Embed Size (px)
Citation preview

Rom J Morphol Embryol 2016, 57(2 Suppl):681–689
ISSN (print) 1220–0522 ISSN (online) 2066–8279
OORRIIGGIINNAALL PPAAPPEERR
Ultrastructure and histomorphometric analysis of human umbilical cord vessels in preeclampsia: a potential role of VEGF, VEGFR-1 and VEGFR-2
SHAIMA M. ALMASRY1,2), AMR K. ELFAYOMY3,4), HALA E. HASHEM5)
1)Department of Anatomy, Taibah University, Al-Madinah Al-Munawarah, Kingdom of Saudi Arabia 2)Department of Anatomy and Embryology, Almansoura University, Almansoura, Egypt 3)Department of Obstetrics and Gynecology, Taibah University, Al-Madinah Al-Munawarah, Kingdom of Saudi Arabia
4)Department of Obstetrics and Gynecology, Zagazig University, Zagazig, Egypt 5)Department of Histology and Cell Biology, Zagazig University, Zagazig, Egypt
Abstract Aim: This study aimed to analyze the ultrastructure and histomorphometric changes of the human umbilical cord vessels of preeclampsia compared to healthy pregnancies and the possible role of vascular endothelial growth factor (VEGF) and its receptors. Materials and Methods: Umbilical cord (UC) specimens were collected between August, 2014 and July, 2015. Histomorphometric analysis of UC vessels was performed utilizing an image analysis system. Cellular localization of VEGF, VEGFR-1 (VEGF receptor-1) and VEGFR-2 (VEGF receptor-2) were examined in immunohistochemically-stained sections of UC from 45 pregnancies with preeclampsia (PE) and 40 healthy pregnancies. Results: Compared with healthy pregnancies, UC venous measurements were significantly higher in PE; total venous area (p<0.001), luminal venous area (p<0.001) and luminal venous index (p=0.005). Arterial measurements except for the total arterial area were significantly lower in the PE compared to healthy pregnancies, luminal arterial area (p=0.32) and luminal arterial index (p=0.004). Histological and ultrastructural examination of UC from PE revealed discontinuity of vascular endothelium and disorganized edematous widely spaced smooth muscle cells. We demonstrated a significant increase in tissue expression of VEGF in PE (16.6±0.1) compared to healthy pregnancies (12±0.8) (p=0.001). Also, significant higher VEGFR-1 expression in PE (20.5±2.5) compared to healthy pregnancies (9.5±1.2) (p<0.001) has been observed. However, tissue expression of VEGFR-2 was decreased significantly in PE (10.5±0.7) compared to healthy pregnancies (13.8±1.6) (p=0.043). Conclusions: Altered tissue expression of VEGF and its receptors in the UC vessels could play a crucial role in disturbing the UC vascular endothelium, smooth muscles and measurements and this may underlie the existence of preeclampsia.
Keywords: VEGF, VEGFR-1, VEGFR-2, umbilical cord vessels.
Introduction
The umbilical cord (UC) is an extension of the fetal weave and consequently it allows assessing the influence of clinical diseases and/or intrinsic fetal sickness affecting pregnancy. Human umbilical vessels vary from the major vessels of the same caliber in the body for many causes. Transudation of fluid occurs in these vessels and contri-butes to the formation of the amniotic fluid [1]. Umbilical vessels are not supplied by vasa vasorum and thus depend on their own oxygen supply, making them more susceptible to alterations in hemodynamic condition [2, 3]. In spite of complete and accurate descriptions of the histology of UC vessels, the histomorphometry of these structures have been inadequately studied, despite the wide broad spectrum of pathologies affecting the mother and the fetus.
Preeclampsia (PE), a syndrome characterized by the presence of hypertension and proteinuria and is believed to be due to endothelial cell dysfunction [4]. Endothelial dysfunction cause vascular manifestations like hyper-tension, proteinuria, edema, hepatic hemorrhage, renal morbidity and others. Still, the causes lie behind endothelial dysfunction in PE remains uncertain. Some investigators proposed placental factors to have a key role, but the exact nature of these constituents is however not clear [5].
Vascular endothelial growth factor (VEGF) is a dimeric glycoprotein with potent angiogenic properties. The role of VEGF is to stimulate endothelial cell proliferation, migration, and survival [6]. VEGF employs its biologic effect through two high-affinity receptor tyrosine kinases: VEGFR-1 and VEGFR-2 [7]. Previous reports suggest that the circulating levels of VEGF are elevated significantly in PE [8]. In addition, a positive correlation between VEGF concentrations and systemic or uterine vascular resistance was described [9].
To date, little is known about the expression of VEGF and VEGF receptors in the tissues and the vessels of the umbilical cord from healthy and PE pregnancies and its relation to the measurements of the UC vessels. Thus, ultrastructural, histomorphometry and immunohisto-chemical analysis was conducted in this study to explore the immunoexpression of VEGF/VEGFR-1/VEGFR-2 in the tissues and the vessels of the UC from healthy and PE pregnancies.
Materials and Methods
This observational study included 85 consecutive women with singleton pregnancies that delivered between 29 to 40 weeks gestation, 40 cases were healthy pregnancies
R J M ERomanian Journal of
Morphology & Embryologyhttp://www.rjme.ro/

Shaima M. Almasry et al.
