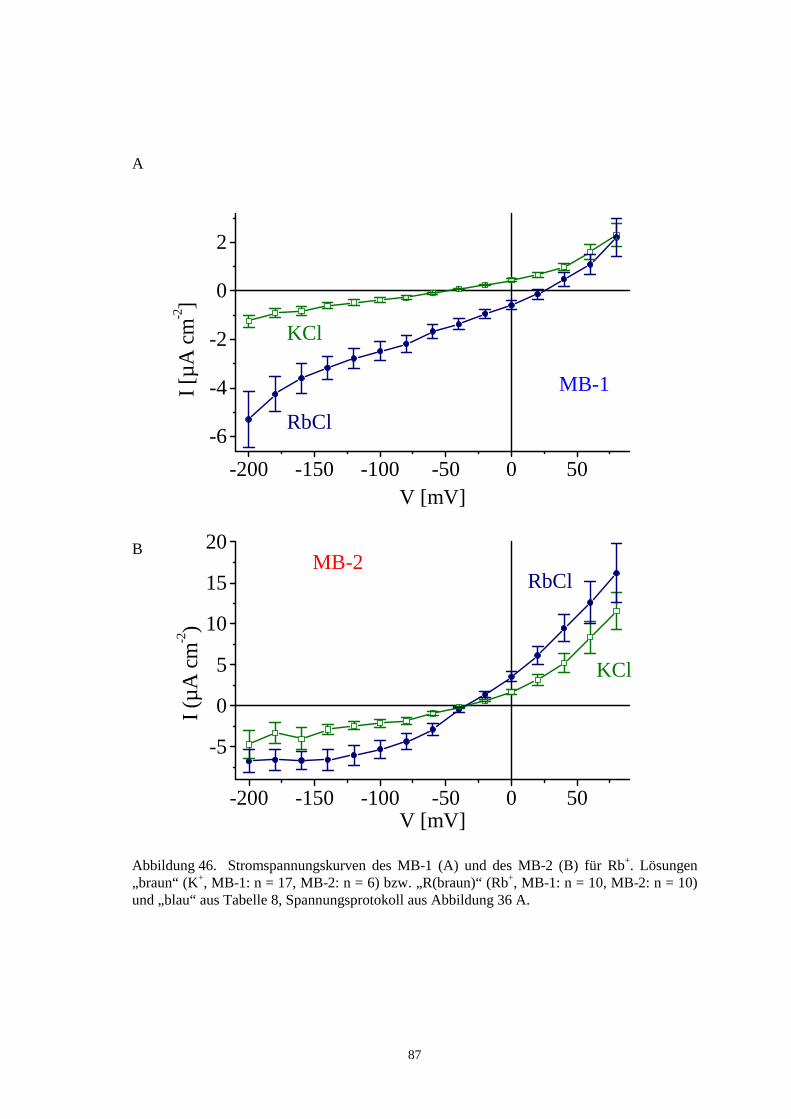
87
A
B
Abbildung 46. Stromspannungskurven des MB-1 (A) und des MB-2 (B) für Rb+. Lösungen„braun“ (K+, MB-1: n = 17, MB-2: n = 6) bzw. „R(braun)“ (Rb+, MB-1: n = 10, MB-2: n = 10)und „blau“ aus Tabelle 8, Spannungsprotokoll aus Abbildung 36 A.
-200 -150 -100 -50 0 50
-5
0
5
10
15
20MB-2
RbCl
KCl
I (µA
cm
-2)
V [mV]
-200 -150 -100 -50 0 50-6
-4
-2
0
2
MB-1
KCl
RbCl
I [µA
cm
-2]
V [mV]

88
A
B
Abbildung 47. Aufspaltung des Whole-Cell-Stromes vom MB-2 in die instantane (A) und diezeitabhängige Komponente (B). Während der MB-KI2 die Whole-Cell-Leitfähigkeit für Rb+
des MB-2 widerspiegelt, ist der MB-AOR in seinem Leitfähigkeitsverhalten für K+ und Rb+
nicht zu unterscheiden.
-200 -150 -100 -50 0 50-10
-5
0
5
10
15MB-KI2 RbCl
KCl
I [µA
cm
-2]
V [mV]
-200 -150 -100 -50 0 50
0
2
4
6 MB-AOR
RbCl
KCl
I (µA
cm
-2)
V (mV)

89
Der Anstieg im Bereich <-140 mV in Abbildung 47 B ist artifiziell: da dieHyperpolarisationsspannung nach dem Protokoll aus Abbildung 36 bei – 140 mV lag(um die Zellen nicht zu stark zu belasten), dort aber noch nicht die minimaleOffenwahrscheinlichkeit erreicht war, waren bei Beginn der Messung noch zu vieleTransporter offen, die dann nach und nach schlossen. Somit war der instantangemessene Strom kleiner als der am Ende der Messung, sa daß die Differenz einenpositiven Wert ergab. Läge die Vorspannung bei –200 mV, wäre der Effektverschwunden. Dies bedeutet, daß die Trennung zwischen MB-KI2 und MB-AOR nichtsauber ist, denn ein Einzelkanalstrom kann der Spannung nicht entgegenlaufen(negativer Sehnenwiderstand). Der Endwert der Testspannung ist unabhängig von derVorspannung (selbst bei positiver Testspannung von 40 mV wurden die gleichen Werteerreicht).
Der MB-1 verhält sich genau entgegengesetzt zum CKC1in (coleoptile K+ channelinward rectifier) aus Zea mays Koleoptilen (Hedrich et al., 1995), der Rb+ nicht durch-läßt. Da beide aus Maispflanzen stammen, zeigt sich die große genetische Variation vonK+-Kanälen in einer Pflanze.
Die Rechtfertigung für die Benutzung von Rb+ als Tracer für K+ in den Messungenmit LAMMA (Kapitel 3) ist dennoch gegeben: Es gibt in den Bündelscheidenzellen(insbesondere bei MB-2, die vermutlich ans Xylem angrenzen) Kanäle, die Rb+ durch-lassen. Insbesondere hat MB-1 (vermutlich) zwar keinen Xylemkontakt, nimmt aberRb+ deutlich besser auf als MB-2 (mit Xylemkontakt). Das verleiht dem Ergebnis, daßdie Aufnahme in die Bündelscheidenzellen (über MB-2) noch schneller als in das vas-kuläre Parenchym geschieht, besondere Aussagekraft.

90
8.7 pH-Effekt bei Bündelscheidenzellen
Veränderungen des pH-Wertes haben vielfältigen Einfluß auf die Pflanze, und es gibtviele Faktoren, die den pH-Wert beeinflussen. So treten räumliche pH-Gradienten in-nerhalb des Blattes (Canny, 1988, 1990 a, b) und um die Stomata (Grignon und Sen-tenac, 1991) auf. Ursache zeitlicher pH-Veränderungen sind zum Beispiel Photosynthe-se (Mühling et al., 1995, Plieth et al., 1998) oder Trockenheit (Gollan et al., 1992,Schurr und Schulze, 1996). Daher wurde der pH-Wert-Einfluß auf Kanäle relativ häufiguntersucht (Übersicht in Tabelle 14). Trotzdem beschränken sich die verfügbaren Datenvon Blättern nahezu ausschließlich auf Schließzellen oder Vakuolen (58 von 84 Unter-suchungen, Kapitel 1)
Somit besteht ein großer Bedarf für pH-Messungen an Bündelscheidenzellen, denngerade in ihren Arbeitsbereich, der Grenze zu den Xylemelementen (Kapitel 3), fandCanny (1987, 1988, 1990a, b) eine starke Ansäuerung. Er vermutete einen Zusammen-hang zwischen der Aufnahme von Substanzen und H+-getriebenem Cotransport.
Abbildung 48. pH-Effekt bei Bündelscheidenzellen des MB-2. Ein- und Auswärtsströme sindverändert. Verwendete Lösungen: „+(rot)“, „rot-braun“, „braun“ und „blau“ aus Tabelle 8. n =11 bei pH 6.15, n = 8 bei pH 6.5, n = 11 bei pH 7.2.
-160 -80 0 80
0
20
pH7.26.56.15
MB-2
V (mV)

91
A
B
Abbildung 49. pH Effekt aufgespalten nach instantanem (MB-KI2, A) und zeitabhängigemAnteil (MB-AOR, B). Die Richtung des pH-Effektes ist bei beiden gleich.
-160 -80 0 80
-20
0
20I (
µA c
m-2)
pH6.156.57.2
MB-KI2
V (mV)
-160 -80 0 80
0
4
8MB-AOR 6.15
6.5
7.2
I (µA
cm
-2)
V (mV)

92
Bei Bündelscheidenzellen (und zum Vergleich auch bei Mesophyllzellen, Kapitel 9)wurden mit dem in Abbildung 36 A gezeigten Protokoll Strom-Spannungskurven auf-genommen. Die extreme Fragilität der Whole-Cell-Präparate verhinderte allerdings, daßLösungswechsel durchgeführt werden konnten. Deshalb mußte für jeden pH-Wert einneuer Protoplast genommen werden, so daß sich eine größere Streuung ergab. DieseStreuung konnte vermindert werden, indem der Strom auf die Protoplastenoberflächenormiert wurde. Dennoch wurden 5 bis 15 (im Mittel 10) Messungen pro pH-Wertdurchgeführt, um signifikante Ergebnisse zu erhalten.
Abbildung 48 zeigt Messungen am MB-2 von Bündelscheidenzellen, identifiziertdurch die zeitabhängige Aktivierung (Abbildung 36). Ein pH-Effekt tritt in gleicherRichtung sowohl bei Auswärts- als auch bei Einwärtsströmen auf. In beiden Bereichenführt eine Ansäuerung zu einer Zunahme des Ein- und Auswärtsstromes.
Eine Aufspaltung der Stromantwort in einen instantanen und einen zeitabhängigenAnteil (direkt zu Beginn und zum Ende der jeweiligen Spannung) zeigt Abbildung 49.Der zeitabhängige MB-AOR ist nur bei positiven Spannungen zu erkennen (Abbildung49 B), zeigt aber die gleiche pH-Abhängigkeit wie der MB-KI2 (Abbildung 49 A).
Abbildung 50. pH Effekt auf den MB-1. Der pH-Wert hat keinen so großen Einfluß wie beimMB-2, aber die Reihenfolge der Veränderung im negativen Spannungsbereich stimmt überein.Lösungen: „rot“, „+(rot)“, „rot-braun“, „braun“ und „blau“ aus Tabelle 8 n = 5 bei pH 5.8, n =13 bei pH 6.15, n = 10 bei pH 6.5, n = 15 bei pH 7.2.
-160 -80 0 80
-2
0
2
4MB-1
I (µA
cm
-2)
pH 7.26.5 6.155.8
V (mV)

93
Der MB-1 (identifiziert durch die instantane Stromantwort, Abbildung 36) in Bün-delscheidenzellen (Abbildung 50) verhält sich ähnlich wie der MB-2 in Abbildung 48.Allerdings ist der pH-Effekt beim MB-2, bei dem die Ströme insgesamt größer sind,wesentlich deutlicher (Abbildung 48). Dennoch ist die Tendenz trotz des geringen Ef-fektes im negativen Bereich ersichtlich (Abbildung 50).
Auf die Bedeutung der pH-Abhängigkeit für die Rolle der Zellen im Ferntransportwird im Kapitel 10 noch näher eingegangen.

94
9 Whole-Cell-Messungen an Mesophyllzellen
Bündelscheidenzellen sind der Ort des ersten Übertritts der Nährlösung vom Xylem inden Symplasten. Von ihnen geht es weiter über die hier sehr zahlreichen Plasmodesmen(Abschnitt 2.4.1, Evert et al., 1977) in die Mesophyllzellen. Die Untersuchung derMesophyllzellen dient verschiedenen Zwecken. Zum einen werden durch den Vergleichzwischen den Transporteigenschaften von Bündelscheiden- und Mesophyllzellen dieBesonderheiten der Bündelscheidenzellen deutlich, und aus dem Unterschied ergebensich Hinweise auf die Anpassung an die physiologische Funktion. Zum anderen sind dieMesophyllzellen an sich auch von Interesse, denn die Apoplasten von Mesophyll- undBündelscheidenzellen sind durch die Suberinlamelle, die die Bündelscheide umgibt,quasi getrennt (Hattersley and Browing, 1981, Botha, et al. 1982, Canny 1995, Evert etal. 1977, 1985, 1996). Durch die Mesophyllzellen kann also der Nährstoffstrom in denBlattapoplasten hinter der Suberinschicht gelangen.
Abbildung 51. Whole-Cell-Ströme einer Mesophyllzelle. Überlagerung der vom Spannungs-protokoll in Abbildung 36 A hervorgerufenen Ströme. Deutlich zu erkennen ist, daß es sichhierbei um einen Auswärtsgleichrichter handelt. Der Strom setzt sich zusammen aus eineminstantanen und einem zeitabhängigen Anteil. Analog zu den Bezeichnungen bei den Bündel-scheidenzellen werden diese Anteile MM-KI2 und MM-AOR genannt (gesamte Antwort: MM-2). Lösungen „braun“ und „blau“ aus Tabelle 8.

95
Abbildung 51 zeigt Whole-Cell-Antworten von Mesophyllzellen. Es trat bei diesenMessungen nur ein Typ auf, der dem MB-2 (Mais Bündelscheidenzellen Typ 2, Kapitel8) entspricht. Daher haben wir den gefundenen Kanal MM-2 genannt (Maize MesophyllTyp 2).
Abbildung 52. Stromspannungskurven für Mesophyllzellen. Die Antwort am Ende derangelegten Testspannung (MM-2) und ihre Komponente (MM-KI2) haben ein Umkehrpotential(-41± 3 mV, -42 ± 3 mV), das in der Nähe der K+-Nernstspannung (-56 mV) liegt (für MM-AOR läßt es sich nicht bestimmen, kann aber nicht stark von den anderen abweichen, da ja dievon MM-2 und MM-KI2 sonst nicht gleich sein könnten). Die positiven Ströme des MM-AORbei negativen Werten liegen wieder an der Testspannung. Lösung “braun” und “blau” ausTabelle 8.
Die Stromspannungskurven sind in Abbildung 52 dargestellt. Wie bereits inAbbildung 51 zu erkennen, besteht die Whole-Cell-Antwort aus einem instantanen(MM-KI2, Mais Mesophyll K+ Kanal Typ 2) und einem zeitabhängigen Anteil (MM-AOR, Mais Mesophyll aktivierter Auswärtsgleichrichter). Der Gesamtstrom und derinstantane Anteil haben beide ein Umkehrpotential (Schnittpunkt mit derSpannungsachse), das bei EK liegt, es ist also ein K+-Kanal.
Die kleinen positiven Ströme des MM-AOR für Spannungen <-120 mV resultierenaus der Wahl der Vorspannung (Spannungsprotokoll aus Abbildung 36 A). Es gilt dasgleiche wie schon für Abbildung 47 und 49: Bei Spannungen, die negativer als die
-200 -150 -100 -50 0 50-5
0
5
10
15
MM-KI2
MM-AOR
MM-2
I (µA
cm
-2)
V (mV)
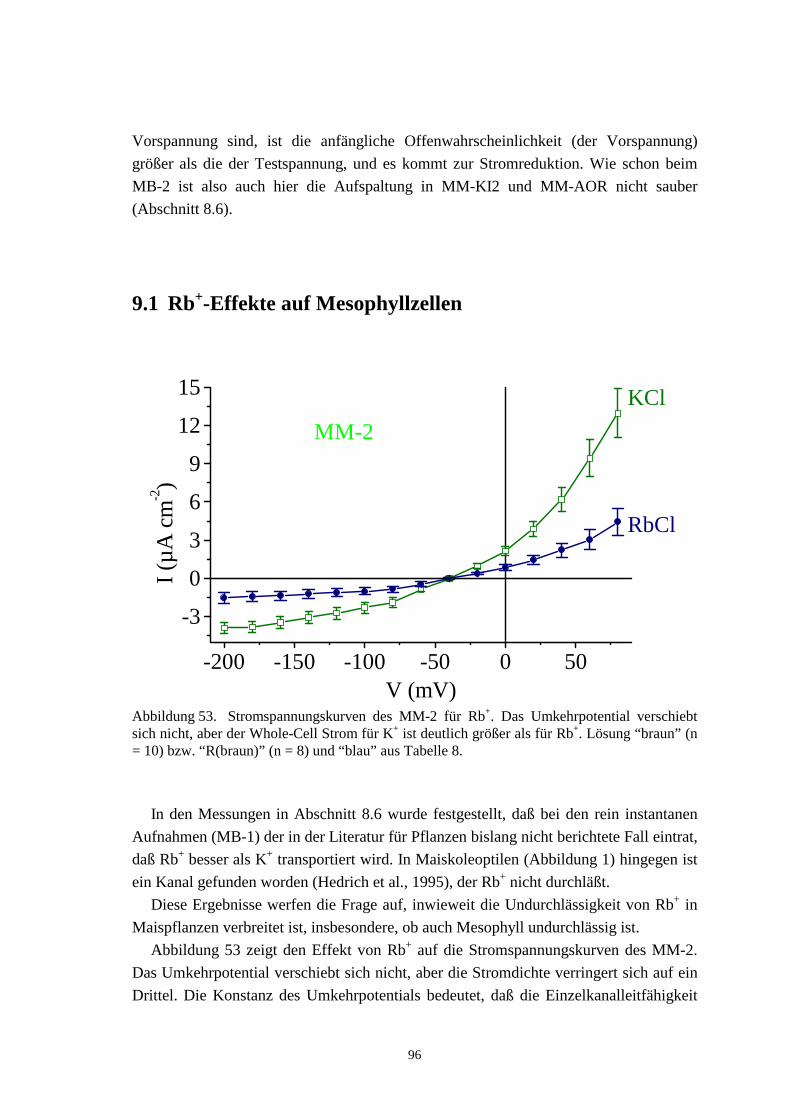
96
Vorspannung sind, ist die anfängliche Offenwahrscheinlichkeit (der Vorspannung)größer als die der Testspannung, und es kommt zur Stromreduktion. Wie schon beimMB-2 ist also auch hier die Aufspaltung in MM-KI2 und MM-AOR nicht sauber(Abschnitt 8.6).
9.1 Rb+-Effekte auf Mesophyllzellen
Abbildung 53. Stromspannungskurven des MM-2 für Rb+. Das Umkehrpotential verschiebtsich nicht, aber der Whole-Cell Strom für K+ ist deutlich größer als für Rb+. Lösung “braun” (n= 10) bzw. “R(braun)” (n = 8) und “blau” aus Tabelle 8.
In den Messungen in Abschnitt 8.6 wurde festgestellt, daß bei den rein instantanenAufnahmen (MB-1) der in der Literatur für Pflanzen bislang nicht berichtete Fall eintrat,daß Rb+ besser als K+ transportiert wird. In Maiskoleoptilen (Abbildung 1) hingegen istein Kanal gefunden worden (Hedrich et al., 1995), der Rb+ nicht durchläßt.
Diese Ergebnisse werfen die Frage auf, inwieweit die Undurchlässigkeit von Rb+ inMaispflanzen verbreitet ist, insbesondere, ob auch Mesophyll undurchlässig ist.
Abbildung 53 zeigt den Effekt von Rb+ auf die Stromspannungskurven des MM-2.Das Umkehrpotential verschiebt sich nicht, aber die Stromdichte verringert sich auf einDrittel. Die Konstanz des Umkehrpotentials bedeutet, daß die Einzelkanalleitfähigkeit
-200 -150 -100 -50 0 50
-3
0
3
6
9
12
15
MM-2KCl
RbCl
I (µA
cm
-2)
V (mV)

