Embed Size (px)
Citation preview

WS 13/14
Strukturbiologie, Transcription 1
TRANSCRIPTION
a) Transcription in prokaryotes
b) Transcription in eukaryotes

2
Fig.
Stryer
Transcription in prokaryotes and eukaryotes
The fundamental mechanism of transcription is conserved among cellular RNA
polymerases, yet there are also marked differences between prokaryotes and eukaryotes:
Transcription and translation are coupled in bacteria; wherease transcription and
translation are uncoupled in eukarya.
-Stages of Transcription: Initiation, Elongation, Termination
-“transcription bubble,” unwound region of about 15 base pairs of the DNA template and
some eight residues of the RNA transcript hybridized with the DNA in the center of the
bubble.

3
Transcription cycle in prokaryotes
INITIATION of transcription
1. Binding of polymerase as a
holoenzyme (s factor plus core
polymerase)
2. Open complex formation
(transcription “bubble”).
Unwinding of DNA, forming single
strandedness within the active site.
3. Initial RNA synthesis. Up to 10
bp of RNA is synthesized. During
this initial step the polymerase is
not very efficient and can easily fall
off. www.bmb.psu.edu/courses/bmmb501/bmmb597a_fa
03/reese/16_lect_gene_reg_1_.pdf -
3 distinct phases:
INITIATION,
ELONGATION,
TERMINATION.

4
Transcription cycle in prokaryotes
4. Release of the s factor and
synthesis beyond 10 bp of RNA
(transition from initiation to
elongation Durniak, K.J., Bailey, S.,
Steitz, T.A. (2008) The structure of a
transcribing T7 RNA polymerase in transition
from initiation to elongation Science). A
structural change in the polymerase
occurs. The “jaws” tighten down on
the DNA and the elongation
complex is MUCH more stable.
5. Highly processive ELONGATION
phase
6. TERMINATION. A termination
signal in the DNA forms a RNA
hairpin in the emerging transcript.
Its binding to polymerase stimulates
a change in conformation (opening
of the “jaws”) and release.
www.bmb.psu.edu/courses/bmmb501/bmmb597a_fa
03/reese/16_lect_gene_reg_1_.pdf -

Strukturbiologie, Transcription 5
a) Transcription in prokaryotes
In prokaryotes, transcription and translation are closely coupled.
Gene transcription is regulated by protein transcription factors that bind to
operator DNA and thus influence the ability of RNA polymerase to bind to
a promoter region and initiate transcription.
Protein transcription factors are regulated by cellular environmental factors
(e.g. transcription factors, allosteric effectors), which can include small
molecules, another protein or metal ions. Transcription can be blocked by
binding of a specific repressor (e.g. lac) protein at a DNA site called an
operator. These DNA binding proteins recognize specific DNA sequences via
distinct DNA-binding domains.
Gene transcription in bacteria, Schreiter, 2007

Strukturbiologie, Transcription 7
Transcription in prokaryotes - RNAP
Y.W. Yin and T.A. Steitz, Structural basis for the transition from initiation to
elongation transcription in T7 RNA polymerase. Science 298 (2002).

Strukturbiologie, Transcription 8
Transcription in prokaryotes
Crystal structures of the T.
thermophilus elongation
complex (ttEC) with the
non-hydrolysable
substrate analogue
AMPcPP (Vassylyev et al,
nature 2007, 3Å)
Overall view of the ttEC/AMPcPP
complex.
The DNA template, non-template and
RNA strands are in red, blue and yellow,
respectively.
The BH, the TH and the rest of the RNAP
molecule are in magenta, cyan and
grey, respectively.
The insertion and preinsertion NTP
analogues and Stl are designated by
green, orange and black, respectively.
The catalytic Mg2+ ions (MgI and MgII)
are shown as magenta spheres. a, b,

9
-Eukaryotes: Transcription and Translation are uncoupled
b) Transcription in eukaryotes
-Eukaryotes: 3 different
RNA polymerases (Pol I,
Pol II, Pol III):
Regulatory elements of
eukaryotic transcription
(TATA-box, -25)

Strukturbiologie, Transcription 10
3D structure of the nucleosome
Surface representation of the histone
octamer
Structure of the nucleosome core
particle; (14 independent DNA-binding
locations)
Review, Karolin Luger
-In eukaryotes: Chromatin is composed of nucleosomes, which consist of an
octamer of histones around which 147 base pairs of DNA are wrapped.
Structure of the nucleosome, T. Richmond Lab, 1997