682
and 45 were suffering from PE. Preeclampsia was defined as elevated blood pressure (≥140/90 mmHg on two measurements ≥6 hours apart or ≥160/105 mmHg on presentation) after 20 weeks gestation with the onset of proteinuria (≥0.3 g in 24 hours urine specimen, or ≥2+ on urinary dipstick) [2]. Severe PE was defined as severe hypertension (diastolic blood pressure ≥110 mmHg) and proteinuria (a 24-hour urine sample containing 5 g protein or urine specimen ≥3+ protein by dipstick measurement). Women with chronic hypertension, diabetes mellitus, renal and liver diseases or smokers were excluded. For all patients, arterial umbilical flow velocity waveforms (FVW) from 24 weeks until one week before delivery were recorded with Toshiba SSA 270A/HG pulse-waved Doppler, using abdominal probe 5 MHz. Normal umbilical Doppler indices were defined as a systolic/diastolic values between 5% and 95% with respect to the gestational age. Values out of this range were taken as abnormal [10]. Demographic data and routine clinical investigations were recorded.
Tissue sampling
Umbilical cord samples were collected after delivery from Al-Madinah Maternity Hospital (Taibah University-Teaching Hospital), Al-Madinah Al-Munawarah Province, Kingdom of Saudi Arabia, between August, 2014 and July, 2015. Each patient was given an informed consent to contribute in the study. The study protocol was approved by the Medical and Health Sciences Research Committee Involving Human Subjects of our Hospital (No. 413/ 20-8-2014), which conforms to the provisions of the Declaration of Helsinki.
Segments of UC 5 cm away from the placental end were obtained and fixed in buffered formaldehyde (pH 7.0), processed to wax using routine laboratory practices. Random choice of tissue blocks was done and the blocks were cut at 5 μm thicknesses and were processed for Hematoxylin–Eosin (HE) staining, carried out according to conventional procedures [11].
Morphometric analysis
Morphometric measures were taken from 85 blocks: 40 from UC of normal pregnancies and 45 from UC of preeclampsia. Entire sections including the arteries and the vein were digitalized and used for histomorphometry at 200× magnification. The same investigator performed all analyses blind. Morphological measurements were taken using an Image Analyzer (Leica Q Win standard, digital camera CH-9435 DFC 290, Germany). The following measurements were obtained: (a) venous measurements: total venous area, luminal venous area and luminal venous index = luminal area/total area; (b) arterial measurements: total arterial area, luminal arterial area and luminal arterial index = luminal area/total area.
Immunohistochemistry
Three μm thick sections were collected onto coated slides and examined using an immunoperoxidase according to the conventional methods. Immunohistochemical staining was done using primary rabbit monoclonal anti-human VEGF, primary rabbit polyclonal anti-human VEGFR-1 and anti-human VEGFR-2 (Table 1).
Table 1 – Specifications of the primary antibodies
Antigen Source Catalog Clonality Code Dilution
VEGF Spring
BioscienceM3281 Monoclonal Sp 28
Ready to use
VEGFR-1Spring
BioscienceE2801 Polyclonal –
Ready to use
VEGFR-2Spring
BioscienceE3711 Polyclonal –
Ready to use
To localize binding of the primary antibodies, the slides were incubated with the secondary anti-rabbit antibodies (IgG1) universal kits obtained from Zymed Corporation. For negative controls, the incubation was done by a non-specific mouse IgG1 antibody. Then, the sections were incubated in 3,3’-diaminobenzidine (DAB) reagent and counterstained with Hematoxylin and coverslipped using Protex ting media (DAB-Stock Stain box; Boster Bio-technology) [12]. All the immunostainings were performed under the same conditions, including temperature, incu-bation periods of primary and secondary antibodies, and exposure period to DAB, to avoid the potential influence on the staining intensity in the different groups.
Evaluation of immunostaining
Tissue sections were tested using an optical microscope at 4× and 10× magnification for the initial screening. A positive reaction for VEGF protein was detected as a brown-yellow granulation in the cellular membranes, cytoplasm and extracellular matrix while a positive reaction for VEGFR-1 and VEGFR-2 proteins was detected as a brown-yellow granulation in the cytoplasm. The area % of the VEGF, VEGFR-1 and VEGFR-2 immunopreci-pitates was measured utilizing the Image Analyzer that allowed avoiding subjectivity and selection bias in judgment of the degree of staining through adjustment of the image detection tools at a fixed point detecting a certain point of brown coloration in all examined slides.
TEM processing
Small fragments of umbilical cord tissues were fixed in 2% glutaraldehyde, in 0.085 M sodium cacodylate buffer (pH 7.4) for 24 hours, then washed in phosphate-buffered saline (PBS) and post-fixed in 1% osmium tetroxide for one hour, dehydrated and embedded in Epoxy resin. Ultrathin sections (50 nm) were cut, mounted on copper grids and stained with uranyl acetate after that, lead citrate, and then assessed by transmission electron microscope (JEOL 1200 EXII) in Research Laboratory, College of Science, Ain Shams University, Egypt [13].
Statistical analysis
All data were expressed as mean ± SEM (standard error of mean). Significant differences of average values between groups were counted on using independent Student’s t-test or Mann–Whitney U-test. The threshold for statistical significance was set at p<0.05. IBM SPSS (version 21) statistical software was used.
Results
Clinical data
Mean maternal age, maternal parity and infant sex were not significantly different between the two groups,
Ultrastructure and histomorphometric analysis of human umbilical cord vessels in preeclampsia: a potential role…
683
however, body mass index, mode of delivery and Apgar score was significantly higher in preeclampsia patient. On the other hand, gestational age and birth weight of newborn babies was significantly lower in the PE group as indicated in Table 2. Among preeclamptic patients, 35 (77.8%) were categorized as having severe PE, 10 (22.2%) patients experienced a previous history of PE and 27 (60%) had abnormal umbilical artery FVW.