97
für K+ und Rb+ gleich ist, aber die Verringerung des Stromes bedeutet, daß sich das Ak-tivierungsverhalten (Gating) unterscheidet. Die Stromverminderung in Abbildung 53 istdeshalb damit zu erklären, daß das Verhältnis von Offen- zu Geschlossenwahrschein-lichkeit bei Rb+ sinkt.
Die Aufspaltung in den instantanen und den zeitabhängigen Anteil (Abbildung 54)zeigt einen weiteren Unterschied zum MB-2: Während beim MB-2 die schlechtereDurchlässigkeit für Rb+ allein auf den instantanen Anteil zurückgeführt werden konnte,sind beim Mesophyll beide Komponenten beteiligt.
Es lassen sich somit alle drei Whole-Cell-Antworten (MB-1, MB-2 und MM-2) al-lein durch ihre unterschiedlichen Transporteigenschaften für K+ und Rb+ eindeutig un-terscheiden.
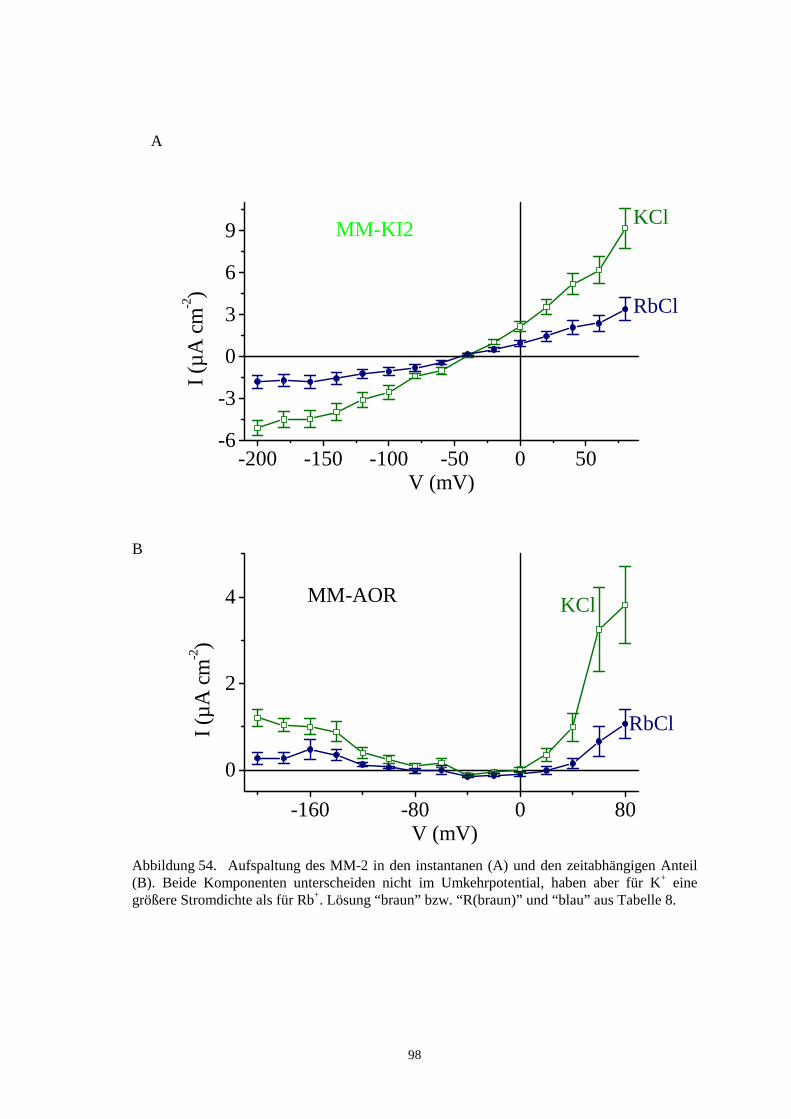
98
A
B
Abbildung 54. Aufspaltung des MM-2 in den instantanen (A) und den zeitabhängigen Anteil(B). Beide Komponenten unterscheiden nicht im Umkehrpotential, haben aber für K+ einegrößere Stromdichte als für Rb+. Lösung “braun” bzw. “R(braun)” und “blau” aus Tabelle 8.
-200 -150 -100 -50 0 50-6
-3
0
3
6
9 MM-KI2 KCl
RbCl
I (µA
cm
-2)
V (mV)
-160 -80 0 80
0
2
4 MM-AOR KCl
RbClI (µA
cm
-2)
V (mV)
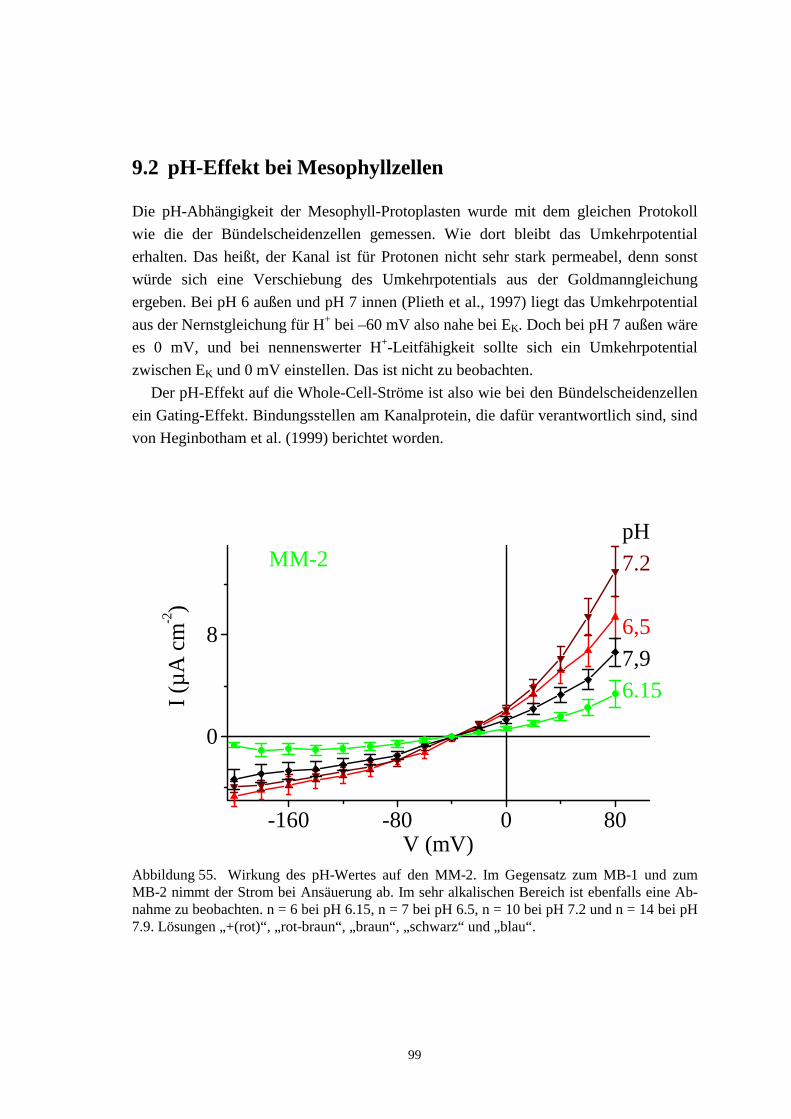
99
9.2 pH-Effekt bei Mesophyllzellen
Die pH-Abhängigkeit der Mesophyll-Protoplasten wurde mit dem gleichen Protokollwie die der Bündelscheidenzellen gemessen. Wie dort bleibt das Umkehrpotentialerhalten. Das heißt, der Kanal ist für Protonen nicht sehr stark permeabel, denn sonstwürde sich eine Verschiebung des Umkehrpotentials aus der Goldmanngleichungergeben. Bei pH 6 außen und pH 7 innen (Plieth et al., 1997) liegt das Umkehrpotentialaus der Nernstgleichung für H+ bei –60 mV also nahe bei EK. Doch bei pH 7 außen wärees 0 mV, und bei nennenswerter H+-Leitfähigkeit sollte sich ein Umkehrpotentialzwischen EK und 0 mV einstellen. Das ist nicht zu beobachten.
Der pH-Effekt auf die Whole-Cell-Ströme ist also wie bei den Bündelscheidenzellenein Gating-Effekt. Bindungsstellen am Kanalprotein, die dafür verantwortlich sind, sindvon Heginbotham et al. (1999) berichtet worden.
Abbildung 55. Wirkung des pH-Wertes auf den MM-2. Im Gegensatz zum MB-1 und zumMB-2 nimmt der Strom bei Ansäuerung ab. Im sehr alkalischen Bereich ist ebenfalls eine Ab-nahme zu beobachten. n = 6 bei pH 6.15, n = 7 bei pH 6.5, n = 10 bei pH 7.2 und n = 14 bei pH7.9. Lösungen „+(rot)“, „rot-braun“, „braun“, „schwarz“ und „blau“.
-160 -80 0 80
0
8
MM-2pH7.2
6,57,96.15I (
µA c
m-2)
V (mV)

100
Abbildung 56. Maximum im Whole-Cell-Strom für den Auswärtsgleichrichter MM-AOR. DieStröme wurden aus Abbildung 55 bei V= + 80 mV entnommen.
Die pH-Wert-Abhängigkeit in Abbildung 55 unterscheidet sich grundsätzlich von derbei den Bündelscheidenzellen gefundenen (Abschnitt 8.7). Wird vom physiologisch zuerwartenden leicht sauren Bereich in den alkalischen Bereich gewechselt, so nimmt hierder Strom bis zum neutralen Bereich hin zu. Anschließend nimmt der Strom wiederleicht ab. Das Durchlaufen eines Maximums ist für den Strom bei der festen Spannungvon +80 mV in Abbildung 56 noch einmal separat dargestellt.
Eine Aufspaltung in den instantanen (MM-KI2, Mais Mesophyll K+ instantaner Typ2, Abbildung 57 A) und zeitabhängigen Teil (MM-AOR, Mais Mesophyll aktivierterAuswärtsgleichrichter, Abbildung 57 B) liefert auch hier die gleiche Abhängigkeit wiefür den gesamten Strom des MM-2 (Abbildung 55). Wie beim MB-AOR ist auch hierein artifizieller Anstieg im Bereich <-140 mV in Abbildung 57 B zu sehen, der aufunvollständiges Schließen bei der Vorspannung von –140 mV zurückzuführen ist.
6 7 80
5
10
I (µA
cm
-2)
pH

101
A
B
Abbildung 57. Wirkung des pH-Wertes auf den instantanen Anteil des MM-2 (MM-KI2, A)und auf den zeitabhängigen Teil (MM-AOR, B). Wie schon bei den Rb+-Messungen verhaltensich beide Komponenten gleich.
-160 -80 0 80-6
0
6
7.26.57.9
MM-KI2I (
µA c
m-2)
V (mV)
-160 -80 0 80
0
2
4MM-AOR
7.26.57.9
I (µA
cm
-2)
V (mV)

102
10 DiskussionIn diesem Kapitel werden die Methoden und Ergebnisse von Einzelkanal- und Whole-Cell-Untersuchungen gegenübergestellt (Abschnitte 10.1 bis 10.5.). Die Ergebnissewerden dann mit den Ergebnissen anderer Autoren verglichen, die an Gerstenwurzelnund Maiskoleoptilen gemessen haben (Abschnitte 10.6 und 10.8).
10.1 Vergleich der Einzelkanal- und Whole-Cell-Messungen
Der hier erreichte Stand der Patch-Clamp-Technik an Maisblattzellen ermöglicht zweiArten von Messungen: Einzelkanal- und Whole-Cell. Allerdings wurde bei den Ein-zelkanalmessungen vorrangig mit sehr hohen Ionenkonzentrationen (100 – 200 mM)gemessen, da die Ströme bei Bündelscheidenzellen klein und das Schalten schnell war.Einzelkanalmessungen mit geringeren Konzentrationen, die den physiologischen Bedin-gungen eher entsprechen (Abschnitt 7.7), führten zu einer Meßlücke um das Umkehr-potential, da dort die Einzelkanalströme im Rauschen verschwanden. Dennoch war dieBestimmung des Umkehrpotentials selbst mit der Spannungsrampenmethode (Abschnitt7.4) möglich.
Whole-Cell-Messungen erbrachten in dieser Arbeit die interessanteren Ergebnisse.Sie haben den Vorteil (und den Nachteil, je nach Fragestellung), daß sie einen Überblicküber das Gesamtverhalten der Zelle liefern. Rauschen stellt hier kein Problem dar, unddie Effekte lassen sich unter physiologischen Bedingungen messen (z.B. Na+-Block,Abschnitt 8.5, pH-Effekt Abschnitte 8.7 und 9.2, Rb+-Effekt, Abschnitte 8.6 und 9.1).Eine Messung liefert bereits die gesamte Stromspannungskurve, während bei Einzelka-nalmessungen jeder Spannungswert einzeln aufgenommen werden muß. Zudem ist dasZeitverhalten auf den ersten Blick erfaßbar: MB-1 hat kein zeitaktiviertes Verhalten,MB-2 und MM-2 hingegen schon.
Warum würde es sich dennoch lohnen, die Einzelkanalmessungen weiterauszubauen? Gute Einzelkanalmessungen (das sind solche mit einen hohen Signal- zuRauschverhältnis, die zur Zeit bei diesem Objekt noch nicht erreichbar sind) könneninsbesondere biophysikalische Informationen liefern. So kann bei den Einzelkanalmes-sungen das Schalten genau einem Transporter (einem Kanal) zugeordnet werden (SCoder IR). Die Zeitreihe kann detailliertere Informationen über die Zeitabhängigkeitliefern als Whole-Cell-Messungen, die Trennung von Gating und Leitfähigkeitseffektenist sehr viel besser möglich, die Offenwahrscheinlichkeit ist aus der idealisiertenZeitreihe (Draber und Schultze, 1994a,b, Keunecke, 1995) direkt bestimmbar und die

103
Ratenkonstanten des Markov-Modells des Kanals mit mehreren Offen- undGeschlossenzuständen (Lühring, 1999, Farokhi et al., 1999) ergeben sich aus denVerweildauerhistogrammen (Blunck et al., 1998) oder anderen Fitverfahren (Albertsenund Hansen, 1994). Diese Zustände können dann mit neuesten molekularbiologischenErkenntnissen zusammengebracht werden (Nonner und Eisenberg, 1998, Nonner et al.,1998, Nonner et al., 1999, Immke et al., 1999, Ogielska und Aldrich, 1998, 1999).Leider war aber bei den Einzelkanaldaten von Mais das Signal- zu Rauschverhältnis füreine solche Analyse zu schlecht. Dennoch tauchten, wie bei den Na+-Daten noch zubesprechen, Ansatzpunkte für sehr aktuelle Fragestellungen auf (Abschnitt 10.7).
Das Herausfinden von Korrespondenzen zwischen den in Einzelkanal- und Whole-Cell-Messungen auftretenden Transportern, SC und IR auf der einen Seite und MB-1und MB-2 auf der anderen, scheitert an der geringen Datenmenge, die bis jetzt von denEinzelkanalmessungen zu erhalten war. Der Vergleich der Häufigkeit des Auftretenseinzelner Transporter, der z. B. bei Tabelle 9 eine große Rolle spielte, müßte sich aufdie Gegenüberstellung folgender Zahlen stützen: Auftreten von SC (92%) und IR (8%)verglichen mit dem von Whole-Cell-Typen MB-1 (57%) und MB-2 (43%). Dies läßtkeine Zuordnung zu, denn es muß bedacht werden, daß die Statistik sich dadurch ver-schiebt, daß der MB-2 zur Aktivierung ATP benötigt, das in den Einzelkanalmessungennicht eingesetzt wurde.
Ein anderer Test wäre die Konstruktion der Whole-Cell-Stromspannungskurven ausden IU-Kurven der Einzelkanäle gewichtet mit ihrer spannungsabhängigen Offenwahr-scheinlichkeit. Die IU-Kurven für den SC sind vorhanden, auch gibt es einige Informa-tionen über das Gating. Ein Vergleich bei + 100 und - 100 mV zeigte, daß hier die Of-fenwahrscheinlichkeiten des SC identisch sind, während die des IR für negative Span-nung größer ist. Für die Whole-Cell-Messungen kann man aus den Stromspannungskur-ven entnehmen, daß die Offenwahrscheinlichkeit abhängig von der Spannung ist: Insbe-sondere MB-2 und MM-2 haben ein „Gedächtnis“ an die Vorspannung (-140 mV) undbrauchen eine gewisse Zeit, bis sie ihren Strom an die Testspannung angepaßt haben.Einen zweiten Hinweis auf die Spannungsabhängigkeit der Offenwahrscheinlichkeitgeben die Stromspannungskurven aller Whole-Cell-Typen: Sie sind im Gegensatz zuden SC-Kurven nicht symmetrisch um das Umkehrpotential (keine Punktspiegelungmöglich). Eine gute Beschreibung der Offenwahrscheinlichkeit des Einzelkanals hätteallerdings einer Zeitreihenanalyse bedurft, für die das gegenwärtige Signal- zuRauschverhältnis nicht ausreicht.
Letztendlich hätte noch die Möglichkeit des Vergleiches der Empfindlichkeit gegen-über Ionen und Pharmaka herangezogen werden können. Die Rb+-, Na+- und pH-Messungen zeigen, daß dies bei Whole-Cell-Experimenten möglich ist, doch bei denEinzelkanalmessungen hätte der Zeitbedarf alle Grenzen gesprengt.

