Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SANTA CATARINA
CENTRO DE CIÊNCIAS FÍSICAS E MATEMÁTICAS
COORDENADORIA ESPECIAL DE OCEANOGRAFIA
GRADUAÇÃO EM OCEANOGRAFIA
Victória Silvestre Corrêa
CRESCIMENTO E MORFOLOGIA DA DIATOMÁCEA Asterionellopsis glacialis EM
CONDIÇÕES CONTROLADAS DE CULTIVO EM LABORATÓRIO
Florianópolis – SC
2021

1
Victória Silvestre Corrêa
CRESCIMENTO E MORFOLOGIA DA DIATOMÁCEA Asterionellopsis glacialis EM
CONDIÇÕES CONTROLADAS DE CULTIVO EM LABORATÓRIO
Trabalho de Conclusão do Curso de Graduação
em Oceanografia do Centro de Ciências Físicas
e Matemáticas da Universidade Federal de
Santa Catarina como requisito básico para a
obtenção do Grau de Bacharel do Curso de
Oceanografia.
Orientador: Leonardo Rubi Rörig
Florianópolis – SC
2021

2

3
Victória Silvestre Corrêa
Crescimento e morfologia da diatomácea Asterionellopsis glacialis em condições
controladas de cultivo em laboratório
Este Trabalho de Conclusão de Curso foi julgado adequado para obtenção do Título de
Bacharel em Oceanografia e aprovado em sua forma final pelo Curso de Oceanografia.
Florianópolis, 28 de abril de 2021
____________________________
Prof. Felipe Mendonça Pimenta, Dr.
Coordenador do curso
Banca Examinadora:
_________________________
Prof. Leonardo Rubi Rörig, Dr.
Orientador
Universidade Federal de Santa Catarina
_________________________
Prof. Paulo Horta Júnior, Dr.
Avaliador
Universidade Federal de Santa Catarina
_______________________
Ana Gabriela Itokazu Canzian da Silva, Msc.
Avaliadora
Auburn University

4
AGRADECIMENTOS
Em primeiro lugar, gostaria de agradecer aos meus pais, que durante todo o tempo
moveram montanhas por mim para me propiciar as melhores oportunidades em todos os
aspectos da minha vida, e que também sempre me apoiaram e me incentivaram a seguir meus
sonhos, independentemente de quais fossem.
Às amigas que a Oceanografia me apresentou: Rúbia Avelar, Thais Konrad, Ariane
Paulutti e Karina Albuquerque. Embora presentes em fases diferentes da minha graduação,
todas tiveram e têm uma grande importância na minha vida. Agradeço por estarem sempre ao
meu lado e me auxiliarem nos mais diversos momentos. Com certeza o processo da graduação
foi mais agradável ao lado das risadas e dos abraços delas.
A uma amiga extremamente especial, Giulia Gaia, por me acolher em sua família e me
fazer sentir como se fosse sua irmã. Agradeço por sempre ouvir minhas reclamações e me
ajudar, e também pelas incontáveis risadas que demos ao longo desse tempo. Todos esses anos
em Florianópolis não teriam sido tão marcantes sem a presença dela.
A meu orientador Leonardo Rörig, sempre muito paciente e dedicado no processo de
ensinar, quase como um segundo pai, tratando todos os alunos com muito carinho e atenção. E
também aos outros membros da banca, Prof. Paulo Horta e a Gabii, por disporem de tempo e
atenção para me ajudar a completar essa fase.
E por último, a todas as outras (muitas) pessoas que, direta ou indiretamente, me
ajudaram e me incentivaram ao longo dessa jornada.

5
Dedico este trabalho à minha mãe Sonita, que sempre me incentivou na busca pelo
conhecimento, dizendo que essa sim é nossa maior herança, pois ninguém pode nos tirar. E a
meu pai, Edvaldo (in memorian), que me apresentou o mar e era tão fascinado por ele assim
como eu sou, também me incentivando a ser uma grande cientista e a desvendar os mistérios
que o oceano carrega.

6
RESUMO
O efeito da variação de alguns fatores ambientais nos cultivos de microalgas pode ser refletido
em seu crescimento e morfologia celular. No Brasil, a diatomácea Asterionellopsis glacialis é
um organismo muito importante para as praias dissipativas da região Sul, servindo de alimento
para diversos organismos da teia trófica. No presente trabalho foi realizado um experimento
para analisar o crescimento e desenvolvimento de A. glacialis sob diferentes intensidades de
turbulência, sendo criados dois grupos, um sob aeração e o outro estático, durante 14 dias. A.
glacialis foi cultivada em meio de cultivo F/2. Este estudo buscou responder quais foram os
fatores que influenciaram o crescimento, a morfologia, e as diferenças entre os organismos,
quando aconteceram estas mudanças e por quais motivos. Durante o período experimental,
semanalmente, foram retiradas alíquotas dos frascos e os espécimes foram fixados em
microtubos em formol. Posteriormente, foram retiradas fotografias dos espécimes fixados para
comparação entre os grupos com e sem aeração. Ao longo de todas as 168 células examinadas,
foram vistas diferenças no crescimento e na morfologia celular, sendo vistas até, deformações
celulares e contaminações no meio de cultivo por outras espécies. As maiores células
encontradas em termos de tamanho foram das amostras que estavam em aeração, entretanto, as
menores também.
Palavras-chave diatomáceas de zona de arrebentação, frústula, morfologia, teratologia,
agregação, sedimentação, nutrientes.

7
ABSTRACT
The effect of the variation on some environmental factors in microalgae cultivations affects
their growth and cellular morphology. In Brazil, the diatom Asterionellopsis glacialis is an
essential organism for the dissipative beaches of the South region, serving as food for several
organisms of the trophic web. In the present study, an experiment was performed to analyze the
growth and development of A. glacialis under different intensities of turbulence, creating two
groups, one under aeration and the other static, for 14 days. A. glacialis was grown in F/2 culture
medium. This study sought to answer the factors that influenced growth, morphology, and the
differences between organisms, when did these changes occur, and for what reasons. During
the experimental period, weekly samples were taken from the flasks and fixed with
formaldehyde. Subsequently, photographs of the fixed specimens were taken for comparison
between the groups with and without aeration. Throughout all 168 cells examined, cell growth
and morphology differences were noted, including cell deformations and culture contamination
by other species. The largest cells found in terms of size were from the samples that were in
aeration; however, the smallest ones also.
Keywords: surf diatoms, frustule, morphology, teratology, aggregation, sedimentation,
nutrients.

8
LISTA DE FIGURAS
Figura 1 - Aspecto de um fenômeno de acumulação da diatomácea de zona de arrebentação
Asterionellopsis glacialis na Praia do Rincão, Içara, SC, Brasil ............................................ 18
Figura 2 - Dados morfométricos adotados para a caracterização morfológica das células de
Asterionellopsis glacialis cultivadas. .................................................................................... 21
Figura 3 - Disposição dos frascos experimentais de crescimento de Asterionellopsis glacialis
na sala de cultivos do LAFIC. (A) Cultivos recém inoculados (t = 0). (B) Cultivo (t = 1) antes
da homogeneização. (C) Cultivo (t = 1) depois da homogeneização. (D) Cultivo (t = 2) antes
da homogeneização. (E) Cultivo (t = 2) depois da homogeneização. ..................................... 22
Figura 4 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras Pré-Experimento.
(A) Cadeia saudável. (B, C) Células com deformações. (D) Célula solitária saudável, com o
cloroplasto destacado ao centro. ........................................................................................... 23
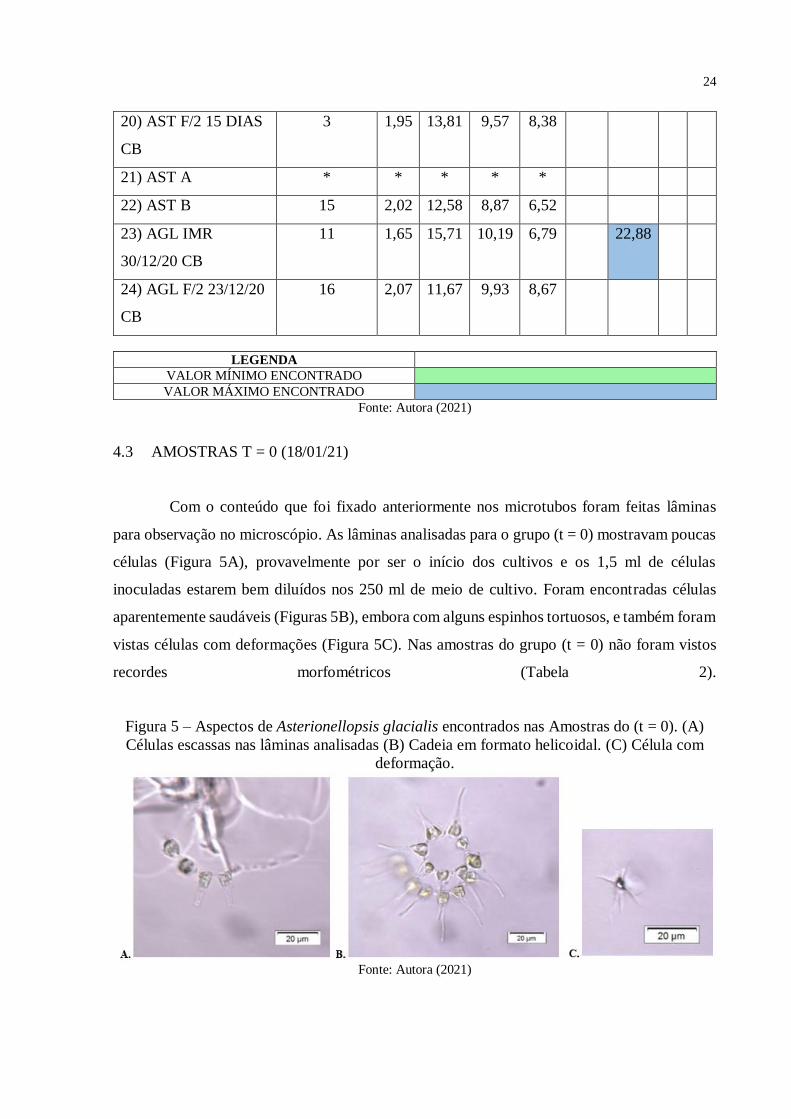
Figura 5 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t = 0). (A)
Células escassas nas lâminas analisadas. (B) Cadeia em formato helicoidal. (C) Célula com
deformação. ......................................................................................................................... 24
Figura 6 - Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t = 1). (A)
Agregados de células. (B) Colônia saudável. (C) Células em vista valvar e em vista
conectival, da esquerda para a direita, respectivamente. (D) Cultivo contaminado por bactéria.
(E) Cadeia com espinhos com crescimento incompleto. ....................................................... 26
Figura 7 - Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t = 2). (A,B)
Células deformadas vistas sob microscópio. ......................................................................... 28
Figura 8 – Aspecto dos cultivos ao final do experimento. (A) Formação de agregados no
fundo dos frascos que estavam em aeração. (B) Formação de agregados no fundo dos frascos
que estavam estáticos. .......................................................................................................... 29
Figura 9 - Classificação dos espinhos de Asterionellopsis glacialis ...................................... 32
Figura 10 - Vista conectival e vista valvar encontradas nas amostras de A. glacialis do
presente estudo. À esquerda, células em vista conectival, e, à direita, células em vista
valvar.........................................................................................................................................33
Figura 11 - Eixo apical e eixo transapical das células, conforme medidas determinadas na
(Figura 1)...................................................................................................................................42
Figura 12 - Distribuição das medidas A, B, C, D dos tratamentos com aeração (CB) e sem
aeração (SB) ao longo do tempo experimental.........................................................................43

9
LISTA DE TABELAS
Tabela 1 - Informações sobre as Amostras Pré-experimento: quantidade de células
fotografadas, média de medidas e recordes de medidas no grupo .......................................... 23
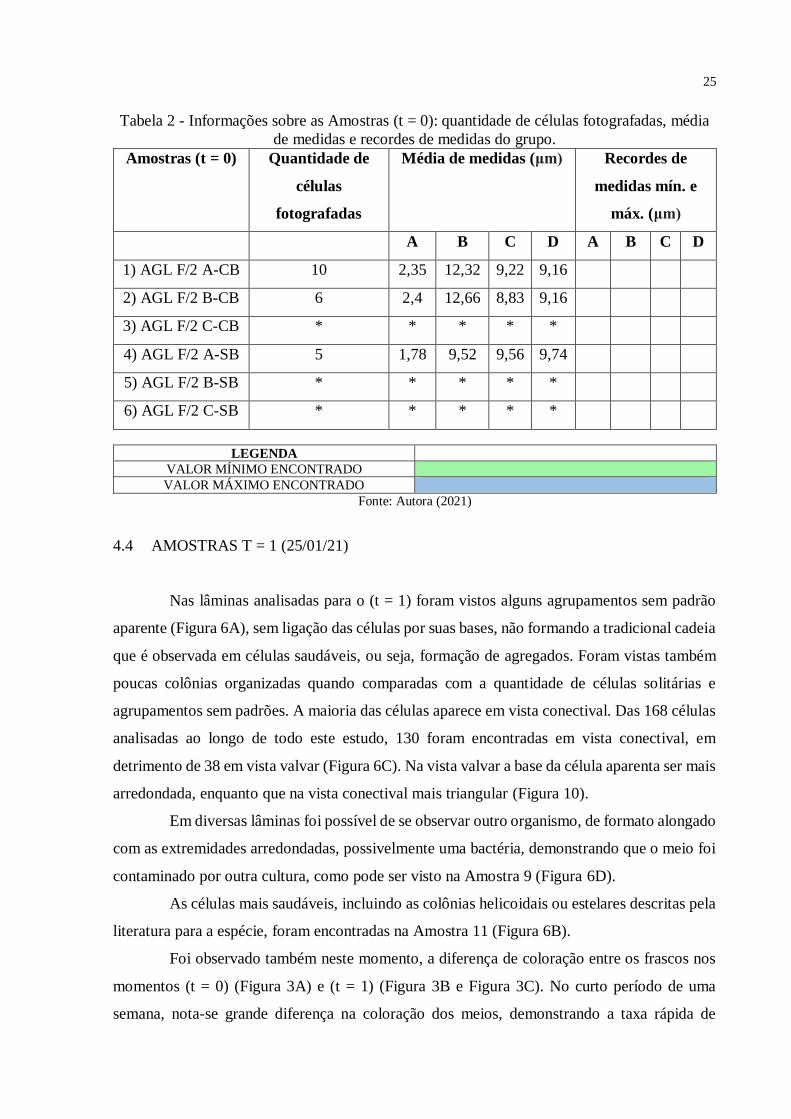
Tabela 2 - Informações sobre as Amostras (t = 0): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo ......................................................................... 25
Tabela 3 - Informações sobre as Amostras (t =1): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo ......................................................................... 27
Tabela 4 - Informações sobre as Amostras (t = 2): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo ......................................................................... 28
Tabela 5 - Reunião de informações sobre todas as amostras: número e código de identificação
da amostra, quantidade de céllas fotografadas, média de medidas e recorde de medidas
encontrados nos grupo ......................................................................................................... 29
Tabela 6 – Comparação entre os dados obtidos no presente trabalho e os dados de Cupp
(1943), para diferentes medidas da célula de Asterionellopsis glacialis ................................ 41