Table 2 – Clinical characteristics of the study population
Parameters Normal (n=40)
Preeclampsia (n=45)
p-value
Maternal age [years] 31.2±1.22 30.9±0.95 0.448
Gestational age [weeks] 39 (37–40) 35 (29–40) <0.001*
Parity 3.1±0.22 2.6±0.19 0.164Body mass index (BMI)
[kg/m2] 27.3±0.34 28.7±0.25 0.001*
Birth weight [g] 3016.6±50.81 2304.2±69.82 <0.001*
Infant sex
Female n (%) 23 (57.5) 25 (55.5)
Male n (%) 17 (42.5) 20 (44.4) 0.322
Mode of delivery
Vaginal 27 (67.5) 22 (48.8)
Caesarean 13 (32.5) 23 (51.1) <0.001*
5 min. Apgar score 8.1±0.7 (8–10) 7.2±0.4 (6–9) 0.033*
Data are expressed as mean ± SEM (standard error of mean) or number and percentage. *Significant p-value <0.05.
Morphometric analysis of umbilical cord blood vessels
As shown in Table 3, compared to the control group, total venous area (p<0.001), luminal venous area (p<0.001) as well as the luminal venous index (p=0.005) was signi-ficantly higher in the preeclamptic specimens. Arterial measures show significant reduction in the luminal arterial area (p=0.032) and luminal arterial index (p=0.004) in the preeclamptic vessels compared to the controls, however, there no significant difference found in the total arterial area between both groups (p=0.681).
Table 3 – Vascular measurements of the umbilical cord
Measurements Normal (n=40)
Preeclampsia (n=45)
p-value
Venous measurements
Total area [μm2] 2 296 292±
143 794 31 841 801±
3 320 129 <0.001*
Lumen area [μm2] 243 276± 40 283
5 051 625± 401 758
<0.001*
Luminal index 0.1±0.02 0.2±0.02 0.005*
Arterial measurements
Total area [μm2] 1 362 700±
113 025 1 426 333±
104 430 0.681
Lumen area [μm2] 173 588± 72 341
26 859± 4403
0.032*
Luminal index 0.15±0.05 0.02±0.003 0.004* *Significant p-value <0.05 for t-test.
Histological analysis
Light microscopic examination of the human UC sections taken from control group revealed the presence of two arteries and one vein, which are embedded within Wharton’s jelly. The surface of the cord is covered by a single layer of amniotic epithelium. The Wharton’s jelly contained spindle-shaped cells embedded in matrix.
The umbilical arteries have a double-layered muscular wall entails inner longitudinal and outer circular smooth muscle fibers (Figure 1A). The vein has an inner elastic lamina and a single layer of circular smooth muscle. Compared to the arteries, the veins have a larger diameter and thinner muscular coat (Figure 1B). The umbilical vessels are lined by endothelium with flattened pale stained nuclei and resting on subendothelial connective tissue (Figure 1C).
Light microscopic examination of the human UC sections taken from preeclamptic group showed the endothelium of umbilical vessels with areas of discon-tinuation and contained pyknotic darkly stained nuclei. The smooth muscle layer of umbilical vessels showed some areas of thickening. Pale stained areas of separations between smooth muscle layers and in the subendothelial connective tissue of the umbilical vessels and within Wharton’s jelly were detected. Smooth muscle cell sub-endothelial infiltration was also noted (Figure 1, D–F).
Ultrastructural analysis
Electron micrograph of a cross-section of UC from healthy pregnant women showed longitudinal and circular smooth muscle cells with centrally located nucleus and cytoplasm occupied by thin filaments and dense bodies. In that location were no transverse striations (Figure 2, A and B).
Electron micrograph of a cross-section of UC from preeclamptic pregnant women showed the muscular layers of the umbilical vessels with cellular infiltrations, edematous disorganized smooth muscle cells, widened intercellular spaces with collagen fibril deposition (Figure 2, C–E).
