Embed Size (px)
Citation preview

UNIVERSITÀ DEGLI STUDI DI TORINO
Facoltà di Agraria
Corso di laurea specialistica in Biotecnologie agrarie vegetali
TESI DI LAUREA DI II LIVELLO
Sviluppo di marcatori microsatellite e costruzione di una mappa
genetica intra-specifica in Capsicum annuum L. .
Relatore: Prof. SERGIO LANTERI Correlatore: Dott. EZIO PORTIS
Candidato: SARA FALVO
Anno Accademico 2005/2006

II
INDICE
I-INTRODUZIONE………………………………………………….1
1. IL PEPERONE…………………...……………….…………………1
1.1 Origine e diffusione………………………..………………………....1 1.2 Importanza economica…………………………………………...……2 1.3 Classificazione………………………………………………………3
1.3.1 Gruppo Capsicum pubescens…………………………………….....5 1.3.2 Gruppo Capsicum baccatum……………………………………….6 1.3.3 Gruppo Capsicum annuum…………………………………...……8
1.4 Caratteristiche botaniche del gruppo Capsicum anuum L…………………….10 1.5 Caratteristiche organolettiche e nutrizionali……………………………….13 1.6 Varietà tradizionali…………………………………..………………19
1.7 Possibili utilizzi di Capsicum…………………………………………..22
2. MAPPE…………………………………………………………..…25
2.1 Marcatori molecolari: breve excursus……………………………………25
2.1.1 I Microsatelliti…………………………………………………28 2.1.2 I dCAPS………………………………………………………30
2.2 Le mappe genetiche…………………………………………….……32 2.2.1 Test a due punti………………………………...………………34 2.2.2 Test a tre punti…………………………………………………35 2.2.3 Interferenza e Funzione di Kosambi………………………………...37 2.2.4 LOD score………………………………………………….…38
2.3 Popolazioni segreganti di mappaggio……………………………………39 2.3.1 Back-Cross (o reincrocio, BC1)…………………………………….39 2.3.2 La F2…………………………………………………………40
2.3.3 Recombinant Inbred Lines (RILs)………………………………..…41 2.3.4 Di-Haploids (DH)………………………………………………42 2.3.5 Pseudo-Test-Cross (F1)…………………………………..………42
2.4 Software per la costruzione di mappe genetiche……………………………44

III
3. MARCATORI E MAPPE GENETICO MOLECOLARI SVILUPPATE
IN Capsicum spp.………………………….……………………….…..45
3.1 Mappe genetico molecolari inter-specifiche……………………….……....49
3.1.1 Livingstone et al. (1999): “Genome Mapping in Capsicum and the Evolution of
Genome Structure in the Solanaceae” Genetics……………………………...…..50
3.1.2 Kang et al. (2001): “An interspecific (Capsicum annuum x C. chinense) F2
linkage map in pepper using RFLP and AFLP markers.” Theoretical and Applied
Genetics………………………………………………………...………54
3.1.3 Lee et al. (2004): “Characterization and molecular genetic mapping of
microsatellite loci in pepper.” Theoretical and Applied Genetics………………….....57
3.1.4 Sasvári et al. (2004): “Construction of a new interspecific genetic map in pepper
based on AFLP and microsatellite markers.” A.B. Center, Gödöllõ, Ungheria.................61
3.2 Mappe genetico molecolari intra-specifiche…………...……..……………62
3.2.1 Ben Chaim et al. (2001a): “QTL mapping of fruit related traits in pepper
(Capsicum annuum).”……………………………………………………...63
3.2.3 Sugita et al. (2005): “Rapid Construction of a Linkage Map using High-
efficiency Genome Scanning / AFLP and RAPD, Based on a Intraspecific, Doubled-haploid
Population of Capsicum annuum.” Breeding Science……………………………...66
3.2.4 Ogundiwin et al. (2005): “Construction of 2 intraspecific linkage maps and
identification of resistance QTLs for Phytophthora capsici root-rot and foliar-blight diseases
of pepper (Capsicum annuum L.)” Genome……………………………………..68
3.3 Mappe genetiche integrate…………………………………………….72
3.2.2 Lefebvre et al. (2002): “Towards the saturation of the pepper linkage map by
alignment of three intraspecific maps including known-function genes.” Genome……….73
3.3.2 Paran et al. (2004): “An integrated genetic linkage map of pepper (Capsicum
spp.)”…………………………………………………………………...80

IV
II. OBIETTIVI DELLA RICERCA……………...…………..……84
III. MATERIALI E METODI……………….…………………….85
1. Materiale vegetale……………………………………………………85
2. Estrazione del DNA………………………………………………..…87
3. Marcatori molecolari utilizzati nello sviluppo della mappa……………….……88
3.1 Marcatori microsatelliti (SSR)………………………………………88
3.1.1 Microsatelliti sviluppati da Sasvári et al. 2004…………………...…88
3.1.2 Microsatelliti sviluppati da Lee et al. 2004……………………...…88
3.1.3 Sviluppo di marcatori SSR, derivati da EST………………….……90
3.2 Sviluppo di un marcatore STS……………………………………....93
4. Analisi molecolare sulla popolazione segregante……………………………96
4.1 Condizioni di amplificazione………………………………………..96
4.2 Elettroforesi su gel di agarosio…………………………………..…..97
4.3 Quantificazione e aggiunta del tampone di corsa………………….……..97
4.4 Elettroforesi duplex e triplex………………………………………..97
4.5 Elettroforesi su gel di poliacrilamide………………………………….99
4.5.1 Preparazione delle lastre………………………………………99
4.5.2 Preparazione della soluzione di acrilamide………………………...99
4.5.3 Corsa elettroforetica…………………………………………100
4.6 Visualizzazione dei prodotti di amplificazione…………………………101
5. Screening dei marcatori microsatelliti e successiva analisi……………………103
5.1 Screening dei marcatori SSR sviluppati da Sasvári et al. (2004) e da Lee et al.
(2004).…………………………………………………………...…103
5.2 Screening dei marcatori SSR, derivati da EST…………………………103

V
6. Analisi dCAPs del marcatore STS………………………………………104
7. Analisi dei dati……………………………………………………...105
7.1 Valutazione del polymorphic information content (PIC) dei marcatori SSR,
derivati da EST……………………………………….………………105
7.2 Test del χ2……………………………………………………..106
7.3 Analisi di linkage………………………………………………..106
7.4 Assegnazione dei gruppi linkage ai cromosomi di C. annuum……………..109
7.5 Scelta degli individui più informativi.……………………………..…109
IV. RISULTATI…………………………………….…………...…111
1. Sviluppo di marcatori SSR, derivati da EST…………………………...…111
2. Marcatori polimorfici e segregazione…………………………………...121
3. Costruzione della mappa linkage………………………………………129
4. Posizione dei marcatori sui gruppi linkage……………………….………130
5. Attribuzione dei gruppi linkage ai cromosomi……………………………132
6. Identificazione degli individui più informativi……………………………135
V. DISCUSSIONE………………………………………...……….136
1. Popolazione segregante RILs:F5………………………………………138
2. Marcatori molecolari e costruzione della mappa genetica…………………...139
3. Allineamento e confronti con mappe precedentemente sviluppate………….…145
4. Identificazione degli individui RILs più informativi e mappaggio
selettivo………………………………………………….………147

VI
VI. CONCLUSIONI……………………………………………….148
VII. BIBLIOGRAFIA………………..……………………………149
Letteratura non citata.……………………………………………….…165
Ringraziamenti………………………………………………..……169

I-INTRODUZIONE
1
I-INTRODUZIONE
1. IL PEPERONE
1.1 Origine e diffusione.
Il genere Capsicum appartiene alla famiglia delle Solanaceae, come altre importanti
piante coltivate, quali pomodoro, patata, melanzana, tabacco e petunia. La denominazione
Capsicum è stata introdotta da Fuchs nel 1543, ripresa e precisata da Tourneforte (1700) e
infine da Linneo in Species Plantarum (1753, prima edizione). Il termine sembra derivare dal
latino capsa che significa “cassetta”, “custodia” e descrive perfettamente il frutto del
peperone: una piccola scatola con all’interno la placenta, sulla quale sono posizionati i semi.
Il termine può anche derivare dal Greco, kapto, che significa “pungere”: la piccantezza è
infatti una delle caratteristiche del genere anche se una singola mutazione è responsabile della
perdita della capacità di produrre capsacinoidi, le molecole che conferiscono questo carattere.
La maggior parte delle cultivar oggi coltivate in Europa sono infatti caratterizzate da assenza
o scarsa piccantezza.
Il peperone è originario dell’area centrale del Sudamerica (Bolivia). Incisioni rupestri,
risalenti al IX e X secolo d.C., rinvenute sulle Ande, testimoniano che in queste zone erano
conosciute e domesticate diverse specie appartenenti al genere Capsicum. Altri autori
sostengono che il centro di origine sia il Messico dove, ancora oggi, le popolazioni indigene
utilizzano specie con bacche piccole e piccanti.
Il viaggio di Cristoforo Colombo e le successive spedizioni dei Conquistadores in
America hanno permesso la diffusione del peperone in Europa, Asia e Africa. Colombo
denominò erroneamente la nuova spezia “pepper” in quanto ricordava la piccantezza del pepe
nero, Piper nigrum al quale il peperone non è filogeneticamente correlato. Peter Martyr
(Anghiera, 1493) scrisse che il “pepper” di Colombo era “più piccante di quello del Caucaso”;
l’assunzione che fosse un nuovo tipo di pepe nero ha incentivato la rapida introduzione della
spezia nelle tradizioni culinarie locali.
Nelle isole dei Caraibi gli indiani chiamavano il peperone “aji”, termine ancora oggi
impiegato nella lingua Aymara, tipica delle Ande del Perù e della Bolivia. Gli indiani del
Messico utilizzano il termine “chili”; gli Andini del Perù con “uchu” indicano i frutti raccolti
sulle piante selvatiche. Nel Vecchio Mondo sono nate diverse espressioni per denominare le
spezie a base di peperone, importate dalle Americhe: “Pepe Indiano”, “Pepe di Calicut”,

I-INTRODUZIONE
2
“Pepe di Guinea”, “Pepe di Cayenna”, “Pepe del Brasile”. Tra i vari termini, “Pepe dei
Poveri” esprime bene il fatto che questa pianta ha reso democratico l’uso delle spezie, fino ad
allora destinate alle tavole dei nobili e dei ricchi.
Oggigiorno il genere Capsicum ha una vasta dispersione geografica che va dalla cintura
intertropicale, alle latitudini del Nord Europa, ciò lo rende suscettibile a un gran numero di
patogeni. Gli Oomyceti, i nematodi e i virus trasmessi da afidi sono i principali responsabili
delle perdite di produzione della coltura in tutto il mondo (Yoon et al. 1991; Thabuis et al.
2004).
1.2 Importanza economica
Il peperone vanta oggi giorno una produzione mondiale di 23 milioni di tonnellate
annue e una superficie coltivata di 1,7 milioni di ettari, (http://www.FAO.org 2003) che lo
classificano al secondo posto dopo il pomodoro (Fig. 1). Cina e Turchia sono i maggiori
produttori mondiali. A livello Europeo al primo posto figura la Spagna, seguita da Italia e
Paesi Bassi; la Francia è al primo posto per la produzione di sementi (Palloix et al. 2003). In
Italia, nel 2001, sono stati destinati alla produzione in pieno campo 11.336 ha, con una
produzione unitaria di circa 22 t destinata in gran parte all’esportazione nei Paesi dell’Europa
centrosettentrionale. A livello nazionale, le regioni in cui si è realizzata la maggiore
produzione sono: la Sicilia (3.000 ha), il Piemonte (1.520 ha), la Puglia (1.390 ha), la
Basilicata (1.290), la Calabria (1.347 ha), la Campania (920 ha), il Lazio (700 ha), il Veneto
(630 ha), l’Abruzzo (510 ha), l’Umbria (430 ha), le Marche (406), la Toscana (290 ha), la
Sardegna (270 ha), l’Emilia Romagna (140 ha), la Lombardia (130 ha) e infine la Liguria (50
ha) (Valli, 2001).
Fig. 1 Raccolta

I-INTRODUZIONE
3
1.3 Classificazione
In base alla più recente classificazione, il genere Capsicum appartiene al:
Regno Plantae
Divisione Magnoliophyta
Classe Magnoliopsida
Ordine Solanales
Famiglia Solanaceae
All’interno del genere Capsicum tutte le specie sono diploidi, con numero cromosomico
base 12 (2n=2x=24), comune agli altri membri della famiglia delle Solanaceae. Studi
effettuati indipendentemente da Galbraith et al. (1983) e Arumuganathan e Earle (1991),
dimostrano che il genoma aploide del peperone corrisponde a 2,76 pg, ed ha una lunghezza
che va da 2702 a 3420 Mbp. E’ 3-4 volte più grande di quello del pomodoro, della patata e
della melanzana e circa 20 volte quello di Arabidopsis thaliana (Lefebvre 2004). Il genere
Capsicum comprende all’incirca 25 specie selvatiche e 5 specie domesticate (Tabella 1).
Tab. 1 Le diverse specie del genere Capsicum.
Specie di Capsicum. (IBPGR, 1983)
Capsicum: C. annuum C. baccatum
Var. aviculare Var. baccatum
Var. annuum Var. pendulum
C. buforum C. campylopodium
C. cardenasii C. chacoense
C. chinense C. ciliatum
C. coccineum C. cornutum
C. dimorphum C. dusenii
C. eximium C. frutescens
Var. tomentosum C. galapagoense
C. geminifolium C. hookerianum
C. lanceolatum C. leptopodum
C. minutiflorum C. mirabile
C. parvifolium C. praetermissum
C. pubescens C. schottianum
C. scolnikianum C. tovarii
Var. flexuosum
C. villosum

I-INTRODUZIONE
4
Alcune specie selvatiche vengono ancora raccolte e utilizzate dalle popolazioni
indigene: C. lanceolatum, specie individuata solo nelle foreste vergini pluviali del Guatemala;
C. annuum varietà avirculare, il “chiltepin” selvatico (Fig. 2). Tutte le specie selvatiche
condividono caratteri simili: frutti piccoli, di colore rosso vivo, eretti, con un pedicello tenero
che ne permette il facile distacco dal calice a maturità. Questa caratteristica facilita l’azione
degli uccelli frugivori che, attratti dal colore dei frutti e non sensibili ai capsacinoidi, ne
garantiscono la disseminazione. Così si spiega il termine “bird pepper” usato nel linguaggio
comune per designare queste specie (Bosland e Votava, 1999).
Fig. 2 C. annuum var. avirculare
Dalle specie native, le popolazioni indigene del Messico, Centro e Sud America hanno
addomesticato indipendentemente 5 specie, che ora sono le più coltivate (IBPGR, 1983):
- C. pubescens R. & P;
- C. baccatum L.;
- C. annuum L. (il più diffuso e commercialmente importante);
- C. frutescens L. (prevalentemente usato come spezia “tabasco”);
- C. chinense Jacq..
Le diverse specie sono distinte in tre gruppi sulla base di caratteristiche morfologiche e
citologiche. Ogni gruppo comprende sia le specie domesticate, sia i loro progenitori selvatici.
Specie appartenenti ad uno stesso gruppo sono inter-incrociabili, viceversa l’ibridazione tra
specie di gruppi diversi è ostacolata da svariati meccanismi di incompatibilità. I gruppi
rappresentano una sorta di gene pool d’origine della variabilità genetica delle specie di
Capsicum.

I-INTRODUZIONE
5
1.3.1 Gruppo Capsicum pubescens
Il gruppo C. pubescens è costituito da specie relativamente sconosciute. C. pubescens fu
inizialmente descritto da Ruiz e Pavon (1790) come specie originaria degli altopiani della
Bolivia e, in accordo con Heiser (1976), fu domesticato intorno al 6000 A.C. nelle Ande, dove
spesso viene denominato “locato” o “rocoto”. Cresce nell’area compresa tra Messico e Perù,
nelle Ande del Sud America e negli altipiani del Centro America in piccoli appezzamenti
famigliari. Nel resto del mondo è poco diffuso.
La pianta di C. pubescens ha fiori color porpora, numerose foglie pubescenti e semi
neri, che la rende distinguibile dalle altre specie (Fig. 3 e 4). E’ una pianta erbacea, molto
ramificata; può crescere fino a 12 m di altezza e può vivere oltre 10 anni nelle condizioni
climatiche dell’America tropicale. I frutti variano per forma: da allungati a sferici, e per
colore: rossi, arancione o gialli, ma non mostrano lo stesso range di variabilità di C. annuum. I
frutti sono caratterizzati da una polpa densa, difficile da disidratare e da conservare (Fig. 5).
Fig. 3 Fiore viola di C. pubescens Fig. 4 Foglie pubescenti
Il gruppo C. pubescens include anche le specie C. eximium e C. cardenasii.

I-INTRODUZIONE
6
Fig. 5 Variabilità nei frutti di C. pubescens.
1.3.2 Gruppo Capsicum baccatum
C. baccatum, detto “ajì” in dialetto, è una specie diffusa dal Sud, in Brasile, fino
all’Ovest, sulle coste dell’Oceano Pacifico. Si ritiene che il suo centro di domesticazione
comprenda la Bolivia, l’Ecuador, il Perù, e il Cile. Nel Sud America è la specie più
comunemente coltivata.
Fig. 6 C. baccatum cv christmas bell ajì Fig . 7 C. baccatum cv peri peri

I-INTRODUZIONE
7
C. baccatum ha fiori di colore crema con punti gialli, marroni e verde scuro sulla
corolla. Si distinguono due varietà botaniche: C. baccatum var. baccatum , che è la forma
selvatica, e C. baccatum varietà pendulum, che include le forme domesticate. I frutti variano
in funzione alla forma, al colore, alla piccantezza, e esprimono peculiari proprietà
organolettiche (Fig. 6, 7, 8 e 9).
Fig. 8 C. baccatum cv kovinchu
Il gruppo C. baccatum comprende anche le specie C. praetermissium (Fig. 10) e C.
tovarii (Tong, 1998).
Fig. 10 Fiore di C. praetermissum USDA#44165
Fig. 9 C. baccatum cv peru yellow

I-INTRODUZIONE
8
1.3.3 Gruppo Capsicum annuum
Il gruppo C. annuum comprende tre specie domesticate: C. chinense, C. frutescens e C.
annuum, che condividono un gene pool ancestrale comune. Sono le specie più importanti dal
punto di vista commerciale. Ciascuna è stata domesticata in modo indipendente: C. chinense
in Amazzonia o in Perù, C. frutescens nell’America centromeridionale e C. annuum in
Messico (Smith et al. 1987; Bosland, 1992).
Le diverse varietà di C. annuum sono solitamente classificate in base alle caratteristiche
dei frutti: piccantezza, colore, forma, sapore, dimensioni e utilizzo, (Smith et al. 1987;
Bosland, 1992). Il capostipite più probabile del C. annuum coltivato è il selvatico “chiltepin”
(C. annuum varietà avirculare), caratterizzato da un’ampia diffusione, dall’America
meridionale all’Arizona. Si ritiene che sia stato domesticato in Messico e in America centrale
dove, prima della colonizzazione spagnola, gli Atzechi ne avevano già sviluppato una dozzina
di varietà. Senza dubbio i “chiltepin” sono i precursori di un gran numero di varietà oggi
coltivate in Messico (Fig. 11). La specie C. annuum è la più coltivata nel mondo; include
molti dei “chile peppers” messicani, i peperoni piccanti coltivati in Africa e Asia, le diverse
cultivar di peperone dolce coltivato nelle regioni temperate dell’Europa e del Nord America.
Fig. 11 “chiltepin ”

I-INTRODUZIONE
9
La specie C. chinense, come tutte le altre, è originaria dell’America. Fu il fisico
olandese Kikolaus von Jacquinomist che diede il nome “chinense” alla specie nel 1776, ma
non è ancora chiaro il perché pensasse che la Cina fosse il suo centro di origine (Jacquin,
1776). E’ la specie più diffusa in Brasile (Cheng, 1989) e nelle regioni tropicali, soprattutto
quelle Caraibiche. Anche in questa specie i frutti mostrano grande variabilità nella forma e nel
grado di piccantezza; l’“habanero” si distingue come il più piccante dei peperoni al mondo
(Fig. 12).
Fig. 12 Variabilità nei frutti di C. chinense
La specie C. frutescens (sinonimi: C. minimum e C. fastigiatium) comprende un minor
numero di varietà rispetto a C. chinense e C. annuum. In Brasile è famosa la cultivar
“malagueta” che però non è relata a Aframomum melegueta, “melegueta” o “peperone di
Guinea” (Fig.13). C. frutescens comprende anche la nota varietà “Tabasco”, utilizzata per
produrre l’omonima salsa (Fig. 14) Alcune varietà sono coltivate in Africa, India e Asia, dove
sono chiamate “bird peppers” e crescono come piante arbustive e perenni.
Fig. 13 C. frutescens “malagueta” Fig. 14 C. frutescens , “tabasco”

I-INTRODUZIONE
10
1.4 Caratteristiche botaniche del gruppo Capsicum anuum L.
Capsicum annuum, nei nostri climi, è una pianta erbacea annuale, con caratteristiche
prevalentemente brevidiurne: la durata ottimale del fotoperiodo è di circa 12 ore al giorno.
L’apparato radicale è tendenzialmente superficiale e raggiunge approssimativamente i 30 cm
di profondità. E’ abbastanza robusto ma non è in grado di fornire un sufficiente sostegno alla
pianta durante lo sviluppo vegetativo: nella fase di ingrossamento dei frutti può infatti
piegarsi in modo marcato e per evitare questo si fa spesso ricorso a tutori. Nel suo insieme
l’apparato radicale è costituito da numerose radici secondarie, affastellate e di dimensioni
ridotte e da un fittone (prolungamento principale) il cui sviluppo è influenzato dalla
composizione del terreno di coltura.
Il fusto è glabro nelle specie coltivate, pubescente in alcune specie spontanee, angoloso
ed eretto, con un’altezza variabile tra i 40 cm e 1 m. Le ramificazioni che si dipartono dal
fusto sono dicotomiche e in corrispondenza di ciascuna biforcazione si sviluppa un fiore, solo
occasionalmente due, e una foglia.
Fig. 15 Variabilità dei frutti di C. annuum

I-INTRODUZIONE
11
Le foglie sono alterne, lucide, a lamina ovata o ellittica, acuminate e a margine intero;
sono ricche di stomi sulla pagina inferiore (oltre 30 mila/cm2), più povere su quella superiore
(un terzo). Questo comporta ritmi di respirazione molto intensi e continui, e la necessità di
regolari apporti idrici.
I fiori sono ermafroditi, localizzati all’ascella delle ramificazioni e caratterizzati da un
penduncolo breve che si incurva all’antesi, cioè alla schiusura del bocciolo fiorale,
permettendo la caduta del polline sullo stilo.
Il sistema riproduttivo è tendenzialmente autogamo, nonostante siano stati segnalati casi
di allogamia anche sino al 90% (Pickersgill et al. 1979); questo dipende dalla varietà
(lunghezza dello stilo in rapporto agli stami), dalle condizioni ambientali e dall’attività dei
pronubi. Il vento e gli insetti, come Coccinella septem-punctata, Hippodamia tredecim-
punctata, Apis mellifera, Macrosteles sexnotatus, Coleoptera Halticinae, Diptera
Brachycerinae, Hymenoptera Anthocoridae, Lygaeidae, Nabidae, Aphididae, Typhlocybidae,
Lepidoptera Sphingidae, possono essere responsabili di una frequenza di impollinazione
incrociata variabile dall’1-5 % fino a più del 50 % (Franceschetti, 1971; Lorenzetti e Cirica,
1974; Csillery et al. 1986). Tutto ciò impone cautele e regole rigorose per riuscire a
mantenere le cultivar in purezza.
I caratteri morfologici dei fiori sono impiegati per la distinzione delle specie. Il fiore di
C. annuum è completo ed è caratterizzato da: corolla bianca, rotata, con sei petali; calice
angoloso, verde, con 5-6 sepali; androceo composto da 5 stami sporgenti, inseriti alla base
della corolla con antere violacee che si aprono fessurandosi longitudinalmente in
corrispondenza dei sacchi pollinici; gineceo composto da un pistillo con stigma allungato e
piatto; ovario supero e di norma binoculare (raramente è suddiviso in 3-4 loculi).
La fioritura ha inizio con uno o due fiori sul primo nodo, successivamente i fiori si
sviluppano sugli altri nodi seguendo una progressione geometrica. Solitamente si sviluppano
più di cento fiori su ogni pianta. Per ogni singolo fiore l’antesi dura 2-3 giorni, quindi la
fioritura si prolunga, in ambienti colturali favorevoli, per molti mesi.
Il frutto è una bacca carnosa, indeiescente. L’interno è caratterizzato da ampi spazi
vuoti, suddivisi in 2-4 logge da setti placentari bianchi la cui estensione non raggiunge mai
l’apice del frutto. Nella regione peduncolare è presente un tessuto placentare bianco e
spugnoso sul quale sono inseriti i semi. La bacca può avere portamento pendulo o eretto in
base alla disposizione dei fiori. L’epidermide a maturazione può assumere colorazioni
diverse: giallo, rosso, paglierino, arancione o bruno. L’uniformità di colorazione è indice
dell’avvenuta maturazione in condizioni ambientali e colturali buone.

I-INTRODUZIONE
12
In base alla forma della bacca si distinguono:
- i quadrati americani: 8-20 cm x 8-10 cm;
- i quadrati olandesi: 7-8 cm x 8-10 cm;
- i quadrati italiani: 10-12 cm x 10-12 cm;
- i rettangolari ½ lunghi (a sezione quadrata e forma di piramide tronca):
8-10 cm x 15-18 cm;
- i rettangolari ¾ lunghi: 8-10 cm x 15-18 cm;
- i rettangolari lunghi: 9-11 cm x superiore a 18 cm;
- i corni, di lunghezza e dimensione variabile, a sezione quadrata o circolare con
forma conica molto allungata, circa 5-6 cm x 18-30 cm.
A queste tipologie si aggiungono i “cornetti” piccanti, i “topepo”, le “sigarette dolci”, i
“naso di cane”, i “ceraseli” (o frutti a ciliegia) e una svariata serie di ecotipi locali
caratterizzati da un maggiore o minore grado di piccantezza e diffusi in aree limitate e
specifiche.
I semi sono di colore giallo, variabile da crema a paglierino vivo; hanno una forma
tondo-reniforme, fortemente appiattita, con un diametro di 3-5 mm (Fig. 16). Il numero varia
da 120 a 160 per grammo, in funzione alla varietà e alla posizione delle bacche sulla pianta: i
primi frutti allegati producono solitamente semi più grossi e in numero maggiore. La
produzione del seme è anche fortemente influenzata dalle condizioni metereologiche (luce e
temperatura).
Fig. 16 Peperoncini esiccati e relativi semi.

I-INTRODUZIONE
13
1.5 Caratteristiche organolettiche e nutrizionali
Aromi, vitamine e colori partecipano al fascino del peperone (Fig. 17). La composizione
è diversa in funzione alla specie, alla cultivar, alle condizioni di coltivazione e allo stadio
della raccolta. E’ per questo che la variabilità genetica viene conservata e considerata un bene
prezioso dai produttori interessati alla coltura: le diverse varietà non sono assolutamente
interscambiabili.
Come la maggior parte delle piante, i peperoni contengono migliaia di composti: acqua,
acidi grassi, acidi grassi volatili, carotenoidi, capsacinoidi, resine, proteine, fibre ed elementi
minerali. La loro presenza è fondamentale per determinare il valore nutrizionale, il sapore, il
colore e l’aroma del peperone. I composti più importanti sono i carotenoidi, che
contribuiscono al colore del frutto e hanno un importante ruolo nutrizionale, e i capsacinoidi,
alcaloidi che conferiscono piccantezza.
Fig. 17 Piatto di peperoni
ACQUA: è l’elemento più abbondante nei frutti. La quantità presente dipende dall’età e
dalla varietà; si passa dal 90% nei peperoni verdi a maturità, al 70% per quelli che vanno
incontro ad essiccazione sulla pianta. I prodotti disidratati e conservati in magazzino ne
contengono dal 15 al 20%. Dopo la raccolta, la perdita di acqua è rapida; questo rappresenta
un problema soprattutto per il mantenimento della qualità nei prodotti freschi, destinati alla
spedizione (Lownds et al. 1993).

I-INTRODUZIONE
14
CARBOIDRATI: i frutti del peperone contengono zuccheri, pentosani e fibra. Il
glucosio rappresenta il 90-98% degli zuccheri presenti nei frutti maturi (Somos, 1984). La
quantità di zuccheri nel frutto è variabile in funzione alla varietà e al tipo di frutto. Alcuni
zuccheri conferiscono una particolare dolcezza al peperone, mentre altri riducono questa
sensazione. Gli zuccheri totali e riducenti sono presenti in livello massimo nei frutti rossi e
succulenti (Wall e Biles, 1993). La cellulosa e le fibre alimentari rappresentano il 20% del
peso secco del tessuto del pericarpo. McKee nel 1998 osservò che la buccia del frutto
contiene il 77% di fibre solubili e l’80% della fibra alimentare totale; questa quantità è
maggiore di quella presente nel riso o nel grano. Il pericarpo è stato sempre considerato un
sottoprodotto della lavorazione del peperone verde “New Mexican”, venduto inscatolato;
sembra ora essere vantaggiosa la sua presenza nei prodotti cotti al forno in quanto apporta
un’eccellente quantità di fibra alimentare.
LIPIDI: il contenuto lipidico totale dei frutti freschi è pari all’incirca a 400 mg su 100 g
(peso fresco), livello relativamente basso (Kinsella, 1971). L’80 % dei lipidi sono neutri, il 2
% sono fosfolipidi e il 16% glicolipidi. I trigliceridi rappresentano il 60% dei lipidi totali e
sono preponderanti l’acido palmitico, linoleico e linolenico (Lyons e Lippert, 1996). L’acido
linoleico rappresenta il 70% degli acidi grassi totali. La quantità di acidi grassi insaturi nei
mitocondri sembra essere correlata alla maggiore o minore sensibilità delle specie vegetali nei
confronti delle ferite da freddo. Le specie che accumulano maggiori quantità di acidi grassi
insaturi sono più resistenti alle basse temperature. I peperoni sono sensibili alle basse
temperature e il rapporto acidi grassi insaturi/saturi lo conferma.
AMMINOACIDI, PROTEINE, MICROELEMENTI: Somos (1984) ha studiato a fondo
questi composti dimostrando la presenza di lisina, arginina, prolina, tirosina, triptofano,
metionina, valina, fenilalanina, leucina, acido glutammico, glicina, asparagina, treonina e
alanina nei frutti. Dalle ricerche effettuate in Ungheria, ha osservato che il pericarpo è
costituito per il 16-17% da proteine che i semi ne contengono 18%. Dall’analisi dei
microelementi emerge che il ferro è quello presente in concentrazione maggiore, seguito da
bromo e manganese. Altri microelementi presenti sono: cadmio, calcio, cobalto, rame,
magnesio, fosforo, potassio, sodio e zinco.

I-INTRODUZIONE
15
CAPSADIOLO: è una fitoalessina, sesquiterpene biciclico, che viene sintetizzato
attraverso la via metabolica dell’acido mevalonico. Il capsadiolo svolge un’azione
fungostatica nei confronti di Phytophthora capsici (Jones et al. 1975), l’Oomycete più
pericoloso per il peperone. La sua importanza a riguardo e la correlazione diretta tra
capsadiolo e resistenza al patogeno non sono ancora stati delucidati.
FLAVOUR E AROMA: il flavour è una sensazione complessa che si percepisce in
bocca per mezzo delle papille gustative e grazie al nervo olfattivo, che permette di cogliere gli
aromi dei composti volatili. I grandi esperti di peperoni, come i sommellier, sono in grado di
identificare le sottili differenze esistenti nelle varietà: l’“ancho” è dolciastro, il “mulato” è
caratterizzato da un aroma di cioccolato, il “mirasol” è fruttato e il “chipotle” è affumicato. Si
sa ancora poco sulla natura specifica degli aromi del peperone, soprattutto nel caso delle
specie più ricche di varietà come C. chinense e C. baccatum. Nel frutto verde di “Jalapeño”
(C. annuum) pirazine, terpeni, esteri, acidi carbossilici e alcoli partecipano tutti assieme al
tipico flavour di questa varietà messicana molto apprezzata, e di quelle nord-americane che
derivano da essa. Uno dei composti volatili tipico del peperone è la pirazina, 2-metossi-3-
isobutil-pirazina, l’aroma del “green bell pepper”. Buttery e collaboratori (1969) scoprirono
che l’uomo è in grado di percepire questo aroma già quando è presente in quantità pari a 2
parti per trillione! Keller e collaboratori, nel 1981, hanno analizzato i composti volatili del
peperone e identificato 102 composti aromatici in C. annuum e C. frutescens. In studi
analoghi, Haymon e Aurand (1971) hanno caratterizzato 125 composti nell’olio estratto da C.
frutescens cv “Tabasco” e osservato che la loro abbondanza relativa era variabile in funzione
al periodo di raccolta. La composizione aromatica del “Tabasco” è significativamente diversa
da quella del “green bell pepper” e non contiene pirazina. Il suo aroma è determinato
fondamentalmente da tre composti: 4-metil-1-pentil-2-metilbutirrato, 3-metil-1-pentil-3-
metilbutirrato e isoexil-isocaproato.
VITAMINE: la ricchezza in vitamine è uno dei punti forza del peperone. I frutti
presentano alti livelli di vitamina C, provitamina A, E, P (citrina), tiamina (B1), riboflavina
(B2) e niacina (B3). I livelli sono variabili in funzione alla varietà, al grado di maturità del
frutto, alle pratiche di coltivazione, al clima, alla conservazione post-raccolta e ai metodi
analitici impiegati nell’analisi (Mozafar, 1994).
Il peperone è una delle principali fonti di acido ascorbico (vitamina C) e, tra i vegetali,
solo il ribes nero può competere con esso. Il contenuto varia tra i 46 e i 243 mg per 100 g

I-INTRODUZIONE
16
(peso fresco) nei frutti maturi, mentre nel frutto allo stadio arancione è di circa 1 mg
(Wimalasiri e Wills, 1983; Nisperos-Cerriedos et al. 1992; Howard et al. 1994; Lee et al.
1995). L’acido ascorbico aumenta durante la maturazione dei frutti (Osuna-Garcia, 1996), il
livello raggiunto è variabile, ma tale da soddisfare l’RDA, che è di 60 mg (NRC, 1989).
Anche cotto, il frutto conserva più della metà dell’acido ascorbico; nel corso del processo di
essiccazione, la vitamina C, essendo idrosolubile, viene ossidata. La vitamina C è stata isolata
e cristallizzata per la prima volta dai frutti del peperone nel 1928 dall’ungherese Albert Szent-
Györgyi che vinse il premio Nobel (Fig. 18).
Fig. 18 La vitamina C
I peperoni sono anche ricchi di vitamina E (tocoferolo). La polvere rossa, ottenuta dalla
disidratazione del frutto, ha livelli di α-tocoferolo comparabili a quelli di spinaci e asparagi; i
frutti freschi ne contengono livelli quattro volte maggiori rispetto al pomodoro. Osuna-Garcia
(1996) riporta che i semi contengono γ-tocoferolo mentre il pericarpo accumula la forma α. Il
γ-tocoferolo raggiunge il livello massimo nei semi quando i frutti sono allo stadio rosso della
maturazione, poi decresce. Il contenuto di α-tocoferolo dipende dal contenuto lipidico, che
varia in funzione allo stadio di maturazione e al tipo di varietà (Kanner et al. 1979).
La vitamina A appartiene alla classe dei carotenoidi ed è presente nel peperone
sottoforma di provitamina come β-carotene. Nei frutti verdi rappresenta il 95% dei
carotenoidi, nei frutti rossi e maturi il 93% (Howard et al. 1994).
CAROTENOIDI: conferiscono i colori brillanti al peperone e sono accumulati nelle
membrane tilacoidali dei plastidi. Nelle piante sono sintetizzati sia nei cloroplasti presenti nei
tessuti fotosintetici, sia nei cromoplasti di frutti, fiori e radici. Chimicamente, sono molecole
liposolubili appartenenenti alla classe degli idrocarburi. Hanno una struttura simmetrica con
una serie di doppi legami coniugati, responsabile dell’assorbimento della luce visibile. I
carotenoidi sono fotoprotettori indispensabili nelle piante: il β-carotene e altre forme
preservano i cloroplasti dai danni fotoossidativi; nel processo fotosintetico svolgono invece

I-INTRODUZIONE
17
un ruolo accessorio. Nei frutti hanno un importante ruolo attrattivo nei confronti degli uccelli,
indispensabili nel garantire la dispersione dei semi. I colori verde, giallo, arancione e rosso
derivano dai pigmenti prodotti durante la maturazione del frutto; sono stati identificati più di
30 composti differenti in peperone (Matus et al. 1991) tra cui: clorofilla (a e b), luteina,
zeaxantina, violaxantina, anteraxantina, β-criptoxantina e β-carotene, presenti nella maggior
parte dei vegetali. Capsantina, capsarubina, e criptocapsina sono pigmenti unici del peperone.
Il colore rosso del frutto è determinato da capsantina e capsarubina, il giallo e l’arancione dal
β-carotene e dalla violaxantina. La capsantina è il carotenoide più abbondante nel frutto
maturo e rappresenta il 60% dei carotenoidi totali. Capsantina e capsarubina aumentano
proporzionalmente al grado di maturazione del frutto (Harkay-Vinkler, 1974; Kanner et al.
1979) e la loro concentrazione aumenta fino a 100 volte tanto nel giro di due settimane. In
seguito alla raccolta la quantità di pigmenti continua a crescere (post-maturazione).
I carotenoidi hanno funzioni antitumorali nell’uomo: la loro attività anti-ossidante
neutralizza i radicali liberi (radical scavenger). Questa evidenza rinnova l’interesse nei
confronti di cibi ricchi questi composti; gli integratori dietetici infatti non sono sempre in
grado di apportare gli stessi benefici medici e nutrizionali derivanti dal consumo quotidiano di
frutta e verdura freschi.
PICCANTEZZA: è un’altra caratteristica tipica dei peperoni. E’ determinata dai
capsacinoidi, sette molecole omologhe alle catene laterali alchiliche della vanillammide,
esclusive del genere Capsicum (Torabi, 1997). Sono composti inodori, incolori, insapori. La
composizione della capsaicina (C18H27NO3), 8-metil 6-nonenoil vanillammide, è simile a
quella della piperina (C17H19NO3), composto tipico del pepe nero. La sua formula chimica è
stata determinata nel 1923 da Nelson e Dawson (Fig. 19). I capsacinoidi sono sintetizzati in
ghiandole presenti sulla placenta (Fig. 20). I semi, localizzati su di essa, tendono ad assorbire
la capsaicina, risultando così piccanti.
Fig. 19 Capsaicina.