10
LISTA DE ABREVIATURAS E SIGLAS
AGL Asterionellopsis glacialis
AST Asterionellopsis glacialis
CB Com borbulhamento
F/2 Meio de cultivo de microalgas
IMR Meio de cultivo de microalgas
SB Sem borbulhamento
LAFICmu Laboratório de Ficologia Microscópica
IMA Instituto do Meio Ambiente de Santa Catarina
EPS Substâncias Poliméricas Extracelulares
TEP Partículas Exopoliméricas Transparentes

11
SUMÁRIO
1 INTRODUÇÃO .......................................................................................................... 12
2 OBJETIVOS ............................................................................................................... 17
2.1 OBJETIVO GERAL ............................................................................................ 17
2.2 OBJETIVOS ESPECÍFICOS .............................................................................. 17
3 MATERIAIS E MÉTODOS ....................................................................................... 18
3.1 ÁREA DE ESTUDO ............................................................................................ 18
3.2 AMOSTRAGEM IN SITU .................................................................................. 18
3.3 MANUTENÇÃO E ISOLAMENTO DA CEPA ................................................. 19
3.4 AMOSTRAGEM EM LABORATÓRIO ............................................................ 19
3.5 ANÁLISE DOS DADOS ...................................................................................... 19
3.6 DESENVOLVIMENTO DO EXPERIMENTO.................................................. 20
4 RESULTADOS ........................................................................................................... 22
4.1 ASPECTO GERAL DOS CULTIVOS EXPERIMENTAIS: ............................. 22
4.2 ANÁLISE MICROSCÓPICA DOS CULTIVOS ............................................... 23
4.3 AMOSTRAS T = 0 (18/01/21) .............................................................................. 24
4.4 AMOSTRAS T = 1 (25/01/21) .............................................................................. 25
4.5 AMOSTRAS T = 2 (01/02/21) .............................................................................. 27
4.6 MORFOLOGIA DOS ESPINHOS ..................................................................... 31
5 DISCUSSÃO ............................................................................................................... 34
5.1 Morfologia da cepa estudada de acordo com padrões vistos na literatura ........ 34
5.2 Agregação, floculação e sedimentação das células nos cultivos experimentais .. 34
5.3 Considerações sobre a morfologia celular nos cultivos experimentais .............. 38
5.3.1 Dados morfométricos .................................................................................... 41
6 CONCLUSÃO ............................................................................................................. 44
APÊNDICE ........................................................................................................................ 45
REFERÊNCIAS BIBLIOGRÁFICAS .............................................................................. 46

12
1 INTRODUÇÃO
Alga é um termo genérico, sem significado taxonômico, relacionado a organismos que
geralmente apresentam clorofila-a e outros pigmentos atuando em seu metabolismo celular
predominantemente fotoautotrófico, variando de tamanhos macroscópicos a microscópicos. Tal
termo engloba algas procarióticas, como as cianobactérias, bem como as eucarióticas (BONINI,
2012; RAVEN et al., 1996), como as diatomáceas.
As microalgas possuem a capacidade de responder a variações ambientais, como
temperatura e irradiância solar, com mudanças bioquímicas e morfológicas (RENAUD et al,
2002). Diversos processos hidrodinâmicos podem alterar a disponibilidade de nutrientes e de
luz para o fitoplâncton. Oscilações na quantidade de luz podem causar mudanças significativas
nas taxas de fotossíntese e produção primária. Além disso, podem dificultar na absorção de
nutrientes e causar flutuações populacionais (KOOISTRA et al., 2007).
As microalgas, em toda sua diversidade, têm sido objeto de inúmeros estudos ao longo
dos anos devido a sua importância ecológica, econômica e nutricional (ANTELO et al., 2010).
Nas regiões costeiras a predominância é de diatomáceas, pois a maior instabilidade da coluna
d’água disponibiliza nutrientes e turbulência que favorecem a estratégia de vida do tipo ‘’r’’
desse grupo de microalgas (MARGALEF, 1997). O aumento da temperatura atmosférica e a
mudança no padrão de circulação dos ventos é reconhecido como um impacto das mudanças
climáticas globais (SILVA, 2018), e estes podem aumentar ou diminuir o índice das
precipitações e das ressurgências costeiras, de acordo com a região. Com o aumento das chuvas,
a salinidade da água pode ser reduzida. Esses fatores combinados trazem maior quantidade de
nutrientes para a região costeira, causando um aumento da densidade celular e mudanças na
composição do fitoplâncton (VIANNA et al, 2012).
Além da importância de fatores físicos e químicos, é importante salientar a expressiva
interdependência de parâmetros biológicos na determinação do desempenho e estrutura das
assembleias fitoplanctônicas (RAVEN, 1988). Iluminação, temperatura, pH, disponibilidade de
nutrientes, turbulência e regime hidrodinâmico são os aspectos abióticos mais importantes
(RICHMOND, 2004). Em relação aos nutrientes, os principais elementos limitantes do
crescimento destes organismos são: carbono, nitrogênio, fósforo e silício. Entretanto, elementos
como enxofre, potássio, ferro, cálcio e magnésio também são extremamente importantes
(RICHMOND, 2004). Algumas algas também necessitam de vitaminas para seu
desenvolvimento (GUILLARD, 1975). De acordo com Shukla (2013), o tamanho médio das

13
valvas de diatomáceas reflete a disponibilidade de nutrientes, e pode revelar efeitos do clima e
da ciclagem de nutrientes nos oceanos.
Diatomáceas são um grupo de algas unicelulares caracterizado pela presença de um
tipo específico de parede celular, chamada de frústula. Tradicionalmente, essas algas têm sido
identificadas em função da morfologia de suas paredes celulares sob microscopia óptica (COX,
2014). Para a formação das frústulas, é necessária a presença de silício. A frústula é uma
estrutura formada por duas metades, chamadas de valvas ou tecas, que se encaixam
perfeitamente. A teca maior é chamada de epiteca, enquanto que a menor é chamada de hipoteca
(VIEIRA, 2011). Em situações de limitação por silício, pode ser observada a diminuição da
espessura dessa parede celular (LOURENÇO, 2006). De acordo com Werner (1966), muitas
espécies de diatomáceas reagem mais rapidamente à deficiência de ácido ortossilícico
(Si(OH)4) do que à de qualquer outro nutriente. No ambiente marinho e costeiro, a sílica
utilizada para formar as células das diatomáceas se origina principalmente de processos naturais
como o intemperismo das rochas (HUMBORG et al., 2008).
Diatomáceas constituem um dos tipos mais comuns de fitoplâncton. Sua abundância é
tal que as frústulas de diatomáceas mortas, embora microscópicas, formam extensos depósitos
rochosos que podem alcançar vários metros de altura no fundo dos oceanos, compostos
predominantemente por bioclastos silicosos, chamados diatomitos (VIEIRA, 2011). Além de
serem organismos extremamente importantes para o ciclo biogeoquímico do silício, como
pode-se perceber, as diatomáceas possuem também a capacidade de fixar carbono da atmosfera
para grandes profundidades (DONEY, 1997; SMETACEK, 1998), sendo essenciais para o ciclo
biogeoquímico do carbono. Logo, mudanças nas comunidades fitoplanctônicas podem levar a
diferenças temporais e espaciais na quantidade e qualidade da matéria orgânica que chega ao
fundo dos oceanos, o que é um fator importante para a manutenção da ciclagem de nutrientes
(SPILLING et al., 2018).
O gênero Asterionellopsis é caracterizado por células agrupadas em cadeias em forma
estelar ou helicoidal (KOOISTRA et al., 2007). Suas frústulas são inchadas na base e em
formato de agulha ao longo do comprimento, podendo ser encontradas células solitárias ou em
cadeias (CUPP, 1943). São conectadas às células vizinhas pela base, com a ajuda de blocos de
mucilagem excretados pelas mesmas (KOOISTRA et al., 2007). Com ampla distribuição, tais
diatomáceas habitam a zona nerítica (CUPP, 1943) e bêntica (MCLACHLAN & DEFEO,
2017), podendo também ser encontradas em estuários (JOUENNE et al., 2007). Acredita-se
que a formação das cadeias afeta a aptidão das diatomáceas, influenciando a competição por
luz e nutrientes e também as interações com predadores (KENITZ et al., 2020). Estudos

14
sugerem que o regime de luz pode controlar o tipo de produção de espinho em diatomáceas
(BENTLEY, 2012), entretanto, ainda não existem experimentos controlados que determinem
com precisão como condições ambientais podem controlar a mudança de um tipo de produção
de valva para outro (COX, 2014). O comprimento das ondas e a intensidade luminosa podem
também afetar o tamanho das células e o diâmetro e densidade dos poros (SU et al., 2018).
Descrita por Castracane em 1886, Asterionellopsis glacialis foi coletada pela primeira
vez no mar Antártico durante a Expedição Challenger, uma expedição científica cujo objetivo
era analisar fatores biológicos, químicos, físicos e geológicos do oceano. Em alguns estudos,
Asterionellopsis glacialis, inicialmente nomeada como Asterionella glacialis, é considerada um
sinônimo para Asterionella japonica (CLEVE, 1882).
A diatomácea A. glacialis é um organismo importante ecologicamente às praias
dissipativas da costa sul brasileira, ocorrendo com alta frequência na região e apresenta uma
ampla distribuição na América do Sul, variando das latitudes tropicais às temperadas (RORIG
& GARCIA, 2003). Esta espécie pertence a um grupo particular de diatomáceas, chamadas de
diatomáceas de zona de arrebentação (surf diatoms), que formam densas acumulações de algas
de coloração marrom em praias dissipativas expostas (MCLACHLAN & DEFEO, 2017). A
coloração marrom na espuma da zona de arrebentação está associada à presença de matéria
orgânica dissolvida na água, bem como a coloração de um de seus pigmentos típicos, a
fucoxantina (CAMPBELL; BATE, 1997).
Inicialmente essas acumulações eram tidas como florações clássicas e tinham a
aparência de manchas de óleo na superfície. Porém, alguns estudos demonstraram que essas
acumulações não eram florações, mas sim características semipermanentes em zonas de
arrebentação de alta energia onde as células das diatomáceas se dividiam em uma taxa constante
(MCLACHLAN & DEFEO, 2017). De acordo com Odebrecht et al. (1995), em alguns casos,
estudos sugeriram padrões sazonais nas ocorrências, associados à presença de chuvas e ventos.
Franco et al. (2018) sugere que uma maior quantidade de chuvas aumenta o suprimento de
nutrientes disponível para o fitoplâncton da zona de surf, e relata que mais de 80% das
acumulações de A. glacialis aconteceram durante as estações chuvosas, com ventos de
quadrante sul. A passagem de frentes frias vindas da Antártica na região Sul do Brasil, por
exemplo, resulta em ventos em direção à praia, que promovem maior hidrodinâmica (RORIG
& GARCIA, 2003), gerando ressuspensão de estoques bênticos das diatomáceas e acumulando-
as na zona de arrebentação interna (RÖRIG et al., 2017). Kenitz et al. (2020) afirma que
diatomáceas que formam cadeias dominam as assembleias fitoplanctônicas em períodos de
águas mais geladas e eventos de florações e acumulações, enquanto que diatomáceas solitárias

15
são mais dominantes durante períodos nos quais a água está mais quente, associados a passagem
do El Niño e estratificação mais acentuada da coluna d’água, por exemplo.
Essas diatomáceas se acumulam na superfície da água durante o dia, geralmente
associando-se a bolhas de ar (possivelmente devido à ação de camadas mucilaginosas) ou até
formando uma espuma semi-estável. Estando concentradas na superfície da água, podem ser
transportadas por ondas e correntes, e, em determinadas condições, são movidas para a costa,
concentrando-se na zona de arrebentação interna e ficando presas na praia como depósitos de
espuma. As diatomáceas não aparecem só na espuma, mas também são dispersas na coluna
d’água e nos sedimentos, movendo-se entre esses ambientes (MCLACHLAN & DEFEO,
2017). À tarde, um muco mais pegajoso é produzido, e isto pode facilitar a ligação de partículas
finas de sedimento às células (ODEBRECHT et al., 1995). Com isso, sua densidade aumenta,
e, neste período do dia as células afundam com ajuda do peso do sedimento aderido (RÖRIG
& GARCIA, 2003).
Durante a ocorrência de altas concentrações de A. glacialis as taxas de produção
primária podem ser muito altas, podendo assim atender a demanda de matéria orgânica
necessária para o crescimento e desenvolvimento do zooplâncton, bentos e nécton dos
ambientes costeiros (BROWN & MCLACHLAN, 1990; ABREU et al., 2003). A diminuição
de A. glacialis pode impactar atividades econômicas importantes como a pesca (ODEBRECHT
et al., 2010), já que a espécie é o principal item alimentar da tainha (Mugil spp.) (SEELIGER,
2004), que contém 90% do seu conteúdo estomacal composto por diatomáceas.
Quando cultivada em laboratório, após algumas semanas ou meses, A. glacialis
demonstra aberrações em suas frústulas. Uma das hipóteses é a falta ou diminuição de algum
nutriente em seu meio de cultivo ou a qualidade dos mesmos (RÖRIG et al., 2017), ou a
ausência da influência de fatores físicos como turbulência, por exemplo. São vistas formas
teratológicas (ou seja, com deformações) em outras espécies de diatomáceas como: Eunotia sp.,
Frustulia guayanensis (VIANA, 2007) e Fragilaria sp. (SILVA, 2012; WENGRAT, 2011). A
maleabilidade dos parâmetros morfológicos, como nas frústulas por exemplo, em resposta a
mudanças na temperatura ou na concentração de nutrientes é um fenômeno relativamente
comum, e, estas mudanças já foram vistas em outros organismos (MOUSING et al., 2018).
Outra hipótese sobre o aparecimento de variedades morfológicas da espécie ao ser
cultivada em laboratório seria a existência de espécies crípticas. Espécies crípticas são duas ou
mais espécies que, morfologicamente, são indistinguíveis. Porém, geneticamente, possuem
diferenças, sendo assim consideradas espécies diferentes. Podem ser diferenciadas também
ecologicamente, através de alguns comportamentos distintos. Como os estudos anteriores sobre

16
essas florações classificavam sempre a espécie como A. glacialis, não considerando a
ocorrência de espécies crípticas, não é possível saber se os fenômenos de formação de
acumulações na zona de arrebentação de praias arenosas do sul do Brasil são uniespecíficos ou
multiespecíficos (FRANCO et al., 2016).
Com a descrição de quatro espécies crípticas recentemente (KACZMARSKA et al.,
2014), a identidade dos espécimes encontrados em acumulações em zona de surf permanece
incerta. No estudo de Franco et al. (2016), estruturas nas frústulas e sequenciamento
filogenético foram utilizados para estudar amostras, e, uma espécie nova, nomeada
Asterionellopsis tropicalis, foi descrita. A. tropicalis foi diferenciada morfologicamente de
outras espécies do mesmo gênero pelo número de estrias encontradas em 10 μm e outras
características presentes nos polos. Entretanto, A. tropicalis é morfologicamente indistinguível
de A. glacialis. A análise filogenética corrobora a ideia, indicando que não são espécies irmãs.
No processo de especiação, divergências genéticas precedem diferenciação
morfológica, isso explica por que espécies próximas filogeneticamente podem ser
indistinguíveis ao olhar morfológico (LELIAERT et al., 2014).
Diante do exposto, o objetivo principal deste trabalho é investigar se mudanças nos
parâmetros físicos interferem no padrão de crescimento, e, principalmente, na morfologia das
frústulas silicosas da microalga Asterionellopsis glacialis. Nesse sentido, cepas puras foram
isoladas e submetidas a experimentos controlados de cultivo em laboratório. Em função da
incompatibilidade para acesso frequente ao laboratório por conta do regime de pandemia, foi
realizada apenas a análise fotográfica de como estes fatores podem influenciar a morfologia e
o desenvolvimento. A relevância deste esforço técnico-científico parte da premissa de que a
eficiência de cultivos em laboratório poderá ser incrementada com base na compreensão das
alterações de crescimento e morfologia controladas por características físicas.

