Embed Size (px)
Citation preview

1
TRAFICO LINFOCITARIO
• Numerosas puertas de ingreso del antígeno a nuestro cuerpo
Piel (2 m2)
Mucosas:
Tracto urogenital; pulmón ; intestino; (450 m2)
Torrente sanguíneo
• Número reducido de linfocitos con receptores específicos
La recirculación linfocitaria incrementa la probabilidad de encuentro de linfocitos
con su antígeno específico

2
Probabilidad de encuentro entre un linfocito y su Agsería muy baja de no existir rutas de tráfico definidas
# de linfocitos en un individuo no inmunizado: 25-100 millones; específicos
contra un Ag unos pocos miles
Antígenos son transportados desde su sitio de ingreso hasta los órganos linfáticos secundarios
Los linfocitos circulan a través de estos órganos
Concentración espacial de antígenos y linfocitos
3
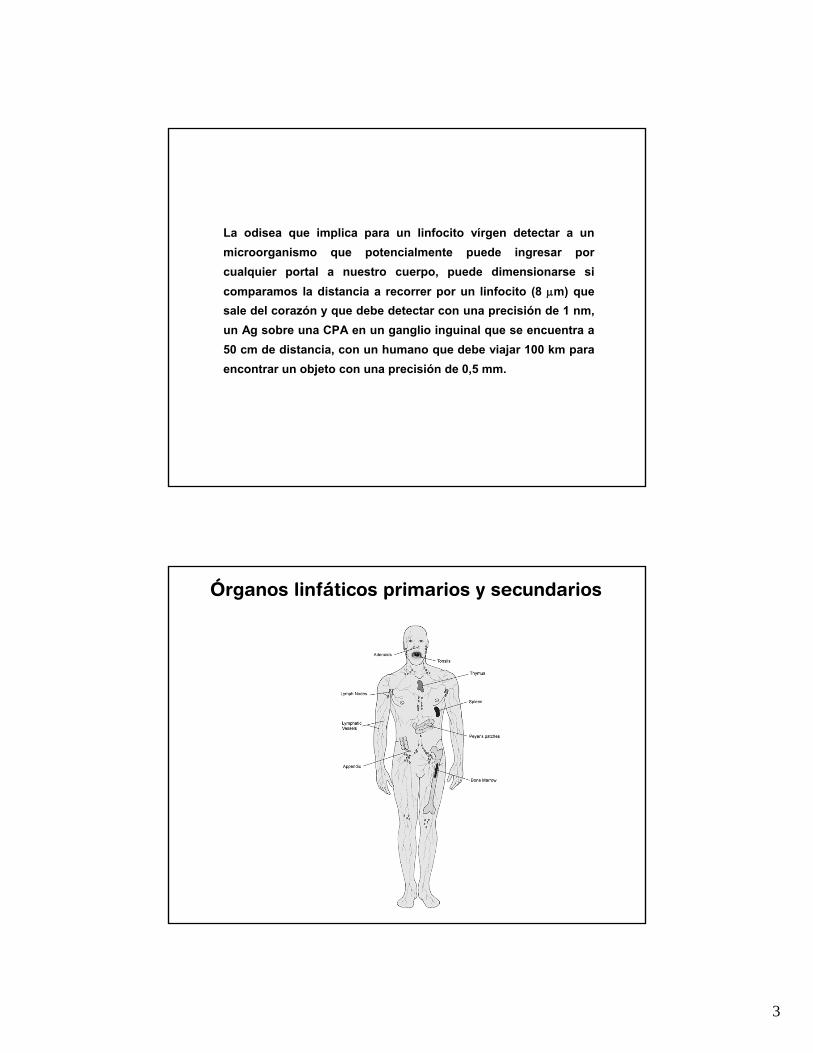
La odisea que implica para un linfocito vírgen detectar a un microorganismo que potencialmente puede ingresar por cualquier portal a nuestro cuerpo, puede dimensionarse si comparamos la distancia a recorrer por un linfocito (8 μm) que sale del corazón y que debe detectar con una precisión de 1 nm, un Ag sobre una CPA en un ganglio inguinal que se encuentra a 50 cm de distancia, con un humano que debe viajar 100 km para encontrar un objeto con una precisión de 0,5 mm.
Órganos linfáticos primarios y secundarios

