Embed Size (px)
Citation preview

RZECZPOSPOLITA POLSKA
Urząd Patentowy Rzeczypospolitej
Polskiej
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2913394
(96) Data i numer zgłoszenia patentu europejskiego: 18.12.2013 15161369.2 (97) O udzieleniu patentu europejskiego ogłoszono: 22.06.2016 Europejski Biuletyn Patentowy 2016/25 EP 2913394 B1
(13) (51)
T3 Int.Cl. A61K 35/15 (2015.01) C12N 5/0784 (2010.01)
(54) Tytuł wynalazku: WSPÓLNE RÓŻNICOWANIE I AKTYWOWANIE MONOCYTÓW OD DAWCÓW ALLOGENICZNYCH DLA DOSTARCZENIA PROZAPALNYCH KOMÓREK DENDRYTYCZNYCH
PL/E
P 29
1339
4 T3
(30) Pierwszeństwo:
18.12.2012 EP 12197687
(43) Zgłoszenie ogłoszono:
02.09.2015 w Europejskim Biuletynie Patentowym nr 2015/36
(45) O złożeniu tłumaczenia patentu ogłoszono:
30.12.2016 Wiadomości Urzędu Patentowego 2016/12
(73) Uprawniony z patentu:
Immunicum AB, Göteborg, SE
(72) Twórca(y) wynalazku:
ALEX KARLSSON-PARRA, Uppsala, SE BENGT ANDERSSON, Mölndal, SE
(74) Pełnomocnik: rzecz. pat. Wojciech Tykarski
SULIMA GRABOWSKA SIERZPUTOWSKA BIURO PATENTÓW I ZNAKÓW TOWAROWYCH SP.J. Skr. poczt. 6 00-956 Warszawa 10
Uwaga: W ciągu dziewięciu miesięcy od publikacji informacji o udzieleniu patentu europejskiego, każda osoba może wnieść do Europejskiego Urzędu Patentowego sprzeciw dotyczący udzielonego patentu europejskiego. Sprzeciw wnosi się w formie uzasadnionego na piśmie oświadczenia. Uważa się go za wniesiony dopiero z chwilą wniesienia opłaty za sprzeciw (Art. 99 (1) Konwencji o udzielaniu patentów europejskich).

SGS-8817/VAL EP 2 913 394 B1 Opis Dziedzina wynalazku
[0001] Niniejszy wynalazek dotyczy sposobu wytwarzania niewyczerpanych (ang. non-exhausted) niedojrzałych ludzkich komórek dendrytycznych (DC) uzyskanych z monocy-tów.
Tło wynalazku 5
[0002] Tradycyjne terapie przeciwnowotworowe, takie jak chirurgia, naświetlanie i che-mioterapia są często niewystarczające do leczenia pacjentów i zazwyczaj powodują po-ważne skutki uboczne. Immunoterapia okazała się obiecująca jako alternatywny sposób le-czenia z mniejszą ilością szkodliwych skutków ubocznych. [0003] Obecnie dobrze wiadomo, że układ immunologiczny zawiera komórki, w szcze-10 gólności cytotoksyczne limfocyty T (CTL) CD8+, które mogą rozpoznawać i potencjalnie zabijać komórki nowotworowe. Niemniej jednak, głównym problemem jest to, że zdolność do zabijania przez te limfocyty T nie jest indukowana lub tylko słabo indukowana u pa-cjentów z nowotworem. Jedną z możliwości jest to, że u pacjentów z nowotworem zacho-dzi niewystarczająca prezentacja antygenów nowotworowych i ko-stymulacja przez ko-15 mórki dendrytyczne (DC), „naturalny adiuwant” do wywoływania funkcjonalnej i specy-ficznej względem nowotworu odporności limfocytów T. [0004] Obecne strategie immunoterapii nowotworów skupiają się głównie na autologicz-nych, pochodzących od pacjentów DC załadowanych antygenem, które poddano różnico-waniu i załadowano antygenem ex vivo. Podstawowym założeniem tego rozwiązania jest 20 to, że wydajność i kontrola zapewniana przez manipulację DC ex vivo wytwarza DC o sil-nej zdolności do prezentacji antygenu i kostymulacji. Jakość odpowiedzi limfocytów T za-leży od zdolności tych autologicznych DC do prezentacji antygenów nowotworowych lim-focytom T w sposób wykazujący restrykcję MHC (komórki DC i T muszą pochodzić od tego samego pacjenta) w odprowadzających węzłach chłonnych, a zatem do tworzenia od-25 powiedzi limfocytów T specyficznej względem nowotworu. [0005] Autologiczne DC pochodzące z monocytów są najpowszechniej stosowanymi DC w badaniach pilotażowych, ponieważ możliwe jest uzyskanie miliardów monocytów z leu-kocytów krwi obwodowej zebranych przez leukaferezę, pracochłonną i czasochłonną pro-cedurę, w której białe krwinki oddziela się od krążącej krwi. Dostępnych jest kilka sposo-30 bów dalszego wzbogacania monocytów i dwa z tych sposobów, elutriację i izolację na przeciwciałach/kulkach, można również przeprowadzić zgodnie z wytycznymi Dobrej Praktyki Wytwarzania (GMP). [0006] Następnie monocyty hoduje się w podłożu uzupełnionym GM-CSF i IL-4 przez 4-7 dni, co prowadzi do ich różnicowania w niedojrzałe DC, które to niedojrzałe DC charak-35 teryzują się swoją wyjątkową zdolnością wytwarzania dużych ilości chemokin i cytokin prozapalnych po późniejszej stymulacji pewnymi typami czynników aktywujących (Sallusto i in., Eur J Immunol, 1999. 29:1617; Napolitani i in., Natuer Immunology 2005. 6: 769). Stymulowane DC są zazwyczaj poddawane wstępnemu impulsowi odpowiednie-go(-ich) antygenu(-ów) nowotworowego(-ych) i aktywowane przez 1-2 dni przed szcze-40 pieniem. Jednak odpowiedzi immunologiczne na takie szczepionki oparte na DC są często słabe, a odpowiedzi kliniczne rzadko są całkowite i długotrwałe. [0007] Niewiele wiadomo na temat losu i funkcji wytwarzanych ex vivo autologicznych DC po ich wstrzyknięciu. U ludzi wzór migracji wstrzykniętej szczepionki z DC został ostatnio prześledzony in vivo i, przede wszystkim, mniej niż 5% wstrzykniętych DC osią-45 gnęło odprowadzające węzły chłonne, podczas gdy większość DC pozostało w miejscu

2
wstrzyknięcia. Te miejscowo zatrzymane DC ze szczepionki szybko traciły swoją żywot-ność, a następnie były usuwane przez rekrutowane komórki prezentujące antygen. [0008] Obecnie dostarczono dane, że wstrzyknięte DC ze szczepionki, które były akty-wowane ex vivo przez określony okres czasu (tj. 6 do 18 h) stają się prozapalnymi (PI) DC, które są w stanie pośrednio preaktywować natywne limfocyty T CD8+ in vivo przez dzia-5 łanie jako wyłącznie miejscowy adiuwant immunologiczny. Ta funkcja adiuwantowa wstrzykniętych PI DC jest ściśle zależna od ich ciągłego wydzielania niektórych chemokin rekrutujących DC i komórki NK w czasie podania (po usunięciu czynników aktywują-cych). Takie PI-DC również eksprymują/wydzielają czynniki, które wywołują aktywację rekrutowanych endogennych komórek NK i DC w miejscu szczepienia. W odróżnieniu od 10 PI-DC, długo (tj. > 24 h) aktywowane DC, które były powszechnie stosowane w badaniach klinicznych, charakteryzują się swoim stanem „wyczerpania” (Langenkamp i in. 2000), a zatem nie są w stanie wydzielać pożądanych chemokin i czynników aktywujących DC w czasie podania. [0009] Podsumowując, PI-DC mogą działać nie tylko jako bezpośrednie stymulatory 15 zgodnych pod względem MHC autologicznych limfocytów T, ale również działają jako ad-iuwant do wytwarzania dużych ilości chemokin i cytokin prozapalnych w czasie podania. Miejscowe wstrzyknięcie PI-DC będzie prowadziło do rekrutacji i aktywacji innych komó-rek immunologicznych, w tym krążących komórek NK i prekursorów DC. Jeśli wstrzyk-nięte PI-DC zostały wcześniej załadowane odpowiednimi antygenami nowotworowymi lub 20 wstrzyknięte bezpośrednio do istniejącej zmiany nowotworowej, rekrutowane endogenne DC pochłoną odpowiednio umierające komórki ze szczepionki eksprymujące odpowiednie antygeny nowotworowe lub umierające komórki nowotworowe eksprymujące antygen. Po aktywacji te rekrutowane, a następnie załadowane antygenem endogenne DC będą migro-wać do odprowadzających węzłów chłonnych, gdzie preaktywują one limfocyty T specy-25 ficzne dla nowotworu w sposób wykazujący restrykcję MHC (Liu i in., 2008). Wniosek ten jest poparty przez dane z kilku ostatnich badań przedklinicznych, w których wzrost guza był istotnie zmniejszony przez szczepienia terapeutyczne niewyczerpanymi, MHC-niezgodnymi allogenicznymi PI-DC (Alder i in. 2008, Siders i in. 2009, Edlich i in. 2010) [0010] Zatem silna funkcja adiuwantowa PI-DC, która, co ważne, nie wymaga zgodności 30 pod względem MHC między PI-DC i limfocytami T pacjenta, wprowadza możliwość sto-sowania wstępnie wytworzonych i przechowywanych w stanie zamrożenia niezgodnych pod względem MHC, allogenicznych, PI-DC jako szczepionek „dostępnych od ręki”, sta-nowiących realną, praktyczną alternatywę dla obecnych wytwarzanych na zamówienie, szczepionek DC specyficznych dla danego pacjenta. Zastosowanie takich niezgodnych pod 35 względem MHC, allogenicznych PI-DC ujawniono w EP 1509244 B1 i WO 2011/098516. [0011] Z przyczyn etycznych, zdobywanie monocytów od zdrowych dawców krwi przez leukaferezę na dużą skalę wyłącznie do celów komercyjnego wytwarzania szczepionki do stosowania klinicznego nie jest możliwe. W praktyce, dostępny surowiec do wytwarzania PI-DC, tj. monocyty, jest zatem ograniczony do monocytów uzyskanych z produktu odpa-40 dowego (kożuszków leukocytarno-płytkowych i/lub użytych filtrów do usuwania leukocy-tów) w przebiegu oddzielania niepożądanych leukocytów od różnych składników pełnej krwi lub monocytów uzyskiwanych z kożuszków leukocytarno-płytkowych w bankach krwi. [0012] Jednakże, całkowita liczba monocytów, która może zostać wyizolowana z każdego 45 kożuszka leukocytarno-płytkowego lub każdego filtra do usuwania leukocytów z worka z krwią jest zazwyczaj mniejsza niż 200 milionów (Ebner S i in., Generation of large num-bers of human dendritic cells from whole blood passaged through leukocyte removal fil-ters: an alternative to standard buffy coats J Immunol Methods 252 (2001), co prowadzi do niedopuszczalnie wysokich kosztów wzbogacania, a następnie różnicowania do DC po-50 szczególnych partii monocytów (każda partia pochodzi od pojedynczego dawcy) sposoba-

3
mi, które są zgodne z wytycznymi Dobrej Praktyki Wytwarzania (GMP) zgodnymi z tą dziedziną. [0013] Zatem, istnieje potrzeba w dziedzinie opracowania wielkoskalowego i opłacalnego sposobu wytwarzania z jakością kliniczną, niewyczerpanych niedojrzałych komórek den-drytycznych z zawierającego monocyty odpadu uzyskanego z banków krwi. 5
Streszczenie
[0014] W konsekwencji, niniejszy wynalazek ma na celu zminimalizowanie, złagodzenie, wyeliminowanie lub przezwyciężenie jednego lub większej liczby z wymienionych powy-żej potencjalnych deficytów w dziedzinie i wad, pojedynczo lub w dowolnym połączeniu, przez dostarczenie sposobu sposobu wytwarzania niewyczerpanych niedojrzałych komórek 10 dendrytycznych (DC) pochodzących od co najmniej dwóch różnych, allogenicznych daw-ców. W sposobie tym dostarcza się mieszaninę allogenicznych leukocytów, które to allo-geniczne leukocyty zostały uzyskane od co najmniej dwóch różnych allogenicznych daw-ców. Następnie allogeniczne monocyty izoluje się z mieszaniny allogenicznych leukocy-tów z dostarczeniem allogenicznych leukocytów wzbogaconych w monocyty. Następnie, 15 niewyczerpane niedojrzałe DC wytwarza się z allogenicznych leukocytów wzbogaconych w monocyty przez wspólne hodowanie allogenicznych leukocytów wzbogaconych w mo-nocyty przez 2 do 7 dni w wodnym podłożu do hodowli komórek wolnym od surowicy in-nej niż ludzka. Podłoże uzupełnia się interleukiną-4 (IL-4) oraz czynnikiem stymulującym tworzenie kolonii granulocytów i makrofagów (GM-CSF). 20 [0015] W przeciwieństwie do powszechnego przekonania, wytworzone niedojrzałe ko-mórki dendrytyczne uzyskane z monocytów są niewyczerpane, a zatem zdolne do wytwa-rzania znacznych ilości chemokin prozapalnych i cytokin prozapalnych w trwały sposób po aktywacji do prozapalnych DC. Zatem, sposób stanowi wielkoskalowy i opłacalny spo-sób wytwarzania, z jakością kliniczną, niewyczerpanych niedojrzałych komórek dendry-25 tycznych. [0016] Kolejny aspekt wynalazku dotyczy mieszaniny allogenicznych niewyczerpanych niedojrzałych komórek dendrytycznych (DC) pochodzących od co najmniej dwóch róż-nych, allogenicznych dawców. Taką mieszaninę uzyskuje się opisanym sposobem. [0017] Kolejny aspekt wynalazku dotyczy sposobu wytwarzania prozapalnych DC. Przez 30 aktywację niewyczerpanych niedojrzałych DC otrzymuje się prozapalne DC. Takim spo-sobem można otrzymać mieszaninę allogenicznych prozapalnych komórek dendrytycz-nych pochodzących od co najmniej dwóch różnych, allogenicznych dawców. Tę mieszani-nę można formułować w kompozycję farmaceutyczną zawierającą ponadto co najmniej je-den farmaceutycznie dopuszczalny nośnik. Tę mieszaninę i kompozycję farmaceutyczną, 35 odpowiednio, można stosować do leczenia nowotworu. [0018] Dalsze korzystne cechy wynalazku są zdefiniowane w zależnych zastrzeżeniach. Ponadto, korzystne cechy wynalazku omówiono w ujawnionych tutaj postaciach.
Szczegółowy opis korzystnych postaci
[0019] Twórcy niniejszego wynalazku przewidywali, że zawierające monocyty populacje 40 leukocytów, które są obecne w kożuszkach leukocytarno-płytkowych lub zatrzymane na filtrach do usuwania leukocytów stosowanych do usuwania leukocytów z pełnej krwi (co jest stosowane do wzbogacania w krwinki czerwone) lub filtrach do usuwania leukocytów stosowanych do usuwania leukocytów z połączonych kożuszków leukocytarno-płytkowych (wykorzystywanych do wzbogacenia w płytki krwi), można potencjalnie sto-45 sować do wielkoskalowego i opłacalnego wytwarzania niewyczerpanych niedojrzałych komórek dendrytycznych. [0020] Jak wcześniej wykazano, leukocyty zatrzymywane przez różne rodzaje filtrów do usuwania leukocytów mogą być odzyskane przez wsteczne przemycie odpowiednim pod-łożem, a następnie wzbogacenie w monocyty (Ebner S i in., Generation of large numbers 50