Strukturbiologie, Transcription 12
Transcription in eukaryotes
4 October 2006
The Royal Swedish Academy of Sciences has decided to
award the Nobel Prize in Chemistry for 2006 to
Roger D. Kornberg
Stanford University, CA, USA
"for his studies of the molecular basis of eukaryotic
transcription".
Kornberg's contribution has culminated in his creation of detailed
crystallographic pictures describing the transcription apparatus in full
action in a eukaryotic cell. In his pictures (all of them created since 2000)
we can see the new RNA-strand gradually developing, as well as the role of
several other molecules necessary for the transcription process. The pictures
are so detailed that separate atoms can be distinguished and this makes it
possible to understand the mechanisms of transcription and how it is
regulated.

Strukturbiologie, Transcription 13
Structure determination of RNA
polymerase II and complexes
1983 2-D protein crystals on lipid layers
1991 2-D crystals seed 3-D crystals (poor diffraction-work under
Argon)
1998 Diffraction phased with heavy atom clusters
2000 Structure of RNA polII at 2.8 Å resolution
2002 Structure of transcribing complex 3.3 Å
2002-ongoing Series of structures of transcribing complexs (2.9-
4.4 Å), complexes with bound inhibitors .....

14
The Pol II transcription machinery (>3 MioDa)
Pol II is capable of unwinding DNA, synthesizing RNA, and rewinding
DNA. But Pol II alone is incapable of recognizing a promoter and
initiating transcription. For these essential functions, the participation
of the General Transcription Factors is required. Mediator is co-
activator, a co-repressor, and a general transcription factor all in one.
Mediator, a megaDalton multiprotein complex, enables the regulation
of transcription; it bridges between gene activator proteins at
enhancers and RNA polymerase II (pol II) at promoters.
Pol II: DNA unwinding
RNA polymerization
proofreading
GTFs (TFIIB,-D,E,F,-H): promoter recognition
Mediator: interaction with activator proteins
and polII; essential for transcription

Strukturbiologie, Transcription 15
Structure determination of the polymerase in
the form of a transcribing complex (3.3Å)
(B) Comparison of structures of
free Pol II (top) and the Pol II
transcribing complex (bottom). The
clamp (yellow) closes on DNA
and RNA, which are bound in the
cleft above the active center. The
remainder of the protein is in gray.

16
DNA can be seen entering the
transcribing complex in duplex
form and unwinding three bases
before the active site. Then the
template strand makes a sharp
bend, and as a result, the next
base is flipped, pointing down
towards the active site. This base
is paired with that of the
ribonucleotide just added to the
RNA strand. The structure reveals
eight more DNA-RNA hybrid base
pairs and one additional base on
the template DNA strand. The
remainder of the template strand,
the RNA, and the nontemplate
DNA strand are not seen, due to
motion or disorder.
Crystal structure of the Pol II transcribing complex
Gnatt et al, Science 2001

Strukturbiologie, Transcription 17
Transcription Initiation mechanisms
How is straight duplex promoter DNA melted, bent, and inserted in the
Pol II active center, enabling the initiation of transcription?
Bushnell, D.A., et al. (2004) Structural basis of transcription: an RNA polymerase II-TFIIB
cocrystal at 4.5 Ångströms. Science
These DNA
transactions are
made by the GTF’s
TFIIB , -D, -E, -F, and
-H.
TFIIB stabilizes an
initial transcribing
complex and the N-
terminal region
forms Zn ribbon and
B finger.

