Embed Size (px)
Citation preview

THE SEXUAL ISOLATION BETWEEN Drosophila melanogasterAND Drosophila simulans
BY AUBREY MANNINGDepartment of Zoology, University of Edinburgh
IntroductionBehavioural barriers to inter-specific hybrid-
ization are widespread in Drosophila, and theireffects have been measured in a number of waysfor a wide range of species, (see Patterson &Stone, 1952, for a thorough review of the liter-ature to that date) . It is usual for females of onespecies to recognise and discriminate against allforeign males, though such discrimination maynot be absolute, and often the males also dis-criminate and do not court foreign females sofrequently as their own . In relatively few cases,however, have the stimuli involved in specificrecognition been analysed . Mayr (1950) showedthat removing the antennae from females of D.pseudoobscura and persimilis almost completelyprevented them discriminating between malesof their own and the other species . Similarly,Spieth (1952) showed that removing the fore-tarsi from males of two strains of D. virilishampered their discrimination of their ownfemales from related ones, which they mustnormally recognise by contact chemical stimuliwhen they tap with the fore-legs prior to court-ship. The tarsi of many Diptera have long beenknown to be richly provided with chemosensoryhairs .
This paper describes some experiments whichanalyse to a certain extent the basis for thestrong sexual isolation between the very closelyrelated D. melanogaster and D. simulans. Thehybridization of these species was first de-scribed by Sturtevant (1920) who found, as havemany workers since, that the cross using melano-gaster males and simulans females is more diffi-cult than the converse one . Drosophila workersgenerally recommend that for either cross agedmales should be used with very young females to
60
obtain the greatest success, although the reasonsfor this are not clear .
Various genetical stocks of closely relatedspecies will almost certainly differ in their degreeof isolation. Koopman's (1950) work withpseudoobscura and persimilis, for example,shows how easily selection can alter isolation, sothe quantitative data provided here are only validfor the stocks used. The qualitative stimuli in-volved in isolation probably vary rather less andone is safer in making generalizations .
Materials and MethodsThe stocks used were all "wild-type" and kept
as outbred as possible. A large number of pairswere used to found each generation and newstocks, including some caught in the wild, wereadded as they were obtained, though not duringthe course of these experiments . For single pairmatings and observation of courtship smallperspex cells were used (see Bastock & Manning,1955) whilst mass matings were made in 3" x 1"vials with food . The flies were reared and theexperiments all made at 25°C .
The Receptiveness of FemalesAs mentioned above, females are said to be morereceptive to foreign males when they are young,and Table I shows the results of one experimentto test this . Five 3-day-old males of one speciesand ten I- or 3-day-old females of the other wereput unetherised into a vial with food, and thefemales dissected at the end of 48 hours todetermine the number inseminated . No femaleswhich were 3 days old at the start of the experi-ment had been inseminated, but a small per-centage of those that were only 1 day old at thestart were fertilized, and melanogaster females
Table I. The Proportion of Females Inseminated by Foreign Males in 48 Hours . Fivedd and 10fl in a 3" X 1" vial with food-a few females were lost during transfers and dissection. Female age refers to that at start of test ; all males were 3 days old
at start.
1-day Y?
3-day 99 Antennaless 1-day 99
Sim. d'd' x mel. 9? 35/147 0/136 15/146
Mel. de x sim. 9?
10/140 0/137 9/134

MANNING : SEXUAL ISOLATION BETWEEN Drosophila melanogaster & Drosophila simulans
61
had accepted about three times as many foreignmales as had simulans . .
It is of interest to see if this apparent fall inreceptivity with age is true of females courted bytheir own males, and to this end, a large numberof single pair matings were, timed using femalesof various ages, but always with mature 3-day-old males. Both males and females were trans-ferred to fresh food every day and not morethan twenty insects were kept in a vial togetherwhilst ageing. Under such conditions almost100 per cent. of males court well within a fewseconds of introduction with a female into anobservation cell . Only courtships which werepractically continuous were recorded and maleswere discarded unless they began to court within2 minutes. At the end of 20 minutes continuouscourtship without copulation the pair was dis-carded and a courtship time of "20+ minutes"recorded. Fig . 1 . shows histograms of the court-ship time to copulation, grouped in 2-minuteintervals, with females of different ages . It is notpossible to detect any differences in the courtshipof males with females of the different ages, andthus the variation due to individual male differ-ences is probably equal for each histogram .Withboth species it is clear that females are mostreceptive, and least variable in this respect,when 3 days from eclosion and that there is agradual increase in receptiveness with ageingup to 3 days. By 4 days from eclosion there aresome signs of increased variability and decreasedreceptiveness, but measures of virgins 10 daysold or more (not illustrated) show very littlechange from the 4-day averages . Receptivenessprobably declines slowly after the slight fallingoff from the peak at 3 days old. Very old virgins--some were tested at an age of 85 days-arevery unreceptive and extrude their ovipositorwhen courted in the manner of fertilized females.