4
of human dendritic cells from whole blood passaged through leukocyte removal filters: an alternative to standard buffy coats J Immunol Methods 252 (2001) 93-104; oraz Meyer T P H i in., Filter Buffy Coats (FBC): A source of peripheral blood leukocytes recovered from leukocyte depletion filters. J Immunol Methods 307 (2005) 150-166). [0021] Kożuszek leukocytarno-płytkowy jest frakcją antykoagulowanej próbki krwi, która 5 zawiera większość leukocytów, w tym neutrofile, bazofile, eozynofile, monocyty i limfo-cyty, oraz płytki krwi po odwirowaniu pełnej krwi w gradiencie gęstości. Kożuszki leuko-cytarno-płytkowe są zazwyczaj stosowane jako surowiec do wytwarzania płytek krwi. Podczas tego procesu, zmniejszenie liczby leukocytów przez filtr usuwający leukocyty prowadzi się po połączeniu 4 do 8 kożuszków leukocytarno-płytkowych. Zazwyczaj, do 10 izolacji płytek krwi z kożuszka leukocytarno-płytkowego stosuje się urządzenie TACSI lub system OrbiSac. [0022] Urządzenie TACSI do wytwarzania płytek krwi z połączonych kożuszków leuko-cytarno-płytkowych zawiera moduł systemowy (umieszczony na wirniku) i wkładkę, którą można usunąć do zamontowania zestawu TACSI. Każdy moduł systemowy jest dostarcza-15 ny z układem tłoczącym, kontrolowanym i monitorowanym przez indywidualny mikropro-cesor. W pierwszej kolejności, połączone kożuszki leukocytarno-płytkowe w zbiorniku do łączenia osadza się przez odwirowanie w położeniu pionowym. W kolejnym etapie war-stwę bogatą w płytki krwi (zawierającą również znaczne ilości leukocytów, włącznie z monocytami i limfocytami) supernatantu połączonego kożuszka leukocytarno-płytkowego 20 przenosi się do pojemnika do przechowywania przez aktywację układu tłoczącego w każ-dym module. Ponadto filtr do usuwania leukocytów jest zintegrowany z zestawem TACSI między workiem do przetwarzania i końcowym zbiornikiem do przechowywania. Następ-nie wyładowuje się filtr do usuwania leukocytów i resztę kożuszka leukocytarno-płytkowego ze zbiornika do łączenia kożuszków leukocytarno-płytkowych, przy czym 25 obydwa zawierają znaczne ilości monocytów. [0023] W alternatywnym systemie OrbiSac do automatycznego wzbogacenia w płytki krwi, pojemnik do łączenia kożuszków leukocytarno-płytkowych jest w kształcie pierście-nia. Po odwirowaniu bogatą w płytki krwi środkową część supernatantu przenosi się do po-jemnika umieszczonego w środku wirówki, a przeniesienie następuje przez zintegrowany 30 filtr do usuwania leukocytów. Następnie wyładowuje się filtr do usuwania leukocytów i resztę kożuszka leukocytarno-płytkowego ze zbiornika do łączenia kożuszków leukocytar-no-płytkowych, przy czym obydwa zawierają znaczne ilości monocytów. [0024] Podsumowując, dostępne w dziedzinie sposoby wzbogacania w płytki krwi z ko-żuszka leukocytarno-płytkowego zapewniają dwa możliwe źródła monocytów, tj. resztę 35 kożuszka leukocytarno-płytkowego pozbawioną płytek krwi, nazywaną również kożusz-kiem leukocytarno-płytkowym pozbawionym płytek kwi, oraz filtr do usuwania leukocy-tów. Kożuszek leukocytarno-płytkowy pozbawiony płytek krwi oraz filtr do usuwania leu-kocytów zawierają mieszaninę leukocytów, w tym do 1 miliarda monocytów. Jednakże monocyty te są allogeniczne w stosunku do siebie, ponieważ pochodzą one od różnych, al-40 logenicznych dawców, ze względu na łączenie kożuszka leukocytarno-płytkowego przed usuwaniem płytek krwi. [0025] Nie połączone kożuszki leukocytarno-płytkowe, zawierające płytki lub filtry uzy-skane po usunięciu leukocytów z pełnej krwi będą dostarczać maksymalnie tylko do około 100 do 200 milionów monocytów na kożuszek leukocytarno-płytkowy lub filtr. Zatem mu-45 szą być one łączone w celu zapewnienia ilości wystarczającej do opłacalnego wytwarzania zgodnie z GMP niewyczerpanych niedojrzałych DC. [0026] Podobnie do leukocytów uzyskanych przy wytwarzaniu z płytek krwi, połączone leukocyty z kożuszków leukocytarno-płytkowych zawierających płytki krwi lub z filtrów stosowanych do usuwania leukocytów z pełnej krwi będą również składać się z mieszanej 50 populacji komórek pochodzących od różnych allogenicznych dawców.

5
[0027] Łączenie leukocytów z co najmniej 5 do 10 kożuszków leukocytarno-płytkowych lub łączenie wymytych leukocytów z co najmniej 5 do 10 filtrów leukocytarnych z pełnej krwi rozwiąże, przynajmniej teoretycznie, problem zapewnienia dostatecznej liczby leuko-cytów do wielkoskalowego i opłacalnego wytwarzania jakości klinicznej niewyczerpanych niedojrzałych DC. 5 [0028] Podobnie, połączone kożuszki leukocytarno-płytkowe pozbawione płytek krwi i/lub filtry do usuwania leukocytów stosowane do wzbogacania płytek krwi z kożuszka leukocytarno-płytkowego mogą być potencjalnie stosowane do dostarczenia wystarczają-cej liczby leukocytów do wielkoskalowego i opłacalnego wytwarzania z jakością klinicz-ną, niewyczerpanych niedojrzałych DC. 10 [0029] Jak wcześniej wykazano, leukocyty zatrzymane przez filtry usuwające leukocyty mogą zostać odzyskane przez wsteczne przemywanie odpowiednim podłożem, a następnie wzbogacane w monocyty (Ebner S i in., Generation of large numbers of human dendritic cells from whole blood passaged through leukocyte removal filters: an alternative to stan-dard buffy coats J Immunol Methods 252 (2001) 93-104; oraz Meyer T P H i in., Filter 15 Buffy Coats (FBC): A source of peripheral blood leukocytes recovered from leukocyte de-pletion filters. J Immunol Methods 307 (2005) 150-166). [0030] Jednakże przewidywanym problemem związanym z późniejszą wspólną hodowlą leukocytów wzbogaconych w monocyty, które pochodzą od różnych, allogenicznych daw-ców, jest to, że uważa się, że ich niezgodność pod względem antygenów głównego układu 20 zgodności tkankowej (MHC) klasy I i klasy II prowadzi do przedwczesnej aktywacji mo-nocytów/niedojrzałych DC od jednego dawcy przez zanieczyszczające alloreaktywne lim-focyty T i/lub naturalne komórki cytotoksyczne od innego dawcy. [0031] Wiadomo, że wspólne hodowanie w standardowym podłożu do hodowli komórek, takim jak RPMI-1640 (RPMI = Roswell Park Memorial Institute, w którym to instytucie 25 podłoże to zostało pierwotnie opracowane przez Moore i in.) z płodową surowicą cielęcą lub wolnym od surowicy podłożu do hodowli komórek, takim jak X-VIVO 15, komórek jednojądrzastych, w tym monocytów, limfocytów i komórek NK, od dwóch allogenicz-nych dawców indukuje wytwarzanie dobrze znanych czynników aktywujących DC, włącz-nie z TNF-alfa (Laurin i in., Transplantation 2004; 77:267; Wallgren i in., Scand J Immu-30 nol 2005; 62:234). Ponadto, wielokrotnie wykazano, że dodanie supernatantów hodowli z takich wspólnych hodowli allogenicznych komórek jednojądrzastych w standardowym podłożu RPMI-1640 z płodową surowicą cielęcą lub w standardowym podłożu wolnym od surowicy, wywołuje aktywację/dojrzewanie uzyskanych z monocytów, niedojrzałych DC (Laurin i in., Transplantation 2004; 77:267; Wallgren i in., Scand J Immunol 2005; 35 62:234). [0032] Ponadto wykazano, że dodanie TNF-alfa do standardowego podłoża RPMI-1640 uzupełnionego GM-CSF i IL-4 stosowanych do różnicowania monocytów do niedojrza-łych DC wywołuje przedwczesną aktywację i następnie wyczerpanie (tolerancję) różnico-wanych DC (Rieser C i in., Differential Deactivation of Human Dendritic Cells by Endo-40 toxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117). [0033] Problem z zanieczyszczającymi limfocytami T i komórkami NK jest istotny nawet po zgodnym z GMP wzbogacaniu (elutriacji i izolacji na przeciwciałach/kulkach) monocy-tów z połączonych kożuszków leukocytarno-płytkowych lub z filtrów do usuwania leuko-45 cytów (Schwanke i in., Journal of Clinical Apheresis 21: 153-157 (2006); Meyer i in., Jo-urnal of Immunological Methods 307 (2005) 150-166), ze względu na trudności w wytwa-rzaniu populacji komórek monocytów zasadniczo wolnych od zanieczyszczających limfo-cytów T i komórek NK. [0034] Ponadto, i co istotne, nie tylko wspólna hodowla allogenicznych limfocytów w 50 standardowym podłożu, ale również wspólna hodowla monocytów z allogenicznymi neu-trofilami w podłożu RPMI-1640 uzupełnionym płodową surowicą cielęcą, powoduje

6
zwiększenie ekspresji błonowych CD40, CD86 oraz ludzkiego antygenu leukocytów (HLA)-DR na DC, tj. przedwczesną aktywację, jak wykazano przez Meggiovanni i in. (patrz Journal of Leukocyte Biology, 2006; 79; 977-988). Wykazano, że zasadnicze usu-nięcie neutrofili z monocytów w połączonych kożuszkach leukocytarno-płytkowych lub leukocytów wymytych z filtrów z użyciem elutriacji (Schwanke i in., Journal of Clinical 5 Apheresis 21:153-157 (2006)) lub izolacji na przeciwciałach/kulkach (Meyer i in., Journal of Immunological Methods 307 (2005) 150-166) jest bardzo trudne. Zazwyczaj takie pro-dukty wzbogacone w monocyty zawierają znaczącą ilość (tj. 25-40% w stosunku do cał-kowitej liczby obecnych komórek) neutrofili. Z punktu widzenia bezpieczeństwa to zanie-czyszczenie neutrofilami nie stanowi jednak problemu. 10 [0035] Ponadto, nawet jeśli możliwe byłoby wytworzenie 100% czystych populacji komó-rek monocytów, to nie wyeliminuje to ryzyka przedwczesnej aktywacji ze względu na ak-tywne oddziaływania między monocytami od różnych, allogenicznych dawców. W nie-dawno opublikowanym artykule przeglądowym pt. „Origin and biology of the allogeneic response” autorstwa wybitnych i cenionych immunologów Fadi G. Lakkis i Robert I. 15 Lechler (patrz Cold Spring Harbor perspectives in medicine, tom 3, nr 8, 2013) stwierdzo-no, że rzeczywiście istnieją wrodzone mechanizmy allorozpoznawania. Twórcy twierdzą, że „odrzucenie alloprzeszczepu nie ogranicza się do zwierząt kręgowych obdarzonych ad-aptywnym układem immunologicznym, ale jest powszechne wśród wielu organizmów bez-kręgowych, które poprzedzają ewolucję odporności adaptywnej (zwierzęta, u których brak 20 jest limfocytów T i B, komórek NK, enzymów somatycznej rearanżacji genów i MHC)”. [0036] Ponadto, a może i bardziej bezpośrednio, allogeniczne odpowiedzi są również ob-serwowane u myszy pozbawionych komórek limfoidalnych. Aktywacja monocytów zależy od różnic w antygenach nie-MHC między monocytami biorcy i wstrzykniętymi alloge-nicznymi leukocytami dawcy, włącznie z allogenicznymi monocytami (Zecher D i in., An 25 Innate Response to Allogeneic Nonself Mediated by Monocytes. J Immunol 83 (2009) 7810-7816). Zecher i in. wykazali, że wstrzyknięcie allogenicznych leukocytów do mał-żowiny usznej myszy RAG2/2, pozbawionych limfocytów T i B, wywołuje istotnie więk-szy obrzęk i naciekanie skóry komórkami szpikowymi gospodarza niż wstrzyknięcie syn-genicznych leukocytów. Odpowiedź na allogeniczne leukocyty występowała niezależnie 30 od komórek NK i pośredniczyły w niej monocyty. Ponadto, odpowiedź monocytów była skierowana przeciw allodeterminantom nie związanym z MHC. [0037] W jeszcze bardziej aktualnej pracy (Zeng Q, i in. „Innate recognition of allogeneic non-self induces monocyte differentiation to mature dendritic cells in vivo.” Am J Trans-plant 12: 148-148, 2012), twórcy wykazali że alloprzeszczepy serca przeszczepione my-35 szom gc2/2RAG2/2, pozbawionym limfocytów T, B i komórek NK, są szybko naciekane przez monocyty gospodarza, które różnicują się w dojrzałe, eksprymujące IL-12 komórki dendrytyczne (DC). Jednakże determinanty na komórkach allogenicznych gospodarza, któ-re wywołują dojrzewanie monocytów gospodarza oraz przypuszczalne receptory monocy-tów, które je rozpoznają nie są jeszcze znane. 40 [0038] Istnieje zatem wyraźny dowód, że monocyty ssaków bezpośrednio odpowiadają na determinanty nie-MHC na allogenicznych komórkach niezależnie od limfocytów T, B i komórek NK. Zatem zgodnie z ogólnie dominującym rozumieniem, alloreaktywność uwa-ża się za ogólną właściwość monocytów. [0039] Podsumowując, oznacza to, że wyraźnie przewiduje się, że wspólne hodowanie 45 wzbogaconych w monocyty populacji komórek pochodzących od różnych allogenicznych dawców prowadzi do wstępnej aktywacji, a następnie wyczerpania monocytów podczas ich różnicowania do DC uzyskanych z monocytów. Zatem DC nie będą w stanie stać się PI-DC wytwarzającymi wymagane czynniki adiuwantowe w sposób trwały, gdy będą po-nownie stymulowane odpowiednimi czynnikami aktywującymi. 50 [0040] To przewidywane wyczerpanie wywołane aktywacją jest podobne do dobrze zna-nego wyczerpania monocytów, makrofagów i DC, które jest wywoływane przez przed-

7
wczesną aktywację czynnikami zapalnymi, takimi jak TNF-α (Park i in., Nat Immunol. 2012; 12:607-615) lub lipopolisacharydami mikroorganizmów (LPS) (Rieser C i in., Dif-ferential Deactivation of Human Dendritic Cells by Endotoxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117; Langenkamp A i in., Kinetics of dendritic cell activation: impact on priming TH1, TH2 and nonpolar-5 ized T cells. Nature Immunol. 1 (2000) 311-316). [0041] Twórcy niniejszego wynalazku jednakże nieoczekiwanie stwierdzili, że niewy-czerpane niedojrzałe DC faktycznie można namnażać z początkowej populacji komórek zawierającej mieszaninę allogenicznych leukocytów wzbogaconych w monocyty od róż-nych, allogenicznych dawców, w sposób podobny do tego, który jest stosowany do nam-10 nażania niewyczerpanych niedojrzałych DC ze wzbogaconych monocytów, które pocho-dzą od jednego dawcy (patrz WO 2011/098516), tj. przez użycie wodnego podłoża do ho-dowli komórek wolnego od surowicy innej niż ludzka, ale uzupełnionego interleukiną-4 (IL-4) oraz czynnikiem stymulującym tworzenie kolonii granulocytów i makrofagów (GM-CSF). 15 [0042] W przeciwieństwie do powszechnego przekonania, nieoczekiwanie wykazano, że namnażanie mieszaniny wzbogaconej w monocyty od różnych, allogenicznych dawców, do niedojrzałych DC w pewnych warunkach, nie powoduje przedwczesnej aktywacji, a na-stępnie wyczerpania DC. Warunki te obejmują wspólną hodowlę w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka oraz uzupełnionym GM-CSF i IL-20 4. [0043] Jednakże wspólna hodowla w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka i nie uzupełnionym interleukiną-4 (IL-4) oraz czynnikiem sty-mulującym tworzenie kolonii granulocytów i makrofagów, a także wspólna hodowla w wodnym podłożu do hodowli komórek zawierającym surowicę inną niż ludzka, np. płodo-25 wą surowicę bydlęcą, i uzupełnionym interleukiną-4 (IL-4) oraz czynnikiem stymulującym tworzenie kolonii granulocytów prowadzi do, jak oczekuje się i uznaje w dziedzinie, przedwczesnej aktywacji, a następnie wyczerpania DC. [0044] Zatem, monocyty można izolować z połączonych populacji leukocytów od różnych allogenicznych dawców, co umożliwia przeprowadzenie wielkoskalowego i opłacalnego, 30 zgodnego z GMP wzbogacania w monocyty, takiego jak elutriacja (Elutra) lub izolacja na przeciwciałach/kulkach (CliniMacs). Następnie, niewyczerpane niedojrzałe DC mogą być wytwarzane z izolowanych allogenicznych leukocytów wzbogaconych w monocyty bez przedwczesnej aktywacji. [0045] Takie niewyczerpane niedojrzałe DC mogą być stosowane do namnażania proza-35 palnych DC, które mają na celu działanie jako szczepionka przeciwnowotworowa po wstrzyknięciu do guza (patrz WO 2011/098516). Ponadto, niewyczerpane niedojrzałe DC od różnych, allogenicznych dawców, mogą być załadowane antygenem(-ami) nowotwo-rowym przed aktywacją, w celu wytworzenia „pełnej” komórkowej allogenicznej szcze-pionki przeciwnowotworowej (patrz EP 1509244 B1), która może być wstrzykiwana w 40 różnych miejscach, w tym do guza, podskórnie, naskórkowo, domięśniowo i/lub dożylnie. [0046] Pewna postać dotyczy zatem sposobu wytwarzania niewyczerpanych niedojrzałych DC z mieszaniny allogenicznych leukocytów wzbogaconych w monocyty. W takim sposo-bie, dostarcza się mieszaninę allogenicznych leukocytów, uzyskaną od co najmniej dwóch różnych, allogenicznych dawców. Zgodnie z jedną postacią, przez dwóch różnych alloge-45 nicznych dawców rozumie się, że ci dwaj osobnicy oddający leukocyty są z tego samego gatunku, ale o różnym składzie genetycznym, tj. odrębni antygenowo. Jak już opisano, al-logeniczne leukocyty można uzyskać z połączonych kożuszków leukocytarno-płytkowych lub przez wymywanie leukocytów ze zużytych filtrów do usuwania leukocytów. Następnie allogeniczne monocyty izoluje się z dostarczonej mieszaniny allogenicznych leukocytów. 50 Następnie, niewyczerpane niedojrzałe DC wytwarza się z izolowanych allogenicznych leukocytów wzbogaconych w monocyty.