17
2 OBJETIVOS
2.1 OBJETIVO GERAL
Analisar o padrão de crescimento e a morfologia das frústulas da diatomácea
Asterionellopsis glacialis cultivadas em laboratório, sob diferentes condições físicas, ao longo
de um determinado período de tempo.
2.2 OBJETIVOS ESPECÍFICOS
- Avaliar os padrões morfológicos possíveis de uma cepa de A. glacialis através de
análise fotográfica.
- Verificar a influência da turbulência por aeração sobre os padrões morfológicos da
frústula de uma cepa de A. glacialis em cultivo laboratorial.

18
3 MATERIAIS E MÉTODOS
3.1 ÁREA DE ESTUDO
Este estudo foi realizado utilizando amostras coletadas na Praia do Rincão (28°50’4’’S
- 49°14’10’’O), localizada no município de Içara, Estado de Santa Catarina, no Sul do Brasil.
É uma praia dissipativa com cerca de 16 km de extensão. A região é caracterizada por clima
subtropical úmido, com temperatura média anual de 19.5°C. No mês mais quente a temperatura
média é de 23.9°C, no mês mais frio a temperatura média é de 15.2 °C. De acordo com o
relatório de balneabilidade Nº: 37 do IMA, publicado em 12/03/21, dos cinco pontos de coleta
de água ao longo da Praia do Rincão, quatro foram classificados como próprios para banho
enquanto um foi classificado como impróprio.
3.2 AMOSTRAGEM IN SITU
No presente trabalho será utilizada uma cepa coletada e isolada a partir de uma
acumulação na praia do Rincão, SC, em 2013. A Praia do Rincão possui registro frequente de
acumulações de diatomáceas de zona de arrebentação, especialmente de A. glacialis. Durante a
ocorrência dessas acumulações, a superfície da água apresenta coloração marrom (Figura 1).
Figura 1 – Aspecto de um fenômeno de acumulação da diatomácea de zona de arrebentação
Asterionellopsis glacialis na Praia do Rincão, Içara, SC, Brasil
Fonte: Honorato (2014)

19
3.3 MANUTENÇÃO E ISOLAMENTO DA CEPA
O isolamento da microalga facilita as observações em microscópio. Para este
procedimento, o primeiro passo é escolher o meio de cultivo. O meio F/2 (GUILLARD, 1975)
é amplamente utilizado no isolamento e manutenção de algas, pois tem se mostrado eficiente
para inúmeras espécies de fitoplâncton. Repiques para manutenção das cepas foram feitos
periodicamente, em intervalos de 15 a 30 dias.
3.4 AMOSTRAGEM EM LABORATÓRIO
Na primeira etapa experimental, foi realizada uma sessão de fotos com os cultivos de
A. glacialis já existentes no cepário do Laboratório de Ficologia Microscópica (LAFICmu) para
análise da condição dos espécimes.
Após isso, dois novos grupos de cultivos foram inoculados, um com aeração e um
estático, com três réplicas cada. Ambos foram cultivados em meio de cultivo F/2, e sob as
mesmas condições de irradiância. A aeração dentro dos frascos impede a sedimentação das
células, melhora a distribuição de nutrientes e induz as células a aproveitarem melhor a luz
(LOURENÇO, 2006).
A agitação dentro dos frascos também influencia o crescimento das células e a
morfologia das cadeias. Culturas de A. glacialis cultivadas em laboratório, sem agitação,
começam a apresentar aberrações celulares como redução dos espinhos ou má-formação (ou
mesmo ausência) das cadeias helicoidais, que são características observadas no meio natural
(RÖRIG et al., 2017), e, experimentos controlados em laboratório já mostraram que
diatomáceas possuem a capacidade de formar longas cadeias quando expostas a fortes
turbulências (DELL’AQUILA et al., 2017).
A partir do dia de inoculação das réplicas, em 7 e 14 dias, foram coletadas fotos para
análise do crescimento e da morfologia das algas.
3.5 ANÁLISE DOS DADOS
Alterações morfológicas e de crescimento foram avaliadas através de fotografias
retiradas com uso do software Olympus cellSens Dimension. Foram analisadas a presença ou
ausência de espinhos e seu tamanho, comprimento da base até o final do espinho,

20
conformação das cadeias de células, vista das células e possíveis deformações celulares; a fim
de verificar se o tratamento com turbulência determina diferenças nesses parâmetros.
3.6 DESENVOLVIMENTO DO EXPERIMENTO
No dia 18 de janeiro de 2021, foram inoculados 1,5 ml de um cultivo em fase
exponencial de A. glacialis no laboratório utilizando-se de 250 ml de meio de cultivo F/2 em
seis erlenmeyers de 500 ml, sendo esses divididos em dois grupos de três frascos cada (Figura
3A). Um grupo foi colocado em aeração (CB) e outro mantido estático (SB), sob as mesmas
condições de luminosidade e temperatura. Os frascos ficaram armazenados no cepário, sob
fotoperíodo de 12 horas claro e 12 horas escuro, com lâmpadas de irradiância de 80
μmol fótons m-2 s-1 e sistema de condicionamento de ar ligado 24 horas.
Neste mesmo dia foram retiradas 6 amostras de 1,5 ml, uma de cada um dos frascos, e
foram colocadas em microtubos do tipo Eppendorf e fixadas com 10 gotas de formol cada. Estas
são as amostras recém-inoculadas (t = 0).
Após isso, em 25 de janeiro de 2021 (Figura 3B e Figura 3C) e em 01 de fevereiro de
2021 (Figura 3D e Figura 3E), foram também recolhidas 6 amostras, uma de cada frasco, e
fixadas em formol, seguindo o mesmo protocolo citado acima. As amostras recolhidas em 25
de janeiro são o (t = 1) e as amostras recolhidas em 01 de fevereiro são o (t = 2). Estas amostras
foram homogeneizadas antes de serem coletadas e fixadas, para melhor seleção dos espécimes.
A temperatura do cepário ao longo das semanas deste estudo variou entre 19,4 °C e 28,3 °C.
Existe também um grupo de amostras chamado de Amostras Pré-Experimento, que
são amostras de cepas que já estavam em cultivo no cepário, anteriores à idealização e execução
deste trabalho. Nessas cepas, temos cultivos com meio de cultivo F/2, meio de cultivo IMR,
cultivos com aeração e cultivos sem aeração. Também foram retiradas alíquotas das Amostras
Pré-Experimento e fixadas em formol, assim como explicado anteriormente, para que fosse
possível realizar fotografias e fazer uma comparação desses cultivos antigos com os cultivos
próprios deste estudo.
As amostras foram fixadas para posterior fotografia e análise celular, ao longo das
semanas seguintes. Todas as fotos deste trabalho foram realizadas no aumento de 400x.
Em função da pandemia e das medidas sanitárias, foram feitas escalas de horário e
cronograma para uso do LAFICmu por todos os alunos, portanto, nem todas as amostras que

21
foram fixadas puderam ser fotografadas, em função do tempo disponível. A quantidade de
fotografias variou entre as amostras.
As amostras que não foram fotografadas estão representadas por um asterisco (*) nas
tabelas, visto que, ausência de dados não necessariamente representa medida zero.
Nas fotografias foram analisados: comprimento e largura dos espinhos, comprimento
e largura da base celular, aspecto dos espinhos e da base (Figura 2), vista celular e também
possíveis deformações celulares. Também foram analisados neste estudo os extremos
fenotípicos das células, nomeados como ‘’recordes morfométricos’’. Os recordes
morfométricos são os valores máximos e mínimos encontrados para as partes medidas dos
organismos, conforme figura abaixo.
Figura 2 - Dados morfométricos adotados para a caracterização morfológica das células de
Asterionellopsis glacialis cultivadas
Fonte: Autora (2021)

22
4 RESULTADOS
4.1 ASPECTO GERAL DOS CULTIVOS EXPERIMENTAIS:
A aparência do cultivo mudou ao longo do experimento. No início (Figura 3A) os frascos
possuíam conteúdo incolor, porém, com o passar dos dias o líquido começa a ganhar cor (Figura
3B e 3C). Ao fim do experimento (Figura 3D e 3E) o meio estava de coloração marrom.
Figura 3 - Disposição dos frascos experimentais de crescimento de Asterionellopsis glacialis
na sala de cultivos do LAFIC. (A) Cultivos recém inoculados (t = 0). (B) Cultivo (t = 1) antes
da homogeneização. (C) Cultivo (t = 1) depois da homogeneização. (D) Cultivo (t = 2) antes
da homogeneização (E) Cultivo (t = 2) depois da homogeneização.
Fonte: Autora (2021)

23
4.2 ANÁLISE MICROSCÓPICA DOS CULTIVOS
4.2.1 CULTIVOS PRÉ-EXPERIMENTAIS
Nas lâminas analisadas para as amostras dos cultivos antes dos experimentos
(inóculos) ocorreram células de todas as conformações, desde células saudáveis (Figura 4A)
incluindo células com tamanho reduzido e com deformações ou formatos estranhos (Figura 4B
e 4C).
Em relação aos dados morfométricos, a menor medida de largura do espinho entre
todas as lâminas analisadas neste estudo, foi encontrado na Amostra 19, com 0,58 μm; e o maior
comprimento de espinho com 22,88 μm na Amostra 23 (Tabela 1). Na Amostra 22 pode-se até
observar o cloroplasto da célula em vista valvar (Figura 4D).
Figura 4 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras Pré-Experimento.
(A) Cadeia saudável. (B, C) Células com deformações. (D) Célula solitária saudável, com o
cloroplasto destacado ao centro.
Fonte: Autora (2021)
Tabela 1 - Informações sobre as Amostras Pré-experimento: quantidade de células
fotografadas, média de medidas e recordes de medidas no grupo.
Amostras pré-
experimento (número
e identificação da
amostra)
Quantidade
de células
fotografadas
Média de medidas (μm) Recordes de
medidas mín. e
máx. (μm)
A B C D A B C D
19) AST IMR 7 DIAS
CB
11 2,13 16,32 9,32 7,17 0,58
24
20) AST F/2 15 DIAS
CB
3 1,95 13,81 9,57 8,38
21) AST A * * * * *
22) AST B 15 2,02 12,58 8,87 6,52
23) AGL IMR
30/12/20 CB
11 1,65 15,71 10,19 6,79 22,88
24) AGL F/2 23/12/20
CB
16 2,07 11,67 9,93 8,67
LEGENDA
VALOR MÍNIMO ENCONTRADO
VALOR MÁXIMO ENCONTRADO
Fonte: Autora (2021)
4.3 AMOSTRAS T = 0 (18/01/21)
Com o conteúdo que foi fixado anteriormente nos microtubos foram feitas lâminas
para observação no microscópio. As lâminas analisadas para o grupo (t = 0) mostravam poucas
células (Figura 5A), provavelmente por ser o início dos cultivos e os 1,5 ml de células
inoculadas estarem bem diluídos nos 250 ml de meio de cultivo. Foram encontradas células
aparentemente saudáveis (Figuras 5B), embora com alguns espinhos tortuosos, e também foram
vistas células com deformações (Figura 5C). Nas amostras do grupo (t = 0) não foram vistos
recordes morfométricos (Tabela 2).
Figura 5 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t = 0). (A)
Células escassas nas lâminas analisadas (B) Cadeia em formato helicoidal. (C) Célula com
deformação.
Fonte: Autora (2021)
25
Tabela 2 - Informações sobre as Amostras (t = 0): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo.
Amostras (t = 0) Quantidade de
células
fotografadas
Média de medidas (μm) Recordes de
medidas mín. e
máx. (μm)
A B C D A B C D
1) AGL F/2 A-CB 10 2,35 12,32 9,22 9,16
2) AGL F/2 B-CB 6 2,4 12,66 8,83 9,16
3) AGL F/2 C-CB * * * * *
4) AGL F/2 A-SB 5 1,78 9,52 9,56 9,74
5) AGL F/2 B-SB * * * * *
6) AGL F/2 C-SB * * * * *
LEGENDA
VALOR MÍNIMO ENCONTRADO
VALOR MÁXIMO ENCONTRADO
Fonte: Autora (2021)
4.4 AMOSTRAS T = 1 (25/01/21)
Nas lâminas analisadas para o (t = 1) foram vistos alguns agrupamentos sem padrão
aparente (Figura 6A), sem ligação das células por suas bases, não formando a tradicional cadeia
que é observada em células saudáveis, ou seja, formação de agregados. Foram vistas também
poucas colônias organizadas quando comparadas com a quantidade de células solitárias e
agrupamentos sem padrões. A maioria das células aparece em vista conectival. Das 168 células
analisadas ao longo de todo este estudo, 130 foram encontradas em vista conectival, em
detrimento de 38 em vista valvar (Figura 6C). Na vista valvar a base da célula aparenta ser mais
arredondada, enquanto que na vista conectival mais triangular (Figura 10).
Em diversas lâminas foi possível de se observar outro organismo, de formato alongado
com as extremidades arredondadas, possivelmente uma bactéria, demonstrando que o meio foi
contaminado por outra cultura, como pode ser visto na Amostra 9 (Figura 6D).
As células mais saudáveis, incluindo as colônias helicoidais ou estelares descritas pela
literatura para a espécie, foram encontradas na Amostra 11 (Figura 6B).
Foi observado também neste momento, a diferença de coloração entre os frascos nos
momentos (t = 0) (Figura 3A) e (t = 1) (Figura 3B e Figura 3C). No curto período de uma
semana, nota-se grande diferença na coloração dos meios, demonstrando a taxa rápida de