It seems very likely that this receptivity peak iscorrelated with the maturation of the ovarieswhich also reach a maximum at about this age .The effect of ovarial growth is unlikely to be adirect one ; no kind of sex hormone has beendemonstrated in insects and Maynard Smith(1958), who finds the same increase in receptive-ness in D. subobscura, has shown that congenit-ally ovariless females go through the same be-havioural cycle as normal ones .
Since the receptivity of females towards theirown males is lowest when they are newly emergedit is unlikely that the increased success offoreign males with young females can be ex-plained in terms of higher receptivity . It is
Fig. 1 . The receptivity of females with agd, expressed asthe percentage copulating within a given time with 3-day-old males. Each block of the histograms represents aspan of 2 minutes and its height denotes the percentageof females which accepted males after the correspondingperiod of continuous courtship . The bracketed figuresafter the age of the females give the number of pairsrecorded for each histogram .
necessary therefore to consider either that youngfemales cannot discriminate so well as older ones,or that males do not court the different agegroups of foreign females equally . We may nowconsider evidence which bears on the second ofthese possibilities .
Discrimination by MalesMales of both melanogaster and simulans from
a few hours of age will approach most movingobjects of roughly the right size and tap withtheir fore-tarsi . They will court their own femaleswhen these are any age from a few hours to
62
ANIMAL BEHAVIOUR, VII, 1 -2
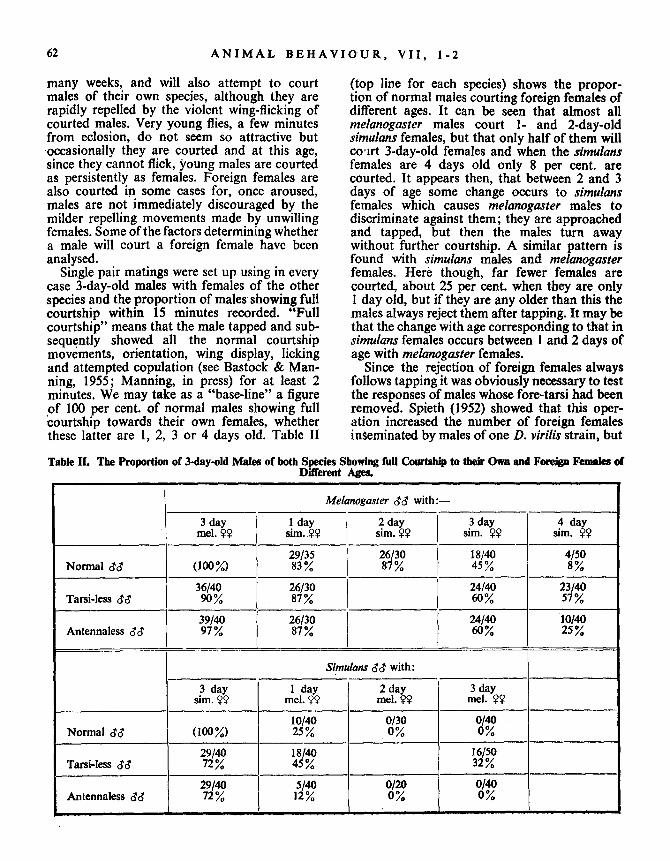
many weeks, and will also attempt to court (top line for each species) shows the propor-males of their own species, although they are tion of normal males courting foreign females ofrapidly repelled by the violent wing-flicking of different ages . It can be seen that almost allcourted males. Very young flies, a few minutes melanogaster males court 1- and 2-day-oldfrom eclosion, do not seem so attractive but simulans females, but that only half of them will
,occasionally they are courted and at this age, co ,irt 3-day-old females and when the simulanssince they cannot flick, young males are courted females are 4 days old only 8 per cent. areas persistently as females. Foreign females are courted. It appears then, that between 2 and 3also courted in some cases for, once aroused, days of age some change occurs to simulansmales are not immediately discouraged by the females which causes melanogaster males tomilder repelling movements made by unwilling discriminate against them ; they are approachedfemales. Some of the factors determining whether and tapped, but then the males turn awaya male will court a foreign female have been without further courtship . A similar pattern isanalysed .
