Embed Size (px)
Citation preview

Présenté pour l’obtention du diplôme de
Magister
Option : Biologie moléculaire et génétique des microorganismes
Président : Pr Senhadji R Examinateur: Pr Fortas Z Examinateur: Pr Belahcene M Encadreur : Pr Bensalah F
Année Universitaire
Recherche des activités antichez les espèces du
isolées de différents biotopes
Mémoire
Présenté pour l’obtention du diplôme de
Magister en Biologie
Option : Biologie moléculaire et génétique des microorganismes
Par
Thème
Université Es-senia OranUniversité Es-senia Oran
Centre Universitaire Ain TimouchentUniversité Es-senia Oran
Année Universitaire
2013-2014
Recherche des activités anti-pathogèness espèces du genre Streptomyces
de différents biotopes
Option : Biologie moléculaire et génétique des microorganismes
senia Oran senia Oran
Universitaire Ain Timouchent senia Oran
pathogènes Streptomyces
BELBELABEDABED BeBellallall
Membres de jury : Soutenue 09 Mars 2014

Remerciements
Ce travail a été effectué au sein du laboratoire de génétique Microbienne, faculté des sciences de la
nature et de la vie, département de biologie, université d’Es-senia Oran.
Je tiens à remercier d’abord, Monsieur le Professeur Bensalah. F qui a bienvoulu diriger ce
travail, pour ses conseils qui m’ont permis de bien mener ce travail.
Je remercie également monsieur le Professeur Oueld Kadda qui n’a pas hésité à tout moment
d’offrir son aide scientifique et ouvrir les portes de son laboratoire à moi et à tous mes collègues, sans
oublier son étudiante Mlle Nabila qui nous a donné l’aide technique nécessaire.
Mes remerciements les plus respectueux s’adressent aussi à Monsieur SENHADJI R, Professeur
à l’université de d’Es-senia qui me fait l’honneur de présider le jury.
Mes plus vifs remerciements s’adressent aux Professeurs : Madame FORTAS Z et Monsieur
BELAHCENE M pour avoir accepté d’être membres de ce jury.
J’associe à mes remerciements à l’équipe du laboratoire de Génétique Microbienne Mlle
Bekenniche et Mme Kouadri pour leur soutien et leur solidarité.

Dédicaces
Je dédié ce modeste travail :
À mes parents, mais aucune dédicace ne serait témoin de mon profond amour, mon immense
gratitude et mon plus grand respect ;
A mes chers frères et sœurs : Amine, Abderrahmane, Haoua et Mekka ;
A mes chers Amis : Abderrahmane, Toufik, Hamza, Kadda, Younes, Zoheir, Boubaker et
Chouaib ;
A mes sœurs et collègues Nahla et Nacima, avec qui j’ai passé les meilleurs 3 ans de ma vie ;
A mes chers enseignants sans exception.

Résumé :
La présence des Streptomycètes dans les différents sols de la région ouest de l’Algérie a
été investiguée par l’isolement de ce genre bactérien à partir des échantillons de sols prélevés
de différents biotopes ; Cet isolement a été fait sur plusieurs milieux de culture comme ISP2,
GLM et Bennett. La mise en évidence de l’activité antibactérienne (premier criblage) des 85
souches isolées et suspectées être Streptomycètes a été faite par la méthode de la double
couche ; plus de 25% des souches isolées avaient un effet antibactérien contre au moins une
seule souche bactérienne pathogène et seulement 3 ont été sélectionnées pour les prochains
tests.
L’extraction des métabolites secondaires des souches sélectionnées (SM2.2, SM1B1 et
TB1) a été faite par solvant (chloroforme) et leur concentration par un Rotavapor.
L’évaluation de l’effet antibactérien de l’extrait était réalisée par la méthode des disques
contre les mêmes souches pathogènes utilisées dans le premier criblage. Seule l’isolat TB1 a
présenté un effet satisfaisant lors du deuxième criblage. Les différents extraits ont subis une
analyse UV-visible ainsi qu’une pré-identification des molécules présentes par
chromatographie sur couche mince (CCM), là où les taches des molécules ont été bien claires
surtout pour l’isolat TB1.
L’extraction de l’ADN des souches sélectionnées a été faite par un kit, et une
amplification (PCR) du gène ADNr 16S a été réalisée avec un programme approprié, des
amorces universelles ont été utilisées et la détection sur gel d’agarose a donné des fragments
de 1500 pb référant à la longueur du gène amplifié.
Mots clé : Streptomycètes, activité antibactérienne, extraction, CCM, PCR.

Abstract :
The presence of Streptomyces in different soils of the Western region of Algeria was
investigated by the isolation of these bacteria from soil samples from different biotopes; the
isolation was made in various media like ISP2, Bennett and GLM. The demonstration of
antibacterial activity (first screening) of the 83 isolates suspected to be Streptomyces was
made by the method of the double layer; more than 25% were actives against at least one
pathogenic strain and only 3 were selected for the next tests.
The extraction of secondary metabolites of the selected strains was made by solvent
(chloroform) and their concentration by a Rotavapor. The evaluation of the effect of the
extract was applied by the method of the discs against the same pathogenic strains used in the
first screening and only the TB1 strain showed an effect in the second screening. The various
samples went through UV-visible analysis and a pre-identification of the molecules present,
by a thin layer chromatography (TLC), where the spots were very clear.
The selected strain’s DNA extraction was made by a kit, and amplification (PCR) of the
16S rDNA gene was carried out with universal primers, in order to make a nucleic acids
chromatography on Agarose gel, it gave fragments of 1500 bp referring to the length of the
amplified gene.
Key words: Streptomyces, antibacterial activity, extraction, TLC, PCR.

:اصخ لم ل
وتسم ىلعىفلتخم أ ونا عا رغلاب ةبرتألا ب زجل ارئيمت Streptomycesاون ایریتكبلع ووج دانع ثحبل
، ISP2ھذا ا زعللىلع مت وأ اسطاھنم ةفلتخم ومن . ازعب ھنم ققحتلھ ل ذا ا ونلعنم وأ اسطةبرتلل ةفلتخم
Bennettو ،GLM . وأابتخر(اةیریتكب دض ةیلاعف نع ثحبل زعم ایریتكبوةلا وكشملكاھنوك يف 85ل لـ ) ا ل
Streptomyces ایریتكبا زعملوةلأ رھظتدض ةیلاعف نما ل % 25أنم رثك . وب متاةینقت ةطسا ةقبطلا زملودةج
ارةقحالل 3وس ى اا یتخ ررضمو ،ة متی مل ىلع ةیریتكبا ىلع لقألاو دحةنم ا ایریتكبلا ل .ابتخالل اھنما ت
وز یكرت مت ) chloroforme(اابكرملا ت ایر یتكبلل ةیو ناثلا اتخمل،ةروب مت اولحم ةطسلبیذم اال خت سص
ا . Rotvaporاابكرملتوب ااھج ةطسز إابثتةیلاعف ا قیرط نع تمت صلختسملا امعتسلةقیرط ا رقألو ص
و ال خل جـدی ة لیة أرھظتاعف TB1طقفا ایریتكبل . سفن دضا ایریتكبلا رضملا ة يف ةلمعتسملا ابتخالا ر ألل
وذك ا ) مرئي -وف امقي جسفنب (احسام ةطس نیوكت لیلحتا اصلختسملا ت الثلثو ب اابتخالا ر يناثلو ، مت دق
. وبارك لیلحت ةطسورغوتامايف احموةلا ققحتلا ألوون نم يلا ع ابكرملا ت اصلختسملل ةنوك ملت
وایریتكبلل ا اتخملةروب مت اطساضمحلا ل اال خت سص اودریثكت مت و مجمعنم ا أل ةونی ر ،ت دعدخسن ا وملث
ADNr 16S اةیملاع و مت رذليوی اھلوط يف قف ووت جمانرب قفايقفو امعتساب لرخذم ت وط فشكا ل وملا ث
. امھ وجوم وديف ا رملاعج
رحسم ،ریثكت : ااملكلا ت ةیحاتفمل ةیر یتكب دض ةیلاعفا ، الختس،صوم ،ث ،.

Liste des Abréviation
Liste des Abréviations :
ADN : Acide Désoxyribonucléique
ARNr : Acide Ribonucléique Ribosomique
ATCC : American Type Culture Collection
ATP :Adénosine triphosphate
BET : Bromured’Ethidium
CCM : Chromatographie sur Couche Mince
CCR : Répression du Catabolite Carboné
CG : Chromatographie à phase Gaseuse
ddNTP :didésoxycléoside triphosphate
dNTP :désoxycléoside triphosphate
EDTA: Ethylene diamine Tetra-acetic acid
GOGAT : Glutamate : 2-OxoGlutarate TransAminase
GS : Glutamate Synthétase
HPLC : Chromatographie Liquide à Haute Pression
mM : milli-molaire
NCTC : National Culture Type Collection
pb : paire de bases.
PCR : Polymerization Chain Reaction
pH : Potentiel d’Hydrogène
qsp : Quantité suffisante pour
Rf ; Rapport frontale
rpm : Rotation par minute
UFC : Unité Formant Colonie
UI : Unité Internationale
V/V : Volume par volume
G+C : Guanine + Cytosine
NCTC : National Culture Type Collection

Liste des tableaux
Liste des tableaux
Tableau 01 : Tableau illustrant les différentes propriétés des Streptomycètes 07
Tableau 02 : Les différentes méthodes moléculaires d’identification des Streptomycètes 12
Tableau 03 : représentation des différents milieux de culture utilisés pour la culture des
Streptomycètes 20
Tableau 04 : Principales différences entre les métabolites primaires et secondaires 23
Tableau 05 : Propriétés physicochimique des solvants utilisés dans l’extraction des
métabolites secondaires 26
Tableau 06 : Tableau qui présente les antibiotiques les plus utilisés 29
Tableau 07 : Lieux de prélèvement des échantillons 34
Tableau 08 : Quelques caractéristiques des souches pathogènes utilisées dans cette étude 39
Tableau 09 : Les ingrédients de la PCR pour chaque échantillon 48
Tableau 10 : Tableau représentant les caractéristiques et le diamètre des zones d’inhibition
en millimètre des souches sélectionnées lors du criblage primaire 63
Tableau 11 : Résultats des tests des 3 souches 67
Tableau 12 : Les pics des courbes d’absorbance des extraits 72

Liste des figures
Liste des figures
Figure 1 : diagramme d’une section verticale à travers le centre de sporulation de S. coelicolor
6
Figure 2 : Colonies de l’espèce type Streptomyces coelicolor 6 Figure 3 : endommagement de la pomme de terre à cause de S. scabies 9 Figure 4 : classification du genre Streptomyces 12 Figure 5 : Schéma du cycle de vie des Streptomyces dans les conditions neturelles
13
Figure 6 : Cycle de vie des Streptomyces 14 Figure 7 : Morphologies rencontrées aux cours de cultures liquides 15 Figure 8 : Couleur du mycélium aérien et des pigments diffusible S. glaucescens
16
Figure 9 : Morphologie du mycélium aérien de Streptomyces 16 Figure 10 : hyphes végétatives et conidiospores de S. coelicolor 17 Figure 11 : développement du mycélium secondaire en spores 17 Figure 12 : Schéma général conduisant à l’obtention des antibiotiques à l’état pur
28
Figure 13 : lieu de prélèvement Ech 02 35 Figure 14 : lieu de prélèvement Ech 03 35 Figure 15 : dilution des échantillons 36 Figure 16 : Rotavapor utilisé pour la concentration des métabolites secondaires
44
Figure 17 : illustration de la CCM 46 Figure 18 : thermocycler (TECHNE TC-312) 49 Figure 19 : exemple sur le résultat d’une éléctrophorèse des acides nucléiques
50
Figure 20 : électrophorèse pour la séparation des fragments d’ADN 51 Figure 21 ; plaque UV 52 Figure 22 : les colonies suspectées être des Streptomycètes 54 Figure 23 : colonies de Streptomyces 54 Figure 24 : observation macroscopique de SB1 54 Figure 25 : observation de la colonie de SB1 54 Figure 26 : observation macroscopique de CB5 55 Figure 27 : observation de la colonie de CB5 55 Figure 28 : observation de la colonie de CQ 55 Figure 29 : observation macroscopique de CQ 55 Figure 30 : observation de la colonie de SB4 55 Figure 31 : observation macroscopique de SB4 55 Figure 32 : observation de la colonie de BN1 55 Figure 33 : observation macroscopique de BN1 55 Figure 34 : observation de la colonie de N3 56

Liste des figures
Figure 35 : observation macroscopique de N3 56 Figure 36 : souche SM2.2 56 Figure 37 : souche BN1 56 Figure 38 : souche SM2.6 56 Figure 39 : souche 2.11 56 Figure 40 : souche SM2.3 56 Figure 41 : souche SM X4.2 56 Figure 42 : souche SM2.8 57 Figure 43 : souche B6 57 Figure 44 : pigments diffusibles produises par les souches de Streptomycètes 58 Figure 45 : Pourcentage des souches isolées selon les types de sol 59 Figure 46 : Coloration de Gram des souches 60 Fihure 47 : l’activité antibactérienne des souches isolées 62 Figure 48 : la non-hydrolyse de l’amidon 64 Figure 49 : la non dégradation de la caséine 64 Figure 50 : l’absence de l’uréase chez SM2.2 65 Figure 51 : l’absence de l’uréase chez TB1 65 Figure 52 : la non dégradation du tween 80 par les 3 souches 65 Figure 53 : dégradation des sucres par la souche SM2.2 66 Figure 54 : extraction des métabolites secondaires 68 Figure 55 : effet des l’extraits sur E.coli 69 Figure 56 : courbe d’absorbance de l’extrait de la souche TB1 70 Figure 57: courbe d’absorbance de l’extrait de la souche SM2.2 71 Figure 58 : courbe d’absorbance de l’extrait de la souche SM1B1 71 Figure 59 : Visualisation de la CCM avec UV de l’extrait de la souche TB1 73 Figure 60 : Chromatographie des extriats des souches SM2.2 et SM1B1 75 Figure 61 : électrophorèse des acides nucléiques sur gel d’Agarose à 1.5% 76

Table de matière
Table de matière
Introduction 1
Partie bibliographique
1 Les Streptomyces : 4
1.1 Les actinomycètes 4
1.2 Historique 4
1.3 Définition 5
1.4 Ecologie des Streptomycètes 7
1.5 Les Streptomycètes comme pathogène 9
1.6 Taxonomie des Streptomycètes 10
1.7 Cycle de développement 13
1.8 Sporulation et germination 16
1.9 Les milieux de culture des Streptomycètes 18
1.9.1 Source de Carbone 18
1.9.2 Source de nitrogène 19
2 Les substances bioactives 21
2.1 Définition 21
2.2 Les métabolites secondaires 21
2.3 Métabolisme des Streptomyces 22
2.4 Métabolisme secondaire des Streptomyces 22
2.5 Production d’antibiotiques par les Streptomycètes 23
2.5.1 Effet de la composition du milieu de culture 24
2.5.2 Effet du pH, de la température, de l’agitation et du temps d’incubation 25
2.5.3 Méthodes d’extraction des métabolites secondaires 26
2.5.4 Purification et Caractérisation métabolites secondaires 27
2.5.5 Production des antibiotiques 27
3 Génétique des Streptomycètes 30
3.1 Le chromosome 30
3.2 Instabilité génétique des Streptomycètes 31
3.3 Plasmides circulaire et linéaire 31
3.4 Identification moléculaire des Streptomycètes 32
Partie expérimentale
1 Prélèvement 34
2 Pré-enrichissement des échantillons 35
3 Dilution et ensemencement 35

Table de matière
4 Isolement des streptomycètes 36
4.1 Purification des souches 37
4.2 Determination de la couleur 37
4.3 Coloration de Gram 37
4.4 Conservation des souches 38
5 Mis en évidence de l’activité antibactérienne des souches isolées 38
5.1 Souches de Streptomyces utilisé 38
5.2 Souches pathogènes 38
5.3 Criblage primaire 49
5.4 Criblage secondaire 40
6 Identification des souches sélectionnées 41
6.1 Tests physiologiques 41
6.1.1 Hydrolyse de l’amidon 41
6.1.2 Hydrolyse de la caséine 41
6.1.3 Dégradation de l’urée 41
6.1.4 Dégradation du tween 80 41
6.2 Tests biochimiques 42
6.2.1 Utilisation des différentes sources de carbone 42
7 Extraction et concentration des métabolites secondaires 42
7.1 L’extraction 42
7.2 Concentration par Rotavapor 43
8 Spectre UV des extraits 44
9 Chromatographie sur couche mince (CCM) 45
10 Identification moléculaire des souches 46
10.1 Extraction d’ADN 46
10.2 La PCR 48
10.3 Electrophorèse des acides nucléiques 49
10.3.1 La préparation du gel d’agarose 49
9.3.2 Dépôt des échantillons 50
9.3.3 Migration 51
9.3.4 Visualisation 52
Résultats et discussion
1 Effet des prétraitements 53
2 Isolement des Streptomyces 53
2.1 Détermination des couleurs 54

Table de matière
2.1.1 Couleurs du mycélium du substrat 54
2.1.2 Les couleurs du mycélium aérien 56
2.1.1 Production des pigments diffusibles 57
2.2 Dénombrement des souches suspectées etre des Streptomycètes 58
3 Mise en évidence de l’activité antibactérienne des souches isolées 61
4 Identification des souches sélectionnée 64
4.1 Tests physiologiques 64
4.1.1 L’hydrolyse de l’amidon 64
4.1.2 Hydrolyse de la caséine 64
4.1.3 Dégradation de l’urée 65
4.1.4 Dégradation du tween 80 65
4.2 Tests biochimiques 66
5 Extraction des métabolites secondaires 68
6 Evaluation de l’effet des extraits 69
7 Courbes d’absorbance des extraits 70
8 Chromatographie sur couche mince 73
9 Electrophorèse des acides nucléiques 76
Conclusion et perspectives 78
Références 80
Annexes 95

Introduction

Introduction
1
Introduction :
La découverte de la tyrothricine par Dubos en 1939 à partir d’une culture de Bacillus
brevis fut le point de départ de recherches systématiques sur la production d’antibiotiques par
les bactéries. De nombreuses souches, particulièrement de germes sporulés, isolés de
différents milieux, furent étudiées durant la décade qui suivit la découverte de cet
antibiotique. Ces antibiotiques avaient un large spectre d’action mais ils été toxiques, ce qui a
pousser les chercheurs vers la prospection des champignons et surtout des actinomycètes :
source de tous nos antibiotiques majeurs. Chaque année encore de nombreux antibiotiques
nouveaux sont isolés à partir des cultures de Streptomyces (Kauffmann, 1962).
Le besoin permanent du développement de nouveaux antibiotiques pour le traitement des
infections bactériennes sévères est évident. En effet, dès qu’une nouvelle molécule est utilisée
en pratique clinique, la résistance ne tarde pas à émerger dans les années qui suivent. Depuis
plus de 20 ans, de nombreux déterminants de résistance ont été identifiés avec l’émergence de
bactéries de plus en plus résistantes. L’impact de cette multi-résistance aux antibiotiques est
important au niveau clinique en termes de morbidité et de mortalité mais aussi sur le plan
économique (durée d’hospitalisation et d’antibiothérapie prolongées) et sur l’écologie des
flores microbiennes (Cattoir & Daurel, 2010).
L’importance thérapeutique des antibiotiques est non discutable, mais n’oublions pas
l’importance économique de l’industrie pharmaceutique basée sur la production de ces
molécules. Il a été estimé que le marché pharmaceutique international est de 60 milliard de
dollar en 2006 (Pearce et al., 2009).
La découverte des antibiotiques produits par les Streptomycètes dans les années 40 du
dernier siècle qui a débutée par le travail de Waksman, a conduit à un criblage extensif de
nouvelles molécules bioactifs et en conséquence ça a donné une sur-classification de ce genre.
Les souches décrites du genre Streptomyces ont évoluées de 40 à plus de 3000 ; plusieurs de
ces souches ont été similaires. De cet effet il été essentiel de mettre des critères bien
déterminés pour l’identification de ce genre bactérien et c’était l’ISP (International
Streptomyces Project). Ça a bien reflété l’importance et l’utilité des Streptomycètes.
En 1930, le microbiologiste allemand Lourens G. M. Baas Becking a dit que « toute chose
est partout, mais l’environnement la choisit », et Mr Noah Fierer de l’université de Colorado
dit « probablement on connait les organismes des océans profonds mieux que les organismes

Introduction
2
qui vivent dans le sol et dans nos jardin » (Pearce et al., 2009). De cet effet le sol est un
milieu très riche en microorganismes et il constitue l’habitat le plus adéquat pour les
Streptomycètes. Selon les estimations 50 % des métabolites secondaires bioactifs depuis
1900 sont produits par les Streptomycètes du sol (Flavia, 2009).
Alors l’exploitation des ressources terrestres non-exploités en Streptomyces jusqu’à
moment reste un souci pour les chercheurs, vue leurs réserve en métabolites secondaires qui
est généralement dû à la taille du génome, ainsi qu’à la forme de ce génome qui est linéaire et
aussi aux mutations qui se manifestent au niveau de l’ADN de ce genre bactérien.
Les travaux qui ont pour but l’identification de nouvelles souches, c’est rarement où ils
peuvent identifier de nouvelles souches ; mais les travaux puissants dans le domaine
d’innovation et de la recherche des nouveaux métabolites secondaires bioactifs ciblent surtout
des souches de Streptomyces bien connues et bien identifiées comme S. coelicolor, S. viridans
…etc.
Le premier but de ce travail est d’explorer le sol algérien et spécialement celui de la région
ouest de l’Algerie en Streptomyces et de mettre en évidence l’activité antibactérienne des
isolats présents dans les différents types de sol, ainsi de les identifier à base de la biologie
moléculaire et l’étude de leurs caractères physiologiques et biochimiques.
Le second but est d’extraire les métabolites secondaires issus des cultures des souches et de
les pré-identifiés afin d’essayer de classer ces métabolites.
Pour cela, on a divisé ce mémoire en plusieurs partie : la partie « bibliographique », là où
on décrit les Streptomyces, leur taxonomie, écologie, mode de reproduction, caractères
culturaux ; les substances bioactives, définition, métabolites secondaires, métabolites
secondaires des Streptomycètes. Après on décrit la production des antibiotiques et enfin la
génétique des Streptomycètes et les différentes méthodes d’identification moléculaire des
souches.
La deuxième partie est la partie « matériels et méthodes », là où les différentes
méthodologies utilisées dans ce modeste travail sont décrites ou illustrées, soit l’isolement,
l’identification ou l’extraction des métabolites secondaires.

Introduction
3
La troisième partie est la partie « Résultats et Discussion », où tous les résultats des
travaux sont présentés et discutés, par rapport à des travaux faits auparavant où bien analysés
selon les informations disponibles.
La fin de ce travail est une conclusion et des perspectives par rapport à ce qui été souhaité ou
bien à ce qui parait intéressant de le faire dans le prochain futur.