I-INTRODUZIONE
18
I capsacinoidi non sono percepiti dall’uomo attraverso le papille gustative ma dai
recettori delle innervazioni trigemine, localizzate nella bocca, nel naso e nello stomaco (Silver
e Maruniak, 1981). Determinati neurotrasmettitori inducono la tipica sensazione di bruciore e
calore.
Fig. 20 Placenta con venatura arancio, luogo di sintesi dei capsacinoidi.
Sono stati sviluppati diversi metodi per misurare la piccantezza. I due più usati sono il
test organolettico di Scoville, che esprime la piccantezza in Scoville heat units (SHU)
(Scoville, 1912), e l’HPLC, che permette un’analisi accurata sia del contenuto sia dei tipi di
capsacinoidi presenti (Collins et al. 1995). Per convertire le unità di misura in SHU si
moltiplicano i valori ppm per 15. La quantificazione dei capsacinoidi è importante in quanto
vengono utilizzati sottoforma di oleoresine in svariati ambiti industriali (paragrafo 1.7).

I-INTRODUZIONE
19
1.6 Varietà tradizionali
Nei mercati locali di frutta e verdura il peperone occupa uno spazio di riguardo e
ciascun Paese ne possiede un assortimento particolare (Fig. 21) (Palloix et al. 2003). In
Spagna, nelle regioni Tutela e Calahorra, si coltiva un peperone cordiforme detto “Morron”,
impiegato soprattutto per la preparazione di conserve. In Ungheria, paese dove il peperone è
un re, vanno matti per i piccoli peperoni conici di color bianco-avorio del tipo “Cecei”. Sono
spesso consumati crudi, sono meno acidi delle varietà di colore verde ma anche meno ricchi
in vitamina C. Esistono sia varietà dolci, sia piccanti. In Romania e Bulgaria sono molto
utilizzati peperoni dolci, a forma di pomodoro, con la polpa spessa, molto utili per la
produzione di concentrati. Nei Balcani, i peperoni più consumati hanno sezione triangolare e
colore verde-giallo prima della completa maturazione. Alcune varietà hanno una
composizione peculiare adatta alla produzione di polvere. In Francia, l’importanza del
peperone è limitata alle aree meridionali.
Fig. 21 Varietà tipica: Ciliegino
Nei Paesi Baschi è famoso il “Piment d’Espelette”, che seccato al sole e ridotto in
polvere, serve ad aromatizzare i prodotti di salumeria locale. Un tempo ornava in ghirlande le
facciate delle case. In Aquitania, il peperone “Doux long des Landes”, dal frutto molto dolce e
dalla forma stretta e allungata, viene consumato tagliato a julienne in insalata o nelle
omelettes. “Doux long des Landes” è un nome celebre nei laboratori che si occupano della
genetica del peperone; da studi su questa varietà, all’INRA di Avignone negli anni ’70, è stata
ottenuta la seria dei trisomici (piante in cui uno dei cromosomi si trova in triplice copia). Il
“Petit Marseillais” è una varietà locale precoce, a polpa fine, di colore arancione a maturità,
che viene consumato soprattutto in inverno. Il peperone “Antibois” ha una forma sferica e un

I-INTRODUZIONE
20
aroma molto particolare. La popolazione valchiusana “Gros carré des Lagnes” ha frutti molto
grossi e da essa si è ottenuto l’ibrido “Lamuyo”, che ha regnato per molto tempo senza
concorrenti nelle colture invernali in Francia, Spagna e Italia. A Nord, nella valle del Rodano
e della Saona, si trovano piccole popolazioni residue come il “Poivron d’Ampuis” e il “Poivre
rouge de Bresse”, attualmente impiegato come condimento per un particolare formaggio
tipico.
In Italia, la regione Piemonte è famosa per la produzione di varietà di peperoni tipici.
Ad Asti prevale il peperone giallo, a forma sferica più o meno solcato, detto “Quadrato
giallo” o il più appiattito “Topepo”. A Cuneo si coltivano varietà caratteristiche che sembrano
derivare da un incrocio naturale tra una vecchia cultivar locale, dalle bacche di dimensione
medio-piccola, a forma di pomodoro con varietà caratterizzate da bacche di dimensioni
elevate riconducibili alla tipologia “Napoletana” e/o “Nocera”. Da selezioni successive sono
nati i contemporanei e apprezzati “Peperoni di Carmagnola” e l’ecotipo “Cuneo”.
Fig. 22 Il Quadrato di “Carmagnola”
Il Peperone di Carmagnola comprende quattro tipologie. “Il Quadrato di Carmagnola”,
(Figura 22), è prelibato per un consumo crudo, ripieno ed al forno. “Il Lungo” o “Corno di
Bue” è molto apprezzato perchè la sua polpa compatta garantisce una buona “tenuta alla
cottura”. “La Trottola”, dalla forma a trottola con punta leggermente estroflessa o con punta
troncata, è una qualità che assicura alte rese in tutte le preparazioni culinarie. Il “Tumaticot”,
ibrido di forma tondeggiante schiacciata ai due poli, ha forma compatta e polpa spessa, che lo
rendono adatto, in particolar modo, alla preparazione di antipasti ed alla conservazione in
edizioni raffinate e particolari (i.e.: peperone in agrodolce). Anche in altre regioni italiane

I-INTRODUZIONE
21
emergono varietà locali di peperone dalle straordinarie qualità organolettiche: “Sigaretta di
Bergamo” (Fig. 23), “Lombardo”, “Senise” (Fig. 24), “Corno calabrese” (Fig. 25), “Corno
pescarese”, “Roggianese”, “Friarello”, “Nocera”, “Braidese”.
Fig. 23 Sigaretta di Bergamo Fig. 24 Senise Fig. 25 Corno calabrese

I-INTRODUZIONE
22
1.7 Possibili utilizzi di Capsicum
Tutti i peperoni possono essere consumati freschi, ma quelli comunemente destinati a
questo tipo di impiego hanno caratteristiche particolari: la polpa è spessa, succosa,
consistente, brillante, priva di ammaccamenti e abrasioni. E’ importante una corretta
conservazione dei frutti post-raccolta per mantenerne le caratteristiche organolettiche ed
evitarne la rapida disidratazione.
Una parte della produzione di peperoni è destinata alla conservazione in scatola. In
questo caso i frutti devono essere ben maturi, rosso brillante, con pareti spesse e croccanti.
Possono essere sbollentati, arrostiti o affumicati e conservati sott’olio, sotto aceto o in
agrodolce.
Le varietà piccanti di C. annuum, C. frutescens e C. chinenese sono spesso utilizzate per
la produzione di salse. Alla polpa viene addizionato sale, circa 14-20% poi viene lasciata
fermentare fino a sviluppare il suo caratteristico aroma. La salsa Tabasco ®, prodotta dalla
McIlhenny Company of Louisiana, è probabilmente quella più famosa. Si ottiene dai frutti di
C. frutescens e viene fatta fermentare per un minimo di tre anni in botti di rovere (Fig. 26).
Fig. 26 La famosa salsa Tabasco ®.
Grandi quantità di peperoni essiccati sono utilizzati in cucina per insaporire i cibi e nelle
industrie che producono alimenti in scatola. Le tipologie utilizzate sono: “New Mexican”,
“cayenne”, “ancho”, “paprika”, “mirasol”, “piquin” e “de arbol” (Bosland, 1992). Fin dai
tempi antichi i peperoni sono stati conservati disidratati. La qualità dei prodotti ottenuti
dipende dal livello di piccantezza, dalla quantità di pigmenti rossi estraibili e dall’aroma. I
frutti devono essere processati e stoccati correttamente per mantenere le caratteristiche
organolettiche nel prodotto finale, che conserverà a lungo le sue proprietà.
Dai peperoni si estraggono le oleoresine, molto ricche in pigmenti e capsacinoidi,
utilizzate dalle industrie agro-alimentari e farmaceutiche. Le oleoresine sono disponibili in

I-INTRODUZIONE
23
due forme: liposolubili e idrosolubili, e si ottengono dal pericarpo essiccato. Le varietà
impiegate sono selezionate in funzione alla ricchezza di pigmenti carotenoidi e alla facilità di
essiccazione naturale o in stufe. Questi prodotti vengono impiegati per standardizzare la
piccantezza, il colore e l’aroma dei prodotti alimentari. A causa dell’elevata concentrazione
dei composti nelle oleoresine è opportuno diluirle in un solvente appropriato, in modo da
ottenere un’essenza. Le oleoresine costituiscono un additivo naturale particolarmente
importante nell’industria alimentare perché risultano un valido sostituto dei coloranti rossi,
sintetici, che sono tutti più o meno tossici. Vengono impiegate in vari prodotti: bibite, lattiero
caseari, pasticceria, confetteria, alimenti per volatili (incrementa il colore giallo dell’uovo).
Fin dall’epoca pre-Colombiana i peperoni sono stati utilizzati come piante medicinali e
ancora oggi sono considerati rimedi naturali. Dai reperti della farmacopea Maya, emerge che i
peperoni venivano utilizzati contro malattie di origine microbica (Cichewicz e Thorpe, 1996).
Al peperone vengono attribuite diverse virtù: antidissenterico, antiparassitario, cicatrizzante,
afrodisiaco, ma sono ancora da confermare. Il peperone ha un ruolo stimolante: aumenta le
secrezioni salivari e gastriche, accelera le contrazioni peristaltiche dell’intestino e quindi
migliora il transito intestinale, stimola l’appetito, riduce l’incidenza delle emorroidi, sembra
promuovere l’attività fibrinolitica riducendo la probabilità di formazione di emboli.
In medicina la capsaicina è stata usata per alleviare il dolore. Viene oggi raccomandata
per medicazioni topiche contro le artriti, per scaldare i muscoli degli atleti prima della
performance o per ristabilire slogature, strappi e distorsioni. Creme contenenti capsaicina
vengono anche consigliate a pazienti che hanno subito mastectomia o amputazione, per
ridurre il dolore post-operazione. L’uso prolungato di queste creme sembra utile per ridurre il
prurito nei pazienti sottoposti a dialisi, il dolore causato dal cosiddetto “fuoco di
Sant’Antonio” (Herpes zoster), e il mal di testa (Carmichael, 1991). La ricerca medica studia
gli effetti della capsaicina per identificare i meccanismi di trasmissione della sensazione di
dolore; le fibre nervose coinvolte nella percezione del bruciore sono le stesse, i recettori
cutanei responsabili sembra siano diversi.
Repellenti a base di capsaicina sono anche consigliati agli escursionisti Nord americani
per evitare contatti troppo diretti con gli orsi; in alcuni stati del Sud-Est Asiatico sono
utilizzati contro i manifestanti; negli Stati Uniti la polizia locale utilizza questi spray contro i
criminali.

I-INTRODUZIONE
24
Fig. 27 C. annuum cv fiesta, ornamentale
Infine, le piante di peperone hanno una grande importanza ornamentale (Fig. 27):
ghirlande di peperoni rossi dette “ristras”, nel Nuovo Messico e nell’Arizona, vengono
utilizzate nelle feste di Halloween, Natale e per il giorno del Ringraziamento (Fig. 28). In
Indonesia, in Giappone e in Cina si utilizzano per le composizioni floreali. In Europa, i
giardinieri propongono un intero assortimento di varietà decorative con frutti eretti e
multicolore, spesso con portamento fascicolato dei rami fioriferi.
Fig. 28 Peperoni rossi, detti “chile ristras”

I-INTRODUZIONE
25
2-MAPPE
Nell’ultimo decennio lo sviluppo delle mappe molecolari è diventato uno strumento
essenziale per la ricerca e il miglioramento genetico delle specie vegetali (Kang et al. 2001).
Le mappe genetiche sature sono rappresentazioni grafiche dei singoli gruppi di associazione,
linkage group, in cui sono indicati l’ordine e la posizione relativa dei geni lungo il
cromosoma. Per sviluppare una mappa genetica è necessario analizzare come segregano i
marcatori molecolari in progenie ottenute con appropriati programmi di incrocio, calcolare i
valori di ricombinazione tra coppie di marcatori e risalire così alle loro posizioni sui gruppi
linkage (Barcaccia et al. 2000).
2.1 Marcatori molecolari: breve excursus
Un marcatore molecolare viene definito come: “un locus genomico rilevabile con sonde
( = probe) o primer ( = inneschi) specifici, che in virtù della sua presenza, permettono di
identificare inequivocabilmente il tratto cromosomico specifico con il quale si identifica e le
regioni che lo circondano alle estremita 3`e 5`” (Barcaccia et al. 2000).
Enrico Pè, del Dipartimento di Scienze Biomolecolari e Biotecnologie dell’Università di
Milano, definisce i marcatori molecolari: “Caratteristiche ereditarie che presentano una certa
variabilità tra gli individui di una popolazione e che contraddistinguono la regione
cromosomica che li contiene”.
Un marcatore molecolare ideale deve rispondere ai seguenti requisiti:
- polimorfico: deve permettere di rilevare la variabilità;
- multiallelico: presentare più di due alleli alternativi in uno stesso locus genico;
- codominante: permettere di distinguere la condizione eterozigote da quella dei due
omozigoti, (il prodotto ibrido ha le caratteristiche di entrambe);
- stabile: non deve essere influenzato dall’ambiente o avere effetti epigenetici;
- neutro: le sostituzioni alleliche a livello del locus non hanno altri effetti fenotipici,
(dunque non comportano effetti selettivi), ma permettono solo di determinare il suo
genotipo;
- non epistatico: i locus analizzzati non devono essere influenzati dall’informazione
genetica presente in un altra zona del genoma, ovvero non devono esserci interazioni
inter o intralocus;

I-INTRODUZIONE
26
- dispersibile nell’intero genoma;
- semplice da rilevare;
- basso costo;
- eredità mendeliana;
- riproducibile.
Si possono distinguere due classi principali di marcatori molecolari: quelli basati su
tecniche di ibridazione e quelli basati sull’impiego della reazione di PCR. In entrambe i casi i
polimorfismi a carico del DNA vengono studiati sfruttando la tecnica dell’elettroforesi, che
permette di separare i frammenti di DNA ottenuti, in funzione della loro dimensione. Il DNA
è infatti carico negativamente e migra verso il polo positivo. L’elettroforesi può essere
effettuata su supporti di agarosio o poliacrilamide.
I marcatori molecolari possono essere classificati anche in funzione al numero di loci
che permettono di analizzare in un singolo esperimento. I marcatori multi-locus, permettono
di analizzare diversi loci, o forme alleliche, contemporaneamente: i.e. RAPD, AFLP, SSR,
ISSR, S-SAP, VNTR. Sono solitamente marcatori dominanti, cioè non permettono la
distinzione tra la situazione di omozigosi (a/a) e eterozigosi (a/-) a livello dei loci analizzati. I
marcatori single-locus permettono l’analisi di un locus unico come nel caso degli RFLP, SSR,
STS, SCAR, STMS. Sono solitamente marcatori co-dominanti cioè permettono la distinzione
tra la situazione di omozigosi (a/a) e eterozigosi (a/-) del locus analizzato (Fig. 29). Nel primo
caso, a/a viene visualizzata un’unica banda sul gel; nel secondo caso a/-, due bande, ciascuna
relativa ad un allele diverso.
Fig. 29 Marcatori co-dominanti: distinzione dell’eterozigote dai due omozigoti.
AA Aa aa

I-INTRODUZIONE
27
I marcatori molecolari maggiormente impiegati nei laboratori di ricerca e le loro
caratteristiche sono riassunti in tabella. Mi soffermerò a descrivere solamente quelli utilizzati
nel mio lavoro sperimentale: i microsatelliti e i dCAPS.
Tab. 2 Principali marcatori molecolari e loro caratteristiche.
MARCATORE POLIMOR-
FISMO DOMINANZA
MULTI / SINGOLO
LOCUS
RIPRODU-
CIBILITA'
COMPLESSI-
TA'
RFLP
(Restiction Fragment
Length Polymorphism)
Medio Co-dominante Multi locus Molto
riproducibile Elevata
CAPS
(Cleaved Amplified
Polymorphic Sequence)
Limitato Co-dominante Singolo locus Riproducibile Semplice
RAPD
(Random-Amplified
Polymorphic DNA)
Basso Dominante Multi locus Poco
riproducibile Semplice
SSR
(Simple Sequenze
Repeats)
Elevato Co-dominante Multi locus Molto
riproducibile Media
AFLP
(Amplification Fragment
Length Polymorphisms)
Basso Dominante Multi locus Molto
riproducibile Complessa
SNP
(Single nucleotide
Polymorphysms)
Elevato Co-dominante Singolo locus Molto
riproducibile Complessa

I-INTRODUZIONE
28
2.1.1 I Microsatelliti.
Sono anche detti Simple Sequence Repeats (SSRs) (Jacob et al. 1991), Short Tandem
Repeats (STR) (Edwards et al. 1991) o Sequence Tag Microsatellite Sites (STMS), sono delle
corte sequenze di basi (1-6 bp) ripetute a tandem e interdisperse nei genomi eucariotici,
soprattutto a livello delle regioni non codificanti (Tautz e Renz 1984). I marcatori SSR
rispondono ai seguenti requisiti: abbondanza, elevato polimorfismo, riproducibilità,
codominanza, alta risoluzione e portabilità all’interno dello stesso genere. Possono essere
distinti in: 1) perfetti: stessa sequenza ripetuta n volte, (CA)n; 2) imperfetti: sequenze ripetute
n volte talvolta interrotte da 7-8 bp casuali, (CA)n GCATTCG (CA)n; 3) composto: due tipi di
sequenze ripetute n volte e alternate, (CA)n(GC)n . I polimorfismi che si osservano sono legati
alla presenza di varianti alleliche che differiscono nella lunghezza della sequenza ripetuta, nel
numero di ripetizioni della sequenza base o nella presenza di mutazioni puntiformi all’interno
del microsatellite (Lee et al. 2004). Le variazioni nel numero di ripetizioni sono dovute alla
strange-slippage replication: scivolamento della polimerasi durante la replicazione del DNA
che comporta l’aggiunta o la delezione di basi ripetute (Schlötterer e Tauz 1992), oppure
derivano da eventi di crossing-over ineguale tra cromatidi omologhi (Innan et al. 1997). Le
regioni fiancheggianti i microsatelliti sono invece molto conservate nei genotipi appartenenti
alla stessa specie. Costruendo su queste regioni primer specifici è possibile amplificare
mediante Polymerase chain reaction, (PCR) (Kary Mullis 1986) frammenti contenenti SSR e
analizzare i polimorfismi esistenti (Lee et al. 2004).
La maggior parte dei loci microsatelliti è definita da un’unica coppia di primer, cosa che
rende più facile lo scambio di informazioni tra laboratori (Fig. 30). Questi marcatori sono
strumenti largamente usati per: i) identificazione varietale di cultivar e accessioni (Thomas e
Scott 1993); ii) costruzione di mappe genetiche (Bowcock et al. 1994; Röder et al. 1998); iii)
studi di genetica delle popolazioni e conservazione delle risorse genetiche (Terauchi e
Konuma 1994; Innan et al. 1996; Zane et al. 2002). I limiti maggiori dei microsatelliti sono
legati al fatto che spesso devono essere isolati de novo, soprattutto nel caso di specie poco
studiate dal punto di vista genomico. Sono disponibili a riguardo diverse tecniche: screening
tradizionali, con sonde SSR, di librerie classiche o arricchite (Nybom et al. 1990), tecniche
basate sulla reazione PCR: l’AFLP (Vos et al. 1995), la MFLP proposta da Yang et al. 2001 e
Albertini et al. 2003, la Microsatellite Amplified library, MAL (Acquadro et al. 2003) e molte
altre tecniche reperibili in letteratura.

I-INTRODUZIONE
29
Fig. 30 Rappresentazione schematica della della natura del polimorfismo single-locus dei marcatori SSR. I modelli di
bande al locus microsatellite sono diversi a seconda della situazione allelica: l'individuo eterozigote Aa mostra due bande
mentre quelli omozigoti AA e aa soltanto una.
Marcatori SSR
AA Aa aa -
+
A
a
CTCTCTCTCTCTCTCTCTC
GAGAGAGAGAGAGAGAG
CTCTCTCTCTC
GAGAGAGAGAG
= Coppia di primer
specifici

I-INTRODUZIONE
30
2.1.2. I dCAPS.
I derived Cleaved Amplified Polymorphic Sequence sono marcatori molecolari utili per
valutare la segregazione di singole mutazioni note in popolazioni segreganti, qualora non
siano localizzate in un sito di restrizione (Neff et al. 1998).
I dCAPS derivano dalla tecnica Cleaved Amplified Polymorphic Sequence (CAPS) che
è vincolata all’analisi di mutazioni associate a siti di restrizione specifici (Fig. 31). In questa
tecnica, infatti, si amplificano determinati tratti di DNA genomico, recanti un single
nucleotide polymorphism (SNP) all’interno di un sito di restrizione. Gli ampliconi vengono
digeriti con l’enzima opportuno e il polimorfismo, assenza o presenza del sito di taglio, viene
rilevato mediante elettroforesi (Konieczny e Ausubel 1993).
Nella strategia dCAPS vengono utilizzati primer particolari, contenenti mismatches, che
amplificano la sequenza CAPS specifica e inseriscono un sito di restrizione nell’amplicone
che presenta la SNP. Il prodotto PCR, così modificato, viene sottoposto a digestione
enzimatica ed a elettroforesi per evidenziare il polimorfismo: assenza o presenza della SNP
(Fig. 32). Il metodo è virtualmente in grado di identificare qualsiasi SNPs. Come la tecnica
CAPS, è semplice, relativamente a basso costo in quanto impiega tecnologie e reagenti
comunemente usati nella maggior parte dei laboratori di ricerca. La strategia dCAPs può
dunque essere sfruttata per sviluppare marcatori molecolari associati a geni di interesse
agronomico qualora presentino polimorfismi di tipo SNPs, peraltro piuttosto frequenti nei
genomi eucarioti.
Fig. 31. Tecnica CAPs.

I-INTRODUZIONE
31
E’ inoltre disponibile un programma online, “dCAPS Finder”, che facilita lo sviluppo di
questi marcatori. Inserendo nel programma la porzione della sequenza di un gene noto,
contenente la SNPs di interesse, “dCAPS Finder” fornisce un elenco di possibili primers
dCAPS in associazione ai relativi enzimi di restrizione, indispensabili per rilevare il
polimorfismo e studiarne la segregazione.
Fig. 32 Esempi di polimorfismi generati dall’uso di marcatori dCAPS.

I-INTRODUZIONE
32
2.2 Le mappe genetiche.
La prima mappa genetica è stata quella del cromosoma X di Drosophila melanogaster,
sviluppata grazie agli studi di Morgan del 1911 (Fig. 33); successivamente sono state costruite
mappe in mais e in pomodoro, mediante analisi della segregazione di caratteri morfologici ad
eredità semplice, il cui numero era molto esiguo.
Fig. 33 Morgan
La mappa fisica indica la localizzazione fisica dei geni sui cromosomi e si ottiene
mediante sequenziamento e assemblaggio dei cloni contenuti in una libreria genomica; l’unità
di misura impiegata è il numero di nucleotidi. La mappa genetica definisce l’ordine dei geni e
di altre strutture, quali centromeri, telomeri e zone N.O.R. (Nucleolar Organizer Regions),
lungo i cromosomi in termini di distanza di ricombinazione. La costruzione delle mappe
genetiche sfrutta le modalità di segregazione dei geni: due geni appartenenti ad uno stesso
cromosoma vengono ereditati assieme e sono detti linked, associati. Talvolta, durante il
processo di meiosi, (profase I), può verificarsi crossing-over, cioè scambio di materiale
genetico tra cromosomi omologhi; si parla di ricombinazione, fenomeno che interrompe
l’associazione tra i due geni (Fig. 34 e 35).

I-INTRODUZIONE
33
Fig. 34 Evento di crossing-over.
La frequenza di ricombinazione, probabilità che avvenga un crossing-over, è
direttamente proporzionale alla distanza tra i geni: maggiore è la distanza, maggiore è la
probabilità che venga interrotta l’associazione tra gli alleli. Le meiosi caratterizzate da un
evento di crossing-over tra due loci danno luogo a un 50% di gameti parentali e un 50% di
gameti ricombinanti. In realtà, non è detto che il crossing-over si verifichi in tutte le meiosi:
in caso di geni linked la percentuale di gameti ricombinanti è inferiore al 50%, (frequenza
attesa in presenza di segregazione indipendente).
Fig. 35 Possibili eventi di crossing-over.
SINAPSI di CROMOSOMI OMOLOGHI
DUPLICAZIONE dei CROMOSOMI e FORMAZIONE delle
TETRADI
FORMAZIONE del
CHIASMA
RISULTATO dell’EVENTO di CROSSING-OVER
CROSSING-OVER
tra w e m
NO CROSSING-OVER
CROSSING-OVER
tra y e w
DOPPIO
CROSSING-OVER

I-INTRODUZIONE
34
Le distanze genetiche possono essere calcolate mediante test a due punti o a tre punti,
sulla base dei confronti fra dati di segregazione di coppie di marcatori molecolari. Le distanze
sono espresse in unità di ricombinazione o unità di mappa o centimorgan (cM). 1 cM è pari a
una distanza fra geni che fa ottenere un gamete ricombinante, portatore di un gamete
crossover, ogni 100 prodotti della meiosi (25 meiosi). 1 cM è la distanza alla quale la
percentuale di ricombinazione tra i due geni e pari all’1%.
2.2.1 Test a due punti
Il test a due punti permette di definire le associazioni e calcolare le distanze genetiche
sulla base delle frequenze di ricombinazione tra coppie di marcatori. In seguito a una serie
numerosa di confronti tra i dati, viene assegnata una posizione a ciascun marcatore in
relazione agli altri geni dello stesso gruppo.
Per spiegare il test verrà descritto l’esperimento di Hutchinson, condotto sul mais.
L’obiettivo è stabilire se il gene dell’aleurone (C/c) e quello per la forma della cariosside
(Sh1/sh1) sono localizzati sullo stesso cromosoma e identificarne la distanza reciproca.
Hutchinson effettuò l’incrocio CCSh1Sh1 x ccsh1sh1. Reincrociò l’ibrido, CcSh1sh1, con il
doppio omozigote recessivo, ccsh1sh1. Nella progenie non ottenne il rapporto fenotipico
1:1:1:1 atteso sulla base della segregazione indipendente dei quattro gameti, ma un rapporto
fenotipico diverso. Siccome due gameti erano presenti in frequenza nettamente superiore
rispetto agli altri due, concluse che i due geni erano situati sullo stesso cromosoma e
appartenevano allo stesso gruppo linkage. In questo esperimento la frequenza di
ricombinazione è pari al 3,6%, cioè i due geni si trovano a una distanza di 3,6 cM. Tale
percentuale è bassa, cioè i geni sono molto vicini sul cromosoma e tendono a segregare
insieme. Il test a due punti permette di risalire, qualora non si conoscessero, ai genotipi dei
parentali del di-ibrido. I gameti parentali prodotti sono riconoscibili in quanto sono sempre
presenti, ciascuno in percentuale superiore al 25%.

I-INTRODUZIONE
35
2.2.2 Test a tre punti.
I cromatidi di un cromosoma possono andare incontro a più crossing-over nello stesso
tempo e questa situazione può influenzare i tipi di gameti che si formano. Il verificarsi
contemporaneo di due crossing-over è detto doppio crossing-over (Fig. 36). Nei cromosomi
non crossover o crossover doppi non si osserva ricombinazione tra i geni terminali, tale
ricombinazione è presente nei cromosomi crossover singoli.
Fig. 36 Doppio crossing-over.
Quando tra due geni la distanza è notevole, superiore a 20 cM, i crossover doppi sono
piuttosto frequenti; siccome però non si manifestano ricombinazioni tra i geni terminali, il test
a due punti fornisce distanze viziate per difetto. Il test a tre punti permette di ovviare a questo
difetto prendendo in esame tre locus genici e calcolando le distanze tra i due geni terminali
come somma di distanze parziali (Fig. 37).
Fig. 37 Doppio crossing-over nel caso di tre locus
Per spiegare il test verrà descritto l’esperimento di Stadler. Egli incrociò una linea di
mais omozigote per gli alleli recessivi: c (cariosside bianca), sh1 (cariosside collassata) e wx
(cariosside cerosa), con una linea omozigote per i tre alleli dominanti: C (cariosside colorata),
Sh1 (cariosside piena), Wx (cariosside con endosperma normale).

I-INTRODUZIONE
36
Reincrociò poi l’ibrido F1 con la linea omozigote recessiva cc sh1sh1 wxwx e ottenne i
seguenti risultati:
- le otto classi fenotipiche non sono ugualmente rappresentate, come atteso in caso di
segregazione indipendente, quindi i geni non sono localizzati su coppie di cromosomi
omologhi;
- i ricombinanti crossover nelle zone C Sh1 e Sh1 Wx sono presenti in percentuale
significativamente diversa dal 50%, cioè i tre geni sono localizzati sullo stesso cromosoma;
- le due classi fenotipiche più numerose: C Sh1 Wx e c sh1 wx, sono le classi parentali,
le altre sei, meno numerose, sono le classi ricombinanti;
- le classi fenotipiche simmetriche: C Sh1 Wx e c sh1 wx; C Sh1 wx e c sh1 Wx; C sh1
wx e c Sh1 Wx; C sh1 Wx e c Sh1 wx, hanno frequenze simili;
- tra le sei classi ricombinanti, le due meno rappresentate derivano da crossing-over
doppi. Queste due classi permettono di stabilire quale dei tre geni occupa la posizione centrale
sul cromosoma: è quello che, in queste classi, ha cambiato la sua posizione relativa rispetto ai
gameti parentali. Nell’esperimento di Stadler è centrale il gene Sh1.
- la distanza tra due geni (esempio C e Sh1) si ottiene dal rapporto tra i ricombinanti
(per C e Sh1) e i fenotipi osservati. C e Sh1 distano 2,31 cM; Sh1 e Wx, 19,93 cM. La
distanza tra C e Wx si calcola sommando le frequenze di ricombinazione parziali appena
calcolate, è dunque 22,24 cM.

I-INTRODUZIONE
37
2.2.3 Interferenza e Funzione di Kosambi
Nel valutare le frequenze di ricombinazione è importante tenere conto del fenomeno
dell’interferenza. La probabilità che avvenga un crossing-over dipende dai crossing-over già
verificatisi in regioni vicine: un evento di crossing-over tende a inibire il verificarsi di una
seconda ricombinazione nella stessa regione cromosomica. Il coefficiente di coincidenza è il
valore che si ottiene dal rapporto tra la frequenza dei doppi crossover osservati e quella dei
doppi crossover attesi, e permette di risalire al valore del coefficiente di interferenza (= 1-
coefficiente di coincidenza), che varia tra 0 e 1.
Sono state elaborate funzioni di mappaggio che tengono conto dell’insorgenza
indipendente di più crossing-over in una stessa regione cromosomica e del fenomeno
dell’interferenza.
La funzione più utilizzata in ambito vegetale è quella di Kosambi (1944):
xAB= ¼ ln [(1 + 2*cAB)/(1 – 2*cAB)]
dove: cAB : è la frequenza di ricombinazione tra i geni A e B.
Il valore di interferenza diminuisce all’aumentare della distanza tra i geni: è zero nel
caso di loci non associati, con c=0,5; xAB ≅ cAB quando la frequenza di ricombinazione è
molto ridotta.
Un’altra funzione disponibile è quella di Haldane:
xAB= - ½ ln (1-2*cAB).
I-INTRODUZIONE
38
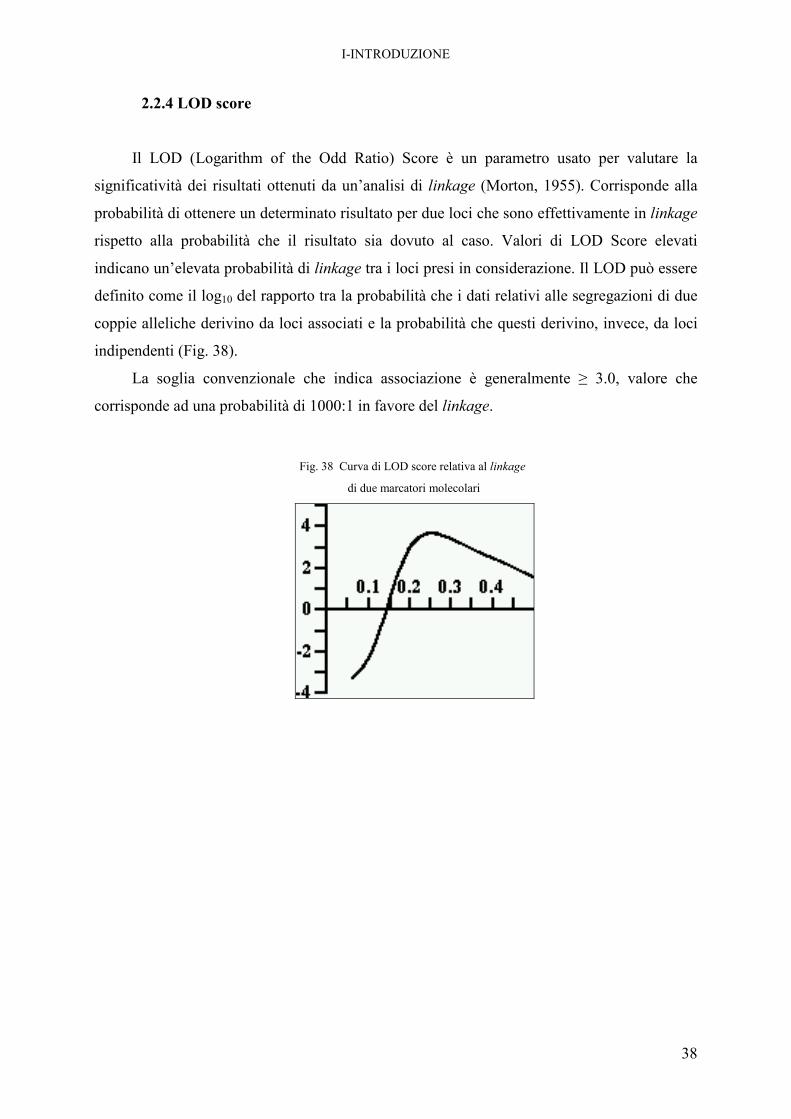
2.2.4 LOD score
Il LOD (Logarithm of the Odd Ratio) Score è un parametro usato per valutare la
significatività dei risultati ottenuti da un’analisi di linkage (Morton, 1955). Corrisponde alla
probabilità di ottenere un determinato risultato per due loci che sono effettivamente in linkage
rispetto alla probabilità che il risultato sia dovuto al caso. Valori di LOD Score elevati
indicano un’elevata probabilità di linkage tra i loci presi in considerazione. Il LOD può essere
definito come il log10 del rapporto tra la probabilità che i dati relativi alle segregazioni di due
coppie alleliche derivino da loci associati e la probabilità che questi derivino, invece, da loci
indipendenti (Fig. 38).
La soglia convenzionale che indica associazione è generalmente ≥ 3.0, valore che
corrisponde ad una probabilità di 1000:1 in favore del linkage.
Fig. 38 Curva di LOD score relativa al linkage
di due marcatori molecolari

I-INTRODUZIONE
39
2.3 Popolazioni segreganti di mappaggio
Mappe genetiche di interi genomi possono essere sviluppate sulla base dei valori di
ricombinazione analizzando la segregazione di marcatori molecolari in progenie ottenute con
appropiati programmi di incrocio: Back-Cross (BC1), F2, Recombinant Inbred Lines (RIL),
Di-Haploids (DH), Pseudo-Test-Cross (F1).
2.3.1 Back-Cross (o reincrocio, BC1)
Il reincrocio è il metodo migliore per stabilire se i due geni marcatori si trovano sullo
stesso marcatore e per calcolare la loro distanza relativa. Infatti i rapporti fenotipici che si
ottengono esprimono fedelmente i rapporti tra i gameti prodotti dall’ibrido. La progenie Back-
Cross si ottiene dall’incrocio tra un ibrido F1, eterozigote a tutti i loci, e un individuo
altamente omozigote. Solitamente viene incrociato con uno dei suoi parentali (detto
ricorrente), entrambe altamente omozigoti (Fig. 39). Nel reincrocio del diibrido si ottengono
due fenotipi parentali e due ricombinanti: questi ultimi, in caso di associazione, sono
immediatamente riconoscibili perché sempre presenti in una misura complessiva inferiore al
50 %. Per una frequenza non significativamente diversa dal 50 %, infatti, si deve ipotizzare
segregazione indipendente.
Fig. 39 Popolazione Back-cross.
P1 x P2
F1 x P2
BC1
Back-Cross

I-INTRODUZIONE
40
Il Back-Cross è utile nel caso di specie annuali autogame. Può essere impiegato anche
per specie allogame, qualora non manifestino un inbreeding troppo marcato quando
autofecondate al fine di ottenere le linee pure, parentali, di partenza. Le popolazioni BC1 non
sono perpetuabili nel tempo; non permettono dunque la saturazione progressiva delle mappe
genetiche, la valutazione di caratteri quantitativi soggetti a diversa influenza ambientale e il
trasferimento delle progenie segreganti tra laboratori.
2.3.2 La F2
La popolazione F2 si ottiene per autofecondazione o incrocio di due individui F1 (Fig.
40) altamente eterozigoti, originati da due parentali altamente omozigoti. In caso di
segregazione indipendente si osserva un rapporto fenotipico 9:3:3:1, con i geni marcatori
localizzati su cromosomi differenti. In caso di segregazione non indipendente, i rapporti
fenotipici delle due classi parentali si avvicinano alla situazione 3:1, (seconda legge di
Mendel). Le classi ricombinanti hanno frequenza inferiore, variabile in funzione alla distanza
dei loci; i geni marcatori analizzati sono in questo caso linked.
Fig. 40 Popolazione F2
P1 x P2
F1 ����
F2
Popolazione F2
F1 x F1

I-INTRODUZIONE
41
La popolazione F2 fornisce una quantità di informazioni teoricamente doppia rispetto
alla BC1, perché i marcatori segregano sia nei gameti maschili che in quelli femminili della
popolazione F1. Risulta tuttavia difficile analizzare in dettaglio le regioni cromosomiche
caratterizzate dalla presenza di marcatori strettamente associati. Come la BC1, anche la F2 e
indicata nel caso di specie autogame o allogame e fornisce popolazioni non perpetue.
2.3.3 Recombinant Inbred Lines (RILs)
Le linee inbred ricombinanti (RILs) si ottengono per autofecondazione a partire dalla
F2, scegliendo in ciascuna generazione un solo individuo, che originerà, per autofecondazione
la generazione successiva. Si parla di discendenza SSD, cioè Single Seed Descendent. Si
effettua inizialmente un incrocio tra due parentali P1 e P2, caratterizzati da certe combinazioni
genetiche, per ottenere un ibrido F1, altamente eterozigote. L’ibrido viene autofecondato per
ottenere una progenie F2 costituita da un insieme di individui che, per quella data coppia di
cromosomi, presenta certe combinazioni, frutto della ricombinazione meiotica. Ad ogni
generazione di autofecondazione, l’eterozigosi media del genoma decresce rapidamente, al
ritmo di 1/2n (dove n è il numero di generazioni dopo F1). In F5 il 93,75 % del genoma è
fissato (1- 1/24), in F6 il 96.88 % e in F10 il 99,80 %. Si può affermare che dopo 6-7
generazioni di autofecondazione, è possibile ottenere linee ricombinanti altamente omozigoti
che avranno fissato gli alleli parentali in brevi blocchi di associazione. Dopo n generazioni,
gli individui ottenuti da un individuo pressoché omozigote, sono pressoché identici
geneticamente e ciascuna famiglia avrà fissato combinazioni alleliche diverse, sulla base delle
ricombinazioni subite. L’analisi finale consiste nel confronto tra la distribuzione degli alleli
nelle linee RI e quella nei parentali. Quando due loci sono strettamente concatenati, gli alleli
parentali sono mantenuti assieme molto frequentemente. Se, viceversa, i loci sono meno
strettamente concatenati, gli alleli parentali si presentano insieme con una frequenza minore.
Il grosso vantaggio delle linee ricombinanti è che possono essere mantenute nel tempo,
mediante autofecondazione, in quanto costituiscono una popolazione perpetua. Offrono
inoltre la possibilità di aggiungere, in tempi successivi, nuovi marcatori a una mappa
precedentemente prodotta. Le linee ricombinanti, infatti, possono essere utilizzate da genetisti
che lavorano indipendentemente e che condividono un database a cui tutti contribuiscono
arricchendolo continuamente. Sono molto utili per sviluppare mappe fini (sature).