26
crescimento celular. Analisando somente o grupo (t =1) é possível de se notar uma diferença
de coloração entre os frascos que estão em aeração, CB, e os frascos que estão estáticos, SB. O
grupo que está em aeração, à esquerda nas Figuras 3B e 3C, apresenta coloração mais escura
do que o grupo que está em repouso. Entretanto, quando foram analisadas as lâminas para o
cultivo em aeração e o cultivo estático no grupo (t = 1), não foram vistas muitas diferenças em
termos de encontrar com maior facilidade as células.
Em termos de dados morfométricos, na Amostra 7 foram encontrados valores recordes
de mínimo e de máximo para largura e comprimento do espinho e largura e comprimento da
base das células de A. glacialis. A largura do espinho, o comprimento da base, e a largura da
base atingiram os valores máximos encontrados de 7,42 μm; 17,42 μm; 18,47 μm;
respectivamente. Já o comprimento do espinho atingiu o valor mínimo recorde de 4,02 μm
(Tabela 3). Em algumas células, os espinhos ainda estão ausentes, ou são pequenos demais para
serem medidos com precisão, mesmo após uma semana de desenvolvimento, como pode se
observar na Amostra 8 (Figura 6E).
Figura 6 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t =1). (A)
Agregados de células. (B) Colônia saudável. (C) Células em vista valvar e em vista
conectival, da esquerda para a direita, respectivamente. (D) Cultivo contaminado por bactéria.
(E) Cadeia com espinhos com crescimento incompleto.
Fonte: Autora (2021)
27
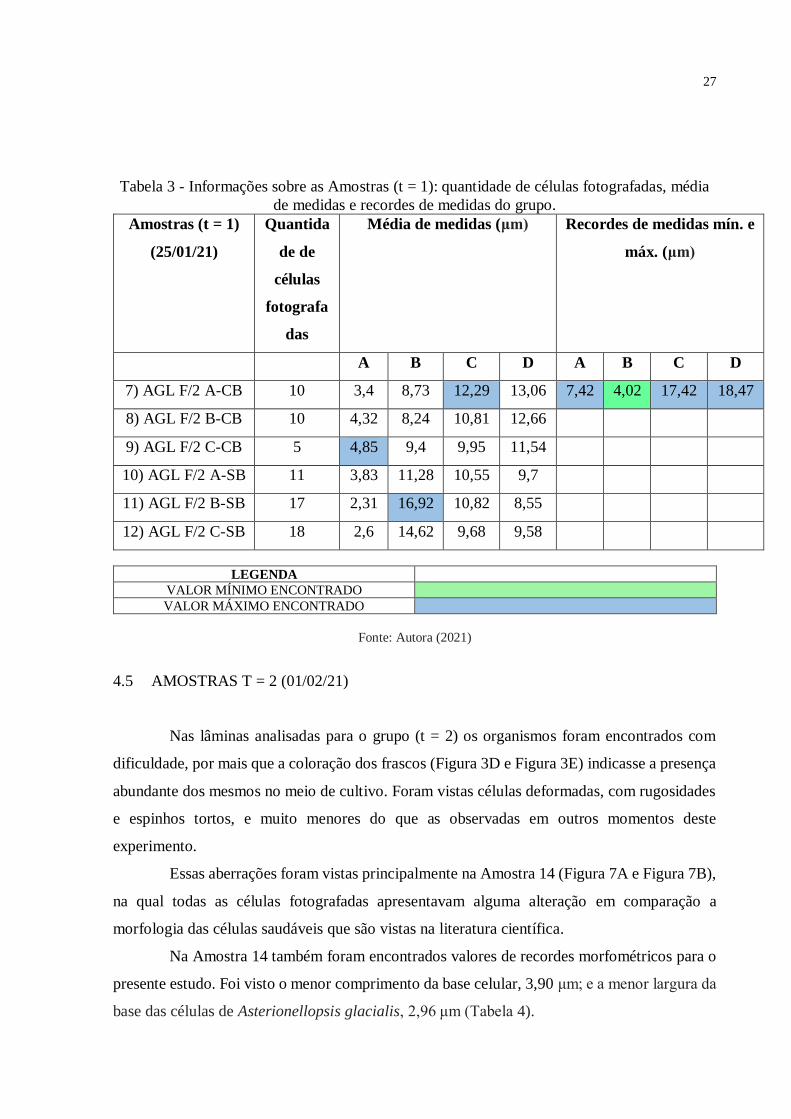
Tabela 3 - Informações sobre as Amostras (t = 1): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo.
Amostras (t = 1)
(25/01/21)
Quantida
de de
células
fotografa
das
Média de medidas (μm) Recordes de medidas mín. e
máx. (μm)
A B C D A B C D
7) AGL F/2 A-CB 10 3,4 8,73 12,29 13,06 7,42 4,02 17,42 18,47
8) AGL F/2 B-CB 10 4,32 8,24 10,81 12,66
9) AGL F/2 C-CB 5 4,85 9,4 9,95 11,54
10) AGL F/2 A-SB 11 3,83 11,28 10,55 9,7
11) AGL F/2 B-SB 17 2,31 16,92 10,82 8,55
12) AGL F/2 C-SB 18 2,6 14,62 9,68 9,58
LEGENDA
VALOR MÍNIMO ENCONTRADO
VALOR MÁXIMO ENCONTRADO
Fonte: Autora (2021)
4.5 AMOSTRAS T = 2 (01/02/21)
Nas lâminas analisadas para o grupo (t = 2) os organismos foram encontrados com
dificuldade, por mais que a coloração dos frascos (Figura 3D e Figura 3E) indicasse a presença
abundante dos mesmos no meio de cultivo. Foram vistas células deformadas, com rugosidades
e espinhos tortos, e muito menores do que as observadas em outros momentos deste
experimento.
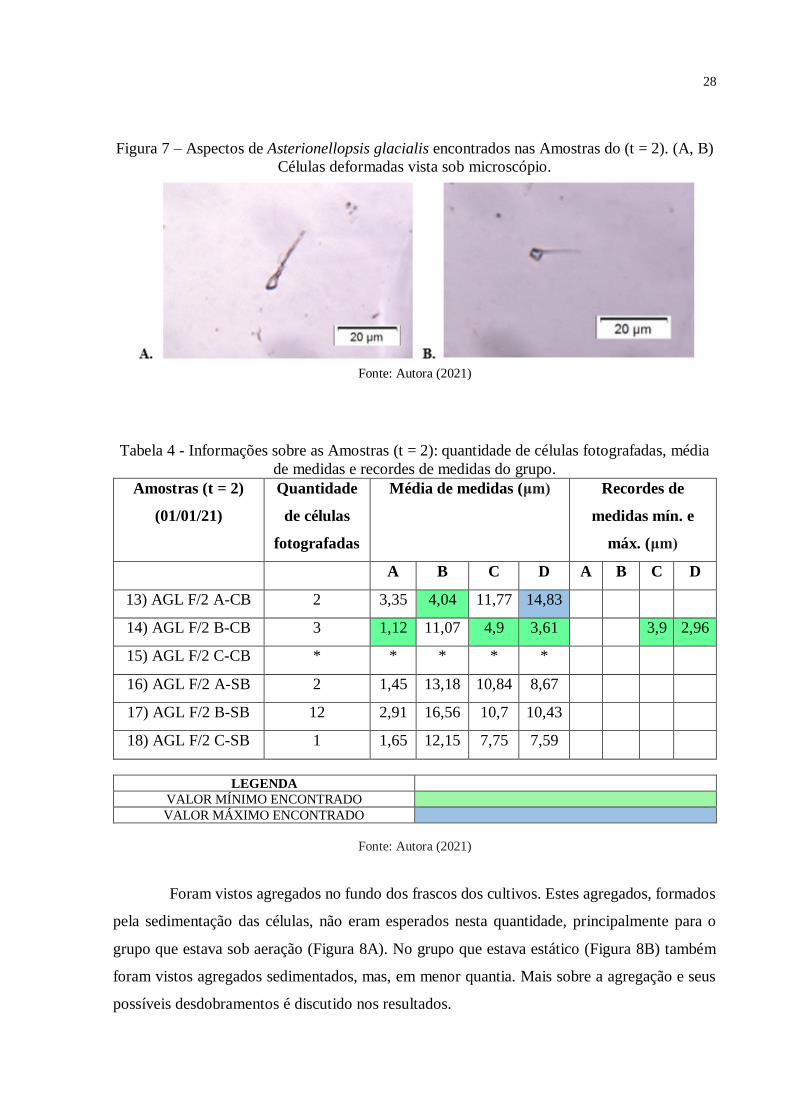
Essas aberrações foram vistas principalmente na Amostra 14 (Figura 7A e Figura 7B),
na qual todas as células fotografadas apresentavam alguma alteração em comparação a
morfologia das células saudáveis que são vistas na literatura científica.
Na Amostra 14 também foram encontrados valores de recordes morfométricos para o
presente estudo. Foi visto o menor comprimento da base celular, 3,90 μm; e a menor largura da
base das células de Asterionellopsis glacialis, 2,96 μm (Tabela 4).
28
Figura 7 – Aspectos de Asterionellopsis glacialis encontrados nas Amostras do (t = 2). (A, B)
Células deformadas vista sob microscópio.
Fonte: Autora (2021)
Tabela 4 - Informações sobre as Amostras (t = 2): quantidade de células fotografadas, média
de medidas e recordes de medidas do grupo.
Amostras (t = 2)
(01/01/21)
Quantidade
de células
fotografadas
Média de medidas (μm) Recordes de
medidas mín. e
máx. (μm)
A B C D A B C D
13) AGL F/2 A-CB 2 3,35 4,04 11,77 14,83
14) AGL F/2 B-CB 3 1,12 11,07 4,9 3,61 3,9 2,96
15) AGL F/2 C-CB * * * * *
16) AGL F/2 A-SB 2 1,45 13,18 10,84 8,67
17) AGL F/2 B-SB 12 2,91 16,56 10,7 10,43
18) AGL F/2 C-SB 1 1,65 12,15 7,75 7,59
LEGENDA
VALOR MÍNIMO ENCONTRADO
VALOR MÁXIMO ENCONTRADO
Fonte: Autora (2021)
Foram vistos agregados no fundo dos frascos dos cultivos. Estes agregados, formados
pela sedimentação das células, não eram esperados nesta quantidade, principalmente para o
grupo que estava sob aeração (Figura 8A). No grupo que estava estático (Figura 8B) também
foram vistos agregados sedimentados, mas, em menor quantia. Mais sobre a agregação e seus
possíveis desdobramentos é discutido nos resultados.
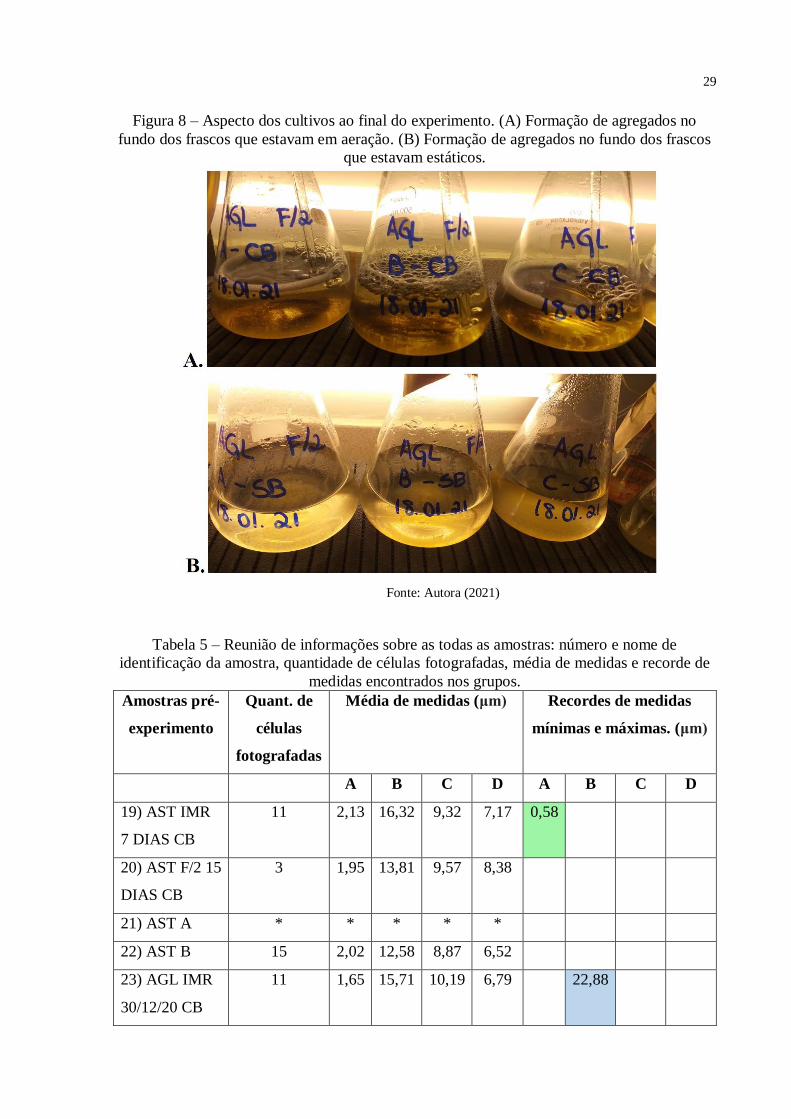
29
Figura 8 – Aspecto dos cultivos ao final do experimento. (A) Formação de agregados no
fundo dos frascos que estavam em aeração. (B) Formação de agregados no fundo dos frascos
que estavam estáticos.
Fonte: Autora (2021)
Tabela 5 – Reunião de informações sobre as todas as amostras: número e nome de
identificação da amostra, quantidade de células fotografadas, média de medidas e recorde de
medidas encontrados nos grupos.
Amostras pré-
experimento
Quant. de
células
fotografadas
Média de medidas (μm) Recordes de medidas
mínimas e máximas. (μm)
A B C D A B C D
19) AST IMR
7 DIAS CB
11 2,13 16,32 9,32 7,17 0,58
20) AST F/2 15
DIAS CB
3 1,95 13,81 9,57 8,38
21) AST A * * * * *
22) AST B 15 2,02 12,58 8,87 6,52
23) AGL IMR
30/12/20 CB
11 1,65 15,71 10,19 6,79 22,88
30
24) AGL F/2
23/12/20 CB
16 2,07 11,67 9,93 8,67
Amostras t = 0
1) AGL F/2 A-
CB
10 2,35 12,32 9,22 9,16
2) AGL F/2 B-
CB
6 2,4 12,66 8,83 9,16
3) AGL F/2 C-
CB
* * * * *
4) AGL F/2 A-
SB
5 1,78 9,52 9,56 9,74
5) AGL F/2 B-
SB
* * * * *
6) AGL F/2 C-
SB
* * * * *
Amostras t = 1
7) AGL F/2 A-
CB
10 3,4 8,73 12,29 13,06 7,42 4,02 17,42 18,47
8) AGL F/2 B-
CB
10 4,32 8,24 10,81 12,66
9) AGL F/2 C-
CB
5 4,85 9,4 9,95 11,54
10) AGL F/2
A-SB
11 3,83 11,28 10,55 9,7
11) AGL F/2
B-SB
17 2,31 16,92 10,82 8,55
12) AGL F/2
C-SB
18 2,6 14,62 9,68 9,58
31
Amostras t = 2
13) AGL F/2
A-CB
2 3,35 4,04 11,77 14,83
14) AGL F/2
B-CB
3 1,12 11,07 4,9 3,61 3,9 2,96
15) AGL F/2
C-CB
* * * * *
16) AGL F/2
A-SB
2 1,45 13,18 10,84 8,67
17) AGL F/2
B-SB
12 2,91 16,56 10,7 10,43
18) AGL F/2
C-SB
1 1,65 12,15 7,75 7,59
LEGENDA
VALOR MÍNIMO ENCONTRADO
VALOR MÁXIMO ENCONTRADO
A LARGURA DO ESPINHO
B COMPRIMENTO DO ESPINHO
C COMPRIMENTO DA BASE
D LARGURA DA BASE
CB COM AERAÇÃO
SB SEM AERAÇÃO
* AMOSTRAS NÃO MEDIDAS
Fonte: Autora (2021)
4.6 MORFOLOGIA DOS ESPINHOS
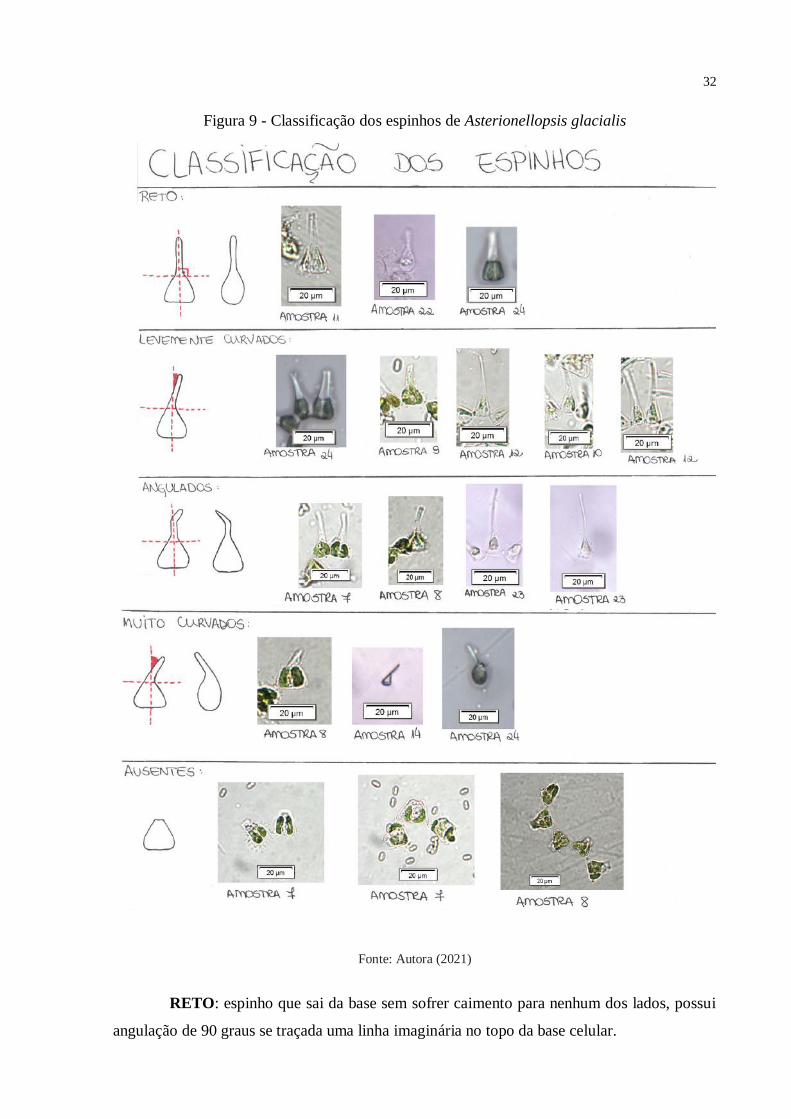
Os espinhos de Asterionellopsis glacialis foram encontrados em 5 morfotipos
diferentes, que foram classificados pela autora como: reto, levemente curvado, muito curvado,
angulado e ausente. Das 168 células analisadas neste estudo, em relação a morfologia dos
espinhos: 102 eram retos, 39 levemente curvados, 17 angulados, 6 muito curvados e em 4
células os espinhos ainda estavam ausentes ou não tinham completado seu processo de
crescimento total. Ou seja, 60,71% das células de Asterionellopsis glacialis analisadas
possuíam espinhos retos. Abaixo, um diagrama (Figura 9) para melhor diferenciação na
classificação dos espinhos:
32
Figura 9 - Classificação dos espinhos de Asterionellopsis glacialis
Fonte: Autora (2021)
RETO: espinho que sai da base sem sofrer caimento para nenhum dos lados, possui
angulação de 90 graus se traçada uma linha imaginária no topo da base celular.