Bibliographie

Bibliographie
4
1 Les Streptomyces 1.1 Les actinomycètes
Les bactéries Gram positif contiennent 2 branches majeurs : les microorganismes à une
valeur G+C faible, contenant les genres tel que Bacillus, Clostridium, Staphylococcus et
Streptococcus ; et les microorganismes à une valeur G+C élevée, sont ceux qui référent aux
actinomycètes. Plusieurs développent un mycélium qui est une marque typique des
actinomycètes au moins dans certains stages de leur vie, mais il y’en a d’autres qui ne le font
pas(Kieseret al., 2000). Cette famille a été créée par Buchanan en 1918 (Dworkinet al.,
2006c).
Auparavant les Streptomycètes ont été un genre intégré des Actinomycètes. Tout un
ensemble d’outils taxonomiques sont utilisés maintenant pour définir les genres et les
différents groupes des Actinomycètes, mais l’analyse séquentielle de l’ARN 16S est le plus
significatif(Kieseret al., 2000).
1.2 Historique
L’histoire des Streptomycètes a commencé avec la découverte du premier antibiotique
issue de ce genre, qui été la streptothricine en 1942 et la découverte de la streptomycine deux
ans plus tard, ça a permis à Mr WaksmanSelmand’avoir le prix Nobel de médecine en 1952
(Emerson et al., 2012 ; Pearce et al., 2009). En 1943 Waksman et Hinrici ont établi la
famille des Streptomycetaceae(Kämpfer, 2006). Alors les premières investigations sur les
actinomycètes incluant les Streptomycètes ont été fortement basées sur la morphologie, qui
été considérée comme l’élément suffisant pour la classification en genres et en
familles(Dworkinet al., 2006c). Ça a conduit à une augmentation du nombre des souches
appartenant à ce genre ; mais plusieurs ont été similaires. Il était nécessaire de mettre des
critères d’identification standards pour éviter la sur-spécification. En 1964, le Projet
Internationale Streptomyces (International Streptomyces Project) « ISP » a été initié pour
introduire des critères standards pour la détermination des espèces et pour diminuer le nombre
des souches qui sont similaires(Annaliesaet al., 2001).
Shirling& Gottlieb(1966) ont décrit les critères standards qui incluent la morphologie des
spores, forme de la surface des spores, la couleur de spores, le mycélium végétatif et les
pigments solubles, production de mélanine et l’utilisation des différentes sources de
carbone(Annaliesaet al., 2001).

Bibliographie
5
Puis les groupes de ce genre ont été divisés selon l’utilisation des différentes sources de
carbone ; ensuite Williams et al(1983)ont proposé la classification numérique (Anderson &
Wellington, 2001). Parmi les programmes utilisés pour cette classification : CHARSEP qui
donne un pourcentage de ressemblance aux critères essentiels du genre (%) et le DIACHAR
qui est basé sur un score (points) et il y’en a un autre programme « BEST » (Brun, 2003).Puis
la classification phylogénétique a pris sa place pour l’identification des streptomycètes,
surtout le séquençage de l’ADNr 16S (Kieseret al., 2000)
1.3 Définition
Les Streptomycètes (du grec Streptos : courbé et Myces : moisissure) (Colombié, 2005)
sont des bactéries Gram positif, aérobies, membres de l’ordre Actinomycetales et la classe
Actinobacteria, elles ont un ADN qui contient un pourcentage G+C qui varie entre 69 et 78
mol%. Les Streptomycètes produisent un substrat extensif ramifié et un mycélium aérien
(Anderson & Wellington, 2001).
Bien qu’ils aient une structure de procaryotes, leur cycle biologique est semblable à celui
de certains champignons et peut conduire à la formation de spores. Ces spores sont isolées,
groupées en chaines ou contenues dans un sporange. De forme variable, elles peuvent être
flagellées ou ornementées. Ces spores permettent la propagation de l’espèce et la survie dans
les conditions défavorables. Dans certains cas il n’y a pas de production de spores, mais le
mycélium se fragmente et donne naissance à de nouveaux hyphes (Saffroy, 2006).
Le genreStreptomyces représentent le genre majoritaire des Streptomycètes (95,34%) il
s’agit de bactéries du sol filamenteuses, dont les hyphes, de longueur variable, ont un
diamètre compris entre 0,5 et 2,0 µm ; quelques espèces sont pathogènes pour l’homme et les
animaux et on trouve des autres qui sont phyto-pathogènes (Saffroy, 2006).
Ils synthétisent le géosmine qui est une molécule volatile, donnant au sol son odeur
typique. David Hopwood(2007) été le premier responsable du choix de S. coelicolor comme
la souche typique des Streptomycètes, motivé par le fait qu’elle produit 2 métabolites
secondaires pigmentés qui sont : prodiginines (Rouge), l’actinorhodine (Bleu) (Dyson, 2011),
et d’autres métabolites comme : l’anthracyclinglycoside, diazaindophenol, naphthoquinone,
phenoxazinone, et prodigiosine(Kutzner, 1981 in Kämpfer, 2006).

Bibliographie
6
Figure 1 : diagramme d’une section verticale à travers le centre de sporulation de S. coelicoloren noir les cellules intactes et en blanc les cellules désintégrées (Dworkinet al., 2006b).
Figure 2 : Colonies de l’espèce type Streptomyces coelicolor. Andrew Davis, John Innes Centre, Norwich (Flavia, 2009)

Bibliographie
7
Tableau 01 : tableau illustrant les différentes propriétés des Streptomycètes (Goodfeilowet al., 1992)
Caractéristiques Propriétés Demande du genre Aérobie Cycle de vie L’hyphe végétatif produit un mycélium extensivement ramifié.
Reproduction par germination de spores formées du mycélium aérien. A la maturité le mycélium aérien se différencie en 3 spores ou plus, sa surface peut être chevelue, rugueuse, régulière et avec contour.
Pigmentation Produisent une large gamme de pigments responsable de la couleur du substrat et du mycélium aérien. Des pigments diffusibles colorés peuvent être formés.
Colonies De discrète et lichenoïde, Demandes nutritives Beaucoup de molécules utilisées comme sources de carbone du sol. Peptidoglycane de la paroi Contient en majorité le LL-diaminopimelique. Acides gras Mélange complexe de molécules iso et antiso saturés. Lipides polaires Composition complexe, qui regroupe : diphosphatidylglycerol,
phosphatidylethanolamine, phosphatidylinositol et phosphatidylinositolmannosides.
La composition de l’ADN 69-78 % mol G+C.
1.4 Écologie des Streptomycètes
Les streptomycètes sont ubiquitaire dans la nature ; leur aptitude de coloniser le sol est très
facilitée par le développement comme des hyphes végétatives et qui peuvent se différencier en
spores qui se dispersent et assurent la persistance. Des cultures de streptomycètes ont été
revivifiées à partir des échantillons de sol de 70 ans ! Plusieurs bactéries de ce genre peuvent
aussi coloniser le rhizosphère, ça est due à leur caractère d’antagonisme contre des autres
bactéries comme Pseudomonas et Bacillus(Kieseret al., 2000).
Le sol contient 104 à 107 UFC de streptomycètes par gram, représentant 1-20% ou même
plus de la totalité de la flore bactérienne (Donadioet al., 2002). Dans les sols riches en
végétation ou ceux riches en matière organique il y’en a un grand nombre de Streptomycètes.
Comme les Streptomycètes du sol, les Streptomycètes aquatiques constituent eux mêmes un
pourcentage assez important, Alan &Stach (2007) estiment le nombre des actinomycètes est
de 35% de la flore bactérienne des larges zones des océans.

Bibliographie
8
La plupart des Streptomycètes peuvent dégrader pas mal de matériaux de plante et
d’animaux comme les polysaccharides (ex : amidon, pectine, cellulose et chitine), protéines
(ex : kératine et l’élastine), lignocellulose et des autres composés
aromatiques(Yikmis&Steinbüchel, 2012).
Les membres du genre Streptomyces sont impliqués dans la biodégradation de plusieurs
polymères qui sont abondants dans le sol ; en basant sur leur capacité de produire des
enzymes extracellulaires, les streptomycètes sont l’un de peu de bactéries qui peuvent
dégrader la lignine (Dworkinet al., 2006c ; Loriaet al., 2003).
La majorité des Streptomycètes sont non exigeantes, ils ne demandent pas de sources
organiques de nitrogène, de vitamines ou bien des facteurs de croissance, le sol comme
habitat donne aux Streptomycètes un support pour la croissance de leur mycélium (Kieser et
al., 2000).
De plus, la formation de spores aide les Streptomycètes à s’adapter aux variations des
conditions physiques dans le sol (variation d’aération, tension hydrique et le pH), périodes de
sécheresse, froid et conditions d’anaérobie qui peuvent se changer dramatiquement et
rapidement (Dworkinet al., 2006c).
Les streptomycètes dans l’environnement marin
La réalité que les océans sont une maison d’une grande population diverse, est
indisputable ; en plus si au maximum 1/1012 de la surface terrestre est analysé pour la
présence des actinomycètes, donc que sera le cas pour les océans ? (Alan&Stach, 2007). Les
Streptomycètes marines constituent une très importante source de molécules bioactifs,
puisque les conditions de l’environnement marin sont différentesde celles de l’environnement
terrestre, elles produisent des antibiotiques de type différent (Daset al., 2010; Bhatnagar&
Kim, 2010)et pas que les antibiotiques, mais des autres molécules bioactives à différents
usages thérapeutique (Solankiet al., 2008). Les Streptomycètes de l’environnement marin
sont plus connues pour leur potentiel à produire les anti-cancers comme les : Borophycine,
Daunorubicine et Gutingimycine (Bhatnagar& Kim, 2010 ; Boopathy&Kathiresan, 2010)
À part leur rôle comme producteurs d’antibiotiques, les Streptomyces de l’environnement
marin participent vivement dans le recyclage et la biodégradation continue des différents
matériaux (Daset al., 2010).

Bibliographie
9
1.5 Les Streptomycètes comme pathogène
L’importance des Streptomycètes est très limitée dans le domaine médical, car les maladies
causées par des actinomycètes sont dues à des infections les espèces de Mycobacterium,
Actinomadura etActinomyces. Mais S. somaliensis et S. sudanensis sont considérées comme
pathogènes appartenant au genre de Streptomyces et qui causent essentiellement
l’Actinomycetoma, qui est une maladies très présente en Afrique et spécialement au
Soudan(E Hamid, 2011 ; Kieseret al., 2000).
Figure 3 : endommagement de la pomme de terre à cause de S. scabies (Loriaet al., 2003)
La plupart des Streptomycètes sont saprophytes ; et que peu de souches de ce genre sont
phyto-pathogènes, comme S. scabiei qui provoque des lésions de surface de plusieurs
végétaux comme la pomme de terre (rougeur de la pomme de terre). La virulence est
dépendante de l’inhibition de la synthèse de la cellulose par la production de thaxtomines.
Chez Streptomyces turgidiscbies, l’ADN code pour la synthèse de thaxtomines qui incluent
des gènes de nécrose par le transfert latéral des gènes de virulence, les souches non-
pathogènes deviennent pathogènes (Dawnet al., 2010 ; Loriaet al., 2008).

Bibliographie
10
1.6 Taxonomie des Streptomycètes
La famille des Streptomycetaceae été d’origine proposée par Waksman et Henrici (1943)
sur la base de la morphologie et le chimotype de la paroi ; et elle contient en ce temps juste 2
genres : le genre Streptomyces et le genre Micromonospora. Les Streptomycètes ont été
décrites comme « Streptomycetaceae » formant des chaines de spores dans le mycélium
aérien, (Dworkinet al., 2006c).
Les Streptomycètes sont définies par des caractères chimotaxonomiques et phénotypique.
Maintenant tous les travaux se concentrent plus sur l’ARNr 16S et l’analyse de la paroi en
acides gras.
Il y’on a 4 types de classifications pour les Streptomycètes : la classification phénotypique,
numérique, la chimiotaxonomie et la taxonomie génétique :
I. Taxonomie phénétique : Depuis la classification proposée par Cohn en 1872 et
jusqu'au début des années 1960, la définition d'une espèce (et d'une manière générale
toute la taxonomie bactérienne) reposait sur une classification phénétique ou
phénotypique. Cette dernière utilise un faible nombre de caractères considérés comme
importants tels que la morphologie, les spores, la mise en évidence d'un caractère
biochimique jugé essentiel, l'habitat, le pouvoir pathogène... etc. Une classification
phénétique a l'inconvénient de ne refléter qu'une quantité d'information réduite
(Smaoui, 2010).
En 1964, le projetinternational des Streptomyces (ISP) a essayé de standardiser les
critères de déterminationde l’espèce. Shirling et Gottlieb (1966, 1969, 1972) ont
décrit des critères standardsclés pour la détermination des espèces :
la forme des chaines de spores (réctiflexible pour les chaines droites ou
flexueuses,spirale pour les chaines hélicoïdales) ;
la couleur du mycélium aérien sporulé avec sept classes de couleur (blanc,
gris, jaune,rouge, bleu, vert et violet) ;
la production de pigments mélanoïdes bruns à noirs ;
l’ornementation de la paroi sporale (lisse, verruqueuses, échinelée ou
chevelue) ;
la présence ou non de pigments solubles ;

Bibliographie
11
la capacité d’utiliser certaines sources de carbone pour se développer (D-
glucose, D-xylose,D-galactose, D-raffinose, D-mannitol, D-fructose, L-
arabinose, L-rhamnose, inositole et saccharose)(Boughachiche, 2012).
La taxonomie des Streptomycètesbasée juste sur le caractère de la morphologie de la
colonie est injuste, car la morphologie des colonies de ce genre est très variable et ne
constituent pas un vrai critère pour la classification ; tandis que la couleur des spores est très
importante (Kämpfer, 2006).
II. Taxonomie numérique : développée dans les années cinquante du dernier siècle en
parallèle du développement des ordinateurs (Garrityet al. 2005). En 1957, Sneath
développe une taxonomie qualifiée de numérique pour les bactéries (Smaoui,
2010)elle repose sur l’étude de plusieurs caractères macroscopiques, microscopiques,
physiologiques et biochimiques ; et à l’aide d’un logiciel on peut déterminer le genre
et l’espèce, généralement le nombre des caractères étudiés est de 134
caractères(Yamaçet al., 2011). Et cette classification a été proposée par Williams et
al (1983) et elle a donné 19 groupes majeurs, 14 groupes mineurs et 18 groupes
constitue chacun d’eux d’une seule, ça a permis de mettre un dendrogramme qui
représente tous les groupes (Annexe) (Anderson & Wellington, 2001).
III. Chemotaxonmie : elle été largement utilisée pour distinguer les Streptomycètes des
Actinomycètes. Le développement des analyses rapides et sensibles comme la
chromatographie, l’électrophorèse et la spectroscopie qui ont mené à une approche
chimotaxonomique alternative pour la classification des Streptomycètes. Parmi ces
analyses : analyse des acides gras, analyse de la paroi cellulaire, sensibilité aux
phages, tests biochimiques, sérologie, profiles protéiques (Anderson & Wellington,
2001).
IV. Taxonomie Génétique : c’est l’application des techniques de biologie moléculaire
sur le génome bactérien, le rôle de ces méthodes dans la classification étant de nous
donner la relation entre les procaryotes soit au niveau des espèces, des genres oudes
familles. Les différentes techniques de biologie moléculaire utilisées pour la
classification des Streptomycètes sont regroupées dans le tableau 2(Anderson &
Wellington, 2001).

Spécificité des méthodes moléculaire d’identification des Streptomycètes
Wellington., 2001) :
Tableau 02 : Les différentes méthodes moléculaires d’identification des StreptomycètesCible Méthode
ADN chromosomique totale Hybridation ADN
Restriction avec les endocucléases
(RFLP, LFRFA)
Isolement et clonage de
Gène de protéine ou fragment de
gene 16S/23S ARNr
Isolement et séquençage du gène ou
de fragments
Analyse comparatif de séquences
La classification du genre Streptomyces
bactériologique est la suivante :
Figure 4 : classification du genre Streptomyces
Actinobacteria
actinobacteria
Actinobacteridae
Actinomycetales
Phylum
Bibliographie
Spécificité des méthodes moléculaire d’identification des Streptomycètes (Anderson &
: Les différentes méthodes moléculaires d’identification des StreptomycètesMéthode Spécificité
Hybridation ADN-ADN Genre aux espèces
Restriction avec les endocucléases
(RFLP, LFRFA)
Espèces à la souche
Isolement et clonage de Espèces à la souche
Isolement et séquençage du gène ou
de fragments de gène
Espèces à la souche
Analyse comparatif de séquences Famille, genre, espèces
Streptomyces selon Bergy’sMannual de classification
Streptomyces selon Bergy'sMannual of SystematicBacteriology(2006)
actinobacteria
Actinobacteridae
Actinomycetales
Streptomycineae
Streptomycetaceae
Streptomyces
Famille
Subordre
Ordre
Subclasse
Classe
Phylum
Bibliographie
12
(Anderson &
: Les différentes méthodes moléculaires d’identification des Streptomycètes
Genre aux espèces
Espèces à la souche
Espèces à la souche
Espèces à la souche
Famille, genre, espèces
selon Bergy’sMannual de classification
selon Bergy'sMannual of SystematicBacteriology
Genre
Famille

Bibliographie
13
1.7 Cycle de développement :
Les Streptomycètes peuvent utiliser un grand nombre de composés organiques comme
source de carbone et d’énergie et la température optimale de croissance se situe entre 25 et
35°C. Elles sont en majorité des souches mésophiles, mais il existe quelques souches
psychrophiles ou thermophiles et la gamme de pH optimale est comprise entre 6,5 et 8,0. La
croissance peut se conduire en milieux liquide ou solide ; cependant, l’étude d’un cycle de
différenciation se réalise préférentiellement en milieu solide(Boughachiche, 2012).
La vie d’un Streptomycète débute par la germination d’une spore, bien sûr quand elle est
dans de bonnes conditions physiques et nutritionnelles. Cette germination produise une
structure morphologique semblable au tube germinatif de moisissure, celle-ci donne naissance
à un envahissement radial c’est-à-dire le mycélium du substrat (ou végétatif)(Manteca&
Sanchez, 2009) ; ce dernier se développe comme on a dit de la germination des spores qui
comprend quatre étapes : l’activation, l’initiation, l’émergence du tube germinatif et sa
croissance, pour lesquelles le degré hygrométrique joue un rôle important. Parfois l’activation
peut être déclenchée par un choc thermique, par exemple un traitement de 5 minutes à 50°C
pour les spores de Streptomyces viridochromogenes. Puis le tube de germination croît et
donne des hyphes qui se ramifient de manière apicale, l’ensemble de la colonie se développe
d’une manière radiale. Le mycélium primaire est ancré dans le support solide dans lequel il
puise les nutriments, cette habitude de formation de mycélium du substrat montre la capacité
des actinomycètes à dégrader la matière organique insoluble grâce à leurs enzymes
extracellulaires (Emerson et al., 2012 ; Kitouni, 2007).
Figure 5: Schéma du cycle de vie deStreptomyces dans les conditions naturelles (Manteca&
Sanchez, 2009)
Sur le mycélium primaire se développe un mycélium aérien ou secondaire, composé
d’hyphes, dressé sur le mycélium du substrat. Ils sont souvent pigmentés et sont enfermés
dans une enveloppe externe hydrophobe. Une fois développé, le mycélium aérien couvre les
MORT PROGRAMMEE DE LA CELLULE
MI
REPRODUCTIVE VEGETATIVE
Mycélium primaire compartimentais Spores Mycélium secondaire

Bibliographie
14
colonies de surfaces donnant un aspect poudreux compact, poilu ou en chou-fleur (Kitouni,
2007)(la nature hydrophobique est probablement présente pour permettre à la cellule de
survivre en absence de l’eau) (Dworkinet al., 2006b). Pour le cas de S. coelicolor qui est la
souche la plus étudiée, la formation du mycélium aérien nécessite l’activation d’un nommé
bld car les souches mutantes au niveau de ce gène ne peuvent pas développer une surface
poudreuse ; en plus ces mutantes ne produisent pas quelques protéines extracellulaires qui
couvrent la surface du mycélium secondaire en lui donnant son caractère hydrophobe (Brun,
2004 ; Kelemen&Buttnert, 1998).
En conséquence de la carence en nutriments, ce mycélium aérien commence à former des
spores, ce mycélium aérien et ces spores donnant à la colonie de Streptomyces une apparence
poudreuse, et chevelue(Dworkinet al., 2006b).
Figure 6 : Cycle de vie des Streptomyces (Scherr& Nguyen, 2009 ; Delaunay et al, 2003)
En milieu liquide :elle nécessite une aération du milieu par agitation, et ou par
injection d’air puisque ces bactéries sont aérobies strictes.En milieu liquide, les
cellules se développent uniquement sous forme de mycéliumprimaire, même si
certaines souches peuvent sporuler dans cet environnement (Madiganet al., 1997).Les

Bibliographie
15
Streptomycètes croissent par élongation des filaments et peuvent par la suite présenter
trois types de morphologies (Saffroy, 2006)(Fig. 07).
Figure 7: Morphologies rencontrées aux cours de cultures liquides de Streptomyces (Olmoset al., 2013)
En milieu solide : lorsqu’elles sont placées dans des conditions favorables, les spores
vont produire un tube germinatif qui va croitre pour donner des hyphes qui vont à leur
tour se ramifier. Il s’agit d’un mycélium primaire (végétatif) ou substrat qui se
développe en surface et à l’intérieur du support. Un mycélium secondaire (aérien) va
ensuite se développer sur la colonie (Saffroy, 2006) (Fig. 06).
Les espèces appartenant au genre Streptomyces peuvent produire une large variété de
pigments responsables de la couleur du mycélium végétatif et aérien. En plus des pigments

Bibliographie
16
diffusables peuvent être formés. Il faut bien noter que la production de ces pigments est
dépendante de la composition du milieu de culture et ainsi des conditions de culture
(Dworkinet al., 2006c).
Starch-casein-nitrate Ager GYM Agar Oatmeal Agar
Figure 8: Couleur du mycélium aérien et des pigments diffusible S. glaucescens(Dworkin et al., 2006c)
Figure 9: Morphologie du mycélium aérien de Streptomyces(Kämpfer, 2006)
1.8 Sporulation et germination :
Comme la sporulation est un moyen d’adaptation aux différentes conditions dans
lesquelles se trouvent les Streptomycètes, les spores donc restent en un état semi-dormant, ce
qui facilite leur survie dans le sol pour de longues périodes. Relativement les Streptomycètes
sont présentes dans le sol sous forme de spores et la germination des spores peut être causée
par la compétition avec des autres microorganismes indigènes ; mais ces spores ne cessent pas
de sporuler de nouveau(Dworkinet al., 2006c). Il faut bien noter que les spores de
Streptomyces sont moins résistantes que les endospores de bacillus et autres genres de
Firmicutes(McCormick &Flärdh, 2012).