4
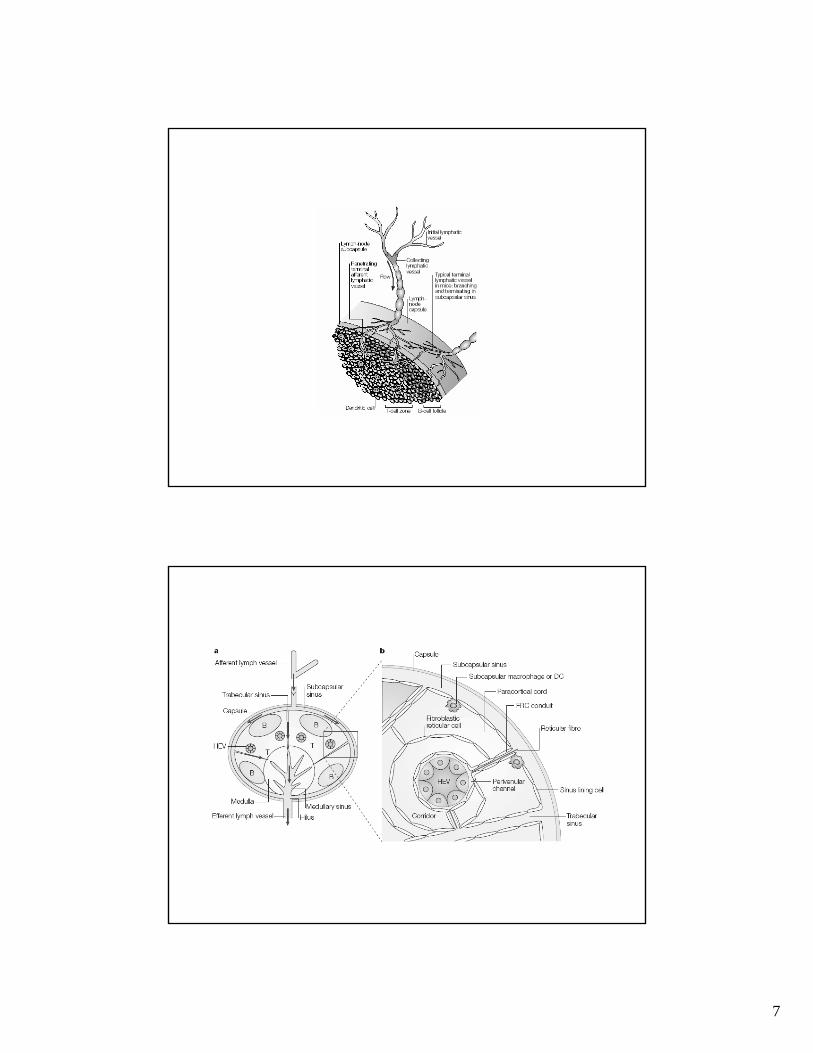
Estructura de un ganglio linfático

5
Cómo llegan los antígenos desde su puerta de ingreso a los órganos linfáticos secundarios?
transportados por CDsvía los vasos linfáticos aferentes
en forma directa por drenaje linfático
Transporte de antígenos por CDs

6
E-cadherina
CCR7 CCL19/CCL21
7

8
Localización de las CDs en los ganglios linfáticos
Ags con PM<70 kDa circulan a través de las fibras reticulares y pueden acceder a las HEVs
Las CD en los ganglios linfáticos se encuentran asociadas a las fibras reticulares

9
Linfocitos B
FRC
Linfocitos T
Localización de las CDs en los ganglios linfáticos
Ingreso de antígenos a los ganglios linfáticos vía drenaje linfático

10
Qué determina la correcta migración de las células del sistema inmune a los tejidos donde
deben desempeñar sus funciones?
Patrón de expresión de moléculas de adhesión
Patrón de expresión de receptores de quimiocinasy la distribución de dichas quimiocinas
FAMILIAS DE MOLECULAS DE ADHESION
SELECTINAS
SIALOMUCINAS
INTEGRINAS
MOLECULAS PERTENECIENTES A LA SUPERFLIA DE Ig
CADHERINAS