I-INTRODUZIONE
42
2.3.4 Di-Haploids (DH)
Le popolazioni Di-aploidi possono essere prodotte mediante androgenesi (coltura di
antere o polline), o ginogenesi (coltura di ovuli) e successivo raddoppiamento cromosomico
delle piante aploidi ottenute. Viene sfruttata la ricombinazione che si verifica durante la
meiosi della popolazione F1, altamente eterozigote. Dopo una sola generazione è possibile
ottenere linee completamente omozigoti, fissate a tutti i loci. Come le linee RI, le DH possono
essere mantenute nel tempo mediante autofecondazione, offrono la possibilità di aggiungere,
in tempi successivi, nuovi marcatori a una mappa precedentemente prodotta e consentono di
sviluppare mappe fini (sature).
2.3.5 Pseudo-Test-Cross (F1)
Per sviluppare le popolazioni di mappaggio precedentemente descritte è indispensabile
avere a disposizione delle linee pure di partenza, altamente omozigoti. Alcune specie sono
molto suscettibili all’inbred e non è possibile disporre di linee pure. In questi casi si ricorre
all’impiego di popolazioni F1, Pseudo-Test-Cross (Fig. 41). Si parla di Two-way Pseudo-Test-
Cross quando la popolazione F1, utilizzata per la mappa, deriva dall’incrocio di due individui
parentali altamente eterozigoti. Si parla di One-way Pseudo-Test-Cross quando la popolazione
F1 deriva dall’incrocio tra un individuo parentale eterozigote e uno altamente omozigote
(Grattapaglia e Sederoff, 1994).
Fig. 41 Popolazione Pseudo-Test-Cross.
P1 x P2
F1
F1
Pseudo-Test-Cross
P1 x linea omozigote
Two-way:
One-way:

I-INTRODUZIONE
43
L’analisi genetica coincide con quella eseguita con il metodo del reincrocio. Nel caso
del Two-way Pseudo-Test-Cross vengono seguite separatamente le segregazioni dei marcatori
in condizione eterozigote in ciascuno dei due parentali; vengono prodotte due mappe distinte,
una per ciascuno dei due parentali utilizzati nell’incrocio iniziale. I loci codominanti
eterozigoti nei parentali, per i medesimi alleli, possono eventualmente fungere da loci-ponte
per eseguire un allineamento tra le due mappe. Nel caso del One-way Pseudo-Test-Cross si
produce una sola mappa genetica, quella per il genotipo parentale eterozigote. L’analisi di
segregazione può essere condotta unicamente per gli alleli marcatori segreganti dal genotipo
parentale altamente eterozigote.
Lo Pseudo-Test-Cross è adatto per specie allogame, altamente eterozigoti, con lungo
ciclo di vita, ad esempio piante arboree, perché la popolazione da mappare si ottiene con una
sola generazione da incrocio.

I-INTRODUZIONE
44
2.4 Software per la costruzione di mappe genetiche.
Sono disponibili diversi programmi informatici per la costruzione di mappe genetiche,
che differiscono in funzione al tipo di algoritmo utilizzato per ordinare i marcatori e al tipo di
popolazione in analisi. Nel caso di progetti di mappa per famiglie relativamente piccole,
genoma umano o bovino, per le quali sono sufficienti informazioni relative a tre generazioni,
si utilizza LINKAGE o CRIMAP. Nel caso di grandi discendenze derivate dall’incrocio di due
linee parentali fissate, tipiche dei genomi vegetali, il software più utilizzato è MAPMAKER
(Lander et al. 1987). Il programma è in grado di elaborare i dati ottenuti da popolazioni BC1,
DH, F2 e RIL. Il mappaggio è sequenziale: in un primo tempo MAPMAKER calcola le
frequenze di ricombinazione e il LOD Score associato a tutte le coppie di marcatori, presi due
a due. E’ necessario fissare un limite superiore alla percentuale di ricombinazione, e un limite
inferiore al valore di LOD Score per stabilire gruppi linkage dove i marcatori sono l’uno
vicino all’altro. La tappa successiva consiste nel definire l’ordine dei loci in ciascun gruppo di
associazione. Infine, si calcola la distanza tra i marcatori che viene espressa in cM, come
distanza di Kosambi o di Haldane. Il programma MAPMAKER / EXP Version 3.0 è stato utilizzato
per ottenere la mappa inter-specifica di Capsicum nell’ambito del lavoro sperimentale di tesi.
E’ stato impiegato anche il software Map.Pop 1.0 (Brown et al. 2000), molto utile per il
mappaggio selettivo. Il comando SAMPLEXP permette infatti di selezionare nella popolazione
di mappaggio, un campione altamente significativo, su cui testare ulteriori marcatori
molecolari da inserire nella mappa ad alta risoluzione, limitando le operazioni di mappaggio.

I-INTRODUZIONE
45
3- Marcatori e Mappe genetico molecolari sviluppate in
Capsicum spp.
Pochard, nel 1970, identificò in peperone la cosiddetta “serie dei trisomici”, essi
rappresentarono il primo materiale utilizzato per il mappaggio in peperone. Pochard attribuì ai
12 individui trisomici i nomi: violet, indigo, bleu, vert, jaune, orange, rouge, pourpre, noir,
brun, bistre, gris. Lo studio effettuato su progenie segreganti, aventi come parentali i
trisomici, permise di localizzare dieci caratteri monogenici al trisomico Violet (cromosoma I),
Jaune (cromosoma XI) e Pourpre (cromosoma XII) (Pochard 1977; Pochard e Dumas de
Vaulx 1982).
Successivamente, lo sviluppo dei marcatori, biochimici e molecolari, ha permesso un
notevole approfondimento degli studi sul genoma di Capsicum (Fig. 41). Nel 1984, Tanksley
analizzò la segregazione di marcatori isoenzimatici in una progenie BC1, ottenuta
dall’incrocio inter-specifico tra i parentali C. annuum cv NM6-4 e C. chinense CA4,
utilizzando il parentale NM6-4 come ricorrente. Dall’analisi vennero identificati 4 isoenzimi
polimorfici nella popolazione segregante che sono stati attribuiti a 4 gruppi linkage.
Negli ultimi 15 anni, l’impiego dei marcatori molecolari ha permesso lo sviluppo di
numerose mappe genetico-molecolari in Capsicum, tra cui le più importanti sono elencate in
tabella 3.
Fig. 42 C. chacoensis

I-INTRODUZIONE
46
Tabella 3 Principali mappe genetiche di Capsicum pubblicate.
Autori Parentali e Tipo
di popolazione
n° marcatori
mappati
n°
Gruppi
Linkage
Taglia
(cM)
Tanksley
(1984)
80-153 BC: (C.annuum cv. NM6-4 x
C.chinense CA4)x NM6-4
85-295 F2: (C.annuum cv. NM6-4 x
C.chinense CA4)
9 isoenzimi 4 ~ 52
Tanksley et
al. (1988)
46 BC: (C.annuum Doux des Landes CA50 x
C.chinense CA4)x CA50
80 marcatori
(RFLP + isoenzimi)
14 634
Prince et al.
(1993)
46 F2: (C.annuum CA133 x C.chinense CA4)
192 marcatori
(RFLP + isoenzimi)
19 720
Lefebvre et
al. (1995)
Mappa integrata intraspecifica, C.annuum:
94 DH Perennial x Yolo Wonder + 44 DH Vat
x CM334 + 31 DH Yolo x CM334
85 marcatori
(RFLP + RAPD)
14 823
Lefebvre et
al. (1997)
94 DH Perennial x Yolo Wonder
98 DH H3 x Vania
83-189 marcatori
(RFLP + RAPD)
15-16 417-1515
Kim et al.
(1997)
86 F2: (C.annuum 2002 x C.chinense 1679)
174 marcatori
(AFLP) 12 n.a.
Livingstone
et al. (1999)
75 F2: (C.annuum NuMex Rnaky x C.chinense
Habanero)
677 marcatori
(isoenzimi + RFLP
+ RAPD + AFLP)
13 1246
Kang et al.
(2001)
107 F2: (C.annuum TF68 x C.chinense
Habanero)
580 marcatori (RFLP
+ AFLP) 16 1320

I-INTRODUZIONE
47
Tabella 3 Principali mappe genetiche di Capsicum pubblicate (continua).
Autori Parentali e Tipo
di popolazione
n° marcatori
mappati
n°
Gruppi
Linkage
Taglia
(cM)
Ben Chaim et
al. (2001b)
180 F2: C.annuum: (Maor x Perennial)
177 marcatori
(RFLP + RAPD +
AFLP + morfologici)
12 1740
Lefebvre et
al. (2002)
Mappa intraspecifica, C.annuum: 101 DH: (H3
x Vania) + 114 DH: (Perennial x Yolo
Wonder) + 151 F2: (Yolo Wonder x CM334)
208-630 marcatori
(RFLP + RAPD +
AFLP + morfologici)
12
685-1668 (H)
Paran et al.
(2003)
75 F2: (C.annuum NuMex Rnaky x C.chinense
PI159234) + 83 BC1: [(C.annuum 100/63 x
C.chinense PI152225) x 100/63] + 180 F2::
(C.annuum Maor x C.annuum Perennial) + 101
DH: (H3 x Vania) + 114 DH C.annuum:
(Perennial x Yolo Wonder) + 151 F2: (Yolo
Wonder x CM334)
2262 marcatori
nella mappa integrata
13 1832
Sasvári et al.
2004
238 F2: C. annuum (linea inbred di P4) x C.
frutescens (cv. Tabasco)
429 (278 AFLP + 85
SSR + 3 morfologici,
63 PCR-based
markers)
13 1280
Sugita et al.
(2005a)
Ogundiwin et
al. (2005)
176 DH: C. annuum L. (K9-11 x AC2258)94
RILs: C. annuum (PSP-11 x PI201234)
94 F2: C.annuum (JEP x CM334)
144
113
518 (AFLP + RAPD +
SSR + SCAR +
morfologici)
16
17
16
1466.1
1089.2
1043.1

I-INTRODUZIONE
48
I primi marcatori molecolari che sono stati sviluppati e impiegati nella costruzione delle
mappe sono stati gli RFLP. Essi sono in grado di identificare un elevato livello di
polimorfismi e sono stati inseriti anche in mappe di peperone di più recente sviluppo.
Successivamente sono state impiegate altre classi di marcatori molecolari quali: RAPD,
AFLP, SSR, e i marcatori di ultima generazione tra i quali i marcatori STS, sviluppati su geni
noti, gli SSAP e le SNPs.
Le popolazioni di mappaggio impiegate per lo sviluppo delle mappe molecolari
comprendono progenie F2, BC1 (Back-cross), RILs (Recombinant Inbred Lines) o DH
(double-haploid), ottenute sia da incroci inter che intra-specifici. Nei paragrafi seguenti
verranno descritte con maggior dettaglio le principali mappe sviluppate in Capsicum (Fig.
43).
Fig. 43 Pianta di Tabasco

I-INTRODUZIONE
49
3.1 Mappe genetico molecolari inter-specifiche
Lo sviluppo di mappe a partire da incroci inter-specifici offre il vantaggio di ottenere
progenie segreganti caratterizzate da elevati livelli di polimorfismo. Per contro, le progenie
inter-specifiche mostrano spesso bassi livelli di fertilità, segregazione distorta e, come
dimostrato negli studi effettuati da Lanteri et al. (1991) e da Lanteri e Pickersgill (1993),
fenomeni di riarrangiamenti strutturali cromosomici insorti durante la differenziazione delle
specie.
La maggior parte delle mappe inter-specifiche sono state sviluppate dall’analisi
molecolare di popolazioni ottenute dall’incrocio tra C. annuum x C. chinense, in quanto è un
incrocio fertile che permette di sviluppare progenie molto polimorfiche. Due mappe inter-
specifiche sono state sviluppate utilizzando una progenie F2 ed una popolazione BC1, ottenute
dall’incrocio C. annuum x C. frutescens.
I risultati più significativi degli studi sopra menzionati sono brevemente riportati a
seguito.

I-INTRODUZIONE
50
3.1.1 Livingstone et al. (1999): “Genome Mapping in Capsicum and the Evolution
of Genome Structure in the Solanaceae” Genetics.
Livingstone et al. (1999) hanno sviluppato una mappa inter-specifica di Capsicum,
utilizzando una popolazione di mappaggio F2, che è stata ottenuta autofecondando gli ibridi F1
prodotti dall’icrocio tra C. annuum “NuMex Rnaky” (Nakayama e Matta, 1985) x “C.
chinense PI 159234” (sviluppata da Tanksley alla Cornell Universisty, Ithaca, NY). Sono stati
impiegati marcatori RFLP, AFLP, RAPD e isoenzimi. La mappa è illustrata in figura 44.
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER,
Version 3.0b (Lincoln et al. 1993); i marcatori molecolari sono stati assemblati in gruppi
linkage mediante il comando “group” con LOD score maggiore a 3.0 e percentuale di
ricombinazione inferiore a 20 cM, e ordinati mediante il comando “order”. Il comando
“ripple” è stato impiegato per verificare la correttezza dei risultati ottenuti. I restanti marcatori
sono stati aggiunti alla mappa col comando “try” e LOD compreso fra 2.0 e 3.0. La funzione
di mappaggio di Kosambi (Kosambi 1944) è stata impiegata per convertire la percentuale di
ricombinazione in distanza di mappa (cM).
Nel complesso sono stati analizzati 1007 marcatori molecolari, di cui 677 sono stati
impiegati per lo sviluppo della mappa. Sono stati identificati 13 gruppi linkage (LGs) totali:
11 large LGs e 2 small LGs, che coprono 1245.7 cM del genoma totale. Si osserva una forte
variabilità nella densità di mappa, dovuta al fatto che i marcatori tendono a localizzarsi in
clusters. In ogni gruppo linkage il 44-66% dei marcatori è disposto in clusters; questa
distribuzione non random è stata osservata anche in pomodoro; analisi citogenetiche
(Tanksley et al. 1992) dimostrano che i clusters corrispondono alle regioni centromeriche,
aree dove la ricombinazione è soppressa. L’impiego di sonde RFLP di pomodoro ha permesso
di effettuare studi di sintenia tra i genomi delle due specie. Sebbene la maggior parte dei cloni
RFLP testati su peperone e pomodoro sia in grado di ibridare reciprocamente col DNA
genomico, l’ordine lineare dei cloni lungo i cromosomi non è conservato nelle due specie. Lo
studio evidenzia che, nel corso dell’evoluzione differenziale delle due specie, si sono
verificate perdite di geni e molteplici riarrangiamenti a livello dei cromosomi, come riportato
anche da Lefebvre et al. (2002). Tuttavia, i cromosomi di peperone mantengono alcuni
segmenti identici a quelli di pomodoro. Sono stati identificati 18 blocchi di associazione in
grado di coprire il 98.1% del genoma di pomodoro e il 95% del genoma di peperone.
Mediante studi di sintenia tra le due mappe è stato possibile identificare i tipi di
riarrangiamenti che si sono verificati durante l’evoluzione, ed è stato evidenziato che sono

I-INTRODUZIONE
51
necessarie almeno 32 rotture cromosomiche per convertire la mappa di pomodoro in quella di
peperone. Dallo studio, inoltre, emerge che loci multipli sono presenti sia in peperone, sia in
pomodoro, ma che l’ipotesi della duplicazione genica non permette di spiegare il contenuto di
DNA, 3-4 volte maggiore in peperone rispetto a pomodoro (Tanksley et al. 1988).
Livingstone e collaboratori ipotizzano che la differenza nel contenuto di DNA nucleare tra le
due specie, poiché non è legato ad un’incremento del numero dei geni codificanti, possa
essere dovuto alla dispersione di retrotrasposoni (Barakat et al. 1997), sia nelle regioni
cosiddette “gene rich” che non, e all’espansione dell’eterocromatina nel genoma di peperone
(Moscone et al. 1993).
L’impiego di una popolazione di mappaggio derivante da un incrocio inter-specifico ha
permesso di evidenziare un gran numero di polimorfismi, nonostante si siano verificati
riarrangiamenti tra i due genomi. E’ stato possibile definire in modo preciso le differenze
esistenti tra C. annuum e C. chinense, informazione molto importante in programmi di
breeding in quanto quest’ultima specie è una fonte di caratteri utili, di resistenza a patogeni.

I-INTRODUZIONE
52
Fig. 44 Mappa di Livingstone et al. 1999

I-INTRODUZIONE
53
Fig. 44 Mappa di Livingstone et al. 1999 (continua)
Fig. 44 Mappa di Livingstone et al. 1999 (continua)

I-INTRODUZIONE
54
3.1.2 Kang et al. (2001): “An interspecific (Capsicum annuum x C. chinense) F2
linkage map in pepper using RFLP and AFLP markers.” Theoretical and Applied
Genetics
Kang et al. (2001) hanno sviluppato una mappa inter-specifica di Capsicum, utilizzando
una popolazione di mappaggio F2 (n= 107), che è stata ottenuta autofecondando gli ibridi F1
prodotti dall’incrocio C. annuum cv “TF68” x C. chinense cv “Habanero” (Fig. 45). “TF68” è
una cultivar originaria del Sud Asia, caratterizzata da frutti rossi, di forma allungata e non
piccanti. “Habanero” è originaria del Sud America ed è caratterizzata da frutti color arancio,
di forma “a campanula”, piccanti e dall’aroma forte.
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER,
Version 3.0/EXP (Lander et al. 1987); per identificare i gruppi linkage dai dati ottenuti con i
marcatori RFLP è stato usato il comando “group” con LOD score maggiore a 3.0 e
percentuale di ricombinazione massima di 25 cM. L’ordine dei marcatori è stato stabilito col
comando “compare” e confermato col comando “ripple”. I marcatori AFLP sono stati
addizionati mediante il comando “assign” e “build”; solo i marcatori con valore di LOD
superiore a 3.0 sono stati inseriti nella mappa. Gli altri marcatori sono stati assegnati mediante
il comando “place”. Le percentuali di ricombinazione sono state convertite in distanza di
mappa (cM) utilizzando la funzione di mappaggio di Kosambi (Kosambi 1944).
Sono stati impiegati nelle analisi linkage 150 marcatori RFLPs e 430 AFLPs. La mappa
finale è costituita da 16 gruppi linkage: 11 large LGs e 5 small LGs, che coprono 1320 cM,
con una distanza di mappa media tra marcatori di 7.5 cM. L’80 % dei marcatori RFLP sono
cloni derivati da librerie di cDNA di peperone e risultano uniformemente distribuiti
all’interno del genoma. La maggior parte dei marcatori AFLP è localizzata in clusters in ogni
gruppo linkage, tale distribuzione è stata rilevata anche in altri studi, e pare corrispondere alle
regioni centromeriche.
Nel lavoro sono stati anche mappati geni coinvolti nella biosintesi di carotenoidi e di
capsacinoidi che permettono di sviluppare ulteriori studi relativi ai metaboliti secondari di
peperone.

I-INTRODUZIONE
55
Fig. 45 Mappa di Kang et al. 2001

I-INTRODUZIONE
56
Fig. 45 Mappa di Kang et al. 2001 (continua)

I-INTRODUZIONE
57
3.1.3 Lee et al. (2004): “Characterization and molecular genetic mapping of
microsatellite loci in pepper.” Theoretical and Applied Genetics
Gli autori hanno inserito nella mappa SNU-RFLP di Kang et al. (2001a)
precedentemente descritta, 46 marcatori microsatelliti. La mappa è stata sviluppata
dall’analisi di una popolazione F2 (n= 107), originata dall’incrocio inter-specifico tra C.
annuum “TF68” e C. chinense “Habanero”. La mappa SNU-2 di Lee et al. (2004) è costituita
da 333 marcatori totali: 46 SSR e 287 RFLPs, raggruppati in 15 gruppi linkage; copre 1761.5
cM del genoma totale, con distanza di mappa media pari a 5.3 cM (Fig. 46).
I marcatori SSR sono stati isolati in parte da due librerie genomiche di peperone,
caratterizzate da inserti di piccole dimensioni (500 bp) e in parte dal database GenBank. E’
stato effettuato uno screening dei 20,000 cloni delle librerie con sonde caratterizzate da
motivi microsatellite del tipo (AT)n, (GA)n, (GT)n, (ATT)n e (TTG)n. I cloni positivi sono stati
sequenziati per validare la presenza del motivo microsatellite; le sequenze ottenute sono state
inserite nel database EMBL per correlarle ad eventuali funzioni biologiche: il 14% dei cloni
mostra omologia significativa con retrotrasposoni, trasposoni e short interpersed repetitive
elements (SINE). I motivi microsatelliti ottenuti dal database GenBank e quelli isolati dalle
librerie sono stati analizzati per valutare la possibilità di disegnare primers sulle sequenze
fiancheggianti. Dei 76 marcatori SSR totali sviluppati, 46 sono stati mappati con successo: 32
loci (29 primers) su 40 marcatori sviluppati dalle librerie di peperone, e 14 loci (13 primers)
su 36 marcatori sviluppati da database GenBank hanno evidenziato polimorfismo nelle linee
parentali.
La mappa SNU-2 è costituita dai 46 SSR isolati e da 287 marcatori RFLPs, di cui 146
nuovi loci e 141 loci già presenti nella mappa SNU-RFLP. I cloni RFLP impiegati sono sonde
di peperone, di pomodoro e di tabacco e permettono l’impiego della mappa SNU-2 come
mappa di riferimento per studi di sintenia. Sono stati mappati anche marcatori associati a geni
coinvolti nella resistenza a patogeni: CaPR, CaLTP e N32, e ai geni che conferiscono
piccantezza.
Le analisi linkage sono state condotte col software MAPMAKER 3.0 / EXP (Lander et al.
1987); i LGs sono stati identificati col comando “group” con LOD score minimo minore di
4.0 e percentuale di ricombinazione massima di 20 cM. L’ordine dei marcatori in ciascun
gruppo linkage è stato stabilito col comando “order” e i marcatori restanti sono stati aggiunti
col comando “try”. L’ordine finale è stato confermato col comando “ripple”. Le percentuali di

I-INTRODUZIONE
58
ricombinazione sono state convertite in distanza di mappa in cM utilizzando la funzione di
mappaggio di Kosambi (Kosambi 1944).
Fig. 46 Mappa SNU-2 di Lee et al. 2004.

I-INTRODUZIONE
59
Fig. 46 Mappa SNU-2 di Lee et al. 2004 (continua).

I-INTRODUZIONE
60
Fig. 46 Mappa SNU-2 di Lee et al. 2004 (continua).

I-INTRODUZIONE
61
3.1.4 Sasvári et al. (2004): “Construction of a new interspecific genetic map in
pepper based on AFLP and microsatellite markers.” A.B. Center, Gödöllõ, Ungheria
Sasvári et al. hanno sviluppato una mappa molecolare inter-specifica, utilizzando una
popolazione F2 ottenuta dall’incrocio tra C. frutescens cv Tabasco e la linea inbred P4 di C.
annuum, selezionata dalla varietà piccante ungherese “Kalocsai 622”.
Sono stati impiegati marcatori AFLP, SSR e marcatori associati a geni di caratteri
fenotipici quali: piccantezza, resistenza al patotipo L2 di TMV, colore del frutto, della corolla
e del pistillo.
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER,
(Lander et al. 1987). I marcatori molecolari sono stati assemblati in gruppi linkage utilizzando
il comando “group”, con LOD score maggiore a 3.0 e percentuale di ricombinazione pari a
0.25. I marcatori AFLP EcoRI/MseI hanno rilevato un forte livello di distorsione e tendenza a
formare clusters nelle regioni centrali e terminali dei cromosomi; per questo è stata
inizialmente sviluppata una mappa di base, inserendo gli altri marcatori molecolari analizzati.
Col comando “order” è stato identificato l’ordine più probabile sui gruppi linkage; in un
secondo tempo, sono stati aggiunti alla mappa i marcatori AFLP EcoRI/MseI, mediante il
comando “assign”. La mappa genetica finale copre 1280 cM del genoma di peperone, è
costituita da 278 marcatori assemblati in 13 gruppi linkage: 12 large LGs e 1 small LGs,
costituito da soli 4 marcatori AFLP. La distanza di mappa media è pari a 14 cM/ marcatore.

I-INTRODUZIONE
62
3.2 Mappe genetico molecolari intra-specifiche.
Le mappe intra-specifiche derivano dall’incrocio tra accessioni appartenenti a C.
annuum.
Gli incroci intra-specifici sono vantaggiosi nel mappaggio perché permettono di evitare
i problemi di bassa fertilità e segregazioni distorte osservati nel caso degli incroci inter-
specifici. Inoltre, la disponibilità di mappe intra-specifiche sature, cioè caratterizzate da un
elevato numero di marcatori molecolari, è molto utile nell’ambito del miglioramento genetico
della specie basate sulla selezione assistita da marcatori (MAS) e che sfruttano essenzialmente
la variabilità esistente all’interno della specie C. annuum. Tuttavia, poiché le progenie
ottenute da incroci intra-specifici sono caratterizzate da bassi livelli di polimorfismo, per lo
sviluppo delle mappe è necessario impiegare popolazioni segreganti costituite da un elevato
numero di individui e mappare un’alta quantità di marcatori molecolari.
Nei paragrafi sono illustrate in dettaglio le mappe intra-specifiche più significative.
Fig. 47 Pianta selvatica di peperone.

I-INTRODUZIONE
63
3.2.1 Ben Chaim et al. (2001a): “QTL mapping of fruit related traits in pepper
(Capsicum annuum).”
Ben Chaim et al. hanno condotto un’analisi di QTLs relativa alle caratteristiche dei
frutti su una popolazione F3 ottenuta dall’incrocio di due genotipi di Capsicum annuum:
“Maor” x “Perennial”. Sono stati utilizzati marcatori RFLPs, AFLPs, RAPDs insieme a
marcatori morfologici, per sviluppare una mappa molecolare comparativa pomodoro-
peperone (Fig. 48). E’ stata valutata anche la segregazione di caratteri quantitativi in 180
famiglie F3.
La mappa è stata sviluppata con l’utilizzo del software MAPMAKER, Version 3.0/EXP
(Lander et al. 1987); i marcatori sono stati assemblati in gruppi linkage col comando “group”
a LOD pari a 4.0 e frequenza di ricombinazione massima di 30 cM. E’ stata sviluppata una
“core map” a LOD score di 3.0 e i restanti marcatori sono stati aggiunti a LOD score di 2.0.
Le distanze di mappa sono state calcolate con la funzione di mappaggio di Kosambi (Kosambi
1944).
Sono stati identificati 55 QTLs mediante “interval analysis” con LOD score minimo di
3.0. I QTLs relativi ai caratteri: diametro e peso dei frutti, spessore del pericarpo e diametro
del pedicello risultano localizzati in regioni cromosomiche simili e riflettono una correlazione
significativa tra questi caratteri.
Sul cromosoma 3 è stato identificato un QTL principale che risulta responsabile del
60% della variabilità fenotipica legata alla forma del frutto. Questo cromosoma contiene
anche altri QTLs, responsabili della maggior parte dei caratteri analizzati nella popolazione.
Alcuni marcatori presenti nei LGs 2, 3, 8 e 10 sono associati a QTLs per diversi caratteri,
sembrano essere, pertanto, loci con un ruolo importante nel controllo dei processi di sviluppo
in peperone. Alcuni QTLs sono correlati a loci di pomodoro che controllano caratteri
analoghi, ciò suggerisce che questi loci possano essere ortologhi nelle due specie.

I-INTRODUZIONE
64
Fig. 48 Mappa di Ben Chaim et al. 2001a

I-INTRODUZIONE
65
Fig. 48 Mappa di Ben Chaim et al. 2001a (continua).

I-INTRODUZIONE
66
3.2.3 Sugita et al. (2005): “Rapid Construction of a Linkage Map using High-
efficiency Genome Scanning / AFLP and RAPD, Based on a Intraspecific, Doubled-
haploid Population of Capsicum annuum.” Breeding Science
Sugita et al. hanno sviluppato una mappa intra-specifica di C. annuum, utilizzando una
popolazione di mappaggio DH. Sono stati impiegati principalmente marcatori AFLP,
applicando la tecnica hight efficient genome scanning (HEGS), insieme a marcatori RAPD,
SSRs, RFLPs, SCARs, CAPS e sono stati individuati tra essi, marcatori strettamente associati
al locus L3, coinvolto nella resistenza a pepper mild mottle virus (PPMMoV), e associati al
locus C, correlato al carattere della piccantezza.
La popolazione DH (n=176) è stata ottenuta mediante la coltura di antere di individui
F1, originati dall’incrocio tra “K9-11” (C. annuum L.) e “AC2258” (C. annuum L.). “K9-11”
è una varietà caratterizzata dalla presenza del locus L3, è suscettibile a “Phytophthora blight”,
non manifesta piccantezza e i suoi frutti immaturi hanno una colorazione verde scuro.
“AC2258” è resistente a “Phytophthora blight” mostra piccantezza e i suoi frutti immaturi
hanno una colorazione giallo pallido.
I primers SSR sono stati disegnati sul gene della fibrillina, rinominato “gene PAP”
(Plastis-lipid-Associated Protein). I prodotti ottenuti sono stati sequenziati per disegnare i
primers, che sono stati marcati con fluorocromi e testati in diverse accessioni e specie di
Capsicum.
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER,
Version 3.0 (Lander et al. 1987) utilizzando condizioni di LOD score minimo minore di 3.0 e
percentuale di ricombinazione pari a 25 cM. Sono stati testati 518 marcatori totali: 382 AFLP,
122 RAPD, 3 RFLP, 7 SCAR e 4 CAPS; di questi, 224 sono stati inseriti nella “core map”.
Sono stati identificati 16 gruppi linkage totali: 11 large LGs e 5 small LGs; la mappa copre
1043.1 cM del genoma totale, la distanza di mappa media è pari a 4.6 cM.
I marcatori AFLP e RAPD hanno la tendenza a raggrupparsi in clusters nei LGs, in
corrispondenza delle presunte regioni centromeriche, come riportato nelle mappe genetiche
precedentemente sviluppate (Livingstone et al. 1999; Kang et al. 2001).
I dati ottenti dall’inoculazione di PMMoV hanno permesso di mappare il locus L3 nella
regione terminale di LG6; mappe precedenti riportano tale locus nella porzione terminale del
cromosoma 11 di peperone (Livingstone et al. 1999; Lefebvre et al. 2002), quindi è stato
possibile assegnare LG6 al cromosoma 11. Inoltre, 3 marcatori AFLPs e un marcatore RAPD
risultano strettamente associati al locus L3 e sono disposti su entrambe i lati di tale locus;

I-INTRODUZIONE
67
possono quindi essere molto utili nella MAS, per selezionare i genotipi desiderati all’interno
di una popolazione segregante.
Il locus C, che conferisce piccantezza, è stato mappato nella zona terminale di LG3; la
mappa sviluppata precedentemente da Ben Chaim et al. (2001b) riporta tale locus nella
porzione centrale del cromosoma 2 di peperone, l’LG3 è stato assegnato quindi al cromosoma
2.
Il marcatore PAP-SSR è stato mappato a 0.6 cM dal locus C; negli individui che
manifestano piccantezza si osservano 25 ripetizioni del motivo TA, negli individui che non
presentano piccantezza, il microsatellite ha 23 ripetizioni. Le sequenze nucleotidiche dei
frammenti PAP-SSR sono stati analizzate in diverse accessioni appartenenti a tre specie di
Capsicum ed è emersa la presenza di molteplici varianti alleliche. Le varietà piccanti
presentano spesso caratteri d’interesse, come ad esempio resistenze a patogeni, questo
marcatore può essere dunque molto utile nell’ambito del breeding.
Gli studi futuri saranno volti a: i) convertire i marcatori legati al locus L3 in marcatori
SCAR; ii) a sviluppare marcatori legati alla resistenza nei confronti del “fuoco” indotto da
Phytophthora e iii) sviluppare marcatori legati alle caratteristiche dei frutti immaturi.

I-INTRODUZIONE
68
3.2.4 Ogundiwin et al. (2005): “Construction of 2 intraspecific linkage maps and
identification of resistance QTLs for Phytophthora capsici root-rot and foliar-blight
diseases of pepper (Capsicum annuum L.)” Genome
Ogundiwin et al. hanno sviluppato due mappe genetiche intra-specifiche di C. annuum
con lo scopo di identificare i QTLs che conferiscono resistenza a Phytophthora capsici (Fig.
49).
Sono state impiegate due popolazioni di mappaggio: RILs, e F2, e i seguenti marcatori
molecolari: AFLPs, RAPDs, SSRs, SCARs insieme a marcatori morfologici.
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER,
Version 2.0 for Macintosh (Lander et al. 1987). I marcatori molecolari sono stati raggruppati
in gruppi linkage utilizzando il comando “group”, con LOD score compreso tra 3.0 e 4.5, e
percentuale di ricombinazione pari a 0.35. E’ stata effettuata un’analisi “pair-wise
comparison”, tra coppie di marcatori per stabilire il loro ordine all’interno di ciascun gruppo.
L’analisi fornisce un sottoinsieme di marcatori, che sono stati ordinati con il comando
“compare” a LOD pari a 3.0. Gli altri marcatori appartenenti allo stesso gruppo sono stati
aggiunti al sottoinsieme ordinato, in un secondo tempo, mediante il comando “try” a LOD
pari a 3.0. La procedura è stata ripetuta per ogni LGs. I marcatori posizionati nei gruppi con
LOD ≥ 3.0 sono stati utilizzati per sviluppare la “core map”. I marcatori che non risultano
linked a questo valore di LOD, sono stati inseriti nella mappa a LOD≥ 2.0. La funzione di
mappaggio di Kosambi (Kosambi 1944) è stata impiegata per convertire la percentuale di
ricombinazione in distanza di mappa (cM).
Le due popolazioni segreganti sono state sottoposte ad inoculazione con ceppi differenti
di P. capsici per valutare la risposta all’infezione e studiare i geni coinvolti nella resistenza al
patogeno. P. capsici è responsabile di diverse malattie in peperone e i sintomi sono variabili
in funzione all’organo colpito. In questo studio, Ogundiwin et al. hanno analizzato isolati di
P. capsici che provocano il cosidetto “foliar blight”, malattia tipica di regioni caratterizzate da
forti piogge o di colture irrigate con aspersori; e ceppi responsabili del cosiddetto “root-rot”,
malattia prevalente nelle regioni semiaride, o in colture irrigate per gocciolamento o mediante
straripamento di canali.
I dati ottenuti sono stati impiegati per identificare i QTLs coinvolti nella resistenza ai
due tipi di sintomi. E’ stato utilizzato il software Windows QTL Cartographer version 2.0
(Basten et al. 1997; Wang et al. 2003) per identificarle e localizzarle sulla mappa genetica.

I-INTRODUZIONE
69
Le caratteristiche dei frutti: habitus eretto (up), forma (fs), frutti raggruppati all’apice
(fa), sono state valutate nei parentali (P1 e P2), nell’F1, nell’F2 e nel backcross (F1 x P2) al fine
di studiarne l’ereditarietà. Questi caratteri sono stati utilizzati come marcatori morfologici
nella costruzione della mappa e sono stati assegnati a gruppi linkage.
La prima mappa è stata ottenuta dall’analisi di una popolazione di 94 RILs, originata
per single seed decendence (SSD), dall’incrocio di “PSP-11”, accessione di peperone
suscettibile a P. capsici, x “PI201234”, resistente. Sono stati utilizzati 183 marcatori
molecolari: 123 AFLPs, 41 RAPDs, 15 SSRs, 1 SCAR e tre marcatori morfologici: up, fs e fa.
Di questi, 144 (91 AFLPs, 34 RAPDs, 15 SSRs, 1 SCAR, e i 3 marcatori morfologici)
mostrano un LOD score minimo ≥ 2.0 e sono stati impiegati per lo sviluppo della mappa. I
marcatori sono stati assemblati in 17 gruppi linkage, con distanza di mappa media pari a
13.83 cM; la mappa copre 1466.1 cM del genoma totale del peperone.
I test di resistenza al “foliar blight” e al “root-rot” hanno permesso l’identificazione di
diversi QTLs raggruppati nei Phyto groups, che corrispondono a 16 regioni cromosomiche
della mappa. Di questi, 7 conferiscono resistenza nei confronti di almeno 2 ceppi di P. capsici
e agiscono sia nei confronti del “foliar blight”, sia del “root-rot”; i restanti 9 sono legati alla
resistenza nei confronti di un ceppo specifico.
I test sono stati svolti sia in Taiwn, sia in California per confermare l’esatta
localizzazione dei QTLs; questo è stato possibile grazie all’uso di una popolazione RILs, che
è una popolazione perenne e permette agevoli scambi di materiale e informazioni tra
laboratori.
La seconda mappa è stata ottenuta dall’analisi di una popolazione di 94 individui F2,
originata dall’incrocio tra la varietà commerciale “JEP”, suscettibile a Phytophthora, con la
varietà resistente “Criollo de Morellos 334”. Sono stati testati 146 marcatori molecolari: 78
AFLPs, 51 RAPDs, 14 SSRs and 3 SCARs. Di questi, 113 (51 AFLPs, 45 RAPDs, 14 SSRs,
and 3 SCARs) sono stati inseriti nella mappa genetica, che è costituita da 16 gruppi linkage.
91 marcatori su 113 sono stati mappati con LOD ≥ 3; i rimanenti sono stati inseriti a valori di
LOD compresi fra 2.0 e 3.0. La mappa F2 copre 1089.2 cM del genoma totale di peperone,
con una distanza di mappa media di 14.52 cM.
Sulla popolazione F2 sono stati effettuati i test relativi alla resistenza al “root-rot”,
inoculando il ceppo M, per identificare i QTLs coinvolti. Sono stati caratterizzati 5 QTLs: due
sono localizzati sul LG JC1, gli altri tre sui LGs JC2, JC4 e Jc5, rispettivamente. I QTLs si
localizzano su regioni cromosomiche denominate Phyto, come nella mappa precedente.