Bibliographie
17
La germination peut dépendre de quelques signaux, il est évident que les nutriments, l’eau
et Ca+2, sont indispensables. De plus l’état des nutriments dans le site de germination
influence l’extension du substrat et en même temps la différentiation en mycélium
aérien(Hopwood et al., 1985)
Il a été démontré que les spores des Streptomycètes contiennent tous les acides
aminésessentiels et une contenance élevée en tréhalose et en calcium (Janeiček et al., 1997).
La plupart des Streptomycètes ont des spores pigmentés, et leur couleur été utilisée comme
critère de classification. Chimiquement il y’on a 2 types de pigmentation de spores ; La
première est très abondante, elle est responsable d’une large gamme de couleur ; tandis que la
deuxième est responsable de la production de la mélanine, sachant que les pigments des
spores sont un outil de protection contre les rayons UV(Chater et al., 2010).
Figure 10 : hyphes végétatives et conidiospores de S. coelicolor, SI : spores immatures,
BP : Bordure de la paroi, SM : spores matures (Chater &Hopwood, 1973)
Figure 11 : développement du mycélium secondaire en spores(Saffroy, 2006) (1) : germination de la spore et développement du mycélium primaire (2) : développement du mycélium secondaire (3-4) : enroulement du mycélium aérien (5-6) : cloisonnement et formation des spores (7) libération des spores

Bibliographie
18
Pridham et al(1957) ont donné une suggestion sur le développent de la forme des spores et
divisent les Streptomycètes suivant la forme de spores en 7 groupes : [ Rectus-Flexibilis(RF) ;
Retinaculum-Apertum(RA) ; Spira(S) ; Monoverticillus(MV) ; Monoverticillus-Spira(MV-
S) ; Biverticillus(BIV) ; Biverticillus-Spira(BIV-S) ].
Et il a divisé ces derniers groupes à leur tour en plusieurs séries suivant la couleur du
mycélium aérien à la maturité qui sont :
- Blanc
- Olive
- Jaune
- Bleu (bleu à bleu-vert au vert)
- Rouge (rose au rouge au violet au violet-gris)
- Gris (gris clair au marron-gris au marron-gris)
Dans un autre travail de Pridham(1965), dans lequel il a essayé de collecter toutes les
publications concernant les Streptomycètes ; il a démontré que les Streptomycètes ont
vraiment une large gamme de couleurs qui atteint 251 couleurs.
1.9 Les milieux de culture des Streptomycètes :
Comme tout microorganisme, les Streptomycètes ont besoin des nutriments pour assurer
leur différentes fonctions vitales ; donc ils ont besoin au minimum d’une source de carbone et
autre de nitrogène.
1.9.1 Source de Carbone :
Il n’y a aucune évidence pour le moment que les Streptomycètes utilisent la voie d’Entner-
Doudoroff(Kieseret al., 2000). Il semble que les sucres sont importés dans la cellule puis
phosphorylés par des kinases spécifiques, comme le cas du fructose où il a un système de
phosphorylation et de transport, ça été prouvé chez S. coelicolor, S. lividans, S.
Griseofuscus(Titgemeieret al., 1995). Il semble que ces voies périphériques du catabolisme
du carbone sont tous induites et liées à la répression du catabolite carboné. La répression du
catabolite carboné (CCR), liée à la l’aptitude et l’efficience du catabolisme de la source du
carbone, comme le glucose, pour éviter l’utilisation d’autres sources alternatives de carbone.
La CCR est connue chez les bactéries, et chez les Streptomycètes elle peut inhiber la
production des antibiotiques (Kieseret al., 2000).

Bibliographie
19
1.9.2 Source de nitrogène :
Chez toutes les Streptomycètes étudiées, il y’on a une seule voie pour l’assimilation
du nitrogène, c’est via le glutamate synthétase (GS) et glutamate : 2-oxoglutarate
transaminase (GOGAT). L’assimilation du nitrogène via GS dépense beaucoup
d’énergie, mais elle est possible à une concentration minimale de l’ammoniac
(Aharonowitz& Friedrich,1980).

Bibliographie
20
Tableau 03 : représentation des différents milieux de culture utilisés pour la culture des Streptomycètes :
Numéro du milieu
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Amidon 10,0 10,0 10,0 10,0 10,0 10,0 10,0 Glucose 1,0 1,0 1,0 Glycérol 5,0 5,0 5,0 5,0 5,0 5,0 12,5 12,5 12,5 Na-propionate 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 Soja 5,0 Caséine 0,3 0,3 0,3 0,3 0,3 0,3 1,0 Na-caséinate 2,0 2,0 2,0 2,0 2,0 2,0 Tryptone 5,0 5,0 5,0 15,0 Asparagine 0,1 0,1 0,1 0,1 0,1 0,1 Arginine 1,0 1,0 1,0 KNO3 2,0 2,0 2,0 NH4NO3 4,0 4,0 4,0 4,0 4,0 4,0 Extrait de levure 2,5 2,5 2,5 2,5 2,5 2,5 2,5 2,5
NaCl 2,0 2,0 2,0 2,0 2,0 2,0 1,0 1,0 1,0 5,0 K2HPO4 1,0 2,0 1,0 2,0 0,5 1,0 0,5 1,0 0,5 1,0 0,5 1,0 0,5 1,0 0,5 1,0 0,5 CaCO3 0,02 0,02 MgSO4-7H2O 0,50 0,05 0,50 0,05 0,50 0,10 0,50 0,10 0,50 0,10 0,50 0,10 0,5 0,1 0,5 0,1 Fe3SO4-7H2O 0,01 0,01 0,01 0,01 0,01 0,01 0,01 0,01 Fe2(SO4)3-6H2O 0,01 0,01 0,01 0,01 0,01 0,01 0,01 0,01
CuSO4-5H2O 0,001 0,001 0,001 0,001 0,001 0,001 0,001 0,001 ZnSO4-7H2O 0,001 0,001 0,001 0,001 0,001 0,001 0,001 0,001 MnSO4-H2O 0,001 0,001 0,001 0,001 0,001 0,001 0,001 0,001
The media compositions (milieu 6 :starch– casein–KNO3 agar; milieu 16 :actinomycete-isolation agar; milieu 21 : glycerol –arginine agar; milieu 23 :tryptone– yeastextract– glucose agar)(Suutariet al., 2002)

Bibliographie
21
2 Les substances bioactives 2.1 Définition
Une substance bioactive veut dire une substance quelque soit sa nature, mais l’essentiel
qu’elle a un rôle important dans un organisme vivant dans lequel elle est introduite, c’est-à-
dire un rôle biologique(Zähner, 1979).
2.2 Les métabolites secondaires
De plus d’un siècle, le botaniste allemand Kossel (1891) a défini les métabolites
secondaires par exclusion (qui n’appartiennent pas aux métabolites primaires) (Karlovsky,
2008, Holgeret al, 2008). Le terme « métabolisme secondaire » a été utilisé par les
physiologistes des plantes au début du 20ème siècle, il a compris tout métabolisme spécifique
pour certains organismes (Zähner, 1979).
Comme définition un métabolite secondaire est une substance produite naturellement mais
qui ne joue pas un rôle explicite dans l’organisme producteur et ils appartiennent à des
différentes classes chimiques ;jusqu’à ce jour leur rôle exacte reste mal étudié, mais
probablement ils fournissent des avantages écologiques pour l’organisme producteur (Liras
& Martín 2004)et ça augmente la possibilité de survie de l’organisme qui les produisent car
ils actent comme un mécanisme alternatif de défense (Holger et al, 2008 ; Maplestoneet al.,
1992). Les métabolites secondaires incluent : antibiotiques, pigments, toxines, phéromones,
enzymes inhibiteurs, agents immuno-modulateurs, pesticides, agents anti-tumorales …
etc.(Lancini&Demain, 2013).
La nature produise une variété et un nombre impressionnant de ces métabolites ; vers la fin
de l’an 1999, plus de 23400 métabolites secondaires connus, parmi ces molécules 42,1 %
produites par des moisissures, 42,2 % par les actinomycètes et 15,5 % par les autres bactéries
(Lazzarini et al. 2001). La découverte des molécules bioactives dérivant des
microorganismes est continue à une ampleur étonnante : 200-300 par an vers la fin des années
70 du siècle précédentet vers 500 par an pendant les années 90 du même siècle
(Lancini&Demain, 2013).
La production microbienne de métabolites secondaires est généralement influencée et
connectée au métabolisme primaire de la souche productrice. Les métabolites intermédiaires à
l'issue du métabolisme primaire servent de précurseurs pour la biosynthèse de ces métabolites

Bibliographie
22
secondaires bioactifs. En effet, la composition du milieu de culture influence les capacités
métaboliques de l'organisme producteur ainsi que la biosynthèse des métabolites
secondaires(Thakuret al., 2009).
2.3 Métabolisme des Streptomyces
La voie d’Embden-meyerhof-Pernas (Glycolyse), voie du catabolisme du glucose est
présente chez plusieurs Streptomycètes, mais aussi la chaine d’hexose monophosphate a été
détectée chez S. antibioticus. Quelques Streptomycètes peuvent changer de la glycolyse à la
chaine hexose monophosphate durant le métabolisme secondaire. Jusqu’à présent aucune
souche de ce genre est connue d’utiliser la voie d’Etner-Doudoroff(Kieseret al., 2000).
2.4 Métabolisme secondaire des Streptomyces
Les Streptomyces sont les meilleurs candidats pour la production des
métabolitessecondaires biologiquement actifs ; En effet, ce genre bactérien est à l’origine
d’environ 70% desmolécules antibiotiques utilisés en médecine et 60% des antifongiques
utilisés en agriculture(Sujathaet al., 2005).Il a été prouvé que 50 % des Streptomyces sont
actives contre les bactéries Gram-positif(Dworkinet al., 2006a). L’étude de la souche type S.
coelicolor a révélé que le génome de cette souche code pour 23 métabolites secondaires
d’intérêt et 30 gènes pour StreptomycesavermitilisMA-4680T (Ventura et al., 2005in
Kämpfer, 2006), c’est très intéressant qu’un génome porte ce grand nombre de gènes.
Plusieurs travaux ont été menés pour augmenter la production demétabolites d’intérêts à
partir des souches appartenant à ce genre bactérien et il a été démontré qu’il y’a une influence
majeure des conditions de culture de la bactérie productrice sur la productiondes métabolites
recherchés(Smaoui, 2010).
Plusdes molécules antibactériennes, les Streptomycètes produisent des
immunosuppresseurs, anti-cancers(Olanoet al., 2009), antidiabétiques(Kulkarni-Almeida,
2011), antivirales (Ara et al., 2012), antifongiques(Oskay, 2009),
herbicides(Dhanasekaranet al., 2010), insecticides (Deshpandeet al., 1988), antiparasitaires
(Pimentel-Elardoet al., 2010) …etc(Weber et al., 2003).

Bibliographie
23
En générale, la production des métabolites secondaires est associée au carence en
nutriments essentiels, et ce manque de nutriments spécifiques qui déclenche un cascade
d’évènements de régulation résultant d’une différentiation chimique (métabolisme secondaire)
et une différentiation morphologique (Jonsbuet al., 2002).
Tableau 04 : principales différences entre les métabolites primaires et secondaires (Delaunay et al.,2003)
Métabolisme primaire Métabolisme secondaire
Synthèse pendant la trophophase Synthétisé pendant l’idiophase
Présent tout au long du cycle cellulaire Apparition à un moment du cycle cellulaire
Nécessaire à la croissance Rôle physiologique mal connu
Turn over élevé Turn-over pratiquement nul
Produits dans des conditions de cultures diverses Produits dans des conditions de cultures bien
définies
Ubiquitaire Spécifique
Enzymes à spécificité étroite Enzymes à spécificité large
Voies de synthèse simple et courte Synthèse longue et complexe
Synthèse d’un produit parfaitement défini Synthèse d’un mélange de produits
Structure chimique généralement simple Structure chimique souvent complexe
Concentration élevé Concentration faible
Le séquençage du génome de S. coelicolor et de S. avermitilis a révélé que les deux
espèces contiennent une vingtaine de gènes codant pour des métabolites secondaires, malgré
qu’avant leurs séquençage, il apparait qu’ils produisaient que 3 à 4 métabolites secondaires et
ça reflète l’importance des méthodes moléculaires d’identification et de la
recherche(Alan&Stach, 2007).
2.5 Production d’antibiotiques par les Streptomycètes
La production d’antibiotique est généralement spécifique pour souches (espèces), Le
génome de S. coelicolor par exemple code un large nombre de protéines secrétés (891),
incluent 60 protéases, 13 chitinases, 8 cellulases/endoglucanases, trois amylases et deux,
pactatolysases(Emerson et al., 2012). La production des antibiotiques coïncide avec le début
de la phase stationnaire en milieu liquide et avec le début de la différenciation morphologique
en milieu solide (Bibb&Hesketh, 2009)la synthèse de ces molécules est contrôlée par des
gènes de régulation qui se trouvent au niveau du chromosome(Komatsu et al., 2009).

Bibliographie
24
Vue le nombre des molécules bioactives produites par les Streptomycètes, un model estime
le nombre totale des molécules antimicrobiennes que ce genre est capables de les produire
peut être de l’ordre de 100.000 et qu’une fraction reste toujours non découverte
(Singh&Pelaez, 2008 ;Watveet al., 2001).
La production à l’échelle industrielle a besoin d’un rendement en (g/l), tandis que la
production des souches typiques est de l’ordre de (mg/l) ou plus moins. Pour dépasser ce
problème au niveau industriel il est nécessaire de faire des mutations et des sélections ou bien
utiliser les méthodes de synthèses (Wohllebenet al., 2012).
2.5.1 Effet de la composition du milieu de culture
La nature et la concentration de certains composés dans le milieu de culture ont un
effetéminent sur la production des métabolites secondaires biologiquement actifs(Jonsbuet
al., 2002 ; Thakuret al., 2009). En faite, il a été montré que la nature des sources : de
carbone, d’azote, de phosphore,de potassium, de magnésium et de trace éléments minéraux
(essentiellement Mn2+, Cu2+ et Fe2+),affectent fortement la production de la molécule
antifongique de type macrolide non polyènique. Également, l’origine d’une même source de
carbone peut affecterla sécrétion des biomolécules par le microorganisme
producteur(Elibol&Mavituna 1998) ; c’est le cas de l’antibiotiqueaminoglycoside, la
néomycine. Il a été démontré par Adinarayana et al(2003), que la productionde cette
molécule par la souche Streptomyces marinensis est en fonction de l’origine céréalière
del’amidon utilisé comme source de carbone.
Pour la source d’azote, la production de l’antibiotique SBR-22 par Streptomyces BT-408 a
étéétudiée sur plusieurs sources d’azotes inorganiques ou sous forme d’acides aminés. Il a
étérapporté que c’est la source d’azote inorganique (nitrate d’ammonium) qui donne la
meilleureproduction de la molécule SBR-22 (Sujathaet al., 2005). Ces mêmes auteurs ont
montréégalement l’importance de la nature de la source de phosphate dont certaines peuvent
conduire àl’arrêt de la production de la biomolécule SBR-22.

Bibliographie
25
L'importance du magnésium et du potassium dans la production des antibiotiques par
différentesespèces de Streptomyces a été rapportée par plusieurs chercheurs,Mellouliet
al(2004) ontconstaté que le meilleur rendement de production de biomolécules de la nouvelle
bactérie deStreptomyces TN58 est obtenu en présence de potassium à 2 mM.
Il est à noter que plusieurs autres additifs comme le sulfate, le chlorure, l'acétate
etl'arginine peuvent influencer la production de plusieurs métabolites secondaires, c’est le cas
desdeux biomolécules, l’antibiotique IM-111-81 et de l’azalomycine B, dont la production par
lasouche Streptomyces hygroscopicus est stimulée par l’ajout de ces additifs (Geshevaet al.,
2005).
2.5.2 Effet du pH, de la température, de l’agitation et dutemps d’incubation
Les conditions de culture comme le pH, la température, l’agitation et le tempsd’incubation,
affectent énormément la production des métabolites secondaires(Olmos et al., 2013 ; Liang
et al., 2008 ; Elibol&Mavituna, 1997). Depuis longtemps,il a été démontré que la variation
du pH a une influence sur la production de plusieurs métabolites organiques dumétabolisme
secondaire. En effet plusieurs travaux comme celui de Lianget al(2008) qui a prouvé la
variation de la production de la natamycine par la souche Streptomyces gilvosporeus LK-196
en fonction du pH, de la température, de la vitesse d’agitation et même du taux d’aération ;
cet influence a été rapporté par Thakuret al(2009).
L'agitation affecte l'aération et le mélange des éléments nutritifs dans le milieu
defermentation et par conséquent elle affecte la production de métabolites secondaires. Il a été
rapporté parMellouliet al(2004), que pour la souche de Streptomyces TN58, qui produit cinq
moléculesbioactives différentes, la meilleure production est obtenue à une agitation comprise
entre 200 et250 rpm. Pour des agitations faibles de l’ordre de 100 rpm ou fortes au voisinage
de 300 rpm, laproduction de biomolécules chute énormément.
Selon le travail de (Matelováet al, 1955 in Janeiček et al, 1997), l’aération est très
importante surtout dans les premières heures de culture, cette phase qui est considérée comme
une phase préparatoire pour le cas de S. aureofacienspour la production de la
chlortetracycline.

Bibliographie
26
Chez les Streptomyces, l’évolution de la production des métabolites secondaires enfonction
du temps, qui est un facteur déterminant, est variable d’une espèce à une autre. En effet,pour
la souche de Streptomyces TN58, la production des biomolécules commence après
60h(Mellouliet al., 2004).
2.5.3 Méthodes d’extraction des métabolites secondaires
Il faut prendre en considération que les métabolites secondaires sont produits en faible
quantité (rendement très faible), et une souche peut produire une mixture de molécules. Les
métabolites peuvent être excrétés totalement ou partiellement, tout dépend de leur présence
dans le milieu extracellulaire ou intracellulaire (Sterner, 2012).
Une variété d’approches d’extraction estappliquée, elles varient selon les souches et les
métabolites qu’on veut extraire. Généralement l’extraction des métabolites secondaires des
microorganismes, est faite après une fermentation et le milieu est clarifié soit par filtration ou
par centrifugation avant l’extraction ; cette clarification donne un avantage surtout quand les
métabolites sont extracellulaires(Seidel, 2006).
Tableau 5 : Propriétés physicochimiques des solvants utilisés dans l’extraction des métabolites secondaires(Seidel, 2006)
Solvant Index de
Polarité
Point d’ébullition
(C°)
Viscosité
(cPoise)
Solubilité dans
l’eau (% w/w)
n-Hexane 0.0 69 0.33 0.001
Dichlorométhane 3.1 41 0.44 1.6
n-Butanol 3.9 118 2.98 7.81
iso-propanol 3.9 82 2.30 100
n-Propanol 4.0 92 2.27 100
Chloroforme 4.1 61 0.57 0.815
Ethyleacétate 4.4 77 0.45 8.7
Acétone 5.1 56 0.32 100
Méthanol 5.1 65 0.60 100
Éthanol 5.2 78 1.20 100

Bibliographie
27
2.5.4 Purification et Caractérisation métabolites secondaires :
Les méthodes de purification des métabolites secondaires sont très variées et dépendent de
la nature des métabolites isolées. La chromatographie en couche mince (CCM) s’est imposée
comme une des techniques de base dans la recherche sur les substances naturelles, en raison
de sa simplicité de mise en œuvre et de sa capacité de séparation et d’analyse des mélanges
complexes que constituent les extraits bruts d’origine naturelle(Poole, 2003).
Elle permet de comparer rapidement et simultanément plusieurs dizaines d’échantillons,
dans des conditions peu onéreuses, et ce avec une grande flexibilité par rapport aux autres
techniques couramment pratiquées dans le domaine des produits naturels (chromatographie
liquide haute pression, chromatographie en phase gazeuse et chromatographie de partage
centrifuge...)(Pradeau& Dauphin, 2002).
Pratiquement toutes les publications relatives à l’isolement des produits d’origine
microbienne font état du rapport frontal (Rf) comme caractéristique de la molécule purifiée.
La révélation est très simple, elle est tout d’abord faite par visualisation sous une lampe UV
(254 et 366 nm) (Arasu et al., 2009).Il faut bien noter que la visualisation par UV est non-
destructive, si les molécules n’apparaissent pas sous UV il est indispensable de passer par la
pulvérisation des réactifs (révélateurs)(Gibbons, 2006) là où chaque type de produits a son
révélateur (ex : vanilline, acide sulfurique, molybdate d’ammonium … etc).
Il y’en a une autre méthode de révélation qui est « la bioautographie », elle est surtout
utilisée quand la substance à révéler est une molécule antibactérienne ou antifongique, elle
consiste à placer la plaque ou le papier au-dessus d’une culture bactérienne ou fongique déjà
ensemencée et laisser là s’incuber pendant 24h. La révélation sera le jour suivant où il
apparaitrait une zone claire autour de l’emplacement de la substance ; bien sûre si la molécule
est biologiquement active, sinon il y’aura pas de zone(Choma &Grzelak, 2011).
2.5.5 Production des antibiotiques À l’échelle industrielle la production des antibiotiques se fait dans des fermenteurs, Le
schéma général consiste à séparer par centrifugation la biomasse du jus defermentation
contenant les molécules d'intérêts. Par ailleurs, il est à noter que dans des cas assezrares,
certaines molécules ne sont pas secrétées en totalité dans le surnageant et une fraction
resteintracellulaire. Dans cette situation il faut traiter la biomasse par des moyens
physiques(Smaoui, 2010).