8
[0047] Oprócz zdolności do przeprowadzania wielkoskalowego i opłacalnego, zgodnego z GMP wzbogacania w monocyty, kolejną zaletą niewyczerpanych niedojrzałych DC, uzy-skanych z mieszaniny allogenicznych monocytów pochodzących od co najmniej dwóch różnych allogenicznych dawców, jest to, że zostanie zmniejszona normalna zmienność bio-logiczna wytwarzania różnych czynników prozapalnych po aktywacji, która, jak wiadomo, 5 istnieje między PI-DC od różnych dawców. [0048] W korzystnej postaci, allogeniczne leukocyty dostarcza się przez połączenie co najmniej dwóch kożuszków leukocytarno-płytkowych zawierających leukocyty. Kożuszki leukocytarno-płytkowe do połączenia uzyskuje się od co najmniej dwóch różnych, alloge-nicznych dawców. Połączone kożuszki leukocytarno-płytkowe mogą zawierać płytki krwi 10 lub mogą być pozbawione płytek krwi. [0049] Allogeniczne leukocyty można również dostarczyć przez wymycie leukocytów z co najmniej dwóch filtrów do usuwania leukocytów, które to filtry, odpowiednio, zostały wcześniej użyte do usuwania leukocytów z pełnej krwi od co najmniej dwóch różnych al-logenicznych dawców. Po wymyciu uzyskane leukocyty łączy się aby otrzymać mieszani-15 nę allogenicznych leukocytów. Oczywiście, ale mniej korzystnie, pełna krew może rów-nież być łączona przed usuwaniem leukocytów. Procedura wymywania leukocytów z filtru do ich usuwania, który to filtr był wcześniej użyty do usunięcia leukocytów z pełnej krwi, została opisana przez Ebner i in. (patrz Journal of Immunological Methods 252 (2001), 93-104). 20 [0050] Podobnie, allogeniczne leukocyty można również dostarczyć przez wymywanie leukocytów z filtra do usuwania leukocytów, który to filtr był użyty do usuwania leukocy-tów z połączonych kożuszków leukocytarno-płytkowych, gdzie połączone kożuszki leuko-cytarno-płytkowe pochodzą od co najmniej dwóch różnych allogenicznych dawców. Pro-cedura wymywania leukocytów z filtra do ich usuwania, który to filtr był wcześniej uży-25 wany do usunięcia leukocytów z kożuszków leukocytarno-płytkowych, została opisana przez Meyer i in. (patrz Journal of Immunological Methods 307 (2005) 150-166). [0051] Podczas gdy takie allogeniczne leukocyty, otrzymane z filtru do usuwania leukocy-tów, mogą być również stosowane do wytwarzania niewyczerpanych niedojrzałych DC, wydaje się, że korzystne jest stosowanie allogenicznych leukocytów dostarczonych przez 30 połączenie co najmniej dwóch kożuszków leukocytarno-płytkowych, otrzymanych od co najmniej dwóch różnych, allogenicznych dawców. Allogeniczne leukocyty wymyte z filtra do usuwania leukocytów mogą wytwarzać nieco mniejsze ilości chemokin, za wyjątkiem MIG, oraz cytokin, po dojrzewaniu w porównaniu z allogenicznymi monocytami uzyska-nymi bezpośrednio z połączonych próbek krwi obwodowej lub z połączonych kożuszków 35 leukocytarno-płytkowych. [0052] Izolacja monocytów z mieszaniny różnych leukocytów jest dobrze znana w dzie-dzinie. Zgodnie z jedną z postaci, monocyty izoluje się z dostarczonej mieszaniny alloge-nicznych leukocytów znanymi, zgodnymi z GMP sposobami wytwarzania. Zatem, mono-cyty można izolować z dostarczonej mieszaniny allogenicznych leukocytów przez elutria-40 cję lub izolację na przeciwciałach/kulkach. [0053] Elutriacja jest techniką, w której ciągła przeciwprądowa elutriacja rozdziela ko-mórki na wiele frakcji. W skrócie, stałej sile odśrodkowej, która rozdziela komórki na pod-stawie gęstości, przeciwdziała stale rosnący przepływ podłoża przepływającego przez osad, dyspergujący komórki według wielkości. Zatem przepływ podłoża wypłukuje ko-45 mórki na kilka produktów, najpierw najmniejsze/najlżejsze oraz jako ostatnie najwięk-sze/najcięższe. [0054] Izolację monocytów na przeciwciałach/kulkach przeprowadza się z użyciem (Im-muno)-magnetycznego aktywowanego sortowania komórek (MACS). MACS jest szeroko stosowaną techniką selektywnej izolacji komórek z pełnej krwi, kożuszków leukocytarno-50 płytkowych lub WBC z aferezy. W skrócie, przeciwciała specyficzne względem CD niosą-ce ferromagnetyczne kulki na swoim końcu Fc są sprzęgane z komórkami pożądanymi (se-

9
lekcja pozytywna) lub niepożądanymi (selekcja negatywna). Tak traktowane komórki mo-gą być zatrzymane w porowatej, powlekanej metalem kolumnie pod działaniem silnego pola magnetycznego. [0055] Po izolacji monocyty poddaje się różnicowaniu w niedojrzałe DC, tj. wytwarza się niedojrzałe DC. Niedojrzałe DC są wytwarzane przez wspólną hodowlę allogenicznych 5 monocytów w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludz-ka i uzupełnionym czynnikiem stymulującym tworzenie kolonii granulocytów i makrofa-gów (GM-CSF), w połączeniu z interleukiną-4 (IL-4), przez 2 do 7 dni, tak jak około 5 dni, co różnicuje monocyty do niedojrzałych DC. [0056] Ponieważ stwierdzono, że podłoże do hodowli komórek zawierające płodową su-10 rowicę cielęcą wywołuje przedwczesną aktywację pomimo uzupełnienia GM-CSF i IL-4, ważne jest to, aby stosowane podłoże było wolne od surowicy innej niż ludzka. [0057] Niedojrzałe DC można także wytwarzać przez hodowanie allogenicznych monocy-tów w wodnym podłożu zawierającym GM-CSF w połączeniu z interleukiną-2 (IL-2), in-terleukiną-15 (IL-15) lub interferonem alfa przez 2 do 7 dni, tak jak 5 dni, co powoduje 15 różnicowanie monocytów do niedojrzałych DC. Zastosowanie GM-CSF w połączeniu z IL-4 jest jednak korzystne, ponieważ wykazano, że zapobiega on alloreaktywności i przedwczesnej aktywacji, gdy jest stosowany w połączeniu z podłożem wolnym od suro-wicy innej niż ludzka. [0058] Specjalista w dziedzinie jest zaznajomiony z podłożami do hodowli komórek i ich 20 składem. Zazwyczaj stosowane podłoże do hodowli komórek zawiera:
co najmniej jedną sól, taką jak NaCl, KCl, MgSO4 i/lub Ca(NO3)2; co najmniej jeden cukier, taki jak glukoza; jeden lub większą liczbę aminokwasów, takich jak L-metionina, L-fenyloalanina, L-prolina, L-seryna, L-treonina, L-tryptofan, L-tyrozyna, L-walina, L-arginina, L-25 asparagina, L-asparaginowy, L-cystyna, L-glutamina, kwas L-glutaminowy, glicyna, L-histydyna, L-hydroksyprolina, L-izoleucyna, L-leucyna i/lub L-lizyna; jedną lub większą liczbę witamin i innych ważnych substancji odżywczych, takich jak glutation, biotyna, witamina B12, D-pantotenian Ca, chlorek choliny, kwas foliowy, mio-inozytol, amid kwasu nikotynowego, kwas p-aminobenzoesowy, pirydoksyna, ry-30 boflawina i/lub tiamina; oraz co najmniej jeden bufor, taki jak sól fosforanowa (np. Na2HPO4) i/lub sól węglanowa (np. NaHCO3).
[0059] Według jednej postaci, podłoże hodowlane zawiera co najmniej jedną sól, taką jak NaCl, co najmniej jeden cukier, taki jak glukoza, jeden lub większą liczbę aminokwasów, 35 jedną lub większą liczbę witamin oraz bufor, taki jak sól fosforanowa (np. Na2HPO4) i/lub sól węglanowa (np. NaHCO3). [0060] Ponadto, podczas gdy podłoże do hodowli komórek ponadto jest wolne od surowi-cy innej niż ludzka, typowo zawiera ono co najmniej ludzki polipeptyd. Zgodnie z jedną postacią, podłoże do hodowli komórek zawiera co najmniej jeden ludzki polipeptyd wy-40 brany z grupy obejmującej transferynę, albuminę i insulinę; korzystnie, podłoże do hodow-li komórek zawiera wszystkie trzy z nich. Ludzki polipeptyd można uzyskać z ludzkiego osocza. Ponadto mogą być one wytwarzane drogą rekombinacji. Przykładowo, insulina może być wytwarzana drogą rekombinacji w komórkach drożdży. [0061] Przykładowo, podłożem do hodowli komórek wolnym od surowicy innej niż ludz-45 ka może być CellGro®, które jest zgodnym z GMP, wolnym od surowicy podłożem dla komórek dendrytycznych (DC) dostarczonym przez CellGenix GmbH. W Stanach Zjedno-czonych podłoże to jest sprzedawane pod znakiem towarowym CellGenix™. [0062] Jak może stwierdzić specjalista, oraz jak tutaj wyjaśniono, określenie izolowany niekoniecznie odnosi się do 100% czystości, ale monocyty otrzymywane w procesie izola-50 cji wykazują preferencję dla monocytów. Monocyty otrzymane takim sposobem mogą być

10
określane allogenicznymi leukocytami wzbogaconymi w monocyty, ponieważ oprócz mo-nocytów obecne będą inne leukocyty. [0063] Zgodnie z jedną postacią, allogeniczne monocyty są wzbogacone z mieszaniny al-logenicznych leukocytów. Allogeniczne leukocyty wzbogacone w monocyty oprócz mo-nocytów zazwyczaj zawierają również allogeniczne neutrofile. Ponadto, mogą one zawie-5 rać także inne granulocyty. [0064] W przeciwieństwie do powszechnego przekonania, nie zaobserwowano oznak przedwczesnej aktywacji, gdy allogeniczne monocyty hodowano wspólnie w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka oraz uzupełnionym GM-CSF/IL-4, pomimo faktu, że niedojrzałe DC otrzymano z mieszaniny allogenicznych 10 leukocytów. Zgodnie z tym, takie niedojrzałe DC są niewyczerpane, a tym samym są w stanie wytwarzać znaczne ilości, tak jak przykładowo ponad 2000, 5000 lub 7500 pg/ml chemokin prozapalnych, w tym MIP-1 alfa, MIP-1 beta, RANTES i MIG, oraz znaczne ilości, tak jak przykładowo ponad 500, 1500 lub 3000 pg/ml cytokin prozapalnych, w tym IL-12p70 i TNF-alfa w sposób trwały po późniejszym usunięciu czynników aktywujących. 15 [0065] Zgodnie z jedną postacią, niedojrzałe oznacza DC, które eksprymują tylko niski poziom markerów dojrzewania DC CD83 i CD86 oraz, które są zdolne do wytwarzania dużych ilości chemokin i cytokin prozapalnych po aktywacji. Niskie poziomy są takimi, według postaci, które należy interpretować tak, że co najmniej 3-krotny, taki jak co naj-mniej 5-krotny wzrost ekspresji CD83 jest obserwowany po aktywacji oraz, że co najmniej 20 5-krotny, taki jak co najmniej 8-krotny wzrost ekspresji CD86 jest obserwowany po akty-wacji. [0066] Ponieważ przewidywano, że zaobserwuje się przedwczesną aktywację, do niektó-rych doświadczeń dodano inhibitor cykloksoygenazy-2 NS-398, czynnik znany z tego, że utrudnia wyczerpanie aktywowanych DC za pośrednictwem prostaglandyny E2 (PGE2). 25 Jednakże obecność NS-398 w czasie namnażania monocytów do DC nie zwiększyła, ale raczej zmniejszyła wywoływane aktywacją wytwarzanie MIG i IL-12p70. Zatem, brak jest oznak wyczerpania aktywowanych zróżnicowanych niedojrzałych DC za pośrednictwem PGE2 we wspólnych hodowlach mieszanych monocytów allogenicznych. [0067] Jak uzna specjalista (patrz np. EP 1509244 B1 i WO2011/098516) niewyczerpane 30 niedojrzałe komórki dendrytyczne (DC) są przydatne do wytwarzania kompozycji farma-ceutycznej do leczenia nowotworu. Zatem, postać dotyczy mieszaniny allogenicznych niewyczerpanych niedojrzałych komórek dendrytycznych (DC) pochodzących od co naj-mniej dwóch różnych, allogenicznych dawców. Takie komórki dendrytyczne (DC) można uzyskiwać takim sposobem jak tutaj ujawniono. Po zróżnicowaniu do niedojrzałych komó-35 rek DC, niedojrzałe DC mogą być aktywowane, tak aby stały się prozapalnymi DC. Akty-wacja może być wywołana na kilka sposobów. Wykazano, że wiele sygnałów wywołuje co najmniej niektóre aspekty aktywacji DC. Wśród najpotężniejszych z nich są produkty mi-krobiologiczne i wirusowe (wzorce molekularne związane z patogenami (PAMP), które są bezpośrednio rozpoznawane przez receptory rozpoznające wzorce (PRR), obejmujące 40 członków rodziny receptora Toll-podobnego (TLR). PRR kontrolują ekspresję wielu ge-nów odpowiedzi wrodzonej i mogą bezpośrednio sygnalizować aktywację DC. Ponadto, sygnalizacja PRR zarówno w komórkach immunologicznych, jak i nieimmunologicznych często prowadzi do syntezy cytokin zapalnych, takich jak czynnik martwicy nowotworu (TNF) i interleukina 1 (IL-1), które mogą także wspierać aktywację DC. Zatem dodanie 45 cytokin zapalnych może również przyczyniać się do aktywacji niedojrzałych DC. [0068] Zgodnie z pewną postacią, niedojrzałe DC są ładowane antygenami przed, lub równocześnie z, aktywacją, w celu dostarczenia komórkowej allogenicznej szczepionki przeciwnowotworowej. Ładowanie antygenem jest dobrze znane w dziedzinie (patrz np. EP 1 509 244 B1) i można je przeprowadzić takimi sposobami jak podanie impulsu, trans-50 fekcja, infekcja lub fuzja. Przykładowo, antygen można zazwyczaj uzyskiwać z nowotwo-ru; zazwyczaj rodzaju nowotworu, przeciw któremu szczepionka ma być skierowana. W