11
SELECTINAS LIGANDOS DE SELECTINAS
• E- y P- selectina unen ligandos que contienen fucosa, galactosa, N-acetilglucosamina y ácido siálico• Las glucosiltransferasas son esenciales para generar ligandos de selectinas (fucosiltransferasas, GlcNac-, Gal- y sialiltransferas). Deficiencia en una fucosiltransferasa ocasiona LAD II, con infecciones bacterianas recurrentes en piel y mucosas• Los ligandos de la L-selectina además están sulfatados por lo que su síntesis requiere la acciónde glucosilsulfotransferasas (GSTs)
12
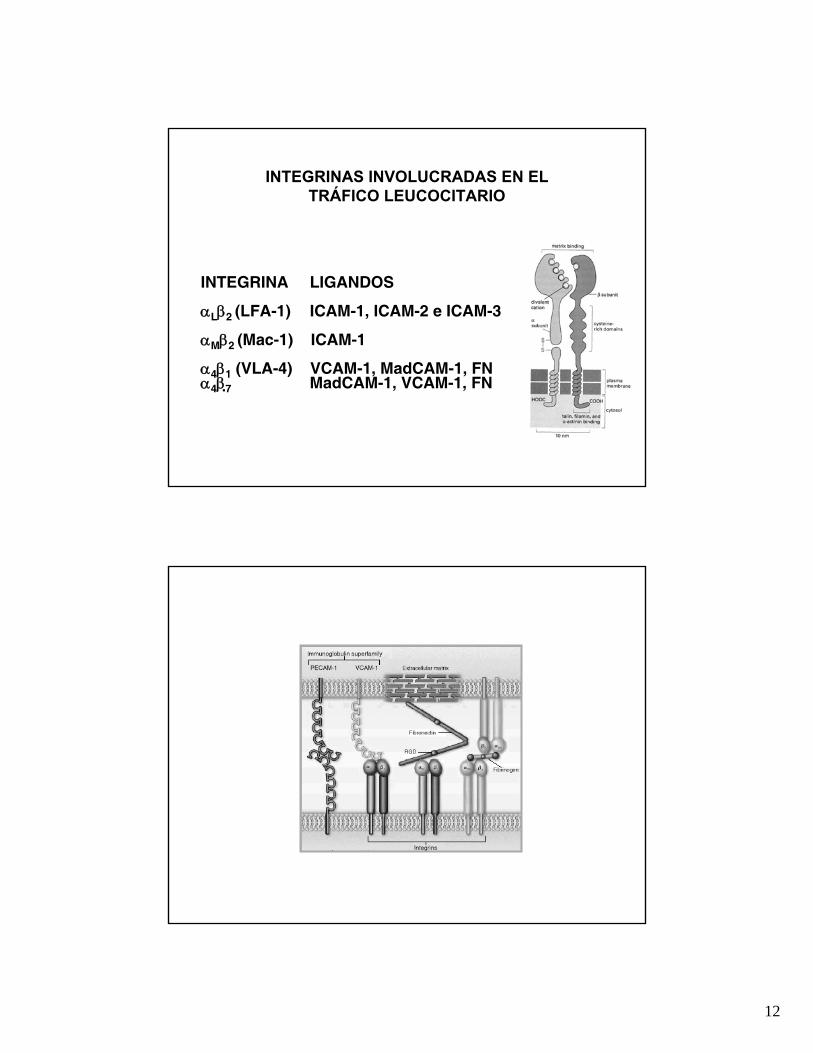
INTEGRINAS INVOLUCRADAS EN EL TRÁFICO LEUCOCITARIO
INTEGRINA LIGANDOS
αLβ2 (LFA-1) ICAM-1, ICAM-2 e ICAM-3
αMβ2 (Mac-1) ICAM-1
α4β1 (VLA-4) VCAM-1, MadCAM-1, FNα4β7 MadCAM-1, VCAM-1, FN

13
CADHERINAS
DEFICIENCIAS DE ADHESIÓN LEUCOCITARIA
LAD-1: defectos en la cadena b2 de las integrinas. defectiva migración de neutrófilos a piel inflamada y peritoneo. Neutrofilia. Infecciones bacterianas recurrentes.
LAD-2: defectos en la fucosiltransferasa y en consecuencia en la síntesis de ligandos de selectinas. Rolling defectivo.