found with simulans males and melanogasterSingle pair matings were set up using in every
females. Here though, far fewer females arecase 3-day-old males with females of the other
courted, about 25 per cent . when they are onlyspecies and the proportion of males showing full
1 day old, but if they are any older than this thecourtship within 15 minutes recorded . "Full
males always reject them after tapping. It may becourtship" means that the male tapped and sub-
that the change with age corresponding to that insequently showed all the normal courtship
simulans females occurs between I and 2 days ofmovements, orientation, wing display, licking
age with melanogaster females .and attempted copulation (see Bastock & Man-
Since the rejection of foreign females alwaysning, 1955 ; Manning, in press) for at least 2
follows tapping it was obviously necessary to testminutes. We may take as a "base-line" a figure
the responses of males whose fore-tarsi had beenof 100 per cent. of normal males showing full
removed. Spieth (1952) showed that this oper-courtship towards their own females, whether
ation increased the number of foreign femalesthese latter are 1, 2, 3 or 4 days old . Table II
inseminated by males of one D . virilis strain, but
Table II. The Proportion of 3-day-old Males of both Species Showing full Courtship to their Own and Foreign Females ofDifferent Ages.
Melanogaster dd with :-
3 daymel . ~~
1 daysim. ??
2 daysim. W
3 daysim . ??
4 daysim. Y?
29/35 26/30 18/40 4/50Normal dd (100%) 83% 87 % 45% 8
36/40 26/30 24/40 23/40Tarsi-less dd 90 % 87% 60% 57
39/40 26/30 24/40 10/40Antennaless dd 97 % 87 % 60 % 25
Simulans dd with :
3 daysim. $?
1 daymel . ??
2 daymel. W
3 daymel . ??
10/40 0/30 0/400Normal do' (100Y) 25 % 0
29/40 18/40 16/50Tarsi-less de 72 0% 45% 32
29/40 5/40 0/40Antennaless dd 72 / 12
0/~

MANNING : SEXUAL ISOLATION BETWEEN Drosophila melanogaster & Drosophila simulans
63
lie did not observe the effect upon the males'behaviour . As shown on Table II, melanogastermales without their fore-tarsi court their ownfemales practically normally, only four out offorty failing to respond . Simulans males doshow a significant reduction (x 2=--10 .55, P<0 .01) following the operation, but nearly three-quarters of them court . Tarsi-less males ap-proach and try to tap the females, usually comingcloser to them than normal and pushing partof the way on to the female's back with thestumps of their fore-legs . Most of the operatedmales are sufficiently aroused for their subse-quent courtship is quite normal, although theylag behind the females somewhat because theirrunning is hampered. It appears that the chem-ical stimuli received by a male on tapping are notessential for arousal in most cases .
The results with foreign females show con-clusively that tarsi-less males do not distinguishforeign females so well as intact ones. Withmelanogaster the proportion of tarsi-less malescourting 3-day-old simulans females is somewhatincreased over normal, and that courting 4-day-old females is very greatly increased by someseven times (X2-23-62, P<0 .001). This patternis even more vivid with operated simulans males .Here the proportion courting 1-day-old melano-gaster females is nearly doubled and nearly onethird of tarsi-less males show full courtship to3-day-old females which are never courted bynormal males. Thus although no essentialpositive stimuli results from tapping conspecificfemales, it appears that inhibitory stimulinormally result from tapping mature foreignfemales. In the absence of these inhibitorystimuli, the other positive stimuli to courtshipwhich such females provide suffice to elicit aresponse in many cases .
