Embed Size (px)
Citation preview

of February 11, 2018.This information is current as
Leishmania donovaniof Antimony-Susceptible and -Resistant StrainsInduces Protection against Both Cell-Mediated Immune Response andFrame F DNA Vaccine Leads to Enhanced Ubiquitin Conjugation of Open Reading
Ankur Sharma and Rentala Madhubala
http://www.jimmunol.org/content/183/12/7719doi: 10.4049/jimmunol.0900132November 2009;
2009; 183:7719-7731; Prepublished online 23J Immunol
average*
4 weeks from acceptance to publicationSpeedy Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
?The JIWhy
Referenceshttp://www.jimmunol.org/content/183/12/7719.full#ref-list-1
, 30 of which you can access for free at: cites 69 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Ubiquitin Conjugation of Open Reading Frame F DNAVaccine Leads to Enhanced Cell-Mediated Immune Responseand Induces Protection against Both Antimony-Susceptible and-Resistant Strains of Leishmania donovani1
Ankur Sharma and Rentala Madhubala2
Resistance of Leishmania donovani to sodium antimony gluconate has become a critical issue in the current, prolonged epidemicin India. Hence, there is an urgent need for a vaccine that is protective against both antimony-susceptible and -resistant strainsof L. donovani. The multigene LD1 locus located on chromosome 35 of Leishmania is amplified in �15% of the isolates examined.The open reading frame F (ORFF), a potential vaccine candidate against visceral leishmaniasis, is part of the multigene LD1 locus.ORFF was expressed as a chimeric conjugate of ubiquitin to elicit an Ag-specific cell-mediated immune response. Analysis of thecellular immune responses of ubiquitin-conjugated ORFF (UBQ-ORFF) DNA-immunized, uninfected BALB/c mice demonstratedthat the vaccine induced enhanced IFN-�-producing CD4� and CD8� T cells compared with nonubiquitinated ORFF DNAvaccine. Higher levels of IL-12 and IFN-� and the low levels of IL-4 and IL-10 further indicated that the immune responses withUBQ-ORFF were mediated toward the Th1 rather than Th2 type. Infection of immunized mice with either the antimony-sus-ceptible (AG83) or -resistant (GE1F8R) L. donovani strain showed that UBQ-ORFF DNA vaccine induced higher protection whencompared with ORFF DNA. UBQ-ORFF DNA-immunized and -infected mice showed a significant increase in IL-12 and IFN-�and significant down-regulation of IL-10. High levels of production of nitrite and superoxide, two macrophage-derived oxidantsthat are critical in controlling Leishmania infection, were observed in protected mice. The feasibility of using ubiquitinated-conjugated ORFF DNA vaccine as a promising immune enhancer for vaccination against both antimony-susceptible and -resistantstrains of L. donovani is reported. The Journal of Immunology, 2009, 183: 7719–7731.
L eishmaniasis constitutes a wide spectrum of diseasesranging from the simple self-limiting cutaneous form tothe debilitating visceral form, which is often fatal if left
untreated. The protozoan parasite Leishmania donovani is the ma-jor causative agent of visceral leishmaniasis. Leishmania is a di-genetic parasite, existing as flagellate promastigote in sand fly vec-tor, which further infect human macrophages and colonize asnonmotile amastigotes. Two million new cases (1.5 million forcutaneous leishmaniasis and 500,000 for visceral leishmaniasis(VL)3 are considered to occur annually, with an estimated 12 mil-lion people presently infected worldwide (World Health Organi-zation, Leishmaniasis Control home page: http://www.who.int/leishmaniasis/en/) (1–3).
Current control measures rely on chemotherapy to alleviate dis-ease and on vector control to reduce the transmission. However,current chemotherapeutic agents are unsuitable, in part because oftheir high toxicity and the emergence of drug resistance (4, 5).Thus, identification of novel immunological targets that can beeffective against both antimony-susceptible and -resistant strains ofLeishmania is of tremendous economic and medical importance(6). Leishmaniasis is considered one of the few parasitic diseaseslikely to be controlled by vaccination, especially because of anuncomplicated leishmanial life cycle and the fact that recoveryfrom infection renders the host resistant to subsequent infection.Considerable efforts have been made to develop a vaccine to in-duce specific antiparasite immune responses (7). Several candidatevaccine molecules have already been evaluated in the mice modelfor leishmaniasis (8–14). Treatment and vaccination that controlthe disease in susceptible mice invariably promote Th1 responsesover Th2 responses (15–18). However, this may or may not holdtrue for other models of leishmaniasis. In mice infected with L.donovani, the differential production of Th1 and Th2 cytokine doesnot control the cure rate; although the Th1 response correlates withresistance to infection, the Th2 response does not determine sus-ceptibility (19). The biological phenotype of the immune responseis determined in a large part by the predominance of one cell typeover the other, not simply the presence or absence of Th1- orTh2-type immune cells.
We have previously examined the ability of the open readingframe F (ORFF) gene from the LD1 locus to induce protectiveimmunity in BALB/c mice infected with L. donovani. The multi-gene LD1 locus is present on chromosome 35 in all Leishmaniaisolates examined (20–22). The ORFF protein is present in both
School of Life Sciences, Jawaharlal Nehru University, New Delhi, India
Received for publication January 14, 2009. Accepted for publication October13, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work is supported by the Life Science Research Board Grant from the DefenseResearch and Development Organization, government of India (New Delhi, India) toR.M. A.S. is supported by the Department of Science and Technology, government ofIndia.2 Address correspondence and reprint requests to Prof. Rentala Madhubala, School ofLife Sciences, Jawaharlal Nehru University, New Delhi 110067, India. E-mailaddress: [email protected] Abbreviations used in this paper: VL, visceral leishmaniasis; ORFF, open readingframe F; rORFF, recombinant ORFF protein; UBQ, ubiquitin; ROS, reactive oxygenspecies; H2DCFDA, 2�,7�-dichlorodihyrofluorescein diacetate; SA, soluble Ag; LDU,Leishman Donovan unit; ROS, reactive oxygen species; UB-ORFF or UBQ-ORFF,ubiquitin-conjugated ORFF.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900132
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

promastigote and amastigote forms of the parasite but is pref-erentially expressed in L. donovani amastigotes (22). ORFF Agwas recognized by the sera of patients infected with L. donovani(23). Our previous studies have shown that immunization withrecombinant ORFF protein (rORFF) resulted in partial protec-tion against visceral leishmaniasis in mice (24). We have alsoshown that the protection induced by the administration of an ORFFDNA vaccine is much stronger than that induced by a rORFF proteinvaccine (25). Immunostimulatory oligodeoxynucleotides have alsobeen used as potent enhancers of protective immunity in mice immu-nized with rORFF Ag (26, 27). Besides this, we have also shown thata heterologous prime-boost vaccination regimen using ORFF DNAand rORFF protein confers protective immunity against visceral leish-maniasis (28). Coexpression of ORFF with IL-12 resulted in protec-tion against challenge with L. donovani (29).
Several molecular strategies have been developed to enhanceimmune responses to DNA vaccines that include coexpression ofcytokines (30), incorporation of CpG motifs (31, 32), fusion withplasminogen activator (33), and so forth. Recently, DNA con-structs have been developed that express proteins conjugated toubiquitin. Ubiquitin is a highly conserved small regulatory proteinthat is ubiquitous to eukaryotes. The main function of this 76-aapeptide is to mark other proteins for destruction, a process referredto as proteolysis. At least four ubiquitin molecules attach to alysine residue on the condemned protein, in a process called polyu-biquitination, and the protein then moves to proteasome whereproteolysis takes place (34). The ubiquitin conjugation enhancesproteasome-dependent degradation of the endogenously synthe-sized Ag and results in an increase of cell-mediated response in-duced in vivo against the conjugated Ag (35–37). A direct corre-lation between rapid Ag processing and enhanced cell-mediatedimmune responses has been established (38–40).
In the present study, we have developed a DNA vaccine ex-pressing ubiquitin conjugated to a leishmanial Ag, ORFF (UBQ-ORFF), and evaluated its immunological activity and ability togenerate protective immunity in vaccinated mice. Hence, studieswere performed to assess and compare the immune responses gen-erated by UBQ-ORFF vs ORFF DNA vaccine. The present dataindicated that a strong Th1-oriented immune response was gener-ated by DNA vaccine expressing a ubiquitin-conjugated protein,resulting in a protective immune response in the experimental VLmodel. We also report the protective effector mechanism of theubiquitin-conjugated ORFF DNA vaccine against both antimony-susceptible and -resistant strains of L. donovani.
Materials and MethodsReagents
The fluorescent probe H2DCFDA (2�,7�-dichlorodihydrofluorescein diac-etate) was obtained from Fluka. Medium M-199 and RPMI 1640, HEPES,penicillin, streptomycin, sulfanilamide, and N-1-naphthylethylenediaminedihydrochloride were obtained from Sigma-Aldrich. FCS was obtainedfrom Life Technologies. [3H]Thymidine was obtained from AmershamBiosciences. PerCP-conjugated rat anti-mouse CD3e, FITC-conjugated ratanti-mouse CD4, PE-conjugated rat anti-mouse CD8, allophycocyanin-conjugated rat anti-mouse IFN-�, and their respective isotype controls wereobtained from BD Biosciences. The Cytofix/Cytoperm Fixation/Permeabi-lization kit (with GolgiStop protein transport inhibitor containing brefeldinA) and mouse Fc Block were obtained from BD Biosciences. OptEIAELISA kits for IFN-�, IL-4, IL-10, and IL-12 and biotinylated rabbit anti-mouse IgG1 and IgG2a mAbs were also obtained from BD Biosciences.
Animals
Female BALB/c mice, all 4–6 wk old, were obtained from the NationalInstitute of Nutrition (Hyderabad, India) and were maintained in the insti-tute facility. All mice were at ages ranging from 6 to 8 wk during the study.Animals were used in accordance with the institutional guidelines and therelevant committee approved the use of mice for this study.
Parasite culture
L. donovani antimony-susceptible strain AG83 (MHOM/IN/1983/AG83)and L. donovani antimony-resistant strain GE1-R (MHOM/IN/89/GE1F8R)were originally isolated from Indian patients with VL and were maintainedin BALB/c mice by repeated passage as described previously (41). Theisolates were provided by Dr. M. Chatterjee (Institute of PostgraduateMedical Education & Research, Kolkata, India) (6, 24, 41). GE1F8R isapproximately eight times more resistant to antimony compared with antimo-ny-sensitive clone AG83 (42). Amastigotes were isolated from infected spleenand then transformed to promastigotes in Medium 199 (Sigma-Aldrich) sup-plemented with 100 �g/ml streptomycin, 100 U/ml penicillin (Sigma-Aldrich),and 25% FCS (Life Technologies). Freshly transformed promastigotes werecultured at 22°C in Medium 199 and 10% heat-inactivated FCS and were thenused for infecting BALB/c mice. Mice were injected i.v. via the tail veinwith100 �l of PBS/mouse with 1 � 108 promastigotes suspended in PBSusing a 22-gauge needle as reported elsewhere (43).