14
QUIMIOCINAS

15
Cuál es el rol que las moléculas de adhesión y las quimiocinas cumplen en la migración leucocitaria?
E-cadherina
CCR7 CCL19/CCL21
Rol de las moléculas de adhesión y las quimiocinas en la migración de las CDs a los ganglios linfáticos

16
Rol de las moléculas de adhesión y las quimiocinas en la migración de las CDs a los ganglios linfáticos
Las moléculas de adhesión y las quimiocinas también determinan la migración de los linfocitos vírgenes a los OLS

17
Homing o extravasación sitio específica de linfocitos determinada por:
Patrón de expresión de moléculas de adhesión sobre el linfocito y el endotelio
Patrón de expresión de receptores de quimoquinassobre el linfocito y la distribución de dichas quimoquinas en los distintos órganos o tejidos
disparan la adherencia al endotelio vascular y/o dirigen la migración a y dentro del espacio intersticial
Extravasación leucocitaria
Videos
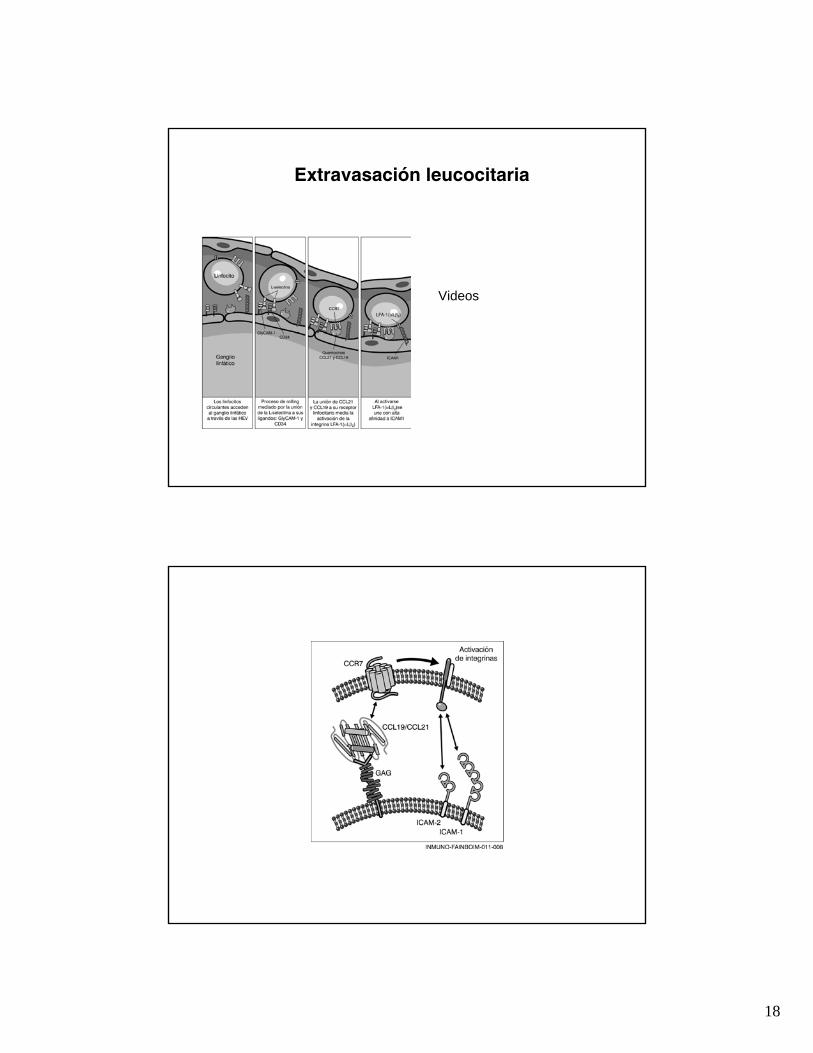
18
Extravasación leucocitaria
Videos

19
Cascada de extravasación leucocitaria
Videos

20
Qué sucede una vez que los linfocitos vírgenes se han extravasado en los OLS?
Migración de los linfocitos en el ganglio en ausencia de antígeno específico
Choreography of Cell Motility and Interaction Dynamics Imaged by Two-Photon Microscopyin Lymphoid Organs. Annu. Rev. Immunol. 2008. 26:585–626.