11
uzyskaniu antygenów, zazwyczaj stosuje się reprezentatywną próbkę rodzaju nowotworu będącego przedmiotem zainteresowania. [0069] Zgodnie z korzystną postacią, aktywacja niedojrzałych DC jest przeprowadzana zgodnie ze sposobem ujawnionym w WO 2011/098516. Dojrzewanie można zatem wywo-łać przez dodanie liganda receptora Toll-podobnego 3 (TLR3) poli I:C, liganda TLR7/8, 5 takiego jak R848 (Rezykwimod) i cytokiny interferonu gamma (IFN-γ). Ligand receptora Toll-podobnego 3 (TLR3) poli I:C jest syntetycznym analogiem dsRNA zawierającym nić poli(I) zhybrydyzowaną z nicią poli(C). Wielkość nici może być różna. Wielkość może wynosić 200 par zasad do 8000 par zasad, przykładowo 200 do 1500 lub 1500 do 8000 par zasad. Ligand TLR7/8 R848 jest również określany w dziedzinie jako Rezykwimod. Jako 10 alternatywę dla Rezykwimodu, jako ligandy TLR7/8 można użyć Gardikwimod lub Imi-kwimod. Zazwyczaj niedojrzałe DC są wystawione na działanie czynników aktywujących przez 8 do 24 godzin, przykładowo 18 godzin. [0070] Aktywacja może ponadto obejmować dodanie co najmniej jednej substancji wy-branej z grupy obejmującej ligandy TLR2, ligandy TLR4, takie jak bakteryjny lipopolisa-15 charyd i monofosforylolipid A, ligandy TLR9, takie jak sekwencje oligonukleotydów CpG (ODN), które odróżniają DNA bakterii od DNA ssaków, interferon alfa (IFN-α), interleu-kinę 1β (IL-1β) i czynnik martwicy nowotworów alfa (TNF-α). Ponadto, aktywacja ko-rzystnie nie obejmuje dodania prostaglandyny E2 (PGE2), w celu zapobieżenia temu by dojrzałe DC stały się migrującymi DC, które szybko opuszczają miejsce wstrzyknięcia 20 (guz), co byłoby niekorzystne w kontekście tego wynalazku. [0071] Po aktywacji, powstałe prozapalne DC mogą być przemywane w celu usunięcia zasadniczo wszystkich czynników aktywujących. Zatem, czynniki aktywujące są zazwy-czaj wymywane przed użyciem prozapalnych DC jako szczepionki. Usuwanie czynników aktywujących pozwala uniknąć jednoczesnego podania czynników aktywujących (prze-25 znaczonych do wywołania prozapalnych DC ex vivo). Jednoczesne podanie czynników ak-tywujących najprawdopodobniej doprowadziłoby do silnej i trwałej aktywacji również re-krutowanych do guza niedojrzałych DC, co prowadziłoby do ich różnicowania w proza-palne dojrzałe DC zamiast pożądanego różnicowania w migrujące dojrzałe DC. [0072] Jak już wcześniej opisano (patrz WO2011/098516), prozapalne komórki dendry-30 tyczne są przydatne w leczeniu nowotworu, ponieważ mogą one aktywować własne DC pacjenta do ich rozwoju w załadowane nowotworowo migrujące DC. Zatem jedna postać dotyczy mieszaniny allogenicznych prozapalnych komórek dendrytycznych pochodzących od co najmniej dwóch różnych allogenicznych dawców. Takie allogeniczne prozapalne komórki dendrytyczne są otrzymywane sposobem takim jak tutaj opisano. Zamrożenie 35 prozapalnych komórek dendrytycznych po aktywacji pozwala na ich przechowywanie. Za-zwyczaj prozapalne komórki dendrytyczne zamraża się w podłożu zawierającym dimetylo-sulfotlenek (DMSO) i surowicę lub osocze. Przed użyciem zamrożone komórki rozmraża się i przemywa DMSO. [0073] Do stosowania w leczeniu nowotworu takie allogeniczne prozapalne komórki den-40 drytyczne można formułować w kompozycję farmaceutyczną. Kompozycja farmaceutycz-na może zawierać co najmniej jeden farmaceutycznie dopuszczalny nośnik, taki jak roz-twór soli buforowany fosforanami, woda i emulsje, takie jak emulsje olej/woda lub wo-da/olej oraz różne typy środków zwilżających. Ponadto może ona zawierać adiuwanty do-puszczalne farmaceutycznie, zaróbki, stabilizatory, środki konserwujące i/lub inne skład-45 niki znane w dziedzinie. Przykładowo, nośnikiem może być roztwór soli fizjologicznej zawierający albuminę surowicy ludzkiej. [0074] Kolejna postać dotyczy takiej mieszaniny allogenicznych prozapalnych komórek dendrytycznych lub takiej kompozycji zawierającej takie allogeniczne prozapalne komórki dendrytyczne do stosowania w leczeniu nowotworu. Podobnie, pewna postać dotyczy za-50 stosowania takiej mieszaniny allogenicznych prozapalnych komórek dendrytycznych do stosowania w wytwarzaniu leku do leczenia nowotworu. Dalsza postać dotyczy sposobu

12
leczenia nowotworu, w którym mieszaninę allogenicznych prozapalnych komórek dendry-tycznych podaje się pacjentowi potrzebującemu takiego leczenia, w dawce skutecznej do aktywowania własnych DC pacjenta do rozwoju w załadowane nowotworowo migrujące DC. [0075] Dalsze ponumerowane postacie wynalazku ujawniono poniżej. 5
1. Sposób wytwarzania niewyczerpanych niedojrzałych komórek dendrytycznych (DC), obejmujący etapy:
- dostarczenia mieszaniny allogenicznych leukocytów, które to allogeniczne leuko-cyty zostały uzyskane od co najmniej dwóch różnych, allogenicznych dawców;
- izolacji allogenicznych monocytów ze wspomnianej mieszaniny allogenicznych 10 leukocytów z dostarczeniem allogenicznych leukocytów wzbogaconych w mono-cyty; i
- wytworzenia niewyczerpanych niedojrzałych DC ze wspomnianych allogenicz-nych leukocytów wzbogaconych w monocyty, przy czym wytworzenie niewy-czerpanych niedojrzałych komórek dendrytycznych (DC) prowadzi się przez 15 wspólną hodowlę wspomnianych allogenicznych leukocytów wzbogaconych w monocyty przez 2 do 7 dni w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka, przy czym wspomniane podłoże jest uzupełnione inter-leukiną-4 (IL-4) oraz czynnikiem stymulującym tworzenie kolonii granulocytów i makrofagów (GM-CSF). 20
2. Sposób według postaci 1, w którym wspomniane podłoże do hodowli komórek zawiera co najmniej ludzki polipeptyd.
3. Sposób według postaci 2, w którym wspomniany ludzki polipeptyd jest wybrany z grupy obejmującej transferynę, albuminę i insulinę.
4. Sposób według którejkolwiek z postaci 1 do 3, w którym wspomniane allogenicz-25 ne leukocyty wzbogacone w monocyty zawierają allogeniczne neutrofile.
5. Postać według którejkolwiek z postaci 1 do 4, w której wspomniana mieszanina al-logenicznych leukocytów jest dostarczana przez łączenie co najmniej dwóch ko-żuszków leukocytarno-płytkowych zawierających leukocyty, które to wspomniane kożuszki leukocytarno-płytkowe do połączenia są otrzymywane od co najmniej 30 dwóch różnych allogenicznych dawców.
6. Sposób według postaci 5, w którym wspomniane połączone kożuszki leukocytar-no-płytkowe zawierają płytki krwi lub są pozbawione płytek krwi.
7. Sposób według którejkolwiek z postaci 1 do 4, w którym wspomniana mieszanina allogenicznych leukocytów jest dostarczana przez: 35 - wymycie leukocytów z co najmniej dwóch filtrów do usuwania leukocytów, któ-
re to filtry, odpowiednio, zostały wcześniej użyte do usunięcia leukocytów z peł-nej krwi, która to pełna krew została uzyskana od co najmniej dwóch różnych al-logenicznych dawców; oraz
- połączenie uzyskanych leukocytów z uzyskaniem wspomnianej mieszaniny allo-40 genicznych leukocytów;
lub przez: - wymycie leukocytów z filtra do usuwania leukocytów, który to filtr został użyty
do usunięcia leukocytów z połączonych kożuszków leukocytarno-płytkowych, gdzie połączone kożuszki leukocytarno-płytkowe pochodzą od co najmniej 45 dwóch różnych allogenicznych dawców.
8. Sposób według którejkolwiek z poprzednich postaci, w którym wspomniane allo-geniczne monocyty izoluje się metodą elutriacji lub przez izolację na przeciwcia-łach/kulkach.
9. Sposób według którejkolwiek z poprzednich postaci, w którym wspomnianą 50 wspólną hodowlę prowadzi się przez około 5 dni.

13
10. Sposób według którejkolwiek z poprzednich postaci, obejmujący ponadto etap ła-dowania niewyczerpanych niedojrzałych DC antygenem.
11. Mieszanina allogenicznych niewyczerpanych niedojrzałych komórek dendrytycz-nych (DC) pochodzących od co najmniej dwóch różnych, allogenicznych dawców, przy czym wspomniane komórki dendrytyczne (DC) są uzyskane sposobem zgod-5 nie z którąkolwiek z postaci 1 do 10.
12. Sposób wytwarzania prozapalnych DC, obejmujący etapy: - dostarczania niewyczerpanych niedojrzałych DC zgodnie z którąkolwiek z posta-
ci 1 do 11; - aktywowania niewyczerpanych niedojrzałych DC dla uzyskania prozapalnych 10
DC. 13. Sposób według postaci 12, w którym wspomnianą aktywację prowadzi się przez
dodanie liganda receptora Toll-podobnego 3 (TLR3) poli I:C, liganda TLR7/8, ta-kiego jak Rezykwimod, i interferonu gamma (IFN-γ) dla wywołania aktywacji.
14. Sposób według postaci 13, w którym wspomniana aktywacja ponadto obejmuje 15 dodanie co najmniej jednej substancji wybranej z grupy obejmującej ligandy TLR2, ligandy TLR4, ligandy TLR9, interferon alfa (IFN-α), interleukinę 1β (IL-1β) i czynnik martwicy nowotworów alfa (TNF-α) dla wywołania aktywacji.
15. Sposób według którejkolwiek z postaci 13 albo 14, w którym wspomniana aktywa-cja nie obejmuje dodania prostaglandyny E2 (PGE2). 20
16. Sposób według którejkolwiek z postaci 13 do 15, w którym niedojrzałe DC są wy-stawione na działanie czynników aktywujących przez 8 do 24 godzin, po czym za-sadniczo wszystkie czynniki aktywujące są wymywane.
17. Mieszanina allogenicznych prozapalnych komórek dendrytycznych pochodzących od co najmniej dwóch różnych, allogenicznych dawców, przy czym wspomniane 25 komórki dendrytyczne są otrzymane sposobem zgodnie z którąkolwiek z postaci 12 do 16.
18. Kompozycja farmaceutyczna zawierająca mieszaninę allogenicznych prozapalnych komórek dendrytycznych zgodnie z postacią 16 i co najmniej jeden farmaceutycz-nie dopuszczalny nośnik. 30
19. Mieszanina allogenicznych prozapalnych komórek dendrytycznych zgodnie z po-stacią 17 albo kompozycja zgodnie z postacią 18, do stosowania do leczenia nowo-tworu.
[0076] Bez dalszego omawiania, uważa się, że specjalista w dziedzinie może, stosując powyższy opis i poniższą część doświadczalną, stosować niniejszy wynalazek w jego naj-35 pełniejszym zakresie. Korzystne, konkretne postacie opisane tutaj należy zatem interpre-tować wyłącznie jako ilustrujące i nie ograniczają one pozostałej części opisu w żaden sposób. Ponadto, chociaż niniejszy wynalazek został opisany powyżej w odniesieniu do konkretnych postaci, to nie ma być on ograniczony do określonej przedstawionej tutaj po-staci. Przeciwnie, wynalazek jest ograniczony jedynie przez załączone zastrzeżenia paten-40 towe, a w zakresie tych załączonych zastrzeżeń patentowych możliwe są również postacie inne niż te konkretne powyższe, np. inne niż te opisane powyżej. [0077] W zastrzeżeniach patentowych określenie „zawiera/zawierający” nie wyklucza obecności innych elementów lub etapów. Ponadto, chociaż poszczególne cechy mogą być włączone w różnych zastrzeżeniach, mogą być ewentualnie korzystnie łączone, a włącze-45 nie w różne zastrzeżenia patentowe nie oznacza, że połączenie cech nie jest wykonalne i/lub korzystne. [0078] Ponadto, odniesienia w liczbie pojedynczej nie wykluczają liczby mnogiej. Odnie-sienia do liczby pojedynczej oraz określenia „pierwszy”, „drugi” itp. nie wykluczają więk-szej liczby. 50

14
Krótki opis rysunków
[0076] Fig. 1 przedstawia ekspresję markerów aktywacji/dojrzewania CD83 i CD86 na nie-dojrzałych DC i PI-DC uzyskanych z pojedynczych lub mieszanych hodowli monocy-tów krwi obwodowej. 5 Fig. 2 przedstawia wytwarzanie prozapalnych chemokin przez niedojrzałe DC (uzy-skane z pojedynczych lub mieszanych hodowli monocytów krwi obwodowej) w ciągu 18 godzin trwałej stymulacji czynnikami aktywującymi („Aktywne wytwarzanie”). Fig. 3 przedstawia wytwarzanie prozapalnych cytokin przez niedojrzałe DC (uzyskane z pojedynczych lub mieszanych hodowli monocytów krwi obwodowej) w ciągu 18 10 godzin trwałej stymulacji czynnikami aktywującymi („Aktywne wytwarzanie”). Fig. 4 przedstawia wytwarzanie prozapalnych cytokin przez niedojrzałe DC (uzyskane z pojedynczych lub mieszanych hodowli monocytów krwi obwodowej) w ciągu 18 godzin trwałej stymulacji czynnikami aktywującymi +/- dodanie inhibitora cykloogy-genazy-2 (Cox-2), NS-398. 15 Fig. 5 przedstawia wytwarzanie prozapalnych chemokin przez niedojrzałe DC (uzy-skane z pojedynczych lub mieszanych hodowli monocytów kożuszka leukocytarno-płytkowego) w ciągu 18 godzin trwałej stymulacji czynnikami aktywującymi („Ak-tywne wytwarzanie”). Fig. 6 przedstawia wytwarzanie prozapalnych cytokin przez niedojrzałe DC (uzyskane 20 z pojedynczych lub mieszanych hodowli monocytów kożuszka leukocytarno-płytko-wego) w ciągu 18 godzin trwałej stymulacji czynnikami aktywującymi („Aktywne wytwarzanie”). Fig. 7 przedstawia wytwarzanie prozapalnych chemokin przez PI-DC (uzyskane z po-jedynczych lub mieszanych hodowli monocytów krwi obwodowej). Te PI-DC prze-25 myto po stymulacji czynnikami aktywującymi przez 18 godzin, a następnie ponownie hodowano przez 24 godziny bez dodania czynników aktywujących („Bierne wytwa-rzanie”). Fig. 8 przedstawia wytwarzanie prozapalnych cytokin przez PI-DC (uzyskane z poje-dynczych lub mieszanych hodowli monocytów krwi obwodowej). Te PI-DC przemyto 30 po stymulacji czynnikami aktywującymi przez 18 godzin, a następnie ponownie ho-dowano przez 24 godziny bez dodania czynników aktywujących („Bierne wytwarza-nie”). Fig. 9 przedstawia wytwarzanie prozapalnych chemokin przez PI-DC (uzyskane z po-jedynczych lub mieszanych hodowli monocytów kożuszka leukocytarno-płytkowego). 35 Te PI-DC przemyto po stymulacji czynnikami aktywującymi przez 18 godzin, a na-stępnie ponownie hodowano przez 24 godziny bez dodania czynników aktywujących („Bierne wytwarzanie”). Fig. 10 przedstawia wytwarzanie prozapalnych cytokin przez PI-DC (uzyskane z po-jedynczych lub mieszanych hodowli monocytów kożuszka leukocytarno-płytkowego). 40 Te PI-DC przemyto po stymulacji czynnikami aktywującymi przez 18 godzin, a na-stępnie ponownie hodowano przez 24 godziny bez dodania czynników aktywujących („Bierne wytwarzanie”). Fig. 11 przedstawia, że mieszane niedojrzałe DC uzyskane z monocytów z filtra wy-twarzają znaczne ilości chemokin prozapalnych w ciągu 18 godzin trwałej stymulacji 45 czynnikami aktywującymi („Aktywne wytwarzanie”). Fig. 12 przedstawia, że mieszane niedojrzałe DC uzyskane z monocytów z filtra wy-twarzają znaczne ilości cytokin prozapalnych w ciągu 18 godzin trwałej stymulacji czynnikami aktywującymi („Aktywne wytwarzanie”). Fig 13 przedstawia, że mieszane PI-DC uzyskane z monocytów z filtra wykazują 50 znaczne wytwarzanie chemokin prozapalnych po usunięciu czynników aktywujących. Te PI-DC przemyto po stymulacji czynnikami aktywującymi przez 18 godzin, a na-