Construction of the ubiquitinated and nonubiquitinated ORFFexpression vectors
Mammalian expression vector pUMVC3 (http://www.med.umich.edu/vcore/plasmids/pUMVC3.htm) fused with the ubiquitin gene was a giftfrom Dr. U. K. Ranga (Jawaharlal Nehru Center for Advanced ScientificResearch, Bangalore, India) (37). Briefly, the 228-bp ubiquitin gene wasamplified from the genomic DNA extracted from mouse liver (GenBanksequence X51703) and cloned directionally between EcoRI and KpnI sitesinto the mammalian expression vector pUMVC3 under the control of aCMV promoter (37). The ORFF gene (GenBank accession no. L38571,http://www.ncbi.nlm.nih.gov/nuccore/703121) was amplified from theORFF DNA expression plasmid construct designated F/pcDNA3.1 (25)using the following primers: forward primer, CATGC CTTAAG ATGCAAAGCGACGCACG and reverse primer, GC TCTAGA CTACTCTGCAACCGCAGAC with flanking AflII and XbaI sites, respectively (under-lined). PCR was performed using PFU TaqDNA polymerase for 35 cycles ina Thermocycler (2720 Thermal Cycler; Applied Biosciences). The conditionsfor PCR were as follows: 94°C for 10 min, 94°C for 30 s, 57°C for 30 s, and72°C for 2 min. Final extension was conducted for 10 min at 72°C. The PCRproduct was digested with AflII and XbaI and the resulting amplicon wascloned directionally into the above ubiquitin vector between AflII and XbaIsites to give the construct pUMVC3-UB-ORFF (Fig. 1A). The cytoplasmicproteasome machinery is known to recognize the ubiquitin-fused protein andcleave the protein at the C-terminal of ubiquitin, thereby exposing the N-terminal amino acid of the tagged Ag. It has been reported earlier that thehalf-life of a protein is determined by the N-terminal amino acid residue ofprotein, i.e., proteins having serine, glycine, or valine as the N-terminal aminoacid were long-lived with a half-life of �20 h and proteins with aspartic acid,tryptophan, or leucine as their N-terminal amino acid have a half-life of only3 min (44). In the present construct, we used AflII as the restriction enzymesince the first triplet of this enzyme codes for leucine. Hence, when ORFF Agis guided to proteasome for its degradation, it would have leucine residue at itsN-terminal which would target the protein for rapid processing.
To obtain the pUMVC3-ORFF construct (without ubiquitin), ORFF wasamplified from the F/pcDNA3.1 plasmid using the following primers: for-ward primer, CAGTGC GGGCCC ATGCAAAGCGACGCACG and re-verse primer, GC TCTAGA CTACTCTGCAACCGCAGAC with flankingApaI and XbaI sites, respectively (underlined). PCR was performed usingPFU TaqDNA polymerase for 35 cycles. The conditions for PCR were asfollows: 94°C for 10 min, 94°C for 30 s, 57°C for 30 s, and 72°C for 2 min.Final extension was conducted for 10 min at 72°C. The PCR product wasdigested with ApaI and XbaI and the resulting amplicon was cloned direc-tionally into the pUMVC3 vector between ApaI and XbaI sites to give theconstruct pUMVC3-ORFF (Fig. 1B). Both the expression cassettes wereconfirmed by restriction enzyme analysis and sequencing. The pUMVC3blank vector was used in all control immunization groups. Plasmids weremaintained and propagated in Escherichia coli strain DH5�.
Source of ORFF Ag
rORFF from L. donovani strain AG83 (GenBank accession no.AAA96728.1, http://www.ncbi.nlm.nih.gov/protein/703122) was producedfrom the full-length fusion clone pET 17b-C14 by induction with isopropyl�-D-1-thiogalactopyranoside followed by sonication and purification usingurea gradient as described elsewhere (24).
Source of soluble Ag (SA)
SA was prepared from stationary phase L. donovani AG83 and GE1-Rpromastigotes as reported earlier (26). Briefly, cells were harvested bycentrifugation at 4000 � g for 15 min at 4°C followed by washing with
7720 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

PBS and were then resuspended in PBS containing protease inhibitors. Celllysis was done by sonication with six pulses of 30 s each (MSE Sonicator).The lysate was centrifuged at 17,000 � g for 15 min at 4°C and the su-pernatant containing SA was used for estimating protein concentration bythe Bradford method (45).
Immunization of mice
Female BALB/c mice (6–8 wk old) were used for all of the experiments.They were randomly divided into four groups (six mice in each group).Group 1 was immunized with saline solution, group 2 with blank vectorconstruct, group 3 with pUMVC3-ORFF, and group 4 with pUMVC3-UB-ORFF. The endotoxin-free plasmids intended for immunizations were pre-pared using EndoFree Giga kits (Qiagen) per the manufacturer’s instruc-tions. For immunization studies, 100 �g of endotoxin-free plasmid DNAconstructs (pUMVC3, pUMVC3-ORFF, pUMVC3-UB-ORFF) diluted inPBS were injected i.m. at the midpoint of the left thigh muscle of BALB/cmice. For vaccination, cell proliferation, cytokine production, and Ab re-sponse studies, BALB/c mice in the above-mentioned four groups wereimmunized twice at 2-wk intervals. Two weeks after the first two doses, asingle booster dose with the same constructs was given. Mice were bled 2wk following the final dose. Spleens were removed aseptically from micein each group. Serum samples and spleens were used for the analysis of thehumoral and cellular responses after immunization.
Assessment of parasite load after challenge
For the vaccination and protection studies, mice were injected i.m. withsaline solution (n � 24), 100 �g of blank vector construct (n � 24), 100�g of pUMVC3-UB-ORFF (n � 24), or 100 �g of pUMVC3-ORFF (n �24) and two booster doses were given at 2-wk intervals. The mice werethen challenged 2 wk after they received the final booster dose. Twelvemice from each of these four groups were challenged with 1 � 108 sta-tionary phase promastigotes of the antimony-sensitive L. donovani AG83strain. Promastigotes were injected i.v. via the tail vein with 100 �l ofPBS/mouse. The remaining 12 mice from each of the above four groupswere similarly challenged with 1 � 108 promastigotes of the antimony-resistant L. donovani GE1F8R strain. Four and 8 wk after infection, micewere euthanized and liver and spleen touch biopsies were microscopicallyexamined after fixing and staining the slides with Giemsa. To quantitate thelevel of infection, Leishman Donovan units (LDU) were calculated as(number of amastigotes/1000 tissue nuclei) � weight of tissue in mg, asreported earlier (25). Assessment of protection was performed using sixmice per group that were sacrificed for determination of parasitemia at 4and 8 wk after infection.
T cell proliferation assay
Spleens from mice of each group were removed aseptically on a sterile dishcontaining RPMI 1640 medium. Single-cell suspensions were prepared by
grinding the spleen with the disk bottom of the plunger of a 10-ml syringe.RPMI 1640 medium (5–10 ml) was added to the suspension and the con-tents were mixed well. The dish was kept undisturbed for 2 min and theclear supernatant was pipetted out slowly. Cells were pelleted by centrif-ugation at 4°C at 250 � g (Sorvall RC-5 centrifuge, HB-4 rotor) for 10min. The pellet was washed once with 0.9% ice-cold ammonium chlorideto lyse the erythrocytes. The remaining cells were resuspended to a densityof 2.5 � 106 cells/ml in RPMI 1640 containing 10% FCS and then dividedinto 200-�l aliquots (5 � 105 cells) in flat-bottom 96-well plates (Nunc).Cells were incubated in the presence or absence of rORFF (5 �g/ml) andincubated for 3 days at 37°C in a 5% CO2 incubator. Proliferation wasmeasured by incorporation of 1 �Ci of [3H]thymidine over the last 16 h ofthe culture. Incorporation of [3H]thymidine was measured by a beta scin-tillation counter (PerkinElmer 1450 LSC and Luminescence Plate Counter).All assays were performed in triplicates.
Intracellular cytokine staining
The assay was performed according to a Cytofix/Cytoperm Plus Fixation/Permeabilization kit (with BD Biosciences GolgiPlug) protocol from BDBiosciences. Briefly, splenocytes from mice immunized with saline, blankvector, pUMVC3-ORFF, or pUMVC3-UB-ORFF were recovered, resus-pended in RPMI 1640 medium supplemented with 10% FCS, and incu-bated in a 96-well flat-bottom plate at a density of 2 � 106 cells/well. Thecells were stimulated with 5 �g/ml rORFF or PMA/ionomycin (as a pos-itive control) or without rORFF (as a negative control). After 2 h of stim-ulation/incubation at 37°C, brefeldin A (1 �g/ml, GolgiPlug; BD Bio-sciences) was added to each well and the incubation was resumed for anadditional 12 h at 37°C. Cells were blocked with 1 �g/ml BD BiosciencesFc Block for 15 min at 4°C and then stained with PerCP-anti-mouse-CD3e(BD Biosciences), FITC-anti-mouse-CD4 (BD Biosciences), and PE-anti-mouse-CD8 (BD Biosciences) or their respective isotype controls (BD Bio-sciences) in 50 �l of staining buffer for 30 min at 4°C. Cells were permeabil-ized and fixed using a BD Biosciences Cytofix/Cytoperm kit. Intracellularstaining was then performed according to the protocol provided by BD Bio-sciences. Allophycocyanin-anti-mouse-IFN-� (BD Biosciences) or its respec-tive isotype control was used for intracellular staining of the cells. Cells werefinally analyzed on a FACS, BD Biosciences FACSCalibur system using BDBiosciences CellQuest software.
Cytokine assays
The concentrations of IFN-�, IL-12, IL-4, and IL-10 in culture supernatantswere determined as described previously (31). Briefly, spleen cells fromBALB/c were isolated from each group of mice at different intervals fol-lowing the final booster and resuspended in RPMI 1640 medium supple-mented with 10% FCS. Cells were then incubated in a 96-well flat-bottomplate at a density of 5 � 105 cell/well and cultured with rORFF (5 �g/ml).
