Embed Size (px)
Citation preview

The molecular basis of eukaryotic transcriptionRoger D. Kornberg*Stanford University School of Medicine, Stanford, CA 94305-5400
Iam deeply grateful for the honor
bestowed on me by the NobelCommittee for Chemistry and theRoyal Swedish Academy of Sci-
ences. It is an honor I share with mycollaborators. It is also recognition ofthe many who have contributed over thepast quarter century to the study oftranscription.
The NucleosomeMy own involvement in studies of tran-scription began with the discovery of thenucleosome, the basic unit of DNA coil-ing in eukaryote chromosomes (1). X-ray studies and protein chemistry led meto propose the wrapping of DNAaround a set of eight histone moleculesin the nucleosome (Fig. 1). Some yearslater, Yahli Lorch and I found that thiswrapping of DNA prevents the initiationof transcription in vitro (2). MichaelGrunstein and colleagues showed nu-cleosomes interfere with transcription invivo (3). The nucleosome serves as ageneral gene repressor. It assures theinactivity of all of the many thousandsof genes in eukaryotic cells except thosewhose transcription is brought about byspecific positive regulatory mechanisms.What are these positive regulatorymechanisms? How is repression by thenucleosome overcome for transcription?Our recent work has shown that pro-moter chromatin is transformed from astatic to a dynamic state upon gene acti-vation (4). Nucleosomes are rapidly re-moved and reassembled in the activatedstate. Promoter DNA is made tran-siently available for interaction with thetranscription machinery.
RNA Polymerase II (Pol II) TranscriptionOur studies have focused on the RNApol II transcription machinery. Pol II isresponsible for all messenger RNA syn-thesis in eukaryotes. As the first step ingene expression, pol II transcription isan end point of a great many signaltransduction pathways. The intricate reg-ulation of pol II transcription underliescell differentiation and development.
Because nucleosomes are removedfrom promoter DNA for transcription invivo, we and others have been able tofractionate the components of the tran-scription machinery guided by transcrip-tion assays performed with naked DNAin vitro. Robert Roeder and colleaguesinitiated the isolation of pol II transcrip-tion proteins from human HeLa cellextracts (5). This effort was brought to
fruition by Ronald and Joan Conaway,who took advantage of the greaterabundance of starting material availablefrom a rat liver extract (6). At Stanford,we isolated the pol II machinery fromyeast, in work begun by Neal Lue in1987, who solved the longstanding prob-lem of preparing an extract active in polII transcription from yeast (7). In retro-spect, our pursuit of the problem inyeast was a fortunate choice. It provedcrucial for unraveling both the structureand regulation of the pol II machinery.There were serious doubts when we be-gan whether findings in yeast wouldprove relevant to human cells. But,upon fractionation of yeast and mamma-lian systems, the results were the same.
Both systems comprise six proteins:pol II, and five general transcription fac-tors, known as TFIIB, -D, -E, -F, and-H (8). Pol II is capable of unwindingDNA, synthesizing RNA, and rewindingDNA. But pol II alone is incapable ofrecognizing a promoter and initiatingtranscription. For these essential func-tions, the participation of the generaltranscription factors is required.
Mediator of Transcriptional RegulationIt was, at first, thought the set of sixproteins constituted a complete tran-
scription system, that it would supportnot only accurately initiated but alsoappropriately regulated transcription.Communication from a regulatory pro-tein to the transcription machinery at apromoter was believed to be direct. Wefound, however, that an additional crudefraction was required for regulation inthe yeast system (Fig. 2). We referred tothis activity as Mediator (9, 10), and in1994, Stefan Bjorklund and Young-JoonKim isolated the active protein as anassembly of more than 20 subunits, witha total mass in excess of a million dal-tons (11). Thirteen of the subunits wereproducts of genes previously identifiedin screens for molecules involved intranscriptional regulation. These weredisparate screens, done on differentpromoters in different laboratories atdifferent times. With the isolation ofMediator, the products of all of thescreens were united in a common bio-chemical entity. Still, the Mediator ideadid not gain wide acceptance, as regula-tion in higher organisms was believed tobe direct, through interaction of geneactivator proteins with the so-calledTAF subunits of the general transcrip-tion factor TFIID. Finally, in 1998, weand others isolated mammalian counter-parts of the yeast Mediator (reviewed inrefs. 12 and 13), and TAFs were shownto be important for promoter recogni-tion, not regulation (14–16). As othershave shown, 22 of 25 yeast Mediatorsubunits have demonstrable homologsin higher cells (17, 18).
We are just beginning to fathom thecomplexity of Mediator, but alreadythree points are clear. First, Mediator isnot only the basis for regulated tran-scription; it is absolutely required foralmost all transcription of almost all polII promoters (19, 20). Mediator is noless essential for transcription than polII itself. Second, Mediator interacts di-
Author contributions: R.D.K. wrote the paper.
*E-mail: [email protected].
Adapted from Les Prix Nobel 2006. © 2006 by the NobelFoundation.