21
Migración de los linfocitos en el ganglio en presencia de antígeno específico
Los linfocitos T exploran el área paracortical por mecanismos de haptotaxis mediados por CCL21 y CCL19
empleando las fibras reticulares como andamiaje

22
VIDEOS

23
Por qué los linfocitos vírgenes se extravasan en OLS y los efectores en
tejidos periféricos?
Linfocito T vírgen: L-selectina + CCR7+
Homing a órganoslinfáticos secundarios
Reconocimiento de addressinas vasculares

24
La L-selectina reconoce un motivo molecular presente en las addressinas vasculares GlyCAM-1, CD34 y MadCAM-1
expresadas en las HEV
Moléculas que median la extravasación de linfocitos T vírgenes en órganos linfáticos secundarios

25
La fiebre incrementa la extravasación de linfocitos vírgenes a los órganos linfáticos secundarios
Células T CD8+ vírgenes upregulanrápidamente CCR5 una vez que entran a los ganglios que drenan un sitio de vacunación. Las APC son inducidas porlas T CD4+ activadas a secretar ligandosde CCR5, aumentando la frecuencia de contacto entre células T CCR5+ CD8+ y la DCs “licenciada”
Co-localización CD4+ y CD mediada por CCL19
Las quimiocinas incrementan la probabilidad de interacción de los linfocitos T con las CD

26
Linfocito T vírgen
L-selectina+
CCR7+
Linfocito T efector
L-selectina-
CCR7-
Homing de linfocitos T
Homing a OL secundarios
Homing a tejidos periféricos (o folículos) según patrones de expresión de moléculas de adhesión y Rc de quimoquinas

27
CLACLA
Deficiencia en CCR9 o el bloqueo de CCL25 (su ligando), disminuye marcadamente el número de linfocitos T CD8+ en intestino delgado luego de inmunización intraperitoneal
CCL25 no se expresa en colon y la deficiencia de CCR9 no afecta la adhesión de linfocitos T a mucosa colónica
Los linfocitos T de memoria pueden clasificarse en base a sus patrones de migración
TCM: Linfocitos T de memoria centrales: L-selectina+ CCR7+ (similar a los nativos). También CXCR4 para migrar a MO y GLP donde se expresa CXCL12.
TEM: Linfocitos T efectores de memoria: L-selectina- CCR7-
TCM: Recirculan vía sangre-órg. linfático secundario-linfa. Son centinelas de tejidos linfoideos
TEM: Recirculan vía sangre-tejidos extralinfoides periféricos-ganglios locales vía linfático aferente-conducto toráxico-sangre. Patrullan tejidos periféricos. Algunos permanecen en tejidos periféricos como Linfocitos TEM residentes.

28
Cómo las células TEFE y las TME aprenden dónde migrar?
Observaciones en humanos mostraron que el sitio de ingreso del Ag influencia el patrón de migración de las TEFE generadas.
Herpes simplex ingresando por piel, prima linfocitos con receptores de homing a piel.
La vacunación oral induce mayores niveles de α4β7 (que confiere tropismo por intestino) sobre las TEFE y TME generadas, que la vacunación intramuscular o subcutánea
Células T activadas en ganglios mesentéricos expresan mayores niveles de α4β7 y CCR9 que las activadas en ganglios que drenan piel
Células T activadas en ganglios que drenan piel expresan receptores de homing que confieren tropismo por piel (ligandos de selectinas)
CD8+ activadas en ganglios cervicales recibiendo antígenos derivados de SNC adquieren altos niveles de α4β1 y ligando de P-selectina que les confiere capacidad de migrar a cerebro
Cuando células T CD8+ fueron estimuladas simultáneamenteen el mismo GLP con dos tumores diferentes, uno administradoip y otro administrado s.c., las células efectoras generadasadquirieron altos niveles de α4 integrinas o ligandos de P-selectina respectivamente. Ello sugirió que la fuente anatómicade la CPA es dominate sobre los factores del ambiente en el ganglio drenante.