FIGURE 1. Schematic diagrams ofnonubiquitin and ubiquitin-ORFF ex-pression vectors. A, The ORFF genewas directionally cloned into the mam-malian expression vector pUMVC3fused with ubiquitin (37). Restrictionenzyme (AflII) was introduced at thejunction of ubiquitin and ORFF so as toget leucine at the N terminus of ORFF,thereby exposing the Ag to rapid intra-cellular processing. B, ORFF alonecloned directionally into the mamma-lian expression vector pUMVC3.
7721The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
After 48 h, supernatants were collected and diluted serially and cytokine con-centrations were quantitated using ELISA. The assay was performed using aBD Pharmingen Opt EIA kit according to the manufacturer’s instructions.
Quantification of NO
Splenocytes, 5 � 105 cells/well, from different groups of experimentalmice were incubated in a 96-well flat-bottom plate with or without rORFF(5 �g/ml) for 48 h in a CO2 incubator at 37°C. In the case of GE1F8R-infected and pUMVC3The culture supernatant was collected and analyzed fortheir nitrite content using Griess reagent containing 1% sulfanilamide in 5%H3PO4 and an aqueous solution of N-1-naphthylethylenediamine dihydrochlo-ride. Griess reagent was added to the culture supernatant at ratio of 1:1 andincubated for 15 min at 37°C in the dark. OD was determined at 548 nm usinga Microplate Reader (SpectraMax M2 from Molecular Devices). Concentra-tion of nitrite (NO2
�) was determined using the standard curve prepared fromthe known concentration of sodium nitrite.
Measurement of reactive oxygen species (ROS)
The cell-permeable fluorescent probe H2DCFDA was used to measure thelevels of ROS. Splenocytes (5 � 105) from different groups of mice wereincubated in a 96-well flat-bottom plate with or without rORFF (5 �g/ml)for 72 h in a CO2 incubator at 37°C and were then incubated withH2DCFDA (1 �g/200 �l/well) at room temperature for 20 min in the dark.Relative fluorescence was measured by using an excitation wavelength of504 nm and emission wavelength of 529 nm in a Microplate Reader (Spec-traMax M2 from Molecular Devices). Fluorometric measurements weremade in triplicate and expressed as fluorescence intensity units.
Measurement of anti-ORFF IgG
Serum Ig isotypes were assayed by ELISA using 100 ng of rORFF/well,isotype-specific secondary Abs (biotinylated rabbit anti-mouse IgG1 andIgG2a), and streptavidin-conjugated HRP (BD Pharmingen). Briefly, 96-well plates (Immulon-4; Nunc) were coated with 100 ng of rORFF in 50 �lof carbonate buffer (pH 9.2) and incubated overnight at 4°C. Plates wereblocked with 5% milk in PBST (PBS containing 0.05% Tween 20) at 37°Cfor 2 h to prevent nonspecific binding. Mouse serum (100 �l) was dilutedto a concentration of 1/300–1/30,000 in PBST and was added to the wellsand plates were further incubated for 2 h at 37°C. Biotinylated-conjugatedrabbit anti-mouse IgG1 and IgG2a were then added for 1 h at 37°C fol-lowed by a 1-h incubation with streptavidin-conjugated HRP. Color wasdeveloped by incubation with a 100-�l aliquot of o-phenylenediamine (5mg/10 ml in 50 mmol/L citrate-phosphate buffer, pH 5.2) and 10 �l of 30%H2O2 for 30 min. The absorbance at 490 nm was determined using a model7520 microplate reader (Cambridge Technology).
Statistical analysis
One-way ANOVA with Bonferroni’s post test was performed using Graph-Pad Prism version 5.0 for Windows. Values of p � 0.05 were consideredstatistically significant. Data are represented as mean � SD. The results arerepresentative of two independent experiments.
Nucleotide and protein sequence accession numbers of ORFFgene
The nucleotide and protein sequences are: gene identification no. L38571(http://www.ncbi.nlm.nih.gov/nuccore/703121) and protein identificationno AAA96728.1, (http://www.ncbi.nlm.nih.gov/protein/703122).
ResultsComparison of immune responses generated by ubiquitinatedORFF (pUMVC3-UB-ORFF) vs ORFF (pUMVC3-ORFF) DNAvaccine
To verify enhanced immune response generated by ubiquitinatedORFF vs nonubiquitinated ORFF DNA vaccine, we expressedORFF gene as a fusion protein with ubiquitin, as indicated inMaterials and Methods. We cloned the ORFF gene directionallydownstream of ubiquitin in the expression vector pUMVC3 (Fig.1, A and B). Status of the cloned genes was checked in mice 2 wkfollowing injection and was analyzed by PCR using ORFF-specificprimers followed by Southern blot analysis as reported earlier (25).A significant amount of ORFF DNA was detected in the immu-nized mice (data not shown).
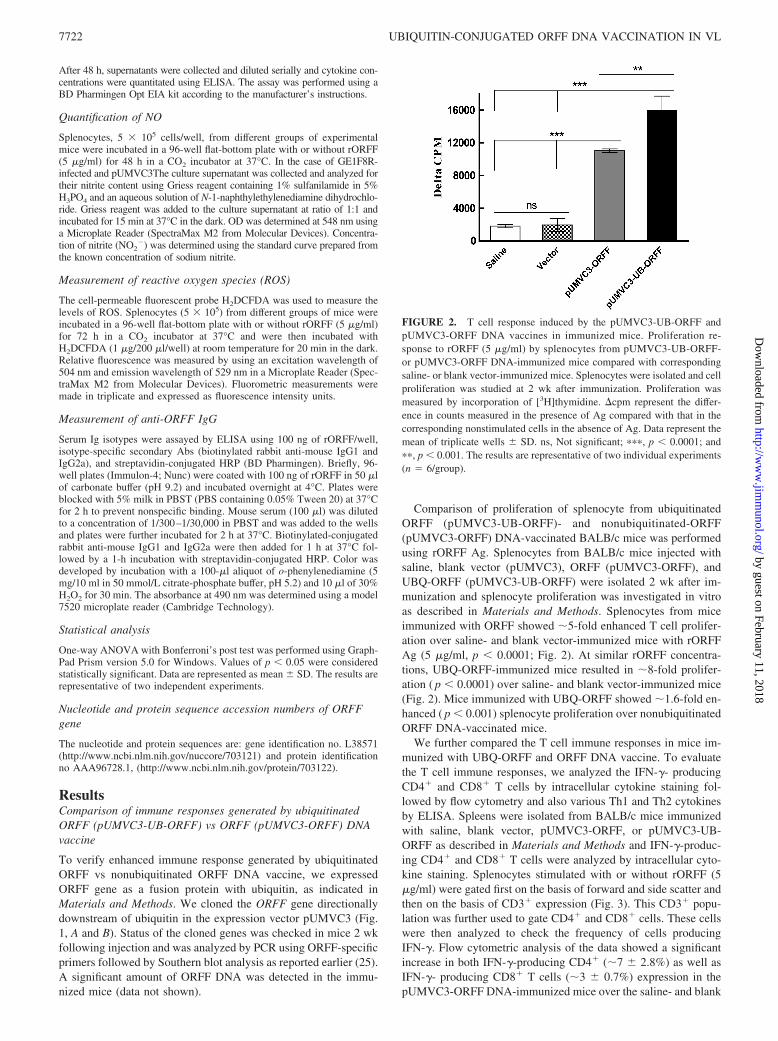
Comparison of proliferation of splenocyte from ubiquitinatedORFF (pUMVC3-UB-ORFF)- and nonubiquitinated-ORFF(pUMVC3-ORFF) DNA-vaccinated BALB/c mice was performedusing rORFF Ag. Splenocytes from BALB/c mice injected withsaline, blank vector (pUMVC3), ORFF (pUMVC3-ORFF), andUBQ-ORFF (pUMVC3-UB-ORFF) were isolated 2 wk after im-munization and splenocyte proliferation was investigated in vitroas described in Materials and Methods. Splenocytes from miceimmunized with ORFF showed �5-fold enhanced T cell prolifer-ation over saline- and blank vector-immunized mice with rORFFAg (5 �g/ml, p � 0.0001; Fig. 2). At similar rORFF concentra-tions, UBQ-ORFF-immunized mice resulted in �8-fold prolifer-ation ( p � 0.0001) over saline- and blank vector-immunized mice(Fig. 2). Mice immunized with UBQ-ORFF showed �1.6-fold en-hanced ( p � 0.001) splenocyte proliferation over nonubiquitinatedORFF DNA-vaccinated mice.
We further compared the T cell immune responses in mice im-munized with UBQ-ORFF and ORFF DNA vaccine. To evaluatethe T cell immune responses, we analyzed the IFN-�- producingCD4 and CD8 T cells by intracellular cytokine staining fol-lowed by flow cytometry and also various Th1 and Th2 cytokinesby ELISA. Spleens were isolated from BALB/c mice immunizedwith saline, blank vector, pUMVC3-ORFF, or pUMVC3-UB-ORFF as described in Materials and Methods and IFN-�-produc-ing CD4 and CD8 T cells were analyzed by intracellular cyto-kine staining. Splenocytes stimulated with or without rORFF (5�g/ml) were gated first on the basis of forward and side scatter andthen on the basis of CD3 expression (Fig. 3). This CD3 popu-lation was further used to gate CD4 and CD8 cells. These cellswere then analyzed to check the frequency of cells producingIFN-�. Flow cytometric analysis of the data showed a significantincrease in both IFN-�-producing CD4 (�7 � 2.8%) as well asIFN-�- producing CD8 T cells (�3 � 0.7%) expression in thepUMVC3-ORFF DNA-immunized mice over the saline- and blank
FIGURE 2. T cell response induced by the pUMVC3-UB-ORFF andpUMVC3-ORFF DNA vaccines in immunized mice. Proliferation re-sponse to rORFF (5 �g/ml) by splenocytes from pUMVC3-UB-ORFF-or pUMVC3-ORFF DNA-immunized mice compared with correspondingsaline- or blank vector-immunized mice. Splenocytes were isolated and cellproliferation was studied at 2 wk after immunization. Proliferation wasmeasured by incorporation of [3H]thymidine. cpm represent the differ-ence in counts measured in the presence of Ag compared with that in thecorresponding nonstimulated cells in the absence of Ag. Data represent themean of triplicate wells � SD. ns, Not significant; ���, p � 0.0001; and��, p � 0.001. The results are representative of two individual experiments(n � 6/group).