Immunohistochemical analysis
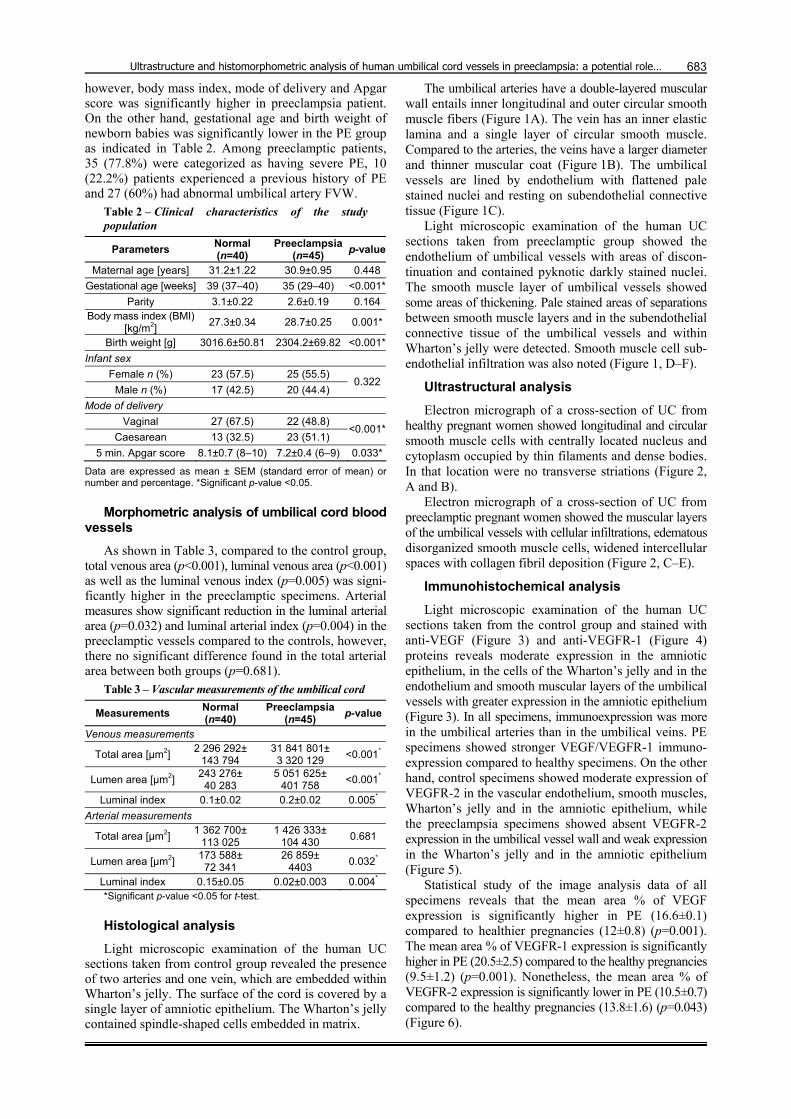
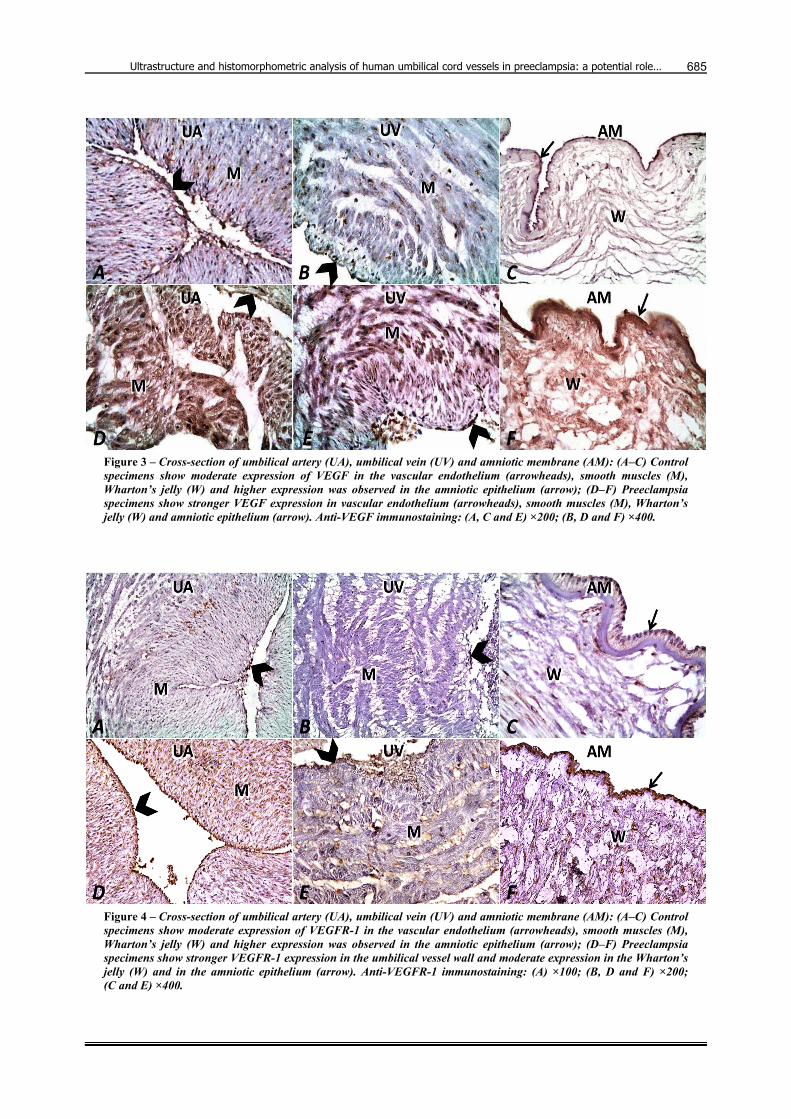
Light microscopic examination of the human UC sections taken from the control group and stained with anti-VEGF (Figure 3) and anti-VEGFR-1 (Figure 4) proteins reveals moderate expression in the amniotic epithelium, in the cells of the Wharton’s jelly and in the endothelium and smooth muscular layers of the umbilical vessels with greater expression in the amniotic epithelium (Figure 3). In all specimens, immunoexpression was more in the umbilical arteries than in the umbilical veins. PE specimens showed stronger VEGF/VEGFR-1 immuno-expression compared to healthy specimens. On the other hand, control specimens showed moderate expression of VEGFR-2 in the vascular endothelium, smooth muscles, Wharton’s jelly and in the amniotic epithelium, while the preeclampsia specimens showed absent VEGFR-2 expression in the umbilical vessel wall and weak expression in the Wharton’s jelly and in the amniotic epithelium (Figure 5).
Statistical study of the image analysis data of all specimens reveals that the mean area % of VEGF expression is significantly higher in PE (16.6±0.1) compared to healthier pregnancies (12±0.8) (p=0.001). The mean area % of VEGFR-1 expression is significantly higher in PE (20.5±2.5) compared to the healthy pregnancies (9.5±1.2) (p=0.001). Nonetheless, the mean area % of VEGFR-2 expression is significantly lower in PE (10.5±0.7) compared to the healthy pregnancies (13.8±1.6) (p=0.043) (Figure 6).