104
Somit wird die Diskussion im Folgenden sich hauptsächlich auf die Whole-Cell-Er-gebnisse stützen.
10.2 Vergleich von Bündelscheiden- und Mesophyllzellen
Ein direkter Vergleich der drei bei Mais aufgetretenen Whole-Cell-Antworten veran-schaulicht die sehr unterschiedlichen Effekte. In Abbildung 58 sind die IU-Kurven unterStandardbedingungen (pH = 7.2 in Lösung „braun“ und „blau“ aus Tabelle 8) darge-stellt. Die Stromspannungskurven der zeitabhängigen Antworten von Mesophyllzellen(MM-2) und Bündelscheidenzellen vom Typ 2 (MB-2) unterscheiden sich nicht, wederin der gesamten Antwort, noch in ihren Komponenten (MB-AOR und MM-AOR,Abbildung 58). Der rein instantane Typ von Bündelscheidenzellen (MB-1) hingegen hateine sehr viel geringere Stromdichte.
Abbildung 58. Vergleich der Whole-Cell-Ströme im K+-Medium für die drei Zelltypen MM-2,MB-KI1 und MB-2 sowie die zeitabhängigen Komponenten MB-AOR und MM-AOR. Lösung„braun“ und „blau“ aus Tabelle 8 (pH = 7,2).
-200 -150 -100 -50 0 50-6
0
6
12
MB-1
MB-2
MM-2
I (µA
cm-2)
V (mV)
-200 -150 -100 -50 0 50
0
2
4
6
MM-AOR
MB-AOR
I (µA
cm
-2)
V (mV)

105
10.3 Wirkung von Na+
Bei den Einzelkanalmessungen mit gemischten K+/Na+ Lösungen (SC, 100 mM KClbzw. 100 KCl + 100 NaCl im Bad, d.h. auf der cytosolischen Seite) trat eine Verminde-rung des Einzelkanalstroms auf (Abbildung 33). Diese Verminderung trat in Whole-Cell-Messungen sowohl beim MB-1 als auch beim MB-2 auf (Abbildung 45, 10 mMKCl bzw. 10 mM KCl + 10 mM NaCl im Bad, d.h. auf der apoplastischen Seite). Ob-wohl der Lösungswechsel auf unterschiedlichen Seiten stattfand (Einzelkanal: Cytosol,Whole-Cell: Apoplast), war der Effekt der gleiche: Einwärtsströme (negative Spannung)blieben unverändert, während Auswärtsströme abnahmen. Das spricht dafür, daß es so-wohl auf der dem Cytosol abgewandten Seite als auch auf der cytosolischen Seite eineBindestelle für Na+ gibt.
Allerdings sind Schlüsse von „makroskopischen“ Strömen (Whole-Cell) auf die Be-setzung von einzelnen Bindungsstellen und von Gating nicht so eindeutig wie bei Ein-zelkanalmessungen.
Die Messungen in reiner Natriumlösung zeigen noch einen besonderen Effekt, der inAbschnitt 10.7 diskutiert wird.
10.4 Einfluß von Rb+
Wie bereits in Abschnitt 9.1 bemerkt, ist Rb+ ein geeignetes Mittel, um alle drei Typen(MB-1, MB-2 und MM-2) zu unterscheiden. Deutlich wird dies in Abbildung 59. Durchden Ersatz von K+ durch Rb+ in der Badlösung unterscheiden sich die Ströme durch dieTransportertypen MB-2 und MM-2 etwa um den Faktor 4, während sie in Kaliumlösungidentisch waren.
Die Erhöhung der Stromdichte in der Rb+-Lösung ist für den MB-2 allein auf den in-stantanen Anteil zurückzuführen (Abbildung 47), bei der Verminderung im MM-2 sindbeide Komponenten beteiligt. Der MM-AOR erniedrigt seine Stromdichte beim Aus-tausch von K+ gegen Rb+, außerdem hat nun die Wahl der Vorspannung keinen so gro-ßen Einfluß mehr. Für negative Spannungen bleibt die Stromdichte etwa Null und steigtnicht, wie in Kaliumlösung, für Werte kleiner als die Vorspannung wieder deutlich an.
Die starke Verschiebung des Umkehrpotentials beim MB-1 durch Rb+ ist so aus derPflanzenwelt noch nicht bekannt. Einen Überblick über die Rubidiumpermeabilitätenbezogen auf K+ (berechnet aus der Verschiebung des Umkehrpotentials nach der Gold-mann-Gleichung, Abschnitt 4.2) geben Tabelle 10 und Tabelle 11 (Zusammenfassungvon Tabelle 10) für pflanzliche Membranen und Tabelle 12 und Tabelle 13 (Zusammen-
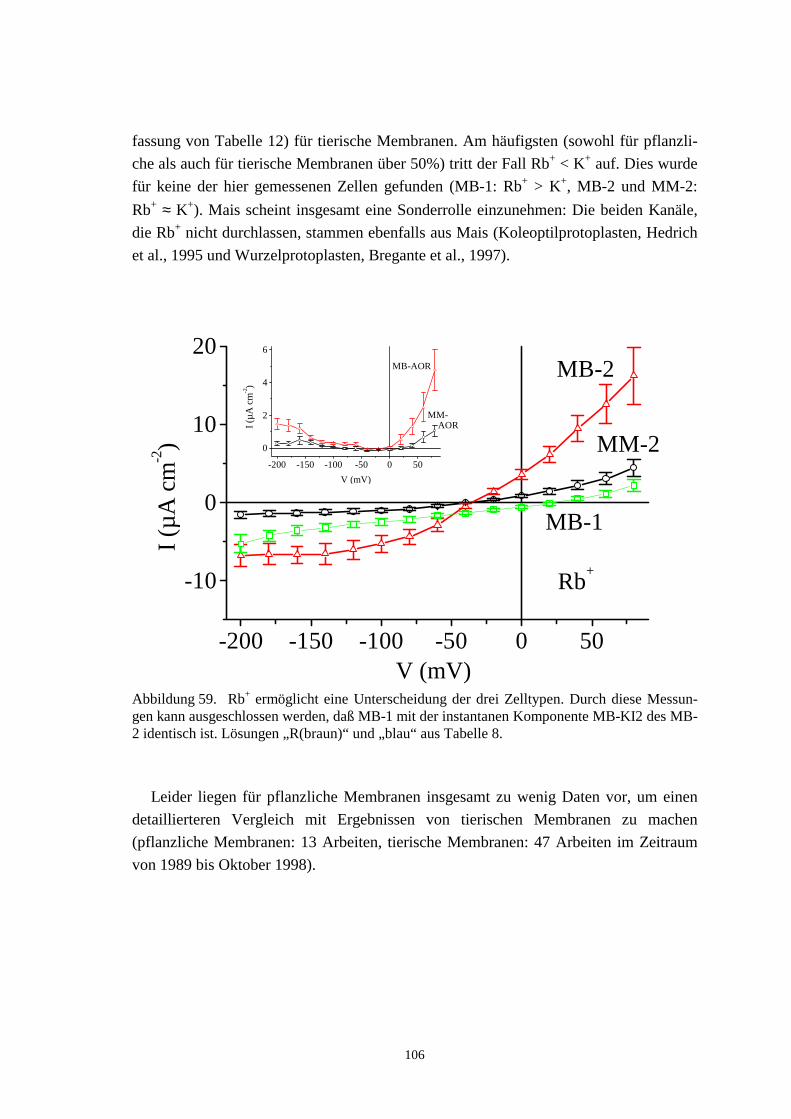
106
fassung von Tabelle 12) für tierische Membranen. Am häufigsten (sowohl für pflanzli-che als auch für tierische Membranen über 50%) tritt der Fall Rb+ < K+ auf. Dies wurdefür keine der hier gemessenen Zellen gefunden (MB-1: Rb+ > K+, MB-2 und MM-2:Rb+ ≈ K+). Mais scheint insgesamt eine Sonderrolle einzunehmen: Die beiden Kanäle,die Rb+ nicht durchlassen, stammen ebenfalls aus Mais (Koleoptilprotoplasten, Hedrichet al., 1995 und Wurzelprotoplasten, Bregante et al., 1997).
Abbildung 59. Rb+ ermöglicht eine Unterscheidung der drei Zelltypen. Durch diese Messun-gen kann ausgeschlossen werden, daß MB-1 mit der instantanen Komponente MB-KI2 des MB-2 identisch ist. Lösungen „R(braun)“ und „blau“ aus Tabelle 8.
Leider liegen für pflanzliche Membranen insgesamt zu wenig Daten vor, um einendetaillierteren Vergleich mit Ergebnissen von tierischen Membranen zu machen(pflanzliche Membranen: 13 Arbeiten, tierische Membranen: 47 Arbeiten im Zeitraumvon 1989 bis Oktober 1998).
-200 -150 -100 -50 0 50
-10
0
10
20
Rb+
MB-1
MM-2
MB-2
I (µA
cm
-2)
V (mV)
-200 -150 -100 -50 0 50
0
2
4
6
AORMM-
MB-AOR
I (µA
cm
-2)
V (mV)

107
Tabelle 10. Vergleich der Permeabilität von Rb+ und K+ bei Pflanzenzellen. Die beiden Kanäle,die Rb+ nicht durchlassen, stammen aus Maiswurzelprotoplasten (Bregante et al., 1997) undMaiskoleoptilprotoplasten (Hedrich et al., 1995). Daten ermittelt mit Winspirs für den Zeitraumvon 1989 bis Oktober 1998.
Sequenz Art desKanals
Zellart Quelle
K+>> Rb+ in Maiswurzelprotoplasten Bregante et al., 1997in Maiskoleoptilprotoplasten Hedrich et al., 1995
K+ > Rb+ Schließzellprotoplasten von Alli-um cepa
Amodeo et al., 1994
in, Isteady Gerstensuspensionszellen Amtmann et al., 1997Weizenwurzelhaare Gassmann et al., 1994Chara corallina Tonoplasten Homble et al.,1990Chara Tonoplasten Klieber et al., 1993Chara australis Tonoplasten Laver & Walker, 1991
out Samanea saman Schließzellen Moran et al., 1990Chara Tonoplasten Pottosin et al., 1993
K+ ≈ Rb+ in, Urev Gerstensuspensionszellen Amtmann et al., 1997Plasmamembran von Hefe Bertl et al., 1998
out Endospermprotoplasten Stoeckel & Takeda, 1989
Tabelle 11. Zusammenfassung von Tabelle 10.
Sequenz PRb/PK HäufigkeitK+ >> Rb+ 0 - 0,4 2 (15%)K+ > Rb+ 0,45 – 0,90 8 (62%)K+ ≈ Rb+ 0,95 – 1,05 3 (23%)K+ < Rb+ ≥ 1,10 (max = 1,40) 0

108
Tabelle 12. Vergleich der Permeabilität für Rb+ und K+ bei tierischen Zellen. Im Gegensatz zupflanzlichen, sind hier Beispiele für alle möglichen Sequenzen (Rb+>K+, Rb+≈K+, Rb+<K+) zufinden (m = membrane, e = expressed in). Daten ermittelt mit Winspirs für den Zeitraum von1989 bis Oktober 1998.
Sequenz Kanal Zellart PRb/PK QuelleK+>>Rb+ basolateral m of intralobular duct cells from
mouse mindibular gland0,38 Dinodom et al., 1994
in guinea pig ventricular cells 0,27 Matsudo, 1996in basolateral m of renal proximal tube Mauerer, 1998
K+>Rb+ cultured embryonic Drosophila myotubes 0,87 Chang et al., 1994cloned renal K+ channels e Oocytes Chepilko et al., 1995human erythrocytes membranes 0,71 Christophersen, 1990malignant lymphocytes (Nb2 cells) 0,75 Cukierman, 1992Drosophila larval muscle Delgado et al., 1995Shaker B K channels e insect cell line Sf9 Gomez-Lagunas & Arm-
strong, 1994out human lung epithelial cell line 0,49 Gillian-Ridge et al., 1997
rabbit superior lacrimal gland 0,83 Herok et al., 1998human submandibular gland duct cell line 0,61 Ishikawa & Cook, 1995NG 108-15 cells 0,63 Ishimoto et al., 1995NG 108-15 cells 0,62 Kaiho et al., 1996carp olfactory receptor cells 0,71 Kolesnikov & Kosolapov,
1993chick sensory neurons 0,45 Lux et al., 1990cultured rat adrenal chromaffin cells 0,81 Park, 1994primary neuronal cultures of Drosophila Saimi & Martinac, 1989principal cells of rat cortical collecting ducts Schlatter et al., 1992rat distal colon crypt cells Seimer & Goegelein,
1992IR bovine pulmonary artery endothelial cells 0,45 Silver et al., 1994
human osteoblasts Steinert & Grissner, 1997cultured human mesangial cells 0,54 Stockand & Sonsom,
1994hair cells isolated of the goldfish saccularmacula
0,80 Sugihara, 1994
resting Necturus gestric exynticopeptic cells Supplisson et al., 1993epithelial cell line 0,88 Tabcharani et al., 1994embryonic chick dorsal root ganglion neurons 0,67 Trequattrini et al., 1996
in erytrhocyte membrane of lamprey Virkki & Nikinmaa, 1996primary neuronal cultures of Drosophila 0,47 Yamamoto & Suzuji,
1989Xenopus oocytes Yang & Sachs, 1990epithelial K channel e Oocytes 0,5 Zhou et al., 1994

109
Sequenz Kanal Zellart PRb/PK
Quelle
K+ ≈ Rb+ out thick ascending limb of Henle’s loop Bleich et al., 1990human T lymphocytes 0,96 Grissmer et al., 1993guinea pig ventricular myocytes Hadley & Hume, 1990Jurat human leukaemic T cell line 0,96 Hanselmann & Grissmer,
1996frog taste receptor cells Kolesnikov & Margol-
skee, 1998mouse cortical collecting duct cells Korbmacher et al., 1995guinea pig cardiac myocytes Matsuoka et al., 1990guinea pig ventricular myocytes Randle et al., 1991
out,in paramecium cell Saitow et al., 1997basolateral membrane of marginal cells disso-ciated from gerbil stria vascularis
Takeuchi et al., 1995
in mouse cortical principal cell Welling, 1997K+<Rb+ rabbit arterial endothelial cells Katnik & Adams, 1995
m of rat inner medullary conducting duct 1,10 Nonaka et al., 1995mouse receptor channesl e Oocytes Tsuzuki et al., 1994rat superior cervical ganglion neurones 1,4 Zhu & Ikeda, 1993Xenopus axon Safronov & Vogel, 1995
abhäng-ig vonRbext
Shaker K channel ? Perez & Begenish, 1994
Tabelle 13. Zusammenfassung der Ergebnisse aus Abbildung 60.
Sequenz PRb/PK HäufigkeitK+ >> Rb+ 0 - 0,4 3 (6%)K+ > Rb+ 0,45 – 0,90 28 (60%)K+ ≈ Rb+ 0,95 – 1,05 11 (23%)K+ < Rb+ ≥ 1,10 (max = 1,40) 5 (11%)

110
10.5 Einfluß des pH-Wertes
Ein direkter Vergleich der pH-Abhängigkeit der K+-Auswärtsgleichrichter von Bündel-scheidenzellen und Mesophyllzellen ergab die in Abschnitt 10.6 besprochene physiolo-gische Bedeutung der Effekte.
A
B
Abbildung 60. Vergleich der pH-Effekte bei Mesophyll- und Bündelscheidenzellen (MB-2).A: pH 6.5 und B: pH 6.15. Lösungen „rot-braun“, „+(rot)“ und „blau“ aus Tabelle 8.
-200 -150 -100 -50 0 50
-10
0
10
20
MB-1
MM-2
pH 6.5 MB-2
I (µA
cm-2)
V / mV
-200 -150 -100 -50 0 50
-10
0
10
20
MB-1
MM-2
pH 6.15MB-2
I (µA
cm-2)
V / mV

111
Während bei neutralem pH-Wert das Aufnahme- und Abgabeverhalten von Bündel-scheidenzellen (MB-2) und Mesophyllzellen vollkommen übereinstimmen (Abbildung58 A), sind bei leichter Ansäuerung Bündelscheidenzellen besser in der Lage, K+-Ionenmit ihrer Umgebung auszutauschen (Abbildung 60 A), und zwar sowohl bei negativenals auch bei positiven Spannungswerten. Eine solche Stimulierung der K+-Kanäle derBündelscheidenzellen im Sauren ist für einige Einwärtsgleichrichter in Blattzellen be-kannt (4/7, Tabelle 15), bislang aber unbekannt für Auswärtsgleichrichter.
Tabelle 14. Übersicht des Einflusses des pH-Wertes auf Blattzellen. Es tritt kein Auswärts-gleichrichter auf, bei dem eine Ansäuerung den Strom erhöht. Kat = Kationen, G = Schließzel-le, V = Vakuole, E = Epidermis, c = Zellkultur, g = geklont, h = heterologe Expremierung. Da-ten ermittelt mit Winspirs für den Zeitraum von 1989 bis Oktober 1998. Der Übersicht halbersind die verschiedenen Wirkungen vom Schriftbild abgehoben (bei Alkalisierung: Abnahme,Zunahme, kein Effekt)
Kanal Zell-typ
Pflanze pH außen pH in-nen
Strom Quelle
K+
(IR)G Solanum tubero-
sum5.6-7.4
(4.5-8.5)7.2 nimmt bei Alkalisierung ab Dietrich et al.
1998, Hoth et al.1997
K+
(IR)G Nicotiana ta-
bacum8.5 - 4.5 7.2 nimmt bei Alkalisierung ab wie oben
K+
(IR)G Vicia faba L. 5.6 - 7.4
(4.5 - 8.5)7.2 kein Effekt wie oben
Kat(inst.)
V Beta vulgaris V: 7.3 7.3 -8.0
nimmt bei Alkalisierung zu Davies et al.1995
Kat (me-chano-
sensitiv)
E onion (bulbcells)
4.5 - 7.2 7.2 nimmt bei Alkalisierung zu Ding et al. 1993
K+
(IR) (OR)c Arabidopsis
thaliana5.2 - 8.2 7 nimmt bei Alkalisierung zu
auf OR kein EffektGiromini et al.
1997K+
(IR) (OR)c Arabidopsis
thaliana? ? -
6.5OR nimmt bei Alkalisierung
zu, IR nimmt abwie oben
K+
(IR)g c Arabidopsis
(Makro patch)7.2 6.2 -
7.2U < -150 mV: nimmt bei
Alkalisierung zu,U > -150 mV Abnahme
Hoshi 1995
K+
(OR)G Vicia faba 4.4 - 8.1 7.2 nimmt bei Alkalisierung zu,
Einzelkanal bleibt gleichIlan et al. 1994
K+
(IR)G Vicia faba 4.4 - 5.5,
5.5 - 8.1? nimmt bei Alkalisierung ab
(erst stark, dann schwach)Ilan et al. 1996
K+
(OR)V Eremosphera
viridis5.1 5.1 -
7.55nimmt bei Alkalisierung zu Linz & Köhler
1994Kat (IR) V Eremosphera
viridis5.1 5.1 -
7.55kein Effekt wie oben
K+ (IR) h Kartoffel 5.5 - 6.6 7.4 nimmt bei Alkalisierung ab Zimmermann etal. 1998