7722 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

vector-immunized groups (Fig. 3). However, mice immunizedwith UBQ-ORFF showed significantly higher expression of CD4
IFN-� (�13 � 1.7%; p � 0.005) and CD8 IFN-� T cells(�12 � 1.2%; p � 0.002) when compared with the values ob-tained in the ORFF DNA-immunized mice group (Fig. 3). Both thecontrol groups had negligible expression of CD4IFN-� andCD8IFN-� T cells. These results are in accordance with theIFN-� levels estimated by ELISA reported below (Fig. 4).
It has been reported earlier that IFN-� is involved in resistanceto infection in murine VL (46), whereas IL-10 correlates with sus-ceptibility (47). Based on the important role played by these cy-tokines, we checked the levels of Th1 and Th2 cytokines byELISA. It is considered that an effective vaccine against VL de-pends on the induction of a strong Th1-type, T cell-mediated re-sponse. To assess the intensity and the cell-mediated immunityprofile induced by ubiquitinated and nonubiquitinated DNA vac-cine, the splenic Ag-specific cells secreting IFN-�, IL-12, IL-4,and IL-10 were evaluated. Spleen cells from all groups of mice 2wk after immunization were stimulated with or without rORFF Agfor 48 h and the supernatants were tested for cytokines. The culture
supernatants from splenocytes of pUMVC3-UB-ORFF DNA- orpUMVC3-ORFF DNA-immunized mice contained significant lev-els of IFN-� and IL-12 compared with the vector- and saline alone-treated groups (Fig. 4, A and B). We found a �4-fold ( p � 0.0001)and �6-fold ( p � 0.0001) increase in IFN-� levels in mice im-munized with pUMVC3-ORFF and pUMVC3-UB-ORFF, respec-tively, compared with the saline and blank vector control groups.Mice immunized with UBQ-ORFF showed an �1.4-fold ( p �0.001) increase in IFN-� levels over nonubiquitinated ORFFDNA-vaccinated mice.
An �4.5-fold ( p � 0.0001) and �8-fold ( p � 0.0001) increasein IL-12 levels in mice immunized with pUMVC3-ORFF andpUMVC3-UB-ORFF, respectively, over the control groups wasobserved. Mice immunized with UBQ-ORFF showed an �1.8-fold( p � 0.0001) increase in IL-12 levels over nonubiquitinated ORFFDNA-vaccinated mice. In contrast to the robust Th1-type immuneresponses observed with pUMVC3-UB-ORFF DNA vaccine in com-parison to the pUMVC3-ORFF DNA vaccine, the levels of Th2 orinhibitory cytokines such as IL-4 and IL-10 were not significantlydifferent from those of the control groups (Fig. 4, C and D).
FIGURE 3. Flow cytometric analysis of ORFF-specific IFN-�-producing CD4 and CD8 T cells in mice immunized with pUMVC3-UB-ORFF orpUMVC3-ORFF DNA vaccines. BALB/c mice were immunized with saline, vector, pUMVC3-ORFF, or pUMVC3-UB-ORFF thrice at 2-wk intervals.Splenocytes were isolated and the emergence of IFN-�-producing CD4 and CD8 T cells was studied after stimulation of splenocytes with rORFF (5�g/ml) followed by blocking with brefeldin A as reported in Materials and Methods. Cells were further blocked with BD Biosciences Fc Block for 15 minat 4°C. The gated CD4 or CD8 cells are shown stained with allophycocyanin-labeled anti-IFN-�. Values in the quadrants represent the percentage ofpositive cells. The results are representative of two individual experiments (n � 6/group). Data represent the mean of triplicate wells � SD. SSC, Sidescatter; FSC, forward scatter.
7723The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In conclusion, pUMVC3-UB-ORFF generated a stronger im-mune response compared with pUMVC3-ORFF DNA vaccine.This is clearly evidenced by a higher T cell proliferation andhigher IFN-�-producing CD4 and CD8 T cells. Enhanced Th1cytokines IL-12 and IFN-� in the pUMVC3-UB-ORFF-immu-nized mice compared with mice immunized with pUMVC3-ORFFfurther strengthens our conclusion.
Parasite load in ORFF- or UBQ-ORFF DNA-vaccinated micefollowing challenge with antimony-susceptible and -resistantstrains of L. donovani
Since pUMVC3-UB-ORFF showed a stronger cell-mediated im-mune response in comparison to pUMVC3-ORFF DNA vaccine,we went ahead to compare the efficacy of pUMVC3-ORFF andpUMVC3-UB-ORFF DNA vaccines on the clearance of splenicand hepatic parasite burden following challenge with antimony-susceptible (AG83) and -resistant (GE1F8R) strains of L. dono-vani. For checking the efficacy of pUMVC3-ORFF or pUMVC3-UB-ORFF DNA vaccination in VL, we immunized and challengedthe mice with either antimony-susceptible (AG83) or antimony-resistant (GE1F8R) strains and the parasite burdens in the spleensand liver were evaluated 4 and 8 wk after challenge. The degree ofprotection against infection was determined by amastigote levelsin the liver and spleen and quantitated as LDU in liver and spleenbiopsies (Fig. 5). As shown in Fig. 5, control mice presented ahigher parasite burden in the liver at 4 and 8 wk after challenge,whereas AG83-infected and pUMVC3-UB-ORFF-vaccinatedmice 4 and 8 wk after infection showed 65% ( p � 0.001) and 80%( p � 0.0001) reduction in parasite load, respectively, in liver whencompared with the corresponding infected and immunized controlmice with saline or blank vector.
In the case of GE1F8R-infected and pUMVC3-UB-ORFF-vac-cinated mice, there was a similar reduction in parasite load of 65%( p � 0.001) and 80% ( p � 0.0001) at 4 and 8 wk, respectively, inliver biopsies. However, the pUMVC3-ORFF- vaccinated and ei-ther AG83- or GE1F8R-infected mice showed a similar reduction inparasite load in the liver of �65% ( p � 0.001) and 70% ( p � 0.0001)after challenge at 4 and 8 wk, respectively, compared with the controlgroups.
Parasite load was also checked in the spleen. The parasite loaddecreased by 75% ( p � 0.001) and 90% ( p � 0.0001) at 4 and 8wk, respectively, in AG83�infected and pUMVC3-UB-ORFF-im-munized mice and by 70% ( p � 0.001) and 85% ( p � 0.0001) at4 and 8 wk, respectively, in GE1F8R-infected and pUMVC3-UB-ORFF-immunized mice. On the other hand, the pUMVC3-ORFF-vaccinated and either AG83- or GE1F8R- infected mice at 4 and 8wk, respectively, after infection showed �70% ( p � 0.0001) and75% ( p � 0.0001) reduction, respectively, in splenic parasite loadcompared with the corresponding infected and immunized controlmice with saline and blank vector. Mice immunized with UBQ-ORFF and challenged with either AG83 or GE1F8R showed an�2.0-fold ( p � 0.01) reduction in parasite load in both spleen andliver at 8 wk after challenge over nonubiquitinated ORFF DNA-vaccinated and AG83- or GE1F8R-challenged mice.
Proliferation of splenocytes and T cell immune response in micevaccinated with ORFF or UBQ-ORFF DNA and challengedwith antimony-susceptible and -resistant strains of L. donovani
In VL, in vitro T cell proliferation has been reported to be impaired(48, 49). To check the status of T cell proliferation in the presentset of experiment, mice were immunized with saline, blank vector,pUMVC3-ORFF, or pUMVC3-UB-ORFF and were then challenged
FIGURE 4. Comparison of produc-tion of cytokines by spleen cells frompUMVC3-ORFF- or pUMVC3-UB-ORFF-immunized mice. BALB/c micewere immunized (i.m.) with saline, vec-tor, pUMVC3-ORFF, and pUMVC3-UB-ORFF thrice at 2-wk intervals.Two weeks after the final dose,spleens were collected as describedin Materials and Methods. Cytokinelevels of (A) IFN-�, (B) IL-12, (C)IL-4, and (D) IL-10 in the culturesupernatant were determined in re-sponse to rORFF (5 �g/ml) bysplenocytes from pUMVC3-UB-ORFF- or pUMVC3-ORFF DNA-immunized mice and were com-pared with corresponding saline- orblank vector-immunized mice. Theresults are representative of two in-dividual experiments (n � 6/group).Data represent the mean of triplicatewells � SD. ��, p � 0.001; ���,p � 0.0001; and ns, not significant.
7724 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

with either the antimony-sensitive strain AG83 or -resistant strainGE1F8R of L. donovani. Eight weeks after the challenge, spleno-cyte proliferation was studied in spleen supernatant as described inMaterials and Methods. Splenocytes from ORFF DNA- vaccinatedand either AG83- or GE1F8R-infected mice showed an �2.5-foldenhanced proliferation with rORFF (5 �g/ml; p � 0.0001) com-pared with the control groups. Splenocytes from AG83-challengedUBQ-ORFF DNA-immunized mice showed an � 4-fold enhancedT cell proliferation with rORFF (5 �g/ml; p � 0.0001) than in-fected and blank vector-immunized mice. A similar �4-fold in-crease in proliferation was observed with rORFF (5 �g/ml) inGE1F8R-challenged and UBQ-ORFF DNA-immunized mice( p � 0.0001) compared with infected and blank vector-immunizedmice (Fig. 6). Mice challenged with either AG83 or GE1F8R andimmunized with UBQ-ORFF DNA showed an �1.5-fold en-hanced ( p � 0.0001) splenocyte proliferation over nonubiquiti-nated ORFF DNA-vaccinated mice.
We further analyzed the IFN-�-producing CD4 and CD8
T cells in the splenocytes of AG83- and GE1F8R-challengedmice by intracellular cytokine staining using flow cytometry. Micewere immunized with saline, blank vector, pUMVC3-ORFF, orpUMVC3-UB-ORFF and then challenged with either the antimo-ny-sensitive strain AG83 or -resistant strain GE1F8R promastig-otes. Eight weeks after the challenge, mice were sacrificed and theIFN-�-producing CD4 and CD8 T cells were analyzed as re-ported in Materials and Methods (Fig. 7). Splenocytes stimulatedwith or without rORFF (5 �g/ml) were gated as reported above.Flow cytometric analysis of the data showed �8 � 1.4% and�7 � 0.7% IFN-�-producing CD4 and CD8 T cells, respec-tively, in the AG83-challenged pUMVC3-ORFF DNA-immunized
FIGURE 5. Protection against an-timony-susceptible and -resistant L.donovani infection in mice immu-nized with pUMVC3-ORFF DNA orpUMVC3-UB-ORFF DNA vaccine.BALB/c mice were immunized withsaline, vector, pUMVC3-ORFF, orpUMVC3-UB-ORFF thrice at 2-wkintervals. Two weeks following thefinal dose, one-half of the mice werechallenged (i.v.) with 1 � 108 pro-mastigotes of either antimony-sus-ceptible AG83 and the remainingwith antimony-resistant GE1F8R L.donovani. Four and 8 wk after chal-lenge infection, mice were euthanizedand LDU was calculated from liverand spleen smears and were com-pared with corresponding saline- orblank vector-immunized mice. Theresults are representative of two indi-vidual experiments. Results are ex-pressed as mean LDU � SD for sixanimals per group. �, p � 0.01; ��,p � 0.001; ���, p � 0.0001; and ns,not significant.