Shaima M. Almasry et al.
684
Figure 1 – Cross-section of the wall of human umbilical cord vessels. Normal pregnancy specimens show: (A) Umbilical artery (UA) inner longitudinal (IL) and outer circular (OC) muscle fibers surround a narrow lumen (L); (B) Umbilical vein (UV) shows a single layer of circular muscle (CM) and an internal elastic lamina (arrowhead); (C) The endothelium of the UA appears with flattened pale stained nuclei (arrowheads) and resting on subendothelial connective tissue (*). Preeclampsia specimens show: (D) Umbilical artery (UA) with thickened wall and area of smooth muscle fibers proliferation (arrow) containing wide spaced muscle cells; (E) Umbilical vein (UV) with thin wall, widened lumen and spaces (arrows) are seen within muscle layer and surrounding Wharton’s jelly; (F) Endothelium of the UV is seen with pyknotic darkly stained nuclei (arrowheads) and subendothelial smooth muscle cell infiltration (*). Multiple spaces (arrows) are seen within the muscle layer. HE staining: insets ×40; (A, B, D and E) ×100; (C and F) ×1000.
Figure 2 – Electron micrograph of cross-section of the wall of human umbilical cord vessels: (A and B) Normal pregnancy specimens show (A) longitudinal smooth muscle cells and (B) circular smooth muscle cells. They have centrally located nucleus (N) and dense cytoplasmic bodies (arrows); (C–E) Preeclampsia specimens show muscular layer with cellular infiltrations (I), edematous disorganized smooth muscle cells (M), widened intercellular spaces (S) with collagen fibrils deposition (arrows). (A–C) ×6000; (D and E) ×9000.
Ultrastructure and histomorphometric analysis of human umbilical cord vessels in preeclampsia: a potential role…
685
Figure 3 – Cross-section of umbilical artery (UA), umbilical vein (UV) and amniotic membrane (AM): (A–C) Control specimens show moderate expression of VEGF in the vascular endothelium (arrowheads), smooth muscles (M), Wharton’s jelly (W) and higher expression was observed in the amniotic epithelium (arrow); (D–F) Preeclampsia specimens show stronger VEGF expression in vascular endothelium (arrowheads), smooth muscles (M), Wharton’s jelly (W) and amniotic epithelium (arrow). Anti-VEGF immunostaining: (A, C and E) ×200; (B, D and F) ×400.
Figure 4 – Cross-section of umbilical artery (UA), umbilical vein (UV) and amniotic membrane (AM): (A–C) Control specimens show moderate expression of VEGFR-1 in the vascular endothelium (arrowheads), smooth muscles (M), Wharton’s jelly (W) and higher expression was observed in the amniotic epithelium (arrow); (D–F) Preeclampsia specimens show stronger VEGFR-1 expression in the umbilical vessel wall and moderate expression in the Wharton’s jelly (W) and in the amniotic epithelium (arrow). Anti-VEGFR-1 immunostaining: (A) ×100; (B, D and F) ×200; (C and E) ×400.

Shaima M. Almasry et al.
686
Figure 5 – Cross-section of umbilical artery (UA), umbilical vein (UV) and amniotic membrane (AM): (A–C) Control specimens show moderate expression of VEGFR-2 in the vascular endothelium (arrowheads), smooth muscles (M), Wharton’s jelly (W) and amniotic epithelium (arrow); (D–F) Preeclampsia specimens show absent VEGFR-2 expression in the umbilical vessel wall and weak expression in the Wharton’s jelly (W) and in the amniotic epithelium (arrow). Anti-VEGFR-2 immunostaining: (A) ×100; (B–F) ×200.
Figure 6 – Box plot analysis of area percent of VEGF, VEGFR-1 and VEGFR-2 expression in the human umbilical cord of normal pregnancy and preeclampsia. Stars indicate significant differences (p<0.05).
Discussion
Preeclampsia is the most common syndrome associated with pregnancy and is known to affect the vascular system of mother as well as the fetus. Assessment of maternal disease during pregnancy as well as intrinsic fetal sickness is facilitated via umbilical cord evaluation by different methods because of its close relation to fetal development.
According to our results, women belonging to PE had a shorter gestation period and delivered newborns with lower birth weight. That may be ascribed to the requirement of induction of labor in preterm and low birth weight fetus by the maternal and fetal indications in PE [14]. The fetal growth restriction and low weight
at birth in different diseases such as PE were reported as a result of fetal adaptive responses.
The imbalance between endogenous regulators of angiogenesis and compounds that modulate vascular tone in the placenta and UC can lead to pregnancy complications [14]. The subject area of endothelial and smooth muscle cells of the UC vessels was reported to await for the alterations involved in the functional vascular changes [15]. Also, the measures of total vascular areas, luminal areas and luminal indices of the umbilical vessels are important from a research perspective as they are the gateway to the fetus from the mother.
In the present work, degenerative changes were found in the wall of umbilical vessels. The vascular endothelium appeared with areas of discontinuation with the presence of pyknotic darker stained nuclei. Also, spaces were determined within the disorganized muscle layers and the surrounding Wharton’s jelly. These findings are inconsis-tent with what’s described by Barnwal et al. [2] on the umbilical vein from PE women. Loss of endothelial cell integrity and endothelial dysfunction could be a key pathogenic feature of PE with a resulting increase in vascular permeability and is considered to underlie hyper-tension, proteinuria, and edema [16]. Along the other point of view, the absence of vasa vasorum in the wall of umbilical vessels enforces them to depend on their own oxygen supply and so being more vulnerable to the influence of hypoxia on the metabolism of both endothelial and smooth muscle cells. Therefore, degenerative changes could reflect the maternal hemodynamic circumstances associated with PE leading to fetal hypoxia, fetal hyper-tension and a discontinuous umbilical blood flow [17].