33
LEVEMENTE CURVADO: o espinho entorta desde o início até o final do seu
crescimento.
MUITO CURVADO: possui um ângulo de inclinação do espinho igual ou maior que
45 graus, se traçada uma linha imaginária no topo da base célula. Ele é muito curvado em
relação à saída do espinho da base.
ANGULADO: o espinho angulado começa a se desenvolver de forma igual ao espinho
reto, porém, durante o processo de desenvolvimento ele passa a entortar para a direita ou para
a esquerda, dependendo da célula. No caso dos espinhos angulados, provavelmente, no início
do desenvolvimento o meio de cultivo apresentava todas as características necessárias para o
organismo crescer de forma saudável, porém, ao longo dos dias, uma das variáveis (nutrientes,
pH, irradiância, temperatura, etc.) sofre uma alteração e faz com que o espinho comece a
apresentar alterações em seu formato.
AUSENTE: como o próprio nome indica, nestas células não encontramos espinhos,
ou ainda não começaram o processo de crescimento. Ou, eram tão mínimos, que não era
possível de se medir com precisão.
As amostras que apresentaram maior diversidade de espinhos foram: Amostra 7 com
células com espinhos retos, levemente curvados, angulados e ausentes; Amostra 24 com células
com espinhos retos, levemente curvados, angulados e muito curvados e a Amostra 22 com
células com espinhos retos, levemente curvados, angulados e muito curvados.
Figura 10 – Vista conectival e vista valvar encontradas nas amostras de A. glacialis do
presente estudo. À esquerda, células em vista conectival, e, à direita, células em vista valvar.
Fonte: Autora (2021)

34
5 DISCUSSÃO
5.1 Morfologia da cepa estudada de acordo com padrões vistos na literatura
Considerando a literatura já existente sobre a diatomácea Asterionellopsis glacialis,
deve-se observar algumas características interessantes para análise no presente estudo. De
acordo com Tomas (1997), o gênero Asterionellopsis pode ser caracterizado por:
- Células com extremidades diferentes nas visualizações valvares e laterais;
- Células unidas por faces valvares dos polos expandidos, em cadeias com formato
estelar;
- Polo basal triangular na vista lateral e mais ou menos arredondados em vista valvar;
- Podem ser encontradas no plâncton de regiões de clima gelado à águas temperadas
costeiras.
Em relação aos dados morfométricos, de acordo com Cupp (1943) a espécie possui as
seguintes características:
- Eixo apical (ou comprimento): 30 – 150 μm;
- Comprimento da parte expandida: 10 - 23 μm;
- Eixo transapical da parte expandida (ou largura): 8 - 12 μm;
- Um ou dois cloroplastos na região basal aumentada das células.
A presença de espinhos e de outros processos acessórios foi fundamental para fornecer
a ancoragem necessária à vida no bentos de mares rasos, naturalmente mais instáveis (VIEIRA,
2011).
5.2 Agregação, floculação e sedimentação das células nos cultivos experimentais
Agregação, floculação e sedimentação são conceitos que estão conectados entre si.
Células de microalgas podem se agregar, ou seja, se unirem, gerando assim flocos, que diferem
das colônias e cadeias. Colônias e cadeias são feições normais em microalgas e apresentam
diferentes funções como aumento de tamanho para diminuir a pressão de pastagem pelo
zooplâncton (KENITZ et al., 2020) e aumento da sua flutuabilidade, quando a forma das células
em colônias gera maior atrito com a água. Os agregados, ou flocos, por sua vez, são feições
anômalas que ocorrem com células em situações de estresse. Os movimentos da água geram
choques e adesões entre as células. Nesse caso, pela falha de processos de flutuação

35
relacionados a troca de íons e formação de vacúolos, ou ainda pela deformação das estruturas
celulares externas, os agregados tendem a sedimentar e aderir ao substrato, o que normalmente
antecede a morte celular.
As diatomáceas, sendo organismos sem motilidade, desenvolveram ao longo do seu
processo evolutivo diferentes meios para regular sua densidade e posição na coluna d’água
(ARRIETA et al., 2020). De acordo com o autor, a regulação da flutuabilidade destes
organismos não muda somente as distribuições populacionais, como também pode modificar a
forma destas microalgas. O afundamento celular deste grupo é facilitado em comparação com
outros grupos de fitoplâncton devido à uma de suas características principais: a parede celular,
que é fina, porém pesada, de sílica. A velocidade do afundamento varia em função do tamanho
das células (ARRIETA et al., 2020).
Agregados de diatomáceas são formados principalmente durante florações. Os gêneros
mais comuns encontrados são: Nitzschia¸ Chaetoceros, Rhizosolenia, Leptocylindrius,
Skeletonema e Thalassionema (THORNTON, 2002). Este autor afirma que esses agregados se
formam quando diatomáceas colidem umas com as outras e se unem. A frequência das colisões
depende da turbulência da água e do tamanho, formato e concentração das células (CROCKER;
PASSOW, 1995).
A capacidade de uma diatomácea de se aderir em outra é afetada pela presença de
substâncias poliméricas extracelulares (Exopolissacarídeos - EPS). Experimentos de
laboratório têm demonstrado que diatomáceas produzem mais EPS sob limitação de nutrientes.
Em habitats bênticos e planctônicos, a produção de EPS leva a diatomácea a se tornar mais
aderente e consequentemente agregar com maior facilidade (THORNTON, 2002).
Partículas exopoliméricas transparentes (TEP), também são importantes para a
agregação de diatomáceas em floração (PASSOW, 1994), embora a presença de TEP na coluna
d’água não necessariamente signifique que a agregação vai acontecer (THORNTON, 2011). A
liberação desses exsudatos depende das condições ambientais (MYKLESTAD, 1995; KAHL,
2008). Logan et al. (1995) apresentou evidências indicando que a TEP, mais do que as próprias
células das diatomáceas, são os agentes primários das agregações, e diferentes espécies de
diatomáceas podem agregar de diferentes formas (ALLDREDGE et al., 1995).
A limitação de nutrientes tem mostrado aumentar a capacidade aderente das
diatomáceas em culturas (DRAPEAU et al., 1994), e a produção de TEP em Chaetoceros
calcitrans foi maior sob limitações de nitrato do que em cultivos com condições de nutrientes
suficientes (CORZO et al., 2000). Concentrações de cálcio (Ca+2) e magnésio (Mg+2) afetam as
propriedades físico-químicas dos EPSs e dos TEPs (DECHO, 1990). Logan e Alldregde (1989)

36
sugeriram que a agregação é uma adaptação para aumentar a absorção de nutrientes durante
condições de escassez de nutrientes, uma vez que os agregados teriam vantagem sobre as
células não agregadas. A maior aderência dos TEPs aumenta a densidade e consequentemente
os índices de sedimentação das células (PASSOW et al., 2001; LA ROCHA et al., 2007).
Nitzschia angularis agrega mais facilmente durante a fase exponencial, entretanto se a
cultura é mantida por mais alguns dias, durante a fase log, a floculação começa a diminuir.
Thalassiosira weissflogii não agrega nem durante a fase de senescência (LOGAN et al., 1994).
Durante o crescimento, células de Chaetoceros gracilis geram TEP (ALLDREDGE et al.,
1993). No estudo de Alldredge et al. (1995) os principais agregados encontrados foram de
Chaetoceros e Thalassiosira.
Bactérias também podem produzir TEP (DECHO, 1990; AZAM, 1994). Bactérias
interagindo com o fitoplâncton podem influenciar a formação de TEPs e a agregação
fitoplanctônica e sua abundância, assim como aumentar a eficiência de processos biológicos
(GÄRDES et al., 2010). Grossart (2006) mostrou que bactérias heterotróficas influenciam o
desenvolvimento e agregação de diatomáceas marinhas como T. rotula e Skeletonema costtum.
As bactérias podem estar ligadas à superfície das algas (KOGURE, 1981) ou podem ocorrer de
forma intracelular como organismos simbiontes (LEWIS, 2001). De acordo com Gärdes (2010),
algumas espécies de bactérias são capazes de induzir agregados em diatomáceas. Sob condições
in vitro definidas cepas de bactérias ligadas à Thalassiosira weissflogii são necessárias para a
produção de TEP e de agregados. Sob condições naturais, bactérias marinhas podem se
beneficiar da fotossíntese das algas interagindo com as mesmas, influenciando na formação de
TEP e agregados ou utilizando a matéria orgânica dissolvida liberada durante a hidrólise dos
exsudatos e TEPs.
Em relação à sedimentação, surpreende a quantidade de material que foi visto nos
meios com aeração. Já que, a aeração e consequente turbulência da água ajuda que as células
não sedimentem com tanta facilidade. Nos meios em repouso é vista menor quantidade de
material sedimentado, entretanto, a quantidade pode ser função da quantidade de células vivas
crescidas, que foi bem maior (crescimento mais rápido) nos cultivos aerados do que nos
estáticos. De qualquer forma, não era esperada essa quantidade de sedimentação, pois com a
turbulência esperava-se uma constante ressuspensão das populações. Por outro lado, a
mortalidade das células e sua decomposição também pode explicar a sedimentação e adesão
dos resíduos celulares ao fundo do frasco.
É plausível que, para uma melhor análise deste experimento, as amostras deveriam ter
sido recolhidas e fixadas a cada 2 dias, ao invés de semanalmente, já que o material cresceu e