FIGURE 6. Splenocyte proliferation in protected pUMVC3-UB-ORFF DNA- or pUMVC3-ORFF DNA-vaccinated mice. Proliferationresponse to ORFF (5 �g/ml) by splenocytes from pUMVC3-ORFF- orpUMVC3-UB-ORFF-immunized and AG83 (antimony-susceptible)- orGE1F8R (antimony-resistant)-challenged mice compared with corre-sponding saline- and blank vector- immunized and -infected mice.Splenocytes were isolated and cell proliferation was studied at 8 wkafter the challenge. Proliferation was measured by incorporation of[3H]thymidine. cpm represent the difference in counts measured in thepresence of Ag compared with that in the corresponding nonstimulatedcells in the absence of Ag. Data represent the mean of triplicate wells �SD. The results are representative of two individual experiments (n �6/group). ���, p � 0.0001.
7725The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

mice (Fig. 7) compared with �16 � 2.1% ( p � 0.005) and �17 �1.4% ( p � 0.005) of IFN-�-producing CD4 and CD8 T cells,respectively, in the UBQ-ORFF-immunized and AG83-challengedmice (Fig. 7). The control groups had negligible expression ofCD4IFN-� and CD8IFN-� T cells.
GE1F8R-challenged and pUMVC3-UB-ORFF DNA- immu-nized mice showed a higher percentage of CD4IFN-� (�17 �2.8%) and CD8IFN-� T cells (�15 � 0.35%) compared withIFN-�-producing CD4 (�8 � 2.1%; p � 0.005) and CD8 Tcells (�5 � 1.4%; p � 0.005) in the ORFF-immunized andGE1F8R-challenged mice (Fig. 7). However, no significant differ-ence was observed in the percentage of CD4IFN-� andCD8IFN-� T cells between the immunized mice challenged
with either the antimony-sensitive strain AG83 or antimony-resis-tant strain GE1F8R.
Experimental and clinical VL have been reported to be accom-panied by the impairment of IL-12 generation (50) and inductionof IFN-� has been found to be involved in resistance to infectionin murine VL (51, 52), while IL-10 is reported to correlate withsusceptibility (53). Based on the important roles of these cyto-kines, we checked the functional activity of these cytokines in bothpUMVC3-UB-ORFF- and pUMVC3-ORFF- immunized and -in-fected mice. Spleen cells from saline-, blank vector-, pUMVC3-ORFF-, or pUMVC3-UB-ORFF DNA-immunized and from 8 wkafter infection were stimulated with rORFF Ag (5 �g/ml) for 48 hand supernatants were tested for cytokine levels (Fig. 8). The
FIGURE 7. Flow cytometric analysis of ORFF-specific IFN-�-producing CD4 and CD8 T cells in pUMVC3-ORFF- or pUMVC3-UB-ORFF DNA-immunized and -challenged mice. Splenocytes were isolated from pUMVC3-ORFF- or pUMVC3-UB-ORFF-immunized and AG83 (antimony-suscepti-ble)- or GE1F8R (antimony-resistant)-challenged mice and the expression of IFN-�-producing CD4 and CD8 T cells was studied in the presence ofrORFF (5 �g/ml) at 8 wk after challenge. Cells were blocked with brefeldin A followed by blocking with BD Biosciences Fc Block. The gated CD cellsare shown stained with allophycocyanin-labeled anti-IFN-�. Values in the quadrants represent the percentage of positive cells. The results are representativeof two individual experiments (n � 6/group). Data represent the mean of triplicate wells � SD.
7726 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

spleen supernatant of pUMVC3-ORFF-vaccinated and either theAG83- or GE1F8R-infected mice showed �4-fold ( p � 0.0001)more IL-12 production over the control groups. However, the cul-tured supernatants from splenocytes of mice challenged withAG83- and pUMVC3-UB-ORFF-immunized cells contained sig-nificantly higher levels of IL-12 (�7-fold; p � 0.0001) comparedwith saline- or blank vector-infected mice. GE1F8R- infected andpUMVC3-UB-ORFF-immunized mice showed �8-fold ( p �0.0001) more IL-12 production compared with the correspondingcontrol groups. The pUMVC3-ORFF-vaccinated and either theAG83- or GE1F8R-infected mice also showed �4-fold ( p �0.0001) more IFN-� production over the control groups. The miceinfected with AG83 or GE1F8R and immunized with pUMVC3-UB-ORFF showed an �7-fold ( p � 0.0001) increase in IFN-�levels over the saline- and blank vector-immunized control mice.Mice challenged with either AG83 or GE1F8R and immunizedwith UBQ-ORFF DNA showed an �1.5-fold ( p � 0.0001) in-crease of IFN-� and IL-12 levels over nonubiquitinated ORFFDNA-vaccinated mice. However, the level of expression of theTh1 suppressive cytokine IL-10 was �5-fold ( p � 0.0001) higherin both AG83- and GE1F8R-infected mice, respectively, than cor-responding infected and vaccinated groups of mice showing ex-treme down-regulation. The Th2 cytokine IL-4 in the AG83- orGE1F8R- infected and pUMVC3-UB-ORFF-vaccinated mice wasalso significantly down-regulated ( p � 0.005) when comparedwith the corresponding blank vector-immunized groups of mice.Thus, pUMVC3-UB-ORFF DNA vaccination resulted in a stron-ger protection in comparison to pUMVC3-ORFF DNA as illus-trated by enhanced induction of a Th1-mediated immune responseagainst both the antimony-susceptible and -resistant strains of L.donovani.
Measurement of NO and ROS in immunized mice challengedwith antimony-susceptible and -resistant strains of L. donovani
In the murine model, both IFN-� and TNF are implicated in themacrophage killing of intracellular L. donovani through the up-regulation of inducible NO synthase and the production of NO, amacrophage-derived oxidant. NO presence has been shown to bevery critical in determining the leishmanicidal activity of murinemacrophages (54) and has been reported to induce a Th1 response(55). On the other hand, IL-10 is reported to down-regulate NOproduction (53). ROS, including free radicals, oxygen ions, andperoxides, have also been implicated in cell damage like NO (56,57). ROS is known to contribute to the efficiency of parasite killing(57, 58). This led to the determination of nitrite and ROS levels invaccinated vs control mice 8 wk after infection. Spleen cells fromsaline-, blank vector-, pUMVC3-ORFF-, or pUMVC3-UB-ORFFDNA-immunized mice and from 8 wk after infection with eitherAG83 or GE1F8R of L. donovani were stimulated with rORFF Ag(5 �g/ml) for 48 h and supernatants were tested for NO and ROSlevels (Fig. 9, A and B).
We found that at 8 wk after infection, AG83-infected andpUMVC3-UB-ORFF DNA-vaccinated mice in the presence ofrORFF Ag showed �36 �M nitrite production and the GE1F8R-challenged mice showed �37 �M nitrite production (Fig. 9A).Both AG83- and GE1F8R-infected mice immunized withpUMVC3-UB-ORFF DNA showed an �12-fold increase in NOproduction ( p � 0.0001) when stimulated with 5 �g/ml rORFF Agover the control groups (Fig. 9A). On the other hand, thepUMVC3-ORFF-vaccinated and both AG83- and GE1F8R- in-fected mice showed an �8-fold ( p � 0.0001) increase in NOproduction over the control groups. In the presence of rORFF, the
FIGURE 8. Cytokine productionby spleen cells from pUMVC3-ORFF-or pUMVC3-UB-ORFF-immunizedinfected mice. BALB/c mice were im-munized with saline, vector, pUMVC3-ORFF, or pUMVC3-UB-ORFF thriceat 2-wk intervals. Two weeks followingthe final dose, one-half of the micewere challenged (i.v.) with 1 � 108
promastigotes of either antimony-sus-ceptible AG83 and the remaining withantimony-resistant GE1F8R L. dono-vani. Eight weeks after challenge infec-tion, spleens were collected as de-scribed in Materials and Methods.Cytokine levels of IFN-�, IL-12, IL-4,and IL-10 in the culture supernatantwere determined in response to rORFF(5 �g/ml) by splenocytes from UBQ-ORFF-immunized and -challengedmice and were compared with cor-responding saline- or blank vector-immunized challenged mice. Theresults are representative of two in-dividual experiments (n � 6/group).Each bar represents the means andSD of triplicate wells. ���, p �0.0001 and �, p � 0.005.
7727The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

nonvaccinated saline or blank vector control groups of miceshowed about �3 �M nitrite production when stimulated with 5�g/ml rORFF Ag. Mice immunized with UBQ-ORFF showed�1.5-fold enhanced levels ( p � 0.0001) of NO production overthose of nonubiquitinated ORFF DNA-vaccinated mice. Mice vac-cinated and challenged with either AG83 or GE1F8R showedhigher levels of NO compared with the saline- or blank vector-vaccinated and -challenged groups even in the absence of rORFFstimulation.
In the presence of rORFF, AG83- and GE1F8R-challenged miceimmunized with pUMVC3-UB-ORFF showed �6-fold ( p �0.0001) increased levels of ROS when compared with correspond-ing nonvaccinated saline or blank vector control groups of mice(Fig. 9B). The pUMVC3-ORFF-vaccinated and both AG83- andGE1F8R-infected mice showed �4-fold ( p � 0.0001) increasedlevels of ROS when compared with the nonvaccinated controlgroups. Mice immunized with UBQ-ORFF showed �1.4-fold en-hanced ( p � 0.0001) ROS production over nonubiquitinatedORFF DNA-vaccinated mice.