112
Bei pH-Werten um 6, die dicht an physiologisch zu erwartenden Werten liegen (pH6.15, Abbildung 60 B), ist der Unterschied so drastisch, daß der Austausch zwischenMesophyllzellen und ihrem Apoplasten sehr gering wird. Die Ströme liegen nun in derGrößenordnung des MB-1. Zudem sind die Einwärtsstöme des MB-2 auch erheblichangestiegen.
Die pH-Werte der Untersuchungen aus Tabelle 15 liegen zwischen pH 4.4 bis pH8.4. Physiologisch ist eher der saure Bereich zu erwarten (Tabelle Apoplast, Xylem etc).Das Cytoplasma wird in den meisten Untersuchungen etwa bei pH 7 gelassen.
Tabelle 15. Wirkung von Alkalisierung auf pflanzliche Membranen. Zusammenfassung derErgebnisse aus Tabelle 14.
Abnahme Kein Effekt ZunahmeIR 4 2 1OR 0 1 3
instantan 0 0 1
10.6 Physiologische Bedeutung der pH-Abhängigkeiten
Die Stimulierung des MB-1 und MB-2 im sauren Milieu weist auf eine wichtige physiolo-gische Funktion bei der Stoffaufnahme vom Xylem in den Symplasten durch Cotransport.Die eindrucksvollsten Experimente zum Nachweis der für den Cotransport notwendigenBedingungen entlang der Blattvenen, stammen von Canny (Sojabohnenblätter: Canny,1987; Bäume, Stäucher und krautartige Pflanzen: Canny 1988; Weizenblätter: Canny1990a; verschiedene Blätter von dicotylen (zweikeimblättrigen) Pflanzen: Canny1990b). Er benutzte den Farbstoff Sulphorhodamine G, um die von H+-Pumpen erzeugteAnsäuerung zu zeigen. Bouché-Pillon et al. (1994) wiesen durch Immunolocalisationebenfalls hohe H+-Pumpenaktivität entlang der kleinen Blattvenen nach.
Die vermutete Rolle des K+-Kanals hierbei wird aus dem Schema in Abbildung 61deutlich: Die linke Seite zeigt die beteiligten Transporter bei der Aufnahme von Anionen(Cl- oder NO3
-) aus dem Xylem in die Bündelscheidenzellen. Die ATP-getriebene H+-Pumpe (ATPase) baut den pH-Gradienten und die Membranspannung auf. Die Driving-Force aus diesem elektroosmotischen Gradienten nutzt ein 2H+/Anionensymporter(Cotransporter, z. B. Sander und Hansen 1981), um Anionen gegen die Membranspan-nung und gegen den Konzentrationsgradienten gekoppelt an H+ zu transportieren. Die H+,die die ATPase herausgepumpt hat, gehen also beide mit dem Cotransporter wieder hin-

113
ein. Nur durch sie würden bei diesem isolierten System elektrische positive Ladungenaußen angereichert. Nach der Formel U = 1/C ∫ I dt würden bald Blitze die Membran zer-stören. Allerdings würde die Spannung als rücktreibende Kraft (– 60 mV mal pH-Unterschied) bei - 450 mV (Energie aus ATP) die Pumpe stoppen. Dennoch ist die Gefahraus den hypothetischen Blitzen nicht unrealistisch: Die Membran schlägt bei ca. -350 mVdurch.
Abbildung 61. Modell für das Zusammenspiel von Cotransport und pH Wert in Bündel-scheidenzellen und Phloem bei Mais. Linkes Schema: Rolle des K+-Kanals MB-2 beim Ladungs-ausgleich für den 2H+/NO3
- (oder 2H+/Cl-) Cotransport (H+ allein: Pumpe). Rechtes Schema, linkeSeite: K+-Einstrom ist bei Zucker (Su) Cotransport (pH 5) nicht notwendig. Rechtes Schema,rechte Seite: Beim Versagen der Pumpe (pH 7) öffnet der K+-Kanal und verhindert, daß dasMembranpotential unter EK fällt.
Um das eine oder das andere zu verhindern, muß ein Kationen-Kanal zum Ladungs-ausgleich für die H+ Pumpe geöffnet werden. Die Ansäuerung aufgrund der Pumpenakti-vität ist für den MB-2 das Signal zu öffnen. Hierbei mag eingewendet werden, daß der vonuns untersuchte Kanal ein K+-Auswärtsgleichrichter zu sein scheint, doch Abbildung 60 Bzeigt, daß auch eine nennenswerte Leitfähigkeit im Einwärtsbereich besteht, die ebenfallseine Stimulierung mit fallendem pH zeigt. Damit scheint der hier nachgewiesene Kanalfür die oben postulierte Aufgabe eingerichtet zu sein.
Obige Rolle des MB-2 soll vor dem Hintergrund eines gegensätzlichen Befundes be-leuchtet werden. Der ZMK1 aus der Maiskoleoptile (Philippar et al., 1999) hat eine entge-gengesetzte pH-Abhängigkeit (Schließen im Sauren). Er kommt auch in Leitbündeln vor.Es erschien erst widersprüchlich, daß zwei Kanäle aus Leitbündeln bzw. deren direkterUmgebung, der ZMK1 und der MB-2, sich so unterschiedlich verhalten. Des Rätsels Lö-sung liegt darin, daß der ZMK1 (wie der entsprechende AKT 1 von Arabidopsis thalliana,Marten et al., 1999) im Phloem lokalisiert ist, und damit ganz andere Aufgaben hat.
Während der Transport Xylem-Bündelscheidenzelle in erster Linie der Aufnahme vonNährsalzen dient, beschäftigen sich die Transporter in der Plasmamembran der Phloem-zellen mit der Aufnahme von Zuckern und Aminosäuren, die häufig elektrisch neutral

114
sind. Abbildung 61 (linke Seite des rechten Schemas) zeigt, daß beim Cotransport vonZuckern oder neutralen Aminosäuren kein Bedarf für den Ladungsausgleich durch einenK+-Kanal besteht. Im Gegenteil, er würde nur Energie verschwenden.
Deshalb wird der ZMK1 (und der AKT1) im Sauren geschlossen. Das Öffnen bei neu-tralem pH kann eine Fail-Safe-Funktion erfüllen (rechte Seite des rechten Schemas inAbbildung 61). Wenn die Pumpe ausfällt (Energiemangel, Parasitenangriff), bricht dieMembranspannung zusammen. Doch die Zelle braucht für viele Funktionen eine Min-destmembranspannung. Jetzt öffnet der K+-Kanal und hält die Membranspannung bei EK
(Nernstspannung, Abschnitt 4.2, -120 mV bis -60 mV) fest. Das Öffnen wird direkt vondem mit dem Ausfall der Pumpe einhergehenden Rückgang der Ansäuerung auf pH 7 aus-gelöst.
Die Frage taucht auf, warum sich der MM-2 wie der ZMK1 verhält. Es ist zu vermuten,daß auch an der Plasmamembran zwischen Blattapoplast und Mesophyllzellen Cotrans-port von Stoffwechselprodukten stattfindet. Der Sinn etwaiger Zuckeraufnahme aus demApoplasten ist unklar. Bentrup (persönliche Mitteilung) äußerte einmal in der Diskussion,warum grüne Algenzellen (also metabolische Selbstversorger) Cotransporter für die Zuk-keraufnahme besitzen, die Vermutung, daß die Zellen sich „putzen“ müssen. Der unver-meidlich aus der Zelle leckende Zucker würde im Apoplasten Parasiten anlocken, und umdies zu verhindern, muß die Zelle diesen Zucker wieder aufnehmen können.
Tabelle 14 und ihre Zusammenfassung in Tabelle 15 zeigt, daß sonst kein Auswärts-gleichrichter aus Blattzellen bekannt ist, bei dem Ansäuerung den Strom erhöht. Füreine solche Stimulierung von Einwärtsgleichrichtern gibt es hingegen einige Beispiele.
10.7 Biophysikalische Aspekte der Na+- und Rb+-Messungen
Die Na+-Messungen am Einwärtsgleichrichter (IR) und die Rb+-Messungen am MB-1brachten noch einen besonderen Effekt: ein zweites Ion, wenn es in Abwesenheit vonK+ vorliegt, verändert die Permeabilität zu seinen Gunsten. Solch ein Effekt wurde fürreines Na+ auch von Immke et al. (1999) für den klonierten K+-Kanal Kv2.1 und vonOgielska und Aldrich (1998, 1999) für den in Oocyten exprimierten K+-Kanal gefunden.
Nach dem Strukturmodell von Doyle et al. (1998) täuschen die Karbonylgruppen derKanalporen den Ionen eine Wasserhülle vor, wenn der Ionenradius genau zum Porenra-dius paßt. Dies ermögliche hohe Durchtrittsraten bei sehr hoher Selektivität. Obige Er-gebnisse unterstützen die Hypothese von Ogielska und Aldrich (1999), daß am Poren-rand Bindungsstellen existieren, die die Selektivität des Kanals beeinflussen: Die Ami-nosäuren T445, W434 und W435 bilden eine Manschette, die bei Besetzung mit K+ sichdem K+- und sonst dem Na+-Ionenradius anpaßt. Ebenso ist ein Lysin im P-Loop des

115
RCK-4 Kanals verantwortlich dafür, daß der Kanal nur dann K+ durchläßt, wenn amP-Loop K+ gebunden wird (Pardo et al., 1992).
Offenbar sind auch die Maiszellen für das Studium solcher biophysikalischen Effekteein geeignetes Objekt.
10.8 Vergleich der Ergebnisse von Wurzel und Blatt
Abbildung 62. Modell für Kanäle in der Gerstenwurzel (linke Seite, nach Wegner undRaschke, 1994) und im Maisblatt (rechte Seite) für physiologische pH- und K+-Werte. Die Plas-modesmen (PD) sorgen für den schnellen diffusiven Austausch zwischen den Blattzellen. DerApoplast ist dort durch die Suberinlamelle in zwei Bereiche geteilt. BS = Bündelscheidenzellen,M = Mesophyllzellen, VP = vaskuläres Parenchym, X = Xylem.
Der Weg der Nährstoffe (Ionen) führt von der Wurzelrinde in den Symplasten derMaiswurzel (Abschnitt 2.3, Abbildung 2), bevor eine Aufnahme in das Xylem stattfin-det. Der Casparysche Streifen, der in der Wurzel die Rinde vom Zentralzylinder trennt(Abbildung 2), ist für Ionen und Wasser undurchlässig und macht daher eine Aufnahmein den Symplasten erforderlich. In der Wurzel kann, anders als im Maisblatt, das Paren-chym eindeutig entweder dem Xylem oder dem Phloem zugeordnet werden. In den Xy-lemparenchymzellen findet die Abgabe der Nährstoffe in das Xylem statt.
Wegner und Raschke (1994) haben diesen Aufnahmevorgang in Gerstenwurzeln zu-nächst für Kationen untersucht. Sie fanden drei Transportertypen: einen K+-Einwärtsgleichrichter (KIRC), einen nicht-selektiven Kationen-Auswärtsgleichrichter(NORC) und einen selektiven K+-Auswärtsgleichrichter (KORC). Die Anatomie derGerstenwurzel ist der von Mais sehr ähnlich, da beide Pflanzen zu den Gräsern gehören.
Abbildung 62 zeigt das Vorkommen der K+-Kanäle in der Grenze zum Wurzelxylem(Wegner und Raschke, 1994) im Vergleich zu denen im Maisblatt in schematischenQuerschnitten. Eingetragen sind auch die K+-Konzentrationen und die pH-Werte. Die

116
Meßbedingungen waren ähnlich, in der Gerstenwurzel wurde mit etwas höheren Kon-zentrationen und apoplastisch in etwas saurerem Milieu gearbeitet.
Die Stromspannungskurven aller am Xylem-Symplasten-Übergang beteiligten K+-Kanäle in der Wurzel und im Blatt zeigt Abbildung 63. Aus dem Maisblatt ist nur derMB-2 eingezeichnet, da nur er vermutlich an das Xylem grenzt und so für die Aufnahmein den Symplasten verantwortlich ist.
KORC ist für große positive und negative Spannungswerte größer als MB-2, aber imphysiologisch relevanten Bereich, gekennzeichnet durch negative Membranspannungzwischen – 50 und – 120 mV, etwa gleich. Da der MB-2 im Sauren aktiviert wird undi.A. zudem höhere Konzentrationen auch zu höheren Strömen führen, ist damit zu rech-nen, daß unter identischen Bedingungen insbesondere die Einwärtsströme des MB-2größer sind, als in Abbildung 63 dargestellt.
Abbildung 63. Schema der Stromspannungskurven der Transporter aus Abbildung 62.Verwendete Lösungen bei NORC, KORC und KIRC: 30 mM KCl + 10 mM HEPES + 2 mMMgCl2 + 1 - 40 mM CaGluconat im Bad und 120 mM KCl + 10 mM Tris + 10 mM EGTA + 2mM MgATP + 1.93 mM MgCl2 + CaGlukonat (insgesamt 1µM Ca2+) in der Pipette. Lösungenbeim MB-2: „+(rot)“ und „blau“ aus Tabelle 8.

117
10.9 Ausblick
Im Fortsetzungsprojekt Ha 712/11-3 (im dritten 2-Jahresabschnitt des DFG-Schwerpunkt-programms SPS 717 „Der Apoplast der Höheren Pflanze, Speicher-, Transport- und Re-aktionsraum“) wird die Verknüpfung zwischen Ionenkanälen und Cotransport als diemöglicherweise beim Xylem/Symplast Übergang im Vordergrund stehende physiologi-sche Funktion im Zentrum des Interesses stehen.
Zwei Fragen stellen sich dabei. Die erste betrifft Milieuunterschiede im Blattapoplastenund im Xylem. Die Xylemsaftzusammensetzung hat sich weitgehend bereits aus den Ar-beiten im Rahmen des DFG-Schwerpunktes SPS 717 ergeben und wird noch vervollstän-digt werden. Einige Ergebnisse sind in Tabelle 2 dargestellt. Auch für den Blattapoplastenliegen Ergebnisse vor (ebenfalls Tabelle 2). Eine Vertiefung der Untersuchungen insbe-sondere unter dem hier angesprochenen Aspekt wird möglich, wenn im Sommer 2000Christoph Plieth aus Edinburgh zurückkehrt und molekularbiologische Methoden mit-bringt, sensorische Farbstoffe an der gewünschten Stelle im Blatt zu exprimieren (Plieth etal., 1999a,b).
Die zweite Frage bezieht sich auf den Nachweis und die meßtechnische Erfassung desCotransportes. Cotransport wird normalerweise dadurch nachgewiesen, daß die Depolari-sation registriert wird, die sich ergibt, wenn die zu transportierende Substanz ins Bad ge-geben wird (Köhler et al., 1991, Maathuis et al., 1997, Maathuis und Sanders, 1997). DieDepolarisation geschieht aufgrund der Belastung der Membranspannung durch den elek-trischen Strom durch den Cotransport.
Für diese Untersuchung haben erste, hier noch nicht dargestellte Ergebnisse gezeigt,daß die Bündelscheidenzellen erstaunlich gut für Elektroden zugänglich sind. Das Einste-chen ist sehr viel einfacher als nach früheren Erfahrungen aus der Arbeitsgruppe mit Holz-strahlzellen (Himpkamp, 1988) oder Aegopodium (Vanselow et al., 1989) erwartet wordenist. Die Membranspannungen sind sehr hoch, im -200 mV-Bereich. Somit solltenCotransportuntersuchungen in situ möglich sein.
Bei Mesophyllzellen stellt sich die Frage, ob die Kopplung zum Cotransport eine ge-ringe Rolle spielt (der Stoffaustausch mit dem Mesophyllapoplasten ist sicher nicht sostark wie mit dem Xylem) oder ob eine Anpassung an einen anderen pH-Wert vorliegt.
Ein dritter Fragenkomplex im neuen Projekt folgt aus der gefundenen Verschieden-artigkeit von MB-1, MB-2 und MM-2: Führt die Aufteilung des Apoplasten des Mais-blattes in mehrere Kompartimente (xylemseitig und mesophyllseitig von der Suberin-schicht um die Bündelscheidenzellen; Leitbündel zweiter und dritter Ordnung, Kontakt-bereich Xylem/BS) zur weiteren Anpassungen der Plasmalemmatransporter an dieseBedingungen? Es ist also zu untersuchen, ob sich die K+-Kanäle noch in weiteren Ei-

118
genschaften außer den in Abb. 48 bis 50 und 60 dargestellten pH-Abhängigkeiten unter-scheiden, und ob die morphologische Zuordnung noch weiter verfeinert werden kann.
In Zusammenarbeit mit der Arbeitsgruppe Hedrich (Würzburg) werde ich versuchen,die Kanäle aus den Zellen zu isolieren. Gelingt dies auch für die zwei Typen (MB-1 undMB-2) aus Bündelscheidenzellen, können diese mit Hilfe von biochemischen Färbe-techniken in Schnitten sichtbar gemacht werden. Damit kann die Frage der in Tabelle 9angesprochenen Zuordnung ohne und mit Xylemkontakt zu MB-1 und MB-2 geklärtwerden.
Kirsch et al. (1992) zeigten, daß der klonierte K+ Kanal Kv2.1 in seiner Rb+-Abhängigkeit in den Kv3.1 überführt werden kann, wenn man die Aminosäure 372austauscht. Diese Aminosäure könnte Teil einer Kationenbindestelle tief in der Kanal-pore sein. Insgesamt unterscheiden sich die beiden Typen im Bereich des Selektivitäts-filters nur in sechs Aminosäuren. Ob sich die Kanäle MB-1, MB-2 und MM-2 in ihrenSequenzen ähnlich gering unterscheiden, und wie eng sie mit den anderen Maiskanälen(ZMK1 und ZMK2, Philippar et al., 1999) verwandt sind, kann ebenfalls bei den ge-planten Untersuchungen in Würzburg geklärt werden.
Offen blieb bislang auch die Frage der Zuordnung von IR, SC und eventuell weiterenTransportern zu den Whole-Cell-Strömen. Dies ließe sich in künftigen Experimentenmit einem besonderen Mittelungsverfahren überprüfen. Das Verfahren ermöglicht eineZuordnung, auch wenn die Einzelkanaldaten ein sehr schlechtes Signalrauschverhältnishaben, welches bislang die Ursache für die spärliche Ausbeute an Einzelkanaldaten ist.Dazu wählt man als vorgegebene Spannung im Excised-Patch keine Konstante, sondernwie bei den Whole-Cell-Aufzeichnungen eine Sprungfolge, mit dem Unterschied, daßimmer derselbe Sprung durchgeführt würde. Eine Überlagerung dieser Sprünge ergäbeeine Art Whole-Cell-Antwort nur dieses einen Kanals (Müller-Röber et al., 1995). Soließe sich ermitteln, ob z.B. das Zeitverhalten vom MB-2 mit dem vom IR überein-stimmt.