18
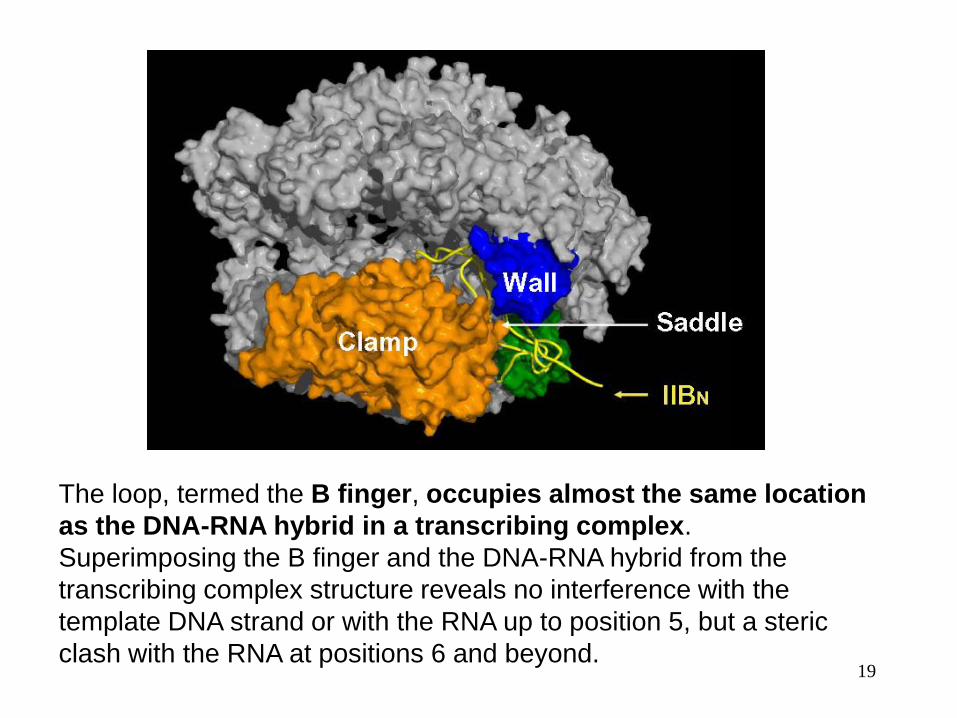
Transcription Initiation mechanisms
The structure of the Pol II-TFIIB complex revealed distinct functions of
the N- and C-terminal domains of TFIIB. The N-terminal domain
(yellow) begins with a Zn ribbon that binds the Pol II surface adjacent
to the clamp and wall.Then the polypeptide continues across the
saddle between the clamp and wall and plunges towards the active
center, from which it loops back and remerges across the saddle.
19
The loop, termed the B finger, occupies almost the same location
as the DNA-RNA hybrid in a transcribing complex.
Superimposing the B finger and the DNA-RNA hybrid from the
transcribing complex structure reveals no interference with the
template DNA strand or with the RNA up to position 5, but a steric
clash with the RNA at positions 6 and beyond.

Strukturbiologie, Transcription 20
B finger is not only compatible
with a hybrid containing five
residues of RNA, but is
required for stability of short
DNA-RNA complex (BiaCore
experiments).
When the RNA grows beyond five or six residues, however, it must
compete with TFIIB for space on the Pol II saddle. If TFIIB wins the
competition, initiation is aborted and must be tried again. If the RNA
wins, TFIIB is ejected and Pol II is released from the promoter to
continue and complete transcription.
The B finger thus explains two crucial but for a long time mysterious
aspects of Pol II transcription, abortive initiation and promoter escape. In
these respects, it resembles the sigma factor in bacterial transcription.

21
Model of open promoter complex
Structure of an RNA polymerase II-TFIIB complex and the transcription
initiation mechanism
Science, 2010, Kornberg Lab