I-INTRODUZIONE
70
Fig. 49 Mappa di Ogundiwin et al. (2005)
Nove gruppi linkage della mappa RIL e 10 della mappa F2 sono stati assegnati ai
cromosomi di peperone sulla base di marcatori comuni a quelli presenti in mappe già
pubblicate in precedenza.
Dal confronto dei QTLs identificati nelle due mappe emergono casi di sintenia: Phyto.U
della mappa F2 e Phyto.P della mappa RIL sembrano essere lo stesso QTL in quanto sono
localizzati entrambe su LGs assegnati al cromosoma 5 di peperone. Gli altri 4 QTLs della
mappa F2 non possono essere direttamente correlati a quelli della mappa RIL, in quanto il
numero di marcatori comuni alle due mappe in tali regioni è troppo basso.
Il fatto che siano stati impiegati ceppi diversi nelle due mappe può spiegare l’assenza di
alcuni QTLs ortologhi. Il maggior numero di QTLs osservato nella mappa RIL è
probabilmente correlato all’uso di un maggior numero di ceppi nei test di resistenza.
Sono state osservate interazioni epistatiche tra alcuni QTLs anche se non sempre si sono
rilevate statisticamente significative.

I-INTRODUZIONE
71
Fig. 49 Mappa di Ogundiwin et al. (2005) (continua).
I QTLs identificati in questo studio possono essere di grande aiuto nel breeding per
elaborare strategie MAS, marker assisted selection, al fine di ottenere varietà di peperone
resistenti a Phytophthora, cosa che non è possibile attuare con i metodi di incrocio
convenzionale attualmente impiegati.

I-INTRODUZIONE
72
3.3 Mappe genetiche integrate.
Le singole mappe genetiche finora sviluppate in Capsicum sono state costituite
utilizzando diverse categorie di marcatori molecolari, cosa che rende difficile l’estrapolazione
di informazioni di linkage da una popolazione all’altra. Sono state create a questo scopo le
cosiddette “mappe integrate” che, sulla base dei marcatori comuni a più mappe, identificano
una mappa consensus che contiene tutti i marcatori presenti nelle singole mappe. Le mappe
integrate sono molto utili ai breeders in quanto, in qualsiasi posizione della mappa, essi hanno
a disposizione un set di marcatori molecolari da impiegare per l’analisi del germoplasma.
Inoltre, possono essere valide core-map in nuovi incroci ed essere arricchite di nuovi
marcatori molecolari, con lo scopo di saturare la mappa genetica o clonare particolari geni.
Nei paragrafi seguenti sono brevemente descritte due studi condotti allo scopo di integrare
mappe sviluppate in peperone.

I-INTRODUZIONE
73
3.3.1 Lefebvre et al. (2002): “Towards the saturation of the pepper linkage map
by alignment of three intraspecific maps including known-function genes.” Genome
Lefebvre et al. hanno sviluppato una mappa consensus intra-specifica di peperone
(Capsicum annuum L.) (Fig. 50), a partire da tre mappe individuali (Tab. 4). Le popolazioni
di mappaggio utilizzate, due DH e una F2, mostrano una notevole segregazione sia per i
marcatori molecolari sia per i caratteri agronomici d’interesse.
Tabella 4: Popolazioni utilizzate nella mappa
Mappa Parentali Popolazione N° individui Marcatori morfologici
HV “H3” x “Vania”
DH 101 C, L, pvr2
PY
“Perennial” x “Yolo
Wonder”
DH 114 C, L, up
YC
“Yolo Wonder” x
“Criollo de Morelos
334”
F2 151 C, Pvr4
Per lo sviluppo delle mappe individuali, sono stati utilizzati marcatori RFLPs (cloni di
librerie di pomodoro e peperone), RAPDs, AFLPs, isoenzimi e marcatori associati a geni di
funzione nota, legati a caratteri di interesse agronomico.
Il software MAPMAKER, Version 3.0 (Lander et al. 1987) è stato utilizzato per la
costruzione delle tre mappe. I gruppi linkage sono stati determinati col comando “group” a
valori di LOD score di 5.0, per HV e YC, e 8.0 per PY, e con percentuale di ricombinazione
pari a 0.3. I comandi “suggest subset” e “order” sono stati utilizzati per determinare l’ordine
più probabile dei marcatori sui LGs. Il comando “ripple” è stato impiegato per confermare
tale ordine. Sono stati inseriti nella “core map” solo quei marcatori che mostravano valore di
LOD maggiore a 2.0 nell’analisi “ripple”.
In un secondo tempo è stata sviluppata una “alignement map” per ciascuna delle tre
mappe singole e sono stati addizionati alle “core map” i marcatori potenzialmente utili per
allineare le mappe individuali a quelle di altre Solanaceae. I marcatori impiegati
comprendono: tutti i marcatori RFLPs utilizzati nell’analisi, 100 marcatori di geni di funzione
nota e tutti i marcatori comuni ad almeno due popolazioni singole. I comandi “try” e

I-INTRODUZIONE
74
“compare” hanno permesso di addizionare i marcatori scelti e determinare l’ordine più
probabile sui LGs. Le percentuali di ricombinazione ottenute sono state convertite in distanza
di mappa mediante la funzione di mappaggio di Haldane (Haldane 1919).
I gruppi linkage delle tre mappe individuali sono stati allineati tra loro, sulla base dei
marcatori comuni; quando almeno 2 marcatori appartenenti ad uno stesso LG di una mappa
individuale mostrano corrispondenza ad un LG di un’altra, i due LGs sono considerati
colineari. Il confronto tra le tre mappe ha permesso di unire LGs che risultavano separati nelle
mappe singole e di identificare 12 LGs consensus, corrispondenti ai 12 cromosomi di
peperone.
Tutti i marcatori AFLP e RAPD comuni alle tre mappe individuali occupano una
posizione corrispondente sui LGs delle stesse e possono essere considerati locus-specifici di
Capsicum annuum. Come osservato nelle mappe precedentemente sviluppate, anche nella
mappa consensus i marcatori molecolari hanno una densità variabile lungo i LGs e tendono a
raggrupparsi in clusters. E’ stato identificato almeno un cluster in ogni LG; poichè ogni
cluster contiene marcatori associati alle regioni centromeriche di pomodoro, si presume che,
anche in peperone, esse corrispondano a tali regioni. I 12 LGs della mappa consensus
mostrano anche colinearità con la mappa di pomodoro.
La mappa consensus contiene 100 marcatori di geni noti e 5 loci di interesse
agronomico (loci che conferiscono resistenza: L, pvr2, pvr4; il locus C, che determina il
contenuto in capsaicina; e il locus up, che controlla l’habitus eretto dei frutti). Sono stati
mappati altri loci di resistenza, grazie a marcatori ad essi associati: Tsw, Me3, Bs3 e il locus y,
che determina la colorazione gialla dei frutti.
La mappa consensus fornisce quindi informazioni importanti per effettuare studi di
sintenia, identificare geni e pathways metabolici di interesse, sviluppare applicazioni MAS e
accrescere le conoscenze relative al genoma del peperone e delle Solanaceae.

I-INTRODUZIONE
75
Fig. 50 Mappa integrata di Lefebvre et al. 2002

I-INTRODUZIONE
76
Fig. 50 Mappa integrata di Lefebvre et al. 2002 (continua)

I-INTRODUZIONE
77
Fig. 50 Mappa integrata di Lefebvre et al. 2002 (continua)

I-INTRODUZIONE
78
Fig. 50 Mappa integrata di Lefebvre et al. 2002 (continua)

I-INTRODUZIONE
79
Fig. 50 Mappa integrata di Lefebvre et al. 2002 (continua)

I-INTRODUZIONE
80
3.3.2 Paran et al. (2004): “An integrated genetic linkage map of pepper
(Capsicum spp.)”
Paran et al. hanno sviluppato una mappa integrata di peperone (Fig. 51) a partire dai dati
relativi a 6 mappe ottenute da altrettante progenie (Tab. 5). Le mappe sono state integrate
utilizzando il software INT_MAP, di proprietà di Keygene (Peleman et al. 2000); il
programma permette di integrare le singole mappe genetiche in un’unica mappa consenso.
Sono stati utilizzati 320 marcatori, “anchor-marker” per l’integrazione: 265 sono comuni a
due mappe; 27 sono comuni a tre mappe, 25 a quattro mappe e 5 marcatori sono condivisi da
5 mappe.
La mappa integrata è costituita da un totale di 2262 marcatori molecolari: 1528 AFLPs,
440 RFLPs (sonde eterologhe di pomodoro), 288 RAPDs, 3 isoenzimi e 3 marcatori
morfologici. I marcatori sono distribuiti in 13 gruppi linkage: 11 corrispondono ai cromosomi
completi di peperone, gli altri due gruppi linkage corrispondono a porzioni diverse del
cromosoma restante. Non è stata identificata una corretta corrispondenza tra gruppi linkage
della mappa e cromosomi di peperone perché non è stata ancora raggiunta la completa
saturazione del genoma. La mappa ha una lunghezza complessiva di 1832 cM ed è maggiore
della lunghezza delle singole mappe individuali.
L’integrazione ha migliorato la copertura del genoma, come dimostrato dal numero di
gap di 10 cM presenti tra i marcatori adiacenti: si osservano da 21 a 50 gap nelle mappe
individuali e soltanto 15 nella mappa integrata. La densità media dei marcatori nella mappa
integrata è di 1 marcatore/0.8 cM ed è maggiore al valore massimo di 1 marcatore/2,1 cM,
assunto nelle mappe individuali. La distribuzione dei marcatori lungo i cromosomi non è
sempre omogenea e si osserva la presenza di clusters a livello delle regioni centromeriche,
fenomeno già osservato nelle mappe sviluppate in peperone e anche in altre piante coltivate
quali mais, pomodoro e orzo (Vuylsteke et al. 1999; Haanstra et al. 1999; Qi et al. 1998). La
mappa integrata offre una grande disponibilità di marcatori molecolari utili nel breeding
vegetale, nella MAS, nell’analisi del genoma di peperone e per le successive operazioni di
mappaggio in Capsicum.

I-INTRODUZIONE
81
Tab. 5 Popolazioni utilizzate per lo sviluppo della mappa integrata, (Paran et al. 2004).
Autori Parentali Popolazione Marcatori
totali
N°
Gruppi
linkage
Lunghezza
mappa
(cM)
Livingston
e et al. 1999
“NuMex
Rnaky”x “PI 159234” F2 672 14 1439
Dati non
pubblicati
“100/63” x “PI
152225” BC1 397 13 1559
Ben Chaim
et al. 2001a
“Maor” x
“Perennial” F2 405 13 1264
Lefebvre
et al. 2002 “H3” x “Vania” DH 490 13 1086
Lefebvre
et al. 2002
“Perennial” x
“Yolo Wonder” DH 616 13 1294
Lefebvre
et al. 2002
“Yolo Wonder”
x “CM334” F2 183 13 736
Mappa
integrata 2262 13 1832

I-INTRODUZIONE
82
Fig. 51 Mappa integrata di Paran et al. 2004

I-INTRODUZIONE
83
Fig. 51 Mappa integrata di Paran et al. 2004 (continua).

II-OBIETTIVI DELLA RICERCA
84
II-OBIETTIVI DELLA RICERCA
L’obiettivo del lavoro sperimentale è stato di contribuire allo sviluppo di una mappa
genetica intra-specifica ad alta risoluzione in Capsicum annuum, già in corso di sviluppo
presso il settore di genetica agraria del DI.Va. P.R.A.
A tale scopo, nell’ambito del lavoro di tesi, sono stati sviluppati marcatori molecolari
simple sequence repeats (SSR = microsatelliti) derivati da EST, mediante approccio
bioinformatico e sono stati individuati marcatori SSR, precedentemente sviluppati in
Capsicum, per poterli inserire nella mappa.
E’ stato inoltre sviluppato un marcatore sequence tagged site (STS) specifico per il gene
pvr4, che conferisce resistenza nei confronti dei potyvirus.
Infine è stata valutata l’efficacia del mappaggio selettivo, al fine di validare l’utilizzo di
questa strategia per lo sviluppo di mappe genetico-molecolari sature, riducendo i tempi e i
costi di tali analisi.

III-MATERIALI E METODI
85
III.MATERIALI E METODI
1. Materiale vegetale
Per l’analisi molecolare volta allo sviluppo della mappa genetica, sono state utilizzate
297 linee inbred ricombinanti (RILs) sviluppate all’INRA (Montflavet – Francia). La
popolazione segregante (RIL:F5) è stata ottenuta dall’incrocio intra-specifico “Yolo Wonder”
X “Criollo de Morellos”. L’ibrido F1 è stato sottoposto a quattro generazioni di
autofecondazione, mediante strategia single seed desendence (SSD). Ciò ha permesso di
fissare gli alleli segreganti in piccoli gruppi di associazione e di raggiungere il 94% di
omozigosi nella popolazione. I parentali utilizzati per l’incrocio sono caratterizzati da
differenze per caratteri produttivi di interesse agronomico quali dimensione e spessore della
bacca, piccantezza, altezza della pianta, utili nell’ambito del breeding. Inoltre, “Yolo
Wonder” è una varietà americana di “bell pepper” (Fig. 52) suscettibile a Phytophthora spp.,
mentre “Criollo de Morellos”, accessione messicana, è dotata di geni di resistenza nei
confronti di Phytophthora spp. e dei nematodi appartenenti alla famiglia Meloydoginae.
Fig. 52 Bacca di Yolo Wonder
Nell’ambito del lavoro relativo allo sviluppo di marcatori SSR derivati da EST,
(paragrafo 3.1.3) sono stati utilizzati 16 genotipi di Capsicum spp., suddivisi in due gruppi
(Tab. 6), allo scopo di verificare la capacità dei marcatori di amplificare prodotti in specie
diverse. Il primo gruppo è composto da 5 accessioni: ‘Tabasco’, appartenente alla specie C.

III-MATERIALI E METODI
86
frutescens, e 4 accessioni appartenenti alla specie C. annuum: ‘P4’, ‘GD1’, ‘Yolo Wonder’
(YW) e ‘Criollo de Morellos’ (CM334). Queste 5 accessioni sono state impiegate come
parentali per lo sviluppo di 3 popolazioni di mappaggio: una inter-specifica, ‘Tabasco’ x ‘P4’
e due intra-specifiche, ‘GD1’ x ‘CM334’ e ‘YW’ x ‘CM334’. Quest’ultima popolazione è
stata impiegata per lo sviluppo della mappa genetica oggetto del lavoro di tesi.
Il secondo gruppo è costituito da 11 accessioni di Capsicum spp., riassunte in tabella 6.
Tab. 6 Accessioni di Capsicum impiegate per l’analisi della variabilità genetica
dei marcatori SSR sviluppati.
Varietà/ Accessione Tassonomia
Gruppo 1 ‘Tabasco’ ‘P4’ ‘GD1’ ‘Yolo Wonder’ (YW) ‘Criollo de Morellos 334’ (CM334)
C. frutescens
C. annuum
C. annuum
C. annuum
C. annuum
Gruppo 2: Per MW1 Fru48 Ch1 Ch7 Pub3 Cha6 Exi1 Bac20 Pen6 Pra1
C. annuum
C. annuum
C. frutescens
C. chinense
C. chinense
C. pubescens
C. chacoense
C. eximium
C. baccatum var. baccatum
C. baccatum var. pendulum
C. praetermissum

III-MATERIALI E METODI
87
2. Estrazione del DNA
Il DNA genomico è stato estratto dal tessuto delle giovani foglie della popolazione
segregante e delle relative linee parentali, secondo il seguente protocollo:
- una quantità pari a circa 2 g di materiale vegetale fresco è stata frantumata in un
mortaio con azoto liquido, fino ad ottenere una polvere bianca, e trasferita in 800 µl di
tampone di lisi (2% CTAB, 0,1 M Tris-HCl pH 9,0, 1.4 M NaCl, 20 mM EDTA pH 8,0, 0,2%
ß-mercaptoetanolo);
- i campioni sono stati incubati per 90 min. a 65°C; miscelati con un volume di
cloroformio:alcol isoamilico (25:24:1 v:v:v) e centrifugati a 14 krpm per 10 min;
- in seguito al trasferimento del surnatante in una nuova provetta, i residui di fenolo
sono stati rimossi mediante l’aggiunta di un volume di cloroformio:alcol isoamilico (24:1 v:v)
ed il DNA presente nella fase acquosa è stato recuperato dopo centrifugazione a 14 krpm per
5 min;
- il DNA è stato precipitato con l’aggiunta di 1.5 volumi di isopropanolo freddo, posto a
–20°C per almeno 30-40 min. Dopo una centrifugazione di 20 min. a 14 krpm a 4°C, il pellet
precipitato è stato sottoposto ad uno o più lavaggi mediante 100 µl di etanolo 70%, lasciato
asciugare all’aria e risospeso in 500 µl di tampone TE (10 mM Tris- HCl pH 8.0, 1 mM
EDTA).
Il DNA estratto è stato quantificato mediante spettrofotometro (Hoefer DyNA Quant
200).

III-MATERIALI E METODI
88
3. Marcatori molecolari utilizzati nello sviluppo della mappa.
La mappa genetico molecolare sviluppata è stata denominata “mappa F5YC” in quanto
è stata ottenuta dall’analisi di una progenie RIL:F5, originata dall’incrocio tra “Yolo Wonder”
(Y) e “Criollo de Morellos” (C). Sono stati impiegati per l’analisi 595 marcatori molecolari;
tra questi: 507 AFLP, 20 SSAP (retrotrasposon based sequenze-specific amplified
polymorphisms), 18 RFLP, 12 STS e 38 SSR.
3.1 Marcatori microsatelliti (SSR)
Per lo sviluppo della mappa F5YC sono stati analizzati: 20 marcatori SSR sviluppati da
Sasvári et al. 2004, 7 sviluppati da Lee et al. 2004 (mappa SNU-2) e, infine, 11 dei 49
sviluppati nell’ambito del lavoro di tesi, mediante approccio bioinformatico.
3.1.1 Microsatelliti sviluppati da Sasvári et al. 2004
Sasvári et al. (2004) hanno sviluppato 75 marcatori SSR presso l’ABC (Agricultural
biotechnology centre, Godollo, Hungary) identificando tra le sequenze EST di Capsicum
presenti in database, quelle contenti motivi microsatelliti. I marcatori SSR sono stati testati
nelle diverse specie di Capsicum da Sasvári et al. (2004). Per lo sviluppo della mappa F5YC,
sono stati sottoposti a screening i marcatori SSR che, dallo studio precedente, risultavano
presenti in C. annuum per valutare quelli polimorfici nella popolazione RIL:F5.
3.1.2 Microsatelliti sviluppati da Lee et al. 2004
23 marcatori SSR descritti da Lee et al. 2004, localizzati sulla mappa SNU-2, sono stati
testati sulla popolazione RIL per valutare la possibilità di inserirli nella mappa F5YC. I
marcatori SSR sono illustrati in tabella 7.

III-MATERIALI E METODI
89
Tab. 7 Marcatori SSR sviluppati da Lee e sottoposti a screening sulla popolazione RIL.
LOCUS MOTIVO RIPETUTO PRIMER TAGLIA
Hpms1–1 (CA)12(TA)4 L:CAACCCAATATTAAGGTCACTTCC
R: CCAGGCGGGGATTGTAGATG 283
Hpms1-111 (AAT)11 L: AAGCTTATCCCTTTCAAATATAA
R: ATATCTCACGTATTGCGGATTCTT 159
Hpms1–172 (GA)15 L: GGGTTTGCATGATCTAAGCATTTT
R: CGCTGGAATGCATTGTCAAAGA 344
Hpms1–173 (GA)16(TG)2 L: TGCTGGGAAAGATCTCAAAAGG
R: ATCAAGGAAGCAAACCAATGC 163
Hpms1–214 (GTTT)2(TTG)9 L: TGCGAGTACCGAGTTCTTTCTAG
R: GGCAGTCCTGGGACAACTCG 100
Hpms1–216 (TTG)7...(TTG) 8 L: TGCTTGTTGTTTTTACCCTCAGC
R: AGTGAAAGGTGGGCAACAGC 108
Hpms1–5 (AT)11(GT)17 L: CCAAACGAACCGATGAACACTC
R: GACAATGTTGAAAAAGGTGGAAGAC 311
Hpms1–6 (AT)2 (GT)4(AT)8 (GT)13 L: TCCATAACTTCACCCATGAGTATGA
R: GCAACACCCACATTCCCTTCTC 197
Hpms1–62 (TG)23(AG)9 L: CATGAGGTCTCGCATGATTTCAC
R: GGAGAAGGACCATGTACTGCAGAG 186
Hpms2–13 (AC)12(AT)4 L: TCACCTCATAAGGGCTTATCAATC
R: TCCTTAACCTTACGAAACCTTGG 259
Hpms2–18 (TTG)11 L: CCTCCCCAGACCCTACTTTATGC
R: TCAACCAAACAAGTCGAAGTCAGC 162
Hpms2–24 (CT)17(CA)5A21 L: TCGTATTGGCTTGTGATTTACCG
R: TTGAATCGAATACCCGCAGGAG 205
AA840739 (TTTC)4 L: CCCTCTCTCTTCCCTTCACC
R: AGTGCTACGATGGCTTAGGG 95
AA842818 (TA)4(TC)3 L: TCCACTGCACCACAACCAATG
R: CCATAAGCAACCAACAGAATTA-GGG 202
AF130118 (AT)11imp L: ATGGATGCTTCGCCATCTCAAC
R: GACACAGCAGCCATATGTATACGC 176
AF208834 T7A11 L: TGCACCAAGGTCCAGTAAGGTTG
R: CCAACCACCATGGTTCATACAAG 201
CAN011109 (GT)5imp(CA)3 L: GGCCTTCAACTCCTCAAAGACG
R: TACTGCATCGGTTCCTGGATTG 195
CM0005 (CCA)8 L: CATGACCACCATGAGGATA
R: GATAGCCACGAGCATAGTATT 160
CM0007 (AG)6 L: TGTTCCTCTCTCTTCTTATCG
R: CCGGAGATAAGATCTTGATAA 103
CM0010 (AT)6 L: TTGGTTTTTGCTACTGGTAAT
R: AAACTGTCATATATTTGTGTGACT 158
CM0011 (AC)5(TA)8 L: TCTGCTTTAAAAACACATACAT
R: CATTCTAACTGAAATTGCATG 116
Hpms1–69 (AC)9(AT)4 L: CGGTGGCATGTAGTTTCTGGAG
R: AAGACATGAAATCCACAAGTTTTC 217
Hpms2–21 (AT)11(AC)9 L: TTTTTCAATTGATGCATGACCGATA R: CATGTCATTTTGTCATTGATTTGG
295

III-MATERIALI E METODI
90
3.1.3 Sviluppo di marcatori SSR, derivati da EST.
I marcatori SSR sono stati sviluppati mediante approccio bioinformatico, utilizzando
l’EMBL Nucleotide Sequence Database, http://www.ebi.ac.uk/embl (Fig. 53). In una
precedente ricerca, condotta nel 2003, il database conteneva 23174 entries appartenenti al
genere Capsicum e permise lo sviluppo di 50 marcatori SSR derivati da EST (Nagy et al.
2004).
Nel luglio scorso è stata effettuata nuovamente una ricerca sul database EMBL che
ha permesso di idenficare 8094 nuove sequenze appartenenti al genere Capsicum (più del
98% EST) e che coprono circa 4.1 Mb. Queste sequenze sono state analizzate al fine di
individuare al loro interno motivi microsatellite per sviluppare marcatori molecolari da
impiegare nel mappaggio.
Fig. 53 Interfaccia dell’EMBL Nucleotide Sequence Database.
In una prima fase del lavoro, sono state rimosse dalle 8094 sequenze, le accessioni
contenenti polyA e polyT, localizzate a circa 10 bp di distanza dalle estremità 5’ e 3’, in
quanto corrispondono alle code di polyA dell’mRNA eucariotico.
Successivamente, le sequenze contenenti motivi microsatellite sono state identificate
mediante l’impiego del software MISA (MicroSAtellite, Thiel et al. 2003), che permette
di analizzare rapidamente centinaia di migliaia di sequenze per identificare la presenza sia

III-MATERIALI E METODI
91
di microsatelliti perfetti sia di quelli composti. Il programma fornisce come output una
lista di sequenze contenenti SSR e specifica, per ciascuna, il numero di ripetizioni presenti
e la loro localizzazione.
In questo studio sono stati considerati motivi microsatellite quelli caratterizzati da
motivi ripetuti compresi tra 1 e 6 nucleotidi. Di conseguenza, è stato stabilito un valore di
lunghezza minima pari a: 10 unità ripetute per i mononucleotidi, 6 unità ripetute per i
dinucleotidi e 5 unità ripetute per le ripetizioni di ordine superiore. Per i microsatelliti
composti è stato impiegato, di default, un valore di massima interruzione tra le unità
ripetute pari a 100 bp.
Una volta identificate le sequenze contenenti i microsatelliti, è stata effettuata
un’analisi di clustering mediante l’impiego del software GDE (Genetic Data Environment,
version 2.2), al fine di eliminare le sequenze multiple e isolare sequenze contenenti SSRs
non-ridondanti. A tale scopo sono stati impiegati 4 criteri: i) nei clusters caratterizzati da 2
o più sequenze identiche, è stata scelta la sequenza più lunga; ii) quando i membri di uno
stesso cluster risultavano simili, ma con lunghezza variabile o facevano parte di sotto-
gruppi, è stato scelto un membro rappresentativo di ciascun sotto-gruppo; iii) i clusters
generati dall’appaiamento di sequenze ambiguamente sovrapposte non sono stati
considerati; iv) le sequenze contenenti SSRs, ma senza regioni fiancheggianti non sono
state considerate, in quanto non permettono lo sviluppo di marcatori molecolari.
Inoltre, le sequenze risultanti sono state allineate con le sequenze ESTs impiegate in
studi precedenti per lo sviluppo di marcatori SSRs (Nagy et al. 2004; Lee et al. 2004) per
rimuovere gli eventuali duplicati, al fine di ottenere un set finale di sequenze non-
ridondanti per la costruzione dei primers.
Le coppie di primers sono state disegnate, sulle sequenze fiancheggianti i motivi
microsatellite, mediante l’impiego del software PRIMER3 disponibile on-line
http://www.genome.wi.mit.edu/genome_software/other/primer3.html, sulla base dei
seguenti parametri:
- lunghezza dei primers: 18-25 bp;
- taglia del prodotto di amplificazione: 150-300 bp;
- contenuto minimo in GC: 40 %;
- CG clamp (n° di Gs e Cs consecutive all’estremità 3’ di ciascun primers): 2;
- Temperatura di melting ottimale: 60°C.

III-MATERIALI E METODI
92
Ciascun primers è stato denominato con la sigla EPMS (EST Pepper MicroSatellite)
seguita da un numero, a partire da EPMS-595. I primers, impiegati per l’amplificazione
PCR, sono stati sintetizzati da Metabion International AG (Martinsried, Germany).

III-MATERIALI E METODI
93
3.2 Sviluppo di un marcatore STS
Il marcatore STS, ASC002, è stato sviluppato mediante strategia dCAPs (vedi paragrafo
2.1.1). Il gene di peperone pvr4, che conferisce resistenza nei confronti dei potyvirus, mostra
polimorfismi di tipo SNPs nelle due varietà parentali: “Yolo Wonder” e “Criollo de
Morellos”. Le rispettive sequenze del gene sono state precedentemente sequenziate (Caranta
et al. 1999) e sono disponibili in database (Tab. 8).
Tab. 8 Sequenza del gene pvr4 nei parentali della popolazione RILs
Parentale
Sequenza del gene pvr4
Yolo
Wonder
GCCTTCTCATGCACAGAATAAAAAAATACGAAGAGAGAAGGTCCCTGTTGGATTCC
TCTGGAAAGCGCAAAGATCGGAATTGCTGAGCCATTGACTAACAAGGACACTGAAG
TAGTGGAGATGCATGACAAGATGAGGTTGACCAAAGTTTGAGGAAATATATAGTAA
AGGAGAGTGCCGTGAATGAAGGACCATTTCAGTCGGTCAAAAGCATTTTTTTCAGAT
CTATTTTTGCCAACATGTAGGCTTTCCTAAATTTGGTGTTTTGTAGGTGATGTAGGAG
TTCCTGCACAAGCATAGCATTATCAGATGCTCTATGATTTTCCAAAAACTTGATTGA
GACGGTTCGATAAGGTCCTTGAGAAAAGGTTTGATTCGGTTGACAATTGTCTTGGTG
ATAATCTTGTATGTAGTGTTACAGAGACAGATAGGACTATACTGGGTAATGGACTCG
GGATTAGATACCTTTGGAATAACCTACCCTGAACTCTGAACCCTGAGCCCTGAATCC
TGAATCTTGAACCCTGA
Criollo
de
Morellos
GCCTTCTCATGCACAGAATAAAAAAATACGAAGAGAGAAGGTCCCTGTTGGATTCC
TCTGGAAAGCGCAAAGAACGGAATTGCTGAGCCATTGACTAACAAGGACACTGAAG
TAGTGGAGATGCATGACAAGATGAGGTTGACCAAAGTTTGAGGGAATATATAGTAA
AGGAGAGTGCCGTGAATGAAGGACCATTTCAGTCGGTCAAAAGCATTTTTTTCAGAT
CTATTTTTGCCAACATGTAGGCTTTCCTAAATTTGGTGTTTTGTAGGTGATGTAAGAG
TTCCTGCACAAGCATAGCATTATCAGATGCTCTATGATTTTCCAAAAACTTGATTGA
GACGGTTCGATAAGGTCCTTGAGAAAAGGTTTGATTCGGTTGACAATTGTCTTGGTG
ATAATCTTGTATGTAGTGTTACAGAGACAGATAGGACTATACTGGGTAATGGACTCG
GGATTAGATACCTTTGGAATAACCTACCCTGAACTCTGAACCCTGAGCCCTGAATCC
TGAATCTTGAACCCTGA

III-MATERIALI E METODI
94
Il gene, nei due parentali, presenta diverse SNPs (evidenziate in rosso e giallo in tabella
8). E’ stata scelta la SNPs evidenziata in giallo e la sequenza che la contiene è stata inserita
nell’apposito box del software on line “dCAPS Finder 2.0” (Fig. 54).
Fig. 54 Interfaccia del software on line dCAPs Finder 2.0
Il software elabora diverse possibili combinazioni primers dCAPs - enzima di
restrizione, per ciascuna SNPs individuata. La scelta della combinazione per lo sviluppo del
marcatore STS è stata effettuata in base a due criteri: i) localizzazione della SNPs sul gene,
che condiziona la successiva costruzione del primer reverse e la possibilità di ottenere, in
seguito alla restrizione, un frammento di dimensioni idonee ad essere rilevato su gel, sia in
assenza sia in presenza del sito di taglio; ii) costo dell’enzima di restrizione necessario per
evidenziare i polimorfismi a carico del DNA.
Sulla base di questi requisiti è stato scelto l’enzima EcoRV, che riconosce il sito di
taglio: GATATC – TGGATTCCTCTGGAAAGCGCAGATA, in associazione al primer forward illustrato
in tabella 9 .
Il programma Bio-OLIGO (Premiersoft) è stato utilizzato per sviluppare il primer
reverse (Tab. 9). Il programma permette di disegnare i primers in posizioni tali da garantire
una corretta amplificazione.

III-MATERIALI E METODI
95
Sono stati impiegati i seguenti criteri:
- temperatura di melting: 60-65°C;
- lunghezza: 18-22 bp;
- stabilità al 5’ > stabilità al 3’(deltaG);
- assenza di hairpins, causate dal ripiegamento della sequenza su sé stessa;
- assenza di associazioni con il primer forward.
Tab. 9 Primers reverse e forward utilizzati
Parentale
Sequenza del primers (5’� 3’)
Yolo Wonder
Forward
TGGATTCCTCTGGAAAGCGCAAAGATCGGAATTGCTGAGCCATTGACTAAC
Reverse
GTTAGTCAATGGCTCAGCAATTCCGATCTTTGCGCTTTCCAGAGGAATCCA
Criollo de
Morellos
Forward
TGGATTCCTCTGGAAAGCGCAAAGAACGGAATTGCTGAGCCATTGACTAAC
Reverse
GTTAGTCAATGGCTCAGCAATTCCGTTCTTTGCGCTTTCCAGAGGAATCCA

III-MATERIALI E METODI
96
4. Analisi molecolare sulla popolazione segregante.
L’analisi molecolare sia dei marcatori SSR che del marcatore STS è stata effettuata in 5
fasi successive:
i) amplificazione PCR (4.1);
ii) elettroforesi su gel di agarosio (4.2);
iii) quantificazione dei prodotti ottenuti (4.3);
iv) elettroforesi su gel di poliacrilamide (4.5);
v) colorazione e visualizzazione dei risultati (4.6).
Nel caso dei marcatori di tipo SSR è stato effettuato uno screening iniziale per
selezionare i marcatori utili nello sviluppo della mappa; nel caso del marcatore STS, ASC002,
è stata effetuata un’analisi di tipo dCAPs. I due procedimenti sono descritti rispettivamente
nei paragrafi 5 e 6.
4.1 Condizioni di amplificazione.
Le reazioni di PCR sono state condotte in volumi finali di 20 µl contenenti 50 ng di
DNA stampo, 0.2 mmol/L di ciascun deossiribonucleotide (dATP, dCTP, dGTP, e dDTP), 0.5
µmol/L di ciascun primers, 1 U di Taq polimerasi (Fermentas), 25 mM di MgCl2 nel Buffer di
reazione NH4+, concentrato 10X.
Tutte le reazioni di amplificazione sono state effettuate per mezzo di un termociclatore
MyCycler ™ BIORAD programmato per il seguente profilo termico, con Touch-Down 60°C-
55°C:
5 min a 94°C (denaturazione iniziale)
30 s a 94°C (denaturazione)
da 60°C a 55°C (-0.5°C ogni ciclo) per 30 s (appaiamento)
60 s a 72°C (estensione)
30 s a 94°C (denaturazione)
30 s a 55°C (appaiamento)
60 s a 72°C (estensione)
7 min a 72°C (estensione finale)
11 cicli
21 cicli

III-MATERIALI E METODI
97
I prodotti di PCR sono stati conservati a 4°C fino al momento della separazione
elettroforetica su gel di agarosio al 2% per controllare l’avvenuta amplificazione.
4.2 Elettroforesi su gel di agarosio.
Un volume totale di 7 µl costituito da 5 µl di campione e 2 µl di tampone di corsa
(0.05% blu di bromofenolo, 40% w/v saccarosio, 0.1 M EDTA), è stato separato mediante
elettroforesi su gel di agarosio al 2%, con 0.35 µl bromuro di etidio, in tampone TBE 0.5X
(1X TBE: 90 mM Trisborato, 2 mM EDTA, pH 8.0) a 90V per circa 20 minuti.
E’ stato utilizzato un marcatore di peso molecolare noto (123 bp ladder) per identificare
e verificare le dimensioni delle bande; i pattern elettroforetici sono stati visualizzati per mezzo
di lampada UV (302 nm) e acquisiti mediante Gel Documentation System (Quantità One
Programme, BioRad).
4.3 Quantificazione e aggiunta del tampone di corsa
La quantificazione dei campioni nel pattern elettroforetico è stata effettuata mediante il
programma Gel Documentation System (Quantità One Programme, BioRad) che, sulla base
delle intesità reciproche rilevate agli UV, ha permesso di stabilire la quantità di loading buffer
(tampone di corsa: 98% fornamide, 10 mmol EDTA/L, 0.01% w/v blu di bromofenolo e
0.01% w/v di xylene cianolo) necessaria per diluire i campioni e renderli tra loro omogenei;
ciò è necessario per una colorazione (Silver Staining) uniforme del gel di acrilammide.
4.4 Elettroforesi duplex e triplex
Quando, a seguito della visualizzazione dell’amplificato su gel di agarosio, i prodotti di
PCR hanno prodotto bande con pesi molecolari sufficientemente distinti, l’elettroforesi su gel
di acrilammide è stata effettuata in “duplex” (due coppie di primers) o “triplex” (tre coppie di
primers) (Fig. 55 e 56), in quanto è stato possibile analizzare contemporaneamente gli
amplificati ottenuti con primers diversi. Ciò permette un notevole risparmio di tempo e costi,
riducendo il numero di corse elettroforetiche da effettuare per l’analisi dei polimorfismi.

III-MATERIALI E METODI
98
Fig. 55 Esempio di corsa elettroforetica duplex.
Fig. 56 Esempio di corsa elettroforetica triplex.
Hpms 1_5 311 bp
Hpms 2_24 205 bp
Hpms 1_214 100 bp
Hpms 1_117
Hpms 2_13

III-MATERIALI E METODI
99
4.5 Elettroforesi su gel di poliacrilamide
La corsa elettroforetica è stata condotta in un sistema verticale dotato di due lastre di
vetro di dimensioni di 38x50 cm e spaziatori, inseriti tra le due, dello spessore di 0.4 mm. Il
gel di poliacrilamide (PAGE), denaturante al 5%, è stato ottenuto miscelando 105 ml di urea 7
M con 15 ml Acrilamide 40%.
4.5.1 Preparazione delle lastre
Le due lastre, base plate e glass plate, sono state lavate accuratamente con detersivo e,
successivamente, con etanolo 100%. La base plate è stata trattata con 0.5 ml di soluzione
Repel silane in modo tale da distribuire in maniera uniforme il prodotto e lasciata asciugare 5
minuti. L’eccesso di Repel silane è stato rimosso con una salvietta inumidita con etanolo. La
glass plate è stata trattata con 1 ml di soluzione Bind silane (5 µl di bind silane in 1 ml di
soluzione 95% etanolo e 0.5% acido acetico) in modo analogo, rimuovendo l’eccesso con una
salvietta inumidita con etanolo.
Sono stati posizionati gli spaziatori, lavati con etanolo, sulla base plate. La glass plate è
stata posizionata al di sopra, cercando di evitare il contatto tra le due lastre. Sono stati inseriti
i clamps ai lati dell’apparato; il sandwich così ottenuto è stato inserito nel tray, chiudendo le
viti e disponendo il tutto su un piano orizzontale.
4.5.2 Preparazione della soluzione di acrilamide
Sono stati miscelati 105 ml di urea 7 M in TBE 1X, a 15 ml Acrilamide 40% in un
becker; successivamente, sono stati aggiunti 500 µl di APS 10% e 60 µl di TEMED, che
garantiscono la polimerizzazione del gel in 30-60 minuti; in tempi brevi, la soluzione è stata
caricata nella siringa, eliminando le eventuali bolle d’aria formatesi. Quindi, la soluzione è
stata iniettata nell’apparato, con pressione costante, in modo da evitare la formazione di bolle.
All’estremità alta del sistema è stato inserito il pettine a denti di squalo, girato al contrario
necessario per creare il fronte. Il gel è stato lasciato polimerizzare overnight.