A further series of tests was performed withmales whose antennae had been removed . Mayr(1946) found that this operation slightly reducedthe isolation between pseudoobscura and per-similis . Begg & Packman (1951) who usedmelanogaster with varying manifestations ofthe gene "antennaless", found that flies withoutantennae produced very few inseminated femaleswhen crossed and there were clear indicationsof reduced sexual activity from males as wellas females. It seems possible that this may bedue to other side effects of the antennalessgene, for I find very little sign of reduction insexual activity with normal melanogaster maleswhose antennae are removed . All but oneof the forty antennaless males showed full
and vigorous courtship towards their ownfemales and they copulated within the normaltime range. There was some indication that theytook a little longer before they approached andtapped females, but lowered general activity istypical of flies without their antennae and is notspecific to sexual behaviour . As with removingthe tarsi, so the operation of removing antennaedid reduce the number of simulans males whichwere aroused (x 2=10.55, P<0 .01) and some25 per cent . did not court their own females .Antennaless simulans are very sluggish indeed,but most that did approach and tap femaleswere aroused and copulated in normal times.
Loss of the antennae had rather differenteffects on isolation with the two species . Withmelanogaster the proportion of antennalessmales that court 1- or 3-day-old simulansfemales is no different from normal . More court4-day-old females than normal (the differenceis just significant at the 5 per cent . level on thesefigures, x 2=3 .81) but a smaller proportion thanwas the case with tarsi-less males (x2 for thedifference-7 .42, P<0 .01). However with simu-lans the antennaless males are more isolated thannormal flies, for only five out of forty such malescourt i-day-old melanogaster females and nonecourt females older than this . This may simplybe due to the raised reaction thresholds ofantennaless simulans males, but it is also possiblethat stimuli perceived by the antennae arerelatively less important for the discriminationof foreign females in simulans than melanogaster .Consequently inhibitory stimuli received by thetarsi are able to prevent courtship even thoughany inhibition via the antennae has been elimin-ated, while in melanogaster the absence of an-tennal inhibitory stimuli decreases discrimin-ation even in the presence of the tarsi . Even if thetwo species do differ in this respect, there is littledoubt that the fore-tarsi are absolutely moreimportant for discrimination in melanogastertoo for, as we have seen, their removal producesa greater reduction in isolation than loss of theantennae .
Summarising, we may conclude that males ofboth species court young foreign females betterthan old ones and that the increasing discrimin-ation against the latter as they age is related tothe strengthening of some species-specific scent .The evidence gained from depriving males ofvarious sense organs indicates that this specificscent is concentrated on the female's body sur-face, but some is probably air-borne and can bedetected by a male without fore-tarsi when he is

64
close to the female. Neither tarsal chemo-receptors or the scent receptors of the antennaeare necessary to arouse courtship, and about 50per cent. of males from which both have beenremoved still court their own females fairly well .However both sets of receptors, and especiallythose of the fore-tarsi, are sources of inhibitinginputs when a male approaches and taps amature foreign female .
It would be of great interest to see whether theparticular chemoreceptor neurones which re-spond to a foreign female's scent correspond tothose described by Dethier (1955) in Phormiawhich react to unpleasant substances . Phormiachemoreceptors have another neurone whichresponds only to sugars and other pleasant sub-stances, but the present experiments do not en-able one to conclude with certainty that femalesof his own species provide a Drosophila male'schemoreceptors with a corresponding sexualstimulating substance . If so such stimulation isnot indispensable for arousing him .
It is reasonable to conclude that the isolationbetween mature males and females of melano-gaster and simulans is as much due to discrimin-ation by the males as that of the females, butthat between males and young foreign females islargely due to the females. In the former case anydiscrimination by the females is masked, sincemales do not try to court them . In fact maturefemales do discriminate strongly, for so far it hasnot been possible to get any inseminated 3-day-old females by putting them with tarsi-less malesthough these certainly court them better .