119
11 Zusammenfassung
Die im Rahmen des DFG Projektes „Der Apoplast der Höheren Pflanze, Speicher-,Transport- und Reaktionsraum“ durchgeführte Arbeit befaßte sich mit den Transport-vorgängen im Maisblatt, insbesondere mit der Ionenaufnahme aus dem Apoplasten inden Symplasten.
Der Apoplast in Maisblättern ist durch die undurchlässige Suberinlamelle um dieBündelscheide in zwei Bereiche geteilt. Es gibt daher zwei Orte der Wechselwirkungenvon Apoplast und Symplast: innerhalb der Suberinlamelle (Bündelscheidenzellen) undaußerhalb (Mesophyllzellen).
Innerhalb der Suberinlamelle mußte zunächst die Frage geklärt werden, an welchemOrt dieser Übergang stattfindet (Kapitel 3). Zwei Zelltypen kamen hierfür in Betracht:die Bündelscheidenzellen und das vaskuläre Parenchym. Hierzu wurden Maisblätter mitRb+ als Tracer für K+ angefüttert. Querschnitte wurden mit Hilfe eines Cryomicrotomserstellt. Die Schnitte wurden dann mit einem Massenanalysator für besonders kleineProben (Laser Mikroproben Massen Analysator = LAMMA) untersucht. Es konntenachgewiesen werden, daß Rb+ sich zunächst in den Bündelscheidenzellen ablagert.Damit waren die Bündelscheidenzellen die Zellen, die für das Projekt von Interesse wa-ren.
Der nächste Schritt mußte nun die Isolation der betreffenden Zellen sein (Kapitel 5).Hier zeigte sich, daß das Maisblatt spezielle Schwierigkeiten aufwies. Die Undurchläs-sigkeit der Suberinlamelle um die Bündelscheide wurde in Farbstoffexperimenten deut-lich. Erst die isolierten Protoplasten waren durch Neutralrot anfärbbar, und damit ihreLebensfähigkeit bewiesen.
Nach erfolgreicher Isolation konnten Einzelkanalmessungen an Bündelscheidenzel-len mit der Patch-Clamp-Technik durchgeführt werden (Kapitel 7). Die Einzelkanal-ströme von Bündelscheidenzellen waren sehr gering. Daher wurde zumeist unter hohenSalzkonzentrationen (100 – 200 mM) gemessen. Es traten zwei Typen von Kanälen auf,ein symmetrischer Kanal (SC) und ein Einwärtsgleichrichter (IR). In 92% der Messun-gen wurde der SC gefunden. Dieser zeichnete sich durch symmetrische Stromspan-nungskurven aus, während der IR bei positiven Spannungen häufig gar nicht öffnete.Unterschiedlich waren daher auch die Offenwahrscheinlichkeiten: Der SC hatte für po-sitive und negative Spannungen die gleiche Verteilung, während der IR eine starkeAsymmetrie zeigte. Anhand des Umkehrpotentials konnte gezeigt werden, daß es sichbei dem SC um einen K+-Kanal handelte, der jedoch nicht sehr selektiv war. Der SCwurde durch Na+ geblockt. Diese Blockierung zeigte sich sowohl in einer Erniedrigungdes Einzelkanalstroms als auch in einer Reduktion der Offenwahrscheinlichkeit (Ab-schnitt 7.6). Der IR konnte durch reine Na+-Lösung aktiviert werden. Der SC war für

120
NH4+ genauso durchlässig wie für K+. Unter physiologischen K+-Bedingungen entstand
aufgrund der geringen Ströme um das Umkehrpotential eine Meßlücke.Mit einem erweiterten Präparationsverfahren war es möglich, auch Whole-Cell-
Messungen mit der Patch-Clamp-Technik durchzuführen (Kapitel 8 für Bündelschei-denzellen und Kapitel 9 für Mesophyllzellen). Bei den Whole-Cell-Messungen ist esmöglich, physiologische Kalium- und Calciumkonzentrationen zu verwenden.
Es traten bei den Whole-Cell-Messungen an Bündelscheidenzellen zwei Typen vonWhole-Cell-Antworten auf: ein rein instantaner (MB-1) und ein zeitabhängiger (MB-2).Die Ströme des MB-1 waren deutlich kleiner als die des MB-2. Der MB-2 setzte sichaus einer instantanen und einer zeitabhängigen Komponente (MB-KI2 und MB-AOR)zusammen. Bei Mesophyllzellen trat nur ein Typ auf (MM-2), der wie der MB-2 auszwei Komponenten bestand (MM-KI2 und MM-AOR).
Die Anatomie des Maisblattes gab Anlaß zu der Vermutung, daß sich die beiden Ty-pen MB-1 und MB-2 darin unterschieden, ob sie Kontakt zum Xylem haben oder nicht.Ein Vergleich des Anteils an Bündelscheidenzellen mit und ohne Kontakt anhand vonausgezählten Schnitten mit dem Anteil des Auftretens bei Patch-Clamp-Messungen be-stätigte diese Hypothese. Auch die unterschiedliche Größenverteilung der Zellen mitund ohne Xylemkontakt zeigten in diese Richtung. Ein eindeutiger Beweis liegt aller-dings noch nicht vor.
Rb+ als Ersatz für K+ auf der Außenseite erwies sich als geeignetes Mittel, die Unter-schiedlichkeit aller drei Typen nachzuweisen. Beim MB-1 verschob sich das Umkehr-potential sehr weit in Richtung Rb+. Bei den beiden zeitabhängigen Typen MB-2 undMM-2 trat keine Verschiebung des Umkehrpotentials auf, allerdings unterschieden sichdie Stromspannungskurven von denen mit K+. Während der MB-2 für Rb+ größereWhole-Cell-Ströme lieferte, wurden sie beim MM-2 für Rb+ geringer als für K+. Diezeitabhängige Komponente des MB-2 unterschied sich für K+ und Rb+ nicht. Insgesamtzeigten alle untersuchten Zelltypen eine Durchlässigkeit für Rb+. Dies ist eine Voraus-setzung für die obigen Messungen mit Rb+ als Tracer für K+.
Auch die Veränderung des pH-Wertes des Außenmediums führte zu unterschiedli-chen Effekten. Eine Ansäuerung hatte bei Bündelscheidenzellen einen Stromanstieg zurFolge, bei Mesophyllzellen hingegen eine Stromverringerung. Dieser Effekt war sehrausgeprägt: während bei etwa neutralem pH-Wert von 7.2 Bündelscheiden- und Meso-phyllzellen (MB-2 und MM-2) identische Stromspannungskurven lieferten, unterschie-den sie sich für einen leicht sauren pH-Wert von pH 6.15 um den Faktor 7 für positiveund sogar Faktor 20 für negative Spannungen. Die Stromspannungskurve des MM-2 lagjetzt in der Größenordnung des MB-1.
Die physiologische Bedeutung des pH-Effektes auf Bündelscheidenzellen wird deut-lich beim Vergleich mit Ergebnissen an Phloemzellen. Phloemzellen aus Maiskoleopti-len (Philippar et al., 1999) werden bei Alkalisierung aktiviert. Ihr Verhalten ist also ge-nau gegenläufig zu den Bündelscheidenzellen (MB-2). Bei den Bündelscheidenzellen

121
wird durch die Ladungsbilanz von Cotransport (2H+ je NO3-) und Pumpe (H+ aus der
Zelle) ein Ladungsausgleich nötig. Dieser wird durch die Ansäuerung der Pumpe voneinfließendem K+ geliefert.
Anders ist die Situation im Phloem und bei Mesophyllzellen. Hier werden mit demCotransporter viele neutrale Aminosäuren und Zucker befördert, wodurch kein Bedarffür einen Ladungsausgleich besteht. Erst wenn die Pumpe versagt, d.h. wenn der pH-Gradient über der Membran verschwindet, tritt der K+-Kanal in Aktion. Er verhindert,daß das Membranpotential zusammenbricht, indem er es auf EK festsetzt.
Ein Vergleich mit Messungen an Gerstenwurzeln (Wegner und Raschke, 1994), dieebenfalls zu den Gräsern gehören und daher eine der Maiswurzel vergleichbare Anato-mie haben, zeigte ein ähnliches Verhalten unter physiologischen Bedingungen (pH-Wert, K+-Konzentration). Während die Kanäle in der Wurzel allerdings die Aufgabehaben, Nährstoffe an das Xylem abzugeben, kommt den Transportern im Blatt die Auf-gabe zu, diese Nährstoffe wieder in den Symplasten aufzunehmen.
Sowohl in ihren pH-Abhängigkeiten als auch in ihrem unterschiedlichen Verhaltenbeim Austausch von K+ durch Rb+ sind die untersuchten Zellen einzigartig. Ihre Unter-suchung bringt einen wesentlichen Fortschritt für das Verständnis von Transportvor-gängen im Maisblatt dar.

122
12 Anhang
12.1 Beschichtung der LAMMA-Grids
Abbildung 64. Zur Beschichtung der Kupfergrids mußzunächst eine Formvarschicht hergestellt werden, diedann auf die Grids übertragen wird. Dazu wird ein neuerObjektträger in eine Formvarlösung eingetaucht, derenAbfluß mit Hilfe eines Hahns geregelt werden kann. ImStopfen, an dem die Halterung befestigt ist, ist ein Loch,um einen gleichmäßigen Abfluß zu gewährleisten.
Die verwendeten LAMMA Grids (G2150C, Plano W. Planet GmbH, Wetzlar) sind ausKupfer und müssen, damit die Proben nicht durch das Gitter fallen, mit einer Formvar-lösung beschichtet werden.
Dazu muß zunächst eine 0.35% Formvar (Sigma-Aldrich Chemie GmbH, Steinheim)in Chloroform Lösung hergestellt werden, die nicht älter als einen Monat sein sollte(alte Lösung in vorgeschriebene Abfallbehälter entsorgen, da es sich um chlorierteWasserstoffe handelt). Das Gefäß (Abbildung 64) wird mit Chloroform gespült und derHahn dann geschlossen. Die Formvarlösung wird bis zur Markierung (soweit der Ob-jektträger eintauchen soll) eingefüllt. Ein neuer, von Staub befreiter Objektträger wirdam Deckel befestigt und in die Lösung getaucht. Nun wird der Hahn etwas geöffnet, sodaß die Lösung unten wieder hinausfließt. Die Zeit, die dabei verstreicht, bestimmt dieSchichtdicke, je länger es dauert, um so dicker ist die Formvarschicht hinterher (da derObjektträger länger von Dämpfen umgeben ist).
Ein Gefäß wird nun mit destilliertem Wasser gefüllt und die Oberfläche von Staubbefreit, indem mit einem sauberen Glasstab überstehendes Wasser (samt Staub) wegge-wischt wird. Die Schicht auf dem Objektträger wird an allen vier Kanten mit einer Prä-pariernadel sorgfältig eingeritzt, angehaucht und danach sofort langsam schräg in dasWasser getaucht. Hierbei löst sich die Beschichtung vom Objektträger. Auf die nun auf

123
der Wasseroberfläche schwimmende Schicht werden die Grids mit der metallisch glän-zenden Seite nach oben (sichtbar) auf den Film gelegt. Ein Streifen aus Parafilm (Ame-rican National Can, Greenwich) wird nun vorsichtig auf den mit Grids bedeckten Filmgelegt und dann samt Film und Grids aus dem Wasser gezogen.
Nach einem Tag trocknen (abgedeckt mit einer Petrischale) können die Grids vomParafilm mit einer Pinzette abgenommen werden. Nun können die Proben darauf gesetztwerden.
12.2 Färben mit Neutralrot
Abbildung 65. Anreicherung von Neutralrot in der Vakuole einer Pflanzenzelle. A. Neutralrotliegt in alkalischer Lösung als lipophiles Molekül vor, in saurer (B) durch Anlagerung einesProtons als Farbkation. C. Ausgangssituation: lebende Zelle in verdünnter Neutraltrotlösung, pH8 (Farbmoleküle als Punkte dargestellt, Farbkationen als Striche). D. Endzustand: Farbmolekülesind in die Vakuole (pH 5) permeiert, die sie als hydrophile Farbionen nicht mehr verlassenkönnen. Gleichgewicht stellt sich erst ein, wenn die Konzentration der Neutralrotlösung in derVakuole gleich jener in der Außenlösung ist. Dann ist aber eine > 1000-fache Anreicherung desNeutralrots – in Ionenform – in der Vakuole erreicht. (Aus: Sitte et al., 1998).

124
Es gibt nur wenige Farbstoffe, die sich zur Vitalfärbung von Pflanzenzellen eignen, diemeisten Farbstoffe schädigen die Zelle irreversibel. Neutralrot liegt in alkalischer Lö-sung als ungeladenes lipophiles Farbstoffmolekül vor (Abbildung 65). Trotz seinerGröße von 252 Dalton kann es durch Membranen diffundieren. In der sauren Vakuolelagern sich dann Protonen an und als nun geladenes Kation kann es die Membran nichtmehr passieren. Die Vakuole stellt somit eine Ionenfalle für Neutralrot dar, die Farb-stoffmoleküle sammeln sich dort an. Daher kann in saurer Lösung eine Neutralrotfär-bung nicht durchgeführt werden.
Zur Anfärbung wird eine Stammlösung von 1 g/l hergestellt. Sie wird dann mit Lei-tungswasser (pH min. 7.2) oder mit einer anderen alkalischen Lösung (ich verwendetedie Whole-Cell-Lösung mit pH 7.9) auf 1:10 verdünnt. Diese Lösung ist braunrot. Nunwerden die Zellen hinzugegeben.
Die Ergebnisse so gefärbter Zellen sind in Abschnitt 5.5 dargestellt.
12.3 Aphidentechnik
Abbildung 66. Blattläuse finden mit ihrem Rüs-sel zielsicher das Phloem (auch Sieböhren ge-nannt), wobei sie die Zerstörung des Gewebesminimieren.
Blattläuse haben die Fähigkeit, zielsicher an den für sie interessanten Phloemsaft zugelangen, indem sie die Phloemgefäße mit ihrem Rüssel anstechen (Abbildung 66). Er-staunlich ist dabei insbesondere, daß sie kaum andere Gefäße verletzen (Winter et al.,1992). Dieses wird sich in der Aphidentechnik zunutze gemacht, indem man Blattläuse

125
auf das gewünschte Blatt setzt und wartet, bis sie sich festgesaugt haben. Ihr Rüsselwird nun mit einem Laserstrahl abgetrennt und danach mit einem Mikromanipulatoreine Kapillare auf den Rüsselrest gesetzt.
Mit etwas Glück kann man sogar über mehrere Tage immer wieder neue Kapillarenauffüllen, und jeweils die wenigen nl analysieren. Es lassen sich so Tagesgänge ermit-teln (Winter et al., 1992).
Allerdings haben Blattläuse Vorlieben für gewisse Blätter. Verschiedene Arten ha-ben sich auf unterschiedliche Pflanzen spezialisiert, aber nicht für jede Pflanze läßt sicheine passende Blattlaus finden. Dies führte, wie in der Einleitung erwähnt, zur Be-schränkung auf die Maispflanze in der Kleingruppe des Apoplastenprojektes.
12.4 Leaching
Um zu untersuchen, ob eine Beregnung für Maispflanzen eine positive oder negativeAuswirkung hat, wurden Maispflanzen kontrolliert beregnet und die Lösung, die inKontakt mit den Blättern war, auf Rückstände untersucht. Verglichen wurden diesePflanzen mit solchen, die eine reine Anfütterung über die Nährlösung bekommen hat-ten.
12.5 Wurzeldruckkammer
Abbildung 67. Pflanzen müssen bereits in der Wurzeldruck-kammer angezogen werden, damit sie das Gefäß luftdicht ver-schließen und dann von außen nicht nur die Nährlösung einge-sprüht werden kann (Pfeil), sondern auch der Druck derartigerhöhbar ist, daß aus angeritzten Leitbündeln eines Blattes Xy-lemsaft exudiert (herausfließt).
Im Xylem herrscht Unterdruck. Daher ist es nicht möglich, ein Blatt einer Pflanze anzu-schneiden und so Xylemsaft zu gewinnen, sondern man würde auf diese Weise Phloem-saft erhalten. Um dennoch Xylemsaft zu gewinnen, muß im Wurzelbereich ein derarti-

126
ger Überdruck erzeugt werden, daß die Pflanze diesen bis in die Blattspitzen behält.Dazu läßt man die Pflanze in ein Gefäß hineinwachsen (Abbildung 67), bis der Wurzel-bereich luftdicht abgeschlossen ist. Die Pflanze wird mit Nährlösung besprüht, so daßleicht ein Überdruck angelegt werden kann. Nun wird ein Leitbündel eingeritzt und mitMikroelektroden pH-Wert, K+- und NO3
--Konzentration bestimmt.Leider ist dies der AG Schurr für Maispflanzen nicht gelungen, so daß nur die Er-
gebnisse für Rizinus vorliegen.