Strukturbiologie, Transcription 23
Transcription Initiation – TATA box binding protein (TBP)
Sigler, Burley, 1993
TBP forms a saddle shaped molecule with stirrups. A b-sheet in TBP forms
the DNA-binding site. TBP binds in the minor groove (strong hydrophobic
interaction, H-bonds) and induces large structural changes in DNA (DNA
helices on both sides form an angle of ca. 110º.
TBP: highly conserved CTD (180 aa); non-conserved NTD
24
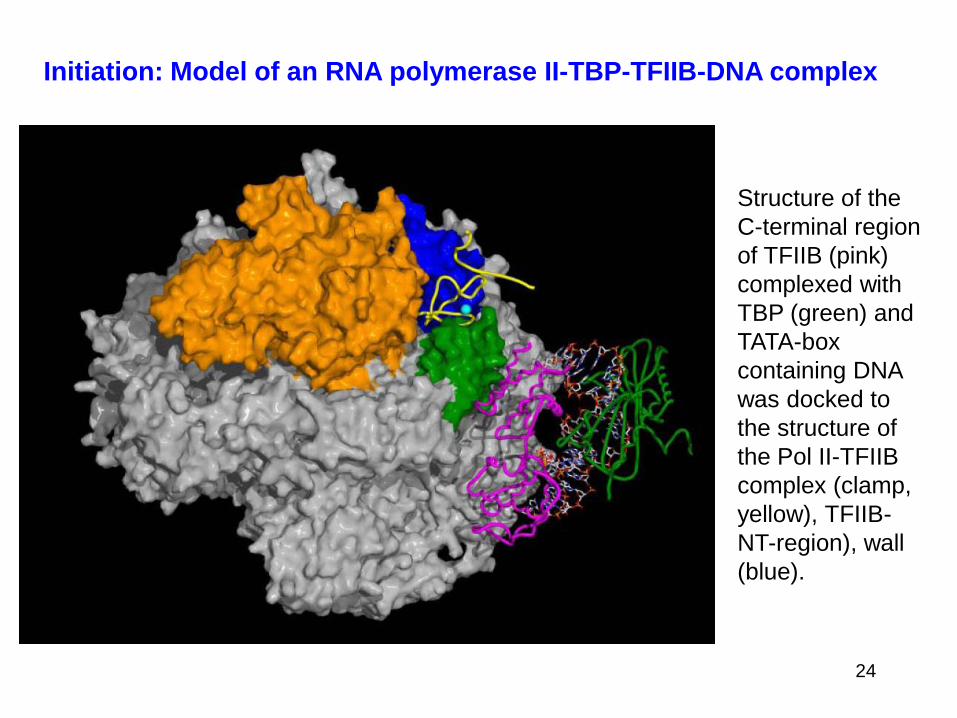
Initiation: Model of an RNA polymerase II-TBP-TFIIB-DNA complex
Structure of the
C-terminal region
of TFIIB (pink)
complexed with
TBP (green) and
TATA-box
containing DNA
was docked to
the structure of
the Pol II-TFIIB
complex (clamp,
yellow), TFIIB-
NT-region), wall
(blue).

25
Model of an RNA polymerase II-TBP-TFIIB-DNA complex
after adding
straight B-form
DNA:
TATA-box-
saddle: 15bp;
saddle-active
site: 12 bp
= ca 27 bp!!
distance TATA-
box to
transcription start
site in promoters
25-30 bp

Strukturbiologie, Transcription 26
Docking a complex of a C-terminal TFIIB fragment, the TATA-binding
protein (TBP) subunit of TFID, and a TATA box DNA fragment:
First, the DNA fit snugly against the protein: – TBP evidently
configures promoter DNA to the contours of the Pol II surface.
Second, the DNA downstream of the TATA box ran past the saddle.
The distance from the TATA box to the saddle is about 1.5 turns of the
double helix, or 15 base pairs (bp).
We know from the transcribing complex structure that about 12
residues are required to cross the saddle to the active site. The
sum of 15 bp from the TATA box and 12 residues to the active site is 27
bp, closely coincident with the spacing of 25–30 bp from the TATA
box to the transcription start site of almost all Pol II promoters. In
this way, Pol II-TFIIB interaction may determine the location of the
transcription start site.

Strukturbiologie, Transcription 27
Initiation: Transcription bubble (complex with TFIIF)
The structure includes a complete
transcription bubble – not only the
template DNA strand with
associated RNA, but also the
nontemplate DNA strand, and the
region upstream of the bubble
where duplex DNA is reformed
following transcription.
yellow: TFIIF; green: coding DNA;
red: RNA; cyan: template DNA
The interaction of the
nontemplate strand with TFIIF
may trap a transient bubble in
promoter DNA, leading to the
initiation of transcription.

Strukturbiologie, Transcription 28
Transcription initation
The structures
of Pol II, TBP, and
TFIIB come from
X-ray
crystallography.
The structures of
TFIIE, TFIIF, and
TFIIH (helicase)
are from electron
crystallography
and from cryo-
electron
microscopy and
single particle
analysis.

Strukturbiologie, Transcription 29
Transcription initation - Complete minimal RNA
polymerase II transcription initiation complex
TBP bends the promoter DNA around
the polymerase and the CTD of TFIIB.
The NTD of TFIIB brings the DNA to a
point on the polymerase surface from
which it need only follow a straight path
and, by virtue of the conserved spacing
from TATA box to transcription start site
in Pol II promoters, the start site is
juxtaposed with the active center.
TFIIE enters the complex and recruits
TFIIH, whose ATPase/helicase
subunit introduces negative
superhelical tension in the DNA.