Editor’s Note: This article is a version of Roger Kornberg’sNobel Lecture, ‘‘The molecular basis of eukaryotic transcrip-tion.’’ The 2006 Nobel Prize in Chemistry was awarded to Dr.Roger Kornberg for his breakthrough progress in the mo-lecular understanding of transcription and its regulation ineukaryotic cells. Kornberg’s contribution has culminated inhis creation of detailed crystallographic pictures describingthe transcription apparatus of the eukaryotic cell in fullaction. The Nobel Foundation has graciously granted uspermission to print this article to help broaden its exposure.
Fig. 1. The nucleosome, the fundamental particleof the eukaryote chromosome. Schematic showsthe coiling of DNA around a set of eight histones inthe nucleosome, the further coiling in condensed(transcriptionally inactive) chromatin, and uncoil-ing for interaction with the RNA polymerase II (polII) transcription machinery.
www.pnas.org�cgi�doi�10.1073�pnas.0704138104 PNAS � August 7, 2007 � vol. 104 � no. 32 � 12955–12961
PE
RS
PE
CT
IVE

rectly both with activator proteins andwith pol II (13). It forms a tight com-plex with an activator at an enhancer,and it subsequently contacts pol II andthe general transcription factors at thepromoter to stimulate the initiation oftranscription (12). Third, Mediator isimportant not only for positive but alsofor negative regulation of transcription.Although Mediator is commonly re-ferred to as a coactivator, this is a mis-nomer. Mediator is a coactivator, acorepressor, and a general transcriptionfactor all in one. Mediator may beviewed as a signal processor. It trans-duces regulatory information from en-hancers to promoters in the entire rangeof organisms from yeast to humans.
Structural Studies of the Pol IITranscription MachineryIt has been said: if you wish to under-stand function, study structure. Thechallenge in the case of the pol II tran-scription machinery lies in the great sizeof the structure. A giant complex ofnearly 60 proteins, with a total mass inexcess of 3 million daltons, assembles atevery pol II promoter before the initia-tion of transcription (Fig. 3). We beganwith structure determination of pol IIbecause it forms the core of the tran-scription complex. In retrospect, thiswas another fortunate choice. It waspreferable to pursuing the smaller sim-pler transcription factors because pol IIis the platform upon which all factors
are assembled. It emerged from ourwork that some of the general factorproteins adopt their fully folded struc-tures only upon interaction with pol II.Knowledge of the pol II structure hasproved key to understanding eukaryoticgene transcription.
Two-DimensionalProtein CrystallographyThe story of the pol II structure beganin my graduate work in physical chemis-try, with nuclear resonance experimentsrevealing the rapid lateral diffusion oflipid molecules in multilayers (21). Someyears later, I thought of exploiting lat-eral diffusion for the formation of sin-gle-layer-thick or two-dimensional (2D)protein crystals. The idea was to bind aprotein to a lipid layer through interac-tion with the lipid head groups. Thebound protein would be constrained in2D but free to diffuse in the plane andcrystallize (ref. 22 and Fig. 4). SethDarst and Al Edwards succeeded informing 2D crystals of pol II in this way(23). The crystals were initially smalland poorly ordered. They were of littleuse for structure determination, but theyprovided a starting point. We could takeadvantage of the ease, rapidity, andsmall amount of material required bythe 2D crystallographic approach anduse it as a structural assay to guide thepreparation of pol II that would formbetter crystals. We soon found that het-erogeneity, due to a substoichiometricamount of two small polymerase sub-units, was the problem: although theyaccounted for only 8% of the mass ofthe enzyme, their presence in a variableamount was the impediment to crystalli-zation. We turned to a deletion mutantof yeast from which we could isolate polII lacking both small subunits. The re-sulting homogeneous pol II preparationformed very large, exceedingly well or-dered 2D crystals (24). Even at the lowprotein concentration used for 2D crys-tal growth, �50 �g/ml, the crystalscould often be seen to add additionallayers in register with the first. This pro-pensity for epitaxial growth could be
exploited and the 2D crystals used toseed the formation of 3D crystals forx-ray analysis.
X-Ray Crystal Structure of Pol IIWe were excited when the first 3D crys-tals were obtained. I also recall a chillof anxiety. The largest x-ray structure ofan asymmetric particle at the time, 17years ago, was a fifth the size of pol II,and for good reason: x-ray beam intensi-ties, detectors, and computational capa-bilities were all limiting. Actually, thelimitations of diffraction technologyneed not have concerned us because thepol II crystals failed to diffract! Theproject would have ended there, were itnot for our noticing the crystals werefaintly tinged with yellow. The problemwas oxidation, and the solution was togrow and maintain the crystals in aglove box under argon.