Bibliographie
28
Figure 12 : Schéma général conduisant à l’obtention des antibiotiques à l’état pur (Smaoui, 2010)
Milieu de culture+ microorganisme Producteur de l’antibiotique
Fermentation
Centrifugation
Culot cellulaire Surnageant contenant l’antibiotique
Etapes d’extraction-séparations
Cristallisation
Dissolution et décoloration
Nouvelle cristallisation
Séchage
L’antibiotique à l’état pur

Bibliographie
29
Après l’obtention de la molécule active à l’état pur et pour élucider sa structure chimique,
il est nécessaire d'appliquer plusieurs techniques spectroscopiques. Le nombre de techniques
àutiliser dépend de la nature de la molécule à savoir sa masse, sa géométrie dans l'espace,
sapolarité …etc. L’élucidation structurale des molécules organiques ayant des activités
biologiquestelles que les antibiotiques est faite à l'aide de mesures de susceptibilité
magnétique, destechniques spectroscopiques (UV-visible, Infrarouge ou Résonance
ParamagnétiqueÉlectronique … etc.)(Smaoui, 2010).
Tableau 06 : Tableau qui représente les molécules bioactives produites par les Streptomycètes (Dyson, 2011 ; Solanki et al, 2008) :
L’antibiotique Producteur Application
Actinolycine D S. spp. Anti tumorale
Antimycine A S .spp. Telocidal
Avermectine S. avermitilis Antiparasitaire
Bambermycine S. bambergiensis Growthpromotant
Bilaphos S. hygroscopius Herbicide
Bleomycine S. verticillus Anti-tumorale
Candicidine S. griseus Antifongique
Chloramphenicol S. venezuelae Antibactérien
Chlortetracycline S. aureofaciens Antibactérien
Acide Clavulanique S. clavuligerus Combiné avec un β-lactam
comme Antibactérien
Cycloserine S. orchidacus Antibactérien
Daptomycine S. roseosporus Antibactérien
Daunorubicine S. peucetius Anti-tumorale
Doxorubicine (adriamycine) S. peucetius var. caesius Anti-tumorale
Fosfomycine S. spp. Antibactérien
Hygromycine B S. hygroscopicus Antihelminthique
Kanamycine S. kanamyceticus Antibactérien
Lasalocid S. lasaliensis Anticoccidien
Lincomycine S. lincolnensis Antibactérien
Milbemycine S. hygroscopicus Antiparasitaire
Mithramycine S. argillaceus Anti-tumorale
Mitomycine C S. caespitosus
S. verticillatus
Anti-tumorale
Monensine S. cinnamonensis Anticoccidien

Bibliographie
30
Natamycine S. nataensis Antifongique
Neomycine S. fradiae Antibactérien
Nikkomycine S. tendae Antifongique ; insecticide
Novobiocine S. niveus Antibactérien
Nystatine S. noursei Antifongique
Oleandomycine S. antibioticus Antibactérien
Oxytetracycline S. rimosus Antibactérien
Phleomycine S. verticillus Anti-tumorale
Polyoxins S. cacaoi var. asoensis Antifongique
Pristinamycine S. pristinaespiralis Antibactérien
Salinomycine S. albus Anticoccidien
Spectinomycine S. spectabilis Antibactérien
Spiramycine S. ambofaciens Antibactérien
Streptogamines S. graminofaciens Antibactérien
Streptomycine S. griseus Antibactérien
Tetracycline S. aureofaciens Antibactérien
Thienamycine S. cattleya Antibactérien
Tobramycine S. tenebrarius Antibactérien
3 Génétique des Streptomycètes 3.1 Le chromosome
Les chromosomes linéaires sont relativement mois présents chez les bactéries et leurs
dominance est chez les Actinimycétales, et chez le genre Streptomyces en particulier (ex : S.
antibioticus, S. moderatus, S. lipmanii, S. parvulus)(Dworkinet al., 2006c),ça pose beaucoup
de questions intéressantes. Ces chromosomes sont limités par des protéines terminales qui
sont liées aux terminaisons 5’ de l’ADN et ces structures sont impliquées dans la réplication
et la protection des terminales de ces réplicons (Bendich&Drlica, 2004).Il ne faut pas oublier
que l’espèce type de la famille Streptomycetaceae : S. coelicolor a un chromosome circulaire
(Dworkinet al., 2006c).
Le chromosome des Streptomycètes est considéré comme le plus large chromosome dans
le monde bactérien. Il est de 8 Mb à plus de 10 Mb avec un pourcentage G+C de 70%
jusqu’au 80% (Medemaet al., 2010). La linéarité du chromosome est reliée à un autre
caractère de Streptomyces qui est leurs tendances à avoir de larges délétions ou bien des

Bibliographie
31
amplifications d’une fréquence de 0,1-1% et même plus que ça lors des traitements qui
endommagent l’ADN ou ceux qui interfèrent avec son réplication, comme l’acridine, promide
d’ethidium ou le choc par le froid et ces délétions peuvent atteindre 2Mb chez Streptomyces
ambofaciens(Desjardin, 2002 ; Hopwood et al., 1985). Cette longueur résulte d’une
augmentation du nombre des gènes ainsi que la longueur énorme des gènes présents. Le grand
nombre des gènes à son tour reflète le mode de vie et la complexité structurale, la variabilité
de l’environnement et la richesse en métabolites secondaires (Kiesseret al., 2000). La région
centrale du chromosome est hautement conservée et intacte, incluant les gènes essentiels pour
la réplication, la transcription, la traduction et la division cellulaire (Zhou et al., 2011).
Depuis pas mal de temps le séquençage du génome complet de 3 espèces de Streptomyces
a été publié, S. coleicor A3 (2) (2002), S. avermitilis(2003), S. griseus(2008)(Gustet al.,
2004 ; Zhou et al., 2011)
Quand le séquençage du génome complet de S. coelicolor a été publié en 2002, il été le
plus grand génome 8,677,507bp comportant 7,912 gènes et 7,769 gènes codant pour des
protéines. Il faut bien noter que jusqu’à ce jour il n’a pas été prouvé que les Streptomyces
possèdent un chromosome circulaire à part S. coelicolor, et tous les souches que leurs
génomes ont été complètement séquencés, leur chromosome est linéaire.
3.2 Instabilité génétique des Streptomycètes
Les bactéries appartenant au genre Streptomyces montrent une instabilité génétique de
haute dégrée, qui est définie comme l’apparence des mutations spontanées avec une fréquence
plus que 10-3.Des observations ont montré que cette instabilité est liée à un stade particulier
du développement des Streptomycètes(Fischeret al., 1997).
3.3 Plasmides circulaire et linéaire :
On trouve souvent des plasmides chez les espèces de Streptomyces, et le plasmide linéaire
semble être le plus commun (SLP2 of Streptomyces lividans et SCP1 de Streptomyces
coelicolor) que le plasmide circulaire (SCP2 deStreptomyces coelicolor) et il y’en a 25
plasmides chez les Streptomycètes (Smillieet al., 2010 ; Zhouet al., 2012). Le plasmide
linéaire partage la même structure générale que le chromosome linéaire (souvent) des
protéines liées aux extrémités, (généralement) un seul point d’origine de réplication. Les
protéines des extrémités des plasmides sont parfois codées par le chromosome ; les deux

Bibliographie
32
formes de plasmides sont généralement transférables, mais ils ne portent que rarement des
gènes de résistance aux molécules toxiques ou des gènes de production d’antibiotiques ou
bien des autre avantages bien reconnus. Il y’a que peu de plasmides qui sont porteurs des
gènes codant pour des métabolites secondaires comme SCP1, pSV1 et pSLA2-L(Hopwood.,
2006).
3.4 Identification moléculaire des Streptomycètes
L’identification génotypique des Streptomycètes inclue toutes les méthodes qui étudient les
molécules d’ADN et de l’ARN. L’utilisation de l’analyse de la séquence de l’ARNr 16S a
développer notre compréhension des relations entre les différentes espèces de ce genre et bien
sûre nous permettre de différencier ce genre des autres taxons. (Hakvåget al., 2008).
Le gène ADNr 16S qui code pour l’ARNr 16S (ARN ribosomique 16S) est l’un des gènes
les mieux conservés parmi lesorganismes procaryotes (Eubactéries et Archaea). Il a été choisi
comme marqueur phylogénétique en constituant une basede comparaison efficace et fiable
pour pouvoir à la fois comparer et différencier entre les bactéries. En effet, L’ADNr16S :
Comporte des séquences internes très conservées qui permettent de sélectionner des
amorces universelles pourl’amplification de l’ADNr 16S de la majorité des bactéries
existantes ;
Comporte des séquences internes variables qui, une fois analysées, permettent de
distinguer les espèces des bactéries entre elles et de les classer en fonction de leur
phylogénie ;
Est d’une taille suffisamment courte (~1500 pb) pour être analyser
rapidement(Boussaberet al., 2013)
Il y’en a 3 régions variables au niveau du gène de l’ARNr 16S, spécifique au genre (la
régiona et b) et spécifique aux espèces (région c). Près du genre ARNr 16S il y’en a l’ARNr
23S, l’ARNr 5S et l’espace l’ARNr 16S-23S qui sont des gènes utilisés pour l’investigation
sur les relations entre les Streptomycètes.
Cette séquence du gène de l’ARNr 16S est même utilisée comme amorce pour identifier le
genre Streptomyces à l’aide d’une amorce spécifique (identification du genre ou de l’espèce
sans séquençage).

Bibliographie
33
L’application des techniques comme l’RFLP (Lanootet al., 2005) et RAPD sur ce gène,
conduisent aussi à l’identification des collections de Streptomyces(Martin et al., 2000 ;
Rintala et al., 2001).
RFLP : Polymorphisme des longueurs de fragments de restriction (Random Fragments
Length Polymorphism), le polymorphisme est utilisé comme marqueur génétique
d’identification des individus, le principe de l’RFLP repose sur la comparaison des profils de
coupure par les enzymes de restriction suite à l’existence d’un polymorphisme dans la
séquence d’une molécule d’ADN(Tagu, 1999).
Il y’on a pas mal de travaux pour l’identification des gènes qui sont spécifique des souches
comme le gène de la virginiamycin chez S. virginiae(Pulsawatet al., 2007).
Muharram(2010) a utilisé la RFLP après polymérisation de la protéine du choc thermique
(HSP) pour identifier pas mal de nombre d’espèces.
RAPD : polymorphisme d’ADN par amplification aléatoire
(RandomAmplifiedPolymorphic DNA), lors de la recherche de polymorphisme d’un génome,
si aucune séquence d’ADN précise n’est ciblée, il est possible d’utiliser des oligonucléotides
de petites tailles (de 6 à 9 bases) de séquence aléatoire. Ces amorces vont se fixer sur l’ADN
au hasard et la nature des produits d’amplification obtenus est inconnue(Martin et al., 2000).
Cette technique permet d’obtenir rapidement des fragments variés d’ADN génomique, qui
seront ensuite analysés. Les conditions de préparation des échantillons et des cycles de PCR
doivent être optimisées pour chaque application afin d’assurer de la reproductibilité des
résultats(Tagu 1999).
L’hybridation ADN-ADN : est l’une des techniques utilisées pour la différentiation entre
les espèces et pour la taxonomie des Streptomyces ; cette technique, selon les études est
presque similaire à la taxonomie numérique. Il a été démontré que les Streptomycètes
présentent une instabilité génétique considérables supportent l’hybridation ADN-ADN. Mais
l’hybridation peut être influencée par la présence de larges plasmides chez les
Streptomyces(Kieser et al., 2000).

Matériels &
Méthodes

Matériels et Méthodes
34
1 Prélèvement
Les prélèvements dans cette étude ont été réalisés comme suit :
Après avoir écarté les trois premiers centimètres de particules terreux, une quantité
suffisante de terre est prélevée du sol (100g), jusqu’à 10 centimètres de profondeur. Puis
déposée à l’aide d’une spatule sur une feuille d’aluminium stérile, les échantillons sont
conservés à 4°C pendant 1-2 jours au maximum pour ne pas affecter sa contenance initiale en
microorganismes afin de les transporter au laboratoire (Chroňákova et al, 2010 ; Awad et
al., 2009 ; Ceylan et al, 2008 ; ahin & U ur, 2003).
Un premier tri des échantillons a été fait en écartant les pierres et les différents débris
végétaux, afin de les stocker dans des flacons stériles en verre.
Onze échantillons ont été prélevés de différents endroits, le tableau suivant présente les
lieux de prélèvement :
Tableau 07 : lieux de prélèvement des échantillons
Echantillon Nature du sol Lieux
Ech 01 Agricole
Sidi Maamar - Saida Ech 02 Semi-aride
Ech 03 Zone humide
Ech 04 Semi-aride Khalfellah-Saida
Ech 05 Aride Elbayadh
Ech 06 Forestier Es-senia - Oran
Ech 07 Agricole Canastel - Oran
Ech 08 Sol de jardin
Ech 09 Sol de jardin
Mostaganem Ech 10 Forestier
Ech 11 Agricole

Matériels et Méthodes
35
Les photos ci-dessous montrent les lieux de prélèvement de deux échantillons :
Figure 13 : lieu de prélèvement Ech 02 Figure 14 : lieu de prélèvement Ech 03
2 Pré-enrichissement des échantillons
Il existe plusieurs méthodes de pré-enrichissement du sol pour l’isolement des
Streptomyces et dans ce travail chaque échantillon a été devisé en 3 parties, chaque partie a
subit un type de traitement comme suit :
1. Traitement par la chaleur : chauffage de l’échantillon à 110°C pendant 10 minutes
après éparpiller l’échantillon sur le papier aluminium pour permettre un bon traitement
(Bouras et al., 2013 ; Awad et al., 2009). Le but étant de réduire la quantité des
bactéries à gram négatif (Jeffrey, 2008) ;
2. Traitement de l’échantillon avec du CaCO3 (carbonate de calcium), une quantité de
CaCO3 égale à 1/10 par apport la quantité de l’échantillon est ajoutée puis le mélange,
est incubé à une température de 26-30°C pendant 2 jours (Kämpfer, 2006 ; Oskay
2009) ;
3. Une partie de chaque échantillon n’a subi aucun traitement.
3 Dilution et ensemencement
Une quantité de 1g de l’échantillon est ajouté à 9 ml d’eau distillée stérile ou bien 9 ml
d’eau physiologique stérile et agiter bien comme il faut à l’aide du vortex deux fois pendant 5
minutes pour permettre une bonne dispersion de la charge microbienne dans le tube et avoir
en fin de compte une solution mère (dilution 10-1) ; puis établir une dilution jusqu’à 10-4.
(Arasu et al., 2009 ; Boudemagh, 2007 ; Kitouni, 2007).

Matériels et Méthodes
36
Prendre un volume 0,1 ml de la dilution 10-4 de chaque échantillon et lui ensemencé à la
surface des boites de Pétri contenant l’un des milieux ISP2, GLM, Bennett à l’aide d’un
étaloir afin d’isoler les souches suspectées être des Streptomycètes (Boughachiche et al.,
2008).
4 Isolement des streptomycètes
Après incubation des boites Pétri pendant 7 jours à une température de 30°C et en
conditions d’aérobiose; les streptomycètes ont été isolés selon leurs critères morphologiques,
macroscopiques (Awad et al., 2009), qui sont en général : morphologie des spores, couleur
des spores, mycélium du substrat, pigments solubles et la production de la mélanine
(Balasubramaniam et al., 2011) et microscopiques (colonies circulaires constituées
d’hyphes) par observation directe sous microscope optique (grossissement × 10) (Boussaber
et al., 2012).
1 1 1
1g d’échantillon
10-1 10-2 10-3
9 ml
9 ml
9 ml
9 ml
10-4
ISP2
Ensemencement
0,1 ml Figure 15 : dilution des échantillons

Matériels et Méthodes
37
4.1 Purification des souches
Afin d’obtenir des souches pures, les différentes colonies obtenues sont repiquées et
ensemencées par la méthode des stries sur des boites Pétri contenant le même milieu
d’isolement, puis incubées pendant 1 semaine à 30°C. Cette dernière opération est répétée
jusqu’à obtention de souches pures. Il est recommandé de réaliser le moins de repiquages
possibles pour conserver la stabilité génétique des souches (Kitouni, 2007 ; Boussaber et al.,
2012).
4.2 Détermination de la couleur
Shirling et Gottlieb (1966) recommandent les milieux IS2, ISP3, ISP4 et ISP5 pour la
détermination de la couleur du mycélium du substrat, du mycélium aérien et des pigments
diffusibles. Des boites de Pétri ensemencées avec les souches cibles et misent en incubation
pendant 3 semaines sont comparées à des milieux non ensemencées incubées dans les mêmes
conditions. La présence des pigments diffusibles est constatée par le changement de la couleur
du milieu autour des colonies.
La différenciation et l’isolement a été basé essentiellement sur les couleurs, et l’utilisation
d’un seul milieu nous a permis de déterminer les différences entre les souches isolées puisque
le changement de la composition du milieu affecte la coloration. Dans cette étude on a utilisé
le milieu ISP 2.
4.3 Coloration de Gram
Elle est effectuée selon la méthode classique :
1) Des frottis de colonies répondant aux caractéristiques des streptomycètes sont
préparés sur lame ;
2) Sécher le frottis au-dessus d’une flamme de bec benzène ;
3) Recouvrir les frottis par le cristal violet et les laisser pendant (1) une minute ;
4) Rincer la lame avec de l’eau ;
5) Recouvrir la lame avec du lugol, laisser agir (1) minute exactement ;
6) Laver avec de l’eau ;
7) Décoloration avec de l’alcool, recouvrir la lame en alcool et laisser agir pendant 30
secondes ;
8) Laver avec de l’eau ;

Matériels et Méthodes
38
9) Recouvrir la lame avec la fushine, laisser agir (1) une minute ;
10) Laver avec de l’eau ;
11) Sécher la lame avec un papier absorbant.
À la fin, observer sous microscope optique (grossissement 10×100) les colonies. Celles qui
présentent une coloration à Gram positif sont toutes repiquées sur ISP 2 (Perrier et al., 1997).
4.4 Conservation des souches
Seules les colonies qui présentent une coloration Gram positive et ayant un aspect
filamenteux caractéristique des Streptomycètes, sont conservées dans des boites Pétri ou des
tubes de gélose inclinés pendant la période d’essai à une température de 4°C (Ben Fguira et
al., 2012). Le dénombrement et la différenciation entre les souches dépendent de : lieu de
prélèvement, couleur du substrat, couleur des spores et la pigmentation (Boughachiche et al.,
2008).
Concernant la conservation à long terme, les souches ont été conservées dans le milieu
ISP9 contenant 15-20 % de glycérol (Shepherd et al., 2010 ; Boughachiche et al., 2008 ;
Hopwood et al., 1985).
5 Mis en évidence de l’activité antibactérienne des souches isolées
5.1 Souches de Streptomyces utilisées
Toutes les souches isolées et suspectées appartenir au genre Streptomyces ont été testées
pour leur capacité de produire des substances antibactériennes vis-à-vis des souches
pathogènes.
5.2 Souches pathogènes
Un certain nombre de souches pathogène utilisées pour le test sont : Escherichia coli,
Staphylococcus aureus, Candida albicans, Klebsiella pneumonea ATCC 4352,
Saccharomyces. cervisiae ATCC 2600, Pseudomonas aeruginosa ATCC 27853, Listeria
monocytogenes NCTC 2026 et Salmonella typhi. Ces dernières ont été données gracieusement
par le laboratoire d’hygiène de la wilaya de Saida.

Matériels et Méthodes
39
Le tableau suivant présente les maladies et les infections causées par les souches
pathogènes citées précédemment :
Tableau 08 : quelques caractéristiques des souches pathogènes utilisées dans cette étude
Souche Infections
Escherichia coli Diarrhée, Myocardites, appendicite, infections du foie,
infections urinaires, méningite … etc.
Staphylococcus. aureus Trachéites, Thyroïdites, bronchite, Endocardites, Diarrhée, appendicite, infections urinaires, abcès cutané, infection de la paupière … etc.
Candida. albicans Laryngite, Thyroïdites, infections urinaires, infections pancréatiques, infections du sang … etc.
Klebsiella pneumonea Bronchite, appendicite, infections pancréatiques, infections
urinaires, gastroentérites … etc.
Saccharomyces cerevisiae Rarement incriminé dans les infections
Pseudomonas. aeruginosa Bronchite, Endocardites, appendicite, infections urinaires,
infections pancréatiques … etc.
Listeria monocytogenes Endocardites, méningite, abcès du cerveau, infections du
sang, choc septique …etc.
Salmonella typhi Diarrhée, fièvre entérique, choc septique, myocardite,
gastroentérite, rare arthrites … etc.
5.3 Criblage primaire
Le criblage primaire a été fait pour toutes les souches isolées et suspectées être des
Streptomycètes a été fait par une technique de diffusion sur gélose.
La technique de la double couche c’est elle qui a été utilisée pour l’évaluation de l’effet
antibactérien de toutes les souches isolées, elle consiste à ensemencer les souches de
streptomycètes en touches à la surface des boites de pétri (sous forme de spot) contenant les
milieux gélosés de ISP2, ISP9 et de GLM (Shomura et al., 1979). Après incubation des
boites de Pétri à 28°C pendant douze jours, une suspension dense (Mueller-Hinton)
faiblement gélosée c’est-à-dire une gélose molle (7g.l-1) contenant les souches tests bien
homogénéisée est ajoutée en double couche. Les concentrations des souches-tests sont de

Matériels et Méthodes
40
106UFC.ml-1 par l’utilisation de l’échelle de McFarland (voir annexe) (Boughachiche, 2012 ;
Khattabi, 2005 ; Peterson, 1954)
Les boites sont mises en incubation à 30°C pendant 24h, la présence d’une zone claire
autour des colonies de Streptomyces indique un antagonisme bactérien, l’absence de cette
zone signifie que la souche testée de Streptomyces ne produit pas de molécules
antibactériennes (Boughachiche, 2012 ; Kitouni, 2007).
5.4 Criblage secondaire
Le criblage secondaire est fait pour les souches qui ont présenté une activité
antibactérienne contre toutes les souches testées, ceci est fait après l’extraction des
métabolites secondaires produits par ces souches actives.
Le criblage secondaire consiste à évaluer l’effet antibactérien des extraits issus des souches
sélectionnées. Ceci a été réalisé par la méthode des disques :
o Des cultures jeunes des souches pathogènes ont été étalées sur une couche du
milieu Mueller-Hinton et on les laisse sécher ;
o Préparer des disques de papier de 6 mm de diamètre, imbiber par différents volume
d’extrait et on les laisse sécher aussi ;
o Après que tout soit sec, on dépose les disques sur les surfaces des boites (4
concentrations différentes pour chaque extrait) et juste un seul extrait par boite.
o Incuber les boites à une température de 37°C pendant 24 heures
o L’effet antimicrobien va être visible par la présence d’halos clairs autour des
disques.

Matériels et Méthodes
41
6 Identification des souches sélectionnées : 6.1 Tests physiologiques 6.1.1 Hydrolyse de l’amidon
Ce test est réalisé sur milieu nutritif gélosé contenant 1% d’amidon soluble selon la
méthode de Gordon et Smith (1953). Après 1 jour d’incubation à 30°C, la gélose est
recouverte d’une solution de lugol. L’hydrolyse est ainsi mise en évidence par l’absence de
coloration autour des colonies. A l’inverse, les zones contant de l’amidon se colorent en brun
(Kitouni, 2007).
6.1.2 Hydrolyse de la caséine
L’hydrolyse de la caséine est étudiée selon la méthode de Williams et Cross, (1971), et de
Gordon et Smith (1953) sur un milieu gélosé contant 5% de lait écrémé. L’apparition de
toute zone claire autour des colonies après 14 jours d’incubation à 30°C témoigne de
l’hydrolyse de la caséine (Kitouni, 2007).
6.1.3 Dégradation de l’urée
La dégradation de l’urée est mise en évidence, après ensemencement du milieu contenant
du rouge de phénol (annexe) par 0.05 mL de l’inoculum général. Le résultat est considéré
positif quand la couleur du milieu vire à l’orangé après incubation pendant sept jours à 28°C
(Boudemagh, 2007).
6.1.4 Dégradation du tween 80
Dégradation du tween 80 (Sierra, 1957) le milieu utilisé est composé de : Tween 80 : 10
ml ; NaNO3 : 1 g ; extrait de levure : 5 g ; solution saline: 50 ml ; CaC12, 2H2O: 0,1 g ; eau
distillée q.s.p. 1000 ml; agar : 18 g. pH 7,2. La dégradation du tween 80 se manifeste par une
auréole opaque autour des colonies (Smaoui, 2010).