III-MATERIALI E METODI
100
4.5.3 Corsa elettroforetica.
In un primo tempo è stata effettuata la “Pre-corsa”. Sono stati preparati 1 L di tampone
TBE 1X e 0.6 L di tampone TBE + 24 g di sodio acetato, e sono stati riscaldati per 4-8 minuti,
in un forno a microonde. Il pettine e i clamps sono stati tolti dall’apparato col gel e sono stati
ripuliti gli eccessi di acrilamide. Il tampone TBE + sodio acetato è stato versato nella base; il
tampone TBE è stato versato tra i due vetri fino a 1 cm sopra il livello del glass plate. Il fronte
del gel è stato accuratamente ripulito con una siringa, eliminando l’urea in eccesso e evitando
la formazione di bolle. Il gel è stato sottoposto a pre-corsa a 110 W fino a raggiungere la
temperatura ottimale di 47°C.
In contemporanea, i campioni diluiti con loading buffer sono stati denaturati per 5 min.
a 95°C e immediatamente raffreddati in ghiaccio per evitare l’appaiamento dei filamenti
complementari.
Al termine della “Pre-corsa” è stata ripulita la parte superiore del gel con una siringa e
sono state fatte cadere, sul fronte del gel, alcune gocce di Blu AFLP; quindi, il gel è stato fatto
correre per alcuni secondi (10-20 W) per rendere visibile la linea superiore creata dal blu. Il
fronte è stato nuovamente pulito con una siringa ed è stato inserito il pettine a denti di squalo
per creare i pozzetti di caricamento. Sono stati caricati 3,5 µl di ciascun campione e il ladder
100 bp. Il gel è stato infine sottoposto a corsa elettroforetica, a 10-15 W, per 5 min. e
proseguita a 110 W per circa 2,5 ore, fino a quando la linea del blu di bromofenolo ha
raggiunto il fondo.
Al termine della corsa i gel sono stati fissati in una soluzione di acido acetico al 10%
(v/v) per 30 min e sono stati dunque colorati.

III-MATERIALI E METODI
101
4.6 Visualizzazione dei prodotti di amplificazione
Per visualizzare le bande ottenute dagli amplificati, i gel sono stati colorati con la
tecnica silver staining (Tab. 10), secondo il protocollo descritto da Bassam et al. (1991). La
colorazione sfrutta la precipitazione degli ioni Ag++, precedentemente legati ai frammenti di
DNA, a seguito della riduzione in ambiente alcalino.
Tab. 10 Soluzioni utilizzate per il Silver Staining
Soluzioni SILVER STAINING
- FIX / STOP SOLUTION: 200 ml acido acetico
1800 ml acqua ultrapura
- STAINING SOLUTION: 2 g nitrato di argento
3 ml formaldeide 37%
2000 ml acqua ultrapura
- DEVELOPING SOLUTION: 60 g carbonato di sodio
2000 ml acqua ultrapura
Raffreddato a 10°C in freezer
Appena prima dell’uso aggiungere: 3 ml formaldeide 37%
400 µl tiosolfato
Il procedimento utilizzato per la colorazione è il seguente: è stata preparata, in anticipo,
la Developing solution (60 g carbonato di sodio e 2000 ml di acqua ultrapura) ed è stata
raffreddata a 10° in freezer. L’apparato di corsa, smontato, è stato raffreddato con acqua e le
due lastre sono state separate in modo tale che il gel rimanga attaccato alla glass plate. Il vetro
è stato posto in un vassoio contenente la Fix/Stop solution (200 ml acido acetico + 1800 ml di
acqua ultrapura) e lasciato in agitazione per 30 minuti, fino alla scomparsa della linea di
xylene cianolo. La soluzione è stata poi recuperata.
Successivamente, la lastra è stata sottoposta a due risciacqui da 5 min. in 1 L di acqua
ultrapura ciascuno, mantenendo il vassoio in agitazione. La lastra è stata poi trasferita per 30
min., in un vassoio contenente la Staining solution (2 g nitrato di argento + 3 ml Formaldeide
37% + 2000 ml acqua ultrapura) in agitazione, per indurre la colorazione.
Nel frattempo è stata completata la Developing solution, aggiungendo 3 ml di
formaldeide 37% e 400 µl di tiosolfato di sodio. La soluzione è stata poi suddivisa 1L+1L in

III-MATERIALI E METODI
102
due vassoi posizionati sotto cappa. Il gel è stato appoggiato su una superficie piana ed
immerso per non più di 4 secondi in un vassoio con 2 L di acqua ultrapura, per un rapido
risciacquo, e immediatamente posto nel vassoio con la Developing solution.
Per indurre lo sviluppo, il vassoio è stato agitato in modo che l’intera superficie del gel
venga a contatto con la soluzione, fino a quando incominciano ad apparire le prime bande,
relative al ladder. A questo punto, il gel è stato trasferito nel secondo vassoio, proseguendo
l’agitazione fino alla completa comparsa delle bande.
La reazione di sviluppo è stata bloccata poco prima dell’imbrunimento del gel,
aggiungendo 1L della soluzione Fix/Stop recuperata, e agitando finchè la soluzione continua a
“friggere”. Infine, la lastra è stata posta in 2 L di acqua ultrapura per 2 min per un risciacquo
finale e lasciata asciugare sottocappa overnight, in posizione verticale.
Le immagini sono state acquisite per mezzo di uno scanner ed i pattern elettroforetici
rilevati mediante Gel Documentation System (Quantity One Programme, BioRad); la lastra è
stata posta in una soluzione di 2 L NaOH 5% (preparata con acqua di rubinetto) per almeno 2
ore. Il vetro è stato lavato con sapone e risciacquato con acqua distillata.

III-MATERIALI E METODI
103
5. Screening dei marcatori microsatelliti e successiva analisi.
5.1 Screening dei marcatori SSR sviluppati da Sasvári et al. (2004) e
da Lee et al. (2004)
I marcatori SSR, sviluppati da Sasvári et al. (2004) e da Lee et al. (2004), sono stati
sottoposti a screening per identificare quali evidenziassero polimorfismo nella popolazione
RILs segregante e potessero essere pertanto utili nello sviluppo della mappa genetica.
12 diverse combinazioni di primers sono state impiegate per effettuare amplificazione
PCR su 8 individui, di cui 5 RILs, i due parentali e l’ibrido F1. I prodotti sono stati esaminati
su agarosio al fine di verificare l’avvenuta amplificazione e, successivamente, è stata
effettuata l’elettroforesi su gel di acrilamide.
5.2 Screening dei marcatori SSR, derivati da EST
Il set finale di marcatori SSR derivati da EST (EST-SSR) è stato sottoposto a screening
per valutare il livello di polimorfismi rilevabile nelle diverse accessioni di C. annuum e C.
frutescens (gruppo 1), descritte al paragrafo 1 (Tab. 6).
I marcatori rivelatisi polimorfici in questo primo gruppo sono stati impiegati per testare
il secondo, costituito da 11 accessioni di Capsicum spp. (Tab. 6, paragrafo 1), ed è stato
valutato il polymorphic information content (PIC) di ciascun marcatore.
Nel progetto di tesi sono stati impiegati i marcatori SSR rivelatisi polimorfici
nell’ambito di C. annuum.

III-MATERIALI E METODI
104
6. Analisi dCAPs del marcatore STS.
I prodotti ottenuti dall’amplificazione col marcatore STS ASC002, associato al gene di
resistenza pvr4, sono stati sottoposti all’analisi dCAPs. I campioni sono stati digeriti con
l’enzima Eco RV: un volume totale di 20 µl, con 2-5 unità di enzima di restrizione (7.87 µl di
acqua + 2 µl di Buffer + 0.3 µl di enzima Eco RV) è stato addizionato a 10 µl di prodotto di
PCR; la reazione di restrizione è stata mantenuta per 2 ore a temperatura ambiente. I campioni
sono stati sottoposti a elettroforesi su gel di agarosio e quantificati per effettuare l’adeguata
diluizione con loading buffer. Infine, sono stati sottoposti a elettroforesi su gel di acrilamide e
colorati (Fig. 57).
Fig. 57 Elettroforesi su acrilamide del marcatore ASC002, legato al gene pvr4.
7. Analisi dei dati.
ASC002 (pvr4)

III-MATERIALI E METODI
105
7. Analisi dei dati
7.1 Valutazione del polymorphic information content (PIC) dei
marcatori SSR, derivati da EST.
Il valore PIC (polymorphic information content) indica la capacità del marcatore nel
rilevare polimorfismi, sulla base del numero di alleli e della loro frequenza di distribuzione
all’interno della popolazione (Bostein et al. 1980). Il PIC è stato stimato, per ciascun
marcatore EST-SSR, mediante l’impiego dell’equazione di Anderson et al. (1993):
dove: Pij è la frequenza del i-esimo allele per il marcatore i, e ∑ Pij la sommatoria estesa per
n alleli, calcolata per ciascun locus SSR.
Ogni prodotto di PCR, ottenuto dall’analisi di screening effettuata con i marcatori EST-
SSR, è stato considerato un singolo locus; il valore ‘1’ è stato attribuito alla presenza del
frammento amplificato, mentre ‘0’ alla sua assenza. La matrice binaria, così generata, è stata
impiegata per calcolare i coefficienti di similarità genetica (GS), mediante analisi pair-wise
(Nei e Li, 1979); le matrici di similarità ottenute hanno permesso di valutare le relazioni
genetiche esistenti tra le accessioni testate, grazie alla costruzione di un dendogramma col
metodo UPGMA (Sneath e Sokal, 1973). Le analisi sono state condotte mediante l’impiego
del software NTSYS-PC versione 2.0 (Rohlf, 2002).

III-MATERIALI E METODI
106
7.2 Test del χχχχ2
Sui dati ottenuti da ciascun marcatore è stato applicato il test del χ2 allo scopo di
evidenziare eventuali distorsioni rispetto al rapporto atteso 1:1. Il test è stato inoltre utilizzato
per verificare che il livello di eterozigosi osservata non si discostasse da quella attesa, che per
una progenie RIL:F5 è di 6,25 %.
L’eterozigosi attesa (HE) è stata calcolata con la formula: HE = 1 - Σ pi2 (Nei 1973),
dove pi rappresenta la frequenza dell’allele i nel campione. L’eterozigosi osservata (HO) è
stata valutata direttamente dall’analisi dei dati del campione.
7.3 Analisi di linkage
Le analisi di linkage sono state condotte mediante l’impiego del software MAPMAKER
EXP 3.0 utilizzando il modello RI, specifico per la popolazione segregante analizzata (Lander
et al. 1987). I dati ottenuti dalla lettura dei gels, relativi a ciascun marcatore, sono stati inseriti
nel file input del software.
In sequenza, sono stati utilizzati i seguenti comandi: i) “group” per identificare i gruppi
di associazione; ii) “order” per determinare l’ordine dei marcatori nei singoli gruppi di
associazione; iii) “ripple” per confermarlo e iv) “try” per attribuire la posizione più probabile
dei marcatori non assegnati precedentemente.
Il comando “group” permette di assemblare i marcatori molecolari in gruppi linkage
sulla base di un’analisi a due punti. Al comando è necessario associare dei parametri soglia,
relativi al LOD score e alla percentuale di ricombinazione, per stabilire le relazioni di linkage
tra i marcatori. In questo studio sono stati utilizzati un LOD score minimo pari a 8.0 e una
percentuale di ricombinazione massima pari a 0.1. Sono state considerate linked tutte le
coppie di loci che hanno una distanza reciproca, minore o uguale a 10 cM e LOD score
maggiore o uguale a 8.0. Il software, nel determinare i gruppi linkage, applica la proprietà
transitiva per cui: se il locus A è associato al locus B, e il locus B è linked al locus C, allora
anche il locus A e il locus C sono associati e verranno quindi assegnati allo stesso LG. La
figura 58 illustra un esempio di report sviluppato dal software, in seguito all’uso del comando
“group”.

III-MATERIALI E METODI
107
Fig. 58 Risultati ottenuti mediante l'uso del comando “group”.
Il comando “order” permette di stabilire l’ordine dei marcatori all’interno del gruppo
linkage. Il comando deve essere associato ai seguenti parametri: LOD score (i.e.: 8.0),
frequenza di ricombinazione (i.e.: 0.1) e numero di marcatori base (i.e: 4). Il software
individua il numero di marcatori stabiliti, che sono i più informativi, e ne stabilisce l’ordine
sul LG; a questa disposizione di base il software aggiunge tutti i marcatori restanti,
appartenenti al LG.
Il comando “ripple” valuta la correttezza dell’ordine dei marcatori precedentemente
stabilito; il comando deve essere associato a due parametri: LOD score (i.e.: 3.0) e numero di
marcatori da considerare (i.e.: 5). Il software effettua permutazioni tra 5 marcatori limitrofi e
tiene conto del LOD score che essi assumuno nelle diverse permutazioni per stabilire le
posizioni più probabili. I dati ottenuti vengono presentati come illustrato nell’esempio della
figura 59.
Fig. 59 Risultati ottenuti mediante l'uso del comando “ripple”.

III-MATERIALI E METODI
108
Il comando “try”permette di inserire un marcatore non mappato su un LG col comando
“order”, o un marcatore aggiuntivo che risulta, da studi precedenti, linked agli altri marcatori
del LG. Il software calcola i valori di LOD assunti dal marcatore in questione in ciascuna
posizione del LG. Le posizioni in cui il marcatore assume LOD pari a 0.00 o poco minori a
questo valore, sono quelle più probabili (Fig. 60).
Fig. 60 Risultati ottenuti mediante l'uso del comando “try”
Le percentuali di ricombinazione sono state convertite in distanze di mappa (cM)
mediante la funzione di mappaggio di Haldane (Haldane, 1919).

III-MATERIALI E METODI
109
7.4 Assegnazione dei gruppi linkage ai cromosomi di C. annuum
I gruppi linkage sono stati assegnati ai cromosomi di C. annuum mediante allineamento
con “anchor marker”, comuni a due mappe precedentemente sviluppate: la mappa F2YC di
Lefebvre (Lefebvre et al. 2002) e la mappa integrata di Paran (Paran et al. 2004), descritte
rispettivamente nei paragrafi 3.3.1 e 3.3.2.
Nel caso in cui due gruppi linkage siano risultati assegnati allo stesso cromosoma, è
stato impiegato il comando “compare” per unirli in un unico gruppo e per stabilire il loro
ordine reciproco più probabile. I nuovi gruppi linkage ottenuti sono stati valutati con il
comando “ripple” per verificarne la correttezza, utilizzando un valore di LOD score pari a 3.0.
7.5 Scelta degli individui più informativi.
Allo scopo di identificare nell’ambito dei 297 individui RIL, quelli più informativi e
utili per un futuro mappaggio di geni di interesse, è stato utilizzato il software MapPop
(http://www.bio.unc.edu/faculty/vision/lab/mappop/). I dati inseriti nel software sono quelli
relativi alla mappa genetica F5YC ottenuta dall’analisi di tutte le 297 RILs. Dalla popolazione
di partenza sono stati eliminati 26 genotipi in quanto risultavano essere particolarmente
difficili da mantenere nel tempo a causa di una ridotta produzione di semi, dovuta ad una forte
manifestazione di depressione da inbreeding.
Il file di input richiesto dal software è il Framework Map File, che riporta i dati relativi
alla “core map” F5 sviluppata. La struttura del file input è quella illustrata in figura 61; sono
presenti tre colonne principali: la prima riporta il nome di ciascun marcatore della “core
map”; la seconda identifica il gruppo linkage di appartenenza; la terza indica la posizione di
ciascun marcatore, in distanza di mappa, rispetto all’estremità del LG; segue una colonna per
ogni individuo analizzato, (da 1 a n), al quale viene associato il genotipo.

III-MATERIALI E METODI
110
Fig. 61 Esempio di un Framework Map File di Map.Pop
Il comando “SAMPLEEXP” di MapPop ha permesso di identificare i genotipi più
rappresentativi. E’ possibile selezionare, ad esempio, individui caratterizzati da un’elevata
frequenza di crossing-overs o con una distanza reciproca, nei blocchi di associazione, meno
variabile rispetto a quella presente nel campione random di partenza.
La sintassi per il comando “SAMPLEEXP” è la seguente:
SAMPLEEXP <size> <time> <choices>
Il parametro <size> corrisponde alla dimensione del campione che si vuole selezionare;
in genere rappresenta una frazione della dimensione della popolazione finale.
Il parametro <time> indica il numero di secondi impiegati dal software, per identificare il
campione; di default vengono selezionati 300 secondi.
Il parametro <choices> indica il numero di campioni alternativi che il software deve
identificare, di default vengono elaborate 5 possibili scelte.
La prima sottopopolazione ottenuta è costituita da 141 individui e corrisponde a circa
1/2 dell’originale popolazione F5YC; la seconda è costituita da 93 individui, corrispondenti a
1/3 dell’originale popolazione F5YC; la terza è costituita da 29 individui ed è stata sviluppata
con l’obiettivo di definire il numero minimo di individui necessari per assegnare
correttamente nuovi marcatori ai gruppi linkage in futuri studi di mappaggio.

IV-RISULTATI
111
IV. RISULTATI
1. Sviluppo di marcatori SSR, derivati da EST.
Il database EMBL ha permesso di identificare 1.325 motivi microsatellite in 899
sequenze delle 8.094 analizzate; 298 sequenze (33%) contengono più di un motivo
microsatellite e, di questi, 164 sono SSR composti (Tab. 11). L’11% (899/8094) delle
sequenze analizzate contiene almeno un motivo SSR e, dal momento che le 8.094 sequenze
rappresentano 4.1 Mb del genoma, è possibile stimare che la distanza media tra i motivi SSRs,
in peperone, sia approssimativamente di 3.1 kb. Si riscontra, cioè, 1 sequenza microsatellite
ogni 9.0 sequenze. Dalla stima sono state escluse 7 sequenze, che coprono circa 120 Kbp del
genoma e sono caratterizzate da una lunghezza superiore a 1500 bp, in quanto sono state
analizzate separatamente.
Per evitare una sovrastima del numero di SSR presenti, è stata effettuata un’analisi della
ridondanza presente nelle 899 sequenze identificate. E’ emerso che 422 sequenze possono
essere raggruppate in 99 clusters; 477 sequenze risultano invece uniche. Sono state pertanto
identificate 576 sequenze uniche, che rappresentano il 7.1% delle sequenze totali, contenenti
783 microsatelliti. Tra queste, il numero di ripetizioni SSR non è uniformemente distribuita:
531 (68%) sono microsatelliti mononucleotidici, 103 (13%) dinucleotidici, 137 (18%)
trinucleotidici, 8 (1%) tetranucleotidici, 1 penta- e 3 exa-nucleotidici (Tab. 11)
Nell’ambito dei microsatelliti dimerici il motivo AG/CT è di gran lunga il più
abbondante, i motivi AC/GT e AT/AT sono presenti a frequenze inferiori mentre il motivo
CG/CG è del tutto assente nelle sequenze analizzate. Il motivo microsatellite trimerico più
frequente è AAG/CTT e quello tetramerico è AAAT/ATTT. La tabella 12 riporta i 783 SSR
identificati nelle sequenze non-ridondanti, classificati in base al loro motivo microsatellite e al
numero di ripetizioni.

IV-RISULTATI
112
Tab. 11 Ricerca di marcatori SSR, EST derivati. NR: sequenze non-ridondanti contenenti SSRs; NA: sequenze non allineate ai marcatori EST-SSR precedentemente sviluppati.
Totale NR NA Sequenze > 1.5 Kbp
Sequenze Totali esaminate: 8094 576 532 7 Sequenze contenenti SSR 899 576 532 7 SSRs Totali identificati: 1325 783 727 57 Taglia Totale delle sequenze contenenti SSR (Kbp):
579.1 283.8 257.7 120.6
Numero di sequenze contenenti più di 1 SSR:
298 157 146 4
Numero di SSRs composti presenti: 164 112 107 7 Mono 936 531 512 39 Di 167 103 88 12 Tri 207 137 116 4 Tetra 11 8 7 2 Penta 1 1 1 - Exa 3 3 3 -
Tale numero è molto variabile nell’ambito dei motivi mononucleotidici, di cui il 51%
appartiene alla classe di 10-15 unità ripetute. Il 65% degli SSR dinuclotidici e l’84% dei
trinucleotidici sono compresi rispettivamente, nelle classi di 6-7 e 5-6 unità ripetute.

IV-RISULTATI
113
Tab.12 Frequenza delle ripetizioni microsatellite uniche.
Motivo SSR Totale NR NA Sequences > 1.5 Kbp
A/T 903 506 488 35 C/G 33 25 24 4 AC/GT 21 16 14 1 AG/CT 99 60 51 2 AT/AT 47 27 23 9 AAC/GTT 20 19 12 1 AAG/CTT 61 40 36 AAT/ATT 25 17 14 1 ACC/GGT 35 15 12 2 ACG/CTG 4 3 3 - ACT/ATG 9 9 9 - AGC/CGT 11 11 9 - AGG/CCT 13 7 6 - AGT/ATC 16 10 10 - CCG/CGG 13 6 5 - AAAG/CTTT 1 1 1 - AAAT/ATTT 4 3 2 1 AAGG/CCTT 1 1 1 - AATT/AATT 3 2 2 - ACAT/ATGT 2 1 1 1 AAGAG/CTCTT 1 1 1 - AAAAAC/GTTTTT 1 1 1 - AACCCG/CTTGGG 1 1 1 - ACGAGG/CCTGCT 1 1 1 -
Total 1325 783 727 57
Una seconda analisi di clustering, per eliminare ulteriori sequenze ridondanti, è stata
effettuata sulle 576 sequenze contenenti SSR ed ha evidenziato che 44 di queste mostrano
omologia significativa con le sequenze EST utilizzate per lo sviluppo di marcatori SSR da
Nagy et al. (2004) e da Lee et al. (2004). In totale dunque, 532 sequenze (6.6% di tutte quelle
analizzate) rappresentano i potenziali candidati per lo sviluppo dei marcatori SSR, EST
derivati.
Sono state analizzate le 7 sequenze, caratterizzate da una lunghezza maggiore a 1500
bp, e sono state identificati 57 motivi microsatellite (Tab. 11). La maggior parte di questi, 39,
sono ripetizioni mononucleotidiche, mentre 12 sono ripetizioni dinucleotidiche. Il motivo
dimerico più frequente è AT/AT, tipico anche delle altre classi di unità ripetute, mentre il
motivo CG/CG risulta assente.

IV-RISULTATI
114
Sulla base delle 532 sequenze candidate, sono state disegnate 319 (60%) combinazioni
di primers che comprendono: 158 microsatelliti mononucleotidici, 34 dinucleotidici, 90
trinucleotidici, 5 tetranucleotidici, 3 exanucleotidici e 29 microsatelliti composti (Tab. 11). Le
sequenze restanti, poichè caratterizzate da sequenze fiancheggianti troppo corte o
inappropriate, non sono state impiegate per il disegno dei primers.
Un secondo set, costituito da 29 coppie di primers, è stato disegnato manualmente su 6
delle 7 sequenze caratterizzate da una lunghezza maggiore a 1500 bp.
Successivamente, è stato selezionato un sottoinsieme di 204 coppie di primers per
effettuare l’analisi dei polimorfismi presenti nei 16 genotipi in esame. I marcatori sono stati
scelti sulla base dei seguenti criteri: i) lunghezza minima di 18 ripetizioni per gli SSRs
mononucleotidici poly(A) e poly(T); ii) 7 ripetizioni per gli SSRs dinucleotidici; sono state
invece impiegate tutte le coppie di primers disegnate per le ripetizioni di ordine superiore e
per i microsatelliti composti. Questi 204 loci SSR sono stati testati sulle 5 accessioni
appartenenti al gruppo 1, descritte in tabella 6.

IV-RISULTATI
115
Tab. 13 Dettagli dei 49 primers SSR utili nel mappaggio.
Nom
e Accession
e ID
a
Descrizione
Motivo SSR
Sequenza dei primers
(forward , reverse)
Tag
lia
(bp)
Test sui
Parentalib
Prodotto
(bp)
N°
alleli
PIC
EPMS-
596
CO907458
C. annuum cDNA
(A) 19
CTCGTGCCGTATTTCTGTCA
AAGGGCGTGTTTGGTATGAA
250
TP
235-240
3 0.64
EPMS-
600
CO776120
C. annuum cDNA
(A) 32
ATGGGTACGTGTTTGGGGTA
ACTTTATTCCTCGTGCCGAA
210
TP
210-218
3 0.56
EPMS-
601
CO776313
C. annuum cDNA
(A) 34
AAATTGAGAACATCGGTGCC
TAAAGAAAGAGCCTCGTGCC
230
TP
230-235
3 0.33
EPMS-
603
CO907570
C. annuum cDNA
(A) 39
GCGGTTCCCTATTTGAAGAA
ATAGGGGGAATTGGGTTCC
164
TP
231-234
2 0.24
EPMS-
628
CO907462
C. annuum cDNA
(T) 19
TGCTCCTTAAGACTGGCACC
GGGTTCGGCTCTGTTATTGA
280
TP
142-148
3 0.64
EPMS-
629
CO908660
C. annuum cDNA
(T) 19
GCTCGAGGGAGAGAGACTGT
GGTCATATGTTTCCATGGGC
228
TP-YC
220-225
3 0.42
EPMS-
642
CO911525
C. annuum cDNA
(AT) 8
CAACTTCGCGTTATTGTCCA
AGGGCGGACAAAGAAGATTT
187
TP
180-210
5 0.71
EPMS-
643
CO908387
C. annuum cDNA
(CT) 17
CCAAGATCAACTCTTACGCTATC
CCCTCAAGAATTCCCTCCAT
212
TP-GC-YC
195-223
9 0.82
EPMS-
648
CO907649
C. annuum cDNA
(GA) 7
TGTAAAATAAAATAAGGCTAAAGGCA
CAAGAAAGTGTGCCCCAAAT
167
TP-YC
165-190
4 0.49
EPMS-
649
CO776131
C. annuum cDNA
(TA) 12
AAGGGTTCTCGAGGAAATGC
TCAATCCCAAAACCATGTGA
263
TP-YC
249-281
8 0.76
EPMS-
650
CO910134
C. annuum cDNA
(TA) 19
CATGGGTGAGGGTACATGGT
AGAGGGAAGGGTTATTTGCC
251
TP-GC-YC
237-261
8 0.86
EPMS-
654
CO776379
C. annuum cDNA
(AAC)5
TTCCACTCTTCGAAGCACCT
GGTAGGGTTTAACACCGCCT
262
TP
263-267
4 0.72
EPMS-
657
CK901687
C. annuum Root cDNA clone similar
to cold-stress inducible protein
(AAG) 5
CTGATCGTGGATGTGGATTG
TAGAATTGCTGTGAGTGCGG
165
TP
159-168
4 0.59

IV-RISULTATI
116
Nom
e Accession
e ID
a
Descrizione
Motivo SSR
Sequenza dei primers
(forward , reverse)
Tag
lia
(bp)
Test sui
Parentalib
Prodotto
(bp)
N°
alleli
PIC
EPMS-
658
CO906736
C. annuum cDNA
(AAG)5
CCTTGAGTAGGCGCACAAAT
TTCCTCATTGCTTTTCCCAC
142
TP
139-160
6 0.58
EPMS-
669
AY488030
C. annuum ly200 protein mRNA
(AGA) 5
AGAGCAGGCGAAACCCTAAT
GGCATCAAGACACATCCCTT
214
TP
290-296
3 0.30
EPMS-
670
CO910307
C. annuum cDNA
(AGA) 5
TCACAAAGATGGAGAAGGGAA
CAATCACTGTCACTGCTACTGCT
102
GC-YC
107-110
2 0.24
EPMS-
676
CO910259
C. annuum cDNA
(ATA) 7
TGGTGAACATGAAGAGCCAA
TTTAGCTGTTTGCCATGCAC
214
TP
201-223
5 0.55
EPMS-
677
CO909968
C. annuum cDNA
(ATA) 8
ATCTGCCCTTATCGATGCAC
CCGAATTGTGGAGGAAACAT
216
TP
200-212
4 0.68
EPMS-
680
CO911150
C. annuum cDNA
(ATT) 6
TGGAATTCACATGGTGAAAAA
TGAAACTTTGTGGGCTATGG
279
TP
267-284
4 0.63
EPMS-
683
CO907354
C. annuum cDNA
(CAA) 7
AAATGGATCCCAACAACCAA
GGAGTTGAAAACGGTGGAGA
200
TP-GC
184-206
5 0.71
EPMS-
686
CK901658
C. annuum Root cDNA clone
(CAG) 6
CAAAGAAGGAAGGGGGTTTC
CTCCCTCTTGTTCCTTGCAC
158
TP-YC
152-158
3 0.58
EPMS-
689
CO910607
C. annuum cDNA
(CAT) 7
AGAGGGGGTACTCATTGGCT
TCGGAGGAAGAGGAAGAACA
272
TP-GC-YC
253-275
4 0.47
EPMS-
694
CO912323
C. annuum cDNA
(CCA) 8
CTAGTACGAGGCAGGGGAGG
CCAGATCCCGCTTTTGACTA
238
TP-GC-YC
228-254
6 0.74
EPMS-
697
CO911728
C. annuum cDNA
(CTG) 5
ATGTCGCTCGCAATTTCACT
CGTAGGGAGGAGCGATAGAG
120
TP-GC-YC
110-120
4 0.54
EPMS-
703
CO776113
C. annuum cDNA
(GAA) 5
AAGATTTGGCGGAGACTTCA
TGCACCAACTTTGTCTCTGC
104
TP
106-118
6 0.69
EPMS-
704
CO910889
C. annuum cDNA
(GAA) 5
GGTCCTCTGATTGGCAACAT
GACCTGAAATTGGAGCAAACA
126
GC-YC
126-134
3 0.52
EPMS-
705
CO909594
C. annuum cDNA
(GAA) 5
TCAACTAGATCCACCACGCA
TAACCCGTTGCTCACACTCA
106
TP
105-114
3 0.65

IV-RISULTATI
117
Nom
e Accession
e ID
a
Descrizione
Motivo SSR
Sequenza dei primers
(forward , reverse)
Tag
lia
(bp)
Test sui
Parentalib
Prodotto
(bp)
N°
alleli
PIC
EPMS-
709
CO909799
C. annuum cDNA
(GAG) 6
ACGCCGAGGACTATGATGAC
TTCTTCATCCTCAGCGTGTG
272
TP-GC
262-280
7 0.79
EPMS-
712
CK902011
C. annuum Root cDNA clone similar
to At1g01160/F6F3_1 protein
(GCA) 6
CCACAAAGGGTTTAAGCAGC
AAGGCAGGAGCAGAGTTCAA
148
TP-GC
140-146
3 0.54
EPMS-
715
CK901642
C. annuum Root cDNA clone similar to
WRKY transcription factor 41 protein
(GGT) 5
GTGCCAAACATCCAAGAGGT
ACCAAAGGTAGTGGTGCTGG
196
TP
180-196
3 0.60
EPMS-
716
CO775942
C. annuum cDNA
(TAA) 12
CAGGAGATCCGCAGGATAAG
TTTTAAAGTTGATCAAGCGTAATTTTT
223
TP-GC-YC
194-232
7 0.84
EPMS-
718
CO912216
C. annuum cDNA
(TAT) 5
TCACTCGTTATATTTTTCTGTCTCG
CTGCTGAAGCGTGGTGAATA
229
TP
210-234
6 0.76
EPMS-
725
CO910566
C. annuum cDNA
(TCT) 8
TTGAATCGTTGAAGCCCATT
ATCTGAAGCTGGGCTCCTTT
158
TP-GC-YC
142-157
5 0.73
EPMS-
745
CO907754
C. annuum cDNA
(ATAC) 7
GTTGTTGGGTGGTACTTGGG
GGAAGATCTCAAATGGGTCG
223
TP-GC-YC
212-231
4 0.69
EPMS-
747
CO907904
C. annuum cDNA
(TTCT) 5
CATTGGACGGTTGGTTCTCT
TGGAATTGGAACTTCAAGCA
274
TP-GC-YC
265-285
4 0.69
EPMS-
749
CO909840
C. annuum cDNA
(AAAACA) 5
TCCCACGAGTCTTTTTGAGG
AAAGGGAATTATGGGGTTGG
270
TP-GC-YC
268-278
5 0.65
EPMS-
750
CO906574
C. annuum cDNA
(CCCGAA) 5
GGGATCCGAATCAGAATACG
TCTAGGTTTCGGGGAGGATT
253
TP
236-257
7 0.73
EPMS-
751
CO776484
C. annuum cDNA
(CTGCTC) 5
AGACTGCACCAGCTGAGGTT
GTAGCCGCAGACTCCTCAAC
262
TP
213-215
2 0.44
EPMS-
755
CO912244
C. annuum cDNA
(A) 12...(T) 11
CGCTCGCTACCCTTTCATTA
AATTTCGGAAGGGCAAAGAT
141
TP-GC-YC
141-150
4 0.68
EPMS-
757
CO776067
C. annuum cDNA
(AG) 6...(GA) 9
CCAACCAAACTTCAACTCCC
GATGCACCCACCATTATTCC
216
TP-GC-YC
205-219
4 0.62
EPMS-
762
CO910085
C. annuum cDNA
(CT) 6...(AT) 6
CGGCGAGATATGGACTTGAT
CCCACGTTATACCATCCAGG
261
TP-GC-YC
344-357
5 0.78

IV-RISULTATI
118
Nom
e Accession
e ID
a
Descrizione
Motivo SSR
Sequenza dei primers
(forward , reverse)
Tag
lia
(bp)
Test sui
Parentalib
Prodotto
(bp)
N°
alleli
PIC
EPMS-
773
CO909133
C. annuum cDNA
(T) 13...(T) 12
CGAGCAACTCCCTCTTATCG
TCGAATAGCACGCACGTTAG
166
TP
163-169
4 0.53
EPMS-
917
AY702979
(CT) 9
TTACCCAACCATCAGAAATGG
GTGTCCTCTCATGAGCATGG
176
TP
170-178
3 0.52
EPMS-
919
AY702979
(T) 12
TTTGGAGTTTGCGAATTGTG
AAGCAACGTGTGTAGGGAGG
231
TP-GC-YC
231-238
4 0.65
EPMS-
921
AY702979
C. annuum clone YAC genom
ic
sequence
(A) 13
AAATTTTTGGATTTTGGGCC
GCGACTTTAGAAACTTCCGC
295
TP
295-302
3 0.64
EPMS-
922
AY308828
(T) 11..(CT) 5
TTTTGGGGTTGTGAAGGAAG
TGTGTCTGCACTTTTGAGCC
492
TP
500-515
3 0.68
EPMS-
923
AY308828
(A) 15
CAAAACCAAATAGGTCCCCC
CGCGCAATAATTCAATATCG
310
TP-GC
261-349
6 0.80
EPMS-
924
AY308828
(CT) 6..(TA) 9..(GT
A) 5
GCCGTCGTCAGAAAAGGTAG
TGCATTTCTGTCAGAGGCTG
287
TP-GC-YC
260-295
8 0.82
EPMS-
925
AY308828
C. annuum salicylic acid-induced
protein 19 gene
(AT) 8
CTCACAAGCAGAAGTGGACC
CCCAGTAAAACTTAACCGCAC
316
TP
314-329
5 0.74
b SSR utili per studi di m
appaggio nelle progenie ‘Tabasco’ x ‘P4’ (TP), ’GD1’ x ‘CM334’ (GC) or ‘Yolo Wonder’ x ‘CM 334’ (YC)

VI-RISULTATI
Il risultato finale è stato lo sviluppo di 49 marcatori SSR, derivati da EST, utili nel
mappaggio molecolare (Tab. 13); di questi, 47 si sono rivelati polimorfici in ‘Tabasco’ e ‘P4’
e possono essere impiegati per il mappaggio inter-specifico, mentre 21 risultano utili in
entrambe le popolazioni intra-specifiche ‘GD1’ x ‘CM334’ e ‘Yolo Wonder’ x ‘Criollo de
Morellos’. In totale, il 49% dei marcatori SSR sviluppati sono polimorfici in una popolazione
segregante, il 18% in due e il 31% in tutte e tre le popolazioni di mappaggio.
Il set di 204 coppie di primers testati è caratterizzato, per la maggior parte, da motivi
trinucleotidici (47%), seguito da un 21% di motivi mono- e un 12% di-nucleotidici.
Nonostante questi ultimi siano poco rappresentati, sono in grado di evidenziare un elevato
livello di polimorfismo (28%) rispetto a entrambe i motivi tri- (23%) e mononucleotidici
(21%). Inoltre, il numero di ripetizioni non risulta correlato al livello di polimorfismo
rilevabile.
Il PIC è stato calcolato per tutti i 49 marcatori SSR analizzati sui 16 genotipi descritti in
tabella 6, allo scopo di ottenere una stima esaustiva circa il numero di alleli e le rispettive
dimensioni dei loci identificati (Tab. 13).
Sono stati amplificati 219 frammenti PCR totali e il numero di alleli osservato varia da
2 a 9, con una media di 4.5. La maggiore variabilità nelle dimensioni dei frammenti è stata
osservata nel marcatore EPMS-923 e va da 261 a 349 bp. I valori di PIC variano da 0.24 a
0.86 con una media di 0.62 ± 0.02. Il marcatore EPMS-650 ha evidenziato il valore di PIC più
elevato, mentre il valore più basso è stato osservato nei marcatori EPMS-603 e EPMS-670. I
marcatori caratterizzati da un valore di PIC elevato hanno un forte potenziale nel rilevare
molteplici varianti alleliche e possono essere impiegati per identificare le linee parentali più
appropriate da impiegare in studi di mappaggio o per valutare la variabilità genetica
all’interno di una popolazione (Anderson et al. 1993).
E’ stata pertanto verificata la possibilità di impiegare i marcatori SSR sviluppati per
analizzare la variabilità esistente in Capsicum spp. Sulla base dei 219 alleli individuati è
emerso che la similarità genetica (GS) tra le accessioni impiegate nello studio, è variabile da
0.09 a 0.81. Le accessioni ‘Tabasco’ e ‘CM334’ evidenziano il GS minimo, mentre ‘Chi1’ e
‘Chi7’ assumono il valore GS massimo. Il dendogramma generato col metodo UPGMA
riassume le relazioni esistenti tra le popolazioni analizzate. Le varietà di C. annuum e le linee
inbred risultano ben differenziate dalle altre accessioni di Capsicum, le quali risultano
distribuite in due clusters: il primo comprende le accessioni che derivano da C. frutescens, C.
chinense e C. eximium, mentre il secondo comprende C. baccatum var. baccatum e var.
pendulum, C. pubescens, C. chacoense e C. praetermissum (Fig. 62).