Strukturbiologie, Transcription 30
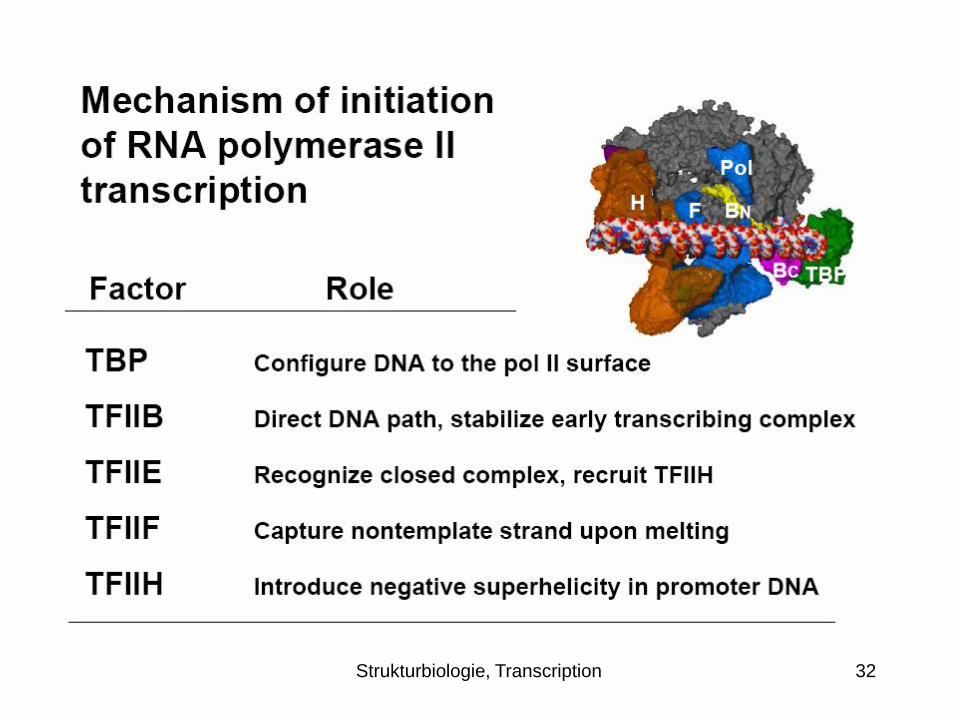
Transcription initation - Complete minimal RNA
polymerase II transcription initiation complex
Thermal unwinding produces a
transient bubble, which is captured
by TFIIF binding to the nontemplate
strand. The DNA can now bend in the
single stranded region and descend into
the Pol II active center.
Initiation and the synthesis of RNA
ensue, initially stabilized by the B finger.
Synthesis of a transcript greater than
about 6 residues in length leads to the
displacement of TFIIB, promoter
escape, and the completion of
transcription.
Strukturbiologie, Transcription 32

Strukturbiologie, Transcription 33
Other essential tasks of transcription:
Translocation
Nucleotide addition
Fidelity of Transcription
RNA escape
Regulation – the role of Mediator

Strukturbiologie, Transcription 34
Translocation: Bridge helix might serve as molecular ratched
Straight and bent states of the bridge helix in RNA polymerase II (yeast) and
bacterial RNA polymerase structures. The bend produces a movement of ≈3 Å in
the direction of the template strand, corresponding to one base pair step along
the strand.

Strukturbiologie, Transcription 35
A cycle of nucleotide addition by RNA polymerase II
At the upper left, the
structure of the
transcriping complex is
shown, omitting all but
the DNA and RNA near
the active center and
the bridge helix
(green).
The ribonucleotide in
the active center, just
added to the RNA
chain, is yellow.
At the lower left is the
structure after
translocation of DNA
and RNA across the
Pol II surface.

Strukturbiologie, Transcription 36
A cycle of nucleotide addition by RNA polymerase II
At the lower right is
the structure with an
unmatched NTP in
the entry (E) site. At
the upper right is
the structure with
NTP, matched for
pairing to the coding
base in the template
strand, in the
addition (A) site.

Strukturbiologie, Transcription 37
A cycle of nucleotide addition by RNA polymerase II
All four NTPs were
seen to bind an entry or
“E” site, whereas only
the NTP correctly
matched for base
pairing with the coding
base in the DNA was
seen to bind in the
active center, at the
nucleotide addition or
“A” site. The orientation
of NTP in the E site
was inverted with
respect to that in the A
site, leading to the
suggestion that NTPs
in the E site rotate to
sample base pairing in
the A site.