15
stępnie ponownie hodowano przez 24 godziny bez dodatku czynników aktywujących („Bierne wytwarzanie”). Fig. 14 przedstawia, że mieszane PI-DC uzyskane z monocytów z filtra wykazują znaczne wytwarzanie cytokin prozapalnych po usunięciu czynników aktywujących. Te PI-DC przemyto po stymulacji czynnikami aktywującymi przez 18 godzin, a następnie 5 ponownie hodowano przez 24 godziny bez dodatku czynników aktywujących („Bierne wytwarzanie”).
Część doświadczalna
[0080] Poniższe przykłady są jedynie przykładowe i nie powinny być w żaden sposób in-terpretowane jako ograniczenie zakresu wynalazku. Przeciwnie, wynalazek jest ograniczo-10 ny jedynie przez załączone zastrzeżenia.
Leukocyty różnego pochodzenia
Izolacja leukocytów z filtrów do usuwania leukocytów (filtrów TACSI) [0081] Filtry leukocytarne (filtry TACSI do usuwania leukocytów stosowane do rutyno-wego usuwania leukocytów z 4 połączonych kożuszków leukocytarno-płytkowych podczas 15 wytwarzania płytek krwi) zebrano w Component Laboratory at the Department of Transfu-sion Medicine, Sahlgrenska University Hospital, Gothenburg i przetransportowano do la-boratorium (Department of Clinical Immunology, Sahlgrenska University Hospital) w lo-dzie. [0082] W laboratorium strzykawkę (Terumo) wypełniono 50 ml buforu PBS/EDTA (Cli-20 niMACS) i połączono z filtrem TACSI przez złączki typu luer-lock. Filtr przemyto wstecznie do sterylnej kolby szklanej, trzykrotnie (całkowicie 150 ml buforu PBS/EDTA). Wymytą zawiesinę komórkową ostatecznie rozcieńczono PBS (PAA, Fisher Scientific) w stosunku 1:2 w probówce typu Falcon (marka Fisher, Fisher Scientific). Kożuszki leukocytarno-płytkowe 25 [0083] Kożuszki leukocytarno-płytkowe od zdrowych dawców krwi zebrano w Depart-ment of Transfusion Medicine i przetransportowano do laboratorium w temperaturze poko-jowej. Krew obwodowa [0084] Krew obwodową od zdrowych dawców zebrano w Department of Transfusion Me-30 dicine i przetransportowano do laboratorium w temperaturze pokojowej. W laboratorium krew zmieszano z PBS w temperaturze pokojowej w stosunku 1:2 w probówce typu Fal-con. Izolacja komórek jednojądrzastych krwi obwodowej (PBMC) [0085] Krew obwodową od zdrowych dawców zebrano w Department of Transfusion Me-35 dicine i przetransportowano do laboratorium w temperaturze pokojowej. W laboratorium krew zmieszano z PBS w temperaturze pokojowej w stosunku 1:2 w probówce typu Fal-con. Zawiesinę komórek ostrożnie przeniesiono do 10 ml probówek do wirówki (Nunc) zawierających 3 ml Lymphoprep (Axis-Shield). Do każdej probówki przeniesiono 5-6 ml, a następnie odwirowano przy 2000 obr/min, 20 min w temperaturze pokojowej, bez ha-40 mowania. Wyizolowane PBMC przeniesiono do wstępnie schłodzonych 10 ml probówek. Komórki dwukrotnie przemyto przez napełnienie probówek zimnym PBS, a następnie od-wirowano przy 1450 obr/min, 10 minut w 4°C. Supernatanty odrzucono, a osady ponownie przeprowadzono w stan zawiesiny w 1 ml zimnego PBS. Kolejnych 9 ml dodano do każdej probówki. 45
Izolacja monocytów [0086] 5 ml frakcji wymytych leukocytów z filtra/leukocytów z kożuszka leukocytarno-płytkowego lub wyizolowanych PBMC z 10-20 ml pełnej krwi obwodowej odwirowano w probówkach przy 1450 obr/min, 10 minut w 4°C. Supernatanty całkowicie usunięto, a osa-dy komórek ponownie przeprowadzono w stan zawiesiny w 80 µl PBS/EDTA (Miltenyi) 50

16
na 107 komórek. Dodano po 20 µl mikrokulek CD14 (Miltenyi) na 107 komórek. Komórki zmieszano i inkubowano przez 15 minut w 4°C, a następnie przemyto przez dodanie 1-2 ml PBS/EDTA, a następnie wirowanie przy 300xg przez 10 minut. Supernatanty całkowi-cie usunięto i pozostałe komórki ponownie przeprowadzono w stan zawiesiny w 500 µl PBS/EDTA. 5 [0087] Separatory MidiMACS (Miltenyi) umieszczono w wielostanowiskowym statywie magnetycznym (Miltenyi) i przemyto 3 ml PBS/EDTA. Zawiesiny komórkowe umiesz-czono na separatorach MidiMACS umożliwiając przejście komórek. Separatory Midi-MACS przemyto trzykrotnie 3 ml PBS/EDTA. Wypływające frakcje nieznakowanych ko-mórek odrzucono. Separatory MidiMACS wyjęto z wielostanowiskowego statywu magne-10 tycznego i umieszczono w probówce typu Falcon. Odpipetowano 5 ml buforu PBS/EDTA na kolumnę i komórki natychmiast przepchano tłokiem. [0088] Stężenie komórek wyznaczono w komorze Bürkera. Zawiesiny komórek zawiera-jące monocyty odwirowano przy 1450 obr/min, 10 minut w 4°C. Supernatanty odrzucono, a komórki ponownie przeprowadzono w stan zawiesiny w podłożu CellGro DC (CellGe-15 nix). Czystość monocytów CD 14+ we wszystkich hodowlach komórkowych izolowanych monocytów wynosiła > 80%, co wyznaczono metodą analizy FACS, patrz poniżej.
Wytwarzanie niedojrzałych DC [0089] Leukocyty pochodzące z filtrów TACSI ponownie przeprowadzono w stan zawie-siny przy stężeniu 300000 komórek/ml w podłożu CellGro DC, które jest podłożem wol-20 nym od surowicy innej niż ludzka i wysiano do płytek 24-studzienkowych (1 ml na stu-dzienkę). Leukocyty wzbogacone w monocyty z kożuszków leukocytarno-płytkowych i krwi obwodowej najpierw ponownie przeprowadzono w stan zawiesiny do stężenia 5 x 105 monocytów/ml w podłożu CellGro. Najpierw do 12 studzienek (A1-6, B1-3, C1-3) w płyt-ce 24-studzienkowej dodano 400 µl podłoża CellGro (bez komórek). Do studzienki A1-3 25 przeniesiono 600 µl zawiesiny komórek wzbogaconej w monocyty od dawcy A (odpo-wiednio kożuszek leukocytarno-płytkowy lub krew obwodowa). Do studzienki B1-3 prze-niesiono 600 µl zawiesiny komórek wzbogaconej w monocyty od dawcy B (kożuszek leu-kocytarno-płytkowy lub krew obwodowa). Do studzienki C1-3 przeniesiono 600 µl zawie-siny komórek wzbogaconej w monocyty od dawcy C (kożuszek leukocytarno-płytkowy 30 lub krew obwodowa). Do studzienki A4-6 przeniesiono 200 µl zawiesiny komórek wzbo-gaconej w monocyty od wszystkich trzech dawców (kożuszek leukocytarno-płytkowy lub krew obwodowa). Ostateczna liczba komórek we wszystkich studzienkach wynosiła 300000 komórek (w objętości 1 ml podłoża CellGro na studzienkę). [0090] W celu różnicowania monocytów do niedojrzałych DC, podłoże hodowlane uzu-35 pełniono 1000 U/ml rekombinowanej ludzkiej IL-4 i 1000 U/ml rekombinowanego ludz-kiego GM-CSF (wszystkie z CellGenix, Freiburg, Niemcy) i komórki następnie hodowano przez 5 dni.
Aktywacja/dojrzewanie niedojrzałych DC [0091] Po 5 dniach hodowli w podłożu CellGro uzupełnionym IL-4 i GM-CSF, aktywa-40 cję/dojrzewanie niedojrzałych DC wywołano przez dodanie 20 µg/ml poliI:C (Sigma, Ste-inheim, Niemcy), środka immunostymulacyjnego specyficznego dla receptora TLR-3, znanego również jako kwas poliinozyno:policytydylowy lub sól sodowa kwasu poliinozy-no-policytydylowego, 2,5 μg/ml R848 (Sigma, Steinheim, Niemcy), ligand receptora Toll-podobnego 7/8, znany również jako rezykwimod, i 1000 U/ml interferonu gamma (IFN-γ, 45 R&D Systems, Minneapolis, USA). Po 18 h inkubacji, komórki przemyto trzykrotnie, a następnie inkubowano w świeżym podłożu AIM-V (bez dodatku egzogennych czynników aktywujących) przez 24 h. Supernatanty z hodowli zbierano zgodnie z protokołami dobrze znanymi specjalistom w dziedzinie. [0092] Analizę ELISA przeprowadzono na supernatantach jak opisano poniżej, w celu 50 analizy poziomów chemokin prozapalnych i cytokin prozapalnych.

17
Ocena poziomów chemokin prozapalnych i cytokin prozapalnych metodą ELISA [0093] Chemokiny prozapalne CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES i CXCL9/MIG i cytokiny prozapalne IL-12p70 i TNF-α zmierzono testem immunoenzyma-tycznym (ELISA) z użyciem Duo Set ELISA Development System z R&D systems, Min-neapolis, USA zgodnie z instrukcjami producenta. 5
Analiza fenotypowa metodą cytometrii przepływowej [0094] Monocyty i DC uzyskane z monocytów wytworzono w sposób opisany powyżej. Częstość występowania monocytów CD14+ po izolacji monocytów oceniono przez wy-barwienie komórek FITC-przeciwciałem przeciw ludzkiemu CD14. Po 5 dniach inkubacji w CellGro z dodatkiem IL-4 i GM-CSF niedojrzałe DC przemyto, a następnie wybarwiono 10 PE-przeciwciałem przeciw ludzkiemu CD86 w połączeniu z FITC-przeciwciałem przeciw ludzkiemu CD83. Niedojrzałe DC, które następnie aktywowano przez 18 godzin czynni-kami aktywującymi również wybarwiono PE-przeciwciałem przeciw ludzkiemu CD86 w połączeniu z FITC-przeciwciałem przeciw ludzkiemu CD83. Mysie IgG1 i IgG2 wybar-wione FITC i PE zastosowano jako kontrolę izotypową (wszystkie z BD Biosciences, Cali-15 fornia, USA). Próbki analizowano metodą cytometrii przepływowej (FACS), stosując oprogramowanie Cell Quest (BD Bioscience, California, USA).
Wyniki [0095] Poniżej omówiono wyniki z części doświadczalnej. [0096] DC uzyskane ze wspólnych hodowli allogenicznych leukocytów wzbogaconych w 20 monocyty nie ulegają fenotypowej aktywacji/dojrzewaniu, gdy są hodowane wspólnie w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka i uzupełnionym GM-CSF i IL-4 [0097] Namnażanie monocytów od pojedynczych dawców krwi w podłożu do hodowli komórek wolnym od surowicy innej niż ludzka i uzupełnionym GM-CSF i IL-4 przez 4-7 25 dni spowodowało wytworzenie niewyczerpanych DC o typowym „niedojrzałym” fenoty-pie, włącznie z niską ekspresją markera dojrzewania CD83 i niską ekspresją cząsteczki ko-stymulującej CD86. Jak widać na Figurze 1 a i b średnia ekspresja zarówno CD83, jak i CD86 dla 3 różnych „pojedynczych” DC była podobna w porównaniu z ekspresją CD83 (Fig. 1a) i CD86 (Fig. 1b) na DC uzyskanych z mieszaniny od wszystkich trzech dawców. 30 Jak widać na Figurze 1c i d silnie zwiększona średnia ekspresja markerów aktywa-cji/dojrzewania CD83 i CD86 dla „pojedynczych” DC (DC od 3 różnych analizowanych dawców krwi obwodowej) po namnożeniu w wodnym podłożu do hodowli komórek wol-nym od surowicy innej niż ludzka i uzupełnionym GM-CSF i IL-4 przez 4 dni, a następnie trwałej aktywacji czynnikami stymulującymi przez 18 godzin była podobna w porównaniu 35 z ekspresją CD83 (Fig. 1c) i CD86 (Fig. 1d) na aktywowanych DC uzyskanych z miesza-niny allogenicznych leukocytów wzbogaconych w monocyty od wszystkich trzech daw-ców. [0098] Podsumowując, wyniki te wskazują, że DC uzyskane z monocytów z mieszanej al-logenicznej populacji monocytów są niedojrzałe po hodowli w GM-CSF i IL-4 przez 5 dni, 40 a zatem nie doświadczyły żadnych sygnałów aktywacji/dojrzewania podczas ich różnico-wania z monocytów do niedojrzałych DC. Ponadto, niedojrzałe DC z mieszanej alloge-nicznej populacji monocytów są co najmniej fenotypowo niewyczerpane, ponieważ silnie odpowiadają one fenotypowym dojrzewaniem po stymulacji czynnikami aktywującymi. [0099] Dane uzyskane metodą cytometrii przepływowej. Odpowiednia oś Y przedstawia 45 średnią intensywność fluorescencji (MFI) dla CD83 i CD86 przed i po trwałej stymulacji czynnikami aktywującymi przez 18 godzin. Na osi x przedstawiono różne zmierzone połą-czenia. [0100] Niedojrzałe DC uzyskane ze wspólnych hodowli mieszanych allogenicznych mono-cytów krwi obwodowej nie są funkcjonalnie wyczerpane. 50

18
[0101] Wiadomo, że namnażanie monocytów (od pojedynczego dawcy krwi) w podłożu hodowlanym uzupełnionym GM-CSF i IL-4 przez 4-7 dni powoduje wytworzenie niewy-czerpanych DC, które odpowiadają silnym wytwarzaniem chemokin prozapalnych (MIP-1 alfa, MIP-1 beta, RANTES i MIG) i cytokin prozapalnych (IL-12p70 i TNF-alfa) po sty-mulacji pewnymi czynnikami aktywującymi. 5 [0102] Jak widać na Figurze 2, wysokie średnie poziomy MIP-1 alfa (Fig. 2a), MIP-1 beta (Fig. 2b), RANTES (Fig. 2c), MIG (Fig. 2d) wytworzone przez „pojedyncze” DC (DC od trzech różnych analizowanych dawców krwi obwodowej) podczas trwałej aktywacji czyn-nikami stymulującymi przez 18 godzin były podobne w porównaniu z DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców. Warto zauważyć, że istnieje znacz-10 na zmienność w wywołanym aktywacją wytwarzaniu chemokin między różnymi DC od pojedynczych dawców. Jak widać na Figurze 4, wysokie średnie poziomy IL-12p70 (Fig. 3a) i TNF-alfa (Fig. 3b) wytworzone przez aktywowane „pojedyncze” DC były podobne w porównaniu z DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu IL-12p70 i TNF-alfa mię-15 dzy różnymi DC od pojedynczych dawców. [0103] Dane uzyskano z analizy ELISA. Przedstawione wyniki są wartościami średnimi ± SD dla trzech osobników i wartością uzyskaną dla mieszaniny od wszystkich trzech daw-ców. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek przez 18 godzin trwałej stymulacji/aktywacji. Na osi x przedsta-20 wiono różne zmierzone połączenia. [0104] Zasugerowano, że prostaglandyna E2 (PGE2) odgrywa centralną rolę w wywoła-nym aktywacją wyczerpaniu niedojrzałych DC (Rieser C i in., Differential Deactivation of Human Dendritic Cells by Endotoxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117). W związku z tym zbadano czy doda-25 nie inhibitora Cox-2 NS-398 (skierowanego do zahamowania potencjalnego wytwarzania PGE2) podczas wspólnego hodowania allogenicznych monocytów, zwiększy wytwarzanie chemokin prozapalnych (reprezentowane przez wytwarzanie MIG) lub cytokin prozapal-nych (reprezentowane przez wytwarzanie IL-12p70) podczas kolejnej aktywacji. Jak widać na Figurze 4, obecność inhibitora Cox-2 NS-398 podczas namnażania monocytów do DC 30 nie zwiększyła, ale raczej zmniejszyła wywoływane aktywacją wytwarzanie MIG i IL-12p70. Zatem, brak jest oznak wyczerpania za pośrednictwem PGE2 zróżnicowanych nie-dojrzałych DC ze wspólnych hodowli mieszanych allogenicznych monocytów. [0105] Dane uzyskano z analizy ELISA. Przedstawione wyniki pochodzą z jednego do-świadczenia z mieszaniny od wszystkich trzech dawców. Odpowiednia oś Y przedstawia 35 ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek, w ciągu 18 go-dzin trwałej stymulacji/aktywacji. Na osi x przedstawiono różne zmierzone połączenia. [0106] Niedojrzałe DC uzyskane ze wspólnych hodowli mieszanych allogenicznych leuko-cytów z kożuszka leukocytarno-płytkowego wzbogaconych w monocyty nie są funkcjonalnie wyczerpane. 40 [0107] Jak widać na Figurze 5, wysokie średnie poziomy wywołanych aktywacją chemo-kin prozapalnych MIP-1 alfa (Fig. 5a), MIP-1 beta (Fig. 5b), RANTES (Fig. 5c), MIG (Fig. 5d) wytworzone przez „pojedyncze” DC (DC od trzech różnych analizowanych daw-ców kożuszka leukocytarno-płytkowego) podczas trwałej aktywacji czynnikami stymulu-jącymi przez 18 godzin były podobne w porównaniu z DC uzyskanymi z mieszaniny mo-45 nocytów od wszystkich trzech dawców. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu chemokin między różnymi DC od pojedynczych dawców. [0108] Jak widać na Figurze 6, wysokie, wywołane aktywacją, średnie poziomy IL-12p70 (Fig. 6a) i TNF-alfa (Fig. 6b) wytwarzanych przez „pojedyncze” DC były podobne w po-równaniu z DC uzyskanymi z mieszaniny leukocytów wzbogaconych w monocyty od 50 wszystkich trzech dawców. Warto zauważyć, że istnieje znaczna zmienność w wytwarza-niu IL-12p70 i TNF-alfa między różnymi DC od pojedynczych dawców.