Then it emerged that the crystalswere profoundly polymorphic, varyingby more than 10 Å along one unit celldirection. This variation, and the sheersize of pol II, posed challenges for phas-ing the diffraction by multiple isomor-phous replacement with heavy atoms.Jianhua Fu, who discovered the bestdiffracting form of pol II crystals, ad-dressed the phase problem by data col-lection from a large number of crystalsand with the use of heavy atom clustersdeveloped by others. He found matchedpairs of native and derivative crystalsfrom which phases to 5-Å resolutioncould be derived (25). The resultingelectron density map correspondedclosely to the structure of pol II at 16-Åresolution determined from 2D crystalsby electron microscopy and 3D recon-struction. This marked a turning pointin the solution of the pol II problem.With reliable 5-Å phases, it was possi-ble, in principle, to locate individualheavy atoms and solve the structure tonear atomic resolution.
Patrick Cramer and Dave Bushnellsought additional heavy atom deriva-tives. When they changed the motherliquor of the crystals for the purpose,the crystals shrank by 11 Å along thepreviously variable unit cell direction,eliminating the problem of polymor-phism and extending diffraction to 2.8-Åresolution. None of the 50 heavy atomcompounds commonly used for phasedetermination gave useful derivatives,but an iridium compound identified byFu and rhenium compounds were even-tually effective. The resulting structurecomprised some 3,500 amino acids, with28,000 nonhydrogen atoms (refs. 26 and27 and Fig. 5).
Where do DNA and RNA bind to polII? The answer came from the structuredetermination of the polymerase in the
Fig. 3. The RNA polymerase II transcription ma-chinery. Masses are round figures for proteins fromthe yeast Saccharomyces cerevisiae.
Fig. 2. Mediator of transcriptional regulation.Schematic shows the transduction of regulatoryinformation from a gene activator protein boundto an enhancer DNA element to the pol II transcrip-tion machinery at a promoter.
Fig. 4. Two-dimensional protein crystallizationon lipid layers. Schematic shows the binding of aprotein of interest (oval objects) to the headgroups (red triangles) of lipid molecules in a mono-layer at the air–water interface. Rapid lateral dif-fusion of the lipids leads to protein crystallization.
12956 � www.pnas.org�cgi�doi�10.1073�pnas.0704138104 Kornberg

form of a transcribing complex. Wetried to crystallize such a complex thewhile we pursued the structure of thepolymerase alone. The problem was thateven very highly purified pol II containsmany inactive molecules and thesewould contaminate any preparation oftranscribing complexes. Finally, AviGnatt discovered a way of removing theinactive molecules and succeeded ingrowing transcribing complex crystals(28). Transcription had been paused bywithholding one of the four nucleosidetriphosphates (NTPs), and upon soakingthe crystals in the missing NTP, tran-scription resumed without loss of crystalmorphology. The crystals were verythin and gave diffraction completeto only �6-Å resolution. After years oftrying, Avi collected a dataset completeto 3.3-Å resolution, which was solved bymolecular replacement with the 2.8-Åpolymerase structure (29).
DNA can be seen entering the tran-scribing complex in duplex form andunwinding three bases before the activesite (Fig. 6). Then the template strand
makes a sharp bend, and as a result, thenext base is f lipped, pointing down to-ward the active site. This base is pairedwith that of the ribonucleotide justadded to the RNA strand. The structurereveals eight more DNA–RNA hybridbase pairs and one additional base onthe template DNA strand. The remain-der of the template strand, the RNA,and the nontemplate DNA strand arenot seen due to motion or disorder.
Fidelity of TranscriptionHow does pol II select the right nucleo-tide for addition to the RNA chain?This is the essence of transcription, theaccurate readout of the genetic code.Our most recent work has shown howaccurate readout is achieved. In theoriginal transcribing complex structure,the nucleotide just added to the RNAwas still in the active center. In subse-quent structures, we were able to trapthe complex following translocation ofthe DNA and RNA across the enzymesurface, creating an empty site at theactive center, available for binding thenext NTP (Fig. 7). Soaking crystals ofthis ‘‘posttranslocation’’ complex withNTPs resulted in additional electrondensity at two sites (30). All four NTPswere seen to bind an entry or ‘‘E’’ site,whereas only the NTP correctlymatched for base pairing with the cod-ing base in the DNA was seen to bind inthe active center, at the nucleotide addi-tion or ‘‘A’’ site. The orientation ofNTP in the E site was inverted with re-spect to that in the A site, leading to thesuggestion that NTPs in the E site ro-tate to sample base pairing in the Asite.
These structures failed to reveal thebasis for the fidelity of transcription.The energy of base pairing, through twoor three hydrogen bonds to the templateDNA, is far less than required to ac-count for the selectivity of the polymer-ase reaction. The mystery remaineduntil this year, when screening manyhundreds of crystals by Dong Wang andDave Bushnell led to improved resolu-tion and data quality. The structure of atranscribing complex with correctlymatched NTP in the A site now in-cluded a feature termed the trigger loopbeneath the A site (ref. 31 and Fig. 8).The trigger loop was seen before inmany pol II structures, but only in thetwo solved this year, with correctlymatched NTP in the A site, did it ap-pear in proximity to the A site. In allprevious structures, it was located 30 Åor more distant from the A site. Thetrigger loop is evidently a mobile ele-ment, swinging like a trap door beneathcorrectly matched NTP in the A site.