37
sedimentou rapidamente. Porém, não havia como saber a velocidade da sedimentação antes que
o experimento fosse realizado. Determinar quais são os fatores que influenciam a sedimentação
é fundamental, pois diversas áreas das ciências naturais utilizam a sedimentação como uma
variável importante (LAROCQUE et al., 1996).
A floculação com sucesso e rápida sedimentação em determinadas espécies de algas
ocorre apenas na presença de Mg+2 e em elevados níveis de pH. O aumento da floculação é
notado com o aumento do pH (SMITH; DAVIS, 2012). O cloreto de cálcio (CaCl2) é um agente
floculante para a Chlorella, estudada por Morioka et al. (2014) e foi escolhido o meio BBM
para seu cultivo. Sua sedimentação foi máxima quando o pH estava entre 9,5 e 11,5. O cloreto
de cálcio também possui efeito floculante nos cultivos de Chlamydomonas reinhardtii. As
células desta espécie flocularam rapidamente com a adição do componente (SCHOLZ et al,
2011). No caso de Chlorella vulgaris, não foi o cálcio que induziu a floculação, e sim o
magnésio, em pH 11. Entretanto, deve ser tomado cuidado para que o alto pH não destrua as
células das algas (VANDAMME et al., 2012). A floculação também pode ser induzida pela
precipitação de hidróxido de magnésio (Mg(OH)2) em valores de pH entre 9,5 e 11,5 (YAHI et
al., 1994).
Foi observado por Nurdogan e Oswald (1995) que em tanques de cultivo de
microalgas, a floculação era atingida pela adição de cálcio, na forma de óxido de cálcio (CaO),
ao meio de cultivo. A autofloculação não ocorria se não existissem íons de cálcio suficientes na
água.
De acordo com Smith et al. (2012) apud Morioka (2014, p. 47), a presença de sais de
cálcio ou magnésio, que apresentam caráter básico, favorece a formação de flocos microalgais
pois a carga positiva do cloreto neutraliza a carga negativa presente na superfície das algas,
induzindo a agregação das células.
Sabe-se, então, que tanto o cálcio quanto o magnésio desempenham um papel
importante para a agregação, floculação e sedimentação algal. No meio de cultivo utilizado para
a Asterionellopsis glacialis temos a presença de cálcio e de magnésio, porém em pouca
quantidade, aparecendo na água do mar que é adicionada ao cultivo. O cálcio na água do mar
se encontra na forma de sulfato de cálcio (CaSO4) e o magnésio na forma de sulfato de magnésio
(MgSO4). O meio F/2 apresenta pH em torno de 8, porém, com o crescimento dos cultivos e
incremento da fotossíntese ocorre incremento gradual do pH nos frascos de cultivo.
O silicato utilizado no laboratório para fazer os meios de cultivo, inclusive o meio F/2
utilizado neste trabalho é silicato sintético, portanto deve-se considerar a solubilidade deste

38
composto como problema na agregação, já que é o elemento base da composição das
diatomáceas, podendo sua indisponibilidade gerar deformações nas frústulas.
Embora o cálcio e o magnésio desempenhem papel importante para a floculação e
sedimentação de Chlorella spp. e de Chlamydomonas reinhardtii isso não significa que serão
estes os fatores a influenciar a agregação, floculação e sedimentação em A. glacialis, visto que,
embora todos esses organismos citados sejam algas, pertencem a grupos diferentes. Enquanto
a Chlorella e a Chlamydomonas são clorófitas, Asterionellopsis glacialis é uma diatomácea, e
portanto, os processos podem ocorrer de formas diferentes.
Um ponto que muitos autores concordam, é que os níveis de floculação aumentam com
o aumento do pH. De acordo com Honorato (2014), o pH do meio de cultura possui a tendência
de aumentar com o aumento da densidade celular e com a idade do cultivo, devido ao consumo
de CO2 por parte das microalgas. Entretanto, os elementos-chave que vimos ser os catalisadores
para induzir a sedimentação ainda são o cálcio e o magnésio. Ora apenas o cálcio é importante,
ora apenas o magnésio, e ora os dois em conjunto são importantes. O aumento de pH de um
meio de cultivo pode levar à floculação, e a floculação tem a capacidade de induzir o
crescimento bacteriano, já visto na (Figura 6D).
Como comentado acima, uma hipótese que pode ser levantada sobre o que ocorreu
durante este experimento seria o aumento natural do pH do meio de cultivo, causando
agregação, floculação e sedimentação das algas. É sabido que o processo fotossintético resulta
no aumento do pH nos meios de cultivo.
Entretanto, não é possível confirmar se o que aconteceu de fato foi o aumento do pH,
pois ele não foi medido após o início do processo de sedimentação, apenas foi medido no
momento em que estavam sendo feitos os meios de cultivo. Porém, como o aumento do pH é
um fator visto em diversos artigos sobre floculação e sedimentação, é algo importante a ser
considerado. Nutrientes também são fatores importantes, mas devem ser observados com
cautela visto que os nutrientes necessários para o desenvolvimento do organismo variam de
espécie para espécie (GUILLARD, 1975; SPILLING et al., 2018).
5.3 Considerações sobre a morfologia celular nos cultivos experimentais
Observam-se morfotipos diferentes dentro de uma mesma espécie. A existência de
diferentes morfotipos, e, em alguns casos, até deformações celulares (também chamadas de
formas teratológicas) pode se dar por problemas no metabolismo das algas, stress,
esgotamento de nutrientes ou contaminação bacteriana. Metais, compostos orgânicos e

39
contaminantes são descritos como possíveis estressores para diatomáceas (LAVOIE et al.,
2017). Múltiplos estressores podem ter efeitos cumulativos, que podem prejudicar funções
especificas e diferentes das células (FALASCO et al., 2021), e a presença de múltiplos
estressores pode aumentar significativamente a proporção de indivíduos deformados
(LAVOIE et al., 2017). Entretanto, quando as deformações acontecem, ou por quais motivos
as mesmas acontecem, ainda não é completamente claro. Crescimentos teratológicos em
culturas de diatomáceas são observados desde o século 19 por Miquel (1890-1892), que
mostrou que mudanças na composição química dos meios de cultura induzia o crescimento de
células anormais. Uma possível explicação para a formação de teratologias envolve o sistema
microtubular, uma parte ativa das células de diatomáceas. Exposição à componentes que
afetem este sistema, levam a formação anormal do núcleo celular durante a divisão das células
e levam também a deformação da nova valva (SCHMID, 1980; LAVOIE et al., 2017).
O crescimento em condições artificiais e a contaminação por metais pesados são as
causas mais conhecidas para o aparecimento de formas teratológicas até o momento. Cobre,
cádmio e zinco aparentam ser os metais traço mais representativos para a produção de células
anormais (FALASCO et al., 2009). Em Asterionella japonica, um sinônimo para
Asterionellopsis glacialis, nota-se deformação dos contornos das valvas das células, com
aumento do tamanho celular, induzido por cobre, zinco e cádmio (FALASCO et al., 2021).
De acordo com o autor, baseado no trabalho de Fisher et al. (1981) concentrações de 5 a 25
microgramas (μg) de cobre por litro, teriam a capacidade de influenciar os cultivos. Na receita
do meio de cultivo F/2, utilizado para a Asterionellopsis glacialis, nota-se a presença de 2,5
μg de sulfato de cobre.
Embora alguns metais traço como ferro, manganês, zinco e cobre, sejam nutrientes
essenciais para a fotossíntese e outros processos ao longo do ciclo de vida das algas
(ANDERSEN, 2005), a presença destes metais nos meios de cultivo deve ser observada com
atenção. Pois, quando estes metais ultrapassam as concentrações ótimas, ou, quando estão
abaixo dos níveis mínimos para o organismo em questão, ou seja, a deficiência ou o excesso
de determinados elementos pode danificar diversos processos metabólicos algais (SU et al.,
2018), podendo levar, até, a morte das células (MASMOUDI et al., 2013; JONES et al.,
2018). Características químicas da água como pH e carbono orgânico dissolvido podem afetar
a biodisponibilidade dos metais (FERNÁNDEZ et al., 2018).
Teratologias ao longo do tempo podem influenciar populações ou espécies. Eventos
temporais longos de deformações podem levar a eventos de especiação. O ganho ou perda de
uma estrutura, ou, mesmo a simples mudança de uma estrutura de posição pode,

40
potencialmente, representar uma nova espécie (LAVOIE et al., 2017). De acordo com Hustedt
(1956) e Granetti (1968), algumas alterações morfológicas não são induzidas por mudanças
genéticas, pois diatomáceas possuem a capacidade de retornar à sua forma ‘’padrão’’ no ciclo
sexual seguinte. O início da deformação das frústulas se dá, quando, uma valva inicial
anormal inicia o efeito cascata de deformações: argumenta-se que a deformidade inicial
aparece durante a reprodução sexual, quando a frústula de uma nova célula é formada sem a
presença da epiteca (HUSTEDT, 1956).
Diatomáceas deformadas em culturas são capazes de se reproduzir, entretanto, a
habilidade de sobrevivência e competição das células anormais no meio ambiente é
potencialmente afetada. Formas teratológicas possuem diferentes impactos em questões
fisiológicas e ecológicas dependendo das estruturas valvares que foram afetadas (LAVOIE et
al., 2017).
Dentro de uma mesma espécie, baseado nos resultados obtidos no presente trabalho,
podem ser vistos tipos de deformidades diferentes. Não seria estranho, então, afirmar que as
formas teratológicas se manifestam de formas diferentes, de acordo com a morfologia da
microalga em questão. Tratando-se de Asterionellopsis glacialis, por exemplo, pode-se
observar deformações de diferentes tipos nos espinhos, entretanto existem diatomáceas que
não possuem esse processo acessório, e mesmo assim podem apresentar deformações. Lavoie
et al. (2017) sugere que a formação de longas e estreitas valvas pode fornecer maiores
possibilidades da ocorrência de erros no desenvolvimento das células. Logo, seguindo esta
linha de pensamento, é possível de se afirmar que os espinhos de A. glacialis, ou seja, as
estruturas mais longas e estreitas deste organismo, são as estruturas mais sensíveis das
células. Os dados obtidos no presente trabalho corroboram com esta ideia, visto que, em
39,29% das amostras foram observados espinhos que não eram retos (que são os espinhos
descritos como saudáveis para esta microalga no meio ambiente), como pode ser observado
na Figura 8. Sob condições de estresse, A. glacialis expressa a mudança de alguma variável
importante para seu desenvolvimento em seus espinhos.
Em relação à base das células, tanto em vista valvar quanto em vista conectival, pelas
imagens que foram analisadas, não foram vistas alterações ou deformações. Apenas na
Amostra 20 (Figura 4C) é possível de se ver uma hachura na base da base, podendo ser uma
deformação. Entretanto, como foi visto este detalhe apenas em uma célula neste estudo, não
seria correto afirmar que é uma deformidade celular, visto que pode ser também fruto da
qualidade do vidro das lâminas ou mesmo de alguma característica do microscópio.

41
Em algumas células é possível de se observar os cloroplastos e em outras não, porém,
isso também pode ser apenas fruto da qualidade das lentes do microscópio e da configuração
do sistema de fotografias utilizado.
5.3.1 Dados morfométricos
A amostra que apresentou o maior número de “recordes” morfométricos em dados
absolutos, em largura e comprimento de espinhos, e, em largura e comprimento da base foi a
Amostra 7. A Amostra 14 também registrou dois recordes de medidas (Tabela 5).
Embora a maioria dos valores recordes morfométricos tenham sido encontrados na
Amostra 7 e Amostra 14, quando feitas médias aritméticas simples, são nas Amostras 13 e 14
que vemos os maiores e menores números. Entretanto, nem sempre uma média aritmética
reflete de forma verdadeira o processo que está acontecendo, já que, não foram tiradas
quantidades de fotos iguais de todas as amostras.
Quando comparados os valores encontrados nas células deste trabalho com os valores
encontrados como base no artigo de Cupp (1943), vemos que:
Tabela 6 – Comparação entre os dados obtidos no presente trabalho e os dados de Cupp
(1943), para diferentes medidas da célula de Asterionellopsis glacialis.
Dados morfométricos médios Cupp (1943) Autora (2021)
Eixo apical médio (B+C) 30 – 150 8,94 – 29,21
Comprimento da parte expandida médio (C) 10 – 23 4,9 – 12,29
Eixo transapical médio (D) 8 – 12 3,61 – 14,83
Dados morfométricos recordes Cupp (1943) Autora (2021)
Eixo apical médio (B+C) 30 – 150 7,92 – 40,3
Comprimento da parte expandida médio (C) 10 – 23 3,9 – 17,42
Eixo transapical médio (D) 8 – 12 2,96 – 18,47
Fonte: Autora (2021)
EIXO APICAL MÉDIO: é a soma das medidas B e C, vistas na (Figura 1). O valor
encontrado foi de 8,94 – 29,21 μm.

42
COMPRIMENTO DA PARTE EXPANDIDA MÉDIO: é a medida C, vista na
(Figura 1). O valor encontrado foi de 4,9 – 12,29 μm.
EIXO TRANSAPICAL DA PARTE EXPANDIDA MÉDIO: é a medida D, vista
na (Figura 1). O valor encontrado foi de 3,61 – 14,83 μm.
Figura 11 – Eixo apical e eixo transapical das células, conforme medidas determinadas na
(Figura 1).
Fonte: Autora (2021)
Ou seja, os dados morfométricos encontrados no presente estudo não foram tão
similares aos conhecidos anteriormente, demonstrando que talvez haja um problema nos
cultivos.
Sabe-se que as microalgas, como diatomáceas, por exemplo, são seres planctônicos e
sem motilidade, portanto, a turbulência no ambiente irá se encarregar de fornecer os nutrientes
necessários para as células, e, em condições turbulências, irá ocorrer um maior fluxo de
nutrientes por área em comparação a organismos de menor tamanho (KARP-BOSS et al., 1996;
PAHLOW et al., 1997). Porém, o fluxo de nutrientes total para uma célula alongada é o mesmo
do que para uma cadeia de células menores. Entretanto, o volume e a demanda efetiva por
nutrientes para células maiores excede o volume e a demanda por nutrientes para células em
cadeia. Portanto, é possível que a formação de uma cadeia de células menores forneça uma
vantagem competitiva em relação às células alongadas maiores (KENITZ et al., 2020).
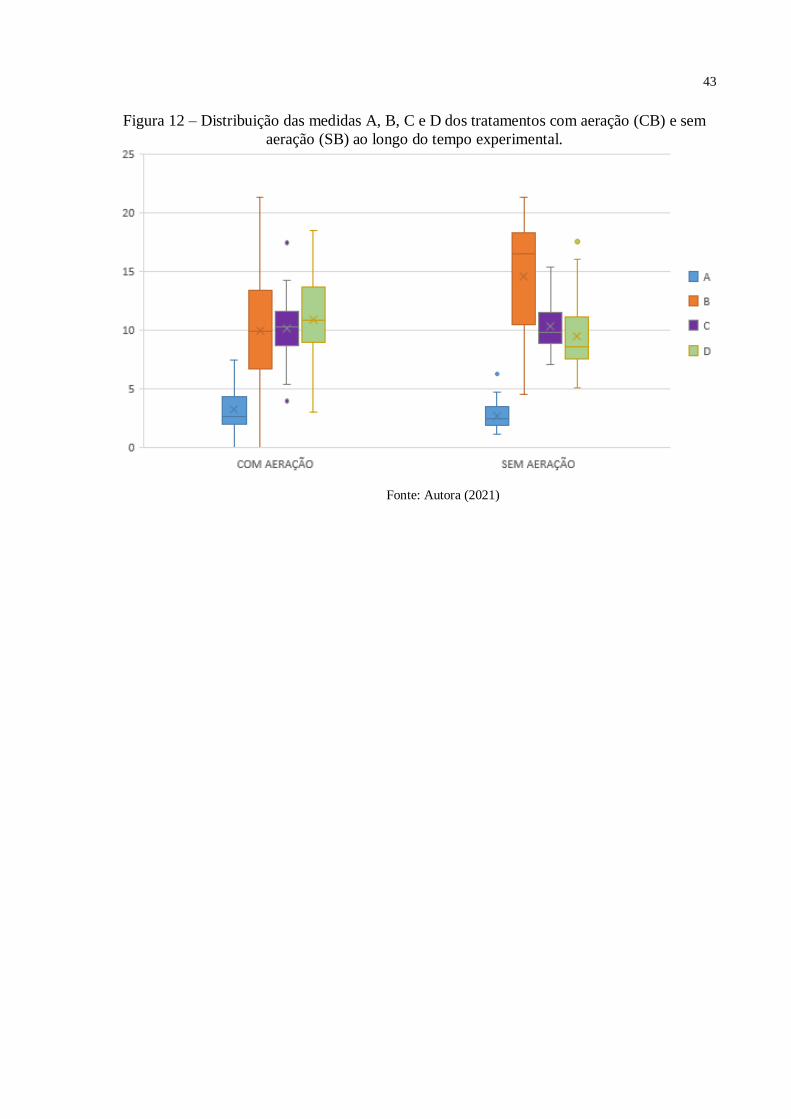
43
Figura 12 – Distribuição das medidas A, B, C e D dos tratamentos com aeração (CB) e sem
aeração (SB) ao longo do tempo experimental.
Fonte: Autora (2021)