127
13 Literatur
Aggarwal SK, MacKinnon R 1996 Contribution of the S4 segment to gating charge in the Shaker K+
channel. Neuron 16: 1169-1177.Albertsen A 1992 Erfassung und Auswertung schneller Schaltvorgänge bei Patch-Clamp Untersuchungen.
Diplomarbeit, Kiel.Albertsen A 1994 Fast Patch-Clamp Data Acquisition and Kinetic Analysis of Multi-Channel Records -
Schnelle Datenerfassung von Patch-Clamp Daten und kinetische Analyse von Mehrkanalaufzeich-nungen. Dissertation Kiel.
Albertsen A, Hansen UP, 1994 Estimation of kinetic rate constants from multi-channel recordings by a directfit of the time series. Biophys. J. 67: 1393-1403.
Amodeo G, Escobar A, Zeiger E 1994 A cationic channel in the guard cell tonoplast of Allium cepa.Plant Physiol. (Rockville) 105: 999-1006.
Amtmann A, Laurie S, Leigh R, Sanders D 1997 Multiple inward channels provide flexibility in Na+/K+
discrimination at the plasma membrane of barley suspension culture cells. J. Exp. Bot. 48: 481-97.Antz C, Geyer M, Fakler B, Schott MK, Guy HR 1997 NMR structure of the inactivation gates from
mammalian voltage-dependent potassium channels. Nature 385: 272-275.Assmann,-SM; Simoncini,-L; Schroeder,-JI 1985 Blue light activates electrogenic ion pumping in
guard cell protoplasts of Vicia faba. Nature 318: 285-287.Backhausen JE, Kitzmann C, Scheibe R 1994 Competition between electron acceptors in photosynthesis:
Regulation of the malate valve during CO2 fixation and nitrite reduction. Photosynthesis Research 42:75-86.
Benz R, Läuger P 1976 Kinetic analysis of carrier-mediated ion transport by the charge-pulse technique.J. Membrane Biol. 27: 171-191.
Bertl A, Bihler H, Reid JD, Kettner C, Slayman CL 1998 Physiological characterization of the yeastplasma membrane outward rectifying K+ channel, DUK1 (TOK1), in situ. J. Membrane Biol. 162:67-80.
Bleich M, Schlatter E, Grger R 1990 The luminal potassium channel of the thick ascending limb ofHenle's loop. Pfluegers Archiv Eur. J. Physiol. 415: 449-460.
Blunck R, Kirst U, Rießner T, Hansen UP 1998 How powerful is the dwell-time analysis of multi-channelrecords? J. Membrane Biol. 165: 19-35.
Bosabalidis AM, Evert RF, Russin WA 1994 Ontogeny of the vasular bundle and contiguous tissues inthe maize leaf blade. Am. J. Bot. 81:745-752.
Botha CEJ, Evert RF, Cross RHM, Marshall DM 1982 The suberin lamella, a possible barrier to watermovement from veins to the mesophyll of Themeda trianda Forsk. Protoplasma 112: 1-8.
Botha CEJ 1992 Plasmodesmatal distribution, structure and frequency in relation to assimilation in C3and C4 grasses in southern Africa. Planta 187: 348-358.
Bouché-Pillon S, Fleurat-Lessard P, Fromont JC, Serrano R, Bonnemain JL 1994 Immunolocalization of theplasma membrane H+-ATPase in minor veins of Vicia faba in relation of phloem loading. Plant Physiol.105: 691-697.
Boyum R, Guidotti G 1997 Glucose-dependent, cAMP-mediated ATP efflux from Saccharomyces cere-visiae. Microbiol. (Reading) 143: 1901-1908.
Bregante M, Carpaneto A, Pastorino F, Gambale F 1997 Effects of mono- and multi-valent cations onthe inward-rectifying potassium channel in isolated protoplasts from maize roots. Eur. BiophysicsJ. 26: 381-391.
Brindley HM 1990 Effects of light/dark and calcium-channel drugs on fluxes of 86Rb+ in 'isolated' guard cellsof Vicia faba L. Planta 181: 440- 447.
Buschmann C, Grumbach K 1985 Physiologie der Photosynthese. Springer Verlag, Berlin.Canny MJ 1985 Water Pathways in Wheat Leaves. III The Passage of the mestome sheath and the func-
tion of the suberised lamellae Physiol. Plant. 66: 637-647.Canny MJ 1987 Locating active proton extrusion in leaves. Plant Cell Environ. 10: 271-274.

128
Canny MJ 1988 Bundle sheath tissues of legume leaves as a site of recovery of solutes from thetrasnpiration stream. Physiologia Plantarum 73: 457-464
Canny MJ, McCully ME 1989 The xylem sap of maize roots: its collection, composition and formation.Australian J. Plant Physiol. 15: 557-566.
Canny MJ 1990a What becomes of the transpiration stream? New Phytol. 114: 341-368.Canny MJ 1990b Fine veins of dicotyledon leaves as sites for enrichment of solutes of the xylem sap. New
Phytol. 115: 511-516.Canny MJ 1993 The transpiration stream in the leaf apoplast: water and solutes. Philos. Trans. Royal
Society London. Series-B, Biol. Sciences 341: 87-100.Canny MJ 1995 Apoplastic water and solute movement: new rules for an old space. Annual Review Plant
Physiol. Plant Mol. Biol. 46: 215-236.Chang G, Spencer RH, Lee AT, Barclay MT, Rees DC 1998 Structure of the MscL homolog from Myco-
bacterium tuberculosis: a gated mechanosensitive ion channel. Science 282: 2220-2226.Chang H, Ciani S, Kidokoro Y 1994 Ion permeation properties of the glutamate receptor channel in
cultured embryonic Drosophila myotubes. J. Physiol. (Cambridge) 476: 1-16.Chepilko S, Zhou H, Sackin H, Palmer LG 1995 Permeation and gating properties of a cloned renal K+
channel. American J. Physiol. 268: C389-C401.Christophersen P 1990 Calcium-activated potassium channel from human erythrocyte membranes:
Single channel rectification and selectivity. J. Membrane Biol. 119: 75-84.Cocucci M, Sacchi GA 1993 Effects of rubidium and potassium on potassium efflux from subapical segments
of maize roots. Plant Physiol. and Biochem. Paris 31: 9-16.Conejero C 1997 Anion exchanger AE1 as a candidate pathway for taurine transport in rat erythro-
cytes. Am. J. Physiol. 272: C1457-C1464.Copello J, Simon B, Segal Y, Wehner F, Ramanujam F, Sadagopa VM, Alcock N, Reuss L 1991 Ba2+
release from soda glass modifies single maxi K+ channel activity in patch clamp experiments. Bio-phys. J. 60: 931-941.
Cukierman S 1992 Characterization of potassium currents in rat malignant lymphocytes (Nb2 cells). J.Membrane Biol. 126: 147-157.
Dainty J, Hope AB 1959 Ionic relations of cells of Chara australis. I. Ion exchange in the cell wall.Austr. J. Biol. Sci. 12: 395-411.
Davies JM, Sanders D 1995 ATP, pH and Mg2+ Modulate a cation current in Beta vulgaris vacuoles: apossible shunt conductance for the vacuolar H+-ATPase J. Membrane Biol. 145: 75-86.
De Boer AH, Van Duijn B, Giesberg P, Wegner L, Obermeyer G, Koehler K, Linz KW 1994 Laser mi-crosurgery: A versatile tool in plant (electro) physiology. Protoplasma 178: 1-10.
Delgado R, Latorre R, Labarca P 1995 Selectivity and gating properties of a cAMP-modulated, K+-selective channel from Drosophila larval muscle. FEBS-Letters 370: 113-117.
Dietrich P, Dreyer I, Wiesner P, Hedrich R 1998 Cation sensitivity and kinetics of guard-cell potassiumchannels differ among species. Planta 205: 277-287.
Ding JP, Badot PM, Pickard BG 1993 Aluminium and hydrogen ions inhibit a mechanosensory calcium-selective cation channel. Austr. J. Plant Physiol. 20: 771-778.
Dinudom A, Young JA, Cook DI 1994 Ion channels in the basolateral membrane of intralobular ductcells of mouse mandibular glands. Pfluegers Archiv Eur. J. Physiol. 428: 202-208.
Doyle DA, Cabral JM, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, MacKinnon R 1998 Thestructure of the Potassium Channel: Molecular Basis of K+ Conduction and Selectivity. Science 280:69-77.
Draber S, Schultze R 1994a Correction for missed events based on a realistic model of a detector. Biophys. J.66: 191-201.
Draber S, Schultze R 1994b Detection of jumps in single-channel data containing subductance levels.Biophys. J. 67: 1404-1413.
Drew MC, Läuchli A 1987 The role of the mesocotyl in sodium exclusion from the shoot of Zea mays L.(cv. Pioneer 3906). J. Exp. Bot. 38: 409-418.
Durell SR, Guy HR 1998 Structural models of the transmembrane region of voltage-gated and other K+
channels in open, closed, and inactivated conformations. J. of Structural Biol. 121: 263-284.Eastman PAK, Dengler NG and Peterson CA 1986 Suberized walls in the bundle sheath of grasses of
differing photosynthetic types: impact on apoplastic permeability. Am. J. Bot. 73: 717.

129
Elzenga JTM, Keller CP, Volkenburgh E van, Van Volkenburgh E 1991 Patch clamping protoplastsfrom vascular plants. Method for the quick isolation of protoplasts having a high success rate ofgigaseal formation. Plant-Physiol. 97: 1573-1575.
Evert RF, Eschrich W, Heyser W 1977 Distribution and structure of the plasmodesmata in mesophyll andbundle-sheath cells of Zea mays L. Planta 136: 77-89.
Evert RF, Eschrich W, Heyser W 1978 Leaf structure in relation to solute transport and phloem loading inZea mays L. Planta 138: 279-294.
Evert RF, Botha CEJ, Mierzwa RJ 1985 Free-space marker studies on the leaf of Zea mays L. Proto-plasma 126: 62-73.
Evert RF, Russin WA 1993 Structurally, phloem unloading in the maize leaf cannot be symplastic. De-partment of Botany, University of Wisconsin, Madison, WI 53706, USA. Am. J. Bot. 80: 1310-1317.
Evert RF, Russin WA, Bosabalidis AM 1996 Anatomical and ultrastructural changes associated withsink-to-source transition in developing maize leaves. Intern. J. Plant Sci. 157: 247-261.
Fairley KA, Walker NA 1989. Patch clamping corn protoplasts: Gigaseal frequency is not improved byCongo Red inhibition of cell wall regeneration. Protoplasma 153: 111-116.
Farokhi A, Keunecke M, Hansen UP 1999 The anomalous mole fraction effect in Chara: Gating at the edgeof temporal resolution. Biophys. J. (submitted).
Ferguson AD, Hofmann E, Couton JW, Diederichs K, Welte W 1988 The structure of the potassiumchannel: molecular basis of K+ conduction and selectivity. Science 280: 69-77.
Fisahn,J, Mikschl E, Hansen UP 1986 Separate oscillations of the electrogenic pump and of a K+-channelin Nitella as revealed by simultaneous measurement of membrane potential and of resistance. J. Exp.Bot. 37: 34-47.
Fritz E, Evert RF, Heyser W 1983 Microautoradiographic studies of phloem loading and transport in theleaf of Zea mays L. Planta, 159: 193-206.
Galli A, Defelice LJ, Duke BJ, Moore KR, Blakely RD 1995 Sodium-dependent norepinephrine-inducedcurrents in norepinephrine-transporter-transfected HEK-293 cells blocked by cocaine andantidepressants. J. Exp. Biol. 198: 2197-2212.
Gambale F, Bregante M, Stragapede F, Cantu AM 1996 Ionic channels of the sugar beet tonoplast areregulated by a multi-ion single-file permeation mechanism. J. Membrane Biol. 154: 69-79.
Gassmann W, Schroeder JI 1994 Inward-rectifying K+ channels in root hairs of wheat. A mechanismfor aluminum-sensitive low-affinity K+ uptake and membrane potential control. Plant Physiol.105: 1399-1408.
Gillian-Ridge FP, Duszyk M, French AS 1997 A large conductance, Ca2+-activated K+ channel in ahuman lung epithelial cell line (A549). Biochi. Biophys. Acta 1327: 249-258.
Giromini L, Camattari G, Cerana R, Colombo R 1997 Effect of apoplastic and cytoplasmic pH on inwardand outward rectifying plasma membrane K+ channels of Arabidopsis thaliana cultured cells. J.Plant Physiol. 151: 682-688.
Goldman DE 1943 Potential, impedance and rectification in membranes. J. Gen. Physiol. 27: 37-60.Gollan T, Schurr U, Schulze ED 1992 Stomatal response to drying soil in relation to changes in the xylem
sap composition of Helianthus annuus. I. The concentrations of cations, anions, amino acids in,and pH of, the xylem sap. Plant Cell Environ 15: 551-559.
Gomez-Lagunas F, Armstrong CM 1994 The relation between ion permeation and recovery from inac-tivation of Shaker B K+ channels. Biophys. J. 67: 1806-1815.
Grignon C, Sentenac H 1991 pH and ionic conditions in the apoplast. Annu. Rev. Plant Physiol. PlantMol. Biol. 42:103-28.
Grissmer S, Nguyen AN, Cahalan MD 1993 Calcium-activated potassium channels in resting and acti-vated human T lymphocytes. J. of General Physiol. 102: 601-630.
Gunning BES, Steer MW 1996 Bildatlas zur Biologie der Pflanzenzelle. Gustav Fischer, Stuttgart, Jena,New York.
Hadley RW, Hume JR 1990 Permeability of time-dependent potassium channel in guinea pig ven-tricular myocytes to cesium, sodium, ammonium and rubidium. Am. J. Physiol. 259: H1448-H1454.
Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ 1981 Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflügers Archiv 392: 85-100.
Hanselmann C, Grissmer S 1996 Characterization of apamin-sensitive Ca2+-activated potassium chan-nels in human leukaemic T lymphocytes. J. Physiol. (Cambridge) 496: 627-637.

130
Hansen UP, Slayman CL, Gradmann D, Sanders D, 1984 Comprehensive description of cotransportkinetics by means of simple carriers with ordered binding. In: Environmental Regulation ofMicrobial Metabolism. (Kulaev IS, Dawes EA, Tempest DW, eds.). Academic Press. London, pp.327-332.
Hansen UP, Keunecke M, Blunck R 1997. Gating and permeation models of plant channels. J. Exp. Bot. 48:365-382.
Hattersley PW, Browing AJ 1981 Occurrence of the suberized lamella in leaves of grasses of different photo-synthetic types. I. In parenchymatous bundle sheaths and PCR ("Kranz") sheaths. Protoplasma 109:371-401.
Hedrich R, Barbier-Brygoo H, Felle H, Flügge UI, Lüttge U, Maathuis FJM, Marx S, Prins HBA, Raschke K,Schnabel H, Schroeder JI, Struve I, Taiz L, Ziegler P 1988 General mechanisms for solute transportacross the tonoplast of plant vacuoles. A patch-clamp survey of ion channels and proton pumps.Botanica Acta 101: 7-13.
Hedrich R, Schröder JI 1989 The physiology of ion channels and electrogenic pumps in higher plants. AnnuRev. Plant Physiol. Plant Mol. Biol. 40: 539-569.
Hedrich R, Bregante M, Dreyer I, Gambale F 1995 The voltage-dependent potassium-uptake channelof corn coleoptiles has permeation properties different from other K+ channels. Planta 197: 193-199.
Heginbotham L, Lu Z, Abramson T, MacKinnon R 1994 Mutations in the K+ channels signature se-quence. Biophys. J. 66: 483-491.
Heginbotham L, LeMasurier M, Kolmakova-Partensky L, Miller C 1999 Single Streptomyces lividans K+
channels. Functional asymmetries and sidedness of proton activation. J. Gen. Physiol. 114: 551-559.Herok GH, Millar TJ, Anderton PJ, Martin DK 1998 Characterization of an inwardly rectifying potas-
sium channel in the rabbit superior lacrimal gland. IOVS 39: 308-314.Hille B, Schwarz W 1978 Potassium channels as multi-ion single-file pores. J. Gen. Bio. 72: 409-442.Himpkamp J 1988 Nachweis und Charakterisierung eines H+/Glucose-Cotransportsystems in den
Holzstrahlzellen von Populus balsamifera L. (Salicacae) und seine jahreszeitliche Veränderung.Dissertation Kiel.
Hocking PJ 1980 The composition of phloem exudate and xylem sap from tree tobacco (Nicotiana glaucaGroh). Ann. Bot. (London) [N.S.] 45: 633-643.
Homble F, Fuks B 1990 Quantitative analysis of single potassium ion channels in the tonoplast ofChara corallina: Selectivity and TEA blockade. J. Plant Physiol. 137: 729-733.
Horn R, Marty A 1988 Muscaric Activation of ionic currents measured by a new whole-cell recordingmethod. J. Gen. Physiol. 92: 145-59.
Hoshi T 1995 Regulation of voltage dependence of the KAT1 channel by intracellular factors. J. Gen.Physiol. 105: 309-328.
Hoth S, Dreyer I, Dietrich P, Becker D, Muller-Rober B, Hedrich R 1997 Molecular basis of plant-specific acid activation of K+ uptake channels. Proc. Natl. Acad. Sci. (USA) 94: 4806-4810.
Ilan N, Schwartz A, Moran N 1994 External pH effects on the depolarization-activated K channels inguard cell protoplasts of Vicia faba. J. Gen. Physiol. 103: 807-31.
Ilan N, Moran N, Schwartz A 1995 The Role of Potassium Channels in the Temperature Control ofStomatal Aperture. Plant Physiol. 108: 1161-1170.
Ilan N, Schwartz A, Moran N 1996 External protons enhance the activity of the hyperpolarisation-activated K+ channels in guard cell protoplasts of Vicia faba. J. Membrane Biol. 154: 169-181.
Immke D, Wood M, Kiss L, Korn SJ 1999 Potassium-dependent changes in the conformation of theKv2.1 porassium channel pore. J. Gen. Physiol. 113: 819-836.
Ishikawa T, Cook DI 1995 A Ca2+-activated 23-pSK+ channel in human submandibular gland duct cellline (HSG). Cellular Physiol. and Biochem. 5: 232-242.
Ishimoto H, Kimura J, Matsuoka I, Kumasaka T, Nakanishi H 1995 Mechanism of Ca2+ influx by ex-tracellular ATP in NG108-15 cells. Folia Pharmacol. Japonica 106: 138P-142P.
Kaiho H, Kimura J, Matsuoka I, Kumasaka T, Nakanishi H 1996 ATP-activated nonselective cationcurrent in NG108-15 cells. J. of Neurochemistry 67: 398-406.
Kamouchi M, Van-Den-Bremt K, Eggermont J, Droogmans G, Nilius B 1997 Modulation of inwardlyrectifying potassium channels in cultured bovine pulmonary artery endothelial cells. J. Physiol.Cambridge. 504: 545-556.
Karlson P, Doenecke E, Koolman J 1994 Kurzes Lehrbuch der Biochemie für Mediziner und Naturwissen-schaftler. Georg Thieme Verlag Stuttgart.