The trigger loop contacts all moieties ofthe NTP: the base, the phosphates, and,through other pol II residues, the sugar aswell (Fig. 9). The resulting network ofinteractions even includes the 2�-OHgroup of the nucleotide just added to theend of the RNA. The importance of theseinteractions is shown by mutations affect-ing transcription (Fig. 9). For example,mutation of asparagine 479, hydrogenbonded to the 3�-OH group of the NTP,results in a loss of discrimination betweenthe normal nucleotide and one lacking a3�-OH group. The discrimination is notgreat, only �10-fold, of the magnitudeexpected for the energy of a single hydro-
Fig. 6. Structure of RNA polymerase II in the actof gene transcription at near atomic resolution.The polypeptide chain is shown in white, orange(mobile ‘‘clamp’’), and green (bridge helix connect-ing the two largest subunits). Backbone models ofthe nucleic acids are shown in blue (template DNAstrand), green (nontemplate DNA strand), and red(RNA).
Fig. 7. A cycle of nucleotide addition by RNApolymerase II. At the upper left, the structure fromFig. 6 is shown, omitting all but the DNA and RNAnear the active center and the bridge helix. Theribonucleotide in the active center, just added tothe RNA chain, is yellow. At the lower left is thestructure after translocation of DNA and RNAacross the pol II surface. At the lower right is thestructure with an unmatched NTP in the entry (E)site. At the upper right is the structure with NTP,matched for pairing to the coding base in thetemplate strand, in the addition (A) site.
Fig. 5. Structure of RNA polymerase II at 2.8-Åresolution. The protein is shown in ribbon repre-sentation, with a color code to the various subunitsand interaction diagram at the upper right. A Mgion at the active center is depicted as a pink sphere.
Fig. 8. The trigger loop. Transcribing complexstructures with purine nucleotide (orange) or py-rimidine nucleotide (dark blue) in the addition site(compare that in the upper right panel of Fig. 7) areshown superimposed. The corresponding triggerloops are purple and yellow, and the bridge helicesare green and light blue.
Kornberg PNAS � August 7, 2007 � vol. 104 � no. 32 � 12957
gen bond. In contrast, discrimination be-tween a normal ribo NTP and a 2�-deoxyNTP is very great, at least 1,000-fold, andis unaffected by mutation of asparagine479. How is such extraordinary specificityfor a single OH group achieved? The an-swer lies in the alignment of the triggerloop with the NTP and the precise posi-tioning of a histidine side chain, 3.5 Åfrom the �-phosphate (Fig. 10). The histi-dine promotes the flow of electrons dur-ing nucleophilic attack of the 3�-OH atthe chain terminus and phosphoanhydridebond breakage. It serves as a proton do-nor for the pyrophosphate leaving group.It literally triggers phosphodiester bondformation. It couples nucleotide selectionto catalysis.
The electronic transactions involvedin trigger loop function require precisealignment of the interacting moieties.This is achieved for a correct NTP byformation of the trigger loop network.In the case of an incorrect NTP, for ex-ample a 2�-deoxy NTP, misalignment isprofound. A double helix formed with a2�-deoxy nucleotide is 2 Å narrowerthan that formed by a ribonucleotide.The resulting misalignment with the cat-alytic histidine residue is as great as inthe case of a pyrimidine-pyrimidine basemismatch, leading to a 1,000-fold reduc-tion in the rate of phosphodiester bondformation.
Nucleic Acid TranslocationThe significance of the trigger loop net-work extends beyond nucleotide selec-tion and catalysis. The network includesmany contacts with the bridge helix that,in turn, contacts the coding base in the
template DNA strand. The structure ofbacterial polymerase, determined bySeth Darst (the first of a multisubunitRNA polymerase at the near atomiclevel), also includes a bridge helix, butin contrast with that in pol II, thebridge helix in the bacterial enzyme isbent (ref. 32 and Fig. 11). The bendproduces a movement of �3 Å in thedirection of the template strand, corre-sponding to one base pair step along thestrand. This led us to suggest that transi-tions of the bridge helix betweenstraight and bent states underlie thetranslocation step in transcription. Thebridge helix may serve as a kind of mo-lecular ratchet, allowing the polymeraseto let go of the DNA and RNA fortranslocation, while retaining a grip onthe growing end of the DNA–RNA hy-brid helix to preserve the register oftranscription. There is now a good dealof biochemical and genetic evidence tosupport this idea.