44
6 CONCLUSÃO
Tendo em vista todos os dados recolhidos por este trabalho, e considerando a literatura
cientifica disponível sobre Asterionellopsis glacialis até o momento, o presente experimento
mostrou que os espinhos são as partes mais suscetíveis a deformidades desta diatomácea,
principalmente quando em cultivos laboratoriais. Pois, quando observa-se o desenvolvimento
da microalga ao longo de um período, é onde aparecem as formas teratológicas. Logo, é a
estrutura mais sensível a sentir as mudanças, quaisquer que sejam, que acontecem no ambiente.
Pôde ser vista a variação da morfologia das células ao longo do experimento. Logo,
fatores abióticos que possuem a capacidade de influenciar cultivos, como: nutrientes,
turbulência, pH, irradiância e temperatura, e até mesmo a presença de bactérias, podem estressar
as células, e, influenciar e induzir a forma como elas desenvolvem suas frústulas nos cultivos.
Entretanto, no experimento realizado não ficou claro que a aeração foi o único fator
associado às formas teratológicas, já que, analisando os resultados, infere-se que não houve
diferença nos tratamentos com e sem aeração. Portanto, outras variáveis que possuem a
capacidade de influenciar cultivos podem estar associadas com o processo de deformação das
frústulas dessas diatomáceas.
Análises multivariadas, que ajudam a revelar padrões de comportamento e
desenvolvimentos dos organismos, e, experimentos com observação em maiores frequências
podem ajudar a elucidar melhor a possível influência da aeração nos cultivos experimentais e
no crescimento e desenvolvimento dos organismos ao longo do tempo. Assim, faz-se necessário
outros tipos de análises para que fique mais claro qual é o fator, ou, quais são os fatores,
causador das formas teratológicas vistas em cultivos de laboratório.

45
APÊNDICE
MEIO F/2 DE CULTIVO PARA MICROALGAS

46
REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, Paulo C. et al. Decoupling between bacteria and the surf-zone diatom
Asterionellopsis glacialis at Cassino Beach, Brazil. Aquatic Microbial Ecology, [S.I.], v. 32,
n. 3, p. 219-228, 2003.
ALLDREDGE, Alice L.; GOTSCHALK, Chris; PASSOW, Uta; RIEBESELL, Ulf. Mass
aggregation of diatom blooms: insights from a mesocosm study. Deep Sea Research Part II:
Topical Studies in Oceanography, [S.L.], v. 42, n. 1, p. 9-27, jan. 1995. Elsevier BV.
ALLDREDGE, A. L., Passow, U., Logan, B. E. (1993) The abundance and significance of a
class of large, transparente organic particles in the ocean Deep Sea Res. 40: 1131-1140.
ANDERSEN, Robert A. Algal culturing techniques. Phycological Society of America.
Cambridge: Elsevier Academic Press, 2005.
ANTELO, F. S.; ANSCHAU, A.; COSTA, J. A. V., KALIL, S. J.. Extraction and purification
of C-phycocyanin from Spirulina platensis in conventional and integrated aqueous two-phase
systems. Journal of The Brazilian Chemical Society, São Paulo, v. 21, n. 5, p. 921-926.
2010.
ARRIETA, J. et al. On the fate of sinking diatoms: the transport of active buoyancy-
regulating cells in the ocean. Philosophical Transactions Of The Royal Society A:
Mathematical, Physical and Engineering Sciences, [S.L.], v. 378, n. 2179, p. 20190529, 3 ago.
2020. The Royal Society. http://dx.doi.org/10.1098/rsta.2019.0529.
AZAM, F, Smith DC, Steward GF, Hagstrom A. (1994). Bacteria—organic-matter coupling
and its significance for oceanic carbon cycling. Microbial Ecology 28: 167–179.
BENTLEY, K.; CLACK, C.; COX, E. J.. Diatom colony formation: a computational study
predicts a single mechanism can produce both linkage and separation valves due to an
environmental switch. Journal Of Phycology. v. 48, p. 716-728. 2012.
BONINI, M. A.. CULTIVO HETEROTRÓFICO DE Aphanothece microscopica Nägeli e
Chlorella vulgaris EM DIFERENTES FONTES DE CARBONO EM VINHAÇA. 2012.
96 f. Dissertação (Mestrado) - Curso de Mestrado em Agricultura e Ambiente, Centro de
Ciências Agrárias, Universidade Federal de São Carlos, Araras, 2012.
BROWN, A. C.; MCLACHLAN, A.. Ecology of Sandy Shores. Amsterdam: Elsevier, 1990.
328 p.
CAMPBELL, Eillen E.; BATE, G.C.. Coastal features associated with diatom discoloration of
surf-zones. Botanica Marina. v. 40, p. 179-185, 1997.
CASTRACANE, F.. Report on the Scientific Results of the Voyage of H. M. S.
Challenger during the years 1873-1876: Report on the Diatomaceae collected by H. M. S.
Challenger during the years 1873-1876. 1886. Disponível em:
https://www.biodiversitylibrary.org/page/818394#page/66/mode/1up

47
http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Bot-04/htm/doc.html Acesso em:
28/10/2020.
CLEVE, P. T. & MÖLLER, J. D. (1882). Diatoms, Part VI, No. 277-324. Upsala: Esatas
Edquists Boktryckeri.
CORZO, A.,Morillo, J.A. & Rodri!guez, S. (2000). Production of transparent exopolymer
particles (TEP) in cultures of Chaetoceros calcitrans under nitrogen limitation. Aquat.
Microb. Ecol., 23: 63±72.
COX, E. J.. Diatom identification in the face of changing species concepts and evidence of
phenotypic plasticity. Journal of Micropaleontology. v. 33, p. 111-120. 2014.
CROCKER, K. M.; PASSOW, U.. Differential aggregation of diatoms. Marine Ecology
Progress Series, [S.L.], v. 117, p. 249-257, 1995. Inter-Research Science Center.
http://dx.doi.org/10.3354/meps117249.
CUPP, E. E.. Marine Plankton Diatoms of the West Coast of North America. Berkeley,
California: Cambridge University Press, 1943. 238 p.
DECHO, A.W. (1990). Microbial exopolymer secretions in ocean environments: their role(s)
in food webs and marine processes. Oceanogr. Mar. Biol. Annu. Rev., 28: 78±153.
DELL’AQUILA, Gianluca; FERRANTE, Maria I.; GHERARDI, Marco; LAGOMARSINO,
Marco Cosentino, D’ALCALÀ, Maurizio Ribera; IUDICONE, Daniele; AMATO, Alberto.
Nutrient consumption and chain tuning in diatoms exposed to storm-like turbulence.
Scientific Reports, [S.L.], v. 7, n. 1, 12 maio 2017. Springer Science and Business Media
LLC. http://dx.doi.org/10.1038/s41598-017-02084-6.
DONEY, Scott C.. The ocean’s productive deserts. Nature, [S.L.], v. 389, n. 6654, p. 905-
906, out. 1997. Springer Science and Business Media LLC. http://dx.doi.org/10.1038/39993.
DRAPEAU, D.T., Dam, H.G. & Grenier, G. (1994). An improved-occulator design for use in
particle aggregation experiments. Limnol. Oceanogr., 39: 723±729.
FALASCO, E., BONA, F., BADINO, G., HOFFMANN, L., ECTOR, L., 2009. Diatom
teratological forms and environmental alterations: a review. Hydrobiologia 623, `-35.
FALASCO, Elisa; ECTOR, Luc; WETZEL, Carlos E.; BADINO, Guido; BONA, Francesca.
Looking back, looking forward: a review of the new literature on diatom teratological forms
(2010-2020). Hydrobiologia, [S.L.], v. 848, n. 8, p. 1675-1753, 4 mar. 2021. Springer
Science and Business Media LLC. http://dx.doi.org/10.1007/s10750-021-04540-x.
FERNÁNDEZ, M. R., G. Martín, J. Corzo, A. de la Linde, E. García, M. López & M. Souza,
2018. Design and testing of a new diatoms-based index for heavy metal pollution. Archives of
Environmental Contamination and Toxicology 74(1): 170-192.
FISHER, Nicholas S. et al. Effects of copper and zinc on growth, morphology, and
metabolism of Asterionella japonica (Cleve) 1. Journal Of Experimental Marine Biology
And Ecology, [S.L.], v. 51, n. 1, p. 37-56, abr. 1981. Elsevier BV.
http://dx.doi.org/10.1016/0022-0981(81)90153-2.

48
FRANCO, A. de O. da R., THEY, N. H., CANANI, L. G. de C., MAGGIONI, R., &
ODEBRECHT, C.. Asterionellopsis tropicalis (Bacillariophyceae): a new tropical species
found in diatom accumulations. Journal of Phycology, v. 52, n. 5, p. 888-895. 2016.
FRANCO, Andréa de Oliveira da Rocha; SOARES, Marcelo de Oliveira; MOREIRA, Maria
Odete Parente. Diatom accumulations on a tropical meso-tidal beach: environmental drivers
on phytoplankton biomass. Estuarine, Coastal And Shelf Science, [S.L.], v. 207, p. 414-421,
jul. 2018. Elsevier BV. http://dx.doi.org/10.1016/j.ecss.2017.07.020.
GÄRDES, Astrid; IVERSEN, Morten H; GROSSART, Hans-Peter; PASSOW, Uta;
ULLRICH, Matthias s. Diatom-associated bacteria are required for aggregation of
Thalassiosira weissflogii. The Isme Journal, [S.L.], v. 5, n. 3, p. 436-445, 9 set. 2010.
Springer Science and Business Media LLC. http://dx.doi.org/10.1038/ismej.2010.145.
GRESSLER, Pablo Diego. DESENVOLVIMENTO DE SISTEMA PARA CULTIVO DE
MICROALGAS E ENSAIO ECOFISIOLÓGICO PARA APLICAÇÃO EM
BIOTECNOLOGIA AMBIENTAL. 2016. 155 f. Tese (Doutorado) - Curso de Programa de
Biotecnologia e Biociências, Universidade Federal de Santa Catarina, Florianópolis, 2016.
GRANETTI, B., 1968. Alcune forme teratologiche compares in colture di Navicula mínima
Grun. e Navicula seminulum Grun. Giorn. Bot. Ital. 102, 469-484.
GROSSART, HP, Kiorboe T, Tang KW, Allgaier M, Yam EM, Ploug H. (2006). Interactions
between marine snow and heterotrophic bacteria: aggregate formation and microbial
dynamics. Aquatic Microbial Ecology 42: 19–26.
GUILLARD, Richard R. L.. Culture of Marine Invertebrates Animals: Culture of
Phytoplankton for Feeding Marine Invertebrates. In: SMITH, W. L.; CHANLEY, M. H.
(Eds.). Plenum Press, New York, USA. p. 29-60, 1975.
HOAGLAND, K.D., Rosowski, J.R., Gretz, M.R. & Roemer, S.C. (1993). Diatom
extracellular polymeric substances: function, ®ne structure, chemistry and physiology. J.
Phycol., 29: 537±566.
HONORATO, Milena Camargo. CARACTERÍSTICAS ECOFISIOLÓGICAS E
PROSPECÇÃO DE ATIVIDADES BIOLÓGICAS E BIOMOLÉCULAS NA
DIATOMÁCEA Asterionellopsis glacialis (CASTRACANE) ROUND. 2014. 105 f.
Dissertação (Mestrado) - Curso de Programa de Mestrado em Biologia de Fungos, Algas e
Plantas, Universidade Federal de Santa Catarina, Florianópolis, 2014.
HUMBORG, Christoph; SMEDBERG, Erik; MEDINA, Miguel Rodriguez; MÖRTH, Carl
Magnus. Changes in dissolved silicate loads to the Baltic Sea – The effects of lakes and
reservoirs. Journal of Marine Systems, [S.L.], v. 73, n. 3-4, p. 223-235, out. 2008. Elsevier
BV. http://dx.doi.org/10.1016/j.jmarsys.2007.10.014
HUSTEDT, F. 1956. Kieselalgen (Diatomeen). Einführung in Die Kleinlebwelt. Kosmos
Verlag, Stuttgart (70 p.).
JONES, J. I., J. F. Murphy, A. L. Collins, K. L. Spencer, P. S. Rainbow, A. Arnold, J. L.
Pretty, A. M. L. Moorhouse, V. Aguilera, P. Edwards, F. Parsonage, H. Potter & P.