In the current work, the smooth muscle layer of umbilical arteries showed some areas of thickening with

Ultrastructure and histomorphometric analysis of human umbilical cord vessels in preeclampsia: a potential role…
687
increased total vascular areas, luminal venous area and luminal venous index. Similar findings were observed by others [2], who proposed that the umbilical vein compensates or increases the effective contractile units only up to a certain limit of rise in pressure and with further rise in luminal pressure, it is unable to recompense and that could explain the dilatation and thinning of the wall of umbilical vein in PE. The expansion in the umbilical artery wall could be due to an increment in both tunica intima and tunica media thickness. Some studies [18, 19] mentioned various theories that hypoxia alone in an autocrine mechanism acts as a stimulus on smooth muscle cells to initiate proliferation. Similar findings were observed in gestational diabetes; Chakraborty et al. [20] found an increase in the cross-sectional area of the Wharton’s jelly accompanied by the appearance of numerous empty spaces within it, which might represent a degenerative effect of diabetes mellitus.
Smooth muscle cell subendothelial infiltration was observed in umbilical vessels of the PE group in the current study. Migration of smooth muscle cells towards the endothelium was accompanied by a splitting of internal elastic lamina, leading to thickening of the tunica intima. This migration is supposed to be due to augmentation of sulphated glycosaminoglycans in UC arteries and reduced expression of elastin. That might represent part of the functional adaptation system of the UC arteries to the altered hemodynamic conditions in PE [19]. That could be attributed to VEGF, which act as a smooth muscle cell chemotactant and stimulates smooth muscle cell prolife-ration [21] or through the combined actions of growth factors, proteolytic agents, and extracellular matrix proteins that are produced by a dysfunctional endothelium following injury or hypoxia induce proliferation and migration of resident smooth muscle cell from the media into the intima leading to intimal hyperplasia [22].
Currently, VEGF expression was significantly elevated in UC tissues of PE. That is in consistence with the finding of Bates [8] on the maternal serum. VEGF has been suggested to be involved in the pathogenesis of PE rather than being an effect of the disease. Rapid placental vascu-larization observed in the second half of pig pregnancy and is positively correlated with VEGF, which have a stimulatory effect in the dynamically growing UC of the pigs [23]. The levels of VEGF were reported to be decreased with gestation and expression is exaggerated in conditions of suggested hypoxia [8, 24]. Increased VEGF expression and decreased occludin expression caused increased permeability of the endothelial barrier. VEGF might be affected in the generalized impairment of endothelium dependent dilatation, which would increase blood pressure in PE [3, 25].
In the current study, the human UC sections obtained from PE pregnancies showed significant higher areas of expression of VEGFR-1 compared to the healthy specimens. Immunohistochemical staining enabled detection of VEGF–VEGF receptor system, in the endothelium and tunica media cells of the umbilical vessels and in allantoic duct and amniotic epithelium on all analyzed days of gestation [26]. That is in agreement with Helske et al. [27] who observed that VEGFR-1 was increased in the placenta in
PE and fetal growth restriction with strong immuno-reaction in endothelial cells of villous capillaries. Padavala et al. [28] reported that in intrauterine growth retardation pregnancies, under restricted oxygen, placental angiogenic potential is suppressed through the overproduction of placental VEGFR-1. This may discourage normal placental vascularization and impact on fetal growth. Interestingly, they stated that measurements of cord reiterate a potential inverse relationship between VEGF and VEGFR-1 in the fetal circulation. Sung et al. [29] confirmed an increase in VEGFR-1 as a potent inhibitor of VEGF in PE preg-nancies. Shibuya [7] suggested that VEGFR-1 is strongly antagonistic to VEGF and neutralizes its effects. Abnormal elevation of circulating levels of the VEGFR-1 has been widely accepted as a biomarker of PE and as a contributor to the systemic endothelial dysfunction and clinical mani-festations of the disease through inhibition of VEGF activity causing failure of the vascular remodeling [29–31]. Moreover, anti-VEGF therapies that either block the extracellular binding of VEGF to its receptor (anti-VEGF antibodies) or VEGF receptor inhibitors, became an innovative target in the treatment of cancers through angiogenesis suppression and leading to hypertension, proteinuria similar to the phenomena seen in PE [32].
The significantly lower expression of VEGFR-2 detected in PE compared to healthy pregnancies is in accordance with Tripathi et al. [33] suggested that the reduced serum levels of soluble VEGFR-2 and the down-regulated expression of membranous VEGFR-2 in placenta, denoted abnormality in VEGF-mediated placental function and thus VEGFR-2 may be a key factor, intimately asso-ciated with PE. They added that VEGFR-1 is considered to be a ‘decoy’ receptor and VEGFR-2, the major mediator of the mitogenic, permeability enhancing endothelial survival effects of VEGF.
VEGFR-2 is believed to be the more important receptor with regard to induction of increased microvascular per-meability and for both physiological and pathological angiogenesis [34, 35]. Although VEGFR-2 has lower affinity for VEGF than VEGFR-1, VEGFR-2 exhibits robust protein–tyrosine kinase activity in response to its ligands [36]. Hence, VEGFR-2 plays a more important functional role in mediating signaling events involved in endothelial cell mitogenesis, migration, survival and vascular permeability [37]. Endothelial cells are one source of plasma soluble VEGFR-2 [38], which is differentially expressed under hypoxia in smooth muscle cells [39].