RNA ReleaseIn the final step of transcription, theRNA is released. The question arises
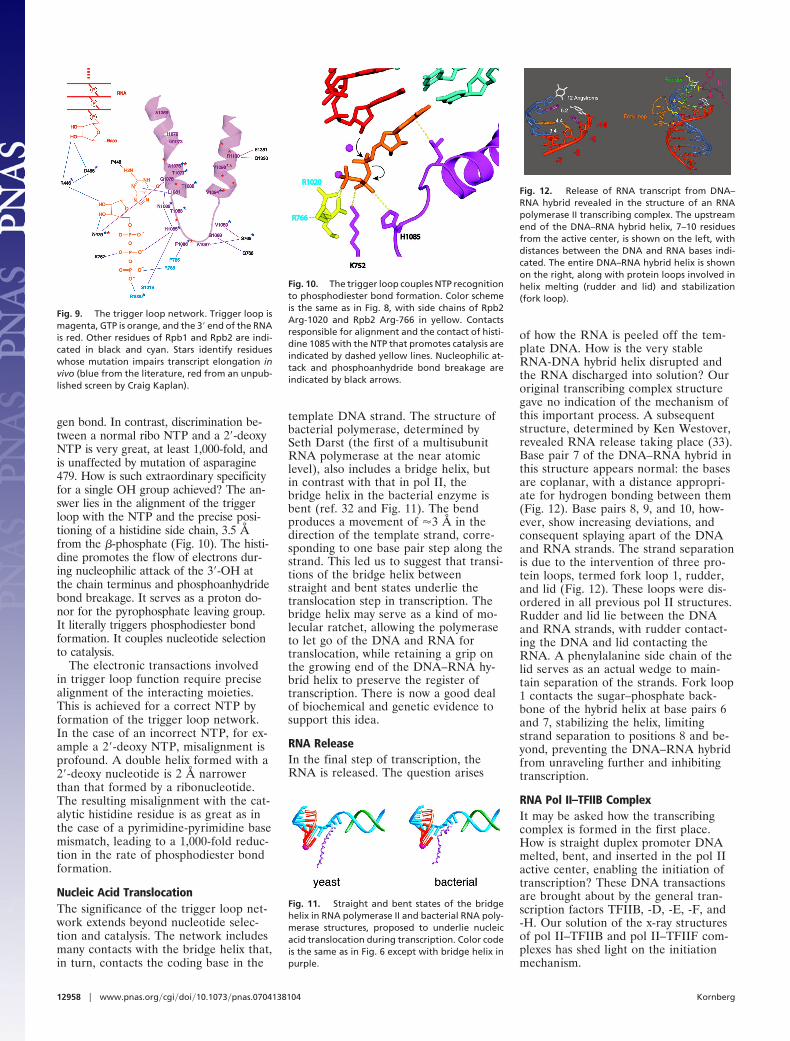
of how the RNA is peeled off the tem-plate DNA. How is the very stableRNA-DNA hybrid helix disrupted andthe RNA discharged into solution? Ouroriginal transcribing complex structuregave no indication of the mechanism ofthis important process. A subsequentstructure, determined by Ken Westover,revealed RNA release taking place (33).Base pair 7 of the DNA–RNA hybrid inthis structure appears normal: the basesare coplanar, with a distance appropri-ate for hydrogen bonding between them(Fig. 12). Base pairs 8, 9, and 10, how-ever, show increasing deviations, andconsequent splaying apart of the DNAand RNA strands. The strand separationis due to the intervention of three pro-tein loops, termed fork loop 1, rudder,and lid (Fig. 12). These loops were dis-ordered in all previous pol II structures.Rudder and lid lie between the DNAand RNA strands, with rudder contact-ing the DNA and lid contacting theRNA. A phenylalanine side chain of thelid serves as an actual wedge to main-tain separation of the strands. Fork loop1 contacts the sugar–phosphate back-bone of the hybrid helix at base pairs 6and 7, stabilizing the helix, limitingstrand separation to positions 8 and be-yond, preventing the DNA–RNA hybridfrom unraveling further and inhibitingtranscription.
RNA Pol II–TFIIB ComplexIt may be asked how the transcribingcomplex is formed in the first place.How is straight duplex promoter DNAmelted, bent, and inserted in the pol IIactive center, enabling the initiation oftranscription? These DNA transactionsare brought about by the general tran-scription factors TFIIB, -D, -E, -F, and-H. Our solution of the x-ray structuresof pol II–TFIIB and pol II–TFIIF com-plexes has shed light on the initiationmechanism.
Fig. 11. Straight and bent states of the bridgehelix in RNA polymerase II and bacterial RNA poly-merase structures, proposed to underlie nucleicacid translocation during transcription. Color codeis the same as in Fig. 6 except with bridge helix inpurple.
Fig. 9. The trigger loop network. Trigger loop ismagenta, GTP is orange, and the 3� end of the RNAis red. Other residues of Rpb1 and Rpb2 are indi-cated in black and cyan. Stars identify residueswhose mutation impairs transcript elongation invivo (blue from the literature, red from an unpub-lished screen by Craig Kaplan).
Fig. 10. The trigger loop couples NTP recognitionto phosphodiester bond formation. Color schemeis the same as in Fig. 8, with side chains of Rpb2Arg-1020 and Rpb2 Arg-766 in yellow. Contactsresponsible for alignment and the contact of histi-dine 1085 with the NTP that promotes catalysis areindicated by dashed yellow lines. Nucleophilic at-tack and phosphoanhydride bond breakage areindicated by black arrows.