29
Rol instructivo de las células dendríticasLas CD son responsables de imprimir a la células T el potencial de homing específico
Ratones deficientes en vitamina A poseen número reducido de linfocitos T efectores y memoria en mucosa intestinal
Experimentos in vitro mostraron que la presencia del metabolito de la vitamina A, el ácido retinoico durante la activación T induce α4β7 y CCR9. Además el ácido retinoico suprime la “up-regulación” por default de la FucT-VII y los ligandos de P- y E-selectina
Similares resultados obtenidos cuando las células T son activadas en presencia de CD obtenidas de PP
CD de PP expresan altos niveles de retinaldehido deshidrogenasa (RALDH), esencial para la biosíntesis de AR desde la vitamina A
Bloqueo de RALDH en CDs o del receptor de AR en células T disminuye la inducción de α4β7 por CD de PP y de GM
CD derivada de GL intestinal
CD derivada de GL de piel
?
Rol instructivo de las células dendríticasLas CD tienen capacidad de imprimir a la células T el potencial de homing específico
CD de GM o PP induce capacidad de homing a
intestino
CD de GLP induce mayores niveles de ligandos para
selectinas
RTA. POR DEFAULT
El microambiente de citoquinas del órgano linfático podría modular la expresión de receptores de homing (por ej. Aquellos asociados a los perfiles Th1 y Th2), sin embargo, la generación de tropismo por un tejido no parece ser determinada por el perfil

30
Moléculas involucradas en el tráfico linfocitario como blanco de drogas anti-inflamatorias
Un anticuerpo monoclonal anti-α4 integrinas, que bloquea la unión de α4β1 (VLA-4) a VCAM-1 de células TH1 infiltrantes de cerebro y la unión de α4β7 a MadCAM-1 sobre células TH1 infiltrantes de intestino es una
estrategia viable para el tratamiento de la esclerosis múltiple y la enfermedad de Crohn
Riesgo de incrementada susceptibilidad a infección

31
Homing de linfocitos B2
Migración a médula ósea y subepitelios
Linfocito B
Foco Primario
Plasmoblasto: Célula secretora de Ac con capacidad de dividirse y con potencial migratorio. Puede madurar transformándose en célula Plasmática.
Plasmocito o célula plasmática: Célula terminalmente diferenciada, inmóvil, incapaz de dividirse y con capacidad de secretar anticuerpos.
CXCR4 es necesario para que los linfocitos B se posicionen en la zona oscura, donde se produce mayoritariamente su ligando, CXCL12, por células estromales locales
Los centroblastos deben down-regular CXCR4 a efectos de migrar a la zona clara guiados por la incrementada expresión de CXCL13 por CFD

32
Homing de plasmoblastos
B de memoria provenientes de centrocitosque sufrieron “switch” a IgG o IgAreexpresan CCR7, CXCR5, CCR6 Y CXCR4. Recirculan por OLS y tejidos no linfoides

33
Tráfico linfocitario en sistemas inmunes asociados a mucosas
Vías de ingreso de antígenos
a través de mucosas

34
Nat Med. 11(4 Suppl):S45-53.; 2005

35
Memoria inmunológica
Los linfocitos T de memoria pueden clasificarse en base a sus patrones de migración
TCM: Linfocitos T de memoria centrales: L-selectina+ CCR7+ (similar a los nativos). No polarizados. También CXCR4 para migrar a MO y GLP donde se expresa CXCL12.
TEM: Linfocitos T efectores de memoria: L-selectina- CCR7-
TCM: Recirculan vía sangre-órg. linfático secundario-linfa. Son centinelas de tejidos linfoideos
TEM: Recirculan vía sangre-tejidos extralinfoides periféricos-ganglios locales vía linfático aferente-conducto toráxico-sangre. Patrullan tejidos periféricos. Algunos permanecen en tejidos periféricos como Linfocitos TEM residentes.