Expression of the VEGF–VEGFR-2 system in UC can be involved in vessel relaxation by interaction with nitric oxide synthase pathways [23]. Takahashi et al. [40] noticed that VEGFR-1 by itself does not generate a strong signal for vascular permeability activity; it cooperates with VEGFR-2 to significantly enhance the permeability and is involved in several diseases.
Conclusions
Modulation in the VEGF/VEGFR-1/VEGFR-2 system expression in the umbilical cord tissues could have a role in pathogenesis of PE through affecting the umbilical vascular diameters. In PE specimens, there was a

Shaima M. Almasry et al.
688
narrowing of the arterial luminal index, widening of the venous luminal index, discontinuation in the vascular endothelium and disorganization of the smooth muscles. Increased VEGF/VEGFR-1 caused endothelial dysfunction and failure of the vascular remodeling. Endothelial cell dysfunction produced down-regulation of VEGFR-2 and thus affecting the endothelial cell migration, survival and vascular permeability.
Conflict of interests The authors declare that they have no conflict of
interests.
References [1] Benirschke K, Kaufmann P, Baergen RN. Pathology of the
human placenta. 5th edition, Springer-Verlag, New York, 2006. [2] Barnwal M, Rathi SK, Chhabra S, Nanda S. Histomorphometry
of umbilical cord and its vessels in pre-eclampsia as compared to normal pregnancies. Nepal J Obstet Gynecol, 2012, 7(1): 28–31.
[3] Feng D, Nagy JA, Brekken RA Pettersson A, Manseau EJ, Pyne K, Mulligan R, Thorpe PE, Dvorak HF, Dvorak AM. Ultrastructural localization of the vascular permeability factor/ vascular endothelial growth factor (VPF/VEGF) receptor-2 (FLK-1, KDR) in normal mouse kidney and in the hyper-permeable vessels induced by VPF/VEGF-expressing tumors and adenoviral vectors. J Histochem Cytochem, 2000, 48(4): 545–556.
[4] Petla LT, Chikkala R, Ratnakar KS, Kodati V, Sritharan V. Biomarkers for the management of pre-eclampsia in pregnant women. Indian J Med Res, 2013, 138:60–67.
[5] Pennington KA, Schlitt JM, Jackson DL, Schulz LC, Schust DJ. Preeclampsia: multiple approaches for a multifactorial disease. Dis Model Mech, 2012, 5(1):9–18.
[6] Niu G, Chen X. Vascular endothelial growth factor as an anti-angiogenic target for cancer therapy. Curr Drug Targets, 2010, 11(8):1000–1017.
[7] Shibuya M. Vascular endothelial growth factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: a crucial target for anti- and pro-angiogenic therapies. Genes Cancer, 2011, 2(12):1097–1105.
[8] Bates DO. An unexpected tail of VEGF and PlGF in pre-eclampsia. Biochem Soc Trans, 2011, 39(6):1576–1582.
[9] Garg N, Polipalli SK, Kapoor S. Genetic conflicts and patho-physiological changes in pregnancy: a risk factor for pre-eclampsia. World J Pharm Pharm Sci, 2015, 4(11):549–583.
[10] Inan S, Sanci M, Can D, Vatansever S, Oztekin O, Tinar S. Comparative morphological differences between umbilical cords from chronic hypertensive and preeclamptic pregnancies. Acta Med Okayama, 2002, 56(4):177–186.
[11] Bancroft JD, Gamble M. Theory and practice of histological techniques. 6th edition, Churchill Livingstone, London, 2008, 125–138.
[12] Kiernan JA. Histological and histochemical methods: theory and practice. Butterworth–Heinemann, Oxford, 1999.
[13] Bozzola JJ, Russell LD. Electron microscopy: principles and techniques for biologists. 2nd edition, Jones & Bartlett Publi-shers, Boston, 1999.
[14] Ahmed A, Ramma W. Unravelling the theories of pre-eclampsia: are the protective pathways the new paradigm? Br J Pharmacol, 2015, 172(6):1574–1586.
[15] Martín de Llano JJ, Fuertes G, Torró I, García Vicent C, Fayos JL, Lurbe E. Birth weight and characteristics of endo-thelial and smooth muscle cell cultures from human umbilical cord vessels. J Translat Med, 2009, 7:30.
[16] Uzan J, Carbonnel M, Piconne O, Asmar R, Ayoubi JM. Pre-eclampsia: pathophysiology, diagnosis, and management. Vasc Health Risk Manag, 2011, 7:467–474.
[17] Junek T, Baum O, Läuter H, Vetter K, Matejevic D, Graf R. Pre-eclampsia associated alterations of the elastic fibre system in umbilical cord vessels. Anat Embryol (Berl), 2000, 201(4):291–303.
[18] Schultz K, Fanburg BL, Beasley D. Hypoxia and hypoxia-inducible factor-1alpha promote growth factor-induced proli-
feration of human vascular smooth muscle cells. Am J Physiol Heart Circ Physiol, 2006, 290(6):H2528–H2534.
[19] Ray JB, Arab S, Deng Y Liu P, Penn L, Courtman DW, Ward ME. Oxygen regulation of arterial smooth muscle cell proliferation and survival. Am J Physiol Heart Circ Physiol, 2008, 294(2): H839–H852.
[20] Chakraborty SK, Ali Yousuf BM, Banu LA, Shamim KM. A gross and histomorphological study of the umbilical cord in gestational diabetes mellitus. Bangladesh J Anat, 2011, 9(1): 21–25.