19
[0109] Dane uzyskano z analizy ELISA. Przedstawione wyniki są wartościami średnimi ± SD dla trzech osobników i wartością uzyskaną dla mieszaniny od wszystkich trzech daw-ców. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek przez 18 godzin trwałej stymulacji/aktywacji. Na osi x przedsta-wiono różne zmierzone połączenia. 5 PI-DC uzyskane ze wspólnych hodowli mieszanych allogenicznych monocytów krwi obwo-dowej wykazują trwałe wytwarzanie chemokin i cytokin prozapalnych [0110] W celu wstrzyknięcia aktywowanych prozapalnych DC (PI-DC) pacjentom, za-zwyczaj muszą być one przemyte przed podaniem. Jeżeli tego się nie przeprowadzi, mogą wystąpić niepożądane skutki uboczne wywołane przez równoczesne podanie środków 10 stymulujących (skierowanych do wywołania PI-DC ex vivo). Zatem, niedojrzałe DC muszą być aktywowane do PI-DC o przedłużonym wytwarzaniu pożądanych czynników, także po usunięciu czynników wywołujących aktywację. Jak widać na Figurze 7, średnie poziomy MIP-1 alfa (Fig. 7a), MIP-1 beta (Fig. 7b), RANTES (Fig. 7c), MIG (Fig. 7d) wytwarza-nych przez „pojedyncze” PI-DC po usunięciu czynników aktywujących (PI-DC z monocy-15 tów krwi obwodowej od trzech różnych analizowanych dawców) były podobne w porów-naniu z PI-DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców krwi obwodowej. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu chemokin między różnymi PI-DC od pojedynczych dawców po usunięciu czynników aktywujących. Średnie wytwarzanie IL-12p70 (Fig. 8a) i TNF-alfa (Fig. 8b) wytwarzanych przez „poje-20 dyncze” PI-DC po usunięciu czynników aktywujących było podobne w porównaniu z przemytymi PI-DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu cytokin między różnymi PI-DC od pojedynczych dawców po usunięciu czynników aktywujących. [0111] Dane uzyskano z analizy ELISA. Przedstawione wyniki są wartościami średnimi ± 25 SD dla trzech osobników i wartością uzyskaną dla mieszaniny od wszystkich trzech daw-ców. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek podczas 24 godzin po usunięciu czynników aktywujących. Na osi x przedstawiono różne zmierzone połączenia. PI-DC uzyskane ze wspólnych hodowli mieszanych allogenicznych obwodowych leukocy-30 tów kożuszka leukocytarno-płytkowego wzbogaconych w monocyty wykazują trwałe silne wytwarzanie chemokin i cytokin prozapalnych [0112] Jak przedstawiono na Figurze 9, średni poziom MIP-1 alfa (Fig. 9a), MIP-1 beta (Fig. 9b), RANTES (Fig. 9c), MIG (Fig 9d) wytwarzanych przez „pojedyncze” PI-DC po usunięciu czynników aktywujących (PI-DC z monocytów z kożuszka leukocytarno-35 płytkowego od trzech różnych analizowanych dawców) był podobny w porównaniu z PI-DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców kożuszków leu-kocytarno-płytkowych. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu chemokin między różnymi PI-DC od pojedynczych dawców. Średnie wytwarzanie IL-12p70 (Fig. 10a) i TNF-alfa (Fig. 10b) wytwarzanych przez „pojedyncze” PI-DC po usu-40 nięciu czynników aktywujących było również podobne w porównaniu z przemytymi PI-DC uzyskanymi z mieszaniny monocytów od wszystkich trzech dawców kożuszków leu-kocytarno-płytkowych. Warto zauważyć, że istnieje znaczna zmienność w wytwarzaniu cytokin między różnymi PI-DC od pojedynczych dawców. [0113] Dane uzyskano z analizy ELISA. Przedstawione wyniki są wartościami średnimi ± 45 SD dla trzech osobników i wartością uzyskaną dla mieszaniny od wszystkich trzech daw-ców. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek podczas 24 godzin po usunięciu czynników aktywujących. Na osi x przedstawiono różne zmierzone połączenia. Mieszane niedojrzałe DC uzyskane z leukocytów z filtra wzbogaconych w monocyty wy-50 twarzają znaczne ilości chemokin i cytokin prozapalnych po aktywacji

20
[0114] Jak widać na Figurze 11, aktywowane mieszane DC uzyskane z leukocytów z filtra wzbogaconych w monocyty (początkową populację leukocytów wymyto z filtra do usuwa-nia leukocytów z 4 kożuszków leukocytarno-płytkowych) wytwarzały znaczne ilości MIP-1 alfa (Fig. 11a), MIP-1 beta (Fig. 11b), RANTES (Fig. 11c), MIG (Fig. 11d). Jak widać na Figurze 12, została również wytworzona znaczna ilość IL-12p70 (Fig. 12a) i TNF-alfa 5 (Fig. 12b). [0115] Dane uzyskano z analizy ELISA. Przedstawione wyniki są wartościami z jednego doświadczenia. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych sub-stancji w pg/ml/1 x 106 komórek podczas 18 godzin trwałej stymulacji/aktywacji. Mieszane PI-DC uzyskane z leukocytów z filtra wzbogaconych w monocyty wykazują 10 znaczne wytwarzanie chemokin i cytokin prozapalnych po usunięciu czynników aktywują-cych [0116] Jak widać na Figurze 13, aktywowane mieszane DC uzyskane z monocytów z filtra (początkową populację leukocytów wymyto z filtra do usuwania leukocytów z 4 kożusz-ków leukocytarno-płytkowych) wytwarzają znaczne ilości MIP-1 alfa (Fig. 13a), MIP-1 15 beta (Fig. 13b), RANTES (Fig. 13c), MIG (Fig. 13d) po usunięciu czynników aktywują-cych. Jak widać na Figurze 14, została również wytworzona znaczna ilość IL-12p70 (Fig. 14a) i TNF-alfa (Fig. 14b). Dane uzyskano z analizy ELISA. Przedstawione wyniki są war-tościami z jednego doświadczenia. Odpowiednia oś Y przedstawia ilość poszczególnych wytworzonych substancji w pg/ml/1 x 106 komórek podczas 24 godzin po usunięciu czyn-20 ników aktywujących. Zastrzeżenia patentowe 1. Sposób wytwarzania prozapalnych komórek dendrytycznych (DC), obejmujący etapy:
- dostarczenia mieszaniny allogenicznych leukocytów, które to allogeniczne leu-kocyty zostały uzyskane od co najmniej dwóch różnych, allogenicznych dawców; - izolacji allogenicznych monocytów ze wspomnianej mieszaniny allogenicznych 25 leukocytów z dostarczeniem allogenicznych leukocytów wzbogaconych w mono-cyty; - wytworzenia niewyczerpanych niedojrzałych DC ze wspomnianych allogenicz-nych leukocytów wzbogaconych w monocyty, przy czym wytworzenie niewy-czerpanych niedojrzałych DC prowadzi się przez wspólną hodowlę wspomnia-30 nych allogenicznych leukocytów wzbogaconych w monocyty przez 2 do 7 dni w wodnym podłożu do hodowli komórek wolnym od surowicy innej niż ludzka, przy czym wspomniane podłoże jest uzupełnione interleukiną-4 (IL-4) oraz czyn-nikiem stymulującym tworzenie kolonii granulocytów i makrofagów (GM-CSF); i aktywowania niewyczerpanych niedojrzałych DC dla uzyskania prozapalnych 35 DC.
2. Sposób według zastrzeżenia 1, w którym wspomniane podłoże do hodowli komórek zawiera co najmniej jeden ludzki polipeptyd wybrany z grupy obejmującej transfery-nę, albuminę i insulinę.
3. Sposób według któregokolwiek z zastrzeżeń 1 albo 2, w którym wspomniane alloge-40 niczne leukocyty wzbogacone w monocyty zawierają allogeniczne neutrofile.
4. Sposób według któregokolwiek z zastrzeżeń 1 do 3, w którym wspomniana mieszani-na allogenicznych leukocytów jest dostarczana przez łączenie co najmniej dwóch ko-żuszków leukocytarno-płytkowych zawierających leukocyty, które to wspomniane ko-żuszki leukocytarno-płytkowe do połączenia są otrzymywane od co najmniej dwóch 45 różnych allogenicznych dawców.
5. Sposób według któregokolwiek z zastrzeżeń 1 do 3, w którym wspomniana mieszani-na allogenicznych leukocytów jest dostarczana przez:

21
- wymycie leukocytów z co najmniej dwóch filtrów do usuwania leukocytów, któ-re to filtry, odpowiednio, zostały wcześniej użyte do usunięcia leukocytów z peł-nej krwi, która to pełna krew została uzyskana od co najmniej dwóch różnych al-logenicznych dawców; oraz - połączenie uzyskanych leukocytów z uzyskaniem wspomnianej mieszaniny al-5 logenicznych leukocytów;
lub przez: - wymycie leukocytów z filtra do usuwania leukocytów, który to filtr został użyty do usunięcia leukocytów z połączonych kożuszków leukocytarno-płytkowych, gdzie połączone kożuszki leukocytarno-płytkowe pochodzą od co najmniej dwóch 10 różnych allogenicznych dawców.
6. Sposób według któregokolwiek z poprzednich zastrzeżeń, w którym wspomniane al-logeniczne monocyty izoluje się metodą elutriacji lub przez izolację na przeciwcia-łach/kulkach.
7. Sposób według któregokolwiek z poprzednich zastrzeżeń, w którym wspomnianą 15 wspólną hodowlę prowadzi się przez około 5 dni.
8. Sposób według któregokolwiek z poprzednich zastrzeżeń, obejmujący ponadto etap ładowania niewyczerpanych niedojrzałych DC antygenem.
9. Sposób według któregokolwiek z poprzednich zastrzeżeń, w którym wspomnianą ak-tywację prowadzi się przez dodanie liganda receptora Toll-podobnego 3 (TLR3) poli 20 I:C, liganda TLR7/8, liganda TLR7/8 wybranego z grupy obejmującej Rezykwimod, Gardikwimod lub Imikwimod, i interferonu gamma (IFN-γ) dla wywołania aktywacji.
10. Sposób według zastrzeżenia 9, w którym wspomniana aktywacja ponadto obejmuje dodanie co najmniej jednej substancji wybranej z grupy obejmującej ligandy TLR4 wybrane z grupy obejmującej lipopolisacharyd i monofosforylolipid A, sekwencje 25 oligonukleotydów CpG (ODN), które odróżniają DNA bakterii od DNA ssaków, in-terferon alfa (IFN-α), interleukinę 1β (IL-1β) i czynnik martwicy nowotworów alfa (TNF-α).
11. Sposób według któregokolwiek z zastrzeżeń 9 albo 10, w którym wspomniana aktywa-cja nie obejmuje dodania prostaglandyny E2 (PGE2). 30
12. Sposób według któregokolwiek z zastrzeżeń 9 do 11, w którym niedojrzałe DC są wy-stawione na działanie czynników aktywujących przez 8 do 24 godzin, po czym zasad-niczo wszystkie czynniki aktywujące są wymywane.
13. Kompozycja farmaceutyczna zawierająca mieszaninę allogenicznych prozapalnych komórek dendrytycznych pochodzących od co najmniej dwóch różnych, allogenicz-35 nych dawców, przy czym wspomniane komórki dendrytyczne są możliwe do otrzy-mania sposobem według któregokolwiek z zastrzeżeń 1 do 12, i co najmniej jeden farmaceutycznie dopuszczalny nośnik.
14. Kompozycja według zastrzeżenia 13, do stosowania do leczenia nowotworu. Uprawniony: Immunicum AB Pełnomocnik: dr inż. Wojciech Tykarski Rzecznik patentowy