Matériels et Méthodes
42
6.2 Tests biochimiques 6.2.1 Utilisation des différentes sources de carbone
Le milieu ISP 9 (annexe) a été utilisé dans cette étude. Ce milieu est exempt de toutes
formes de carbone. Les différentes sources de carbone testées sont celles recommandées par
Shirling et Gottlieb (1966) et par Pridham et Gottlieb (1948), elles sont les suivantes:
saccharose, L-arabinose, D-glucose, lactose, D-mannitol, raffinose et le galactose.
Des solutions de ces sources de carbone, à 10 % (p/v) de concentration, sont préparées et
stérilisées par filtration. L’ensemble des sources de carbones sont additionnées aseptiquement
au milieu de base de manière à obtenir une concentration finale de 1 % (p/v).
Ces différents milieux ont été répartis dans des boîtes de Pétri, ensemencés par 0.01 mL de
l’inoculum lavé préparé précédemment et incubés pendant sept jours à 28°C.
La croissance sur ces milieux est comparée avec un témoin négatif (milieu sans source de
carbone) et un témoin positif (milieu avec glucose). Si la croissance obtenue (nombre de
colonies) est supérieure à celle du témoin négatif, le résultat est considéré positif
(Boudemagh, 2007).
7 Extraction et concentration des métabolites secondaires 7.1 L’extraction
L’extraction est faite comme suit (Bouras et al., 2013 ; Al-Askar, 2012 ; Ben Fguira et
al., 2012 ; Aouiche et al., 2012 ; Kavitha et al., 2010 ; Arasu et al., 2009 ; Ilić et al., 2005)
Trois millilitres (105 spores/ml) de chaque suspension sporale des souches sélectionnées
sont ensemencés dans des Erlenmeyers de 500 ml contenant 150 ml du milieu Bennett (voir
annexe) Les Erlenmeyers sont ensuite placés dans un agitateur thermostaté 28°C et avec
l’assurance de l’aération pendant 5 jours.
Ensuite l’extraction des métabolites secondaires a été réalisée selon les étapes décrites ci-
dessous :
les cultures sont centrifugées à 8000 rpm pendant 10 minutes pour éliminer les
cellules et avoir juste le surnageant.
est extrait par un volume égale de solvant (v/v) comme l’hexane, le chloroforme,
l’éthyle acétate et l’éther, dans ce travail le solvant choisi a été le chloroforme

Matériels et Méthodes
43
il faut bien mélanger le solvant avec le surnageant par une agitation vigoureuse
pendant au moins 20 minutes (Thakur et al., 2009b)
Laisser le mélange se séparer en deux phases, une aqueuse et l’autre organique qui
contient les métabolites secondaires ;
Séparer la phase organique de la phase aqueuse à l’aide d’une ampoule à
décompter ;
7.2 Concentration des métabolites par utilisation d’un Rotavapor
La concentration est faite à l’aide d’un appareil le rotavapor
Avec un rotavapor, une seule étape de séparation rapide s’effectue d’une manière
affectueuse sur le produit (phase organique). La base de cette transformation est d’évaporer
puis condenser le solvant en utilisant un rota-vapeur sous pression. Séparer les composants
sous pression augmente le degré de performance et diminue le risque d’endommager les
molécules présentes.
Dans cette étude on a suivi les étapes suivantes :
1) Faire chauffer l’eau dans un bain marie à une température de 40°C ;
2) Mettre la phase organique dans l’ampoule et faire baigner l’ampoule dans le bain
marie ;
3) Allumer le moteur pour augmenter la pression dans le rotavapor ;
4) N’arrêter pas l’opération jusqu’à que le solvant soit évaporer entièrement ;
5) Récupérer ce qui reste dans l’ampoule (métabolites secondaires) dans 1 mL
d’acétate d’éthyle et mettre le mélange dans un tube Eppendorf ;
6) Conserver le tube à une température de 4°C avant l’utilisation (Arasu et al., 2009).

Matériels et Méthodes
44
Figure 16 : Rotavapor (BÜCHI Rotavapor R-200) utilisé pour la concentration des métabolites secondaires
8 Spectre UV des extraits
Après l’extraction des métabolites secondaires des 3 souches, une quantité suffisante de
chaque extrait a été mise dans une cuve à Spectrophotomètre, et toute une analyse de
l’absorbance de ces extraits a été déterminée de 330 nm à 800 nm par un spectrophotomètre
(Maleki & Mashinchian 2011; Ilić et al., 2005).

Matériels et Méthodes
45
9 Chromatographie sur couche mince (CCM) :
Une première pré-identification des substances présentes dans les extraits a été faite à
l’aide d’un spectrophotomètre, une quantité suffisante (1 ml) est mise dans la cuve afin que le
spectrophotomètre nous donne un graphe représentant les absorbances de 300-600nm (Anzai
et al., 2008 ; Kitouni, 2007 ; Slavica et al., 2005). Cette opération nous oriente sur la nature
des molécules présentes ainsi que leur intervalle d’absorbance.
La deuxième identification préliminaire est faite par une chromatographie sur couche
mince, le but étant de connaitre nature ainsi que le nombre des molécules présentes dans les
dfférents extraits. La CCM a été réalisée comme suit (Bouras et al., 2013 ; Ben Fguira et al.,
2010 ; Boudjella et al., 2006):
1. Une plaque de gel de silice (60 F254) est déposé horizontalement sur la paillasse et
une ligne est tracée loin de 2 cm et en parallèle par rapport au bord inférieur de la
plaque ;
2. Les dépôts des échantillons sont faites sur la ligne tracée et chaque échantillon est
loin de l’autre de 1 cm et la même distance loin des bords latéraux (Arasu et al,
2009 ; Gibbons, 2006) ;
3. Une quantité de 6 µL est déposée à l’aide d’une micro-seringue pour chaque
échantillon (Kitouni 2007, Arasu et al, 2009);
4. Sécher les dépôts à l’aide d’un séchoir ;
5. Préparer la chambre CCM par l’ajout du solvant (chloroforme), et le solvant doit
remplir une hauteur de 1 cm ;
6. Déposer la plaque dans la chambre verticalement comme la montre la figure (17) et
refermer avec le couvercle ;
7. Laisser le chromatogramme se développer ;
8. Après la migration, il faut bien sécher la plaque avant la révélation ;
9. La révélation est faite à l’aide des rayons UV, la plaque est déposé horizontalement
sur une plaque UV (Gibbons, 2006 ; Arasu, 2009 ; Smaoui 2010).

Matériels et Méthodes
46
Figure 17 : illustration de la CCM
10 Identification moléculaire des souches 10.1 Extraction d’ADN
L’analyse du gêne ADNr 16S est un taxon de choix pour l’identification des souches
isolées. L’utilisation d’un kit qui est « Ultra Clean microbial » ; un kit conçu pour isoler une
grande quantité d’ADN génomique des microorganismes. Une grande variété de
microorganismes et de champignons a été testée par ce kit selon le guide.
Protocole :
L’ADN totale bactérien de nos isolats à été préparer selon la méthode d’extraction
de Gevers et al., (2001) et qui se résume dans étapes suivantes :
1. Une colonie bactérienne est suspendue dans 1 ml d’eau distillée stérile dans un
tube Eppendorf stérile,
2. La suspension bactérienne est centrifugée à 3000xg pendant 10 min à 4³C,
3. Après centrifugation congeler à -20°C au moins pendant 1h ;
4. Le culot dégelé, est ensuite laver par 01ml de tampon TES (6.7% sucrose, 50 mM
Tris-HCl, pH 8.0, 1 mM EDTA) ;

Matériels et Méthodes
47
5. Re-suspendre ensuite dans le Tampon STET (8% sucrose, 5% Triton X-100, 50
mM Tris^HCl, pH 8.0, 50 mM EDTA) ;
6. Ajouter 75µl de tampon de lyse (TES contenant 1330U/ml de mutanolysin et
40mg/ml de lysozyme), à la suspension et incuber à 37°C for 1h.
7. Après préchauffage de 37°C, ajouter 40µL de SDS 20% dans du tampon TE (10
mM Tris-HCl, 1 mM EDTA, pH 8.0) en présences de billes en verre ;
8. Agiter au vortex pendant 60 secondes et incuber à 37°C pendant 10min,
9. Continuer l’incubation à 65°C pendant 10min encore ;
10. Ajouter 100µl du tampon TE et le lysat est extraie avec 1 volume de
Phénol/Chloroforme/ isoamylalcohol.
11. Les différentes phases obtenues sont séparé par centrifugation à 12000xg, pendant
10 min ;
12. La phase aqueuse est doucement mixer avec 70µl d’une solution 5 M NaCl et 1ml
d’isopropanol ;
13. L’ADN est précipité dans la glace pendant 15min.
14. Centrifugation à 12000xg pendant 10min afin de collecter l’ADN en culot
15. Le culot est lavé avec l’éthanol 70% froid
16. Re-suspendre le culot dans 250µl d’eau distillé stérile, 250µl de 1.4M sodium
chloride et 50µl de CTAB 10% ;
17. Incuber à 60°C pendant 30 minutes ;
18. La mixture est ensuite extraite par 1volume Phénol/Chloroforme/ isoamylalcohol,
19. Après centrifugation l’ADN est précipiter à partir de la phase aqueuse par 1ml
d’éthanol absolu froid ;
20. Lavage par l’éthanol 70%
21. Centrifugation 12000 x g pendant 10 min afin de collecter l’ADN purifié en culot

Matériels et Méthodes
48
22. le culot d’ADN pure est dissous finalement dans l’eau distillé stérile ;
23. L’ADN est conservé à -20°C jusqu'à son utilisation
Amplification du gène ADNr 16S
Les séquences (amorces) qui permettent d’amplifier le gène ADNr 16S par PCR sont des
primers universels 8UA et 1492 R. Les séquences sont : 8AU, 5´-AGA GTT TGA TCC TGG
CTC AG3´ ; et 1492R, 5´-TAC GGG TAC CTT GTT ACG ACT T-3´.
10.2 La PCR
La PCR (Polymérisation Chain Reaction) est technique permettant d’amplifier in vitro des
séquences d’ADN par répétition des réactions d’élongation en présence d’une ADN
Polymérase et d’amorces nucléotidiques spécifique (Tagu, 1999). La PCR demande pour
chaque réaction les ingrédients suivants présents dans le tableau ci-dessous :
Tableau 09 : les ingrédients de la PCR pour chaque échantillon
Produit Quantité/échantillon
ADN 1 µg
Tampon 2,5 µl
Taq polymérase 0,25 µl
Amorce 1 0,5 µl
Amorce 2 0,5 µl
DNTP 2 µl
Eau bi-distillée stérile 14,25
Après réalisation des mélanges, les microtubes PCR sont déposés dans un « thermocycler »
de type (TECHNE TC-312) et l’amplification s’est faite avec le programme suivant : 15
minutes à 95ºC pour la température initiale d’activation, 35 cycles de polymérisation : 1
minute à 94ºC pour la dénaturation, 1 minute à 60ºC pour l’hybridation, 1 min 30 sec à 72ºC
pour l’élongation et 40 minutes à 72ºC pour l’élongation finale.

Matériels et Méthodes
49
Figure 18 : thermocycler (TECHNE TC-312)
10.3 Electrophorèse des acides nucléiques
Après l’amplification, les échantillons sont déposés dans les puits du gel d’agarose afin de
faire la migration
10.3.1 La préparation du gel d’agarose :
1. Peser une quantité de 1,5 g d’agarose (pour un pourcentage de 1,5 %) et la mettre dans
un Erlenmeyer puis ajouter 100 ml de TBE (45 mM Tris-borate, 1 mM EDTA) et
mélanger le tout ;
2. Mettre l’erlenmeyer dans la microonde pendant 30 secondes jusqu’à la dissolution
complète de l’agarose ;
3. Faire sortir l’erlenmeyer du four microonde et laisser refroidir, mais avant qu’il se
gélifie en ajoute 4 µg de bromude d’éthidium (EtBr) et on mélange jusqu’à
dissolution ;
4. Couler le gel dans un cuve contenant un peigne (pour faire des puits) ;
5. Laisser le gel se refroidir avant d’enlever délicatement le peigne ;
6. Mettre le gel dans une position horizontale par rapport à la base de l’électrophorèse,
cette dernière qui doit être remplie suffisamment de TBE afin de déposer le gel. Les
puits doivent être du côté de la cathode.

Matériels et Méthodes
50
10.3.2 Dépôt des échantillons :
Le dépôt des échantillons se fait comme suit :
1) Mélanger la solution de charge (bleu de bromophénol) 5 µL avec une un volume de
10 µL de chaque échantillon contenant l’ADN déjà amplifié sur une plaque
contenant de petits puits ;
2) Mélanger le marqueur de taille aussi avec la solution de charge (10 µL) avec 5 µL
de la solution de charge ; il faut bien noter qu’il ne faut pas mélanger les
échantillons ;
3) Prendre le total du mélange de chaque échantillon et le déposer prudemment dans
les puits du gel.
La figure ci-dessous présente le dépôt des échantillons ainsi que le sens de migration
Figure 19 : exemple sur le résultat d’une éléctrophorèse des acide nucléiques 1 : marqueur de taille
2, 3, 4 et 5 : des échantillons

Matériels et Méthodes
51
10.3.3 Migration
1) La migration se fait de la cathode vers l’anode, elle demande d’exercer un voltage de
90 à 100 volts entre les deux pôles de l’électrophorèse ;
2) La migration dure de 30-40 minutes jusqu’à ce que le colorant (bleu de bromophénol)
atteigne le bord du gel après migration ;
3) Après migration, le gel est enlevé de l’appareil est mis sous UV pour la visualisation.
Figure 20 : électrophorèse pour la séparation des fragments d’ADN

Matériels et Méthodes
52
10.3.4 Visualisation :
La visualisation de l’ADN après migration se fait à l’aide d’une lampe UV ou bien à l’aide
d’une plaque à UV comme il est le cas pour cette étude. Le gel est déposé horizontalement sur
la plaque et ensuite la plaque est allumée pour voir les bandes d’ADN, des photos sont prises.
Figure 21 ; plaque UV utilisée pour la visualisation des acides nucléiques

Résultats &
Discussion

Résultats et Discussion
53
Beaucoup de travaux s’intéressent à l’isolement et la caractérisation de nouvelles souches
de Streptomyces qui produisent de nouveaux antibiotiques ou bien de nouvelles molécules
bioactives d’intérêt, et cela depuis la découverte de ce genre jusqu’à ce jour et ces travaux
montrent l’importance de ce sujet, nous énumérons quelques travaux (Al-Askar, 2012 ;
Bouras et al., 2013 ; Atta, 2012 ; Boussaber et al., 2012 ; Arasu et al., 2009 ; Thakur et
al., 2007 ; Taddei et al., 2006 ; Sajid et al., 2009 ; Awad et al., 2009 ; Ceylan et al., 2008).
1 Effet des prétraitements :
Une variété de milieux sélectifs et de traitements ont été utilisés pour avoir un bon
isolement des streptomycètes des différents habitats (Bouras et al., 2013). Il a été rapporté
que les prétraitements sont extrêmement utiles pour l’isolement de ce genre bactérien. Le
traitement avec le CaCO3 a donné de bons résultats (au moyenne 1,5x105 UFC/ml) par
rapport au traitement avec la chaleur (en moyenne 4x104 UFC/ml), soit le nombre de
Streptomyces présents dans les boites qui étaient plus élevés pour les échantillons traités par
les carbonate de calcium, soit qu’il y’avait moins de contamination par les moisissures.
N’oublions pas que les échantillons qui n’ont pas étaient traités du tout ont donnés moins de
Streptomycètes (en moyenne 1x104 UFC/ml) que les échantillons traités soit par la chaleur ou
par le CaCO3.
Ceci est similaire à des travaux antécédents (Kitouni, 2007 ; Boudemagh, 2007) et montre
que le meilleur traitement des échantillons pour l’isolement des Streptomycètes est le
traitement avec le CaCO3. Selon El-Nakeeb & Lechevalier (1963), l’ajout du CaCO3 conduit
à l’augmentation du nombre des Streptomycètes dans les échantillons lors de l’isolement.
Jeffrey (2008) cite que le CaCO3 réduit le nombre des bactéries à gram négatif dans les
échantillons, donc il y a plus de chance pour isoler les Streptomycètes qui sont des bactéries à
gram positif
2 Isolement des Streptomyces
L’isolement a été fait suivant les critères décrits dans la partie « matériel et méthodes »
(page 37), les observations macroscopiques et microscopiques ont permis l’isolement d’un
nombre de 85 souches des différents échantillons, les figure du N° 24 à N° 35 présentent
quelques souches avec les observations microscopiques des colonies.

Résultats et Discussion
54
La figure ci-dessous (Figure 22) présente le résultat d’un ensemencement d’un échantillon
de sol après une uncubation de 5 jours, les flèches indiquent les colonies suspectées être des
Streptomycètes, cela selon la forme de la colonie et sa surface, ainsi que la coloration de
Gram.
La forme des colonies présentes dans cette photo ci-dessus (figure 22) sont presque
similaires à des photos qui sont dans le travail de (Oskay, 2009) (figure 23) où elles sont bien
sélectionnées.
2.1 Détermination des couleurs :
Durant le développement des Streptomyces, la couleur du mycélium du substrat est
diffèrent de la couleur du mycélium aérien.
2.1.1 Couleurs du mycélium du substrat
- Les couleurs qui ont été enregistré pour le mycélium du substrat sont les suivantes :
Beige, Rouge, Marron, Vert olive, Rose, Jaune et Pourpre ; ceci a été constaté par des
autres chercheurs dans plusieurs travaux comme (Arasu et al., 2009 ; Taddei et al.,
2006)
Figure 22 : les colonies suspectées être Streptomycètes
Figure 25 : observation de la colonie de SB1 Figure 24 : observation macroscopique de SB1
Figure 23 : Colonies de Streptomyces (Oskay, 2009)
Observation macroscopique Observation de la colonie (G 10×10)
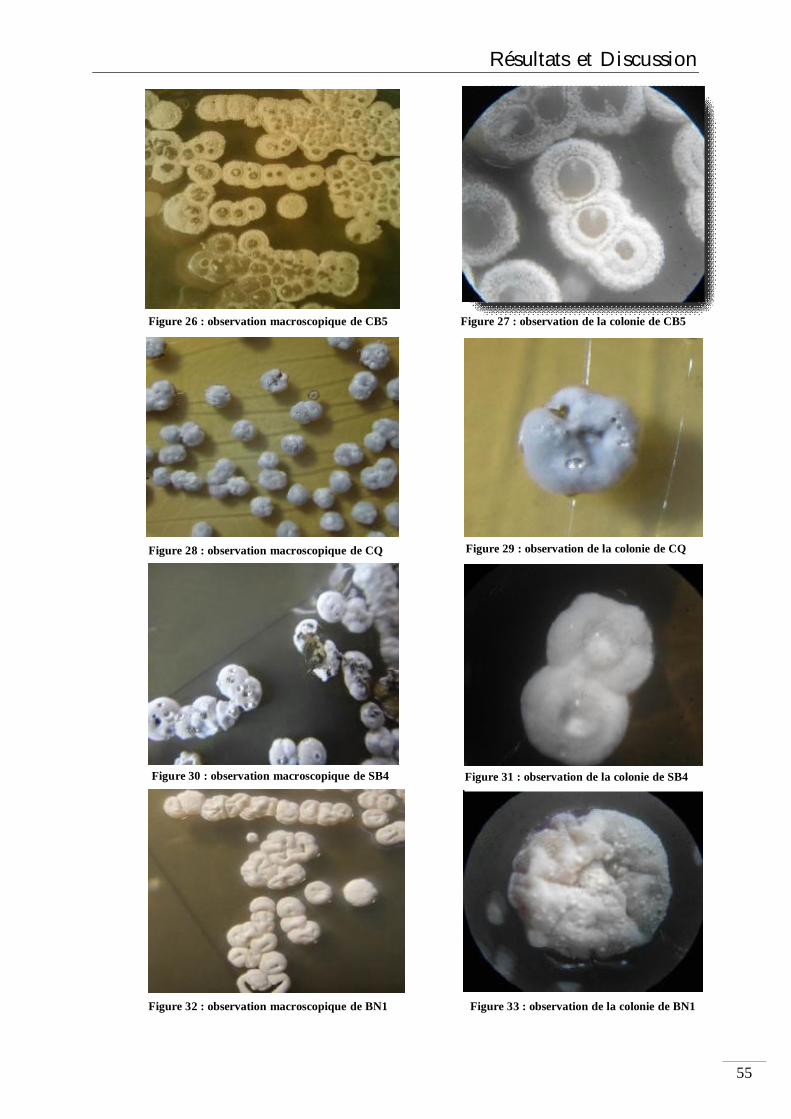
Résultats et Discussion
55
Figure 27 : observation de la colonie de CB5
Figure 29 : observation de la colonie de CQ
Figure 30 : observation macroscopique de SB4
Figure 32 : observation macroscopique de BN1
Figure 26 : observation macroscopique de CB5
Figure 28 : observation macroscopique de CQ
Figure 31 : observation de la colonie de SB4
Figure 33 : observation de la colonie de BN1

Résultats et Discussion
56
2.1.2 Les couleurs du mycélium aérien :
Figure 36 : souche SM2.2 Figure 37 : souche BN1
Figure 38 : souche SM2.6 Figure 39 : souche SM2.11
Figure 40 : souche SM2.3 Figure 41 : souche SM X4.2
Figure 34 : observation macroscopique de N3 Figure 35 : observation de la colonie de N3

Résultats et Discussion
57
Généralement les couleurs du mycélium aérien ne sont pas les même que le mycélium du
substrat et la plupart ont une couleur blanche et des autres couleurs comme le : gris, violet,
beige, noir, marron, vert, jaune, rose blanc et ça a été constaté par (Balasubramaniam et al,
2011)
2.1.1 Production des pigments diffusibles :
La production des mélanoïdes était présente chez quelques souches isolées et il y’avait
d’autres souches qui diffusaient des pigments de couleur différentes : rouge, jaune violet, vert
et marron. Il faut bien noter que la production des pigments dépend généralement de la
composition du milieu et des conditions de culture (Kämpfer, 2006).
La production des pigments est un des caractères des Streptomycètes, la coloration des
mycéliums du substrat et celui de l’aérien est due à des pigments mais qui ne sont pas
diffusibles. La mélanine est un substrat commun produit par les animaux, les plantes et les
microorganismes, elle a un poids moléculaire très élevé formée par la polymérisation
oxydative des produits phénoliques ou indoliques (Vasanthabharathi et al, 2011) on peut la
voir généralement en couleur marron.
Figure 42 : souche SM2.8 Figure 43 : souche B6

Résultats et Discussion
58
2.2 Dénombrement des souches suspectées être des Streptomycètes
D’après les résultats obtenus, les Streptomycètes sont omniprésentes dans tout type de sol
prélevé, quel que soit sa nature et sa composition. D’après (Goodfeilow et al 1992 ; Rintala
2003) les Streptomycètes sont ubiquitaires et colonisent facilement le sol, ceci est due à leur
faculté de croitre comme une masse d’hyphe végétative qui se différencie en spores
permettant la dispersion et la persistance.
Un grand nombre de travaux sur l’isolement des Streptomycètes à partir de
l’environnement terrestre et de nombreux lieux comme : sol agricole, forets, rhizosphère
(Ceylan et al, 2008 ; charbon (Chroňákova et al, 2010), caves (Groth & Saiz-Jimenez,
2010) et même de l’environnement marin ont été décrits (Karthik et al, 2010 ; Solanki et al,
2007)
Williams et al (1972) rapporte que les Streptomycètes résistent à la dessiccation, alors
elles peuvent rester une longue période en état de dormance, c’est ce que cite Kämpfer
(2006) qui dit que les spores de streptomycètes peuvent résister pendant 70 ans en état de
dormance.
Figure 44 : pigments diffusibles produits par les souches de Streptomycètes A: SM2.6, B: TB1 et C: SM2.4
A B
C