131
Katnik C, Adams DJ 1995 An ATP-sensitive potassium conductance in rabbit arterial endothelial cells.J. Physiol. (Cambridge) 485: 595-606.
Keunecke M 1995 Untersuchung des anomalen Mofraktionseffektes an der Grünalge Chara corallina.Diplomarbeit, Kiel.
Keunecke M, Sutter JU, Sattelmacher B, Hansen UP 1997 Isolation and patch clamp measurements ofxylem contact cells for the study of their role in the exchange between apoplast and symplast ofleaves. Plant Soil 196: 239-244.
Keunecke M, Hansen UP 1999a Different pH-dependences of K+ channel activity in bundle sheath andmesophyll cells of maize leaves. Planta (in press).
Keunecke M, Lindner B, Seydel U, Schulz A, Hansen UP 1999b Bundle sheath cells in maize are thelocation of uptake from the xylem. Submitted.
Ketchum KA, Poole RJ 1991 Cytosolic calcium regulates a potassium current in corn (Zea mays)protoplasts. J. Membr. Biol. 119: 277-88.
Kirsch GE, Drewe JE, Taglialatela M, Joho RH, DeBiasi M, Hartmann HA, Brown AM 1992 S singlenonpolar residue in the deep pore of related K+channels acts as a K+:Rb+ conductance switch. Biophys.J. 62: 136-144.
Klieber HG, Gradmann D 1993 Enzyme kinetics of the prime potassium channel in the tonoplast ofChara: Selectivity and inhibition. J. Membrane Biol. 132: 253-265.
Kolesnikov SS, Margolskee RF 1998 Extracellular K+ activates a K+ - and H+-permeable conductancein frog taste receptor cells. J. Physiol. Cambridge 507: 415-432.
Kolesnikov SS, Kosolapov AV 1993 Cyclic nucleotide-activated channels in carp olfactory receptorcells. Biochimica Biophysica Acta 1150: 63-72.
Köhler J, Fritz E, Orlich G, Komor E 1991 Microautoradiographic studies of the role of mesophyll andbundle tissues of the Ricinus cotyledon in sucrose uptake. Planta (Heidelberg) 183: 251-257.
Köhler B, Raschke K 1999 The delivery of salts to the xylem: three types of anion conductance in theplasmalemma of the xylem parenchyma of roots of Hordeum vulgare L. Plant Physiol. (in Druck)
Koolman J, Röhm KH 1994 Taschenatlas der Biochemie. Georg Thieme Verlag Stuttgart, 22. Auflage.Korbmacher C, Volk T, Segal AS, Boulpaep EL, Froemter E 1995 A calcium-activated and nucleotide-
sensitive nonselective cation channel in M-1 mouse cortical collecting duct cells. J. MembraneBiol. 146: 29-45.
Kourie JI 1996a. Voltage-dependence of extracellular Ca2+-induced modification in properties of the inwardrectifying K+ channels in the plasma membrane of mesophyll protoplasts of Avena sativa. Australian J.of Plant Physiol. 23: 349-359.
Kourie JI 1996b. Interaction of extracellular potassium and cesium with the kinetics of inward rectifying K+
channels in the plasma membrane of mesophyll protoplasts of Avena sativa. Plant and Cell Physiol. 37:770-781.
Kreusch A, Schulz GE 1994 Refined Structure of the porin from Rhodopseurodomonas blastica.Comparison with the porin from Rhodobacter capsulantus. J. Mol. Biol. 243: 891-905.
Krömer S 1995 Respiration during photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46: 45-70.Kunert M 1998 Aufnahme und Auswertung von Strom/Spannungskurven mit Rampenfunktionen bei
Patch-Clamp Messungen. Diplomarbeit, Kiel.Kurnikova MG, Coalson RD, Graf P, Nitzan A 1999 A lattice relaxation algorithm for three-dimensional
Poisson-Nernst-Planck theory with application to ion transport through the gramicidin A channel.Biophys. J. 76: 642-656.
Langmann D 1990 Dalanco Spry Model 250 Data Acquisition and Signal Processing Board Documentation.Rochester, NY.
Larsson HP, Baker OS, Dhillon DS, Isacoff EY 1996 Transmembane movement of the Shaker K+ channelS4. Neuron 16: 287-397.
Läuger P 1980 Kinetic properties of ion carriers and channels. J. Membrane Biol. 57: 163-178.Laver DR, Fairley-Grenot KA 1994 Surface potentials near the mouth of the large-conductance K+
channel from Chara australis: a new method of testing for diffusion-limited ion flow. J. MembraneBiol. 139: 149-65.
Laver DR, Walker NA 1991 Activation of calcium ion and block by divalent ions of the potassium ionchannel in the membrane of cytoplasmic drops from Chara australis. J. Membrane Biol. 120:131-140.

132
Leonhardt N, Marin E, Vavasseur A, Forestier C 1997 Evidence for the existence of a sulfonylurea-receptor-like protein in plants: Modulation of stomatal movements and guard cell potassium channels by sulfo-nylureas and potassium channel openers. Proc. Natl. Acad. Sci. (USA) 94: 14156-14161.
Levitt DG 1999 Modeling of ion channels. J. Gen. Physiol. 113: 789-794.Linz KW, Köhler K 1993. Isolation of protoplasts from the coccal green alga Eremosphaera viridis De
Bary for patch-clamp measurements. Botanica Acta 106: 469-472.Linz KW, Köhler K 1994 Vacuolar ion currents in the primitive green alga Eremosphera viridis: the electri-
cal properties of both the characeae and higher plants. Protoplasma 179: 34-45.Lohaus G, Winter H, Riens B, Heldt HW 1995 Further studies of the phloem loading process in leaves of
barley and spinach. The comparison of metabolite concentrations in the apoplastic compartmentwith those in the cytosolic compartment and in the sieve tubes. Bot. Acta 108: 270-275.
Lopez S, Murison SD, Travis AJ, Chesson A 1993 Degradability of parenchyma and sclerenchyma cellwalls isolated at different developmental stages from a newly extended maize internode. Acta Bot.Neerlandica 42: 165-174.
Lucas WJ 1997 Application of microinjection techniques to plant nutrition. In: Ando T, Fujita K, Mae T,Matsumoto H, Mori S, Sekiya J (editors) Proceedings of the XIII International Plant NutritionColloquium, Tokio.
Lühring H 1999 pH-sensitive gating kinetics of the maxi-K channel in the tonoplast of Chara australis. J.Membrane Biol. 168: 47-61.
Lux HD, Carbone E, Zucker H 1990 Sodium ion currents through low-voltage-activated calcium ionchannels of chick sensory neurons: Block by external calcium ion and magnesium ion. J. Physiol.(Cambridge) 430: 159-188.
MacRobbie EAC 1995 Effects of ABA on 86Rb+ fluxes at plasmalemma and tonoplast of stomatal guardcells. Plant J. 7: 835-843.
Maathuis FJM, Sanders D 1992 Plant membrane transport. Current Opinion in Cell Biology 4: 661-669.Maathuis JM, Sanders D 1997 Regulation of K+ absorption in plant root cells by external K+: interplay
of different plasma membrane K+ transpoters. J. Exp. Bot. 48: 451-458.Maathuis FJM, Sanders D, Gradmann D 1997 Kinetics of high-affinity K+ uptake in plants, derived
from K+-induced changes in current-voltage relationships: A modelling approach to the analysisof carrier-mediated transport. Planta (Heidelberg) 203: 229-236.
Marschner H 1995 Mineral nutrition of higher plants. Academic Press. Harcout Brace & Company,Publishers.
Marten I, Hoth S, Deeken R, Ache P, Ketchum KA, Hoshi T, Hedrich R 1999 AKT3, a phloem-localizedK+ channel, is blocked by protons. Proc. Natl. Acad. Sci. (USA) 96: 7581-7586.
Matsuda H 1996 Rb+, Cs+ ions and the inwardly rectifying K+ channels in guinea-pig ventricular cells.Pfluegers Archiv Eur. J. Physiol. 432: 26-33.
Matsuoka S, Ehara T, NOMA A 1990 Chloride-sensitive nature of the adrenaline-induced current inguinea-pig cardiac myocytes. J. Physiol. (London) 425: 579-598.
Mauerer UR, Boulpaep EL, Segal AS 1998 Properties of an inwardly rectifying ATP-sensitive K+
channel in the basolateral membrane of renal proximal tubule. J. Gen. Physiol. 111: 139-160.Möller C 1998 Transportmodelle zur Analyse von Patch-Clamp Daten. Diplomarbeit, Kiel.Moran N, Fox D, Satter RL 1990 Interaction of the depolarization-activated K+ channel of Samanea
saman with inorganic ions: a patch-clamp study. Plant-Physiol. 94: 424-431.Moran N 1996 Membrane-delimited phosphorylation enables the activation of the outward-rectifying K
channels in motor cell protoplasts of Samanea saman. Plant Physiol 111: 1281-1292.Mühling KH, Plieth C, Hansen UP, Sattelmacher B 1995 Apoplastic pH of intact leaves of Vicia faba as
influenced by light. J Exp. Bot. 46: 377-382.Müller-Röber B, Ellenberg J, Provart N, Willmitzer L, Busch H, Becker D, Dietrich P, Hoth S,
Hedrich R 1995 Cloning and electrophysiological analysis of KST1, an inward rectifying K+
channel expressed in potato guard cells. EMBO J. 14: 2409-2416.Noctor G, Foyer CH 1998 A re-evaluation of the ATP: NADPH budget during C3 photosynthesis: a
contribution from nitrate assimilation and its associated respiratory activity? J. Exp. Bot. 49: 1895-1908.Nonaka T, Matsuzaki K, Kawahara K, Suzuki K, Hoshino M 1995 Monovalent cation selective chan-
nel in the apical membrane of rat inner medullary collecting duct cells in primary culture. Bio-chimica et Biophysica Acta 1233: 163-174.
Nonner W, Eisenberg B 1998 Ion permeation and glutamate residues linked by Poisson-Nernst-PlanckTheory in L-type calcium channels. Biophys. J. 75: 1287-1305.

133
Nonner W, Chen DP, Eisenberg B 1998 Anomalous mole fraction effect, electrostatics and binding inionic channels. Biophys. J. 74: 2327-2334.
Nonner W, Chen DP, Eisenberg B 1999 Progress and prospects in permeation. J. Gen. Physiol. 113: 773-782.
Noodén LD, Mauk CS 1987 Changes in the mineral composition of soybean xylem sap duringmonocarpic senescence and alterations by podding. Physiol. Plant. 70: 735-742.
Numberger M, Draguhn A 1996 Patch-Clamp-Technik. Spektrum Akademischer Verlag. Heiderlberg, Berlin,Oxford.
Ogielska EM, Aldrich RW 1998 A mutation in S6 of Shaker potassium channels decreases the K+ affinity ofan ion binding site revealing ion-ion interactions in the pore. J. Gen. Physiol. 112: 243-257.
Ogielska EM, Aldrich RW 1999 Functional consequences of a decreased potassium affinity in a potassiumchannel pore: Ion interaction and C-type inactivation. J. Gen. Physiol. 113: 347-358.
Pardo LA, Heinemann SH, Terlau H, Ludewig U, Lorra C, Pongs O, Stuehmer W 1992 Extracellularpotassium modulates a rat brain potassium channel. PNAS 89: 2466-2470.
Park Y 1994 Ion selectivity and gating of small conductance Ca2+-activated K+ channels in cultured ratadrenal chromaffin cells. J. Physiol. (Cambridge) 481: 555-570.
Pate JS, Wallace W, Die van J 1964 Petiole bleeding sap in the examination of the circulation of introgenoussubstances in plants. Nature (London) 204: 1073-1074.
Perez CP, Begenisich T 1994 The multi-ion nature of the pore in Shaker K+ channels. Biophys. J. 66:1929-1938.
Philippar K, Fuchs I, Lüthen H, Hoth S, Bauer CS, Haga K, Thiel G, Ljung K, Sandberg G, Böttger M,Becker D, Hedrich R 1999 Auxin-induced K+ channel expression represents an essential step incoleoptile growth and gravitropism. PNAS 96: 12186-12191.
Plieth C 1995 Bestimmung von Ionenkonzentrationen und ihrer Änderungen in grünen pflanzlichenZellen und Geweben mit Hilfe der bildverarbeitenden ratiometrischen Fluoreszenz-Mikroskopie undder Laser-Doppler-Anemometrie. Dissertation Kiel.
Plieth C, Sattelmacher B, Hansen UP 1997 Cytoplasmic Ca2+-H+ exchange buffers in green algae.Protoplasma 198: 107-124.
Plieth C, Sattelmacher B, Hansen UP 1998 Light-induced cytosolic calcium transients in green plantcells. II. The effect on a K+ channel as studies by a kinetic analysis in Chara corallina. Planta 207:52-59.
Plieth C, Hansen UP, Knight H, Knight MR 1999a Temperature sensing by plants: the primary characteristicsof signal perception and calcium response. The Plant Journal 18: 491-497.
Plieth C, Sattelmacher B, Hansen, UP, Knight MR 1999b Low-pH-mediated elevations in cytosolic calciumare inhibited by aluminium: a potential mechanism for aluminium toxidity. The Plant Journal 18: 643-650.
Pottosin II, Andjus PR, Vucelic D, Berestovsky GN 1993 Effects of D-2O on permeation and gating inthe Ca2+-activated potassium channel from Chara. J. Membrane Biol. 136: 113-124.
Randle JCR, Oliet SHR, Renaud JF 1991 Alkali cation permeability and cesium blockade of cromaka-lim-activated current in guinea-pig ventricular myocytes. British J. Pharmacol. 103: 1795-1801.
Rains DW 1969 Cation absorption by slices of stem tissues of bean and cotton. Experimenta 25: 215-216.Riens B, Lohaus G, Heineke D, Heldt HW 1991 Amino acid and sucrose content determined in the
cytosolic, chloroplastic and vacuolar compartments and in the phloem sap of spinach leaves. PlantPhysiol. 97: 227-233.
Roberts SK, Tester M. 1995. Inward and outward K+-selective currents in the plasma membrane ofprotoplasts from maize root cortex and stele. Plant-J. 8: 811-825.
Robson L, Hunter M 1997 Two K+-selective conductances in single proximal tubule cells isolated fromfrog kidney are regulated by ATP. J. Physiol. (Cambridge) 500: 605-616.
Russell SH, Evert RF 1985 Leaf vasculature in Zea mays L. Planta 164: 448-458.Russin WA, Evert RF, Vanderveer PJ, Sharkey TD, Briggs SP 1996 Modification of a specific class of
plasmodesmata and loss of sucrose export ability in the sucrose export defective1 maize mutant.Plant Cell 8: 645-658.
Safronov BV, Vogel W 1995 Modulation of delayed rectifier K+ channel activity by external K+ ionsin Xenopus axon. Pfluegers Archiv Eur. J. of Physiology 430: 879-886.
Saimi Y, Martinac B 1989 Calcium-dependent potassium channel in Paramecium studied under patchclamp. J. Membrane Biol. 112: 79-90.