Strukturbiologie, Transcription 38
Bridge helix update
Cheung et al, Structural basis of initial RNA polymerase II transcription, EMBO
J, 2011

Strukturbiologie, Transcription 40
But 3D structure did not explain the fidelity of transcription: The
energy of base pairing, through two or three hydrogen bonds to the
template DNA, is far less than required to account for the selectivity of
the polymerase reaction.
2006:
New structures of RNA polymerase II (Pol II) transcribing complexes
reveal a likely key to transcription. The trigger loop swings beneath a
correct nucleoside triphosphate (NTP) in the nucleotide addition site,
closing off the active center, and forming an extensive network of
interactions with the NTP base, sugar, phosphates, and
additional Pol II residues. A His side chain in the trigger loop,
precisely positioned by these interactions, may literally “trigger”
phosphodiester bond formation. Recognition and catalysis are
thus coupled, ensuring the fidelity of transcription.

Strukturbiologie, Transcription 41
Fidelity of transcription: Trigger loop contacts NTP in the A site
Template DNA RNA
Trigger Loop
NTP in A site
(purine,
pyrimidine NT)
The trigger loop
swings beneath a
correct nucleoside
triphosphate
(NTP) in the
nucleotide addition
site, closing off the
active center, and
forming an
extensive
network of
interactions with
the NTP base,
sugar,
phosphates, and
additional Pol II
residues.

Strukturbiologie, Transcription 42
Fidelity of transcription: Trigger loop contacts NTP in the A site
Template
DNA RNA
Trigger
Loop
NTP in A site
(purine,
pyrimidine
NT)
A His side chain in
the trigger loop,
precisely positioned
by these
interactions, may
literally “trigger”
phosphodiester
bond formation.
Recognition and
catalysis are thus
coupled, ensuring
the fidelity of
transcription.
Strukturbiologie, Transcription 43
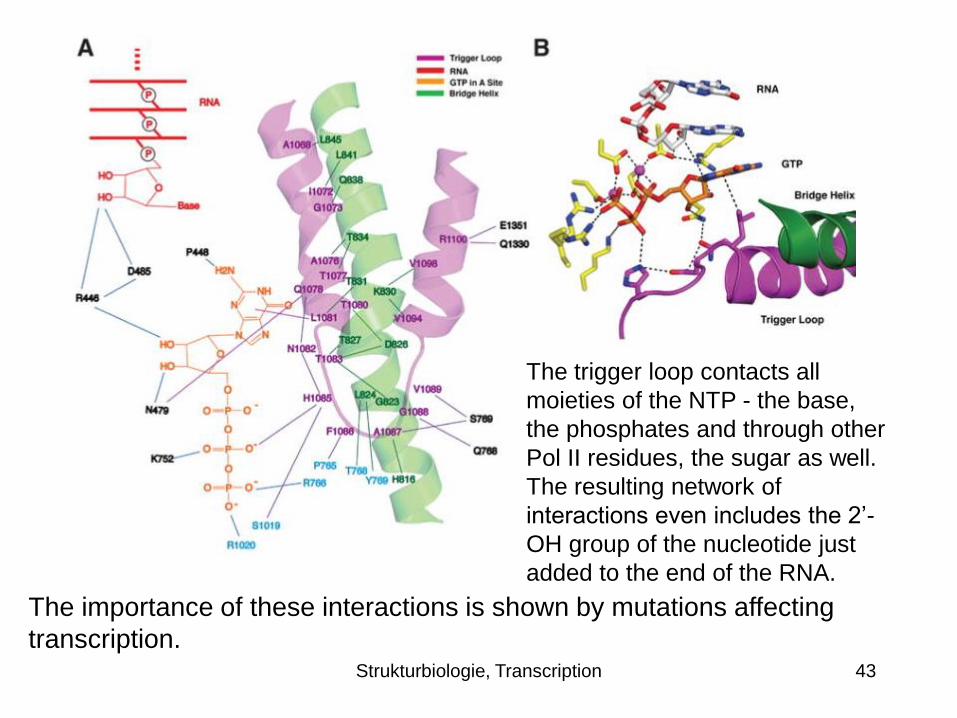
The trigger loop contacts all
moieties of the NTP - the base,
the phosphates and through other
Pol II residues, the sugar as well.
The resulting network of
interactions even includes the 2’-
OH group of the nucleotide just
added to the end of the RNA.
The importance of these interactions is shown by mutations affecting
transcription.