49
Whitehouse, 2018. The impact of metal-rich sediments derived from mining on freshwater
stream life. In: de Voogt, P. (ed) Reviews of Environmental Contamination and Toxicology,
vol 248. Springer, Champ, pp 111-189.
JOUENNE, F., LEFEBVRE, S., VÉRON, B. and LAGADEUC, Y.. Phytoplankton
community structure and primary production in small intertidal estuarine-bay ecosystem
(eastern English Channel, France). Marine Biology, v. 151, p. 805-825. 2007.
KACZMARSKA, I.; MATHER, L.; LUDDINGTON, I. A.; MUISE, F.; EHRMAN, J. M..
Cryptic diversity in a cosmopolitan diatom known as Asterionellopsis glacialis
(Fragilariaceae): Implications for ecology, biogeography, and taxonomy. American Journal
Of Botany. v. 101, p. 267-286. 2014.
KAHL, LA, Vardi A, Schofield O. (2008). Effects of phytoplankton physiology on export
flux. Marine Ecology-Progress Series 354: 3–19.
KARP-BOSS, L. et al. NUTRIENT FLUXES TO PLANKTONIC OSMOTROPHS IN THE
PRESENCE OF FLUID MOTION. Oceanography And Marine Biology: An Annual
Review, [S.].], v. 34, p. 71-107, 1996.
KENITZ, Kasia M.; ORENSTEIN, Eric C.; ROBERTS, Paul L. D.; FRANKS, Peter J. S.;
JAFFE, Jules S.; CARTER, Melissa L.; BARTON, Andrew D.. Environmental drivers of
population variability in colony-forming marine diatoms. Limnology And Oceanography,
[S.L.], v. 65, n. 10, p. 2515-2528, 26 maio 2020. Wiley. http://dx.doi.org/10.1002/ lno.11468.
KOGURE, K, Simidu U, Taga N. (1981). Bacterial attachment to phytoplankton in sea-water.
Journal of Experimental Marine Biology and Ecology 56: 197–204.
KOOISTRA, W. H. C. F.; GERSONDE, R.; MEDLIN, L. K, MANN, D. G.. The origin and
evolution of the diatoms: their adaptation to a planktonic existence. In: Evolution of
primary producers in the sea. (Eds) FALKOWSKI, Paul G.; KNOLL, Andrew H. 2007.
LAROCQUE, Isabelle et al. Sedimentation of algae:: relationships with biomass and size
distribution. Canadian Journal Of Fisheries And Aquatic Sciences, [S.I.], v. 53, n. 5, p.
1133-1142, maio 1996.
LA ROCHA, Christina L. de et al. Factors influencing the sinking of POC and the efficiency
of the biological carbon pump. Deep Sea Research Part II: Tropical Studies in
Oceanography, [S.L.], v. 54, p. 639-658, mar. 2007. Elsevier BV.
http://dx.doi.org/10.1016/j.dsr2.2007.01.004.
LAVOIE, Isabelle; HAMILTON, Paul B.; MORIN, Soizic; TIAM, Sandra Kim; KAHLERT,
Maria; GONÇALVES, Sara; FALASCO, Elisa; FORTIN, Claude; GONTERO, Brigitte;
HEUDRE, David. Diatom teratologies as biomarkers of contamination: are all deformities
ecologically meaningful? Ecological Indicators, [S.L.], v. 82, p. 539-550, nov. 2017.
Elsevier BV. http://dx.doi.org/10.1016/j.ecolind.2017.06.048.
LELIAERT, F.; VERBRUGGEN, H.; VANORMELINGEN, P.; STEEN, F.; LOPEZ-
BAUTISTA, J. M.; ZUCCARELLO, G. C.; CLERCK, O.. DNA-based species delimitation in
algae. European Journal Of Phycology. v. 49, p. 179-196. 2014.

50
LEWIS, J, Kennaway G, Franca S, Alverca E. (2001). Bacterium-dinoflagellate interactions:
investigative microscopy of Alexandrium spp. (Gonyaulacales, Dinophyceae). Phycologia 40:
280–285.
LOGAN, B.E. & Alldredge, A.L. (1989). Potential for increased nutrient uptake by
flocculating diatoms. Mar. Biol., 101: 443±450.
LOGAN, B.E., Passow, U., Alldredge, A.L., Grossart, H.P. & Simon, M. (1995). Rapid
formation and sedimentation of large aggregates is predictable from coagulation rates (half-
lives) of transparent exopolymer particles (TEP). Deep-Sea Res. II, 42: 203±238.
LOGAN, B. E., Passow, U., Alldredge A. L. (1994). Variable retention of diatoms on screens
during size separations Limnol. Oceanogr. 39: 390-395
LOURENÇO, S. O.. Cultivo de microalgas marinhas – princípios e aplicações. São
Carlos: RIMA. 2006.
MARGALEF, R. Turbulence and marine life. Scientia Marina, v. 61, p. 109-123. 1997.
MASMOUDI, S., N. Nguyen-Deroche, A. Caruso, H. Avadi, A. Morant-Manceau, G.
Tremblin, M. Bertrand & B. Schoefs, 2013. Cadmium, copper, sodium and zinc effects on
diatom: from heaven to hell – a review. Cryptogamie, Algologie 34(2): 185 – 225.
MCLACHLAN, A.; DEFEO, O.. 2017. The Ecology of Sandy Shores: Beach and Surf-Zone
Flora. 3. ed. Academic Press, 2017. 572 p.
MIQUEL, P. (1890-1892). De la culture artificielle des diatomées. Le Diatomiste 1 73-75, 93-
99, 121-128, 149-156, 165-172.
MORIOKA, Luiz Rodrigo Ito et al. Floculação de Chlorella sp. produzida em concentrado de
dessalinização e estudo de método de extração de lipídeos intracelulares. Química Nova,
[S.L.], v. 37, n. 1, p. 44-49, 2014. FapUNIFESP (SciELO). http://dx.doi.org/10.1590/s0100-
40422014000100009.
MOUSING, Erik Askov et al. Global patterns in phytoplankton biomass and community size
structure in relation to macronutrients in the open ocean. Limnology And Oceanography,
[S.L.], v. 63, n. 3, p. 1298-1312, 9 jan. 2018. Wiley. http://dx.doi.org/10.1002/ lno.10772.
MYKLESTAD, SM. (1995). Release of extracellular products by phytoplankton with special
emphasis on polysaccharides. Science of the Total Environment 165: 155–164.
NURDOGAN, Yakup; OSWALD, William J.. Enhanced nutrient removal in high-rate
ponds. Water Science And Technology, [S.L.], v. 31, n. 12, p. 33-43, 1 jun. 1995. IWA
Publishing. http://dx.doi.org/10.2166/wst.1995.0453.
ODEBRECHT, C., SEGATTO, A. Z., & FREITAS, C. A.. Surf-zone Chlorophyll a
Variability at Cassino Beach, Southern Brazil. Estuarine, Coastal and Shelf Science, v. 41,
n. 1, p. 81–90. 1995.

51
ODEBRECHT, C.; BERGESCH, M.; RÖRIG, L. R.; ABREU, P. C.. Phytoplankton
Interannual Variability at Cassino Beach, Southern Brazil (1992-2007), with Emphasis on the
Surf Zone Diatom Asterionellopsis glacialis. Estuaries and coasts. v. 33, p. 570-583. 2010.
PAHLOW, Markus et al. Impact of cell shape and chain formation on nutrient acquisition by
marine diatoms. Limnology And Oceanography, [S.L.], v. 42, n. 8, p. 1660-1672, dez. 1997.
Wiley. http://dx.doi.org/10.4319/ lo.1997.42.8.1660.
PASSOW, U., Alldredge, A.L. & Logan, B.E. (1994). The role of particulate carbohydrate
exudates in the ¯flocculation of diatoms blooms. Deep-Sea Res. I, 41: 335±357.
PASSOW, U; SHIPE, R.F; A MURRAY,; PAK, D.K; BRZEZINSKI, M.A; ALLDREDGE,
A.L. The origin of transparent exopolymer particles (TEP) and their role in the sedimentation
of particulate matter. Continental Shelf Research, [S.L.], v. 21, n. 4, p. 327-346, mar. 2001.
Elsevier BV. http://dx.doi.org/10.1016/s0278-4343(00)00101-1.
RAVEN, J. A. Limits to growth. In: BOROWITZKA, M. A.; BOROWITZKA, L. J..
Microalgal Biotechnology, Cambridge: Cambridge University, 1988. p. 331-356.
RAVEN, H. P.; EVERT, F. R.; EICHHRON, E. S.. Biologia vegetal. 5. ed., 1996. 728 p.
RENAUD, S. M.; THINH, L..; LAMBRINIDIS, G..; PARRY, D. L.. Effect of temperature on
growth, chemical composition and fatty acid composition of tropical Australian microalgae
grown in batch cultures. Aquaculture. v. 211, p. 195–214. 2002.
RELATÓRIO DE BALNEABILIDADE DO INSTITUTO DO MEIO AMBIENTE DE
SANTA CATARINA. Disponível em:
https://balneabilidade.ima.sc.gov.br/relatorio/relatorioBalneabildade. Acesso em: 30/03/2021.
RICHMOND, A. (ed.). Handbook of microalgal culture: biotechnology and applied
phycology. Oxford: Blackwell Science, 2004. 566 p.
RÖRIG, L. R.; HONORATO, M. C.; ITOKAZU, A. G.; DE MACEDO, C. X.;
DESCHAMPS, F.; LINS, J. V. R.; MARASCHIN, M.; RAMLOV, F.; GRESSLER, P. D.;
PEREIRA FILHO, J.. Ecophysiological and biochemical variation of the surf zone diatoms
Asterionellopsis glacialis sensu lato from Santa Catarina, Southern Brazil. Brazilian Journal
of Oceanography, v. 65, n. 4, p. 695-708. 2017.
RÖRIG, L. R.; GARCIA, V. M. T.. Accumulations of the surf-zone diatom Asterionellopsis
glacialis (CASTRACANE) ROUND in Cassino Beach, Southern Brazil, and its relationship
with environmental factors. Journal of Coastal Research. v. 35, p. 167–177. 2003.
RÖRIG, L. R.; DE ALMEIDA, T. C. M.; GARCIA, V. M. T.. Structure and succession of the
surf-zone phytoplankton in Cassino beach, Southern Brazil. Journal of coastal research. p.
1246- 1250. 2006.
ROUND, F. E., CRAWFORD, R. M., & MANN, D. G.. Diatoms: biology and morphology
of the genera. Cambridge University Press, 2007.

52
SEELIGER, Ulrich. Areias do Albardão: um guia ecológico ilustrado do litoral no
extremo sul do Brasil. (Eds) SEELIGER, Ulrich; CORDAZZO, César; BARCELOS, Lauro.
Rio Grande: Ecoscientia. 2004. 96 p.
SCHMID, A. M. M., 1980. Valve morphogenesis in diatoms: a pattern-related filamentous
system in pennates and the effect of APM, colchicine and osmotic pressure. Nova Hedwigia
33, 811-847.
SCHOLZ, Matt et al. Flocculation of wall-deficient cells of Chlamydomonas reinhardtii
mutant cw15 by calcium and methanol. Biomass And Bioenergy, [S.L.], v. 35, n. 12, p.
4835-4840, dez. 2011. Elsevier BV. http://dx.doi.org/10.1016/j.biombioe.2011.08.020.
SHUKLA, S. K.; CROSTA, X.; CORTESE, G.; NAYAK, G. N.. Climate controlled size
variability of diatom Fragilariopsis kerguelensis in the Southern Ocean. Quaternary Science
Reviews. v. 69, p. 49-68. 2013.
SILVA, Pryscilla Denise Almeida da. Biodiversidade e distribuição das diatomáceas
planctônicas e de sedimento superficial nas represas de abastecimento do Sistema Alto
Cotia (SP). 2012. 98 f. Dissertação (Mestrado) - Curso de Biodiversidade Vegetal e Meio
Ambiente, Instituto de Botânica da Secretaria do Meio Ambiente do Estado de São Paulo, São
Paulo, 2012.
SILVA, C. F. M.. Impactos das mudanças climáticas sobre diatomáceas perifíticas: como
o aumento da temperatura e de eventos de “brownification” afetam a comunidade? 2018. Tese (Doutorado). Universidade Federal do Rio Grande, 2018.
SMETACEK, Victor. Diatoms and the silicate factor. Nature, [S.L.], v. 391, n. 6664, p. 224-
225, jan. 1998. Springer Science and Business Media LLC. http://dx.doi.org/10.1038/34528.
SMITH, Benjamin T.; DAVIS, Robert H.. Sedimentation of algae flocculated using naturally-
available, magnesium-based flocculants. Algal Research, [S.L.], v. 1, n. 1, p. 32-39, maio
2012. Elsevier BV. http://dx.doi.org/10.1016/j.algal.2011.12.002.
SPILLING, Kristian; OLLI, Kalle; LEHTORANTA, Jouni; KREMP, Anke; TEDESCO,
Letizia; TAMELANDER, Tobias; KLAIS, Riina; PELTONEN, Heikki; TAMMINEN, Timo.
Shifting Diatom – Dinoflagellate Dominance During Spring Bloom in the Baltic Sea and its
Potential Effects on Biogeochemical Cycling. Frontiers In Marine Science, [S.L.], v. 5, 19
set 2018. Frontiers Media SA. http://dx.doi.org/10.3389/fmars.2018.00327.
SU, Yanyan; LUNDHOLM, Nina; ELLEGAARD, Marianne. Effects of abiotic factors on the
nanostructure of diatom frustules – ranges and variability. Applied Microbiology And
Biotechnology, ]S.L.], v. 102, n. 14, p. 5889-5899, 26 maio 2018. Springer Science and
Business Media LLC. http://dx.doi.org/10.1007/s00253-018-9087-1.
THORNTON, Daniel C. O.. Diatom aggregation in the sea: mechanisms and ecological
implications. European Journal Of Phycology, [S.L.], v. 37, n. 2, p. 149-161, maio 2002.
Informa UK Limited. http://dx.doi.org/10.1017/s0967026202003657.
TOMAS, C. R. (ed.). Identifying marine phytoplankton. Elsevier, 1997.

53
VANDAMME, Dries et al. Flocculation of Chlorella vulgaris induced by high pH: role of
magnesium and calcium and practical implications. Bioresource Technology, [S.L.], v. 105,
p. 114-119, fev. 2012. Elsevier BV.
VIANA, João Cláudio Cerqueira. DIATOMÁCEAS (BACILLARIOPHYCEAE)
EPILÍTICAS COMO BIOMONITORES DA QUALIDADE DE ÁGUA DOS RIOS
CUMBUCA, MUCUGÊ E PIABINHA (CHAPADA DIAMANTINA-BA). 2007. 148 f.
Dissertação (Mestrado) - Curso de Ecologia e Biomonitoramento, Universidade Federal da
Bahia, Salvador, 2007.
VIANNA, S. C.; SANQUETTA, C. R.. Mudanças climáticas e o fitoplâncton marinho: uma
revisão. Enciclopédia Biosfera, v.18, n.15, p. 1883-1901. 2012.
VIEIRA, C. E. L.. Diatomáceas. In: CARVALHO, Ismar de Souza (ed.). Paleontologia –
Volume 2: Microfósseis e Paleoinvertebrados. 3. ed. Interciência, 2011. Cap. 6., p. 109-
138.
WENGRAT, Simone. Biodiversidade e distribuição das diatomáceas no Complexo
Billings, São Paulo: influência da compartimentalização espacial e do estado trófico.
2011. 113 f. Dissertação (Mestrado) - Curso de Biodiversidade Vegetal e Meio Ambiente,
Instituto de Botânica da Secretaria do Meio Ambiente do Estado de São Paulo, São Paulo,
2011.
WERNER, Dietrich. Die Kieselsãure in stoffchsel von Cyclutella cryptica Reimann, Lewin
und Guillard. Arch Mikrobiol. v. 55, p. 278-308. 1966.
YAHI, H. et al. Algal flocculation-sedimentation by pH increase in a continuous
reactor. Water Science And Technology, [S.L.], v. 30, n. 8, p. 259-267, 1 out. 1994. IWA
Publishing. http://dx.doi.org/10.2166/wst.1994.0421.





