134
Saitow F, Nakaoka Y, Oosawa Y 1997 A calcium-activated, large conductance and non-selective ca-tion channel in paramecium cell. Biochi. Biophys. Acta 1327: 52-60.
Sakman B, Neher E (Editors) 1985 Single channel recording. Plenum Press, New York und London.Sakman B, Neher E 1995 Geometric parameters of pipettes and membrane patches. Single channel
recording, Plenum Press, New York and London, pp 637-62.Sala S, Parsey RV, Cohen AS, Matteson DR 1991 Analysis and use of the perforated patch technique for
recording ionic currents in pancreatic ß-cells. J. Membrane Biol. 122:177-87.Sanders D, Hansen UP 1981 Mechanism of C1- Transport at the plasma membrane of Chara corallina. II.
Transinhibition and the determination of H+/C1- binding order from a reaction kinetic model. J.Membrane Biol. 58: 139-153.
Sanders D 1988 Solute transport in plant cells and tissues. Monographs and surveys in the biosciences.Wilkens M (ed.) Longman Scientific & Technical.
Sansom MS, Sadcock C, Smith GR 1998 Modelling and simulation of ion channels: applications to thenicotinic acetylcholine receptor. J. Struct. Biol. 121: 246-262.
Schauf CL, Wilson JJ 1987 Effects of abscisic acid on K+ channels in Vicia faba L. guard cell protoplasts.Biochem. Biophys. Res. Comun. 145: 284-290.
Schlatter E, Lohrmann E, Greger R 1992 Properties of the potassium conductances of principal cells ofrat cortical collecting ducts. Pfluegers Archiv Eur. J. of Physiology 420: 39-45.
Schnepf E 1977 Bau der Zelle (Prokaryonten, Eukaryonten). Biophysik. Herausgeber W. Hoppe.Schoppa NE, McCormack K, Tanouye ME, Sigworth FJ 1992 The size of gating charge in wild type and
mutant Shaker potassium channels. Science 255: 1712-1715.Schroeder JI, Ward JM, Gassmann W 1994 Perspectives on the physiology and structure of inward-
rectifying K+ channels in higher plants: Biophysical implications for K+ uptake. Annu. Rev.Biophys. Biomol. Struct. 23: 441-447.
Schurr U, Schulze ED 1996 Effects of drought on nutrient and ABA transport in Ricinus communis. PlantCell Environ 19: 665-674.
Seimer C, Goegelein H 1992 Activation of nonselective cation channels in the basolateral membraneof rat distal colon crypt cells by prostaglandin E-2. Pfluegers Archiv Eur. J. Physiol. 420: 319-328.
Seoh SA, Sigg D, Papzian DM, Bezanilla F 1996 Voltage sensing residues in the S2 and S4 segments ofthe Shaker K+ channel. Neuron 16: 1159-1167.
Seydel U, Haas M, Rietschel ET, Lindner B 1992 Laser microprobe mass spectrometry of individual bacterialorganisms and of isolated bacterial compounds: a tool in microbiology. J. Microbiol. Methods 15: 167-183.
Sigg D, Stefani E, Bezanilla F 1994 Gating current noise produced by elementary transitions in Shakerpotassium channels. Protein Engin. 8: 397-401.
Silver MR, Shapiro MS, Decoursey TE 1994 Effects of external Rb+ on inward rectifier K+ channels ofbovine pulmonary artery endothelial cells. J. Gen. Physiol. 103: 519-548.
Singer SJ, Nicolson GL 1972 The fluid mosaic model of cell membranes. Science 173: 720-731.Sitte P, Ziegler H, Ehrendorfer F, Bresinsky A 1998 Strasburger Lehrbuch der Botanik. Gustav Fischer
Verlag.Skerrett M, Tyerman SD 1994 A channel that allows inwardly directed fluxes of anions in protoplasts
derived from wheat roots. Planta 192: 295-305.Spanswick RM, Stolarek J, Williams EJ 1967 The membrane potential of Nitella translucens. J. Exp. Botany
18: 1-16.Starkus JG, Kuschel L, Rayner MD, Heinemann SH 1997 Ion conduction through C-type inactivated Shaker
channels. J. Gen. Physiol. 110: 539-550.Starkus JG, Kuschel L, Rayner MD, Heinemann SH 1998 Macroscopic Na+ currents in the "nonconducting"
Shaker potassium channel mutant W434F. J. Gen. Physiol. 112: 85-93.Steinert M, Grissmer S 1997 Novel activation stimulus of chloride channels by potassium in human
osteoblasts and human leukaemic T lymphocyte. J. Physiol. (Cambridge) 500: 653-660.Stockand JD, Sansom SC 1994 Large Ca2+-activated K+ channels responsive to angiotensin II in cul-
tured human mesangial cells. American J. Physiol. 267: C1080-C1086.Stoeckel H, Takeda K 1989 Calcium-activated, voltage-dependent, non-selective cation currents in
endosperm plasma membrane from higher plants. Proc. Royal Soc. London B Biol. Sci. 237: 213-231.

135
Suenaga A, Komeiji Y, Uebayasi M, Meguro T, Saito NM, Yamato I 1998 Computational observation ofan ion permeation through a channel protein. Biosci. Rep. 18: 39-48.
Sugihara I 1994 Calcium-activated potassium channels in goldfish hair cells. J. Physiol. (Cambridge)476: 373-390.
Supplisson S, Loo DDF, Sachs G 1993 Whole-cell currents in isolated resting Necturus gastric oxynti-copeptic cells. J. Physiol. (Cambridge) 463: 57-82.
Sutter JU 1996 Biophysikalische Untersuchungen an Xylemkontaktzellen: Präparation, Meßtechnik,Transporteigenschaften und Modellentwicklung. Diplomarbeit, Kiel.
Sze H 1985 H+ translocating ATPases: advances using membrane vesicles. Annual Rev. Plant Physiol. 36:175-208.
Tabcharani JA, Harris RA, Boucher A, Eng JWL, Hanrahan JW 1994 Basolateral K channel activatedby carbachol in the epithelial cell line T-84. J. Membrane Biol. 142: 241-254.
Takeuchi S, Ando M, Kozakura K, Saito H, Irimajiri A 1995 Ion channels in basolateral membrane ofmarginal cells dissociated from gerbil stria vascularis. Hearing Research 83: 89-100.
Thiel G, Blatt, MR 1991 The mechanism of ion permeation through K+ channels of stomatal guard cells:voltage-dependent block by Na+. J. Plant Physiol. 138: 326-334.
Thomine S, Zimmermann S, Guern J, Brabiergrygoo H 1995 ATP-dependent regulation of an anion channelat the plasma membrane of protoplasts from the epidermal cells of Arabidopsis hypocotyls. Plant Cell 7:2091-2100.
Titievsky AV, Takeo T, Tepikin AV, Petersen OH 1996 Decrease of acidity inside zymogen granulesinhibits acetylcholine- or inositol trisphosphate-evoked cytosolic Ca2+ spiking in pancreatic acinarcells. Pfluegers Archiv Eur. J. Physiol. 432: 938-940.
Tittor J, Hansen UP, Gradmann D 1983 Impedance of the electrogenic Cl- pump in Acetabularia; electir-cal frequency entrainements, voltage sensitivity and reaction kinetic interpretation. J. Membr. Biol.75: 129-139.
Trapp S, Tucker SJ, Ashcroft FM 1998 Mechanism of ATP-sensitive K channel inhibition by sulfhydrylmodification. J Gen Physiol 112: 325-332.
Trequattrini C, Petris A, Franciolini F 1996 Characterization of a neuronal delayed rectifier K currentpermeant to Cs and blocked by verapamil. J. Membrane Biol. 154: 143-153.
Truernit E, Stadler R, Baier K, Sauer N 1999 A male gametophyte-specific monosaccharide transporterin Arabidopsis. Plant-Journal 17: 191-201.
Tsuzuki K, Mochizuki S, Iino M, Mori H, Mishina M, Ozawa S 1994 Ion permeation properties of thecloned mouse epsilon-2/zeta-1 NMDA receptor channel. Mol. Brain Research 26: 37-46.
Tucker EB 1990 Calcium loaded 1,2-bis(2-aminophenoxy)ethane-N, N, N, N’, N’-tetraaceticacid blockscell-to-cell diffusion of carbofluorescein in staminal hairs of Setcreasea purpurea. Planta 182: 34-38.
Tyerman SD, Skerrett M, Garrill A, Findlay GP, Leigh RA, Sanders D 1997 Pathways for the permeation ofNa+ and Cl- into protoplasts derived from the cortex of wheat roots. J. Exp. Botany 48: 459-480.
Umrath K 1932 Die Bildung von Plasmalemma (Plasmahaut) bei Nitella mucronata. Protoplasma 16:173-188.
Vanselow KH, Kolbowski J, Hansen UP 1989 Further evidence for the relationship betweenlight-induced changes of plasmalemma transport and transthylakoid proton uptake. J. Exp. Bot. 40:239-245.
Virkki LV, Nikinmaa M 1996 Conductive ion transport across the erythrocyte membrane of lamprey(Lampetra fluviatilis) in isotonic conditions is mainly via an inwardly rectifying K+ channel.Comparative Biochem. Physiol. A 115: 169-176.
Vogelzang SA, Prins HBA 1992 Plasmalemma patch clamp experiments in plant root cells: procedure forfast isolation of protoplasts with minimal exposure to cell wall degrading enzymes. Protoplasma171: 104-109.
Welling PA 1997 Primary structure and functional expression of a cortical collecting duct Kir channel.Am. J. Physiol. 273: F825-F836.
Wegner LH, Raschke K 1994 Ion channels in the plasmalemma of xylem parenchyma cells from barleyroots: a procedure to isolate protoplasts from this tissue and a patch-clamp exploration of saltpassageways into xylem vessels. Plant Physiol. 105: 799-813.
Wegner LH, De Boer AH, Raschke K 1995 Properties of the K+ inward rectifier in the plasma membraneof xylem parenchya cells from barley roots: Effects of TEA+, Ca2+, Ba2+ and La3+. J. MembraneBiol. 142: 363-379.

136
White PJ, Lemtiri-Chlieh F 1995 Potassium currents across the plasma membrane of protoplasts derivedfrom rye roots: a patch-clamp study. J. Exp. Bot. 46: 497-511.
White PJ 1997 Cation channels in the plasma membrane of rye roots. J. of Exp. Botany 48: 499-514.Winter H, Lohaus G, Heldt HW 1992 Phloem transport of amino acids in relation to their cytosolic
levels in barley leaves. Plant Physiology 99: 996-1004.Wu WH, Assmann SM 1995. Is ATP required for K+ channel activation in Vicia guard cells? Plant Physiol.
107: 101-10.Yamamoto D, Suzuji N 1989 Characterization of single non-inactivating potassium channels in pri-
mary neuronal cultures of Drosophila. J. Exp. Biol. 145: 173-184.Yang XC, Sachs F 1990 Characterization of stretch-activated ion channels in Xenopus oocytes. J.
Physiol. (Cambridge) 431: 103-122.Yang PK, George AI Jr, Horn R 1996 Molecular basis of charge movement in voltage gated sodium
channels. Neuron 16: 113-122.Zhou H, Tate SS, Palmer LG 1994 Primary structure and functional properties of an epithelial K chan-
nel. American J. Physiol. 266: C809-C824.Zhu Y, Ikeda SR 1993 Anomalous permeation of sodium through a putative potassium channel in rat
superior cervical ganglion neurones. J. Physiol. (Cambridge) 468: 441-461.Zimmermann S, Thomine S, Guern J, Barbier-Brygoo H 1994 An anion current at the plasma membrane of
tabacco protoplasts shows ATP-dependent voltage-regulation and is modulated by auxin. Plant J. 6:707-716.
Zimmermann S, Talke I, Ehrhardt T, Nast G, Muller-Rober B 1998 Characterization of SKT1, an in-wardly rectifying potassium channel from potato, by heterologous expression in insect cells. PlantPhysiol. 116: 879-890.
Zitate aus den Berichten zu den DFG-Schwerpunktstagungen “Der Apoplast der höheren Pflanze:Speicher-, Transport- und Reaktionsraum”
Canny MJ 1998 Transport of water and ions in the xylem and apoplast of leaves. Vortrag bei der DFG-Tagung “Der Apoplast der höheren Pflanze: Speicher-, Transport- und Reaktionsraum” in Würzburg.
Felle HH, Peters WS 1996 Apoplastisches Ionenmilieu bei Zea mays. In: Kurzfassungen der Vorträge zum 1.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 6.-8. November 1996 in Kiel.
Hecht-Buchholz C, Höflich G 1997 Bedeutung von phytoeffektiven Bakterien im Interzellularraum von Maisfür die Stickstoffernährung. In: Kurzfassungen der Vorträge zum 2. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 23.-24. Oktober 1997 in Hannover.
Heyser W 1997 Die Bedeutung des Apoplasten bei Nährstoffaufnahme und –transport in mycorrhizierten undnicht mycorrhizierten Wurzeln der Kiefer und Pappel. In: Kurzfassungen der Vorträge zum 2.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 23.-24. Oktober 1997 in Hannover.
Köhler B, Wegner LH, Raschke K 1999 Apoplastisches Nitrat modifiziert die Schalteigenschaften von X-QUAC. In: Kurzfassungen der Vorträge zum 4. Berichtskolloquium des DFG-Schwerpunktes Apoplast,11.–13. November 1999 in Bonn.
Lohaus G, Hußmann M, Heldt HW 1996 Arbeitsbericht. In: Kurzfassungen der Vorträge zum 1.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 6.-8. November 1996 in Kiel.
Lohaus G, Hußmann M, Heldt HW 1997 Die Bedeutung des Phloemrücktransportes im Zusammenhang mitder Nährstoffauswaschung aus dem Blattapoplasten. In: Kurzfassungen der Vorträge zum 2.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 23.-24. Oktober 1997 in Hannover.
Lohaus G, Hußmann M, Heldt HW 1998 Die Bedeutung des Phloemrücktransportes im Zusammenhang mitder Nährstoffauswaschung aus dem Blattapoplasten. In: Kurzfassungen der Vorträge zum 3.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 12.-14. November 1998 in Würzburg.
Pennewiß K, Mühling KH, Sattelmacher B 1997 Bedeutung des Apoplasten als Zwischenspeicher für Ionenunter besonderer Berücksichtigung der Nährstoffauswaschung. In: Kurzfassungen der Vorträge zum 2.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 23.-24. Oktober 1997 in Hannover.
Pennewiß K, Sattelmacher B 1998 Charakterisierung der Ionenverhältnisse im Apoplasten von Blöttern unterbesonderer Berücksichtigung der Nährstoffauswaschung. In: Kurzfassungen der Vorträge zum 3.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 12.–14. November 1998 in Würzburg.

137
Plieth C, Sattelmacher B, Hansen UP 1999c Determination of [Ca2+]apo and hints of its homeostasis. In:Kurzfassungen der Vorträge zum 4. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 11.–13.November 1999 in Bonn.
Raschke K, Köhler B, Peters W 1998 Beladung und Entladung des Xylems. In: Kurzfassungen der Vorträgezum 3. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 12.–14. November 1998 in Würzburg.
Sattelmacher B, Mühling KH, Pennewiß C 1996 Charakterisierung der Ionenverhältnisse im Apoplasten vonBlättern unter besonderer Berücksichtigung der Nährstoffauswaschung. In: Kurzfassungen der Vorträgezum 1. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 6.-8. November 1996 in Kiel.
Schreiber L, Hartmann K 1998 Einfluß der Ernährung auf die apoplastischen Transportbarrieren derWurzeln Höherer Pflanzen. In: Kurzfassungen der Vorträge zum 3. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 12.–14. November 1998 in Würzburg.
Schulz S, Boyle C, Götz M, Dammann U, Mertens E 1998 Einfluß apoplastisch wachsenderendophytischer und pathogener Pilze auf den Ernährungszustand ihrer Wirte. In: Kurzfassungen derVorträge zum 3. Berichtskolloquium des DFG-Schwerpunktes Apoplast, 12.–14. November 1998 inWürzburg.
Türk S, Fink S 1997 Die Bedeutung des Apoplasten als Transport- und Reaktionsraum für dieKompartimentierung von Calciumoxalat bei Koniferen. In: Kurzfassungen der Vorträge zum 2.Berichtskolloquium des DFG-Schwerpunktes Apoplast, 23.-24. Oktober 1997 in Hannover.

138
Bedanken möchte ich mich bei:
Ulf-Peter Hansen, der mich hervorragend betreute, mir die DFG-Stellegab, und mir bei der Bearbeitung viel Freiräume ließ, ohne meine Arbeit ausdem Blick zu verlieren;Jan Abshagen, der auch in den Tiefs meine Launen ertrug und mir wieder zuneuen Kräften verhalf und der immer für Diskussionen offen war;meiner Mutter, die mir vormachte, wie man mit viel Geduld beim Patchen zuschönen Ergebnissen kommt;meinem Vater, der mich sehr früh mit der Laborumgebung vertraut machte;Katrin Schinner, die mit mir Zimmer, Freuden und Sorgen und die Schoko-leidenschaft teilte;meinem Bruder Arne, Sabine Berg und Julia Baade, die besonders überEmail Hilfestellungen in allen Lebenslagen gaben;Justus Krisch, der mich spielerisch auf neue Ideen brachte;Hanni Härtel, Mirko Goschütz, Anja Kinny und Tom Janneck, die meine Ab-wehrkräfte stärkten;Gethyn Allen, der mir einen Einblick in kalifornische Arbeits- und Lebens-verhältnisse gab;Thilo Rießner, Uwe Kirst und Oliver Radomski, die die Programme zur Da-tenauswertung erweiterten und mir Antworten auf viele Computerfragengaben;Michael Kunert und Clemens Möller, die nicht an Mais verzweifelten;Afshin Farokhi und Tobias Huth, die mit und ohne Klimagerät für ein nettesKlima beim Patchen sorgten;Roy Bendixen, der in seinen Vorträgen versuchte, uns auf das Leben vorzu-bereiten;Barbara Köhler, die mir bei der Entwicklung der Whole-Cell-Präparationhalf;Frau Thiessen, die durch die gewissenhafte Anzucht der Maispflanzen die-se Arbeit ermöglichte;Burkhard Sattelmacher, der den DFG-Schwerpunkt einrichtete und für unsdie Pflanzen anzüchten ließ;Alexander Schulz, der mir den Umgang mit dem Cryoverfahren beibrachte,und durch seine Hilfestellung und Diskussion die Tracerarbeiten erst er-möglichte;

139
Ulrich Seydel, der mir in seinem Seminar und im Praktikum LAMMA näher-brachte und mich in Borstel messen ließ;Buko Lindner, der mich in die Geheimnisse des LAMMAs einwiesallen, die zu den netten Teerunden beitrugen;und insbesondere meinem super Team bestehend aus meinen Eltern, SabineBerg, Katrin Schinner, Jan Abshagen, Karl Heine, Anja, Urmel und Alexan-der Kinny, Jessica Spors und Julia Baade für’s Korrekturlesen.

Lebenslauf
Angaben zur Person:Name: Maike KeuneckeGeboren: 13.01.1971 in KielFamilienstand: ledig, keine KinderStaatsangehörigkeit: deutsch
Ausbildung:1977-1981 Besuch der Grundschule Strohbrück1981-1990 Besuch des Gymnasiums Kronshagen
Austauschschülerin in: Finnland (Helsinki), Frankreich (Rennes),Israel (Tel Aviv) und Norwegen (Askim)
1990-1995 Physikstudium an der Christian-Albrechts-Universität zu KielVordiplom, Note: gutAbschluß mit Diplom, Note: sehr gutTitel: Untersuchung des anomalen Molfraktionseffektes an derGrünalge Chara corallina.
1995-1999 Promotionsstudium an der Christian-Albrechts-Universität zuKiel
![Visualizing Loops and Data Structures in Xylem: The …ejw/papers/VisualizingLoopsandData...Visualizing Loops and Data Structures in Xylem: ... objectives [16]; this work ... Xylem](https://img.dokumen.tips/doc/110x75/5ae3d7807f8b9a595d8ef8ac/visualizing-loops-and-data-structures-in-xylem-the-ejwpapersvisualizingloopsanddatavisualizing.jpg)




