Fig. 12. Release of RNA transcript from DNA–RNA hybrid revealed in the structure of an RNApolymerase II transcribing complex. The upstreamend of the DNA–RNA hybrid helix, 7–10 residuesfrom the active center, is shown on the left, withdistances between the DNA and RNA bases indi-cated. The entire DNA–RNA hybrid helix is shownon the right, along with protein loops involved inhelix melting (rudder and lid) and stabilization(fork loop).
12958 � www.pnas.org�cgi�doi�10.1073�pnas.0704138104 Kornberg

The structure of the pol II–TFIIBcomplex (34) revealed distinct functionsof the N- and C-terminal domains ofTFIIB. A polypeptide chain trace of theN-terminal domain (yellow in Fig. 13)begins with a Zn ribbon that binds thepol II surface adjacent to the clamp andwall. Then the polypeptide does anamazing thing: rather than pass backinto solution, it continues across thesaddle between the clamp and wall andplunges toward the active center, fromwhich it loops back and remerges acrossthe saddle. The loop, which we havetermed the B finger, occupies almost thesame location as the DNA–RNA hybridin a transcribing complex. Superimpos-ing the B finger and the DNA–RNAhybrid from the transcribing complexstructure reveals no interference withthe template DNA strand or with theRNA up to position 5, but a steric clashwith the RNA at positions 6 and beyond(Fig. 14).
Biochemical experiments show thatthe B finger is not only compatible witha hybrid containing five residues ofRNA, but is required for stability ofsuch a complex. When the RNA growsbeyond five or six residues, however, itmust compete with TFIIB for space onthe pol II saddle. If TFIIB wins thecompetition, initiation is aborted andmust be tried again. If the RNA wins,TFIIB is ejected and pol II is releasedfrom the promoter to continue andcomplete transcription. The B fingerthus explains two crucial but until nowmysterious aspects of pol II transcrip-tion: abortive initiation and promoterescape. In these respects, it resemblesthe sigma factor in bacterial transcrip-tion (35, 36).
Turning to the C-terminal domain ofTFIIB, its location in the structure ofthe pol II–TFIIB complex served as aguide for docking a structure deter-mined long ago (37) of a ternary com-
plex of a C-terminal TFIIB fragment,the TATA-binding protein (TBP) sub-unit of TFIID, and a TATA box DNAfragment (Fig. 15). This modeling exer-cise led to a moment of truth. TheTATA box DNA fragment is sharplybent by TBP. What would happen if theends of the bent fragment were ex-tended with straight B-form DNA? Theresult was remarkable in two respects.First, the DNA fit snugly against theprotein (Fig. 15 Left): TBP evidentlyconfigures promoter DNA to the con-tours of the pol II surface. Second, theDNA downstream of the TATA box ranpast the saddle (Fig. 15 Right). The dis-tance from the TATA box to the saddleis �1.5 turns of the double helix, or 15bp. We know from the transcribingcomplex structure that �12 residues arerequired to cross the saddle to the ac-
tive site. The sum of 15 bp from theTATA box and 12 residues to the activesite is 27 bp, closely coincident with thespacing of 25–30 bp from the TATA boxto the transcription start site of almostall pol II promoters. In this way, polII–TFIIB interaction may determine thelocation of the transcription start site.
RNA Pol II–TFIIF ComplexA cocrystal structure of a transcribingcomplex with the central subunit ofTFIIF is also informative about the ini-tiation of transcription. This work inprogress of Guillermo Calero was madepossible by his mastery of the biochemi-cal behavior of TFIIF. The structureincludes a complete transcription bub-ble, not only the template DNA strandwith associated RNA seen in previousstructures, but also the nontemplateDNA strand, and the region upstreamof the bubble where duplex DNA is re-formed following transcription (Fig. 16).The nontemplate strand and upstreamduplex DNA were revealed because ofinteraction with TFIIF, constrainingtheir mobility. The interaction of thenontemplate strand with TFIIF may trapa transient bubble in promoter DNA,leading to the initiation of transcription.
RNA Pol II Transcription InitiationComplexThe available structural informationmay be assembled in a preliminary pic-ture of the pol II transcription initiationcomplex (Fig. 17). The structures of polII, TBP, and TFIIB come from x-raycrystallography, as I’ve described. Thestructures of TFIIE, TFIIF, and TFIIHare from electron crystallography andfrom cryo-electron microscopy andsingle-particle analysis.
The picture of the complete complexprovides a solution, in outline, of thetranscription initiation problem. Each ofthe general factors is seen to play a sim-ple, essential role in the initiation pro-cess: TBP bends the promoter DNAaround the polymerase and the C-termi-
Fig. 15. Model of an RNA polymerase II–TBP–TFIIB–DNA complex. The structure of the C-terminal region of TFIIB (pink) complexed with TBP(green) and TATA-box containing DNA (red/white/blue atomic model) was docked to the structure ofthe pol II–TFIIB complex (as shown in Fig. 13). Theviews shown at the left and right are related by a90° rotation about an axis between them, as indi-cated by the curved arrow. The direction of view atright is the same as that in Fig. 13.