Strukturbiologie, Transcription 44
Trigger loop couples nucleotide selection to catalysis
Alignment of the trigger loop
with the NTP and the
precise positioning of a
histidine side chain, 3.5 Å
from the β-phosphate. The
histidine promotes the flow
of electrons during
nucleophilic attack of the 3′-
OH at the chain terminus
and phosphoanhydride bond
breakage. It serves as a
proton donor for the
pyrophosphate leaving
group. It literally triggers
phosphodiester bond
formation.

45
Nucleotide selection by alignment with the trigger
loop, coupling recognition to catalysis
The electronic transactions involved in trigger loop function require precise alignment of the
interacting moieties. This is achieved for a correct NTP by formation of the trigger loop
network. In the case of an incorrect NTP, for example a 2′-deoxy NTP, misalignment is
profound. A double helix formed with a 2′-deoxy nucleotide is 2 Å narrower than that formed
by a ribonucleotide.

Strukturbiologie, Transcription 46
Separation of RNA transcript from the template
- 3D structure in the posttranslocation state
Westover, K.D., et al. (2004) Structural basis of transcription:
separation of RNA from DNA by RNA polymerase II. Science.
-7
-8
-9
-10
Fork
loop
Rudder Lid
Release of RNA transcript
from DNA -RNA hybrid
revealed in the structure of
an RNA polymerase II
transcribing complex. The
upstream end of the DNA -
RNA hybrid helix, 7-10
residues from the active
center, is shown on the
left, with distances
between the DNA and RNA
bases indicated. The entire
DNA -RNA hybrid helix is
shown on the right, along
with protein loops involved in
helix melting (rudder and lid)
and stabilization (fork loop).

Strukturbiologie, Transcription 47
Separation of RNA transcript from the template
- 3D structure in the posttranslocation state
Westover, K.D., et al. (2004) Structural basis of transcription:
separation of RNA from DNA by RNA polymerase II. Science.
-7
-8
-9
-10
Fork
loop
Rudder Lid
Base pair 7 of the DNA-
RNA hybrid in this
structure appears normal
– the bases are
coplanar, with a distance
appropriate for hydrogen
bonding between them.
Base pairs 8, 9, and 10,
however, show
increasing deviations,
and consequent splaying
apart of the DNA and
RNA strands. The strand
separation is due to the
intervention of three
protein loops, termed
fork loop 1, rudder, and
lid.

Strukturbiologie, Transcription 48
Separation of RNA transcript from the template
- 3D structure in the posttranslocation state
Westover, K.D., et al. (2004) Structural basis of transcription:
separation of RNA from DNA by RNA polymerase II. Science.
-7
-8
-9
-10
Fork
loop
Rudder Lid
Rudder and lid lie
between DNA and RNA.
Rudder contacts DNA,
Lid RNA. A Phe side
chain of the lid serves as
wedge to maintain
separation of the strands.
Fork loop contacts the
sugar-phosphate
backbone of the hybrid
helix at base pairs 6 and
7, stabilizing the helix,
preventing the DNA-RNA
hybrid from unraveling
further and inhibiting
transcription.

Strukturbiologie, Transcription 49
Transcription
regulation:
the role of
Mediator
Mediator is a key
regulator of eukaryotic
transcription,
connecting activators
and repressors bound
to regulatory DNA
elements with Pol II.

Strukturbiologie, Transcription 50
Transcription regulation: the role of Mediator
Cryo-EM structure, 35 Å resolution, Asturias Lab
2002; Extension of the structure to atomic
resolution will one day reveal the regulatory
mechanism…
In the yeast Saccharomyces
cerevisiae, Mediator comprises
25 subunits with a total mass of
more than one megadalton and is
organized into three modules,
called head, middle/arm and tail.
Architecture of the Mediator head module, nature
2011; x-ray structure of mediator head; 4.3 A

51
Transcription regulation: the role of Mediator
In the yeast Saccharomyces cerevisiae, Mediator comprises 25 subunits with a
total mass of more than one megadalton and is organized into three modules,
called head, middle/arm and tail.
Structure of the Mediator head module:
Laurent Larivière, et al, Nature, 492, 448–451, (20 December 2012); 3.4 Å





