IV-RISULTATI
120
Fig. 62 Dendogramma UPGMA delle 16 accessioni di Capsicum analizzate sulla base dei 219 alleli EST-SSR.
Coefficient0.23 0.42 0.61 0.81 1.00
CM334
YW
MW1
Per
GDI
P4
CM334
Fru48
Chi1
Chi7
Exi
Tab
Pub3
Pen6
Bac20
Pra1
Cha6

IV-RISULTATI
121
2. Marcatori polimorfici e segregazione.
Lo screening di 90 marcatori SSR ha rilevato che 38 di essi sono polimorfici nella
popolazione segregante e possono essere utilizzati nel mappaggio. In tabella 14, 15 e 16 sono
riportati rispettivamente i 7 marcatori SSR polimorfici, sviluppati da Lee et al. (2004):
Hpms1_69, Hpms1_172, Hpms1_214, Hpms1_5, Hpms1_62, Hpms2_13, Hpms2_24; i 20
marcatori SSR polimorfici, sviluppati da Sasvári et al. (2004): EPMS_310 , EPMS_340,
EPMS_353, EPMS_372, EPMS_376, EPMS_377, EPMS_386, EPMS_391, EPMS_402,
EPMS_409, EPMS_410, EPMS_419, EPMS_421, EPMS_426, EPMS_429, EPMS_441,
EPMS_472, EPMS_480, EPMS_497, EPMS_561; e gli 11 marcatori SSR polimorfici,
sviluppati nell’ambito del lavoro di tesi: EPMS_643, EPMS_649, EPMS_650, EPMS_670,
EPMS_686, EPMS_694, EPMS_704, EPMS_725, EPMS_747, EPMS_755, EPMS_919.
Tab.14 Primers sviluppati da Lee et al. 2004 e utilizzati nella mappa.
Primer SSR Motivo
microsatellite Sequenza Taglia
Hpms1_69 (AC)9(AT)4 L: CGGTGGCATGTAGTTTCTGGAG
R: AAGACATGAAATCCACAAGTTTTC 217
Hpms1_172 (GA)15 L: GGGTTTGCATGATCTAAGCATTTT
R: CGCTGGAATGCATTGTCAAAGA 344
Hpms1_214 (GTTT)2(TTG)9 L: TGCGAGTACCGAGTTCTTTCTAG
R: GGCAGTCCTGGGACAACTCG 100
Hpms1_5 (AT)11(GT)17 L: CCAAACGAACCGATGAACACTC
R:ACAATGTTGAAAAAGGTGGAAGAC 311
Hpms1_62 (TG)23(AG)9 L: CATGAGGTCTCGCATGATTTCAC
R:GGAGAAGGACCATGTACTGCAGAG 186
Hpms2_13 (AC)12(AT)4 L: TCACCTCATAAGGGCTTATCAATC
R: TCCTTAACCTTACGAAACCTTGG 259
Hpms2_24 (CT)17(CA)5A21 L: TCGTATTGGCTTGTGATTTACCG
R: TTGAATCGAATACCCGCAGGAG 205

IV-RISULTATI
122
Tab. 15 Primers SSR di Sasvári et al. 2004 polimorifici nella popolazione RIL.
SSR Riferimento Taglia (bp)
EPMS_310 300/330
EPMS_340 BM064640 273/260
EPMS_353 BM067271 299/305
EPMS_372 CA515633 326/324
EPMS_376 CA516334 260/256
EPMS_377 CA516439 156/162
EPMS_386 CA517699 149/143
EPMS_391 CA519548 187/185
EPMS_402 CA523427 203/201
EPMS_409 CA525246 174/164
EPMS_410 CA525390 187/184
EPMS_419 CA523208 226/224
EPMS_421 CA514272 256/247
EPMS_426 CA517376 116/108
EPMS_429 CA525246 174/162
EPMS_441 CA847465 118/124
EPMS_472 BM061910 293/320
EPMS_480 BM062655 256/253
EPMS_497 CA522759 251/250
EPMS_561 BM068460 114/117

IV-RISULTATI
123
Tab. 16 Sequenza dei primers SSR, da noi sviluppati, polimorifici nella popolazione RIL.
SSR Motivo
microsatellite
Taglia
(bp) Riferimento Sequenza
EPMS_643 (CT)17 212 CO908387
F: CCAAGATCAACTCTTACGCTATC R: CCCTCAAGAATTCCCTCCAT
EPMS_649 (TA)12 260/240 CO776131_c43
F:AAGGGTTCTCGAGGAAATGC R:TCAATCCCAAAACCATGTGA
EPMS_650 (TA)19 250 CO910134
R:CATGGGTGAGGGTACATGGT F:AGAGGGAAGGGTTATTTGCC
EPMS_670 (AGA)5 102 CO910307
R:TCACAAAGATGGAGAAGGGAA F:CAATCACTGTCACTGCTACTGCT
EPMS_686 (CAG)6 158 CK901658
R:CAAAGAAGGAAGGGGGTTTC F:CTCCCTCTTGTTCCTTGCAC
EPMS_694 (CCA)8 238 CO912323
R:CTAGTACGAGGCAGGGGAGG F:CCAGATCCCGCTTTTGACTA
EPMS_704 (GAA)5 160/150 CO910889
R:GGTCCTCTGATTGGCAACAT F:GACCTGAAATTGGAGCAAACA
EPMS_725 (TCT)8 158 CO910566
R:TTGAATCGTTGAAGCCCATT F:ATCTGAAGCTGGGCTCCTTT
EPMS_747 (TTCT)5 280 CO907904
R:CATTGGACGGTTGGTTCTCT
F:TGGAATTGGAACTTCAAGCA
EPMS_755 (A)12...(T)11 150/140 CO912244
R:CGCTCGCTACCCTTTCATTA F:AATTTCGGAAGGGCAAAGAT
EPMS_919 (A)12...(T)11 231 AY702979
R:TTTGGAGTTTGCGAATTGTG F:AAGCAACGTGTGTAGGGAGG

IV-RISULTATI
124
Le analisi AFLP (Tab. 17), condotte presso il Di. Va. P.R.A. (Università degli Studi di
Torino) e l’I.N.R.A (Montflavet – Francia), hanno rilevato 507 polimorfismi: 284 sono stati
generati applicando la combinazione di primers (PCs) EcoRI/MseI, 202 sono stati generati
applicando PstI/MseI PCs e 21 applicando SacI/MseI PCs. EcoRI/MseI PCs ha prodotto da 7
(E33/M56) a 24 (E38/M60) bande: 15 in media; PstI/MseI PCs ha prodotto da 13 (P19/M42)
a 29 (P14/M33) bande: 18 in media; due SacI/MseI PCs hanno prodotto rispettivamente 15 e
16 polimorfismi.
261 AFLP analizzati derivano dal genoma di “Criollo de Morellos” e 245 da quello di
“Yolo Wonder”.
Tab. 17 Marcatori AFLP e polimorfismi
Combinazione di primers N° di bande
polimorfiche
Combinazione di
primers
N° di bande
polimorfiche
C33/M54 15 E42/M48 19
C36/M49 6 E43/M53 13
E31/M58 18 E43/M54 13
E32/M55 10 E44/M51 14
E33/M56 7 E44/M61 12
E34/M53 16 P11/M49 14
E36/M47 14 P14/M33 29
E36/M52 19 P14/M39 27
E37/M54 9 P14/M41 21
E38/M60 24 P15/M40 16
E38/M61 18 P15/M43 17
E40/M47 18 P17/M32 16
E40/M49 7 P17/M39 11
E41/M48 20 P19/M42 14
E41/M54 19 P19/M45 18
E41/M61 16 P25/M42 19

IV-RISULTATI
125
Il set di primers SSAP ha generato 20 bande polimorfiche (Tab. 18). Il marcatore
T135E ha prodotto 11 polimorfismi: 7 derivano dal genoma di “Yolo Wonder” e 4 da quello
di “Criollo de Morellos”. Il marcatore TntC ha prodotto 9 bande polimorfiche di cui 6
derivano dal genoma di “Yolo Wonder” e 3 da quello di “Criollo de Morellos”.
Tab. 18 Marcatori SSAP e polimorfismi.
Tipo
Primer/adattatore
Sequenza (5’ 3’)
N° bande
polimorfiche
Primer SSAP per il
retrotrasposone
Primer SSAP
adattatore
T135E
E00 (Eco RI)
TGGTCAAAAATGGGTTGGTCC
GTAGACTGCGATCCAATTC
11
Primer SSAP per il
retrotrasposone
Primer SSAP
adattatore
TntC
C00 (Csp 6I)
TTCCCACCTCACTACAATATCGC
GACGATGAGTCCTGAGATAC
9
I 18 marcatori RFLP impiegati sono cloni di cDNA che derivano da librerie di lievito,
Arabidopsis, tabacco, patata e pomodoro (Tab. 19); i 12 marcatori STS sono stati sviluppati
da marcatori CAPs, costruiti su geni di peperone (Tab. 20). Entrambe hanno generato bande
polimorfiche nella popolazione RIL.

IV-RISULTATI
126
Tab. 19 Marcatori RFLP utilizzati per lo sviluppo della mappa.
RFLP DESCRIZIONE SPECIE
CD35 Hypothetical yeast protein Lievito
CT080 Serine Hydroxymethil Transferase Arabidopsis thaliana
CT100 Xyloglucan endptransglycosilase Pomodoro
CT145 G subunit of the Vacuolar-type
ATPase Tabacco
CT224 Alpha amylase Patata
GC015_1 Acid class III chitinose-lysozime Tabacco
GC207 Unknow protein
PG101 Unknow protein
PG108 Unknow protein
PT001c Unknow protein
T_1430 Putative phospholipase Arabidopsis thaliana
TG29 Subtilisin-like proteinase (EC 3.4.21.-)
4 Pomodoro
TG46 Unknow protein Arabidopsis thaliana
TG123 Phosphatidylglycerolophosphate
synthase (PGS1) Arabidopsis thaliana
TG132 S-locus protein kinase family Arabidopsis thaliana
TG281 Guanine nucleotide-exchange protein-
like Arabidopsis thaliana
C2_At3g52220 Unknow protein Pomodoro
C2_At3g58470 Unknow protein Pomodoro
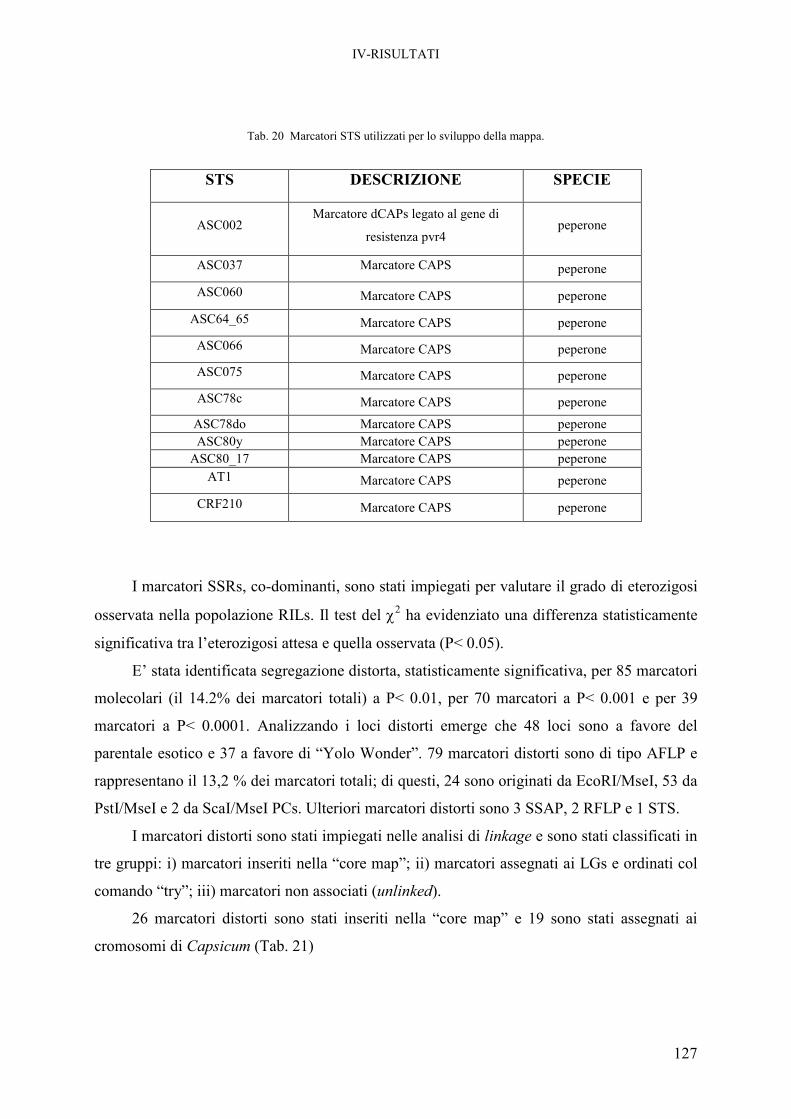
IV-RISULTATI
127
Tab. 20 Marcatori STS utilizzati per lo sviluppo della mappa.
STS DESCRIZIONE SPECIE
ASC002 Marcatore dCAPs legato al gene di
resistenza pvr4 peperone
ASC037 Marcatore CAPS peperone
ASC060 Marcatore CAPS peperone
ASC64_65 Marcatore CAPS peperone
ASC066 Marcatore CAPS peperone
ASC075 Marcatore CAPS peperone
ASC78c Marcatore CAPS peperone
ASC78do Marcatore CAPS peperone ASC80y Marcatore CAPS peperone ASC80_17 Marcatore CAPS peperone
AT1 Marcatore CAPS peperone
CRF210 Marcatore CAPS peperone
I marcatori SSRs, co-dominanti, sono stati impiegati per valutare il grado di eterozigosi
osservata nella popolazione RILs. Il test del χ2 ha evidenziato una differenza statisticamente
significativa tra l’eterozigosi attesa e quella osservata (P< 0.05).
E’ stata identificata segregazione distorta, statisticamente significativa, per 85 marcatori
molecolari (il 14.2% dei marcatori totali) a P< 0.01, per 70 marcatori a P< 0.001 e per 39
marcatori a P< 0.0001. Analizzando i loci distorti emerge che 48 loci sono a favore del
parentale esotico e 37 a favore di “Yolo Wonder”. 79 marcatori distorti sono di tipo AFLP e
rappresentano il 13,2 % dei marcatori totali; di questi, 24 sono originati da EcoRI/MseI, 53 da
PstI/MseI e 2 da ScaI/MseI PCs. Ulteriori marcatori distorti sono 3 SSAP, 2 RFLP e 1 STS.
I marcatori distorti sono stati impiegati nelle analisi di linkage e sono stati classificati in
tre gruppi: i) marcatori inseriti nella “core map”; ii) marcatori assegnati ai LGs e ordinati col
comando “try”; iii) marcatori non associati (unlinked).
26 marcatori distorti sono stati inseriti nella “core map” e 19 sono stati assegnati ai
cromosomi di Capsicum (Tab. 21)
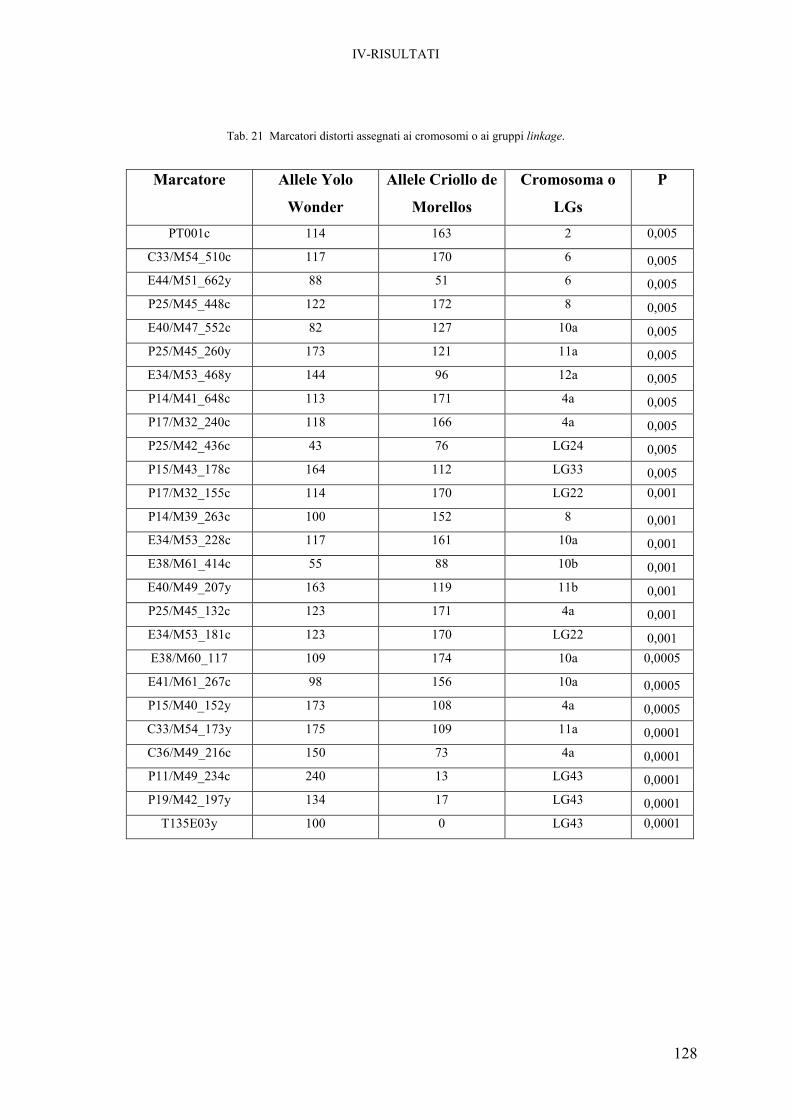
IV-RISULTATI
128
Tab. 21 Marcatori distorti assegnati ai cromosomi o ai gruppi linkage.
Marcatore Allele Yolo
Wonder
Allele Criollo de
Morellos
Cromosoma o
LGs
P
PT001c 114 163 2 0,005
C33/M54_510c 117 170 6 0,005
E44/M51_662y 88 51 6 0,005
P25/M45_448c 122 172 8 0,005
E40/M47_552c 82 127 10a 0,005
P25/M45_260y 173 121 11a 0,005
E34/M53_468y 144 96 12a 0,005
P14/M41_648c 113 171 4a 0,005
P17/M32_240c 118 166 4a 0,005
P25/M42_436c 43 76 LG24 0,005
P15/M43_178c 164 112 LG33 0,005
P17/M32_155c 114 170 LG22 0,001
P14/M39_263c 100 152 8 0,001
E34/M53_228c 117 161 10a 0,001
E38/M61_414c 55 88 10b 0,001
E40/M49_207y 163 119 11b 0,001
P25/M45_132c 123 171 4a 0,001
E34/M53_181c 123 170 LG22 0,001
E38/M60_117 109 174 10a 0,0005
E41/M61_267c 98 156 10a 0,0005
P15/M40_152y 173 108 4a 0,0005
C33/M54_173y 175 109 11a 0,0001
C36/M49_216c 150 73 4a 0,0001
P11/M49_234c 240 13 LG43 0,0001
P19/M42_197y 134 17 LG43 0,0001
T135E03y 100 0 LG43 0,0001

IV-RISULTATI
129
3. Costruzione della mappa linkage
La mappa genetica è stata sviluppata mediante l’utilizzo di 595 marcatori molecolari:
507 AFLPs, 18 RFLPs, 20 SSAPs e 12 STSs e 38 SSR. Di questi, 499 marcatori, che
comprendono 428 AFLP, 15 RFLP, 19 SSAP, 12 STS e 25 SSR, sono stati raggruppati in 49
gruppi linkage: 14 large LGs, comprendenti da 10 a 60 marcatori, e 35 small LGs,
comprendenti da 2 a 9 marcatori. I gruppi sono stati determinati usando un LOD score
minimo pari a 8 e un valore massimo di ricombinazione pari a 0.1. La mappa finale copre
1857 cM del genoma di peperone, con una distanza di mappa media di 5,71.
La mappa genetica di base, “core map”, è stata sviluppata dall’analisi molecolare di 325
marcatori (54.4%) il cui ordine sui LGs è confermato col comando “ripple” a LOD score
minimo di 3. Di questi, 251 marcatori: 223 AFLPs, 10 SSRs, 2 SSAP, 13 RLFP e 3 STS sono
localizzati sui 22 gruppi linkage assegnati ai 12 cromosomi di peperone; coprono 1553 cM del
genoma totale, con una distanza di mappa media tra i marcatori pari a 6,18 cM.

IV-RISULTATI
130
4. Posizione dei marcatori sui gruppi linkage
I marcatori assegnati ai gruppi linkage sono 499: 428 AFLP, 19 SSAP, 12 STS, 15
RFLP e 25 SSR. I marcatori non assegnati ai gruppi linkage sono 96 e comprendono: 79
AFLP, 1 SSAP, 3 RFLP e 13 SSR. Le posizioni che i marcatori occupano sulla mappa sono
riassunte nelle tabelle 22 e 23.
Tab. 22 Posizione dei marcatori STS e RFLP sulla mappa.
STS
CROMOSOMA
DI PEPERONE
RFLP
CROMOSOMA
DI PEPERONE
ASC002 P10b (LG16) CD35 P2
ASC037 P5 CT080 P5
ASC060 LG23 CT100 unlinked
ASC64_65 P5 CT145 unlinked
ASC066 P5 CT224 P4b
ASC075 P7 GC015_1 P9
ASC78c P5 GC207 P4a
ASC78do P5 PG101 P3
ASC80y P5 PG108 P6
ASC80_17 P5 PT001c P2
AT1 LG48 T_1430 P5
CRF210 P12 TG29 P1
TG46 unlinked
TG123 P5
TG132 P4a
TG281 P8
C2_At3g52220 P12
C2_At3g58470 unlinked

IV-RISULTATI
131
Tab. 23 Posizione dei marcatori SSR sulla mappa.
SSR CROMOSOMA
DI PEPERONE
GRUPPO
LINKAGE
SSR CROMOSOMA DI
PEPERONE
GRUPPO
LINKAGE
EPMS_310 P8 LG4 EPMS_643 - unlinked
EPMS_340 P4a LG1 EPMS_649 P19 LG25
EPMS_353 - unlinked EPMS_650 P1 LG14
EPMS_372 P9 LG2 EPMS_670 - unlinked
EPMS_376 P18 LG24 EPMS_686 - unlinked EPMS_377 P29 LG39 EPMS_694 - unlinked EPMS_386 - unlinked EPMS_704 - LG21
EPMS_391 P20 LG27 EPMS_725 - LG14
EPMS_402 P34 LG45 EPMS_755 P27 LG37
EPMS_409 P2 LG5 EPMS_747 - unlinked
EPMS_410 P11a LG6 EPMS_919 - unlinked
EPMS_419 - unlinked Hpms 1_172 - unlinked
EPMS_421 - unlinked Hpms 1_214 P8 LG4
EPMS_426 P22 LG29 Hpms 1_5 P18 LG24
EPMS_429 - LG5 Hpms 1_62 P19 LG25
EPMS_441 P11b LG26 Hpms 2_13 - unlinked
EPMS_472 P3 LG3 Hpms 2_24 P15 LG19
EPMS_480 - unlinked Hpms 1_69 - unlinked
EPMS_497 P2 LG5
EPMS_561 - LG6

IV-RISULTATI
132
e33/m56_363y0,0
e44/m51_318y7,1
e31/m58_516y11,7
e40/m47_221y16,6
e38/m60_099y21,3
e36/m47_206y27,7
e36/m52_158y31,5e43/m54_256y32,4e36/m52_205c37,4
p14/m41_094c45,8e43/m54_195c49,1
e43/m53_286c53,7
e36/m52_225c55,6e40/m49_111c57,7
e34/m53_077c65,7
PG10195,7
p14/m39_221y130,5
p14/m33_242c137,9
e37/m54_260c146,4
p11/m49_196y153,0
c33/m54_198y160,7
EPMS_472174,1
3
c36/m49_216c0,0
p15/m40_152y30,8
p25/m45_160y38,6p14/m39_417y42,4GC20744,5e41/m48_226y49,3
e36/m52_392y56,4
e38/m60_109y63,4
e41/m48_078y70,1
EPMS_34076,6p17/m32_240c80,9p14/m41_648c85,3p25/m45_132c90,6Tntc01c93,8
TG132102,0
p25/m45_109y111,8
e41/m48_080c148,2
e38/m61_168c156,3e38/m61_158y161,1
e42/m48_116y170,4e41/m48_123y172,9
e37
/m5
4_
120
c
p15/m43_118c0,0p15/m43_195y3,3
CT22411,6
4a
CD0350,0
p25/m42_268c17,8p15/m43_158c18,5p11/m49_343c22,2
p14/m41_246y36,3p11/m49_268y40,7
EPMS_40953,6
p15/m43_254c62,4e36/m52_206c67,1e36/m52_116c70,2
e36/m47_146c83,4
e44/m51_258c91,1p14/m33_580c93,6PT001c95,9EPMS_497101,5e41/m54_221y104,6
e34/m53_189y110,7
e38/m
60_445
2
TntC09y0,0
e34/m53_233c7,4
e41/m48_159y18,1
e36/m52_190y22,8
p14/m39_210c46,6c33/m54_327y51,0
e40/m49_148y59,1e41/m61_199y62,9e38/m60_221y64,6e38/m60_220c67,2e38/m60_285c70,2e38/m61_167c73,4
EPMS_65085,3
TG29
1
p19/m42_126c0,0
c33/m54_299c7,3
e44/m61_125c15,0c36/m49_321y19,1T_143028,2p11/m49_638y33,3p14/m39_247y35,3e41/m48_112y37,2TG12341,8p17/m39_228y51,6p17/m32_118y55,8e38/m61_139y60,1e41/m54_117y62,2ASC64_6563,9e42/m48_155y66,9e40/m47_210c69,9e43/m54_405y72,3e36/m47_362c74,3
e36/m47_172y89,5e36/m47_258y97,6e31/m58_219y101,9e34/m53_211y104,0p15/m40_267y106,1e38/m61_295y108,5e38/m61_220c112,3e41/m54_105c114,2e41/m54_353c116,7e41/m61_384c119,4e40/m60_216c123,7e40/m60_215y128,7e43/m54_291y134,6p11/m49_140y137,7p14/m41_224y142,6p14/m41_223c145,2e41/m61_525c148,7
e44/m51_327y180,8p25/m42_129y184,9p15/m43_397y188,6CT080192,4p15/m43_238y197,8
p17/m32_318c205,4
5
e33/m56_363y0,0
e44/m51_318y7,1
e31/m58_516y11,7
e40/m47_221y16,6
e38/m60_099y21,3
e36/m47_206y27,7
e36/m52_158y31,5e43/m54_256y32,4e36/m52_205c37,4
p14/m41_094c45,8e43/m54_195c49,1
e43/m53_286c53,7
e36/m52_225c55,6e40/m49_111c57,7
e34/m53_077c65,7
PG10195,7
p14/m39_221y130,5
p14/m33_242c137,9
e37/m54_260c146,4
p11/m49_196y153,0
c33/m54_198y160,7
EPMS_472174,1
3
e33/m56_363y0,0
e44/m51_318y7,1
e31/m58_516y11,7
e40/m47_221y16,6
e38/m60_099y21,3
e36/m47_206y27,7
e36/m52_158y31,5e43/m54_256y32,4e36/m52_205c37,4
p14/m41_094c45,8e43/m54_195c49,1
e43/m53_286c53,7
e36/m52_225c55,6e40/m49_111c57,7
e34/m53_077c65,7
PG10195,7
p14/m39_221y130,5
p14/m33_242c137,9
e37/m54_260c146,4
p11/m49_196y153,0
c33/m54_198y160,7
EPMS_472174,1
3
c36/m49_216c0,0
p15/m40_152y30,8
p25/m45_160y38,6p14/m39_417y42,4GC20744,5e41/m48_226y49,3
e36/m52_392y56,4
e38/m60_109y63,4
e41/m48_078y70,1
EPMS_34076,6p17/m32_240c80,9p14/m41_648c85,3p25/m45_132c90,6Tntc01c93,8
TG132102,0
p25/m45_109y111,8
e41/m48_080c148,2
e38/m61_168c156,3e38/m61_158y161,1
e42/m48_116y170,4e41/m48_123y172,9
e37
/m5
4_
120
c
p15/m43_118c0,0p15/m43_195y3,3
CT22411,6
4a
c36/m49_216c0,0
p15/m40_152y30,8
p25/m45_160y38,6p14/m39_417y42,4GC20744,5e41/m48_226y49,3
e36/m52_392y56,4
e38/m60_109y63,4
e41/m48_078y70,1
EPMS_34076,6p17/m32_240c80,9p14/m41_648c85,3p25/m45_132c90,6Tntc01c93,8
TG132102,0
p25/m45_109y111,8
e41/m48_080c148,2
e38/m61_168c156,3e38/m61_158y161,1
e42/m48_116y170,4e41/m48_123y172,9
e37
/m5
4_
120
c
p15/m43_118c0,0p15/m43_195y3,3
CT22411,6
4a
CD0350,0
p25/m42_268c17,8p15/m43_158c18,5p11/m49_343c22,2
p14/m41_246y36,3p11/m49_268y40,7
EPMS_40953,6
p15/m43_254c62,4e36/m52_206c67,1e36/m52_116c70,2
e36/m47_146c83,4
e44/m51_258c91,1p14/m33_580c93,6PT001c95,9EPMS_497101,5e41/m54_221y104,6
e34/m53_189y110,7
e38/m
60_445
2
CD0350,0
p25/m42_268c17,8p15/m43_158c18,5p11/m49_343c22,2
p14/m41_246y36,3p11/m49_268y40,7
EPMS_40953,6
p15/m43_254c62,4e36/m52_206c67,1e36/m52_116c70,2
e36/m47_146c83,4
e44/m51_258c91,1p14/m33_580c93,6PT001c95,9EPMS_497101,5e41/m54_221y104,6
e34/m53_189y110,7
e38/m
60_445
2
TntC09y0,0
e34/m53_233c7,4
e41/m48_159y18,1
e36/m52_190y22,8
p14/m39_210c46,6c33/m54_327y51,0
e40/m49_148y59,1e41/m61_199y62,9e38/m60_221y64,6e38/m60_220c67,2e38/m60_285c70,2e38/m61_167c73,4
EPMS_65085,3
TG29
1
TntC09y0,0
e34/m53_233c7,4
e41/m48_159y18,1
e36/m52_190y22,8
p14/m39_210c46,6c33/m54_327y51,0
e40/m49_148y59,1e41/m61_199y62,9e38/m60_221y64,6e38/m60_220c67,2e38/m60_285c70,2e38/m61_167c73,4
EPMS_65085,3
TG29
1
p19/m42_126c0,0
c33/m54_299c7,3
e44/m61_125c15,0c36/m49_321y19,1T_143028,2p11/m49_638y33,3p14/m39_247y35,3e41/m48_112y37,2TG12341,8p17/m39_228y51,6p17/m32_118y55,8e38/m61_139y60,1e41/m54_117y62,2ASC64_6563,9e42/m48_155y66,9e40/m47_210c69,9e43/m54_405y72,3e36/m47_362c74,3
e36/m47_172y89,5e36/m47_258y97,6e31/m58_219y101,9e34/m53_211y104,0p15/m40_267y106,1e38/m61_295y108,5e38/m61_220c112,3e41/m54_105c114,2e41/m54_353c116,7e41/m61_384c119,4e40/m60_216c123,7e40/m60_215y128,7e43/m54_291y134,6p11/m49_140y137,7p14/m41_224y142,6p14/m41_223c145,2e41/m61_525c148,7
e44/m51_327y180,8p25/m42_129y184,9p15/m43_397y188,6CT080192,4p15/m43_238y197,8
p17/m32_318c205,4
5
p19/m42_126c0,0
c33/m54_299c7,3
e44/m61_125c15,0c36/m49_321y19,1T_143028,2p11/m49_638y33,3p14/m39_247y35,3e41/m48_112y37,2TG12341,8p17/m39_228y51,6p17/m32_118y55,8e38/m61_139y60,1e41/m54_117y62,2ASC64_6563,9e42/m48_155y66,9e40/m47_210c69,9e43/m54_405y72,3e36/m47_362c74,3
e36/m47_172y89,5e36/m47_258y97,6e31/m58_219y101,9e34/m53_211y104,0p15/m40_267y106,1e38/m61_295y108,5e38/m61_220c112,3e41/m54_105c114,2e41/m54_353c116,7e41/m61_384c119,4e40/m60_216c123,7e40/m60_215y128,7e43/m54_291y134,6p11/m49_140y137,7p14/m41_224y142,6p14/m41_223c145,2e41/m61_525c148,7
e44/m51_327y180,8p25/m42_129y184,9p15/m43_397y188,6CT080192,4p15/m43_238y197,8
p17/m32_318c205,4
5
5. Attribuzione dei gruppi linkage ai cromosomi.
La mappa sviluppata è stata allineata alle due mappe genetiche integrate di peperone,
sviluppate e pubblicate da Lefebvre et al. (2002) e da Paran et al. (2004). Dal confronto è
emerso che 42 marcatori molecolari: 33 AFLPs e 9 RFLPs sono marcatori comuni alle tre
mappe e sono stati utilizzati come “anchor marker”, per assegnare i gruppi linkage ai
cromosomi. E’ stato possibile assegnare 22 LGs ai 12 cromosomi di peperone, e coprire così
1553 cM del genoma totale; i restanti 27 LGs, che coprono 304 cM, non sono stati attribuiti ai
cromosomi (Fig. 63, 64, 65 e 66).
Ai cromosomi 4, 10, 11 e 12 sono stati assegnati due gruppi linkage ciascuno, ma non è
stato possibile unire i LGs a valori di LOD score maggiori a 3.0.
La taglia dei cromosomi è variabile: da 74.8 cM per il cromosoma 6, a 205.4 cM per il
cromosoma 5.
Fig. 63 Mappa gentica ad alta risoluzione F5YC.