Fig. 13. Structure of an RNA polymerase II–TFIIBcomplex. A surface representation of pol II isshown, with the clamp and wall as in Fig. 6, apolypeptide chain trace of the amino terminal re-gion of TFIIB, designated IIBN, in yellow, and theregion of the pol II surface interacting with IIBN ingreen.
Fig. 14. Superposition of the DNA-hybrid helixfrom an RNA polymerase II transcribing complex(Fig. 6) and the B finger from an RNA polymeraseII–TFIIB complex (Fig. 13). Conserved region of theB finger is green.
Fig. 16. Structure of an RNA polymerase II tran-scribing complex with the central subunit of TFIIF(work in progress of Guillermo Calero). Pol II andthe nucleic acids only are shown on the left, whilethe structure including the TFIIF subunit (known asTfg2, in yellow) is shown on the right. Direction ofview and color scheme as in Fig. 6.
Kornberg PNAS � August 7, 2007 � vol. 104 � no. 32 � 12959

nal domain of TFIIB. The N-terminaldomain of TFIIB brings the DNA to apoint on the polymerase surface fromwhich it need only follow a straight pathand, by virtue of the conserved spacingfrom TATA box to transcription startsite in pol II promoters, the start site isjuxtaposed with the active center. TFIIEenters the complex and recruits TFIIH,whose ATPase/helicase subunit intro-duces negative superhelical tension inthe DNA. Thermal unwinding producesa transient bubble, which is captured byTFIIF binding to the nontemplatestrand. The DNA can now bend in thesingle stranded region and descend intothe pol II active center. Initiation and
the synthesis of RNA ensue, initiallystabilized by the B finger. Synthesis of atranscript greater than about six residuesin length leads to the displacement ofTFIIB, promoter escape, and the com-pletion of transcription.
Mediator and the Regulationof TranscriptionIt remains to solve the Mediator, andthus to understand transcriptionalregulation. The structure of a pol II–Mediator complex has been determinedat low resolution by Francisco Asturias,by cryo-electron microscopy and singleparticle analysis (ref. 38 and Fig. 18).The portion of the structure due to polII could be identified by docking theatomic model of the polymerase. Theremainder of the structure, due to Me-diator, was in the form of a crescent,largely enveloping pol II, with manypoints of contact through which regula-tory information may be transmitted.Extension of the structure to atomic res-olution will one day reveal the regula-tory mechanism.
The charge for this lecture was to tell the storyof the work leading to the Prize. It is also thestory of many students and postdoctoral fel-lows, whose skill, great effort, and willing sus-pension of disbelief, over a quarter century,transformed a scientific dream into a reality. Icould only mention a few by name without in-terrupting the flow of the story, but the contri-butions of others were not less important. I pay
heartfelt tribute to them, and to all who haveshared part of their scientific lives with me (seebox below), in pursuit not only of the pol IIstructure, but also of chromatin, transcriptionbiochemistry, and EM methodology. I shouldadd that this is a lecture, not a review, and so ispersonal rather than referential. The pol IIstructure is a culmination of research done inmany laboratories, and I am deeply indebted tothose involved.
Fig. 18. Cryo-EM structure of an RNA polymeraseII–Mediator complex. The pol II structure wasdocked in the central density, and is shown in asimilar direction of view and color scheme as Fig. 5.
With thanks to . . .
Chris AckersonFrancisco AsturiasMaia AzubelBrad BaerAvital Baraket-SamishStefan BjorklundHinrich BoegerTilman BorggrefeRick BramAndy BuchmanDave BushnellBrad CairnsGuillermo CaleroWei-hau ChangDan ChasmanJoan ConawayPatrick CramerSeth DarstPeter DavidRalph DavisAl EdwardsAndy EhrensbergerJohn FeaverMartha FedorPeter FlanaganJianhua FuBrian Gibbons
Opher GileadiAvi GnattDave GoldfarbJoachim GriesenbeckClaes GustafssonLynn HenryJohn HeumannGlenn HornPablo JadzinskyGrant JensenYi-Wei JiangCraig KaplanLin-Woo KangDon KatcoffRay KelleherYoung-Joon KimHirofumi KomoriGuy Lorch KornbergLiz KubalekYoshikazu KurashimaJan LaPointeKarl-Magnus LarssonKerstin LeutherAlan LevineYang LiYahli LorchNeal Lue
Barbara Maier-DavisLynne MercerGavin MeredithLarry MyersKazuko NishikuraHiroshi NojimaClaudia PoglitschAriel PrunellJeff ReidlerHans RibiPhil RobinsonAlan SachsMike SayreJonny SextonDong ShinHenrik SpahrSeth StrattanNori SugimotoJesper SvejstrupYuro TakagiHerbert TschochnerEd UzgirisPhilippe VeschambreDaguang WangDong WangKen WestoverMincheng Zhang
Fig. 17. Model of an RNA polymerase II preini-tiation complex. Structures shown at the upper leftwere assembled in the complex shown at the lowerright. Direction of view as in Figs. 5 and 13. ‘‘4/7’’indicates pol II subunits Rpb4 and Rpb7, whichwere omitted from the original structure (Fig. 5).