Résultats et Discussion
59
L’isolement de types différents de sol a révélé que le sol des régions aride de la wilaya de
Saida contient plus de Streptomycètes par rapport aux autres types de sol, même s’ils sont de
la même région, et en plus les sols agricoles contiennent plus de Streptomycètes que les autres
types de sol.
Selon le travail de Boussaber et al. (2012), le plus grand nombre de souches
d’actinomycétales se trouvent dans les sols où le taux d’humidité est minime (vers 14%)
contrairement au sol très humides (50-61%) où il y’a moins de souches de cet ordre. Sachant
que dans ce travail le chercheur n’a pas utilisé un sol désertique. Ces résultats sont presque
similaires aux résultats obtenus dans ce travail, le sol de la région humide a donné moins de
souches par rapport aux autres types de sol comme cela a été rapporté aussi par Routien
(1960) où il a démontré que il y’a une corrélation entre le manque d’humidité dans le sol et la
haute présence des Streptomycètes.
Les sols agricoles ont donné un nombre de Streptomycètes plus élevé par rapport aux
autres types de sol, ceci a été déterminé par des travaux antécédents comme celui de Jeffrey
(2008). Mais selon le travail de Oskay (2009) il n y’a pas de différence entre le sol agricole et
non-agricole en terme de contenance en actinomycètes, le problème réside au niveau du
milieu de culture utilisé et sa contenance en éléments minérales lors de l’isolement. Alors il
été préférable de faire tout une étude comme celle de Suutari et al (2002) sur les milieux de
culture appropriés à l’isolement des streptomycètes.
Histogramme présentant le pourcentage des souches isolées de chaque type de sol :
sol agricole Saida35%
zone humide Saida6%
sol desertique Bayadh
7%
sol agricole Mostaganem
18%
sol agricole Oran20%
autres types de sol14%
souches
Figure 45 : Pourcentage des souches isolées selon les types de sol

Résultats et Discussion
60
Coloration de Gram
La coloration de Gram des souches isolées a montré qu’elles sont toutes des bactéries Gram positif, ainsi qu’elles représentent la même forme des Streptomyces qui se présentent sous microscope (grossissement 10 x 100).
Souches Observation Macroscopique Coloration de Gram
N1
Mos 5
CQ1
Figure 46 : coloration de Gram de quelques souches

Résultats et Discussion
61
3 Mise en évidence de l’activité antibactérienne des souches
isolées
L’activité antibactérienne était très remarquable en fonction des zones d’inhibition lors des
tests d’antagonisme des souches de Streptomyces vis-à-vis des souches pathogènes.
Concernant le pourcentage des souches actives contre un pathogène, il y a eu plus de
25,88% du nombre global des souches suspectées être des Streptomycètes. Dans le travail
d’Anzai et al (2008) le nombre global des souches qui ont une activité antimicrobienne est de
l’ordre de 30 % ; sachant que Ben Fguira et al. (2010) dans leur travail, ils ont trouvé que
58,84 % des souches isolés des oasis ont une activité antibactérienne ; ainsi que le travail de
Jeffrey et al., (2008) a montré que 38,8% des souches isolées dans l’étude ont une activité
antibactérienne contre les souches gram positif et gram négatif utilisant presque la même
méthode que celle utilisée dans ce travail.
Il est bien clair que les résultats obtenus dans ce travail sont moindres que les résultats des
travaux cités précédemment, cela peut s’expliquer par différence dans la méthode utilisée
dans l’évaluation de l’effet antibactérien sachant que les deux chercheurs cités précédemment
d’Anzai et al (2008) & Ben Fguira et al (2010) ont utilisé 2 méthodes différentes pour
l’évaluation de l’effet (criblage primaire), la méthode de la double couche ainsi que la
méthode des stries perpendiculaires tandis que dans ce travail, on a utilisé que la première
méthode.
Il sera souhaitable dans les prochains travaux d’intégrer au moins deux méthodes en même
temps dans l’évaluation de l’effet antibactérien. Après ce criblage, 3 souches ont été
sélectionnées pour les prochains tests (tests physiologiques, biochimiques, extraction et pré-
identification des molécules élaborés et l’identification moléculaire).
Ces 3 souches ont présenté un effet antibactérien très remarquable par rapport aux autres
souches, c’est le critère pour lequel ces souches ont été choisis, le même paramètre a été
choisi par Jeffrey et al., (2008).

Résultats et Discussion
62
Figure 47 : l’activité antibactérienne des souches isolées

Résultats et Discussion
63
Tableau 10 : tableau représentant les caractéristiques et le diamètre des zones d’inhibition en millimètre des souches sélectionnées lors du criblage primaire
Souches sélectionnées
SM2.2 SM1B1 TB1
Souc
hes p
atho
gène
s
S. aureus 27 20 32
S. typhi 35 25 27
C. albicans 00 00 00
K. pneumonea 10 10 15
P. aeruginosa 09 05 12
E. feacalis 30 25 20
E. feacium 22 20 33
L. monocytogenes 00 00 00
S. cervisiae 00 00 00
Origine* Ech 1 Ech1 Ech6
Couleur du mycélium végétatif Grise Marron pale Crème
Couleur du mycélium aérien Blanche Blanche Blanche
Couleur des spores Grise Blanche Blanche
Parmi les 3 souches sélectionnées et suspectées être des streptomycètes, la souche TB1 a
présenté une activité antibactérienne plus que les deux autres souches. Il est bien claire que
toutes les souches sélectionnés n’ont pas exercé un effet antimicrobien contre C. albicans et S.
cerviseae.
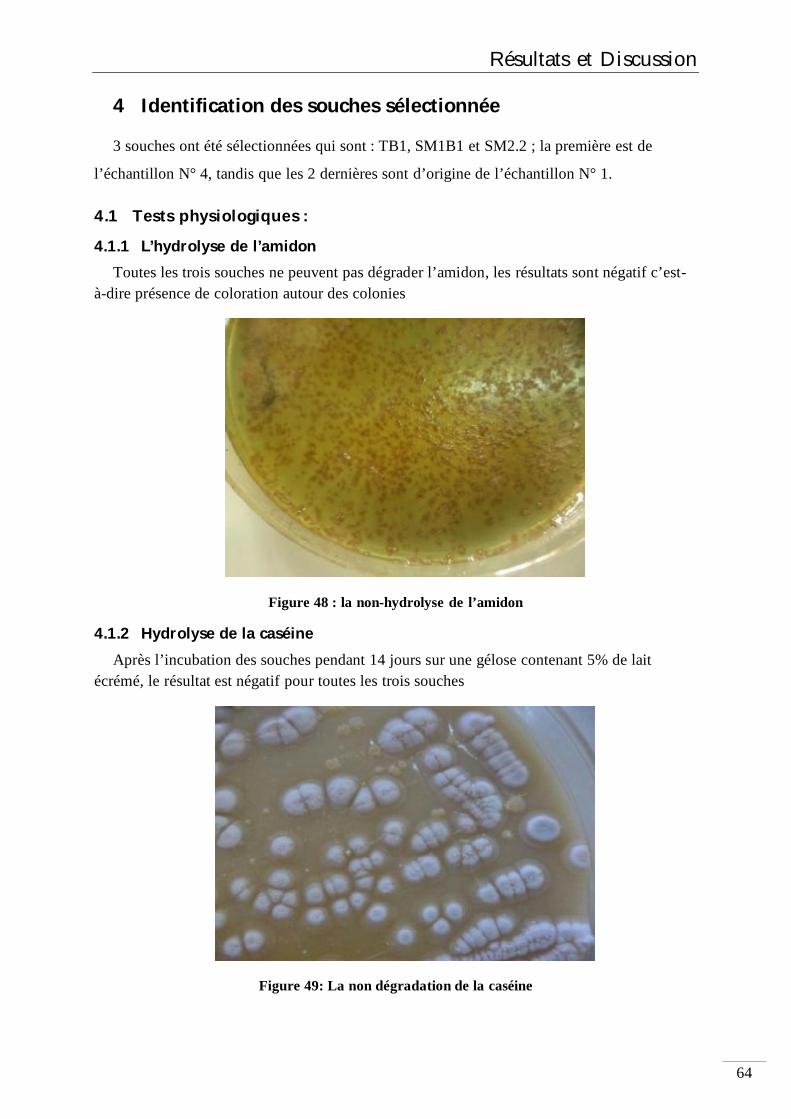
Résultats et Discussion
64
4 Identification des souches sélectionnée
3 souches ont été sélectionnées qui sont : TB1, SM1B1 et SM2.2 ; la première est de
l’échantillon N° 4, tandis que les 2 dernières sont d’origine de l’échantillon N° 1.
4.1 Tests physiologiques :
4.1.1 L’hydrolyse de l’amidon
Toutes les trois souches ne peuvent pas dégrader l’amidon, les résultats sont négatif c’est-à-dire présence de coloration autour des colonies
Figure 48 : la non-hydrolyse de l’amidon
4.1.2 Hydrolyse de la caséine
Après l’incubation des souches pendant 14 jours sur une gélose contenant 5% de lait écrémé, le résultat est négatif pour toutes les trois souches
Figure 49: La non dégradation de la caséine
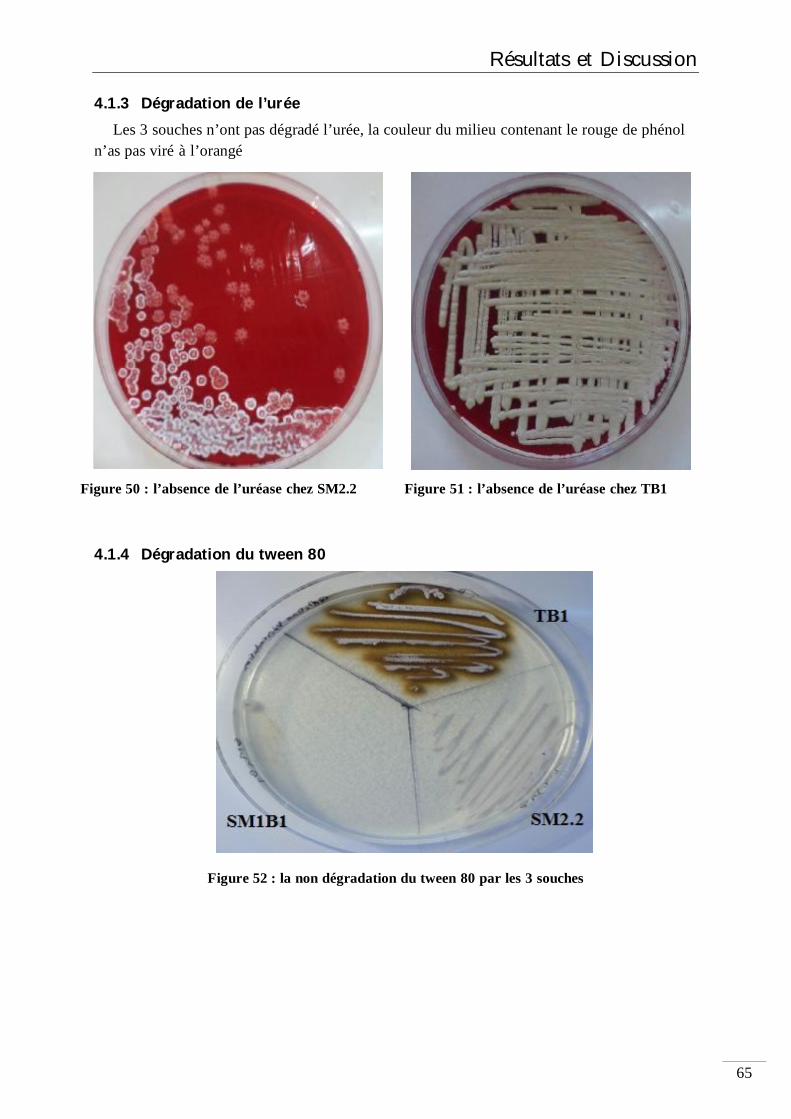
Résultats et Discussion
65
4.1.3 Dégradation de l’urée
Les 3 souches n’ont pas dégradé l’urée, la couleur du milieu contenant le rouge de phénol n’as pas viré à l’orangé
4.1.4 Dégradation du tween 80
Figure 52 : la non dégradation du tween 80 par les 3 souches
Figure 50 : l’absence de l’uréase chez SM2.2 Figure 51 : l’absence de l’uréase chez TB1

Résultats et Discussion
66
4.2 Tests biochimiques
Dégradation des sucres
Comme il a été décrit dans la partie « matériels et méthodes », les souches ont dégradé presque tous les sucres.
Figure 53 : dégradation des sucres par la souche SM2.2
Les tests concernant l’utilisation des différentes sources de carbone n’indique pas
l’appartenance ou non des souches isolées aux Streptomycètes, mais ils nous renseignent sur
les habitudes et les caractères des souches selon Shirling et Gottlieb (1966). La plupart des
études taxonomiques ou bien ceux de criblage qui concernent les Streptomycètes utilisent ces
tests pour juste connaitre le métabolisme et bien caractériser les souches pour des études
ultérieures (Bouras et al., 2013 ; Suneetha et al., 2011 ; Nonoh et al., 2010 ; Dehnad et al.,
2010)

Résultats et Discussion
67
Le tableau 11 : présente les habitudes et les caractéristiques des 3 souches TB1, SM1B1 et
SM2.2
Tableau 11 : résultats des tests des 3 souches
Souches
TB1 SM1B1 SM2.2
Test
s
Production de mélanoïdes
+ - -
Mannitol + + -
Saccharose + + +
Lactose - + -
Raffinose + + +
D-galactose + + +
D-glucose + + +
L-arabinose + + +
Amidon - - -
Caséine - - -
Uréase - - -
Tween 80 - - -
(+) : positif, (-) : négatif

Résultats et Discussion
68
5 Extraction des métabolites secondaires
Après une incubation de 7 jours avec
agitation, la culture est centrifugée et le
surnageant est récupéré. Le surnageant
est mélangé avec le même volume de
chloroforme et agité pendant 20 min,
pour bien extraire les métabolites
présentes dans ce m mélange.
La séparation entre les deux phases
(phase aqueuse et phase organique) a été
faite par une ampoule à décanter.
La phase organique est récupérée
dans un flacon stérile, ensuite cette
phase organique qui contient les
molécules extraites dissoutes dans le
solvant est concentrée à sec à l’aide d’un
Rotavapor afin d’avoir l’extrait brut.
Culture liquide
Phase aqueuse +
Phase organique
Extraction & Séparation
Purification
Figure 54 : extraction des métabolites secondaires

Résultats et Discussion
69
Toutes les études ciblant l’extraction des métabolites secondaires à partir des cultures de
Streptomycètes utilisent de différents solvants. Le choix du chloroforme comme solvant
d’extraction dans cette étude est venu après la consultation de différents travaux dans lesquels
le chloroforme présentait le meilleur solvant pour l’extraction des métabolites secondaires
(Arasu et al., 2009). Selon Gibbons (2006), l’extraction des métabolites comme les poly-
phénoliques et les alcaloïdes nécessitent un solvant assez fort et polaire, sachant que le
constant diélectrique du chloroforme est de 4,8 et il est assez élevé par rapport à l’hexane,
pentane, cyclohexane, benzène, toluène, Diethyl éther et le Diméthyle sulfoxide.
La meilleure façon pour avoir une bonne quantité de métabolites secondaires est d’utiliser
un fermentateur lors de la culture, l’augmentation du volume de la culture implique une bonne
quantité de molécules extraites et vis vers ça.
Il faut bien noter que l’utilisation de plusieurs types de solvants pour l’extraction, donne
une idée plus claire sur le métabolisme secondaire des souches, car chaque solvant a ses
propres caractéristiques (polarité, température d’ébullition).
6 Evaluation de l’effet des extraits
Les extraits ont présenté un effet antibactérien assez remarquable surtout envers E. coli et S
aureus, cet effet a été très limité pour des souches K. pneumeuniae, P. aeruginosa et les
autres souches pathogènes utilisées dans ce travail.
Figure 55: effet des extraits sur E. coli A : extrait de TB1 contre E.coli ; B : extrait de SM1B1 contre E.coli
A B

Résultats et Discussion
70
Le meilleur extrait qui a présenté un effet antibactérien est celui de la souche TB1, avec
une moyenne de 15 mm pour le diamètre de la zone d’inhibition, tandis que les autres extraits
n’ont pas pu exercer le même effet que celui réalisé lors des tests d’antagonisme.
Il bien clair d’après ces résultats que l’extraction n’a pas extrait les molécules qui ont un
effet antibactérien du fait que l’extraction a été réalisée juste par un seul solvant, il y’en a pas
mal d’hypothèses qui justifient ça. Le solvant n’a pas pu rattacher aux molécules présentes
dans le surnageant des cultures, car il est de nature polaire et qui peut être au contraire de la
nature des molécules secrétées par les souches; ou bien les souches ont besoin d’un précurseur
externe qui n’as pu être mis en place lors des cultures ; et autres éléments qui peuvent être la
cause de l’absence des molécules antibactériennes dans les extraits.
L’étude des Streptomycètes est très difficile à cause de leur métabolisme, qui est
généralement dépendant des conditions de culture, soit les nutriments ou bien les autres
conditions de culture comme l’aération.
7 Courbes d’absorbance des extraits
Figure 56 : courbe d’absorbance de l’extrait de la souche TB1

Résultats et Discussion
71
Figure 57: courbe d’absorbance de l’extrait de la souche SM2.2
Figure 58 : courbe d’absorbance de l’extrait de la souche SM1B1

Résultats et Discussion
72
Les extraits ont donné des courbes presque similaires d’absorbance, ce qui montre que
probablement les extraits contiennent des molécules de même nature, mais ça ne veut pas dire
qu’elles ne sont pas différentes. Les pics des extraits sont différents comme le montre le
tableau suivant.
Tableau 12 : les pics des courbes d’absorbance des extraits
Souche Pics
TB1 1,580 à 330 nm
SM2.2 0,276 à 330nm
SM1B1 0,188 à 330 nm
D’après les courbes d’absorbance, on note que la lecture sur spectrophotomètre a donné la
valeur la plus élevée à sa limite de détection (330 nm), ce qui veut dire que normalement le
pic des trois graphes se trouve dans l’UV. Donc il est impératif d’utiliser des méthodes de
détection et d’analyses qui ciblent ce genre de molécules.
Ces résultats sont presque similaires aux résultats de Ilić et al., (2005) où ils ont tracé les
courbes d’absorbance de 9 souches suspectées être des Streptomycètes, et l’absorbance à 330
nm n’as pas dépassé le 0,7 alors que les pics ont été dans l’intervalle de [220-260] nm. Le
travail de Maleki & Mashinchian (2011) rapporte les mêmes résultats et l’absorbance ne
dépasse pas 0,4 et les pic sont dans l’intervalle [200-245] nm.
Il y’en a une grande différence entre les courbes en ce qui concerne le pic suivant la limite
du spectrophotomètre, TB1 a présenté une absorbance maximale de 1,580 alors que les deux
autres extraits ont une absorbance qui ne dépasse pas 0,3. En comparaison avec les résultats
obtenus lors du criblage secondaire, la courbe d’absorbance confirme la carence des extraits
de SM1B1 et SM2.2 en molécules.

Résultats et Discussion
73
8 Chromatographie sur couche mince
La chromatographie sur couche mince est faite sur une plaque de gel de silice, c’est-à-dire
une chromatographie d’adsorption, la séparation demande que l’adsorption d’un composé qui
est plus polaire que les autres composés.
Figure 59 : Visualisation de la CCM avec UV de l’extrait de la souche TB1 A : Visualisation à 254 nm, B : visualisation à 366 nm
B A

Résultats et Discussion
74
Le gel de silice est l’absorbant le plus connu et le moins cher, et il a été utilisé par pas mal
de chercheurs dans différentes chromatographies, mais pour une meilleure séparation il faut
connaitre auparavant la nature des molécules à extraire, pour choisir le solvant qui convient
selon (Cieśla & Waksmundzka-Hajnos 2009). Les travaux ciblant l’extraction et
l’identification des métabolites secondaires produites par les souches du genre Streptomyces
sont nombreux.
Dans le cas du gel de silice, les molécules polaires migrent lentement en comparaison avec
les non-polaires, c’est-à-dire que la séparation des composés est faite suivant la différence de
polarité entre les molécules présentes dans le mélange. Dans le gel de silice il y a un
indicateur (manganese-activated zinc-silicate) qui absorbe la lumière à 254 nm et donne une
fluorescence à ~520 nm (Tanvir et al., 2013 ; Tuzimski, 2011).
Le choix du chloroforme comme éluant (phase mobile) est basé sur la polarité élevée de
cet éluant, car les métabolites polaires demandent un développement dans une phase mobile
polaire. La séparation demande aussi un système éluant mono ou bien binaire, c’est-à-dire
l’utilisation d’un seul éluant ou bien un mélange de deux éluant en même temps (Gibbons,
2006).
La méthode de révélation par les rayons UV est non destructive par rapport à des autres
méthodes qui utilisent des réactifs corrosifs (Fuchs et al., 2011) ainsi que pas mal de
chercheurs qui ont utilisé les UV pour la détection des molécules présentes dans les extraits
(Arasu et al., 2009 ; Sajid et al., 2009). Le seul inconvénient de cette méthode c’est que il y’
en a des molécules qui n’absorbent pas la lumière à 254 ou à 366nm et elles ne seront pas
visible et demandent une détection par pulvérisation (Gibbons, 2006).
La CCM de l’extrait de la souche TB1, en comparaison avec le travail d’Arasu et al
(2009), où il a utilisé les mêmes solvants pour l’extraction et pour le développement de la
CCM, a donné les mêmes taches que ce dernier. Les flèches en orange indiquent la présence
des alcaloïdes, les flèches en jaune indiquent la présence des composés non connus et les
autres flèches indiquent les composés qui absorbent la lumière UV.
Il est très difficile de pouvoir identifier la nature exacte des molécules présentent dans la
plaque, certains composés ne sont pas visibles par UV et demandent une autre détection. Les
composés présents dans la plaque ne sont pas tous des molécules antibactériennes, ou par une
autre façon on ne peut jamais estimer qu’elles sont bioactives ou non ; alors il été préférable

Résultats et Discussion
75
de faire une bioautographie (Sajid et al., 2009) avant de déterminer le Rf de chaque molécule
active et pouvoir au moins estimer la nature ou bien la famille de cette molécule ou bien
toutes les molécules qui donnent un effet antibactérien lors de la bioautographie.
Figure 60 : Chromatographie des extraits des souches SM2.2 et SM1B1

Résultats et Discussion
76
On observe dans la figure 60 qu’il y’a qu’une seule tache, et elle est sombre, ça veut dire
qu’elle absorbe de la lumière UV mais on ne peut pas savoir sa nature exacte. Il sera
souhaitable dans les prochaines études de réaliser des autres méthodes de détection surtout par
pulvérisation afin de savoir au moins les fonctions que ces molécules les portent.
Pour une meilleure caractérisation des molécules présentes dans les extraits, plusieurs
méthodes et techniques récentes et plus poussées que celle utilisé dans ce travail, comme la
Chromatographie à phase gazeuse (CPG), la Chromatographie Liquide à Haute Pression
(HPLC) (Boudjella et al., 2006), Résonance Magnétique Nucléaire (Chu et al., 2001) …
etc ; il sera souhaitable de les utilisées pour bien améliorer les résultats de ce travail.
9 Électrophorèse des acides nucléiques
L’amplification a été réalisée sur le gène ADNr 16S des 3 souches sélectionnées (TB1,
SM2.2 et SM1B1), et la plupart des études qui ont pour but l’identification des souches non
connues ciblent le gène ADNr 16S.
Le séquençage de l’ADNr 16S selon Kämpfer (in Dworkin et al., 2006c), nous indique que
la souche peut être déterminée au niveau du genre et même au niveau de l’espèce. La taille du
fragment issu de l’amplification est de 1500 paires de bases (bp) ce qui est similaire aux
autres amplification faites d’autres auteurs (Valli et al., 2012 ; Valanarasu et al., 2010 ;
Boudjella et al., 2006 ; Ben Fguira et al., 2005)
M S1 S2 S3
1500 pb
Figure 61 : électrophorèse des acides nucléiques sur gel d’Agarose à 1.5% :d’un fragment de 1500 pb amplifié par les amorces universelles pour les souches SM1B1 (S1),
SM2.2 (S2) et TB1 (S3), les autres souches sont des souches du laboratoire M : Marqueur de taille

Résultats et Discussion
77
Comme le montre la figure (61) que la bande de 1500 pb de la souche TB1 (S3) qui est
visible, tandis que rien n’est visible pour les 2 autres souches (S1 et S2). Il peur être que
l’amplification chez les 2 souches n’a pas ciblé l’ADNr 16S, ou bien l’extraction de l’ADN
ou l’amplification du gène n’ont pas été achevées d’une façon correcte ; mais la suggestion la
plus probable, est la mauvaise extraction de l’ADN puisque le gène de l’ADNr 16S de la
souche TB1 a été bien amplifié.