IgG1 and IgG2a levels in immunized mice challenged withantimony-susceptible and -resistant strains of L. donovani
To evaluate the humoral immune responses induced by pUMVC3-UB-ORFF vaccine, anti-ORFF IgG1 and IgG2a titers were mea-sured in infected and pUMVC3-UB-ORFF-immunized mice. Inthe murine model, IgG1 and IgG2a class switching is governedby the presence of IL-4 and IFN-�, respectively (6). Eight weeks afterthe infection, sera from pUMVC3-UB-ORFF DNA- and blankvector-immunized mice challenged with either AG83 or GE1F8Rand corresponding infected controls were analyzed for ORFF-spe-cific anti-IgG1 and IgG2a Ab titers by ELISA as described inMaterials and Methods. A 10-fold increased level of IgG2a in bothAG83- and GE1F8R-infected and pUMVC3-UB-ORFF-immu-nized mice was observed compared with the infected controlgroups (Fig. 10; p � 0.0001). Higher levels of IgG1 were observedin the infected and immunized mice over the infected controlgroups; however, the levels of IgG1 were very low compared withthe levels of IgG2a as shown in Fig. 10. The higher levels of IgG2aAbs in the pUMVC3-UB-ORFF- vaccinated mice show that theimmune responses are further skewed toward a IFN-�-mediated,
cell-mediated immune response. We also measured the IgG iso-types in the above groups using 100 ng of Leishmania SA/well. Anincrease in the IgG2a in both AG83- and GE1F8R-infected andpUMVC3-UB-ORFF-immunized mice was observed comparedwith the infected control groups (data not shown).
DiscussionVaccination strategies are based upon our current understanding ofthe characteristics of an effective anti-Leishmania immune re-sponse, as they have been determined from human and murinestudies (59). Current data suggest that protection to Leishmaniainfection requires the effective activation of several cell popula-tions, including DC, Ag-specific CD4 and CD8 T cells, andmacrophages (8, 51, 60). Resistance to leishmaniasis has been as-sociated with a predominant IL-12 and IFN-� production from theAg-specific CD4 Th lymphocyte population termed as Th1 im-mune response (61, 62). These cells along with CD8 T cytotoxiclymphocytes are then effective in promoting macrophage activa-tion, and the intracellular Leishmania are killed in a NO-dependentmanner (63–65).
Previous studies in our laboratory involving the ORFF genefrom the LD1 locus have shown DNA vaccination with ORFFinduced protective immunity in BALB/c mice (25). In the presentstudy, we evaluated the efficacy of ORFF DNA vaccine-expressingORFF protein fused at the N terminus with ubiquitin in inducing aspecific cellular immune response in an experimental VL murinemodel. From the understanding of several previous studies, adirect correlation between ubiquitin-mediated rapid processingof Ags and induction of a strong cellular immune response hasbeen postulated (37). In the present study, we exploited theN-end rule (44) and modified the N-terminal amino acid to re-duce the half-life of our Ag so that it can undergo rapid deg-radation by the proteasome machinery. Higher rates of intra-cellular Ag turnover should increase the number and variety offragments and peptides available for MHC class I and class IIbinding that may result in an increase of the cell-mediated im-mune response to the expressed Ag (36, 39, 40).
Our results are clearly in accordance with the above-mentionedconcept. We found that vaccination with ubiquitinated ORFF re-sulted in an enhanced cell-mediated immune response compared
FIGURE 9. NO and ROS production in pUMVC3-ORFF- or pUMVC3-UB-ORFF-immunized and -challenged mice. BALB/c mice were immunizedwith saline, vector, pUMVC3-ORFF, or pUMVC3-UB-ORFF thrice at 2-wk intervals. Two weeks following the final dose, one-half of the mice werechallenged (i.v.) with 1 � 108 promastigotes of antimony-susceptible AG83 and the remaining with antimony-resistant GE1F8R L. donovani. Eight weeksafter the challenge infection, spleens were collected as described in Materials and Methods. A, The culture supernatant of splenocytes was used to evaluateNO generation in the presence and absence of rORFF (5 �g/ml) from pUMVC3-UB-ORFF-immunized and -challenged mice and was compared withcorresponding saline- or blank vector-immunized challenged mice. B, ROS generation was measured by H2DCFDA probe staining of the splenocytes fromdifferent groups of mice in the absence and presence of 5 �g of rORFF. Each bar is shown with its respective SD. �, p � 0.005 and ���, p � 0.0001. Theresults are representative of two individual experiments (n � 6/group).
7728 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

with a nonubiquitinated ORFF-based vaccine. Our vaccine strat-egy was found to trigger both the humoral and the cell-mediatedimmune responses. We checked for the splenocyte proliferation bymeasuring the [3H]thymidine incorporation into the spleen cellsstimulated with and without rORFF. We found an enhanced pro-liferation in the mice vaccinated with ubiquitinated ORFF com-pared with the mice vaccinated with nonubiquitinated ORFF. Thisshows that the ubiquitin conjugation indeed increased the cell-mediated response. It has been proved that the resistance againstleishmaniasis is conferred by Th1 cells while susceptibility is con-ferred by Th2 cells. Therefore, we went ahead to do a cytokineprofiling and check for the expression of IL-12, IFN-� IL-10, andIL-4 from the spleens of mice immunized with our vaccine. After2 wk of immunization, we found an enhanced level of expressionin IL-12 and IFN-� over that of the control groups. Besides andimportantly enough, the mice that were given ubiquitinated ORFFshowed higher levels of both IL-12 and IFN-� compared withthose that were given nonubiquitinated ORFF. This was furthersupported by enhanced levels of IFN-�-producing CD4 andCD8 T cells. However. there was no increase in the levels ofIL-10 and IL-4 both in ubiquitinated and nonubiquitinated vaccinegroups over those of the control groups of mice. This gives a clearand positive indication that the immune responses are directed to-ward a Th1 type rather than Th2 type.
We also checked for the Ab levels in the blood serum of vac-cinated and nonvaccinated mice. We found very high levels ofIgG2a in the mice immunized with UBQ-ORFF compared with themice immunized with ORFF (data not shown). IgG2a has beenfound to skew the immune responses further toward an IFN-�-mediated Th1 type of immune response.
Protective efficacy of pUMVC3-UB-ORFF DNA was evaluatedagainst both the antimony-susceptible and -resistant strains of L.
donovani. This was relevant because of the failure of conventionalchemotherapeutic agents due to the emergence of resistance topentavalent antimonials (5, 66). Mice were immunized withpUMVC3-UB-ORFF DNA vaccine and challenged with antimo-ny-sensitive AG83 or -resistant GE1F8R promastigotes of L. do-novani and checked for the parasite burden in spleen and liver ofinfected mice. Four weeks after the infection, there was �70%reduction in parasite load, both in spleen and liver. The protectiveefficacy went up to 85–90% by the end of 8 wk in spleens ofinfected mice. Ubiquitination of ORFF resulted in enhanced re-duction of parasite load in mice compared with ORFF DNA-im-munized and -infected mice. Further to check the long-lasting ef-fect of the immunity conferred by the ubiquitinated constructpUMVC3-UB-ORFF, we again checked for various immunologi-cal parameters, such as IFN-�-producing CD4 and CD8 T cells,splenocyte proliferation, and cytokine profiling after 8 wk of im-munization and challenge. It was evident from the present studythat all of the pUMVC3-UB-ORFF DNA-immunized mice chal-lenged either with the AG83 or GE1F8R strain showed a specificand active T cell response characteristic of the induction of theTh1-type immune response. However, in infected and nonimmu-nized and blank vector-treated mice, the response was impaired asis typical in VL patients showing marked T cell anergy in responseto Leishmania-specific Ags (67). An increase in IFN-�-producingCD4 T cells in UBQ-ORFF DNA-vaccinated and AG83- orGE1F8R-infected mice in the present study is again characteristicof induction of the Th1-type immune response. These data werefurther supported by the enhanced levels of cytokines IFN-� andIL-12, which are known to be associated with protection againstleishmaniasis (50, 51) in AG83- and GE1F8R-infected andpUMVC3-UB-ORF-vaccinated mice. Enhanced levels of thesetwo cytokines may have a synergistic role in killing the parasite. It
FIGURE 10. ORFF-specific Ab isotype profile in pUMVC3-UB-ORFF-immunized and -challenged mice. BALB/c mice were immunized thrice at 2-wkintervals with saline, vector, or pUMVC3-UB-ORFF. Two weeks following the final dose, one-half of the mice were challenged with 1 � 108 AG83promastigotes (A) and the remaining with GE1F8R promastigotes (B) of L. donovani. Mice were then bled 8 wk following the challenge and sera from themice in each group were taken and analyzed by ELISA for the presence of IgG1 and IgG2a. All data here represent the mean and SD from individual mice.The results are representative of two individual experiments (n � 6/group).
7729The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

has been reported recently that IL-10 plays a suppressive role inparasite infection and dictates the susceptibility of L. donovaniinfection and is required for high parasite persistence in bothC57BL/6 and susceptible BALB/c mice (68, 69). Accordingly,the present data support this because we observed low levels ofIL-10 in pUMVC3-UB-ORFF DNA-immunized and AG83- andGE1F8R-challenged mice when compared with the saline- orblank vector-infected mice. Another Th2 or inhibitory cytokine,IL-4, was low in contrast to the robust Th1-type immune re-sponses observed with UBQ-ORFF-vaccinated and -infectedmice. In the murine model of cutaneous leishmaniasis thathealed the primary lesion, the presence of CD8 T cells hasbeen reported. The present data also showed the presence ofhigher levels of IFN-�-producing CD8 T cells in mice immu-nized with UBQ-ORFF and infected with either AG83 orGE1F8R when compared with ORFF-immunized and -infectedmice. In a viral system where CTL induction is a correlate ofprotective immunity, DNA vaccines expressing Ags cotrans-lated with ubiquitin have been used to increase the CD8-medi-ated CTL response (35, 37, 39, 40).
The Leishmania parasite has the remarkable ability to residein the host macrophage and for its elimination from the macrophage;the mechanism involves the need for macrophage activation (70).The activated macrophage then releases various toxic metabolitesof oxygen, which may include superoxide anion (O�
2), hydrogenperoxide (H2O2), and NO, which kill the parasite (63). Accord-ingly, increased production of NO and ROS by the splenocyteswas observed in mice immunized with pUMVC3-UB-ORFFand infected with AG83 or GE1F8R L. donovani promastigotescompared with the saline- and blank vector-immunized and-infected mice.
The present data were further supported by the presence ofsignificantly higher levels of UBQ-ORFF-specific IgG2a Ab ti-ters over IgG1 Ab titers in pUMVC3-UB-ORFF-vaccinated and-infected mice in comparison to infected and blankvector-immunized mice.