22
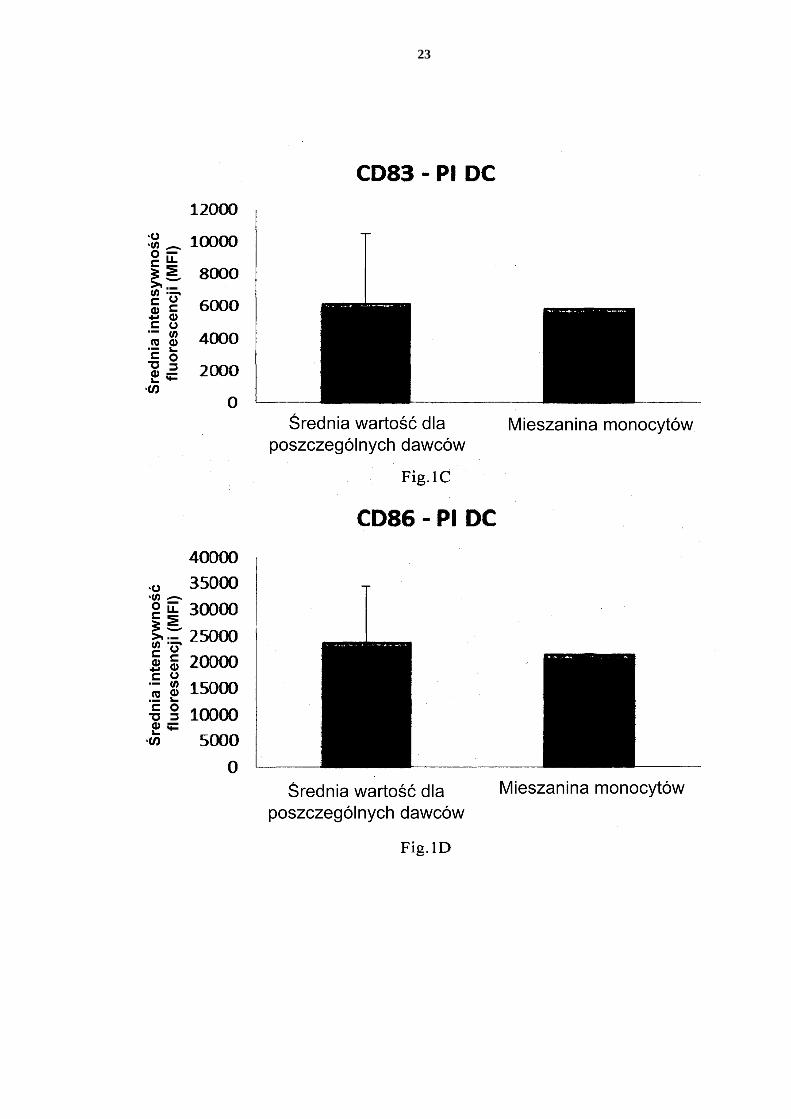
23

24

25
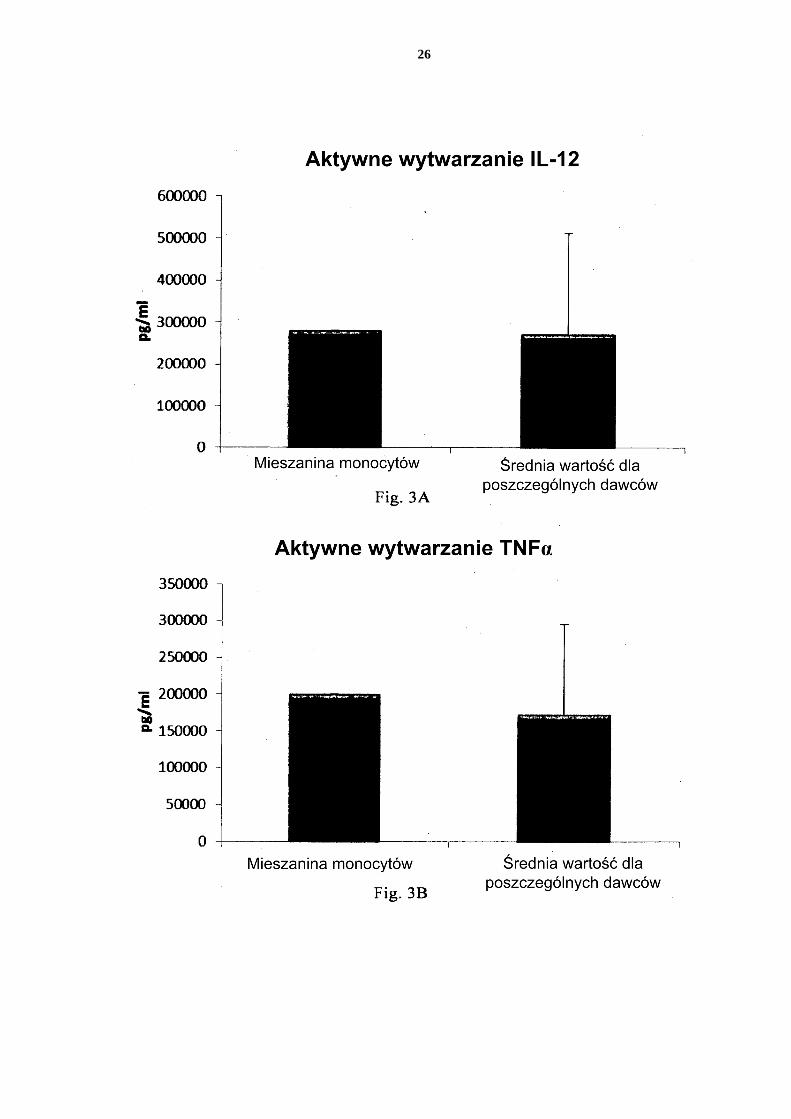
26
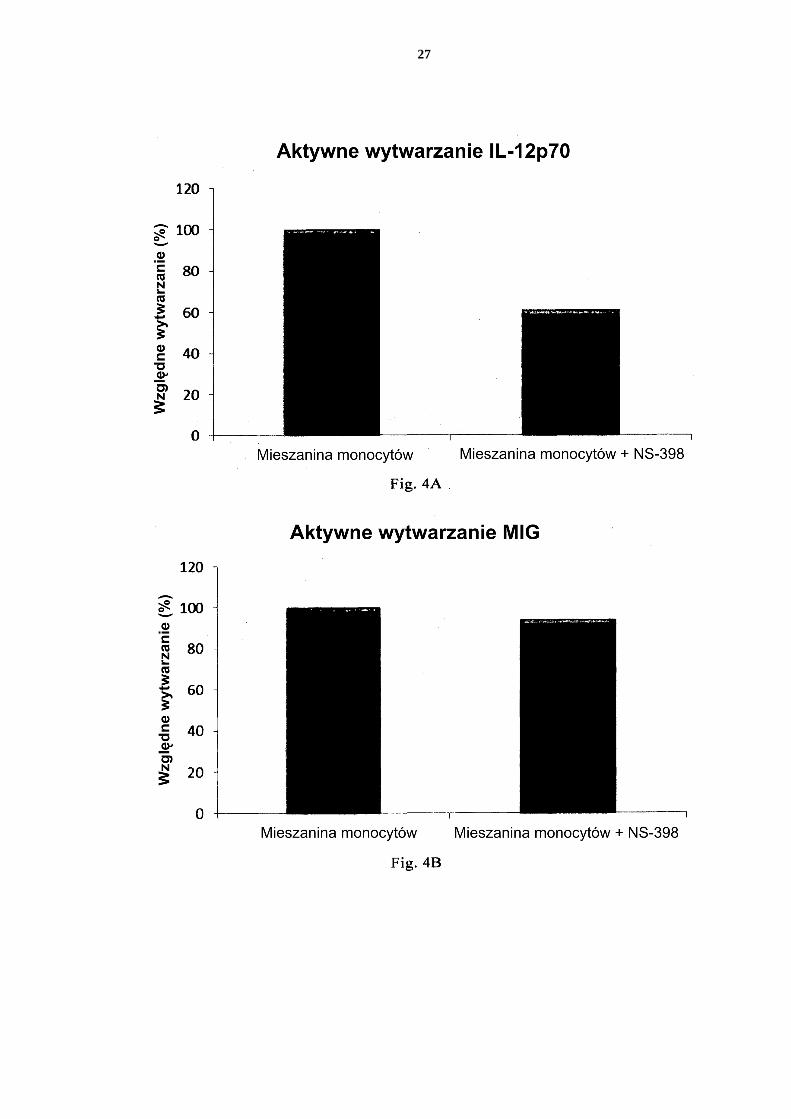
27
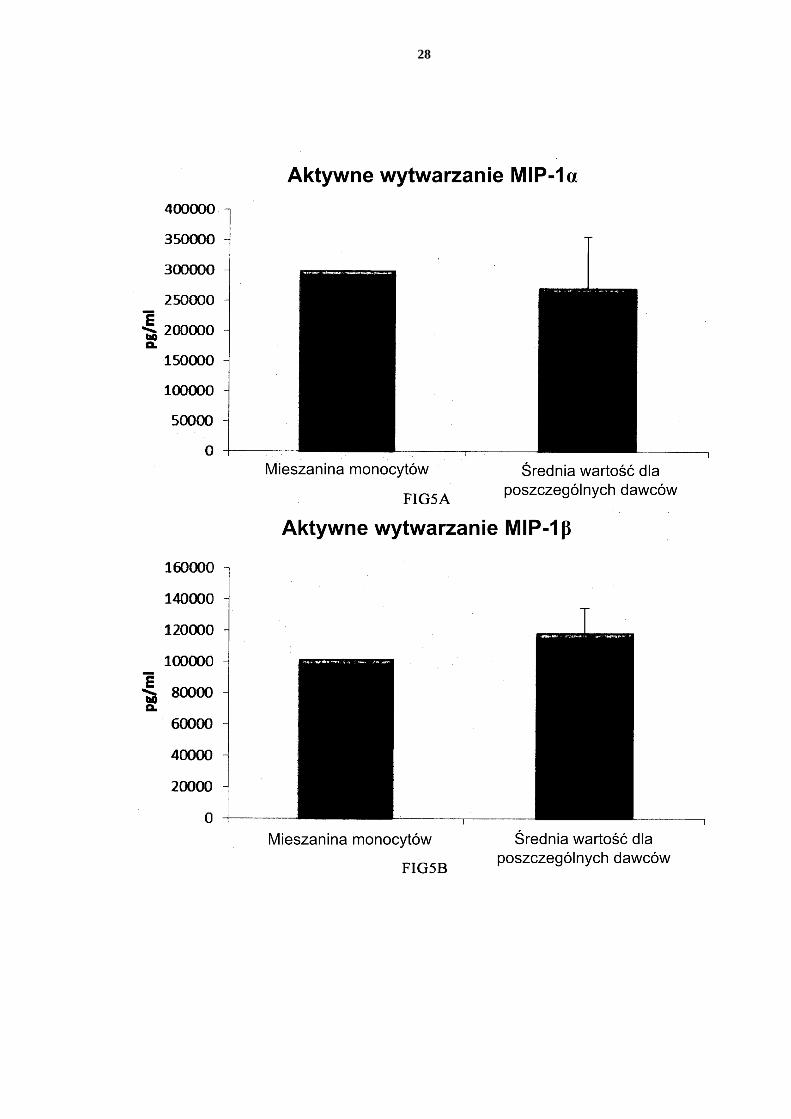
28