Conclusion &
Perspectives

Conclusion & Perspectives
78
Conclusion et perspectives
Les sols agricoles, d’après cette étude sont riche en Streptomyces car ils
contiennent plus de matière organique, cette dernière qui est le milieu favorable de la
plupart des souches de ce genre; n’oublions pas leur richesse en enzymes de
dégradation des différents composés. L’isolement et l’identification ont été faits
suivant les critères décrites et recommandés par Shirling et Gottlieb (1966).
Le pourcentage des souches isolées présentant une activité antibactérienne sur au
moins une seule souche bactérienne pathogène utilisée dans cette étude été de 26 %
du nombre globale des isolats suspectés être des Streptomycètes qui est 85 isolats.
Parmi ces isolats, trois (3) ont présenté une activité antibactérienne assez satisfaisante
vis-à-vis presque toutes les souches pathogènes utilisées, les codes de ces isolats sont :
TB1, SM2.2 et SM1B1, ces derniers isolats ont été le sujet des tests physiologiques et
biochimiques faits dans ce travail.
La recherche des différents enzymes qui dégrade la caséine, l’amidon, l’urée et le
tween 80 a été achevée ; ainsi que l’utilisation des différents sucres par ces 3 isolats.
L’extraction des métabolites secondaires par solvant (chloroforme) et leur
concentration par le Rotavapor a facilité la pré-identification des molécules qui
compose les extraits bruts des isolats sélectionnées. La pré-identification a été faite
par une analyse de l’absorbance des extraits (absorbance UV-visible) et par une
chromatographie sur couche mince (CCM).
La dernière étape se résume dans l’identification par biologie moléculaire
(méthode PCR). L’extraction de l’ADN des isolats sélectionnés a été réalisée par un
kit, et l’amplification par le thermocycler a ciblé le gène ADNr 16S par l’utilisation
des amorces « primers » universels (8UA et 1492 R), à la fin une électrophorèse des
acides nucléiques a été réalisée sur gel d’agarose. Ces fragments d’ADN doivent être
séquencés et analysés par un logiciel (le BLAST) par rapport à une banque de
données afin de donner une classification fiable.

Conclusion & Perspectives
79
D’après les résultats obtenus il parait que le sol est toujours riche en
microorganismes et surtout les Actinomycètes et les Streptomycètes ; et la recherche
de nouvelles molécules bactéricides capables de lutter contre les infections
bactériennes est un volet de recherche illimité et on aura toujours besoin d’aller vers
la nature en tout cas afin d’avoir la guérison souhaitée.
Les souches isolées ont présentées une activité antibactérienne qui est satisfaisante,
mais il faut l’exploitée d’une manière scientifique juste et bien sûre suivre les étapes
indispensables afin d’arriver à une vrai exploitation qui apporte de plus à la
communauté et à la vie humaine.
Malheureusement la recherche de l’activité antibactérienne est devenue moins
attractive par rapport aux autres activités comme l’anti-tumorale, et ça ne veut pas
dire que les molécules qui ont une activité antibactérienne n’ont pas d’autres activités,
au contraire ils peuvent conférer des activités très intéressantes.
Parmi les perspectives qu’il faut tirer de cette étude :
◊ Exploiter les sols non encore explorés et surtout le sol agricole et celui qui
contient plus de débris organiques
◊ Il sera préférable d’utiliser plusieurs méthodes d’évaluation de l’effet
antibactérien des souches isolées comme la méthode du tri perpendiculaire
◊ Une approche complémentaire se concentre sur l’évaluation de l’activité
antifongique des souches de Streptomyces; ainsi que les autres rôles bioactives
suspectés être présentes chez les souches.
◊ Utiliser de divers solvants pour l’extraction des métabolites secondaires et même
utiliser autres méthodes d’extraction que celle utilisée dans cette étude.
◊ L’identification de la structure chimique de chaque constituant des extraits par
l’utilisation des méthodes plus poussées comme HPLC, RMN … etc.
◊ Concernant l’identification moléculaire, il sera souhaitable d’utiliser des amorces
spécifiques au genre ou bien à l’espèce, ainsi que la détection de la présence des gènes
d’intérêt.

Références bibliographiques

Références bibliographiques
80
References
1. Adinarayana K., Ellaiah P., Srinivasulu B., Bhavani Devi R. &Adinarayana G.
(2003) Response surface methodological approach to optimize the nutritional
parameters for neomycin production by Streptomyces marinensisunder solid-state
fermentation. Process Biochem. 38: P 1565-1572.
2. Aharonowitz Y & Friedrich CG (1980), Alanine dehydrogenase of lactam antibiotic
producer Streptomyces clavuligerus. Arch. Microbiol 125: P 137-142.
3. Alan T. Bull &StachJames E.M (2007), Marine actinobacteria: new opportunities for
natural product search and discovery, TRENDS in Microbiology Vol.15 No.11: P 491-
499, RoyaumeUnie.
4. Al-Askar A. A (2012), Microbiological studies on the in vitro inhibitory effect of
Streptomyces collinusalbescens against some phytopathogenic fungi, African Journal
of Microbiology Research Vol. 6(13) : P 3277-3283.
5. Anderson Annaliesa. S & Wellington Elizabeth M. H (2001), The taxonomy of
Streptomyces and related genera, International Journal of Systematic and Evolutionary
Microbiology 51 : P 797–814.
6. Anzai Kozue, OhnoMichiyo, Nakashima Takuji, KuwaharaNatsumi, Suzuki
Rieko, Tamura Tomohiko, Komaki Hisayuki, Miyadoh Shinji,
HarayamaShigeaki& Ando Katsuhiko (2008), Taxonomic distribution of
Streptomyces species capable of producing bioactive compounds among strains
preserve dat NITE/NBRC, ApplMicrobiolBiotechnol 80: P 287–295.
7. AouicheA, Sabaou N, Meklat A, Zitouni A, Mathieu F &Lebrihi A(2012), Activité
antimicrobienne de Streptomycessp.PAL111 d’origine saharienne contre
diversmicroorganismes cliniques et toxinogènes résistantsaux antibiotiques, Journal de
Mycologie Médicale 22 :P 42-51.
8. AraIsmet, BukhariNajat .A, Aref N. M, ShinwariMahera M. A &Bakir M. A
(2012), Antiviral activities of streptomycetes against tobacco mosaic virus (TMV) in
Datura plant: Evaluation of different organic compounds in their metabolites, African
Journal of Biotechnology Vol. 11(8): P 2130-2138.
9. ArasuMariadhasValan, VeeramuthuDuraipandiyan, SavarimuthuIgnacimuthu
(2013), Antibacterial and antifungal activities of polyketide metabolite from marine
Streptomyces sp. AP-123 and its cytotoxic effect, Chemosphere 90: P 479-487.

Références bibliographiques
81
10. ArasuValan M, Duraipandiyan V, Agastian P, Ignacimuthu S (2009), In vitro
antimicrobial activity of Streptomyces spp. ERI-3 isolated from Western Ghats rock
soil (India), Journal de MycologieMédicale19: P 22-28.
11. Atta M Houssam (2012), Biochemical studies on antibiotic production from
Streptomyces sp.: Taxonomy, fermentation, isolation and biological properties, article
in press, Journal of Saudi Chemical Society : P 01-11.
12. Awad M. Hassan, Kamel Y.I. El-Shaedet ABD El-Monaem M. El-Nakkadi
(2009), Isolation, Screening and Identification of Newly Isolated Soil Streptomyces
(Streptomyces sp. NCR-35) for β-Lactamase Inhibitor Production, World Applied
Sciences Journal 7 (5): P 637-646, Egypt.
13. Balasubramaniam V, Ganesh S, Karunanithi V, Perumal P (2011), Improved
Culturing, Screening and Fermentation of Soil Actinomycetes for Antimicrobial
Agents, Int. J.Pharm& Ind. ResVol (01) Issue (02): P 153-159.
14. Ben Fguira Lilia Fourati, Bejar Samir &MellouliLotfi (2012), Isolation and
screening of Streptomyces from soil ofTunisian oases ecosystem for non-polyenic
antifungalmetabolites, African Journal of Biotechnology Vol. 11(29): P 7512-7519.
15. Bendich Arnold J &Drlica Karl (2004), “Chromosome, Bacterial”, The Desk
Encyclopedia of Microbiology, Elsevier Academic Press, P 259-270.
16. Bhatnagar Ira & Kim Se-Kwon (2010), Marine Antitumor Drugs: Status, Shortfalls
and Strategies, Review, Mar. Drugs, 8:P 2702-2720.
17. Bibb Mervyn and Hesketh Andrew (2009), Analyzing the Regulation of Antibiotic
Production in Streptomycetes, in David A. Hopwood, editor, Methods in Enzymology,
Vol. 458, Burlington: Academic Press: P 93-116.
18. Boopathy N. Sithranga&Kathiresan. K (2010), Anticancer Drugs fromMarine
Flora: An Overview, Journal of Oncology Volume 2010, Article ID 214186: P 1-18.
19. BoudemaghAllaoueddine (2007), Isolement, à partir des sols Sahariens, de bactéries
actinomycétales productrices de molécules antifongiques, identification moléculaire
de souches actives, Thèse de Doctorat d’Etat : P 1-132, Université Mentouri
Constantine, Algerie.
20. BoudjellaHadjira, BoutiKarima, ZitouniAbdelghani, Mathieu Florence, Lebrihi
Ahmed, SabaouNasserdine (2006), Taxonomy and chemical characterization of
antibiotics of StreptosporangiumSg 10 isolated from a Saharan soil, Microbiological
Research161 : P 288—298.

Références bibliographiques
82
21. Boughachiche F, Reghioa S, Oulmi L, Erizer H, Kitouni M, Boudemagh A,
Boulahrouf A (2008), isolement d’actinomycetales productrices de substances
antimicrobiennes à partir de la sebkha de Ain Mlila, Sciences &Technologie C –
N°23 :P 5-10.
22. BoughachicheFaiza (2012), Étude de molécules antibiotiques secrétées par des
souches appartenant au genre Streptomyces, isolées de Sebkha, thèse de Doctorat,
UniversitéMentouri- Constantine, Algerie :P 1-141.
23. BourasNoureddine, MeklatAtika, ToumatiaOmrane, MokraneSalim, Holtz D
Michael, Strelkov E Stephen &SabaouNasserdine (2013), Bioactive potential of a
new strain of Streptomyces sp.PP14 isolated from Canadian soil, African Journal of
Microbiology Research, Vol 7 (25): P 3199-3208.
24. BoussaberElarbi, El IdrissiSidiBrahim Salem, MeftahKadmiriIssam,
HilaliLahoucine, and HilaliAbderraouf (2013), A molecular approach to the
identification of actinomycetes producing antimicrobialactivities strains by sequencing
of the 16S rDNA, International Journal of Innovation and Applied Studies, Vol. 3 No.
4: P 1053-1065.
25. BoussaberElarbi, MeftahIssamKadmiri, HilaliLahoucine, HilaliAbderraouf
(2012), Comparaison de l’activité antimicrobienne des souches d’actinomycètes
isolées de milieu variés, Science Lib Editions Mersenne : Volume 4, N ° 121203: P
01-21.
26. Brun Yves V (2004), The Desk Encyclopedia of Microbiology “Developmental
processes in bacteria”, Elsevier Academic Press: P 314-325.
27. Cattoir. V, Daurel. C (2010), Quelles nouveautés en antibiothérapie ? « Update on
antimicrobialchemotherapy », Médecine et maladies infectieuses 40 : P 135–154.
28. CeylanOzgur, GultenOkmen et AyselUgur (2008), Isolation of soil Streptomyces as
source antibiotics active against antibiotic-resistant bacteria, EurAsia J BioSci 2: P 73-
82.
29. Chater F. Keith, Biro Sandor, Lee KyeJoon, Palmer Tracy & Schrempf
Hildgund (2010), The complex extracellular biology of Streptomyces, FEMS
Microbiol Rev 34: P 171–198.
30. Chater K. E& D. A. Hopwood (1973), Differentiation in actinomycetes. In: J. M.
Ashworth and J. E. Smith (Eds.) Microbial Differentiation. Cambridge University
Press. London: P 143–160.

Références bibliographiques
83
31. Choma Irena M &GrzelakEdyta M (2011), Bioautography detection in thin-layer
chromatography, Journal of Chromatography A, 1218 : P 2684–2691.
32. ChroňákovaAlica, KrištůfekVáclav, Tichký Martin, Dana Elhottová (2010),
Biodiversity of streptomycetes isolated from a succession sequence at apost-mining
site and their evidence in Miocene lacustrine sediment, Microbiological Research 165:
P 594-608.
33. Chu Min, Mierzwa Ronald, He Ling, Xu Ling, Gentile Frank, Terracciano
Joseph, Patel Mahesh, Miesel Lynn, Bohanon Samantha, Kravec Cynthia,
Cramer Christina, Fischman Thierry O, Hruza Alan, RamanathanLata,
ShipkovaPetia& Chan Tze-Ming (2001), Isolation and structure elucidation of two
novel deformylase inhibitors produced by Streptomyces sp.Tetrahedron Letters 42, P :
3549–3551.
34. CieślaŁukasz, Waksmundzka-Hajnos Monika (2009), Two-dimensional thin-layer
chromatography in the analysis of secondary plant metabolites, Journal of
Chromatography A, 1216: P 1035–1052.
35. Colombié Vincent (2005), Description de la production de spiramycine par
Streptomycesambofaciens. Modélisation métabolique, simulation et capteur logiciel,
mémoire Doctorat, L’Institut Nationale des Sciences Appliquées de Toulouse : P 1-
163, France.
36. Das Surajit, Ward Louise R & Chris Burke (2010), Screening of marine
Streptomyces spp. for potential use as probiotics in aquaculture, Aquaculture 305: P
32–41.
37. Dawn R. D. Bignell, José C. Huguet-Tapia, Madhumita V. Joshi, Gregg S. Pettis,
Rosemary Loria (2010), What does it take to be a plant pathogen: genomic insights
from Streptomyces species, Antonie van Leeuwenhoek 98: P 179–194.
38. DehnadAlireza, ParsaLalle, BakhshiRouhollah,
SoofianiSamadAbdi&MokhtarzadehAhad (2010), Investigation antibacterial
activity of Streptomycetesisolates from soil samples, West of Iran, African Journal of
Microbiology Research Vol. 4 (14): P 1542-1549.
39. Delaunay S, Rondags E et Germain P (2003), production d’antibiotiques par
biotechnologies, techniques de l’ingénieur. Opérations unitaires, génie de la réaction
chimique J6 008 : P 1-12.

Références bibliographiques
84
40. Deshpande B. S, Ambedkar S. S &Shewale J. G (1988), Biologically active
secondary metabolites from Streptomyces, Enzyme Microb. Technol, 1988, vol. 10: P
455-473.
41. Desjardin Valérie (2002), Réduction du chrome (VI) par la souche
Streptomycesthermocarboxydus NH50 isolée à partir d’un sol pollué, Thèse Doctorat,
L’institut national des sciences appliquées de Lyon, France, P 1-236.
42. DhanasekaranaDharumadurai,
ThajuddinaNooruddin&PanneerselvamAnnamalai (2010), Herbicidal agents from
actinomycetes against selected crop plants and weeds, Taylor & Francis, Natural
Product Research, Vol. 24, No. 6: P 521–529.
43. DjaballahChamsseddine, Biodiversité des Actinomycètes Halophiles et Haltolerants
Isolés de la Sabkha de Ain M’lila, Mémoire de Magister, Université de Mentouri,
Constantine, Algérie, P 1-73, 2010.
44. Donadio Stefano, Monciardini Paolo, Alduina Rosa, Paola Mazza, Claudia
Chiocchini, Linda Cavaletti, Margherita Sosio& Anna Maria Puglia (2002),
Journal of Biotechnology 99 : P 187-198.
45. Donadio Stefano, Paolo Monciardini, Rosa Alduina, Paola Mazza, Claudia
Chiocchini, Linda Cavaletti, Margherita Sosio, Anna Maria Puglia (2002),
Microbial technologies for the discovery of novel bioactive metabolites, Journal of
Biotechnology99 : P 187-198, Italie.
46. Dworkin Martin, Falkow Stanly, Rosenberg Eugene, Schleifer Karl-Henz et
StackebrandyErko, The Prokaryotes, 3ème edition, Volume 3, A Handbook on the
Biology of Bacteria : Archea. Bacteria :Firmicutes, Actinomycetes, edition Springer,
USA. (c)
47. Dworkin Martin, Falkow Stanly, Rosenberg Eugene, Schleifer Karl-Henz et
StackebrandyErko, The Prokaryotes, 3ème edition, Volume 1, A Handbook on the
Biology of Bacteria : Archea. Bacteria :Firmicutes, Actinomycetes, edition Springer,
USA. (a)
48. Dworkin Martin, Falkow Stanly, Rosenberg Eugene, Schleifer Karl-Henz et
StackebrandyErko, The Prokaryotes, 3ème edition, Volume 2, A Handbook on the
Biology of Bacteria : Archea. Bacteria :Firmicutes, Actinomycetes, edition Springer,
USA. (b)
49. Dyson Paul (2011), Streptomyces Molecular biology and Biotechnology, Caister
Academic Press : P 1-260, Royaume-Uni.

Références bibliographiques
85
50. E Hamid Mohamed (2011), Variable antibiotic susceptibility patterns among
Streptomyces species causing actinomycetoma in man and animals, Annals of Clinical
Microbiology and Antimicrobials, 10:24.
51. Elibol Murat &MavitunaFerda (1998), Effect of sucrose on actinorhodin production
by Streptomyces coelicolor A3(2), Process Biochemistry Vol. 33, No. 3: P 307-311.
52. El-Nakeeb, M. A. & H. A. Lechevalier (1963).Selective isolation of aerobic
actinomycetes. Appl. Microbiol. 11: P 75–77.
53. Emerson Rudi de Lima Procópioa, Reis Ingrid da Silvaa, Kassawara Mayra
Martinsa, LúcioJoão de Azevedoa&MagaliJanete de Araújo (2012), Antibiotics
produced by Streptomyces, Braz J Infect Dis 16(5), P 466–471.
54. Fischer G, Kyriacou A, Decaris B, Lebiond P (1997), Genetic instabi|ity and its
possible evolutionary implications on the chromosomal structure of Streptomyces,
Sociétéfrançaise de biochimie et biologiemoléculaire, Biochimie 79: P 555-558,
Elsevier, Paris, France.
55. FlaviaMarinelli (2009), Antibiotics andStreptomyces: the future of antibiotic
discovery, microbiology today, feb 09: P 20-23.
56. Fuchs Beate, SußRosmarie, Teuber Kristin, Eibisch Mandy & Schiller Jurgen
(2011),Lipidanalysis by thin-layer chromatography - A review of the current state,
Journal of Chromatography A, 1218 : P 2754–2774.
57. Garrity George M, Brenner Don J, Krieg Noel R & Staley James T (2005),
Bergey’s Manual of Systematic Bacteriology, Deuxième Edition, Volume Deux The
Proteobacteria, Partie A : Introductory Assays: P 1-304, Springer Edition, ÉtatsUnis.
58. Geoffrey E. Dixon & Emma L. Tilston, Soil Microbiology and Sustainable Crop
Production: P 1-45,edition Springer, Londres, Grand Bretagne, 2010.
59. GeshevaV., Ivanova V &Gesheva R (2005),Effects of nutrients on the production of
AK-111-81 macrolide antibiotic by Streptomyces hygroscopicus. Microbiol. Res. 160:
P 243-248.
60. Gibbons Simon (2006), An Introduction to PlanarChromatography, Methods in
Biotechnology, Vol. 20, Natural Products Isolation, 2nd ed : P 77-116.
61. Goodfeilow Michael, Elayne V. Ferguson & Jean-Jacques Sanglier (1992),
Numerical classification and identification of Streptomyces species - a review,
Elsevier Science Publishers, Gene, 115 P 225-233, RoyaumeUnis.
62. Groth Ingrid, Saiz-Jimenez C (1999), Actinomycetes in Hypogean Environments,
Geomicrobiology Journal, 16-1: P 1-8.

Références bibliographiques
86
63. Gust Bertolt, Chandra Govind, JakimowiczDagmara, YuqingTian, Bruton Celia
J &Chater Keith F (2004), λ Red-Mediated Genetic Manipulation of Antibiotic-
Producing Streptomyces, Advances in Applied Microbiology, Volume 54: P 107-124.
64. Hakvåg Sigrid, FjærvikEspen, JosefsenKjell D, Ian Elena, EllingsenTrond E
&Zotchev Sergey B (2008), Characterization of Streptomyces spp. Isolated from the
Sea Surface Microlayer in the Trondheim Fjord, Norway, Mar. Drugs 2008, 6 : P 620-
635.
65. HolgerJenke-Kodama, Rolf Müller et ElkeDittmann (2008), Evolutionary
mechanisms underlying secondary metabolite diversity, Progress in Drug Research,
Vol. 65 : P 120-140.
66. Hopwood D.A, Bibb M. J, Chater K.F,Kieser T, Bruton C.J, Kieser H.M, Lydiate
D.J, Smith C. P, Ward J.M&Schremph H (1985) Genetic Manipulation of
Streptomyces: A Laboratory Manual. Norwich, UK: John Innes Foundation.
67. Hopwood David (2006), Soil To Genomics: The Streptomyces Chromosome, Annu.
Rev. Genet.40 : P 1-23.
68. IlićSlavica B., Sandra S. Konstantinović, Zoran B. Todorović (2005), UV/VIS
analysis and antimicrobial activity of Streptomyces isolates Medicine and Biology
Vol.12, No 1: P 44 – 46.
69. Janeiček J, Tichy P, Spižek J et Z. Vanĕk (1997), Constitution of the Metabolic
Type of Streptomycetes during the First Hours of Cultivation, Folia Mierobiol. 42 (2):
P 75-96.
70. Jeffrey, L. S. H (2008), Isolation, characterization and identification ofactinomycetes
from agriculture soils at Semongok,Sarawak, African Journal of Biotechnology Vol. 7
(20):P 3697-3702.
71. JonsbuEinar, McIntyre Mhairi& Nielsen Jens (2002), The influence of carbon
sources and morphology on nystatin production by Streptomyces noursei, Journal of
Biotechnology 95, P 133–144.
72. Kämpfer Peter (2006), The Family Streptomycetaceae, Part I: Taxonomy,
Prokaryotes 3: P 538–604.
73. KarlovskyPetr (2008), Secondary Metabolites in Soil Ecology. Soil Biology 14 : P
01-19.
74. Karthik L, Kumar Gaura V, BhaskaraRao K. V (2010), Diversity of marine
actinomycetes from nicobar marine sediments and its antifungal activity, International
Journal of Pharmacy and Pharmaceutical Sciences, Vol 2, Issue 1.