12960 � www.pnas.org�cgi�doi�10.1073�pnas.0704138104 Kornberg

1. Kornberg RD (1974) Science 184:868–871.2. Lorch Y, LaPointe JW, Kornberg RD (1987) Cell
49:203–210.3. Han M, Grunstein M (1988) Cell 55:1137–1145.4. Boeger H, Griesenbeck J, Strattan JS, Kornberg
RD (2003) Mol Cell 11:1587–1598.5. Matsui T, Segall J, Weil PA, Roeder RG (1980)
J Biol Chem 255:11992–11996.6. Conaway JW, Reines D, Conaway RC (1990)
J Biol Chem 265:7552–7558.7. Lue NF, Kornberg RD (1987) Proc Natl Acad Sci
USA 84:8839–8843.8. Conaway R, Conaway J (1997) Prog Nucleic Acid
Res Mol Biol 56:327–346.9. Kelleher RJ, III, Flanagan PM, Kornberg RD
(1990) Cell 61:1209–1215.10. Flanagan PM, Kelleher RJ, III, Sayre MH,
Tschochner H, Kornberg RD (1991) Nature350:436–438.
11. Kim YJ, Bjorklund S, Li Y, Sayre MH, KornbergRD (1994) Cell 77:599–608.
12. Kornberg RD (2005) Trends Biochem Sci 30:235–239.
13. Malik S, Roeder RG (2005) Trends Biochem Sci30:256–263.
14. Moqtaderi Z, Bai Y, Poon D, Weil PA, Struhl K(1996) Nature 383:188–191.
15. Walker SS, Reese JC, Apone LM, Green MR(1996) Nature 383:185–188.
16. Shen W-C, Green MR (1997) Cell 90:615–624.17. Conaway RC, Sato S, Tomomori-Sato C, Yao T,
Conaway JW (2005) Trends Biochem Sci 30:250–255.18. Bourbon HM, Aguilera A, Ansari AZ, Asturias
FJ, Berk AJ, Bjorklund S, Blackwell TK, Borg-grefe T, Carey M, Carlson M, et al. (2004) Mol Cell14:553–557.
19. Thompson CM, Young RA (1995) Proc Natl AcadSci USA 92:4587–4590.
20. Takagi Y, Kornberg RD (2006) J Biol Chem281:80–89.
21. Kornberg RD, McConnell HM (1971) Proc NatlAcad Sci USA 68:2564–2568.
22. Uzgiris EE, Kornberg RD (1983) Nature 301:125–129.
23. Edwards AM, Darst SA, Feaver WJ, ThompsonNE, Burgess RR, Kornberg RD (1990) Proc NatlAcad Sci USA 87:2122–2126.
24. Darst SA, Kubalek EW, Edwards AM, KornbergRD (1991) J Mol Biol 221:347–357.
25. Fu J, Gnatt AL, Bushnell DA, Jensen GJ, Thomp-son NE, Burgess RR, David PR, Kornberg RD(1999) Cell 98:799–810.
26. Cramer P, Bushnell DA, Fu J, Gnatt AL, Maier-Davis B, Thompson NE, Burgess RR, Edwards
AM, David PR, Kornberg RD (2000) Science288:640–649.
27. Cramer P, Bushnell DA, Kornberg RD (2001)Science 292:1863–1876.
28. Gnatt A, Fu J, Kornberg RD (1997) J Biol Chem272:30799–805.
29. Gnatt AL, Cramer P, Fu J, Bushnell DA, Korn-berg RD (2001) Science 292:1876–1882.
30. Westover KD, Bushnell DA, Kornberg RD (2004)Cell 119:481–489.
31. Wang D, Bushnell DA, Westover KD, Kaplan CD,Kornberg RD (2006) Cell 127:941–954.
32. Zhang G, Campbell EA, Minakhin L, Richter C,Severinov K, Darst SA (1999) Cell 98:811–824.
33. Westover KD, Bushnell DA, Kornberg RD (2004)Science 303:1014–1016.
34. Bushnell DA, Westover KD, Davis RE, KornbergRD (2004) Science 303:983–988.
35. Murakami KS, Masuda S, Campbell EA, MuzzinO, Darst SA (2002) Science 296:1285–1290.
36. Murakami KS, Masuda S, Darst SA (2002) Science296:1280–1284.
37. Nikolov DB, Chen H, Halay ED, Usheva AA,Hisatake K, Lee DK, Roeder RG, Burley SK(1995) Nature 377:119–128.
38. Davis JA, Takagi Y, Kornberg RD, Asturias FA(2002) Mol Cell 10:409–415.
Kornberg PNAS � August 7, 2007 � vol. 104 � no. 32 � 12961





