Our studies suggest that DNA vaccines encoding LeishmaniaORFF Ag cotranslated with ubiquitin modulate the immune re-sponse. The ubiquitinated construct UBQ-ORFF induces a cell-mediated response rather than a humoral response, resulting in80–90% control of L. donovani growth in vivo compared withcontrol mice. The protective efficacy of pUMVC3-UB-ORFF Agcorrelated with significant up-regulation of IFN-� and IL-12 alongwith down-regulation of IL-10, which in turn might lead to theobserved NO and ROS generation. We also report a promising roleof the ubiquitin-conjugated leishmanial gene ORFF (pUMVC3-UB-ORFF) as a partially protective vaccine against both the anti-mony-susceptible and -resistant strains of L. donovani. However,there was no difference in the modulation of immune response andprotective efficacy of the pUMVC3-UB-ORFF vaccine in antimo-ny-susceptible AG83 and -resistant GE1F8R-infected mice. A sim-ilar observation has been reported earlier with KMP-11 DNA vac-cination in antimony-sensitive and -resistant infected hamsters (6).This apparent homogeneity in the mechanism and protection wasreported to be the due to the possibility of the limited extent ofMHC polymorphism (71).
In view of the increasing reports of drug resistance in clinicalisolates of L. donovani, our results in the murine L. donovanimodel show a promising role of the ubiquitin-conjugated leishma-nial gene ORFF as a DNA vaccine that can be exploited to createa protective vaccine against both drug-susceptible and -resistantstrains of L. donovani.
AcknowledgmentsWe acknowledge the technical help provided by Bhavana Iyer. We thankDr. Uday Kumar Ranga (Jawaharlal Nehru Center for Advanced ScientificResearch) for providing us the ubiquitin-conjugated-pUMVC3 vector thatwas used throughout our studies.
DisclosuresThe authors have no financial conflict of interest.
References1. Murray, H. W., J. D. Berman, C. R. Davies, and N. G. Saravia. 2005. Advances
in leishmaniasis. Lancet 366: 1561–1577.2. 1998. Advances in battle against leishmaniasis. Trop. Disease Res. News 57: 2.3. 1997. Special Programme for Research and Training in Tropical Diseases: Leish-
maniasis. In World Health Report: Thirteenth Programme Report. World HealthOrganizations Publications, Geneva.
4. Murray, H. W. 2001. Clinical and experimental advances in treatment of visceralleishmaniasis. Antimicrob. Agents Chemother. 45: 2185–2197.
5. Sundar, S., D. K. More, M. K. Singh, V. P. Singh, S. Sharma, A. Makharia,P. C. Kumar, and H. W. Murray. 2000. Failure of pentavalent antimony in vis-ceral leishmaniasis in India: report from the center of the Indian epidemic. Clin.Infect. Dis. 31: 1104–1107.
6. Basu, R., S. Bhaumik, J. M. Basu, K. Naskar, T. De, and S. Roy. 2005. Kineto-plastid membrane protein-11 DNA vaccination induces complete protectionagainst both pentavalent antimonial-sensitive and -resistant strains of Leishmaniadonovani that correlates with inducible nitric oxide synthase activity and IL-4generation: evidence for mixed Th1- and Th2-like responses in visceral leish-maniasis. J. Immunol. 174: 7160–7171.
7. Handman, E. 2001. Leishmaniasis: current status of vaccine development. Clin.Microbiol. Rev. 14: 229–243.
8. Coler, R. N., Y. Goto, L. Bogatzki, V. Raman, and S. G. Reed. 2007. Leish-111f,a recombinant polyprotein vaccine that protects against visceral leishmaniasis byelicitation of CD4 T cells. Infect. Immun. 75: 4648–4654.
9. Campos-Neto, A., R. Porrozzi, K. Greeson, R. N. Coler, J. R. Webb, Y. A. Seiky,S. G. Reed, and G. Grimaldi, Jr. 2001. Protection against cutaneous leishmaniasisinduced by recombinant antigens in murine and nonhuman primate models of thehuman disease. Infect. Immun. 69: 4103–4108.
10. Rafati, S., A. A. Baba, M. Bakhshayesh, and M. Vafa. 2000. Vaccination ofBALB/c mice with Leishmania major amastigote-specific cysteine proteinase.Clin. Exp. Immunol. 120: 134–138.
11. Abdelhak, S., H. Louzir, J. Timm, L. Blel, Z. Benlasfar, M. Lagranderie,M. Gheorghiu, K. Dellagi, and B. Gicquel. 1995. Recombinant BCG expressingthe Leishmania surface antigen Gp63 induces protective immunity against Leish-mania major infection in BALB/c mice. Microbiology 141: 1585–1592.
12. Cox, F. E. 1997. Designer vaccines for parasitic diseases. Int. J. Parasitol. 27:1147–1157.
13. Gurunathan, S., D. L. Sacks, D. R. Brown, S. L. Reiner, H. Charest,N. Glaichenhaus, and R. A. Seder. 1997. Vaccination with DNA encoding theimmunodominant LACK parasite antigen confers protective immunity to miceinfected with Leishmania major. J. Exp. Med. 186: 1137–1147.
14. Khamesipour, A., S. Rafati, N. Davoudi, F. Maboudi, and F. Modabber. 2006.Leishmaniasis vaccine candidates for development: a global overview. IndianJ. Med. Res. 123: 423–438.
15. Awasthi, A., R. K. Mathur, and B. Saha. 2004. Immune response to Leishmaniainfection. Indian J. Med. Res. 119: 238–258.
16. Miralles, G. D., M. Y. Stoeckle, D. F. McDermott, F. D. Finkelman, andH. W. Murray. 1994. Th1 and Th2 cell-associated cytokines in experimentalvisceral leishmaniasis. Infect. Immun. 62: 1058–1063.
17. Kemp, M., J. A. Kurtzhals, K. Bendtzen, L. K. Poulsen, M. B. Hansen,D. K. Koech, A. Kharazmi, and T. G. Theander. 1993. Leishmania donovani-reactive Th1- and Th2-like T-cell clones from individuals who have recoveredfrom visceral leishmaniasis. Infect. Immun. 61: 1069–1073.
18. Campos-Neto, A. 2005. What about Th1/Th2 in cutaneous leishmaniasis vaccinediscovery? Braz. J. Med. Biol. Res. 38: 979–984.
19. Kaye, P. M., A. J. Curry, and J. M. Blackwell. 1991. Differential production ofTh1- and Th2-derived cytokines does not determine the genetically controlled orvaccine-induced rate of cure in murine visceral leishmaniasis. J. Immunol. 146:2763–2770.
20. Tripp, C. A., W. A. Wisdom, P. J. Myler, and K. D. Stuart. 1992. A multicopy,extrachromosomal DNA in Leishmania infantum contains two inverted repeats ofthe 27.5-kilobase LD1 sequence and encodes numerous transcripts. Mol. Bio-chem. Parasitol. 55: 39–50.
21. Sunkin, S. M., P. McDonagh, M. L. Cunningham, S. M. Beverley, K. Stuart, andP. J. Myler. 2001. Conservation of the LD1 region in Leishmania includes DNAimplicated in LD1 amplification. Mol. Biochem. Parasitol. 113: 315–321.
22. Ghosh, A., R. Madhubala, P. J. Myler, and K. D. Stuart. 1999. Leishmania do-novani: characterization and expression of ORFF, a gene amplified from the LDIlocus. Exp. Parasitol. 93: 225–230.
23. Raj, V. S., A. Ghosh, V. S. Dole, R. Madhubala, P. J. Myler, and K. D. Stuart.1999. Serodiagnosis of leishmaniasis with recombinant ORFF antigen.Am. J. Trop. Med. Hyg. 61: 482–487.
24. Dole, V. S., V. S. Raj, A. Ghosh, R. Madhubala, P. J. Myler, and K. D. Stuart.2000. Immunization with recombinant LD1 antigens protects against experimen-tal leishmaniasis. Vaccine 19: 423–430.
7730 UBIQUITIN-CONJUGATED ORFF DNA VACCINATION IN VL
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

25. Sukumaran, B., P. Tewary, S. Saxena, and R. Madhubala. 2003. Vaccination withDNA encoding ORFF antigen confers protective immunity in mice infected withLeishmania donovani. Vaccine 21: 1292–1299.
26. Tewary, P., J. Pandya, J. Mehta, B. Sukumaran, and R. Madhubala. 2004. Vac-cination with Leishmania soluble antigen and immunostimulatory oligode-oxynucleotides induces specific immunity and protection against Leishmania do-novani infection. FEMS Immunol. Med. Microbiol. 42: 241–248.
27. Tewary, P., B. Sukumaran, S. Saxena, and R. Madhubala. 2004. Immunostimu-latory oligodeoxynucleotides are potent enhancers of protective immunity in miceimmunized with recombinant ORFF leishmanial antigen. Vaccine 22:3053–3060.
28. Tewary, P., M. Jain, M. H. Sahani, S. Saxena, and R. Madhubala. 2005. A het-erologous prime-boost vaccination regimen using ORFF DNA and recombinantORFF protein confers protective immunity against experimental visceral leish-maniasis. J. Infect. Dis. 191: 2130–2137.
29. Tewary, P., S. Saxena, and R. Madhubala. 2006. Co-administration of IL-12DNA with rORFF antigen confers long-term protective immunity against exper-imental visceral leishmaniasis. Vaccine 24: 2409–2416.
30. Billaut-Mulot, O., T. Idziorek, M. Loyens, A. Capron, and G. M. Bahr. 2001.Modulation of cellular and humoral immune responses to a multiepitopic HIV-1DNA vaccine by interleukin-18 DNA immunization/viral protein boost. Vaccine19: 2803–2811.
31. Walker, P. S., T. Scharton-Kersten, A. M. Krieg, L. Love-Homan, E. D. Rowton,M. C. Udey, and J. C. Vogel. 1999. Immunostimulatory oligodeoxynucleotidespromote protective immunity and provide systemic therapy for leishmaniasis viaIL-12- and IFN-�-dependent mechanisms. Proc. Natl. Acad. Sci. USA 96:6970–6975.
32. Cafaro, A., F. Titti, C. Fracasso, M. T. Maggiorella, S. Baroncelli, A. Caputo,D. Goletti, A. Borsetti, M. Pace, E. Fanales-Belasio, et al. 2001. Vaccination withDNA containing TAT coding sequences and unmethylated CpG motifs protectscynomolgus monkeys upon infection with simian/human immunodeficiency virus(SHIV89.6P). Vaccine 19: 2862–2877.