Références bibliographiques
87
75. KavithaAlapati, PrabhakarPeddikotla,
VijayalakshmiMuvva&VenkateswarluYenamandra(2010), Purification and
biological evaluation of the metabolites produced by Streptomyces sp. TK-VL_333,
Research in Microbiology 161 : P 335-345.
76. Kelemen Gabriella H &Buttnert Mark J (1998), Initiation of aerial mycelium
formation in Streptomyces, Current Opinion in Microbiology 1: P 656-662.
77. Khattabi A (2005), Isolement et étude taxonomique de deux souches d’actinomycètes
d’origine marine productrices de substances antifongiques. Thèse de doctorat :
Université ChouaibDoukkali. Faculté des sciences, Maroc.
78. Kieser, T., M. J. Bibb, M. J. Buttner, K. F. Chater, and D. A. Hopwood (2000).
Practical Streptomyces Genetics : P 1-613, The John Innes Foundation. Norwich, UK.
79. Kitouni Mahmoud (2007), Isolement de Bactéries Actinomycétales productrices
d’Antibiotiques à Partir d’Ecosystèmes Extrêmes. Identification Moléculaires des
Souches Actives et Caractérisation des Substances Elaborées, thèse Doctorat,
Université de Mentouri, Constantine, Algérie : P 1-176.
80. Komatsu Mamoru, Uchiyama Takuma, Omura Satoshi, Canec David E
&IkedaaHaruo (2009), Genome-minimized Streptomyces host for the heterologous
expression of secondary metabolism, PNAS vol. 107 no. 6 : P 2646–2651.
81. Kozue Anzai, MichiyoOhno, Takuji Nakashima, NatsumiKuwahara, Rieko
Suzuki, Tomohiko Tamura, Hisayuki Komaki, Shinji Miyadoh,
ShigeakiHarayama& Katsuhiko Ando (2008), Taxonomic distribution of
Streptomyces species capable of producing bioactive compounds among strains
preserved at NITE/NBRC, ApplMicrobiolBiotechnol 80: P 287–295.
82. Kulkarni-Almeida Asha A, Brahma Manoja K, PadmanabhanPrabhu, Mishra
Prabhu D, ParabRajashri R, GaikwadNitin V, ThakkarChandni S,
TokdarPradipta, RanadivePrafull V, Nair Amrutha S, DamreAnagha A,
BahiratUmakant A, DeshmukhNitin J, DoshiLalit S, Dixit Amol V, George Saji
D, Vishwakarma Ram A, Nemmani Kumar VS and MahajanGirish B (2011),
Fermentation, Isolation, Structure, and antidiabetic activity of NFAT-133 produced by
Streptomyces strain PM0324667, AMB Express 1:42 : P 1-12.
83. Lancini Giancarlo &Demain L Arnold (2013), Bacterial Pharmaceutical Products,
The Prokaryotes – Applied Bacteriology and Biotechnology, Springer-Verlag Berlin
Heidelberg : P 557-580.

Références bibliographiques
88
84. Lanoot Benjamin, Vancanneyt Marc, Hostea Bart, VandemeulebroeckeKatrien,
Cnockaert Margo C, Dawyndt Peter, Liu Zhiheng, Huang Ying, Swings Jean
(2005),Research in Microbiology 156: P 755–762.
85. Lazzarini A, Cavaletti L, Toppo G, Marinelli F (2001), Potentialities of rare
actinomycetes as producers of new antibiotics. Ant v Leeuwenhoek 79: P 399–407
86. Liang Jingle, XuaZhinan, Liu Tongfeng, Jianping Lin, Cen Peilin(2008), Effects
of cultivation conditions on the production of natamycin with Streptomyces
gilvosporeus LK-196, Enzyme and Microbial Technology 42 (2008) 145–150.
87. Liras Paloma& Martín Juan F (2004), Assay Methods for Detection and
Quantification of Antimicrobial Metabolites ProducedbyStreptomyces clavuligerus,
Methods in Biotechnology, Vol. 18: Microbial Processes and Products,
HumanaPressInc: P 149-163.
88. Loria R, Bignell DR, Moll S, Huguet-Tapia JC, Joshi MV, Johnson EG, Seipke
RF & Gibson DM (2008),Thaxtomin biosynthesis: the path to plant pathogenicity in
the genus Streptomyces. Antonie van Leeuwenhoek 94, P 3–10.
89. Loria Rosemary, Coombs Justin, Yoshida Masahiro, Kers Johan,
BukhalidRaghida (2003), A paucity of bacterial root diseases: Streptomyces
succeeds where others fail, Physiological and Molecular Plant Pathology 62: P 65–72.
90. Madigan M. T., Martinko J.M. & Parker J (1997), Biology of microorganisms.
Printice, Hall International Inc, New York: P 146-147.
91. MalekiHadi&OmidMashinchian (2011), Characterization of Streptomyces
Isolateswith UV, FTIR Spectroscopy and HPLC Analyses, BioImpacts, 2011, 1(1) : P
47-52.
92. Manteca Angel & Sanchez Jesus(2009), Streptomyces Development in Colonies and
Soils, Applied and Environmental Microbiology, Vol 75, N° 9: P 2920–2924.
93. Maplestone Rachael A, Martin J. Stone et Dudley H. Williams (1992), The
evolutionary role of secondary metabolites - a review, Elsevier Science Publishers,
Gene, 115: P 151-157, RoyaumeUnis.
94. Martin Patricia, Dary Annie, André Axelle&Decaris Bernard (2000),
Identification and typing of Streptomyces strains: evaluation of interspecific,
intrspecific and intraclonal differences by RAPD fingerprinting, Res. Microbiol. 151 :
P 853–864.
95. McCormick Joseph. R &FlärdhKlas (2012), Signals and Regulators that Govern
Streptomyces Development, FEMS Microbiol Rev. 2012 January; 36(1): P 206–231.

Références bibliographiques
89
96. MedemaMarnix H, Trefzer Axel, KovalchukAndriy, Marco van den Berg, Müller
Ulrike, Heijne Wilbert, Wu Liang, Alam Mohammad T, Catherine Ronning M,
Nierman William C, BovenbergRoel A. L, Breitling Rainer & Eriko Takano,
(2010), The Sequence of a 1.8-Mb Bacterial Linear Plasmid Reveals a Rich
Evolutionary Reservoir of Secondary Metabolic Pathways, Genome Biol. Evol. 2: P
212–224.
97. Mellouli L., Karray-Rebai I., Sioud S., Ben Ameur-Mehdi R., Naili B., Bejar S.
(2004). Efficient transformation procedure of a newly isolated Streptomyces sp. TN58
strainproducingantibacterialactivities. CurrentMicrobiology. 49: P 400-403.
98. Min J. Kang, Janice L. Strap et Donald L. Crawford (2010), Isolation and
characterization of potent antifungal strains of the Streptomyces violaceusniger clade
active against Candida albicans, J IndMicrobiolBiotechnol 37: P 35-4.
99. Muharram Magdy (2010),Identification of Aerobic Actinomycetes Species Based on
HSP-65 Gene Polymorphism Analysis by PCR-RFLP, Journal of Applied Sciences
Research, 6(6): P 609-615.
100. Mythili B et Ayyappa Das (2011), Studies on Antimicrobial Activity of
Streptomyces spp. Isolates from Tea Plantation Soil, Research Journal of Agricultural
Sciences 2(1): P 104-106,.
101. Nonoh James Oluoch, Lwande Wilber, Masiga Daniel, Herrmann Rafi, Presnail
James K, Schepers Eric, Okech Matilda Angela, Bagine Richard, Mungai Paul,
NyendeAggrey Bernard &BogaHamadiIddi (2010), Isolation and characterization
of Streptomyces species with antifungal activity from selected national parks in
Kenya, African Journal of Microbiology Research Vol. 4(9): P 856-864.
102. Olano Carlos, Méndez Carmen et José A. Salas (2009), Antitumor Compounds
from Marine Actinomycetes, Mar. Drugs 7 : P 210-248.
103. Olmos. E, Mehmood N, Haj Husein L, Goergen J-L, Fick M et Delaunay S
(2013), Effects of bioreactor hydrodynamics on the physiology of Streptomyces,
Bioprocess BiosystEng 36: P 259–272.
104. Olov Sterner (2012), Isolation of Microbial Natural Products, Natural Products
Isolation, Methods in Molecular Biology, vol. 864: P 393-413.
105. OskayMustafa (2009), Comparison of Streptomyces diversity betweenagricultural
and non-agricultural soils by using variousculture media, Scientific Research and
Essay Vol.4 (10): P 997-1005.

Références bibliographiques
90
106. Peterson E.A. (1954), A study of cross antagonisms among some
actinomycetesactiveagainstStreptomyces scabies and
Helminthsporiumsativum,AntibiotChemother. 4: P 145-149.
107. Pimentel-Elardo Sheila Marie, KozytskaSvitlana, Bugni Tim S, Ireland Chris
M, Moll Heidrun et Hentschel (2010), Anti-Parasitic Compounds from Streptomyces
sp. Strains Isolated from Mediterranean Sponges, Mar. Drugs 8: P 373-380.
108. Poole Colin F (2003), T hin-layer chromatography: challenges and opportunities,
Journal of Chromatography A, 1000: P 963–984.
109. Pradeau Dominique & Dauphin Chantal (2002), Chromatographie planaire :
applications, Éditions Techniques de l’Ingénieur 1476 : P 01-10.
110. Pridham Thomas. G (1965), Color and Streptomycetes “Report of an International
Workshop on Determination of Color of Streptomycetes”, Applied Microbiology, Vol
13, N° 1: P 43-61.
111. Pridham. T. G, Hesseltine. C. W et Benedict. R. G (1957), A Guide for the
Classification of Streptomycetes According to Selected Groups “Placement of Strains
in Morphological Sections” meeting of the Society of American Bacteriologists held at
Detroit, Michigan 28 avril-2 Mai, EtatsUnis: P 52-79.
112. PulsawatNattika, Kitani Shigeru, Nihira Takuya (2007), Characterization of
biosynthetic gene cluster for the production of virginiamycin M, a streptogramin type
A antibiotic in Streptomyces virginiae, Gene 393: P 31–42.
113. Rintala H,Nevalainen A, Rönkä E & M. Suutari (2001), PCR primers targeting
the 16S rRNA gene for the specificdetection of streptomycetes, Molecular and
Cellular Probes 15 :P 337–347.
114. Rintala Helena (2003),Streptomycetes in Indoor Environments PCR Based
Detection and Diversity, Kuopio University Printing Office, Kuopio:P 14-26, Finland.
115. Routien John B (1961), distribution of Streptomycetesthatproduceantibiotics,
MedicalResearchLaboratories, Chas. Pfizer & Co., Inc., Groton, Connecticut Vol 81:
P 218-225.
116. Saffroy Sophie (2006), Etude du Métabolisme Carboné chez
Streptomycespristinaespiralis, Thèse Doctorat, Institut Polytechnique de Lorraine,
France, P 6-14.
117. ahinNurretin&U urAysel (2003), Investigation of the Antimicrobial Activity of
Some Streptomyces Isolates, Turk J Biol 27: P 79-84.

Références bibliographiques
91
118. Sajid Imran, Blandine Clarisse, FondjaFotso Yao, ShaabanKhaledAttia,
HasnainShahida et LaatschHartmut(2009), Antifungal and antibacterial activities
of indigenous Streptomycesisolates from saline farmlands: prescreening,
ribotypingand metabolic diversity, World J MicrobiolBiotechnol 25: P 601–610.
119. Scherr Nicole & Nguyen Liem (2009), Mycobacterium versus
Streptomyces—we are different, we are the same, Current Opinion in Microbiology
12: P 699–707.
120. Seidel Véronique (2006), Initial and Bulk Extraction, Methods in Biotechnology,
Vol. 20, Natural Products Isolation, 2nd ed.
121. Shepherd Micah D, KharelMadan K, Bosserman Mary A &Rohr Jürgen (2010),
Laboratory Maintenance of Streptomyces species, CurrProtocMicrobiol. 2010 August;
CHAPTER: Unit–10E.1: P 01-10.
122. Shirling E. B & Gottlieb D (1966), Methods for Characterization of
Streptomyces Species, International Journal of Systematic Bacteriology, Vol. 16, No.
3 : P 313-340.
123. Sierra G., (1957) A Simple method for the detection of lipolytic activity of
microorganisms and some observations on the influence of the contact between cells
and fatty substrates. Antonie Van Leeuwenhoek, 23: P 15-22.
124. Singh L. Shantikumar, BaruahIndra, Bora T.C (2006), Actinomycetes of
LoktakHabitat: Isolation and Screening for Antimicrobial Activities, Biotechnology 5
(2), Asian Network for Scientific Information, Inde: P 217-221.
125. SinghSheo B &Pelaez Fernando (2008), “Biodiversity, chemical diversity and drug
discovery”,Progress in Drug Research, Vol. 65 : P 142-172.
126. SmaouiSlim (2010), Purification et Caractérisation de Biomolécules à partir de
microorganismes nouvellement isolés et identifiés, thèse de doctorat, Université de
Toulouse : 1-187, France.
127. Smillie Chris, Garcillán-Barcia M. Pilar, Francia M. Victoria, Rocha Eduardo
P. C, & Fernando de la Cruz (2010), Mobility of Plasmids, Microbiology AND
MolecularBiologyReviews : P 434–452.
128. SolankiRenu, KhannaMonisha&Lal Rup (2008), Bioactive compounds from
marine actinomycetes, Indian J. Microbiol48: P 410–431.
129. Sujatha P, Bapi-Raju K. V. V. S. N. &Ramana T. (2005) Studies on a new marine
streptomycete BT-408 producing polyketide antibiotic SBR-22 effective against
methicillin resistant Staphylococcus aureus. Microbiol. Res. 160: P 119-126.

Références bibliographiques
92
130. Suneetha V, Raj Karthick&Prathusha .K (2011), Isolation and identification of
Streptomyces ST1 andST2 strains from Tsunami affected soils: Morphologicaland
biochemical studies, Journal of Oceanography and Marine Science Vol. 2(4): P 96-
101.
131. SuutariMerja, Lignell Ulla, Hyvärinen Anne, NevalainenAino (2002), Media for
cultivation of indoor streptomycetes, Journal of Microbiological Methods 51: P 411–
416.
132. TaddeiAntonieta , Rodríguez María José, Márquez-Vilchez Ernesto, Castelli
Cristina (2006), Isolation and identification of Streptomycesspp.from Venezuelan
soils: Morphological andbiochemical studies. I.Microbiological Research 161: P 222-
231.
133. Tagu Denis (1999), Principes des techniques de biologie moléculaire, Institut
nationale de la recherche agronomique, Paris, France: P 1-131.
134. TanvirRabia, Sajid Imran &HasnainShahida (2013), Screening of
endophyticStreptomycetes isolated from Parthenlumhysterophorus L. against
nosocomial pathogens, Pak. J. Pharm. Sei., Vol.26, No.2: P 277-283.
135. Tara Devi Gurung, Chringma Sherpa, Vishwanath Prasad
Agrawal&BinodLekhak (2009), Isolation and Characterization of Antibacterial
Actinomycetes from Soil Samples of Kalapatthar, Mount Everest Region, Nepal
Journal of Science and Technology 10: P 173-182, Népal.
136. Thakur D, Bora T.C, Bordoloi G.N, Mazumdar S (2009), Influence of nutrition
and culturing conditions for optimum growth and antimicrobial metabolite production
by Streptomyces sp. 201, Journal de MycologieMédicale (2009) 19: P 161-167.
137. Thakur D, Yadav A, Gogoi B.K, Bora T.C (2007), Isolation and screening of
Streptomyces in soil of protected forest areas from the states of Assam and Tripura,
India, for antimicrobial metabolites, Journal de MycologieMédicale 17 : P 242-249.
138. Titgemeier, F., J. Walkenhorst, J. Reizer, M. H. Stuiver, X. Ciu, and M. H.
Saier. (1995). Identification and characterization of phosphoenolpyruvate: fructose
phosphotransferase system in three Streptomyces species. Microbiology 141: P 51–58.
139. Titgemeyer F, Walkenhorst J, Reiser J, Stuvier MH, Cui X, Saier MH (1995),
Identification and characterization of
phosphoenolpyruvate:fructosephosphotransferase systems in Streptomyces species,
Microbiol., 141: P 51-58.

Références bibliographiques
93
140. TomaszTuzimski (2011), Application of different modes of thin-layer
chromatography and massspectrometry for the separation and detection of large and
small biomolecules, Review, Journal of Chromatography A, 1218 : P 8799-8812.
141. ValanarasuM, KannanP, EzhilvendanS, GanesanG,IgnacimuthuS &Agastian P
(2010), Antifungal and antifeedant activities of extracellularproduct of Streptomyces
spp. ERI-04 isolated fromWestern Ghats of Tamil Nadu, Journal de
MycologieMédicale20 : P 290—297.
142. Valli S, SugasiniSuvathi S, Aysha OS, Nirmala P, Kumar Vinoth P, Reena A
(2012), Antimicrobial potential of Actinomycetes species isolated from
marineenvironment, Asian Pacific Journal of Tropical Biomedicine : P 469-473.
143. WatveMilind G, TickooRashmi, Jog Maithili M &BhalachandraBhole D (2001),
How many antibiotics are produced by the genus Streptomyces?, Arch Microbiol 176
:P 386–390.
144. Weber T, Welzel. K, Pelzer. S, Vente. A &Wohlleben. W (2003), Exploiting the
genetic potential of polyketide producing streptomycetes, Journal of Biotechnology
106: P 221–232, Allemagne.
145. Wohlleben Wolfgang, Mast Yvonne, MuthGünther, Röttgen Marlene,
StegmannEvi, Weber Tilmann (2012), Synthetic Biology of secondary metabolite
biosynthesis in actinomycetes: Engineering precursor supply as a way to optimize
antibiotic production: Review, FEBS Letters 586: P 2171–2176.
146. Yamaç Mustafa, IşikKamil et ŞahinNevzat (2011), Numerical classification of
streptomycetes isolated from karstic caves in Turkey, Turk J Biol, 35:P 473-484.
147. YikmisMeral&Steinbüchel Alexander (2012), Historical and Recent Achievements
in the Field of Microbial Degradation of Natural and Synthetic Rubber, Applied and
Environmental Microbiology Vol 78 N° 13: P 4543–4551.
148. Zähner H (1979), What Are Secondary Metabolites?Folia Microbiol. 24 : P 435-
443.
149. Zhou Zhan, GuJianying, Li Yong-Quan, Wang Yufeng (2012), Genome plasticity
and systems evolution in Streptomyces, BMC Bioinformatics 13 (Suppl 10): P 1-17.
150. Zhou Zhan, JianyingGu, Yi-Ling Du, Yong-Quan Li and Yufeng Wang (2011),
The -omics Era- Toward a Systems-Level Understanding of Streptomyces, Current
Genomics 12: P 404-416.

Annexes

Annexes
95
Annexes
Milieux de culture
Bennett :
D-Glucose anhydre 10 g
Casaminoacides 2 g
Extrait de levure 1 g
Extrait de viande 1 g
Eau distillée 1000 ml
Agar 15 g
pH = 7,3
ISP 2 :
Extrait de levure 4 g
Extrait de malt 10 g
Glucose 4 g
Eau distillée 1000 ml
Agar 20 g
pH = 7,3
ISP 3 :
Solution d’avoine 1000 ml
Solution d’oligo-éléments 1 ml
Agar 20 g
pH = 7,2

Annexes
96
ISP 4 :
Amidon soluble 10 g
K2HPO4 1 g
MgSO47H2O 1 g
NaCl 1 g
(NH4)2SO4 2 g
CaCO3 2 g
Solution d’oligo-éléments 1 ml
Eau distillée 1000 ml
Agar 20 g
pH = 7,0-7,4
ISP 5 :
Glycérol 10 g
L-Asparagine 1 g
Solution d’oligo-éléments 1 ml
Eau distillée 1000 ml
Agar 20 g
pH = 7,0-7,4
Muller Hinton :
Extrait de viande 2 g
Hydrolysat acide de caséine 17,5 g
Amidon 1,5 ml
Eau distillée 1000 ml
Agar 20 g
pH = 7,4

Annexes
97
Solution d’oligo-éléments :
FeSO4 7H2O 0,1 g
MnCl2 4H2O 0,1 g
ZnSO4 7H2O 0,1 g
Eau distillée 100 ml
Échelle McFarland :
Volume par mL
Standard 1% BaCL2 1% H2SO4 108 UFC
représenté 0.5 0.5 99.5 1.5 1 1.0 99.0 3 2 2.0 98.0 6 3 3.0 97.0 9 4 4.0 96.0 12 5 5.0 95.0 15 6 6.0 94.0 18 7 7.0 93.0 21 8 8.0 92.0 24 9 9.0 91.0 27
10 10.0 90.0 30

Annexes
98
Dendrogramme simplifié présentant les groupes taxonomiques chez le genre Streptomyces utilisant la taxonomie numérique (Anderson & Wellington, 2001) :

Résumé :
La présence des Streptomycètes dans les différents sols de la régionouest de l’Algérie a été investiguée par l’isolement de ce genrebactérien à partir des échantillons de sols prélevés de différentsbiotopes ; Cet isolement a été fait sur plusieurs milieux de culturecomme ISP2, GLM et Bennett. La mise en évidence de l’activitéantibactérienne (premier criblage) des 85 souches isolées et suspectéesêtre Streptomycètes a été faite par la méthode de la doublecouche ; plus de 25% des souches isolées avaient un effetantibactérien contre au moins une seule souche bactérienne pathogèneet seulement 3 ont été sélectionnées pour les prochains tests.L’extraction des métabolites secondaires des souches sélectionnées(SM2.2, SM1B1 et TB1) a été faite par solvant (chloroforme) et leurconcentration par un Rotavapor. L’évaluation de l’effet antibactériende l’extrait était réalisée par la méthode des disques contre les mêmessouches pathogènes utilisées dans le premier criblage. Seule l’isolatTB1 a présenté un effet satisfaisant lors du deuxième criblage. Lesdifférents extraits ont subis une analyse UV-visible ainsi qu’une préidentification des molécules présentes par chromatographie sur couchemince (CCM), là où les taches des molécules ont été bien clairessurtout pour l’isolat TB1. L’extraction de l’ADN des souchessélectionnées a été faite par un kit, et une amplification (PCR) du gèneADNr 16S a été réalisée avec un programme approprié, des amorcesuniverselles ont été utilisées et la détection sur gel d’agarose a donnédes fragments de 1500 pb référant à la longueur du gène amplifié.
Mots clés:
Streptomycètes; Activité Antibactérienne; Extraction; CCM; PCR;ADN; Sol; Isolement; Métabolites; UV.





