[21] Louis SF, Zahradka P. Vascular smooth muscle cell motility: from migration to invasion. Exp Clin Cardiol, 2010, 15(4):e75–e85.
[22] Muto A, Fitzgerald TN, Pimiento JM, Maloney SP, Teso D, Paszkowiak JJ, Westvik TS, Kudo FA, Nishibe T, Dardik A. Smooth muscle cell signal transduction: implications of vascular biology for vascular surgeons. J Vasc Surg, 2007, 45(Suppl A):A15–A24.
[23] Chruściel M, Andronowska A, Postek A. Expression patterns of endothelial and inducible nitric oxide isoforms in the porcine umbilical cord. Reprod Domest Anim, 2009, 44(4):621–630.
[24] Maynard SE, Karumanchi SA. Angiogenic factors and pre-eclampsia. Semin Nephrol, 2011, 31(1):33–46.
[25] Conklin BS, Vito RP, Chen C. Effect of low shear stress on permeability and occludin expression in porcine artery endothelial cells. World J Surg, 2007, 31(4):733–743.
[26] Chruściel M, Ziençik AJ, Andronowska A. Expression of the vascular endothelial growth factor (VEGF-A) and its receptors in the umbilical cord in the course of pregnancy in the pig. Reprod Domest Anim, 2011, 46(3):434–443.
[27] Helske S, Vuorela P, Carpén O, Hornig C, Weich H, Halme-smäki E. Expression of vascular endothelial growth factor receptors 1, 2 and 3 in placentas from normal and compli-cated pregnancies. Mol Hum Reprod, 2001, 7(2):205–210.
[28] Padavala S, Pope N, Baker P, Crocker I. An imbalance between vascular endothelial growth factor and its soluble receptor in placental villous explants of intrauterine growth-restricted pregnancies. J Soc Gynecol Investig, 2006, 13(1): 40–47.
[29] Sung JF, Fan X, Dhal S, Dwyer BK, Jafari A, El-Sayed YY, Druzin ML, Nayak NR. Decreased circulating soluble Tie2 levels in preeclampsia may result from inhibition of vascular endothelial growth factor (VEGF) signaling. J Clin Endocrinol Metab, 2011, 96(7):E1148–E1152.
[30] Roberts JM, Rajakumar A. Preeclampsia and soluble fms-like tyrosine kinase 1. J Clin Endocrinol Metab, 2009, 94(7): 2252–2254.
[31] Dwyer BK, Krieg S, Balise R, Carroll IR, Chueh J, Nayak N, Druzin M. Variable expression of soluble fms-like tyrosine kinase 1 in patients at high risk for preeclampsia. J Matern Fetal Neonatal Med, 2010, 23(7):705–711.
[32] Müller-Deile J, Schiffer M. Renal involvement in preeclampsia: similarities to VEGF ablation therapy. J Pregnancy, 2011, 2011:176973.
[33] Tripathi R, Rath G, Ralhan R, Saxena S, Salhan S. Soluble and membranous vascular endothelial growth factor receptor-2 in pregnancies complicated by pre-eclampsia. Yonsei Med J, 2009, 50(5):656–666.
[34] Agostini H, Boden K, Unsöld A, Martin G, Hansen L, Fiedler U, Esser N, Marmé D. A single local injection of recombinant VEGF receptor 2 but not of Tie2 inhibits retinal neovascula-rization in the mouse. Curr Eye Res, 2005, 30(4):249–257.
[35] Shibuya M. Differential roles of vascular endothelial growth factor receptor-1 and receptor-2 in angiogenesis. J Biochem Mol Biol, 2006, 39(5):469–478.
[36] Chrusciel M, Bodek G, Kirtiklis L Lewczuk B, Hyder CL, Blitek A, Kaczmarek MM, Ziecik AJ, Andronowska A. Immor-talization of swine umbilical vein endothelial cells (SUVECs) with the simian virus 40 large-T antigen. Mol Reprod Dev, 2011, 78(8):597–610.
[37] Shibuya M. Structure and dual function of vascular endothelial growth factor receptor-1 (Flt-1). Int J Biochem Cell Biol, 2001, 33(4):409–420.
[38] Ebos JM, Bocci G, Man S Thorpe PE, Hicklin DJ, Zhou D, Jia X, Kerbel RS. A naturally occurring soluble form of vascular endothelial growth factor receptor 2 detected in mouse and human plasma. Mol Cancer Res, 2004, 2(6):315–326.

Ultrastructure and histomorphometric analysis of human umbilical cord vessels in preeclampsia: a potential role…
689
[39] Chanakira A, Dutta R, Charboneau R, Barke R, Santilli SM, Roy S. Hypoxia differentially regulates arterial and venous smooth muscle cell proliferation via PDGFR-β and VEGFR-2 expression. Am J Physiol Heart Circ Physiol, 2012, 302(5): H1173–H1184.
[40] Takahashi H, Hattori S, Iwamatsu A, Takizawa H, Shibuya M. A novel snake venom vascular endothelial growth factor (VEGF) predominantly induces vascular permeability through prefe-rential signaling via VEGF receptor-1. J Biol Chem, 2004, 279(44):46304–46314.
Corresponding author Shaima Mohamad Almasry, Department of Anatomy and Embryology, Taibah University, Al-Madinah Al-Munawarah, Kingdom of Saudi Arabia; Phone 009660594380057, Box 41411, e-mail: [email protected] Received: March 25, 2016
Accepted: August 14, 2016





