IV-RISULTATI
133
p15/m43_393y0,0e34/m53_091y2,8
e33/m56_193y12,6
e44/m51_662y24,5PG10832,2e40/m47_178c38,7c33/m54_510c44,4e43/m54_452c48,1e43/m53_206c51,0p14/m33_341c53,3e42/m48_379y57,8e43/m53_205y60,4p14/m41_060y62,5p25/m45_185y65,4c33/m54_207y68,3e36/m52_114y70,7e44/m61_248y74,8
6
e41/m48_666c0,0
p25/m42_433y9,0
e42/m48_521c16,4e38/m60_093y20,6e41/m54_368y24,4e34/m53_379c28,3e32/m55_135c31,2e41/m48_140c34,1c33/m54_126c36,1e43/m53_135c38,5e38/m60_297c45,1
e41/m61_140c54,4
e32/m55_247y83,0
7
TG2810,0
p14/m39_263c10,3p25/m45_448c13,2
p11/m49_378y18,7
p11/m49_274c35,2
p11/m49_244y40,0p14/m33_267y45,3p14/m33_273c47,5p14/m39_335c52,9EPMS_31055,7
HPMS1_21486,7
e37
/m54_440
y
8
p25/m42_240y0,0
CT1458,8e37/m54_92c13,7
EPMS_37241,4
e44/m61_545c55,6
e43/m53_146c76,7
e36/m47_153c90,4e36/m47_211c93,4e38/m61_320c100,1e41/m54_110c107,0p14/m33_349c108,3p15/m40_321c109,5p25/m42_683c110,6c33/m49_081c111,9e42/m48_317c114,6e36/m52_136c115,7e44/m61_187y119,2e36/m52_220y121,2e32/m55_191y124,3p14/m41_672y128,3e38/m60_157y132,1e44/m61_480y133,7e44/m61_515y136,2p14/m39_324y139,6e41/m48_120y144,0e41/m54_210y148,7e31/m58_117y154,0e32/m55_392y157,1e31/m58_400y168,5
e43/m54_708c180,5e43/m53_711c185,3e43/m53_451c189,5e36/m52_434c194,0
e41/m61_463c205,2
9
e40/m49_247c0,0
e34/m53_145y9,2
e31/m58_367y20,3
e36/m47_145y28,9
e34/m53_228c43,9
e38/m60_117c50,2
e36/m52_132c56,0
e40/m47_552c61,0
e41/m61_267c67,1
e41/m61_266y72,4
c36/m49_282y81,8
e41/m54_137y0,0
e38/m60_264c11,9
p14/m41_215c16,7
e38/m61_403c25,2
ASC00234,6
10a
p15/m43_393y0,0e34/m53_091y2,8
e33/m56_193y12,6
e44/m51_662y24,5PG10832,2e40/m47_178c38,7c33/m54_510c44,4e43/m54_452c48,1e43/m53_206c51,0p14/m33_341c53,3e42/m48_379y57,8e43/m53_205y60,4p14/m41_060y62,5p25/m45_185y65,4c33/m54_207y68,3e36/m52_114y70,7e44/m61_248y74,8
6
p15/m43_393y0,0e34/m53_091y2,8
e33/m56_193y12,6
e44/m51_662y24,5PG10832,2e40/m47_178c38,7c33/m54_510c44,4e43/m54_452c48,1e43/m53_206c51,0p14/m33_341c53,3e42/m48_379y57,8e43/m53_205y60,4p14/m41_060y62,5p25/m45_185y65,4c33/m54_207y68,3e36/m52_114y70,7e44/m61_248y74,8
6
e41/m48_666c0,0
p25/m42_433y9,0
e42/m48_521c16,4e38/m60_093y20,6e41/m54_368y24,4e34/m53_379c28,3e32/m55_135c31,2e41/m48_140c34,1c33/m54_126c36,1e43/m53_135c38,5e38/m60_297c45,1
e41/m61_140c54,4
e32/m55_247y83,0
7
e41/m48_666c0,0
p25/m42_433y9,0
e42/m48_521c16,4e38/m60_093y20,6e41/m54_368y24,4e34/m53_379c28,3e32/m55_135c31,2e41/m48_140c34,1c33/m54_126c36,1e43/m53_135c38,5e38/m60_297c45,1
e41/m61_140c54,4
e32/m55_247y83,0
7
TG2810,0
p14/m39_263c10,3p25/m45_448c13,2
p11/m49_378y18,7
p11/m49_274c35,2
p11/m49_244y40,0p14/m33_267y45,3p14/m33_273c47,5p14/m39_335c52,9EPMS_31055,7
HPMS1_21486,7
e37
/m54_440
y
8
TG2810,0
p14/m39_263c10,3p25/m45_448c13,2
p11/m49_378y18,7
p11/m49_274c35,2
p11/m49_244y40,0p14/m33_267y45,3p14/m33_273c47,5p14/m39_335c52,9EPMS_31055,7
HPMS1_21486,7
e37
/m54_440
y
8
p25/m42_240y0,0
CT1458,8e37/m54_92c13,7
EPMS_37241,4
e44/m61_545c55,6
e43/m53_146c76,7
e36/m47_153c90,4e36/m47_211c93,4e38/m61_320c100,1e41/m54_110c107,0p14/m33_349c108,3p15/m40_321c109,5p25/m42_683c110,6c33/m49_081c111,9e42/m48_317c114,6e36/m52_136c115,7e44/m61_187y119,2e36/m52_220y121,2e32/m55_191y124,3p14/m41_672y128,3e38/m60_157y132,1e44/m61_480y133,7e44/m61_515y136,2p14/m39_324y139,6e41/m48_120y144,0e41/m54_210y148,7e31/m58_117y154,0e32/m55_392y157,1e31/m58_400y168,5
e43/m54_708c180,5e43/m53_711c185,3e43/m53_451c189,5e36/m52_434c194,0
e41/m61_463c205,2
9
p25/m42_240y0,0
CT1458,8e37/m54_92c13,7
EPMS_37241,4
e44/m61_545c55,6
e43/m53_146c76,7
e36/m47_153c90,4e36/m47_211c93,4e38/m61_320c100,1e41/m54_110c107,0p14/m33_349c108,3p15/m40_321c109,5p25/m42_683c110,6c33/m49_081c111,9e42/m48_317c114,6e36/m52_136c115,7e44/m61_187y119,2e36/m52_220y121,2e32/m55_191y124,3p14/m41_672y128,3e38/m60_157y132,1e44/m61_480y133,7e44/m61_515y136,2p14/m39_324y139,6e41/m48_120y144,0e41/m54_210y148,7e31/m58_117y154,0e32/m55_392y157,1e31/m58_400y168,5
e43/m54_708c180,5e43/m53_711c185,3e43/m53_451c189,5e36/m52_434c194,0
e41/m61_463c205,2
9
e40/m49_247c0,0
e34/m53_145y9,2
e31/m58_367y20,3
e36/m47_145y28,9
e34/m53_228c43,9
e38/m60_117c50,2
e36/m52_132c56,0
e40/m47_552c61,0
e41/m61_267c67,1
e41/m61_266y72,4
c36/m49_282y81,8
e41/m54_137y0,0
e38/m60_264c11,9
p14/m41_215c16,7
e38/m61_403c25,2
ASC00234,6
10a
e40/m49_247c0,0
e34/m53_145y9,2
e31/m58_367y20,3
e36/m47_145y28,9
e34/m53_228c43,9
e38/m60_117c50,2
e36/m52_132c56,0
e40/m47_552c61,0
e41/m61_267c67,1
e41/m61_266y72,4
c36/m49_282y81,8
e41/m54_137y0,0
e38/m60_264c11,9
p14/m41_215c16,7
e38/m61_403c25,2
ASC00234,6
10a
p25/m45_274y0,0
p14/m41_237c6,9
e41/m54_242c19,6e32/m55_079c26,8e40/m49_131y34,1e40/m47_306y43,0e43/m54_428y45,9e38/m60-160c49,6
e43/m53_260c50,4
e41/m61_329c53,3e36/m52_229c56,5EPMS_41063,7e36/m52_335c68,3e41/m61_270c71,6e41/m61_137c74,8e40/m47_223c77,6e33/m56_199c79,3e41/m54_166c82,9e43/m53_121y86,4e40/m47_142y89,1c33/m54_249y91,4e41/m48_253y92,7p25/m45_260y95,6c33/m54_173y104,8
p14/m33_168c124,4
e40/m49_207y0,0
p14/m39_117y7,8
EPMS_44114,7
e41/m54_208y19,9
e44/m51_340c25,6
11a
p25/m45_274y0,0
p14/m41_237c6,9
e41/m54_242c19,6e32/m55_079c26,8e40/m49_131y34,1e40/m47_306y43,0e43/m54_428y45,9e38/m60-160c49,6
e43/m53_260c50,4
e41/m61_329c53,3e36/m52_229c56,5EPMS_41063,7e36/m52_335c68,3e41/m61_270c71,6e41/m61_137c74,8e40/m47_223c77,6e33/m56_199c79,3e41/m54_166c82,9e43/m53_121y86,4e40/m47_142y89,1c33/m54_249y91,4e41/m48_253y92,7p25/m45_260y95,6c33/m54_173y104,8
p14/m33_168c124,4
e40/m49_207y0,0
p14/m39_117y7,8
EPMS_44114,7
e41/m54_208y19,9
e44/m51_340c25,6
11a
CRF2100,0
C2_At3g203906,5
p15/m43_090c11,4e43/m54_27513,2
p17/m32_134y23,1
e44/m51_263c30,9
e41/m61_214c40,2e41/m54_412c44,0c36/m49_353c46,9
p25/m45_087c54,9
e36/m47_237c62,0
e43/m53_241y83,1
e38/m60_186y85,4
12
CRF2100,0
C2_At3g203906,5
p15/m43_090c11,4e43/m54_27513,2
p17/m32_134y23,1
e44/m51_263c30,9
e41/m61_214c40,2e41/m54_412c44,0c36/m49_353c46,9
p25/m45_087c54,9
e36/m47_237c62,0
e43/m53_241y83,1
e38/m60_186y85,4
12
e38/m60_224y0,0p25/m2_131y3,3
LG15
e38/m60_224y0,0p25/m2_131y3,3
LG15
e38/m61_144y0,0
e40/m47_239y9,1
p15/m40_381c15,1p14/m33_851c18,2e43/m54_572c23,1
LG17
e38/m61_144y0,0
e40/m47_239y9,1
p15/m40_381c15,1p14/m33_851c18,2e43/m54_572c23,1
LG17
e31/m58_410y0,0p15/m43_213y1,8
Hpms2_2411,6
LG19
e31/m58_410y0,0p15/m43_213y1,8
Hpms2_2411,6
LG19
e34/m53_181c0,0
p17/m32_155c6,1
p14/m33_206c15,0
LG22
e34/m53_181c0,0
p17/m32_155c6,1
p14/m33_206c15,0
LG22
e32/m55_202c0,0p14/m33_729c2,7ASC0608,2
LG23
e32/m55_202c0,0p14/m33_729c2,7ASC0608,2
LG23
e41/m54_184y0,0
e40/m47_125y9,0
EPMS_37613,6
p11/m49_197y18,7
HPMS1_529,6
Lg24
e41/m54_184y0,0
e40/m47_125y9,0
EPMS_37613,6
p11/m49_197y18,7
HPMS1_529,6
Lg24
p25/m42_436c0,0
HPMS1_6212,4e44/m61_267c17,2e44/m51_646y19,3p15/m40_091c22,1e41/m54_351c26,4
EPMS_64933,3
LG25
p25/m42_436c0,0
HPMS1_6212,4e44/m61_267c17,2e44/m51_646y19,3p15/m40_091c22,1e41/m54_351c26,4
EPMS_64933,3
LG25
e40/m49_305c0,0
e41/m61_169c8,7
EPMS_39115,5
LG27
e40/m49_305c0,0
e41/m61_169c8,7
EPMS_39115,5
LG27
e41/m54_221c0,0
e44/m51_467c5,8
LG28
e41/m54_221c0,0
e44/m51_467c5,8
LG28
e41/m48_070c0,0
p25/m45_434c9,1p25/m45_441y11,9
EPMS_42621,7
LG29
e41/m48_070c0,0
p25/m45_434c9,1p25/m45_441y11,9
EPMS_42621,7
LG29
e41/m48_249c0,0e44/m51_376c1,6e41/m48_150c7,1
LG30
e41/m48_249c0,0e44/m51_376c1,6e41/m48_150c7,1
LG30
p15/m43_178c0,0
p17/m39_330y8,4
LG33
p15/m43_178c0,0
p17/m39_330y8,4
LG33
Fig. 64 Mappa gentica ad alta risoluzione F5YC.
Fig. 65 Mappa genetica ad alta risoluzione F5YC.

IV-RISULTATI
134
p17/m32_344c0,0
p14/m41_132c8,3
LG38EPMS_3770,0
TntC07y9,4
p17/m32_383y24,5
LG39
p11/m49_234c0,0T135E03y2,1p19/m42_197y9,0
LG40
p19/m42_858y0,0p19/m42_874y2,8
LG41
p25/m45_335c0,0
p14/m39_345c7,0
LG42
p14/m39_195c0,0
p11/m49_105y6,5
LG44
c33/m54_118c0,0
EPMS_4029,5
p14/m39_225c20,4
LG45
p14/m33_2780,0p14/m33_294c4,6
p14/m33_268c9,8
LG46
p14/m33_311c0,0p14/m33_313y1,9
LG47
c33/m54_201y0,0
AT110,0
LG48
CD0350,0TD003_2c3,7
LG49
T135E08y0,0
T135E10y8,9
LG50
T135E09c0,0T135E11c1,4
LG51
p15/m43_180y0,0GPMS_1623,3
LG34
p15/m40_200y0,0p15/m40_209c2,1
LG36
p15/m40_319c0,0
EPMS_7558,7
LG37
p17/m32_344c0,0
p14/m41_132c8,3
LG38
p17/m32_344c0,0
p14/m41_132c8,3
LG38EPMS_3770,0
TntC07y9,4
p17/m32_383y24,5
LG39
EPMS_3770,0
TntC07y9,4
p17/m32_383y24,5
LG39
p11/m49_234c0,0T135E03y2,1p19/m42_197y9,0
LG40
p11/m49_234c0,0T135E03y2,1p19/m42_197y9,0
LG40
p19/m42_858y0,0p19/m42_874y2,8
LG41
p19/m42_858y0,0p19/m42_874y2,8
LG41
p25/m45_335c0,0
p14/m39_345c7,0
LG42
p25/m45_335c0,0
p14/m39_345c7,0
LG42
p14/m39_195c0,0
p11/m49_105y6,5
LG44
p14/m39_195c0,0
p11/m49_105y6,5
LG44
c33/m54_118c0,0
EPMS_4029,5
p14/m39_225c20,4
LG45
c33/m54_118c0,0
EPMS_4029,5
p14/m39_225c20,4
LG45
p14/m33_2780,0p14/m33_294c4,6
p14/m33_268c9,8
LG46
p14/m33_2780,0p14/m33_294c4,6
p14/m33_268c9,8
LG46
p14/m33_311c0,0p14/m33_313y1,9
LG47
p14/m33_311c0,0p14/m33_313y1,9
LG47
c33/m54_201y0,0
AT110,0
LG48
c33/m54_201y0,0
AT110,0
LG48
CD0350,0TD003_2c3,7
LG49
CD0350,0TD003_2c3,7
LG49
T135E08y0,0
T135E10y8,9
LG50
T135E08y0,0
T135E10y8,9
LG50
T135E09c0,0T135E11c1,4
LG51
T135E09c0,0T135E11c1,4
LG51
p15/m43_180y0,0GPMS_1623,3
LG34
p15/m43_180y0,0GPMS_1623,3
LG34
p15/m40_200y0,0p15/m40_209c2,1
LG36
p15/m40_200y0,0p15/m40_209c2,1
LG36
p15/m40_319c0,0
EPMS_7558,7
LG37
p15/m40_319c0,0
EPMS_7558,7
LG37
Fig. 66 Mappa genetica ad alta risoluzione F5YC.

IV-RISULTATI
135
6. Identificazione degli individui più informativi.
Al fine di ottimizzare i tempi e i costi necessari per il mappaggio di marcatori
aggiuntivi, sono state identificate tre sottopopolazioni all’interno delle 297 RILs. E’ stato
impiegato a tale scopo il software MapPop.
L’informatività delle sottopopolazioni ottenute è stata testata effettuando un’analisi
molecolare con marcatori aventi localizzazione nota sui cromosomi di peperone.
La sottopopolazione di 141 individui è stata analizzata con 6 marcatori SSRs:
EPMS_649, EPMS_650, EPMS_704, EPMS_725, EPMS_747 e EPMS_755 e col marcatore
STS ASC002 legato al gene di resistenza pvr4. Tutti questi marcatori sono stati assegnati ai
gruppi linkage previsti, mediante l’impiego del software MAPMAKER EXP 3.0. I 6 marcatori
SSR sono stati assegnati e ordinati con il comando “try”; il marcatore STS ASC002 è stato
assegnato al cromosoma 10 della “core map”; l’ordine rispetto agli altri marcatori è stato
confermato col comando “ripple” a LOD score di 3.0.
Anche la sottopopolazione di 93 individui è stata analizzata con i marcatori sopra
elencati, per valutare la sua rappresentatività rispetto all’intera popolazione. I marcatori
molecolari, anche in questo caso, sono stati assegnati ai LGs corretti, ma con una precisione
di mappaggio inferiore; infatti, utilizzando il comando “try” per attribuire i marcatori ai LGs,
il software non identifica una posizione unica, ma diverse posizioni probabili per ciascun
marcatore.
La sottopopolazione di 29 individui, identificata con MapPop, è stata testata per
valutare il numero minimo di campioni necessari in un’analisi molecolare, al fine di poter
assegnare correttamente marcatori aggiuntivi ai LGs di una “core map” precedentemente
sviluppata. La rappresentatività della sotto-popolazione è stata valutata testando 13 marcatori
molecolari (12 AFLPs e 1SSR), precedentemente assegnati ai 12 cromosomi di peperone. I
dati molecolari originali, relativi alla popolazione RIL totale, sono stati sostituiti coi dati
ottenuti dall’impiego di tali marcatori sulla sottopopolazione di 29 individui. Dieci marcatori
su 13, 9 AFLPs e 1 SSR, sono stati mappati sui corretti LGs, mediante l’impiego del software
MAPMAKER EXP/3.0, a LOD di 3.0 e percentuale di ricombinazione 0.1. E’ stato impiegato
un valore di frequenza di ricombinazione stringente per garantire un’alto livello di precisione
nell’identificazione dei gruppi linkage, il valore di LOD è stato invece ridotto a 3.0, in quanto
la sotto-popolazione analizzata è caratterizzata da un basso numero di individui.

V-DISCUSSIONE
V. DISCUSSIONE
Negli ultimi 15 anni sono state sviluppate mappe genetico-molecolari per la maggior
parte delle specie coltivate (Paterson et al. 2000). Queste mappe sono state impiegate per
effettuare studi di sintenia, gene tagging e per clonare geni d’interesse (Paran et al. 2004). La
disponibilità di mappe genetico-molecolari sature, inoltre, è un prerequisito indispensabile per
l’applicazione di programmi di miglioramento genetico basati sulla MAS (selezione assistita
da marcatori).
In peperone sono state sviluppate numerose mappe genetico-molecolari sia inter- che
intra-specifiche, ma non è ancora stata raggiunta la completa saturazione del genoma.
Nell’ambito del genere Capsicum, infatti, sono stati riscontrati due problemi principali che
limitano/ostacolano/frenano il mappaggio: i) il limitato (basso) livello di polimorfismo
osservato nelle progenie segreganti ottenute da incroci intra-specifici; ii) la segregazione
distorta dei marcatori molecolari, in popolazioni ottenute da incroci inter-specifici, che riduce
considerevolmente la precisione nel mappaggio.
L’obiettivo di questo lavoro di tesi è stato di contribuire allo sviluppo di una mappa
genetica intra-specifica di peperone ad alta risoluzione, cioè caratterizzata da un’alta densità
di marcatori molecolari. A tale scopo è stata impiegata una popolazione di mappaggio
costituita da 297 RILs. Un fattore fondamentale che condiziona lo sviluppo di una mappa fine
di un genoma è, non solo la scelta di una popolazione idonea, ma anche di un ampio numero
di marcatori molecolari, che siano il più possibile distribuiti uniformemente nel genoma.
Per la costruzione della mappa genetica di peperone sono stati utilizzati 595 marcatori
molecolari, tra cui RFLPs, AFLPs, SSAP, STS e, nel lavoro di tesi, marcatori SSR, che
risultano particolarmente utili in quanto co-dominanti, altamente polimorfici e facili da
analizzare mediante amplificazione PCR (Lee et al. 2004). I marcatori SSR utilizzati sono
stati in parte ottenuti dalla letteratura (Lee et al. 2004; Sasvári et al. 2004) ed in parte isolati
da sequenze EST di Capsicum reperibili in database EMBL. Tutti i marcatori SSR rivelatisi
polimorfici nella popolazione RIL, e cioè: 20 SSR di Sasvári et al. (2004), 9 SSR di Lee et al.
(2004) e 13 SSR sviluppati da EST, sono stati impiegati per lo sviluppo della mappa genetico-
molecolare.
Inoltre, è stato sviluppato un marcatore STS, con strategia dCAPs, legato alla
resistenza ai potyvirus, con lo scopo di favorire l’applicazione futura di programmi di MAS
per il trasferimento del carattere a cultivar commerciali.

V-DISCUSSIONE
137
Infine, per ridurre i tempi e i costi relativi all’analisi molecolare è stata validata la
possibilità di impiegare la strategia del mappaggio selettivo.

V-DISCUSSIONE
138
1. Popolazione segregante RILs:F5.
La popolazione segregante intra-specifica (RILs:F5), impiegata per lo sviluppo della
mappa genetica, è stata prodotta mediante 4 generazioni di autofecondazione di un ibrido F1,
ottenuto dall’incrocio tra l’accessione commerciale “Yolo Wonder” e l’esotica “Criollo de
Morellos”, appartenenti alla specie C. annuum.
La progenie RILs è vantaggiosa nel mappaggio perché, essendo sviluppata da un
incrocio intra-specifico, rispecchia il gene-pool di peperone utilizzato dai breeders nei
programmi di selezione (Lefebvre et al. 2004). L’impiego di una popolazione intra-specifica
consente, inoltre, di ridurre la probabilità di segregazioni distorte dei marcatori molecolari e
quindi di incrementare la precisione del mappaggio. Lo sviluppo della mappa ha richiesto un
ampio lavoro di genotyping a causa del basso livello di polimorfismi rilevabili, dovuti alla
limitata differenziazione genetica dei parentali e quindi alla ridotta segregazione nella
popolazione di mappaggio. E’ stata sviluppata, pertanto, una popolazione di 297 RILs che è
stata testata con un elevato numero di marcatori molecolari (595), incrementando così la
possibilità di identificare polimorfismi. La popolazione RIL offre anche il vantaggio di poter
essere mantenuta nel tempo e, pertanto, rende possibile aggiungere alla mappa nuovi
marcatori, in tempi successivi, sino a giungere alla sua completa saturazione. Inoltre, è
possibile analizzare la popolazione segregante in ambienti diversi, ciò permette di identificare
con maggiore efficienza i quantitative traits loci (QTLs). I QTLs hanno infatti un ruolo
chiave nella biologia della specie, in quanto regolano caratteri d’interesse agronomico e
caratteri che conferiscono resistenza a malattie, i.e: nei confronti di Phytophthora spp,
Verticillium spp e Nematodi (Lefebvre e Palloix, 1996; Thabuis et al. 2003).

V-DISCUSSIONE
139
2. Marcatori molecolari e costruzione della mappa
genetica.
La mappa genetica di peperone è stata sviluppata utilizzando 595 marcatori molecolari:
507 AFLPs, 18 RFLPs, 20 SSAPs, 12 STSs e 38 SSRs. L’elevato numero di marcatori e il
tipo di popolazione impiegati in questo studio hanno permesso di ottenere una mappa ad alta
risoluzione ed elevato grado di confidenza nella localizzazione dei marcatori, di gran lunga
superiore alle mappe finora sviluppate in Capsicum.
La prima fase del lavoro è consistita nello sviluppo di una “core map”, mappa genetico-
molecolare che rappresenta lo scheletro base della mappa ad alta risoluzione e comprende il
54% (325) dei marcatori molecolari totali. Questi marcatori, detti framework markers,
comprendono tutti i marcatori molecolari che risultano assemblati in gruppi linkage alle
condizioni di massima stringenza (LOD> 8.0 e frequenza di ricombinazione minima di 0.1)
ed il cui ordine sui LGs è stato confermato, col comando “ripple”, a valori di LOD≥ 3.0. I
marcatori nella “core map” sono generalmente ben distanziati tra loro; tuttavia, è stata
osservata la presenza di raggruppamenti in clusters sui cromosomi 3, 5, 9 e 11. Il “clustering”
è un evento comune e sembra verificarsi più frequentemente, ma non in via esclusiva, a
livello delle regioni centromeriche, probabilmente in quanto caratterizzate da una minore
percentuale di ricombinazione (Boersma et al. 2005). La “core map” è stata il punto di
partenza per lo sviluppo della mappa F5YC finale.
Il lavoro di tesi è consistito, in particolare, nell’isolamento e nel mappaggio di marcatori
molecolari di tipo SSR.
I marcatori SSR sviluppati sono stati isolati da sequenze ESTs e pertanto rilevano i
polimorfismi presenti in regioni espresse del genoma; questa peculiarità offre la potenziale
opportunità di impiegare i marcatori in specie diverse, o come “anchor marker” per effettuare
analisi di sintenia o, ancora, per effettuare studi di variabilità genetica. Nel lavoro di tesi,
infatti, i marcatori EST-SSR sviluppati sono stati anche impiegati per valutare il grado di
similarità genetica esistente tra accessioni differenti di Capsicum, ed è stato ottenuto un
dendogramma che evidenzia le relazioni filogenetiche tra esse.
Per quanto riguarda le caratteristiche dei motivi microsatellite identificati, le ripetizioni
trimeriche sono risultate più frequenti rispetto a quelle dimeriche, in accordo con quanto
riportato da studi precenti (Nagy et al. 2004; Portis et al. 2006) effettuati in diverse specie
coltivate. Quanto osservato è presumibilmente dovuto al fatto che, nelle sequenze codificanti,

V-DISCUSSIONE
140
si assiste al fenomeno della soppressione dei motivi ripetuti non trimerici, che permette di
evitare il rischio di slippage dell’enzima polimerasi e la conseguente produzione di mutazioni
frameshift indotte dalla presenza di motivi microsatellite dinucleotidici alternati e separati tra
loro da poche basi (bp).
Nei genomi vegetali sembrano prevalere i microsatelliti con ripetizioni AT (Lagercrantz
et al. 1993; Morgante e Olivieri 1993) mentre le ripetizioni CA/GT, che sono abbondanti nei
genomi dei mammiferi, sono in genere meno frequenti e meno polimorfiche (Powell et al.
1996). In Arabidopsis, il motivo CA/GT è poco rappresentato (Depeiges et al. 1995) e mostra
bassa variabilità. Nella barbabietola da zucchero il motivo GT fa parte di elementi ripetuti
complessi, presenti in copie multiple a livello dei centromeri, quindi poco utili nello sviluppo
di mappe genetico-molecolari (Schmidt e Heslop-Harrison 1996). Nonostante queste
evidenze, sono stati sviluppati con successo marcatori SSR con motivo CA/GT in mais (Chin
et al. 1996), grano (Röder et al. 1998) riso (Temnykh et al. 2000) e anche quelli impiegati per
l’analisi molecolare in peperone mostrano tale ripetizione (Nagy et al. 1998; Lee et al. 2004).
Il motivo TTG è molto abbondante in peperone, nonostante sia raro in altre specie vegetali
(Morgante et al. 2002) e potrebbe pertanto risultare di interesse effettuare studi di sintenia per
valutare se tale motivo è presente con elevata frequenza anche in altri membri della famiglia
delle Solanaceae, quali melanzana, patata, pomodoro.
Gli SSR testati, sviluppati da Sasvári et al. (2005) e da Lee et al. (2004), mostrano
motivi sia di tipo CA/GT, sia TTG. I marcatori sviluppati durante il lavoro di tesi evidenziano
un’elevata frequenza del motivo AG/CT, seguito da AC/GT e AT/AT, mentre il motivo
CG/CG non è stato identificato.
Dei 38 marcatori SSR polimorfici, testati nella popolazione RIL:F5, 25 sono stati
assegnati ai gruppi linkage della mappa e sono stati impiegati per valutare il grado di
eterozigosi nella popolazione.
Nell’ambito del lavoro di tesi è stato anche sviluppato il marcatore STS ASC002,
associato alla resistenza a pvr4 mediante strategia dCAPs. ASC002 è stato mappato sul
cromosoma 10, a conferma di quanto ottenuto precedentemente da Lefebvre et al. (2002); ciò
offre prospettive molto utili nell’ambito della selezione assistita da marcatori. Il risultato
dimostra che la strategia dCAPs può essere sfruttata per sviluppare marcatori molecolari
associati a geni di interesse agronomico, qualora siano caratterizzati dalla presenza di
polimorfismi di tipo SNPs, peraltro piuttosto frequenti nei genomi eucarioti.

V-DISCUSSIONE
141
Nello sviluppo della mappa F5YC, a differenza di altre mappe genetiche sviluppate, non
sono stati impiegati marcatori RAPD. La scelta di non utilizzarli è dovuta al fatto che,
nonostante questi marcatori siano in grado di rilevare un alto livello di polimorfismo e siano
di facile impiego, hanno una bassa riproducibilità e limitano quindi la precisione nello
sviluppo delle mappe. Nonostante le analisi di segregazione permettano di identificare le
bande RAPD utili per il mappaggio, numerosi studi (Williams et al. 1990; Arnold et al. 1991;
Carlson et al. 1991; Ellsworth et al. 1993 e Schweder et al. 1995) dimostrano che i
polimorfismi RAPD sono sensibili a lievi modificazioni nelle condizioni di amplificazione e
Jones et al. (1997) riportano che utilizzando la stessa serie di marcatori molecolari RAPD in
laboratori differenti, non si ottengono pattern elettroforetici identici. Inoltre, è stata
evidenziata la possibilità di osservare bande RAPD, conseguenti alla formazione di
heteroduplex tra i prodotti di amplificazione (Ayliffe et al. 1994).
La maggior parte dei polimorfismi osservati nelle analisi molecolari sono generati dai
marcatori AFLP, che rappresentano l’85% dei marcatori totali. Nonostante questo, l’impiego
di marcatori RFLP, SSR e STS è stato fondamentale per assegnare i LGs identificati ai
cromosomi di Capsicum e per allineare la mappa F5YC a quelle precedentemente sviluppate
da Lefebvre et al. (2002) e Paran et al. (2004).
I 20 marcatori SSAP utilizzati sono risultati tutti polimorfici: 11 sono stati assemblati in
LGs e assegnati ai cromosomi, 1 non è stato assegnato ad alcun gruppo di associazione. Due
marcatori SSAP: Tntc09y e Tntc01c sono stati inseriti nella “core map”.
Circa il 14 % dei marcatori molecolari testati hanno evidenziato segregazione distorta,
cioè rapporto fenotipico che si discosta da quello atteso e che è a favore di uno dei due
parentali. Tale percentuale è superiore ripetto a quella osservata nello sviluppo di altre mappe
intra-specifiche; e cioè del 7% di quella di Lefebvre et al. (2002), e del 10% di quella di
Sugita et al. (2005); ma è significativamente inferiore ai valori riportati nelle mappe inter-
specifiche di Kang et al. (2001) e di Livingstone et al. (1999), pari a 28.8% e 50.7%
rispettivamente. Le percentuali di segregazione distorta risultano pressochè analoghe nelle
diverse classi di marcatori molecolari: 15% per gli AFLP, da 11% a 15% per gli RFLP e gli
SSAP, ad eccezione dei marcatori SSR che non hanno manifestato distorsioni. La causa di
tale segregazione può essere attribuita ad errori nel genotyping o nella lettura dei dati
(scoring), ma è principalmente conseguenza di fattori biologici quali: i) perdita di cromosomi,
ii) associazione non random dei gameti, iii) diverso grado di sopravvivenza degli zigoti, iv)
modificazioni nel flusso genico (Bradshaw e Stettler, 1994) o presenza di alleli nulli
(Pekkinen et al. 2005). Poiché, studi precedenti (Kuang et al. 1999) hanno dimostrato che i

V-DISCUSSIONE
142
marcatori distorti possono risultare utili nell’identificazione dei gruppi linkage, si è deciso di
includere tutti questi marcatori nelle analisi di associazione. E’ stato anche osservato che il
maggior numero dei marcatori che mostrano segregazione distorta sono stati mappati ad un
valore di LOD score pari a 3, in due regioni specifiche dei cromosomi 4 (C36/M49_216c,
P14/M41_648c, P15/M40_152y, P17/M32_240c, P25/M45_132c) e 10 (E34/M53_228c,
E38/M60_117c, E38/M61_414c, E40/M47_552c, E41/M61_267c) (Fig. 66).
Le due regioni sono risultate caratterizzate dalla presenza di marcatori distorti a favore
del parentale Criollo de Morellos 334. Ciò conferma che il fenomeno della segregazione
distorta sia causato da un meccanismo biologico, piuttosto che dall’accumulo di errori di
scoring o da effetti casuali (Fishman et al. 2001).

V-DISCUSSIONE
143
Fig. 66 Regioni specifiche dei cromosomi 4 e 10, ricche di marcatori molecolari distorti.
c36/m49_20,
p15/m40_130,
p25/m45_138,p14/m39_442,GC20744,e41/m48_249,e36/m52_356,
e38/m60_163,
e41/m48_070,
EPMS_376,p17/m32_280,p14/m41_685,p25/m45_190,Tntc093,
TG132102,
p25/m45_1111,
e41/m48_0148,
e38/m61_168c
156,e38/m61_158y
161,
e42/m48_1170,e41/m48_1172,
e37/m54_
p15/m43_10,p15/m43_13,
CT224
11,
4a
e40/m49_20,
e34/m53_19,
e31/m58_320,
e36/m47_128,
e34/m53_243,
e38/m60_150,
e36/m52_156,e40/m47_561,
e41/m61_267c
67,e41/m61_266y
72,
c36/m49_281,
e41/m54_10,
e38/m60_211,p14/m41_216,
e38/m61_425,
ASC00234,
10a
4b
10b

V-DISCUSSIONE
144
Per garantire un elevato grado di confidenza nella posizione dei marcatori, sono stati
impiegati valori soglia di LOD score > 8.0 e percentuale di ricombinazione massima di 0.1.
Queste condizioni hanno permesso di identificare 49 LGs, 21 dei quali sono caratterizzati da
un basso numero di marcatori. In seguito al mappaggio di nuovi marcatori molecolari tali LGs
minori potranno essere associati ad altri gruppi maggiori, colmando così i “gap” ancora
presenti nella mappa. L’elevato numero di LGs ottenuto è dovuto alla scelta di utilizzare
condizioni stringenti; valori soglia meno restrittivi avrebbero permesso di ottenere un minor
numero di LGs, corrispondente al numero dei cromosomi attesi, ma la precisione del
mappaggio sarebbe stata sensibilmente inferiore. Nelle mappe di peperone sviluppate in
precedenza (Livingstone et al. 1999; Kang et al. 2001; Paran et al. 2004; Sugita et al. 2005 e
Ogundiwin et al. 2005) sono stati infatti utilizzati valori di LOD score variabili tra 3 e 4.5 e
frequenza di ricombinazione massima variabile tra 20 e 40 cM. L’unica mappa di peperone
sviluppata con condizioni analoghe a quelle applicate nel nostro lavoro, è stata quella
pubblicata da Lefebvre et al. (2002).

V-DISCUSSIONE
145
3. Allineamento e confronti con mappe precedentemente
sviluppate
La mappa intraspecifica F5YC è stata allineata alla mappa precedentemente pubblicata
da Lefebvre et al. (2002) e a quella integrata, pubblicata da Paran et al. (2004). Sulla base dei
marcatori comuni, “anchor marker”, 22 gruppi linkage sono stati assegnati a specifici
cromosomi di peperone.
Sono stati identificati marcatori AFLP comuni alle tre mappe, che risultano localizzati
nella stessa posizione; questo fatto conferma l’ipotesi che siano locus-specifici, come
precedentemente riportato da altri autori (Lefebvre et al. 2002).
L’allineamento alle mappe precedentemente sviluppate è stato possibile per tutti i
cromosomi, eccetto per i cromosomi 1 e 8. Il cromosoma 1 della mappa integrata di Paran et
al. (2004) risulta separato in due LGs sia nella mappa integrata di Lefebvre et al. (2002), che
nella mappa sviluppata in questo lavoro di tesi. Il LG1 e il LG8 della presente mappa F5YC
sono co-lineari alle estremità del LG1 della mappa integrata di Paran et al. (2004).
Reciprocamente, il cromosoma 8 della mappa integrata risulta co-lineare al cromosoma 1
della mappa intra-specifica di Lefebvre et al. (2002), sebbene non sia stato possibile allineare
ad esso alcuno dei LGs della mappa F5YC, in quanto non sono risultati disponibili “anchor
marker”. I risultati ottenuti, come evidenziato in precedenza da Pickersgill et al. (1991),
confermano che le traslocazioni cromosomiche reciproche avvenute tra C. annuum e C.
chinense sono responsabili di pseudo-linkage tra i marcatori a livello del punto di inter-
scambio, che sembra essere localizzato approssimativamente a 75 cM dal cromosoma 1 della
mappa integrata di Paran et al. (2004), e che corrisponde ad un’estremità del cromosoma 8
nelle mappe intra-specifiche.
La lunghezza totale del genoma di peperone è stimata tra 1498 cM e 2268 cM e la
lunghezza della mappa F5YC è di 1857 cM; il valore ottenuto è simile a quello riportato nella
mappa integrata di Paran et al. (2004) pari a 1832 cM, e maggiore rispetto ai 1466 cM della
mappa recentemente sviluppata da Ogundiwin et al. (2005) a partire da una popolazione RILs.
Mappe precedenti, basate su popolazioni F2 (Sugita et al. 2005) e DH (Ogundiwin et al. 2005)
coprono, rispettivamente, una lunghezza di 1043 e 1089 cM; la mappa genetica sviluppata da
Lefebvre et al. (2002) copre solo 685 cM, in quanto è stato impiegato un basso numero di
marcatori molecolari. La mappa F5YC sviluppata risulta dunque particolarmente valida nel
simulare l’assetto cromosomico di peperone e può essere la base per un ulteriore

V-DISCUSSIONE
146
arricchimento di marcatori molecolari al fine di saturare completamente il genoma di
Capsicum e fornire così grossi input al miglioramento genetico della specie.
La mappa F5YC sviluppata è costituita da 9 marcatori RFLP, (CT080, CT145, CT224,
CD035, TG29, TG123, TG132, TG281 e C2_At3g20390), comuni alla mappa di pomodoro,
sviluppata in precedenza. Nonostante il numero di “anchor markers” sia esiguo, è stato
possibile effettuare una, per quanto approssimativa, valutazione della sintenia tra le due
mappe genetiche. Il cromosoma 1 di peperone risulta corrispondere al cromosoma 1 di
pomodoro, il 2 al 2; il cromosoma 4 di peperone è risultato in parte co-lineare al cromosoma 4
ed in parte al 3 di pomodoro. Il 5 è ortologo ai cromosomi 5 e 4 di pomodoro, mentre l’8 è
ortologo all’estremità superiore del cromosoma 1. Infine, il cromosoma 9 contiene il
marcatore RFLP CT145, che è stato mappato sul cromosoma 9 da Lefebvre et al. (2002) e sul
cromosoma 5 da Tanksley et al. (1992). L’allineamento effettuato è poco informativo ma
permette di confermare quanto già osservato negli studi più dettagliati già effettuati
(Livingstone et al. 1999; Lefebvre et al. 2002).

V-DISCUSSIONE
147
4. Identificazione degli individui RILs più informativi e
mappaggio selettivo.
Le mappe genetico-molecolari precedentemente sviluppate in Capsicum sono state
ottenute dall’analisi di un numero di individui variabile da 72, nel caso di progenie F2
(Livingstone et al. 1999), a 176 nel caso di progenie DH (Sugita et al. 2005). E’ importante
tener presente che, come riportato da Isidore et al. (2003), più numerosa è la popolazione di
mappaggio, maggiore è il numero di prodotti di meiosi analizzati e, maggiore è il numero di
marcatori molecolari testati, migliore è la risoluzione della mappa genetica. Per queste ragioni
la mappa F5YC è stata sviluppata analizzando 297 RILs. L’elevato numero degli individui da
genotipizzare ha tuttavia comportato tempi e costi notevoli; pertanto è stato utilizzato il
software MapPop (Brown e Vision, 2000), al fine di identificare all’interno delle 297 RILs,
gli individui più rappresentativi da utilizzare per studi fenotipici e di mappaggio futuri. Il
software, basandosi sui dati relativi alla mappa già sviluppata, permette di identificare le
sotto-popolazioni di individui più significativi per il mappaggio. Sono state individuate tre
sotto-popolazioni, costituite rispettivamente da 141, 93 e 29 individui ed è stata valutata la
loro attendibilità nell’attribuire i marcatori a specifici LGs. Le popolazioni di 141 e 93
individui hanno dimostrato di garantire una buona precisione di mappaggio, che risulta
maggiore per la prima. La sotto-popolazione di 29 individui ha permesso di assegnare
correttamente 10 marcatori su 13 ai LGs attesi, nonostante la loro localizzazione sul gruppo
risulti imprecisa. Questo sottoinsieme, molto piccolo, è stato prodotto per valutare il numero
minimo di campioni necessari in un’analisi molecolare per poter assegnare correttamente,
marcatori aggiuntivi ai LGs di una mappa precedentemente sviluppata. Lo scopo principale è
quello di impiegare la sotto-popolazione in rapide analisi di screening per identificare nuovi
marcatori molecolari e valutare a quale LGs vengono assegnati. I marcatori che risulteranno
localizzati su gruppi linkage di interesse, ad esempio quelli strategici per completare la
saturazione della mappa genetica, verranno impiegati per testare la sotto-popolazione di 141
individui. I risultati ottenuti confermano che il mappaggio selettivo può essere considerato
una strategia importante per lo sviluppo di mappe genetico-molecolari sature, e anche
un’alternativa per il mappaggio di QTLs e per effettuare test fenotipici, come
precedentemente riportato da Xu et al. (2005) e Howad et al. (2005) in Prunus.

VI-CONCLUSIONI
VI. CONCLUSIONI
Questo lavoro di tesi ha contribuito allo sviluppo della mappa genetico-molecolare
F5YC, che risulta la mappa intra-specifica a più alta risoluzione tra quelle finora sviluppate in
Capsicum. Inoltre, ha permesso lo sviluppo di 49 marcatori SSR, derivati da EST, utili in
studi di mappaggio nell’ambito del genere Capsicum.
La mappa genetica ottenuta dimostra la validità della scelta di una popolazione RILs
numerosa e di un elevato numero di marcatori molecolari per le analisi di genotyping.
I risultati riportati evidenziano l’utilità dell’impiego di marcatori SSR nel mappaggio
dovuto al loro alto livello di polimorfismo, alla loro dispersione uniforme nel genoma e al
fatto che, a differenza dei marcatori AFLP, non producono cluster. Inoltre, i marcatori SSR,
poiché co-dominanti, hanno permesso di valutare il grado di eterozigosi della popolazione in
esame e possono essere utili nel valutare le relazioni filogenetiche esistenti tra diverse
accessioni, appartenenti ad una stessa specie o tra specie diverse, appartenenti ad uno stesso
genere.
Il corretto mappaggio del marcatore ASC002 evidenzia la possibilità di impiegare la
strategia dCAPs per sviluppare marcatori molecolari di tipo STS, legati a geni di interesse. La
tecnica dCAPs permette infatti di evidenziare polimorfismi SNPs, esistenti a livello di geni
coinvolti nella manifestazione di caratteri d’interesse agronomico, quale in questo caso la
resistenza nei confronti dei potyvirus, e di incrementare il numero di marcatori disponibili per
la MAS.
Infine, l’analisi delle sotto-popolazioni ottenute mediante l’impiego del software
Map.Pop ha permesso di dimostrare l’efficacia del mappaggio selettivo, strategia che potrà
essere impiegata per raggiungere la completa saturazione del genoma e l’obiettivo futuro di
localizzare QTLs d’interesse agronomico nella mappa F5YC, studio che è già in corso di
sviluppo presso il settore di genetica del DI.VA.PRA dell’Università degli Studi di Torino e
che è condotto in collaborazione con l’INRA (Montflavet-Francia).