29
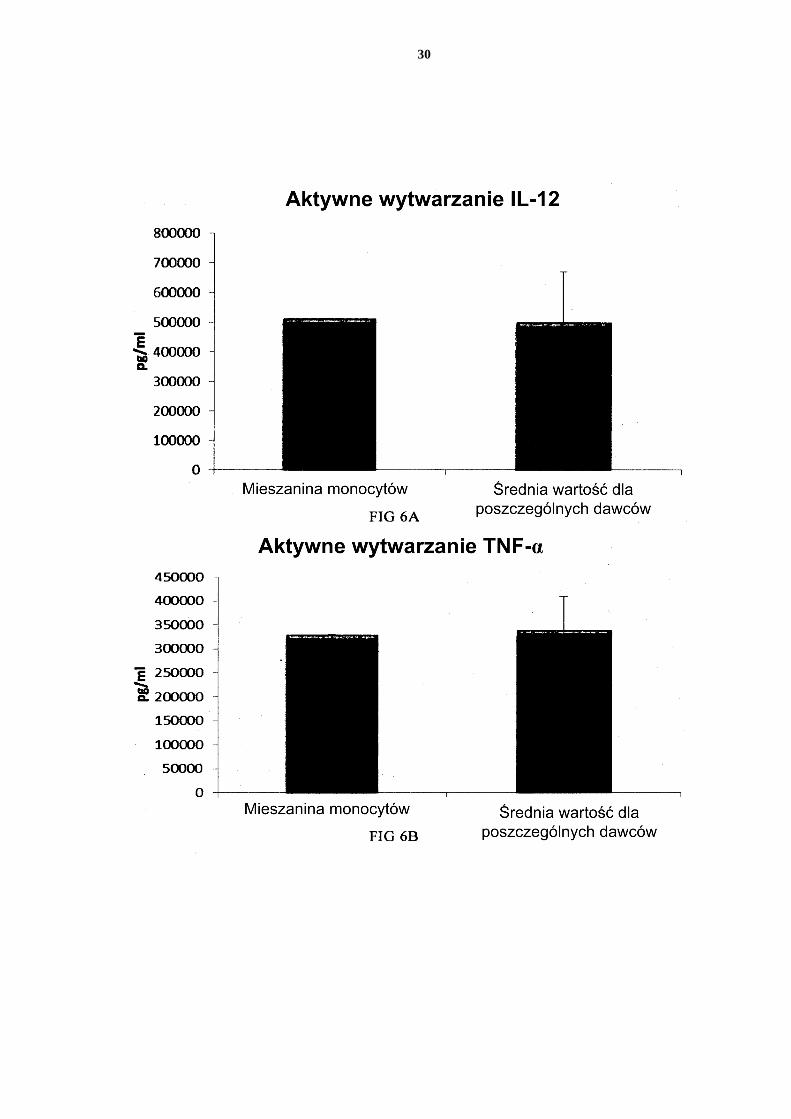
30

31
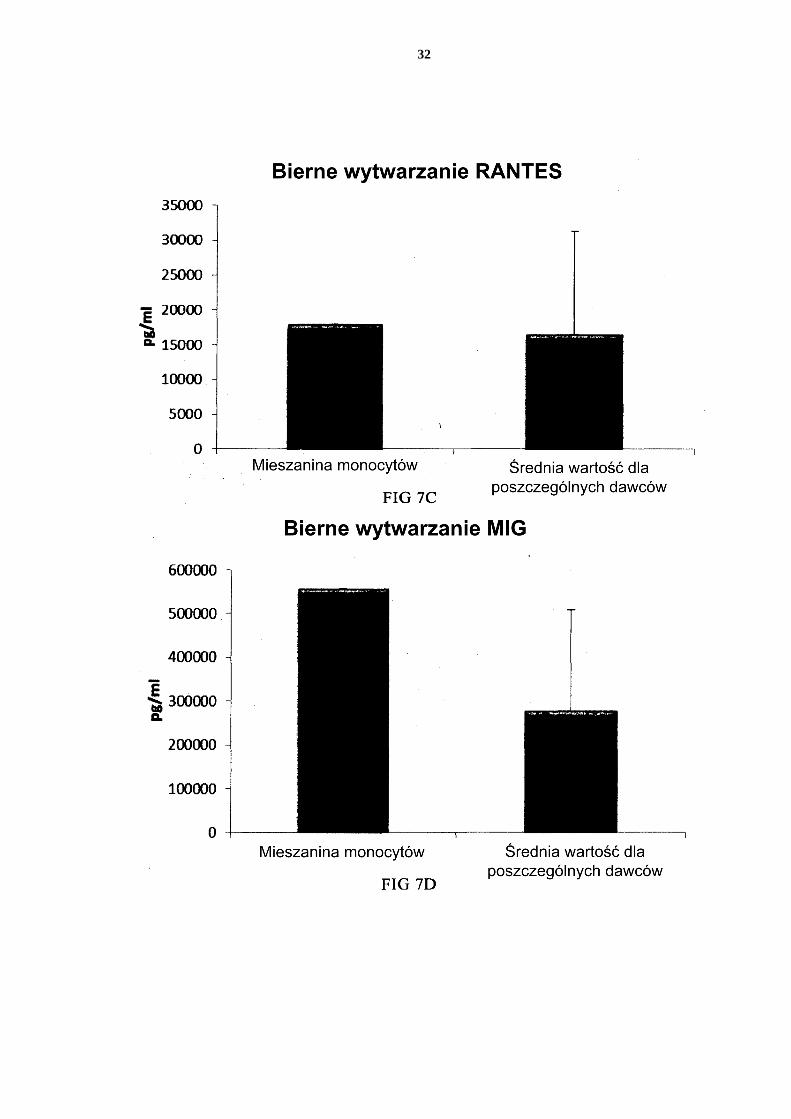
32

33
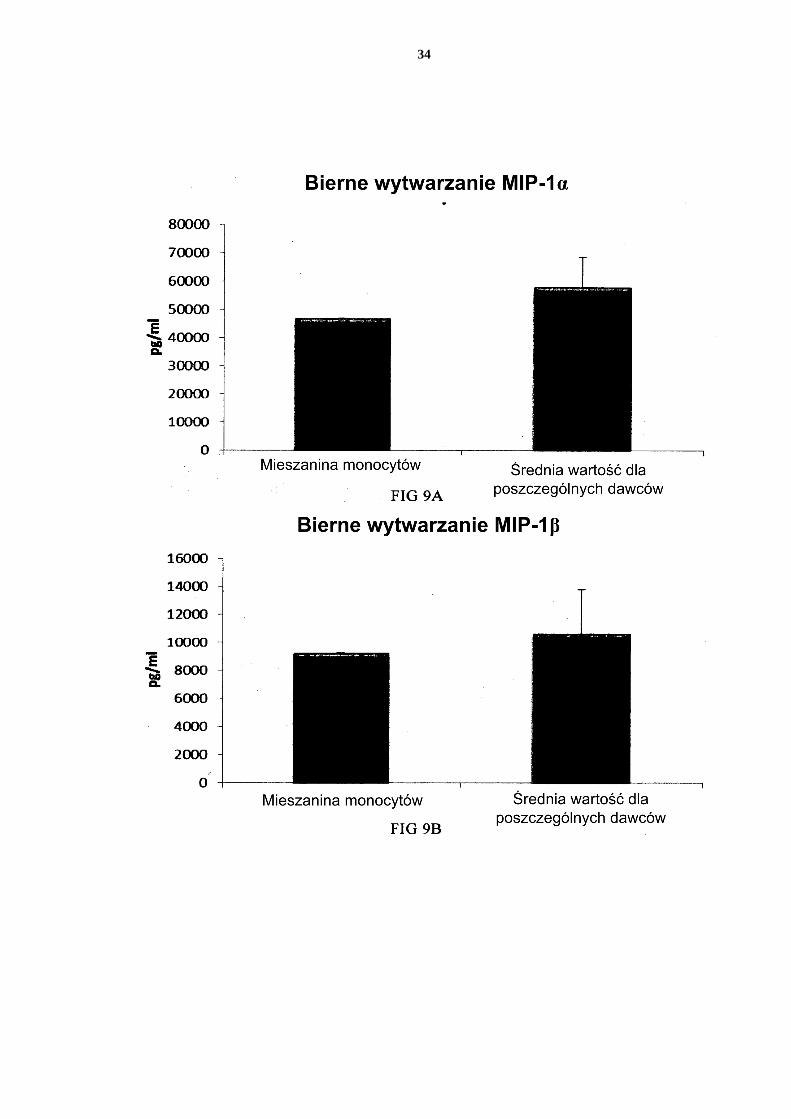
34
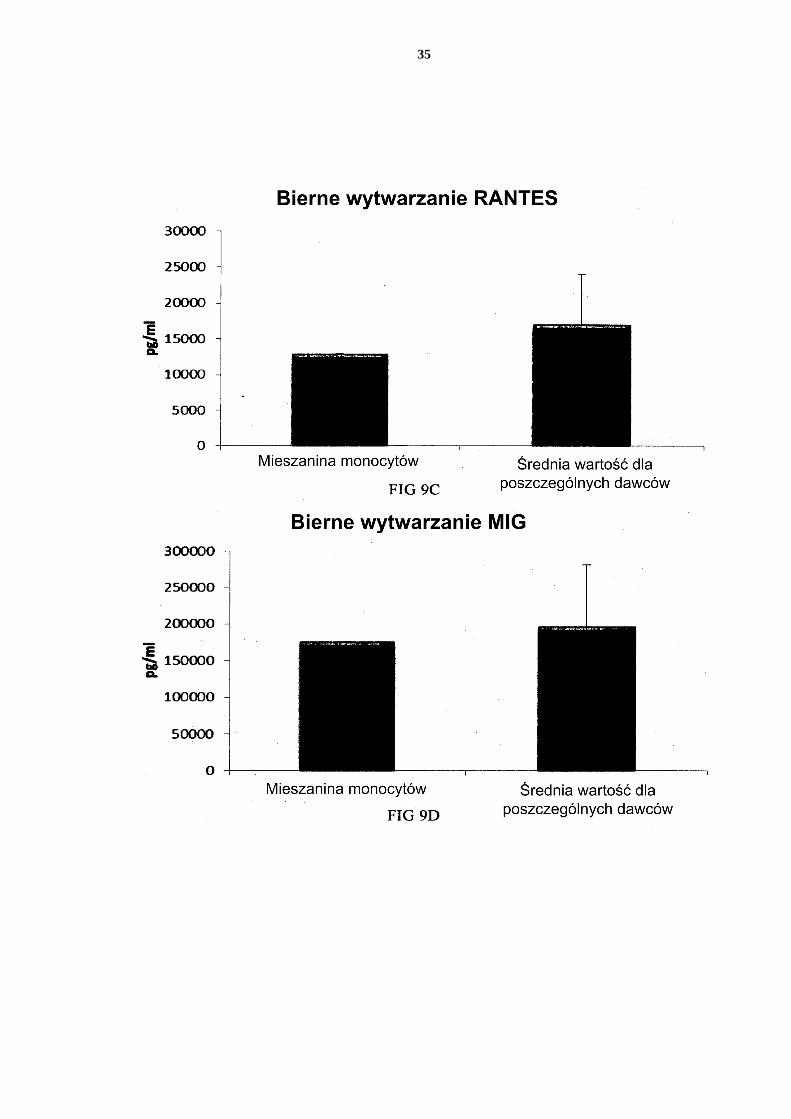
35
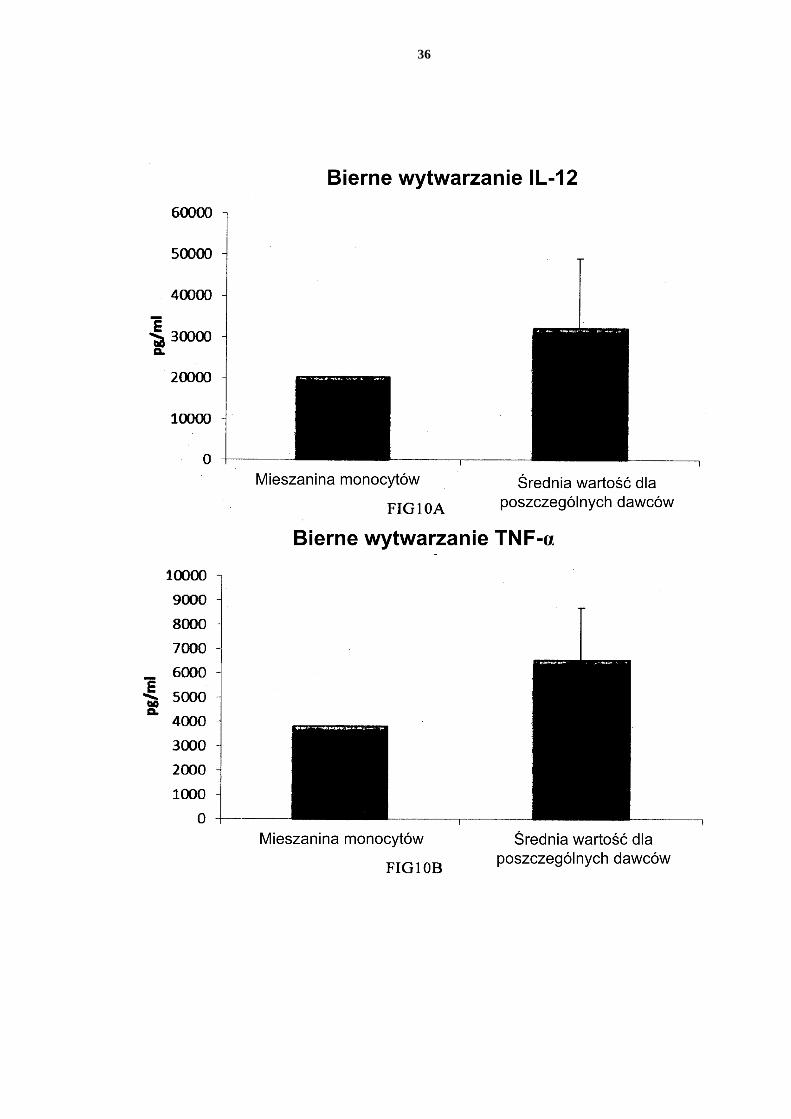
36
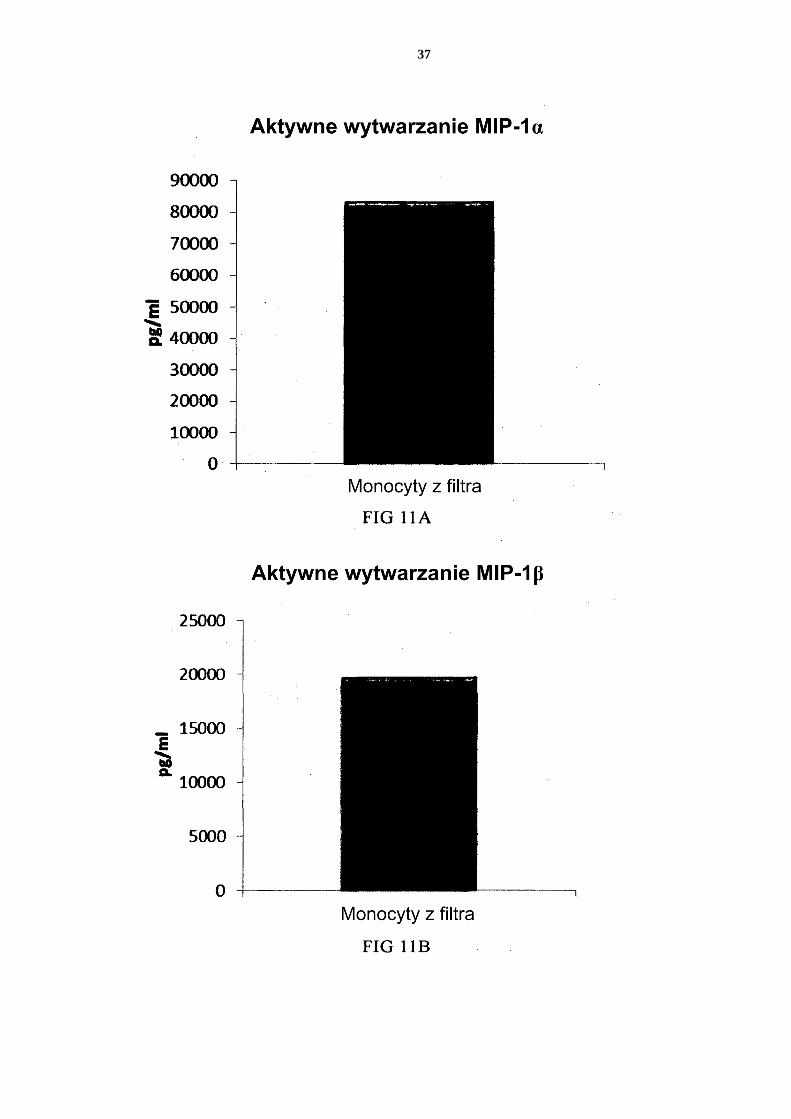
37
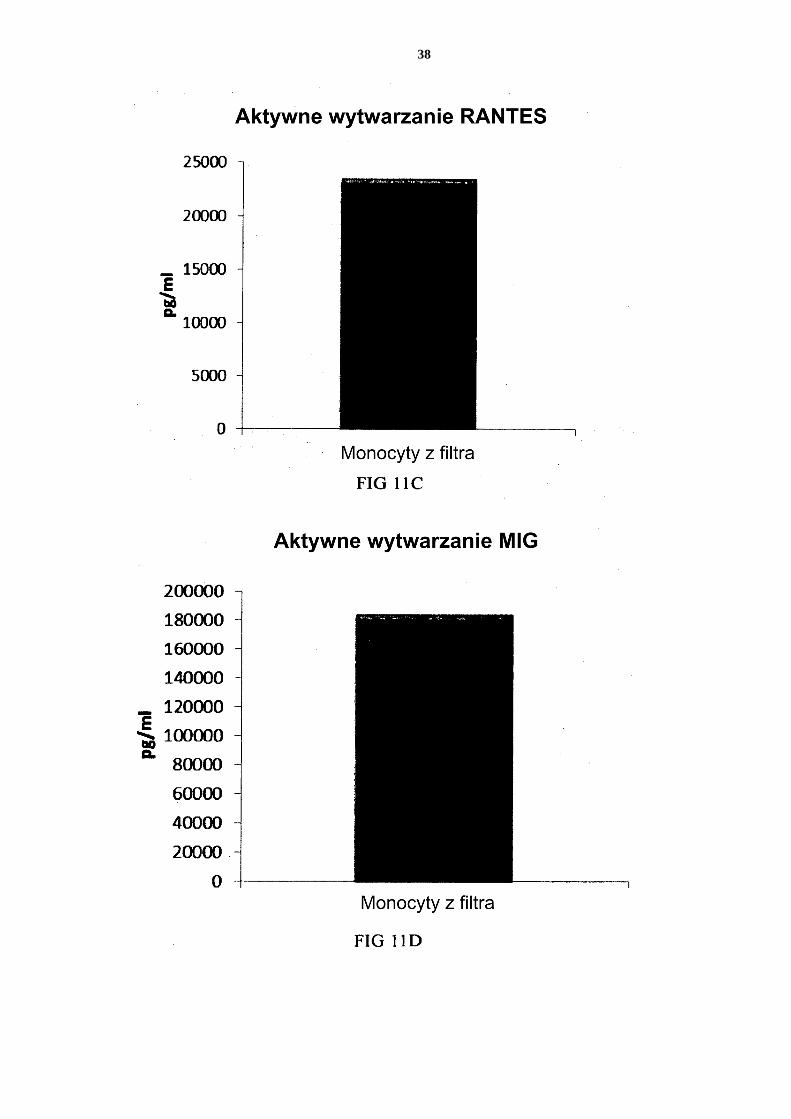
38
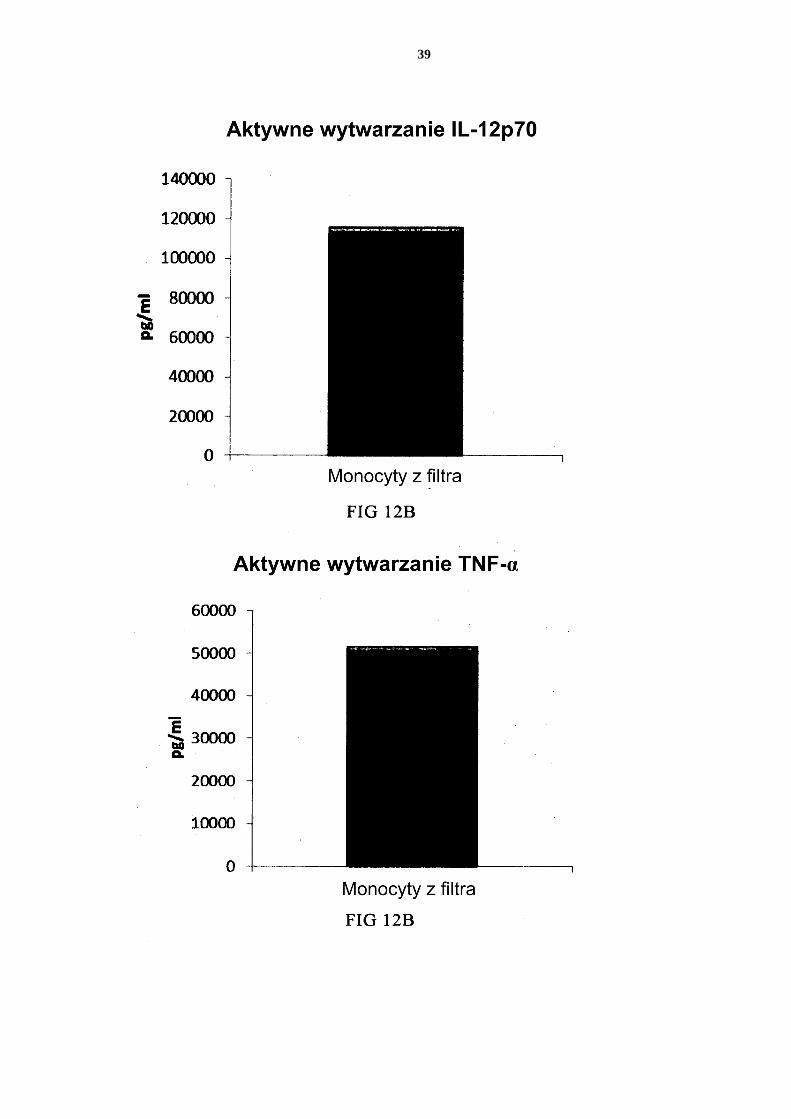
39
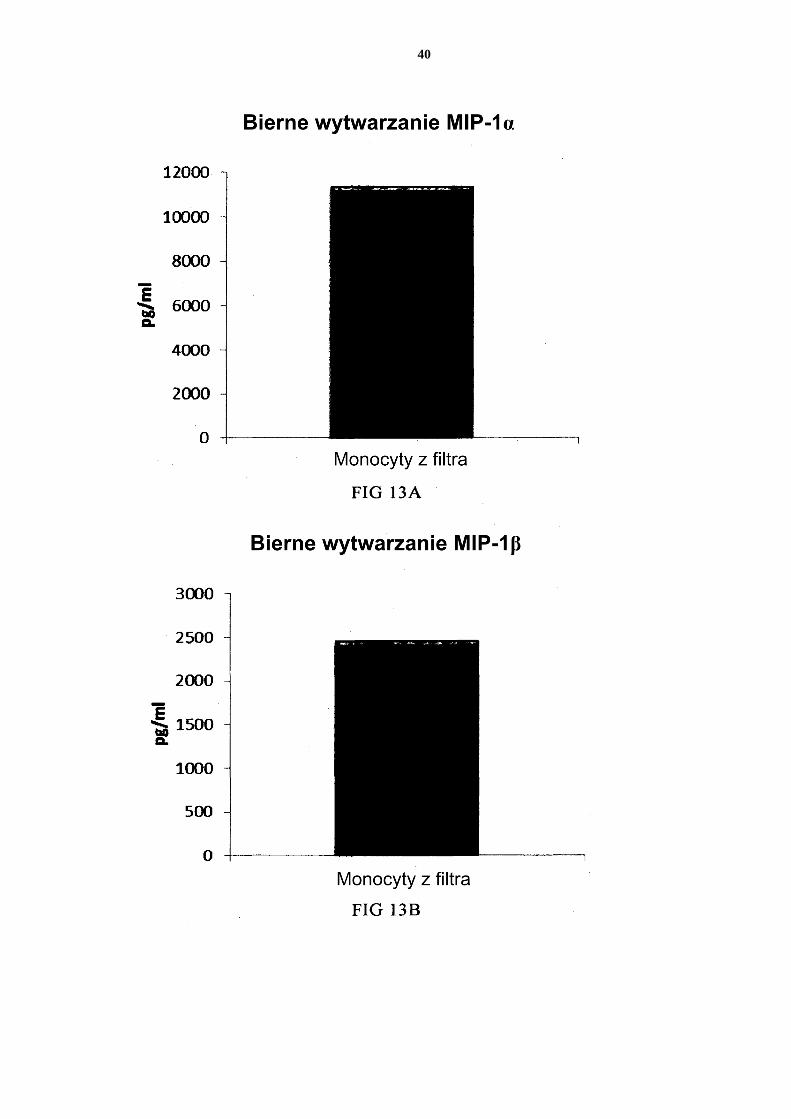
40
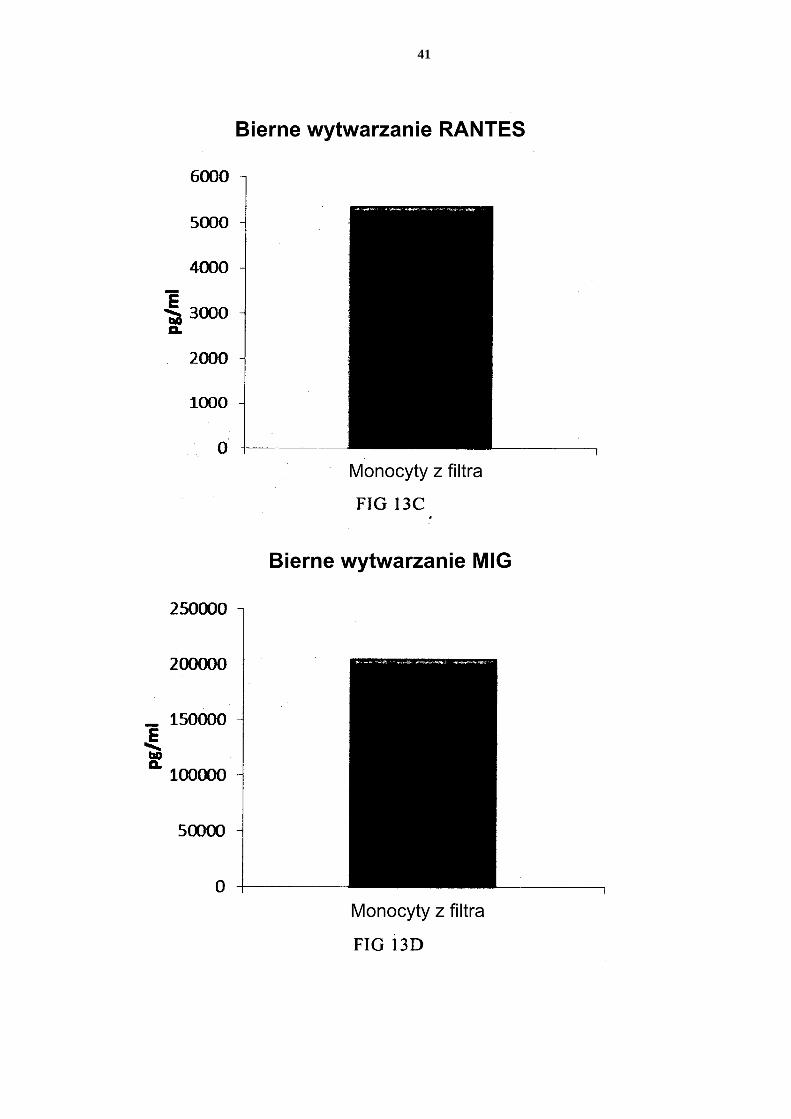
41

42





