36
Dado que sólo a lo sumo unos cientos de linfocitos T vírgenes serán específicos para un Ag dado, se requiere una potente respuesta proliferativa de las células específicas frente a un patógeno a fin de combatir una
infección ocasionada por él. Un precursor origina más de 10.000 células hijas (más de 13 divisiones celulares en 5-8 días)
Cómo se generan los linfocitos T de memoria?
Modelos posibles para la diferenciación de células T de memoria

37
Características del pool de linfocitos T periféricos
Los linfocitos T de memoria CD8+ se mantienen casi indefinidamente en número constante
Los linfocitos T de memoria CD4+ disminuyen gradualmente con el tiempo
Señales que regulan la sobrevida del poolde linfocitos T vírgenes y T de memoria
Immunity 29, 848-862, 2008
IL-7 es producida por la CFR de los OLS: Es utilizada intermitentemente por los linfocitos nativos (1) porque su Rc se down-regula por la propia acción de la IL-7; y (2) por la salida de los
linfocitos de los OLS en su tráfico por el organismo

38
Propiedades de las células TMC y TME
Cinética de la respuesta T CD8 a la infección o vacunación
CD62Llow
CD127low
CCR7low
granzimaBhi
IFNγhi
TNFαmid
CD62Lhi
CD127hi
CCR7hi
granzimaB-
IFNγhi
TNFαhi
IL-2mid
CD127: cad. α Rcpara IL-7
CD62L: L-selectina
Inmediatamente después de la respuesta, el pool de células T de memoria Ag-específicas está constituido ppalmente por TME, las cuales
gradualmente se convierten en TMC

39
Dinámica de la respuesta T secundaria en tejidos periféricos
Cinética de la respuesta CD8 primaria y secundaria a una infección e impacto del número de encuentros con el antígeno
La re-exposición incrementa el
número de cél T de memoria
El número de encuentros con el Ag tiene un dramático impacto en las características de la población de memoria resultante:
La mayoría de las CD8 de memoria inducidas luego de estimulación primaria se convertirán en CD62L+ en semanas (L. monocitógenes) o meses (LCMV) después de la infección primaria. Una fracción (30-40%) producirá IL-2.
Siguiendo al desafío secundario, las TCD8 de memoria primarias se expandirán y convertirán en T CD8 efectoras secundarias, que posteriormente sufrirán una fase de contracción para establecer T CD8 memoria secundarias en # mayores que los de la T CD8 memoria primarias. Estas tienen turn-over más lento, dificultades para incrementar la expresión de CD62L (tardan años en expresarlo) y para producir IL-2: Mayor # TME

40
1) Antígeno dependiente:
Re-estimulación perpetua: (reservorios de antígenos en el individuo o patógeno endémico o latente y se reactiva)
2) Antígeno independiente:
Células de memoria de vida media larga Proliferación homeostática por citoquinas(IL-7 e IL-15)
Modelos para explicar el mantenimiento de la memoria inmunológica T
Modelo aceptado cuando el Ag es eliminado luego de la respuesta primaria
No es necesario parael mantenimiento dememoria frente apatógenos que estableceninfecciones agudasOcurre frente a patógenosque establecen infecciones persistentes
Mantenimiento de los niveles de anticuerpos en ausencia de estimulación antigénica
Posibilidades:
1.Células plasmáticas de muy larga vida media
2.Proliferación continua homeostática de células B de memoria y diferenciación a células plasmáticas de larga vida media (activación bystander?)

41
Bibliografía sobre memoria inmunológica
• Migration, maintenance and recall of memory T cells in peripheral tissues. Woodland DL, Kohlmeier JE. Nat Rev Immunol. 2009 Mar;9(3):153-61.
• Shaping and reshaping CD8+ T-cell memory. Harty JT, Badovinac VP. Nat Rev Immunol. 2008 Feb;8(2):107-19.
• The precursors of memory: models and controversies. Rafi Ahmed, Michael J. Bevan, Steven L. Reiner and Douglas T. Fearon. Nat. Rev. Immunol. 2009; 9: 662-668.