VII-BIBLIOGRAFIA
VII. BIBLIOGRAFIA
Acquadro A., Portis E. e Lanteri S., (2003). “Isolation of microsatellite loci in artichoke
(Cynara cardunculus L. var. scolymus).” Molecular Ecology Notes, 3: 37-39.
Anderson J.A., Churchill G.A., Autrique J.E., Tanksley S.D. e Sorrells M.E., (1993).
“Optimizing parental selection for genetic linkage maps.” Genome, 36: 181-6.
Andrews J., (1984). “Peppers: the Domesticated Capsicums.” University of Texas
Press, Austin, Texas.
Anghiera P. M., (1493). “Decadas de nuevo mundo.” Cited in Andrews, 1984.
Arnold M.L., Buckner C.M., Robinson J.J., (1991). “Pollen-mediated introgression and
hybrid speciation in Louisiana Irises.” Proc Natl Acad Sci, USA, 88: 1398-1402.
Arumuganathan K. e Earle E. D. (1991). “Nuclear DNA content of some important
plant species.” Plant Mol Biol Rep, 9: 208-219.
Ayliffe A.A., Lawrence G.J., Ellis J.G. e Pryor A.J., (1994). “Heteroduplex molecules
formed between allelic sequences cause nonparental RAPD bands.” Nucl Acids Res, 22:
1632–1636.
Barakat A., Carels N. e Bernardi G., (1997). “The distribution of genes in the genomes
of the Gramineae.” Proc. Natl. Acad. Sci, USA, 94: 6857–6861.
Barcaccia G., Lucchin M., Parrini P. (2000). “Analisi del genoma mediante marcatori
molecolari: II. Principali applicazioni.” Riv. Di Sem. El., 5: 17-34.
Bassam B. J., Caetano-Anolles G., Gresshoff P. M., (1991). “Fast and sensitive silver
staining of DNA in polyacrylamide gels.” Analytic Biochemistry, 19: 680-683.

VII-BIBLIOGRAFIA
150
Basten C.J., Weir B.S. e Zeng Z.B., (1997). “QTL CARTOGRAPHER: a reference
manual and tutorial for QTL mapping.” Department of Statistics, North Carolina State
University, Raleigh ùn.C..
Ben Chaim A., Grube R., Lapidot M., Jahn M. e ., Paran I., (2001b). “Identification of
quantitative trait loci associated with resistance to cucumber mosaic virus in Capsicum
annuum.” Theor. Appl. Genet., 102: 1213-1220.
Ben Chaim A., Paran I., Grube R., Jahn M., Van Wijk R. e Peleman J., (2001a). “QTL
mapping of fruit related traits in pepper (Capsicum annuum).” Theor. Appl. Genet., 102:
1016-1028.
Boersma J.G., Pallotta M., Li C., Buirchell B.J., Sivasithamparam K., Yang H. (2005)
“Construction of a genetic linkage map using MFLP and identification of molecular markers
linked to domesticatoion genes in narrow-leafed lupin (Lupinus angustifolius L.).” Cellular &
Molecular Biology Letters, 10: 331-344.
Bosland P.W. (1992) “Chiles: a diverse crop.” HortTechnology, 2 (1): 6-10.
Bosland P.W. e Votava E.J., (2000). “Peppers: vegetable and spice Capsicums.”,
Departement of Agronomy and Horticulture, New Mexico State University, Las Cruces,
USA. CABI publishing.
Bostein D., White R.L., Skolnick M., Davis R.W., (1980). “Construction of a genetic
linkage map in man using restriction fragment lenght polymorphisms.” American Journal
Human Genetics, Chicago, vol.32, 3: 314-331.
Bowcock A.M., Ruizlinares A., Tomfohrde J., Korol A., Schulman A.H. e Gill B.S.,
(1994). “High-resolution of human evolutionary trees with polymorphic microsatellites.”
Nature, 368 (6470): 455-457.
Bradshaw Jr. H.D. e Stettler R.F., (1994). “Molecular genetics of growth and
development in Populus. II. Segregation distortion due to genetic load.” TAG Theoretical and
Applied Genetics, Vol. 89, Number 5: 551–558.

VII-BIBLIOGRAFIA
151
Brown D. G., Vision T. J., Tanksley S.D., (2000). “Selective mapping: a discrete
optimization approach to selecting a population sample for use in a high-density linkage
mapping project.” Proceedings of the 11th SIAM-ACM Symposium on Discrete Algorithms.
Brown D. e Vision T., (2000). “MapPop 1.0: Software for selective mapping and bin
mapping.” http://www.bio.unc.edu/faculty/vision/lab/mappop/.
Buttery R.G., Seifert R.M., Guadagni D.G., Ling L.C., (1969). “Characterization of
some volatile constituents of bell peppers.” Journal of Agriculture, Food and Chemistry, 17:
1322-1328.
Caranta C., Thabuis A. e Palloix A., (1999). “Development of a CAPS marker for Pvr4
locus: Q tool for pyramiding potyvirus resistance genes in pepper.” Genome, 42: 111-116.
Carlson J.E., Tulsieram L.K., Glaubitz J.C., Luk V., Kauffeldt C., Rutledge R., (1991).
“Segregation of random amplified DNA markers in F1 progeny of conifers.” Theor Appl
Genet, 83: 194-200.
Carmichael J.K., (1991). “Treatement of herpes Zoster and postherpetic neuralgia.”
American Family Physician, 44 (1): 203-210.
Cheng S.S., (1989). “The use of Capsicum chinense as sweet pepper cultivars and
source for gene transfer.” In: Griggs, T.D. e McLean B.T. (eds.) Tomato and Pepper
Production in the Tropics. Asian Vegetable Research & Developement Center, Taiwan, pp.
55-62.
Chin E.C.L., Senior M.L., Shu H., Smith J.S.C., (1996). “Maize simple repetitive DNA
sequences: abundance and allele variation.” Genome, 39: 866-873.
Cichewicz R.H., Thorpe P.A., (1996). “The antimicrobical properties of chile peppers
(Capsicum species) and their use in Mayan medicine.” Journal of Ethnopharmacology, 52:
61-70.

VII-BIBLIOGRAFIA
152
Collins M.D., Wasmund L.M., Bosland P.W., (1995). “Improved method for
quantifying capsaicinoids in Capsicum using high-performance liquid chromatography.”
HortScience, 30 (1): 137-139.
Csillery G., Quagliotti L., Rota A., (1986). “Natural cross-pollination experiment on
pepper (Capsicum annuum L.) in Piedmont, Italy, in 1986.” Capsicum Newsletter, 5: 38-39.
Depeiges A., Goubely C., Lenoir A., Cocherel S., Picard G., Rayanal M., Grellet F.,
Delseny M., (1995). “Identification of the most represented repeated motifs in Arabidopsis
thaliana microsatellite loci” Theor Appl Genet, 91: 160-168.
Edwards A., Civitello H., Hammond H.A., Caskey C.T., (1991). “DNA typing and
genetic mapping with a trimeric tetrameric tandem repeat.” Am. J. Hum. Genet., 49: 746-756.
Ellsworth, D.L., Rittenhouse, K.D. and Honeycutt, R.L. (1993). “Artifactual variation in
randomly amplified polymorphic DNA banding patterns.” Biotechniques, 2: 214–7.
Franceschetti U., (1971). “Natural cross-pollination in pepper (Capsicum annuum).”
Eucarpia Meeting “Genetic and Breeding of Capsicum”, Torino, 346-353.
Fishman L., Kelly A.J., Morgan E., Willis J.H., (2001). “A genetic map in the Mimulus
guttatus species complex reveals transmission ratio distortion due to heterospecific
interactions.” Genetics 159: 1701–1716.
Galbraith D. W., Harkins K. R., Maddox J. M., Ayres N. M., Sharma D. P., Firoozabaty
E. (1983). “Rapid flow cytogenetic analysis of the cell cycle in intact plant tissues.” Science,
220: 1049-1051.
Grattapaglia D. e Sederoff R., (1994). “Genetic-linkage maps of Eucalyptus grandis and
Eucalyptus urophylla using a pseudo test-cross-mapping strategy and RAPD markers.”
Genetics, 137 (4): 1121-1137.
Haanstra J. P. W., Wye C., Verbakel H., Meijer-Dekens F., van der Berg P., van
Heusden A.W., Tanksley S., Lindhout P. e Peleman J., (1999). “An integrated high-density

VII-BIBLIOGRAFIA
153
RFLP-AFLP map of tomato based on two Lycopersicon esculentum x L. pennellii F2
populations”. Theor. Appl. Genet., 99: 254-271.
Haldane J. B. S., (1919). “The mapping function” J. Genet., 8: 299-309.
Harkay-Vinkler M., (1974). “Storage experiments with raw material seasoning paprika,
with particular reference to the red pigment components.” Acta Alimentaria Academiae
Scientarium Hungaricae, 3: 239-249.
Haymon L.W., Aurand L.W., (1971). “Volatiles constituents of Tabasco peppers.”
Journal of Agriculture, Food Chemistry 19, 1131-1134.
Heiser C.B. (1976). “Pepper Capsicum (Solanaceae).” In: Simmonds, N.W. (ed.)
Evolution of Crop Plants. Longman Press. London, pp.265-268.
Howad W., Yamamoto T., Dirlewanger E., Testolin R., Cosson P., Cipriani G.,
Monforte A. J., Georgi L., Abbott A. G. and Arus P., (2005). “Mapping with a few plants:
using selective mapping for microsatellite saturation of the Prunus reference map.” Genetics
171: 1305-1309.
Howard L.R., Smith R.T., Wagner A.B., Villalon B., Burns E.E., (1994). “Provitamin A
and ascorbic acid content of fresh pepper cultivars (Capsicum annuum) and processed
jalapeños.” Journal of Food Science, 59: 362-365.
IBPGR (1983). “Genetic Resources of Capsicum: A Global Plan of Action.”
International Board for Plant Genetic Resources. AGPG/IBPGR/82/12, Rome, Italy, pp. 49.
Innan H., Tajima F., Terauchi R., Miyashita N.T., (1996). “Intragenic recombination in
the Adh locus of the wild plant Arabidopsis thaliana.” Genetics, 143: 1761-1770.
Innan H., Terauchi R., Miyashita N.T., (1997). “Microsatellite polymorphism in a
natural populations of the wild Arabidopsis thaliana.” Genetics, 146: 1441-52.
Isidore E., van Os H., Andrzejewski S., Bakker J. , Barrena I., Bryan G. J., Caromel B.,
van Eck H., Ghareeb B., de Jong W., van Koert P., Lefebvre V., Milbourne D, Ritter E.,
Rouppe van der Voort J., Rousselle-Bourgeois F., van Vliet J. e Waugh R., (2003). “Toward a

VII-BIBLIOGRAFIA
154
Marker-Dense Meiotic Map of the Potato Genome: Lessons From Linkage Group I” Genetics,
Vol. 165: 2107-2116.
Jacob H. J., Lindpaintner K., Licopaintner K., Lincoln S.E., Kusumi K., Bunker R.K.,
Mao Y.P., Ganten D., Dzau V.J., Lander E.S., (1991). “Genetic mapping of a gene causing
hypertension in the stroke-prone spontaneously hypertensile rat.” Cell, 67: 213-224.
Jacquin N.J., (1776). “Hortus Botanicus Vindobonensis.” Vienna.
Jones D.R., Unwin C.H., Ward E.W.B., (1975) “Capsidiol induction in pepper fruit
during interactions with Phytophthora capsici and Monilinia fructicola.” Phytopathology, 65:
1417-1419.
Jones C.J., Edwards S., Castiglione M.O., Winfield F., Sala C., van der Wiel G.,
Bredemeijer B., Vosman M., Matthes A., Maly R., Brettschneider P., Bettini M., Buiatti E.,
Maestri A., Malcevschi N., Marmiroli R., Aert G., Volckaert J., Rueda R., Linaacero A.,
Vazque A. e Karp A., (1997). “Reproducibility testing of RAPD, AFLP and SSR markers in
plants by a network of European laboratories.” Molecular Breeding, 3: 381–390.
Kang B. C., Nahm S.H., Huh J.H., Yoo H.S., Yu J.W., Lee M.H., Kim B.D., (2001).
“An interspecific (Capsicum annuum x C.Chinese) F2 linkage map in pepper using RFLP and
AFLP markers.” Theor Appl Genet, 102: 531-539.
Kanner J., Harel S., Mendel H., (1979). “Content and stability of α-tocopherol in fresh
and dehydrated pepper fruits (Capsicum annuum L.).” Journal of Agriculture, Food and
Chemistry, 27: 1316-1318.
Keller U., Flath R.A., Mon T.R., Terranischi R., (1981). “Volatiles from red pepper
(Capsicum spp.).” In: Terranschi R., Barrera-Berirtz H. (eds) “Quality of selected Fruit and
Vegetables of North America”, ACS Symposium Series 170. American Chemical Society,
Washington DC, Chapter 12.
Kinsella J.E., (1971). “Composition of the lipids of cucumber and peppers.” Journal of
Food Science, 36: 865-866.

VII-BIBLIOGRAFIA
155
Konieczny A. e Ausubel F.M., (1993). “A procedure for mapping Arabidopsis
mutations using co-dominant ecotypes estimated by DNA sequence comparison.” Plant
Journal, 4: 403-410.
Kosambi D.D., (1944). “The estimation of map distances from recombination values.”
Annals of Eugenics, 12: 172-175.
Kuang H, Richardson T, Carson S, Wilcox P, Bongarten B (1999). “Genetic analysis of
inbreeding depression in plus tree 850.55 of Pinus radiata D.Don. I. Genetic map with
distorted markers.” Theor Appl Genet, 98:697–703.
Lagercrantz U., Ellengren H., Andersson L., (1993). “The abundance of various
polymorphic microsatellite motifs differs between plants and vertebrates.” Nucleic Acids Res,
21: 1111-1115.
Lander E.S., Green P, Abrahamson J, Barlow A, Daly MJ, Lincoln SE, Newburg L.,
(1987). “MAPMAKER: an interactive computer package for constructing primary genetic
linkage maps of experimental and natural populations.” Genomics., 1(2) :174-81.
Lanteri S. e Pickersgill B., (1993). “Chromosomal structural changes in Capsicum
annuum L. and Capsicum annuum Jacq.” Euphytica, 67: 155-160.
Lanteri S., (1991). “Lack of a karyotype class and skewed chromosome segregation in
two back-cross progenies of Capsicum.” Journal of Genetics & Breeding, 45: 51-58.
Lee Y., Howard L.R., Villalon B., (1995). “Flavonoids and antioxidant activity of fresh
pepper (Capsicum annuum) cultivars.” Journal of Food Science, 60: 473-476
Lee, J.M., Nahm S. H., Kim Y.M., Kim B. D., (2004). “Characterization and molecular
genetic mapping of microsatellite loci in pepper.” Theor Appl Genet, 108: 619-627.
Lefebvre V. e Palloix A., (1996). “Both epistatic and additive effects of QTLs are
involved in polygenic-induced resistance to disease: a case study, the interaction pepper-
Phytophthora capsici Leonian.” Theor Appl Genet, 93: 503-511.

VII-BIBLIOGRAFIA
156
Lefebvre V., Caranta C., Pflieger S., Moury B., Daubèze A.M., Blattes A., Ferrière C.,
Phaly T., Nemouchi G., Ruffinatto A. e Palloix A., (1997). “Updated intraspecific maps of
pepper.” Capsicum and Eggplant Newsletter, 16: 35-41. Invited paper.
Lefebvre V., Palloix A., Caranta C., Pochard E., (1995). “Construction of an
intraspecific integrated linkage map of pepper using molecular markers and doubled-haploid
progenies.” Genome, 38: 112-121.
Lefebvre V., Pflieger S., Thabuis A., Caranta C., Blattes A., Chauvet J.C., Daubèze
A.M. e Palloix A., (2002). “Towards the saturation of the pepper linkage map by alignment of
three intraspecific maps including known-function genes.” Genome, 45: 839-854.
Lefebvre, V. (2004). “Molecular Markers for Genetics and Breeding: Development and
Use in Pepper (Capsicum spp.).” Biotechnology in Agriculture and Forestry Molecular
Marker Systems (ed. By H. Lörz and G. Wenzel) © Springer-Verlag Berlin Heidelberg 2004.
Lincoln S. E:, Daly M. J., e Lander E. S., (1993). “Constructing genetic linkage map
with mapmaker/exp v.3.0: a tutorial and reference manual.” Whitehead Institute Technical
Report, Cambridge, MA.
Linnaeus C., (1753). “Species plantarum.”, Stockholm.
Livingstone K.D., Lackney V.K., Blauth J.R., van Wijk R e Jahn M.K., (1999).
“Genome Mapping in Capsicum and the Evolution of Genome Structure in the Solanaceae.”
Genetics, 152: 1183-1202.
Lorenzetti F., Cirica B., (1974). “Quota d’incrocio, struttura genetica delle popolazioni
e miglioramento genetico del peperone (Capsicum annuum L.).” Genetica Agraria, XXVIII,
2: 191-203.
Lownds N.K., Banaras M., Bosland P.W., (1993). “Relationship between postharvest
water loss and physical properties of pepper fruit (Capsicum annuum L.).” HortScience, 28:
1182-1184.

VII-BIBLIOGRAFIA
157
Lyons J.M., Lippert L.F., (1996) “Characterization of fatty acids from roots and shoot
lipids of Capsicum species.” Lipids, 1 (2): 136.
Matus Z., Deli J., Szabolcs J.J., (1991). “Carotenoid composition of yellow pepper
during ripening-isolation of B-cryptoxanthin 5,6-epoxide.” Journal of Agriculture, Food and
Chemistry, 39: 1904-1914.
McKee L., (1998a). “Chile skins as a fiber source.” NMSU Chile Conference, Las
Cruces, New Mexico.
Morgante M., Hanafey M., Powell W., (2002), “Microsatellite are preferentially
associated with non-repetitive DNA in plant genomes.” Nature Genet, 30: 194-200.
Morgante M., Olivieri A.M., (1993). “PCR-amplified microsatellites as markers in plant
genetics.” Plant J., 3: 175-182.
Morton N.E., (1955). “Sequential tests for the detection of linkage.” Am J Hum Genet,
7 (3): 277-318.
Moscone E.A., Lambrou M., Hunziker A.T. e Ehrendorfer F., (1993). “Giemsa C
banded karyotypes in Capsicum (Solanaceae). ” Plant Syst. Evol., 186: 213–229.
Mozafar A., (1994). “Plant Vitamins: Agronomic, Physiological and Nutritional
Aspects.” CRC Press, Boca Raton, Florida.
Mullis K.B., Faloona F.A., Scharf S.J., Saiki K., Horn G.T., Herlich H.A., (1986).
“Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction.” Cold
Spring Harbor Symp. Quant. Biol., 51: 263-273.
Nagy I., Polley A., Ganal M., (1998). “Development and characterization of
microsatellite markers in pepper.” Xth Meeting on Genetics and Breeding of Capsicum and
Eggplant, pp 235-237.

VII-BIBLIOGRAFIA
158
Nagy I., Sasvári S., Stágel A., Ács S., Bárdos G., (2004). “Occurrence and
polymorphism of microsatellite repeats in the sequence databases in pepper”. Institute of
Genetics, Agricultural Biotechnology Center, Szent-Györgyi A u. 4. H-2100 Gödöllõ,
Hungary.
Nakayama R. M., e Matta F. B., (1985). “ “NuMex Naky” chile pepper.” HortScience,
20: 961-962.
Neff M.M., Neff J.D., Chory J. e Pepper A.E:, (1998). “dCAPS, a simple technique for
the genetic analysis of single nucleotide polymorphisms: experimental applications in
Arabidopsis thaliana genetics.” The Plant Journal, 14 (3): 387-392.
Nei M., (1973). “Analysis of gene diversity in subdivided populations.” Proc. Natl.
Acad. Sci. USA, 70: 3321-3323.
Nei M. e Li W., (1979). “Mathematical model for studying genetic variance in terms of
restriction endonucleases.” Proc. Nati. Acad. Sci., USA, 76: 5269-5273.
Nicot N., Chiquet V., Gandon B., Amilhat L., Legeai F., Leroy P., Bernard M.,
Sourdille P., (2004). “Study of simple sequence repeat (SSR) markers from wheat expressed
sequence tags (ESTs).” Theor Appl Genet, 109: 800-805.
Nisperos-Cerriedos M.O., Buslig B.S., Shaw P.E., (1992). “Simultaneous detection of
dehydroascorbic, ascorbic, and some organic acids in fruits and vegetables by HPLC.”
Journal of Agriculture and Food Chemistry, 40: 1127-1130.
NRC, National Research Council, Food and Dietary Board, (1989). “Recommended
Dietary Allowances.” National Academy of Sciences, Washington, DC.
Nybom H., Rogstad S.H. e Schaal B.A., (1990). “Genetic variation detected by use of
the M13 DNA fingerprint probe in Malus, Prunus and Rubus (Rosaceae).” Theoretical and
Applied Genetics, 7: 153-156.

VII-BIBLIOGRAFIA
159
Ogundiwin E.A., Berke T.F., Massoudi M., Black L., Huestis G., Choi D., Lee S. and
Prince P., (2005). “Construction of 2 intra-specific linkage maps and identification of
resistance QTLs for Phytophthora capsici root-rot and foliar blight diseases of pepper
(Capsicum annuum L.)” Genome, 48:698-711.
Osuna-Garcia J.A., (1996). “Red color retenction durino paprika (Capsicum annuum L.)
storage as affected by natural antioxidants and moisture content.” PhD dissertation, New
Mexico State University, Las Cruces, New Mexico, USA.
Palloix A., Daubèze A.M., Pochard E. (2003). “Le Piment. In: Pitrat M., Foury C. (eds)
Histoire de legumes: Des origins à l’orée du XXIe siècle.” Edition INRA, Paris, France, pp
278-290.
Paran I., Rouppe van der Voort J., Lefebvre V., Jahn M., Landry L., van Schriek M.,
Tanyolac B., Caranta C., Ben Chaim A., Livingstone K., Palloix A., Peleman J., (2004). “An
integrated genetic linkage map of pepper (Capsicum spp.).” Molecular Breeding, 13: 251-261,
(online 2003).
Paterson A.H., Bowers J.E., Burow M.D., Draye X., Elsik C.G., Jiang C.X., Katsar
C.S., Lan T.H., Lin Y.R., Ming R. e Wright R.J., (2000). “Comparative genomics of Plant
Chromosomes.” Plant Cell, 12: 1523-1540.
Pekkinen M., Varvio S., Kulju K.K.M., Kärkkäinen H., Smolander S., Viherä-Aarnio
A., Koski V., Sillanpää M.J., (2005). “Linkage map of birch, Betula pendula Roth, based on
microsatellites and amplified fragment length polymorphisms.” Genome, Vol. 48, Number 4,
(7): 619-625.
Peleman J., van Wijk R., van Oeveren J. e van Schaik R., (2000). “Linkage map
integration: an integrated genetic map of Zea mays L.” Poster P472. Plant and Animal
Genome Conference VIII, San Diego, California, USA.
Pickersgill B., (1991). “Cytogenetics and evolution of Capsicum.” In: Tsuchiya T.,
Gupta P.K. (eds), “Chromosome engineering in plants: genetics, breeding, evolution.”
Elsevier, Amsterdam, pp 139-160.

VII-BIBLIOGRAFIA
160
Pickersgill B., Heiser C.B., McNeill J., (1979). “Numerical and Taxonomic studies on
variation and domestication in some species of Capsicum.” The Biology and Taxonomy of the
Solanaceae, 7: 679-699.
Pochard E. e Dumas de Vaulx R., (1982). “Localization of vy2 and fa genes by trisomic
analysis.” Capsicum Newsletter, 1: 18-19.
Pochard E., (1970). “Description des trisomiques de piment (Capsicum annuum L.)
obtenus dans la descendance d’une plante haploïde.”Ann Amélior Plantes, 20 : 233-256.
Pochard E., (1977). “Localization of genes in Capsicum annuum L. by trisomic
analysis.” Ann Amélior Plantes, 27: 255-266.
Portis E., Nagy I., Sasvári Z., Stágel A., Barchi L. e Lanteri S., (2006). “Capsicum EST-
derived microsatellite and their (potential) utilità for mapping studies”, 50° Annual Congress,
Società Italiana di Genetica Agraria – Ischia – 10/14 settembre 2006.
Powell W., Morgante M., Andre C.H., Hanafey M., Vogel J., Tingey S., Rafalski A.,
(1996). “The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for
germplasm analysis.” Mol. Breed, 2: 225-238.
Prince J.P., Pochard E. e Tanksley S.D., (1993). “Construction of a molecular linkage
map of pepper and a comparison of synteny with tomato.” Genome, 36: 404-417.
Qi X., Stam P. e Lindhout P., (1998). “Use of locus-specific AFLP markers to construct
a high-density molecular map in barley.” Theor. Appl. Genet., 106: 1457-1466.
Röder M.S., Korzun V., Wendehake K., Plaschke J., Tixier M.H., Leroy P., Ganal
M.W., (1998). “A microsatellite map of wheat.” Genetics, 149: 2007-23.
Rohlf (2002). “Geometric morphometrics in systematics.” Published as a chapter in N.
Macleod and P. Forey (eds.) “Morphology, shape and phylogenetics.” Taylor&Francis:
London.

VII-BIBLIOGRAFIA
161
Ruiz H. e Pavon J., (1790). “Flora Peruviana, et Chilensis”, 2: 30-31” cited in Heiser e
Smith (1958).
Sasvári Z., Bárdos G., Ács S., Stágel A., Nagy I., (2004). “Construction of a new
interspecific genetic map in pepper based on AFLP and microsatellite markers.” Proc. XIIth
Meeting on Genetics and Breeding of Capsicum and Eggplant, Noordwijkerhout, The
Netherland.
Schlötterer C. e Tauz D., (1992). “Slippage synthesis of simple sequence DNA.”
Nucleic Acids Research, 20: 211.
Schmidt T., Heslop-Harrison J.S., (1996). “The physical and genomic organization of
microsatellites in sugar beet.” Proc Natl Acad Sci, USA, 93: 8761-8765.
Schweder M.E., Shatters Jr. R.G., West S.H. e Smith R.L., (1995). “Effect of transition
interval between melting and annealing temperatures on RAPD analyses.” Biotechniques, 19:
38–42.
Scoville W.L., (1912). “Note on Capsicum.” Journal of the American Pharmaceutical
Association, 1: 453.
Silver M.L., Maruniak J.A., (1981). “Trigeminal chemoreception in the nasal and oral
cavities.” Chemical Sensea, 6: 295-305.
Smith P.G., Villalon B., Villa P.L., (1987). “Horticultural classification of pepper
grown in the United States.” HortScience, 22: 11-13.
Sneath P.H.A. e Sokal R.R:, (1973). Numerical taxonomy. The pricipales of numerical
classification. W.F. Freeman, San Francisco.
Somos A., (1984). “The paprika.” Akademiai Kiado, Budapest.
Sugita T., Kinoshita T., Kawano T., Yuji K., Yamaguchi K., Nagata R., Shimizu A.,
Chen L., Kawasaki S. and Todoriki A., (2005). “Rapid construction of a linkage map using

VII-BIBLIOGRAFIA
162
High-efficiency Genome Scanning/AFLP and RAPD , Based on an Intra-specific, Doubled-
haploid Population of Capsicum annuum.” Breeding Science, 55: 287-295.
Tanksley S.D., (1984). “Linkage relationships and chromosomal locations of enzyme
coding genes in pepper, Capsicum annuum.” Chromosoma, 89: 352-360.
Tanksley S.D., Bernatzky R., Lapitan N.L. e Prince J.P., (1988). “Conservation of gene
repertoire but not gene order in pepper and tomato.” Proc Natl Acad Sci, USA, 85: 352-360.
Tanksley S.D., Ganal M.W., Prince J.P., de Vincente M.C., Bonierbale M.W., Broun P.,
Fulton T.M., Giovannoni J.J., Grandillo S. e Martin G.B., (1992). “High Density Molecular
Linkage Maps of the Tomato and Potato Genomes.” Genetics, 132: 1141-1160.
Tautz D., Renz M., (1984). “Simple sequences are ubiquitous repetitive components of
eukaryotic genomes.” Nucleic Acid Research, 12: 4127-4238
Temnykh S., Park W.D., Ayres N., Cartinhour S., Hauck N., Lipovich L., Cho Y.C.,
Ishii T., McCouch S. (2000). “Mapping and the genome porganization of microsatellite
sequences in rice (Oryza sativa L.). Theor Appl Genet, 100: 697-712.
Terauchi R. e Konuma A., (1994). “Microsatellite variation in Dioscorea tokoro, a wild
yam species.” Genome, 37: 794-801.
Thabuis A., Lefebvre V., Bernard G., Daubèze A.M., Pochard E., Palloix A. (2004).
“Phenotypic and molecular evaluation of a recurrent selection program for a polygenic
resistance to Phytophthora capsici in pepper.” Theor Appl Genet, 109: 342-351.
Thabuis A., Palloix A., Pflieger S., Daubèze A.M, Caranta C., Lefebvre V. (2003).
“Comparative mapping of Phytophthora resistance loci in pepper germplasm: evidence for
conserved resistance loci across Solanaceae and for a large genetic diversity.” Theor Appl
Genet, 106: 1473-1485.
Thiel T., Michalek W., Varshney R.K., Graner A., (2003). “Exploiting EST databases
for the development and characterization of gene-derived SSR-markers in barley (Hordeum
vulgare L.)”. Theoretical and Applied Genetics, 106: 411–422.

VII-BIBLIOGRAFIA
163
Thomas M.R., Scott N.S., (1993). “Microsatellite repeats in grapevine reveal DNA
polymorphisms when analysed as sequence-tagged sites (STSs).” Theoretical and Applied
Genetics, 86: 985-990.
Tong N., (1998). “Genetic relationships among Capsicum species.” PhD thesis, New
Mexico State University, Las Cruces, New Mexico, USA.
Torabi H., (1997). “Identification of novel capsacinoids.” MS thesis, New Mexico State
University, Las Cruces, New Mexico, USA.
Tourneforte J.P., (1700). “Institutiones rei Herbariae”, 3 volumes, Paris.
Valli E., (2001). “Il Peperone.” Ed. Agricole. Bologna.
Vos P., Hogers R., Bleeker M., Reijand M., Van de Lee T., Hornes M., Fritjers A., Pot
J., Paleman J., Kuiper M. e Zabeau M., (1995). “AFLP: A new technique for DNA
fingerprinting.” Nucleic Acids Research, 23: 4407-4414.
Vuylsteke M., Mank R., Antonise R., Bastiaanse E., Senior M. L., Stuber C. W.,
Melchinger A. E., Lubberstedt T., Xia X. C., Stam P., Zabeau M. e Kuiper M., (1999). “Two
hight densisty AFLP ® linkage maps of Zea mays L.: analysis of distribution of AFLP
markers.” Theor. Appl. Genet., 99: 921-935.
Wall M.M., Biles C.L., (1993). “Alternaria fruit rot of ripening chile peppers.”
Phytopathology 83: 324-328.
Wang S., Basten C.J. e Zeng Z.B. (2003). “Windows QTL Cartographer 2.0.”
Department of Statistics, North Carolina State University, Raleigh, N.C..
Williams J.G.K., Kubelik A.R., Livak K,J., Rafalski J.A e Tingey1 Scott V., (1990).
“DNA polymorphisms amplified by arbitrary primers are useful as genetic markers”. Nucleic
Acids Res. 25; 18(22): 6531–6535.

VII-BIBLIOGRAFIA
164
Wimalasiri P., Wills R.B.H., (1983). “Simultaneus analysis of ascorbic acid and
dehydroascorbic acid in fruits and vegetables by high-performance liquid chromatography.”
Journal of Chromatography 256: 368-371.
Xu Z., Zou F. e Vision T.J., (2005). “Improving Quantitative Trait Loci Mapping
Resolution in Experimental Crosses by the Use of Genotypically Selected Samples.”
Genetics, Vol. 170: 401-408.
Yang H., Sweetingham M.W., Cowling W.A. e Smith P.M.C., (2001). “DNA
fingerprinting based on microsatellite-anchored fragment length polymorphisms, and isolation
of sequence-specific PCR marker in lupin (Lupinus angustifolius L.).” Molecular Breeding, 7:
203-209.
Yoon J. Y., Green S. K., Talekar N. S., Chen J. T. (1991). “Pepper improvement in the
tropics: problems and the AVRDC approach. Tomato and pepper production in the tropics.”
Proceedings of the international symposium on integrated management practices, Tainan,
Taiwan, 21-26 March 1988, AVRDC, pp 86-98.
Zane L., Bargelloni L. e Patarnello T., (2002). “Strategies for microsatellite isolation: a
review.” Molecular Ecology, 11: 1-16.
http://www.FAO.org 2003

VII-BIBLIOGRAFIA
165
Letteratura non citata
Alberini E., Porceddu A., Marconi G., Barcaccia G., Pallottini L. e Falcinelli M.,
(2003). “Microsatellite-AFLP for genetic mapping of complex polyploids.” Genome 46 (5):
824-832.
Bartual R., Lacasa A., Marsal J.I., Tello J.C., (1994). “Epistassi in the resistance of
pepper to Phytophthora stem blight (Phytophthora capsici L.) and its significance in the
prediction of double cross performances.” Euphytica, 72: 149-152.
Burr B., Burr F.A., Thompson K.H., Albertson M. C. e Stuber C. W., (1997). “Gene
Mapping with Recombinant Inbreds in Maize.” Genetics, 118: 519-526.
Caranta C., Lefebvre V. e Palloix A., (1997). “Polygenic resistance of pepper to
potyviruses consists of a combination of isolate specific and broad spectrum quantitative trait
loci.” Plant-Microbe Interactions, 10: 872-878.
Caranta C., Palloix A., Gerbe-Selassie K., Lefebvre V., Moury B., Daubèze A.M.,
(1996). “A complementation of two genes originating from susceptible Capsicum annuum
lines confers a new and complete resistance to pepper veinal mottle virus.” Phytopathology,
86: 739-743.
Cheng C.M., Palloix A. e Lefebvre V., (2002). “Isolation, mapping and characterization
of allelic polymorphism of Chi3-P1, a class III chitinose of Capsicum annuum L.” Plant
Science, 163: 481-489.
Conicella C., Errico A. e Saccardo F., (1990). “Cytogenetic and isozyme studies of wild
and cultivated Capsicum annuum.” Genome, 33: 279-282.
D. de Vienne (1998).“Les marqueurs moléculaires en génétique et biotechnologies
végétales.” INRA EDITIONS.

VII-BIBLIOGRAFIA
166
Daubèze A.M., Hennart J.W., Palloix A., (1995). “Resistance to Leveillula taurica in
pepper (Capsicum annuum) is oligogenically controlled and stable in Mediterranean regions.”
Plant Breed, 114: 327-332.
Djian-Caporalino C. et al. (2001). “High-resolution genetic mapping of the pepper
(Capsicum annuum L.).” Theor Appl Genet, 103: 592-600.
Djian-Caporalino, C. et al. (1999). “Spectrum of resistance to root-knot nematodes and
inheritance of heat-stable resistance in pepper (Capsicum annuum L.).” Theor Appl Genet,
99: 496-502.
Eshbaugh W. H. (1980). “The taxonomy of the genus Capsicum (Solanaceae).”
Phytologia, 47: 153-165.
Grube R.C., Blauth J.R., Arnedo Andres M.S., Caranta C. e Jahn M.M., (2000).
“Identification and comparative mapping of a dominant potyvirus gene cluster in Capsicum.”
Theor. Appl. Genet., 101: 852-859.
Heiser C.B. e Smith P.G., (1953). “The cultivated Capsicum peppers.” Economic
Botany, 7 (3), 214-217.
Huang S.W., Zhang B.X., Milbourne D., Cardle L., Yang G.M. e Guo J.Z., (2001).
“Development of pepper SSR markers from sequence databases.” Euphytica, 117: 163-167.
Jahn M., Paran I., Hoffman K., Radwansky E.R., Livingstone K.D., Grube R.C.,
Aftergoot E., Lapidot M., Moyer J., (2000). “Genetic mapping of the Tsw locus for resistance
to the same pathogene in tomato.” Mol Plant Microbe Interact, 13: 673-682.
Kang B. C. et al. (2000). “Construction of a molecular linkage map in hot pepper.” Acta
Hort., 521. ISHS.
Knox, M. R. et al (2002). “Excess Heterozygosity Contributes to Genetic Map
Expansion in Pea Recombinant Inbred Populations.” Genetics, 162: 861-873.

VII-BIBLIOGRAFIA
167
Koebner R., Powell W. e Donini P., (2001). “Contributions of DNA Molecular Marker
Technologies to the Genetics and Breeding of Wheat and Barley.” Plant Breeding Reviews,
21.
Lefebvre V., Palloix A. e Rives M., (1993). “Nuclear RFLP between pepper cultivars
(Capsicum annuum L.). Euphytica, 71: 189-199.
Lefebvre V., Kuntz M., Camara B., Palloix A., (1998). “The capsanthin-capsorubin
synthase gene: a candidate gene for the y locus controlling the red fruit color in pepper.” Plant
Mol Biol, 36: 785-789.
Litt M., Luty J. A., (1989). “A hypervariable microsatellite revealed by in vitro
amplification of a dinucleotide repeat within the cardiac muscle action gene. ” Am. J. Hum.
Genet., 44: 388-396.
Rao G.U., Ben Chaim A., Borovsky Y. E Paran I., (2003). “Mapping of yield-related
QTLs in pepper in an interspecific cross of Capsicum annuum and C. frutescens.” Theor Appl
Genet, 106: 1457-1466.
Thabuis A., Palloix A., Servin B., Daubèze A.M., Signoret P., Hospital F., Lefebvre V.,
(2003). “Marker-assisted introgression of 4 Phytophthora capsici resistance QTL alleles into a
bell pepper line: validation of additive and epistatic effects.” Mol Breed (on line 2003), 14: 9-
20.
Thorup T.A., Tanyolac B., Livingstone K.D., Popovsky S., Paran I., Jahn M., (2000).
“Candidate gene analysis of organ pigmentation loci in the Solanaceae.” Proc. Natl. Acad.
Sci. USA, 97: 11192-11197.

VII-BIBLIOGRAFIA
Ringraziamenti
La preparazione della Tesi di II livello è stata per me un’esperienza interessante,
costruttiva e formativa, che mi ha premesso di conoscere in modo concreto il mondo della
ricerca scientifica.
Ringrazio pertanto il Prof. Sergio Lanteri per avermi dato l’opportunità di svolgerla nel
suo dipartimento e per avermi seguita costantemente durante la stesura della tesi.
Ringrazio soprattutto Ezio Portis, Lorenzo Barchi e Alberto Acquadro per la loro grande
disponibilità e pazienza nei miei confronti; mi hanno fornito un grande supporto scientifico,
tecnico, informatico e umano.
Ringrazio inoltre Cinzia, Francesco, Andrea e Diana per i preziosi consigli e
suggerimenti in laboratorio.
Ringrazio mia mamma, Paolo, Sergio, tutti i miei parenti e amici per avermi supportata
durante la mia carriera universitaria.





