33. Li, Z., A. Howard, C. Kelley, G. Delogu, F. Collins, and S. Morris. 1999. Im-munogenicity of DNA vaccines expressing tuberculosis proteins fused to tissueplasminogen activator signal sequences. Infect. Immun. 67: 4780–4786.
34. Roos-Mattjus, P., and L. Sistonen. 2004. The ubiquitin-proteasome pathway.Ann. Med. 36: 285–295.
35. Delogu, G., A. Howard, F. M. Collins, and S. L. Morris. 2000. DNA vaccinationagainst tuberculosis: expression of a ubiquitin-conjugated tuberculosis proteinenhances antimycobacterial immunity. Infect. Immun. 68: 3097–3102.
36. Michalek, M. T., E. P. Grant, C. Gramm, A. L. Goldberg, and K. L. Rock. 1993.A role for the ubiquitin-dependent proteolytic pathway in MHC class I-restrictedantigen presentation. Nature 363: 552–554.
37. Ramakrishna, L., K. K. Anand, M. Mahalingam, K. M. Mohankumar, S. Ramani,N. B. Siddappa, and U. Ranga. 2004. Codon optimization and ubiquitin conju-gation of human immunodeficiency virus-1 Tat lead to enhanced cell-mediatedimmune responses. Vaccine 22: 2586–2598.
38. Grant, E. P., M. T. Michalek, A. L. Goldberg, and K. L. Rock. 1995. Rate ofantigen degradation by the ubiquitin-proteasome pathway influences MHC classI presentation. J. Immunol. 155: 3750–3758.
39. Wu, Y., and T. J. Kipps. 1997. Deoxyribonucleic acid vaccines encoding antigenswith rapid proteasome-dependent degradation are highly efficient inducers of cy-tolytic T lymphocytes. J. Immunol. 159: 6037–6043.
40. Tobery, T., and R. F. Siliciano. 1999. Cutting edge: induction of enhanced CTL-dependent protective immunity in vivo by N-end rule targeting of a model tumorantigen. J. Immunol. 162: 639–642.
41. Mukherjee, A., P. K. Padmanabhan, S. Singh, G. Roy, I. Girard, M. Chatterjee,M. Ouellette, and R. Madhubala. 2007. Role of ABC transporter MRPA, �-glu-tamylcysteine synthetase and ornithine decarboxylase in natural antimony-resis-tant isolates of Leishmania donovani. J. Antimicrob. Chemother. 59: 204–211.
42. Singh, S., A. Mukherjee, A. R. Khomutov, L. Persson, O. Heby, M. Chatterjee,and R. Madhubala. 2007. Antileishmanial effect of 3-aminooxy-1-aminopropaneis due to polyamine depletion. Antimicrob. Agents Chemother. 51: 528–534.
43. Hart, D. T., K. Vickerman, and G. H. Coombs. 1981. A quick, simple method forpurifying Leishmania mexicana amastigotes in large numbers. Parasitology 82:345–355.
44. Varshavsky, A. 1996. The N-end rule: functions, mysteries, uses. Proc. Natl.Acad. Sci. USA 93: 12142–12149.
45. Murray, P. D. 1990. Maintaining protein stability. In Methods in Enzymology.M. Drutscher, ed. Academic, London, pp. 83–92.
46. Lehmann, J., K. H. Enssle, I. Lehmann, A. Emmendorfer, andM. L. Lohmann-Matthes. 2000. The capacity to produce IFN-� rather than the pres-ence of interleukin-4 determines the resistance and the degree of susceptibility toLeishmania donovani infection in mice. J. Interferon Cytokine Res. 20: 63–77.
47. Scott, P., P. Natovitz, R. L. Coffman, E. Pearce, and A. Sher. 1988. Immuno-regulation of cutaneous leishmaniasis. T cell lines that transfer protective immu-nity or exacerbation belong to different T helper subsets and respond to distinctparasite antigens. J. Exp. Med. 168: 1675–1684.
48. Haldar, J. P., S. Ghose, K. C. Saha, and A. C. Ghose. 1983. Cell-mediated im-mune response in Indian kala-azar and post-kala-azar dermal leishmaniasis. In-fect. Immun. 42: 702–707.
49. Gifawesen, C., and J. P. Farrell. 1989. Comparison of T-cell responses in self-limiting versus progressive visceral Leishmania donovani infections in goldenhamsters. Infect. Immun. 57: 3091–3096.
50. Murray, H. W., C. Montelibano, R. Peterson, and J. P. Sypek. 2000. Interleu-kin-12 regulates the response to chemotherapy in experimental visceral leish-maniasis. J. Infect. Dis. 182: 1497–1502.
51. Murray, H. W., J. J. Stern, K. Welte, B. Y. Rubin, S. M. Carriero, andC. F. Nathan. 1987. Experimental visceral leishmaniasis: production of interleu-kin 2 and interferon-�, tissue immune reaction, and response to treatment withinterleukin 2 and interferon-�. J. Immunol. 138: 2290–2297.
52. Carvalho, E. M., R. Badaro, S. G. Reed, T. C. Jones, and W. D. Johnson, Jr. 1985.Absence of � interferon and interleukin 2 production during active visceral leish-maniasis. J. Clin. Invest. 76: 2066–2069.
53. Murphy, M. L., U. Wille, E. N. Villegas, C. A. Hunter, and J. P. Farrell. 2001.IL-10 mediates susceptibility to Leishmania donovani infection. Eur. J. Immunol.31: 2848–2856.
54. Liew, F. Y., S. Millott, C. Parkinson, R. M. Palmer, and S. Moncada. 1990.Macrophage killing of Leishmania parasite in vivo is mediated by nitric oxidefrom L-arginine. J. Immunol. 144: 4794–4797.
55. Niedbala, W., X. Q. Wei, D. Piedrafita, D. Xu, and F. Y. Liew. 1999. Effects ofnitric oxide on the induction and differentiation of Th1 cells. Eur. J. Immunol. 29:2498–2505.
56. Green, S. J., R. M. Crawford, J. T. Hockmeyer, M. S. Meltzer, and C. A. Nacy.1990. Leishmania major amastigotes initiate the L-arginine-dependent killingmechanism in IFN-�-stimulated macrophages by induction of tumor necrosisfactor-�. J. Immunol. 145: 4290–4297.
57. Shiloh, M. U., J. D. MacMicking, S. Nicholson, J. E. Brause, S. Potter,M. Marino, F. Fang, M. Dinauer, and C. Nathan. 1999. Phenotype of mice andmacrophages deficient in both phagocyte oxidase and inducible nitric oxide syn-thase. Immunity 10: 29–38.
58. Fang, F. C. 1997. Perspectives series: host/pathogen interactions: mechanisms ofnitric oxide-related antimicrobial activity. J. Clin. Invest. 99: 2818–2825.
59. Liew, F. Y., and C. A. O’Donnell. 1993. Immunology of leishmaniasis. Adv.Parasitol. 32: 161–259.
60. Ghosh, M., C. Pal, M. Ray, S. Maitra, L. Mandal, and S. Bandyopadhyay. 2003.Dendritic cell-based immunotherapy combined with antimony-based chemother-apy cures established murine visceral leishmaniasis. J. Immunol. 170:5625–5629.
61. Macatonia, S. E., C. S. Hsieh, K. M. Murphy, and A. O’Garra. 1993. Dendriticcells and macrophages are required for Th1 development of CD4 T cells from�� TCR transgenic mice: IL-12 substitution for macrophages to stimulate IFN-�production is IFN-�-dependent. Int. Immunol. 5: 1119–1128.
62. Seder, R. A., R. Gazzinelli, A. Sher, and W. E. Paul. 1993. Interleukin 12 actsdirectly on CD4 T cells to enhance priming for interferon � production anddiminishes interleukin 4 inhibition of such priming. Proc. Natl. Acad. Sci. USA90: 10188–10192.
63. Assreuy, J., F. Q. Cunha, M. Epperlein, A. Noronha-Dutra, C. A. O’Donnell,F. Y. Liew, and S. Moncada. 1994. Production of nitric oxide and superoxide byactivated macrophages and killing of Leishmania major. Eur. J. Immunol. 24:672–676.
64. Stefani, M. M., I. Muller, and J. A. Louis. 1994. Leishmania major-specificCD8 T cells are inducers and targets of nitric oxide produced by parasitizedmacrophages. Eur. J. Immunol. 24: 746–752.
65. Muller, I., P. Kropf, R. J. Etges, and J. A. Louis. 1993. � Interferon response insecondary Leishmania major infection: role of CD8 T cells. Infect. Immun. 61:3730–3738.
66. Perez-Victoria, J. M., F. J. Perez-Victoria, A. Parodi-Talice, I. A. Jimenez,A. G. Ravelo, S. Castanys, and F. Gamarro. 2001. Alkyl-lysophospholipid resis-tance in multidrug-resistant Leishmania tropica and chemosensitization by anovel P-glycoprotein-like transporter modulator. Antimicrob. Agents Chemother.45: 2468–2474.
67. Engwerda, C. R., and P. M. Kaye. 2000. Organ-specific immune responses as-sociated with infectious disease. Immunol. Today 21: 73–78.
68. Belkaid, Y., K. F. Hoffmann, S. Mendez, S. Kamhawi, M. C. Udey, T. A. Wynn,and D. L. Sacks. 2001. The role of interleukin (IL)-10 in the persistence ofLeishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 194: 1497–1506.
69. Stober, C. B., U. G. Lange, M. T. Roberts, A. Alcami, and J. M. Blackwell. 2005.IL-10 from regulatory T cells determines vaccine efficacy in murine Leishmaniamajor infection. J. Immunol. 175: 2517–2524.
70. Reiner, S. L., and R. M. Locksley. 1995. The regulation of immunity to Leish-mania major. Annu. Rev. Immunol. 13: 151–177.
71. McGuire, K. L., W. R. Duncan, and P. W. Tucker. 1985. Syrian hamster DNAshows limited polymorphism at class I-like loci. Immunogenetics 22: 257–268.
7731The Journal of Immunology
by guest on February 11, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
![Ubiquitin and Ubiquitin-like Modifications in Viral ...1].pdf · Ubiquitin and Ubiquitin-like Modifications in Viral Infection and Immunity Abstracts of papers presented at the AUGUST](https://img.dokumen.tips/doc/110x75/5e2d68ba2a69b505b71e58fa/ubiquitin-and-ubiquitin-like-modifications-in-viral-1pdf-ubiquitin-and-ubiquitin-like.jpg)




