Discrimination by FemalesWe may now consider briefly the stimuli by
which females recognise foreign males ; so far allthe evidence is of a negative kind . As was pointedout by Spieth (1952), neither visual stimulinor a "sampling" of the courtship movementsare likely to be involved . Foreign males areusually rejected from the instant they tap thefemale and have no more success in the dark thanthe light . Unlike Mayr's (1950) case with pseudo-obscura and persimilis, removing the antennaefrom females does not affect their ability todiscriminate . The third column of Table I showsthe number of antennaless females, 1-day-old atthe start of the experiment, which were fertilisedby foreign males . The proportion is unchangedfor simulans females, but even shows some re-duction with melanogaster, which is probablycorrelated with the greater reduction in re-ceptiveness which follows the loss of the anten-
ANIMAL BEHAVIOUR, VII, 1- 2
nae in this species compared with simulans(Manning, in press). Since the antennae are thechief receptors for air-borne chemical stimuli,one is left with the conclusion, already suggestedby Spieth (1952) for some species, that it islargely by contact chemoreceptors that thesefemales distinguish their own from foreignmales. It is most likely that the male's act oftapping provides the female with chemicalstimuli and thus a means of discriminating. Itmust be noted that foreign males lacking theirfore-tarsi are rejected just as normal, so thatprobably any contact with the male's body issufficient .
These results do not explain why more simu-lans males are eventually successful with foreignfemales for, in general, they court them less wellthan melanogaster males. Presumably melano-gaster females are not so persistent in theirdiscrimination and resist foreign courtship less .
The Behaviour of HybridsThe hybrids between melanogaster and simu-
lans might be expected to be roughly inter-mediate in their behaviour, but the small amountof information so far available does not confirmthis. Sturtevant (1920) says that hybrid males willcourt either parent or hybrid females but givesno data on preferences . I have always foundthat the responses of male hybrids are scarcelyseparable from those of simulans . The courtshippattern is very similar, and combining severalexperiments, while 28/30 hybrid males courted3-day-old simulans females, none (0/20) courtedmelanogaster females of the same age, though2/10 courted 1-day-old females . A further 1/10hybrid males courted 3-day-old hybrid females .Certainly the hybrid males seem to prefersimulans females and these were the only onesever to accept them ; copulation occurred afterquite normal courtship times . Hybrid females,on the other hand, are only courted well bymelanogaster males and I have not seen simulansmales go beyond tapping them . This result maysimply be due to the lower sexual responsethreshold in melanogaster and need not meanthat the hybrid females produce a melanogastertype scent.
DiscussionWe may deduce that the sexual isolation be-
tween melanogaster and simulans depends mostlyupon female discrimination when the females areyoung, but by 3 days of age both sexes are dis-criminating, and by chemical rather than be-

MANNING : SEXUAL ISOLATION BETWEEN Drosophila niclanogaster & Drosophila simulans
65
havioural means . The specific scents involvedappear to be relatively independent of environ-ment in their development and recognition, forall attempts to influence isolation by rearing thetwo species together, both as larvae and adult,have so far failed . Mayr & Dobzhansky (1945)did succeed in influencing the isolation betweenpseudoobscura and persimilis by rearing them inmixed cultures. They found slightly increasedisolation between flies exposed to the otherspecies during development, but such an effectwould be more difficult to demonstrate withmelanogaster and simulans as their normalisolation is so much greater than that of theother sibling pair.
It is clearly advantageous to have specificrecognition based upon a clear-cut chemicaldifference. Such differences may be expected toarise at a relatively early stage in divergencefrom a common ancestor and provide a rapidmeans for recognising a suitable mate . It is notso clear why the specific scents of the femalesshould take one or two days to become fullyeffective ; rather little is known about the de-velopment of cuticular scent in Drosophila .
As a result of this slow development of scent,males may "waste" some of their attentionsupon unsuitable mates . However in a mixedpopulation under natural conditions, it is veryunlikely that males will court young foreignfemales for long. Such females are giving morewing-flicking and other repelling stimuli and thecourting males will soon desist and approachother females. Obviously it is advantageous formales, who will mate many times in succession,to be easily aroused and approach any fly in thevicinity to test its suitability . Females whichprobably rarely mate more than twice and areunreceptive to males for several days followingcopulation, must choose their mates withcertainty.
Summary1 . Some of the stimuli involved in the sexual
isolation between D . melanogaster and D .simulans have been analysed .
2. Females of both species show a slow rise inreceptiveness to their own males which reaches
a peak at 3 days of age and thereafter falls slow-ly. Foreign males are more successful with young(1-day-old) females than mature ones .
3. Males court young foreign females betterthan mature ones, but removing their fore-tarsiprevents their discriminating against the latter toa marked extent .4. Antennaless melanogaster males also show
some reduction in isolation, but this is not seenwith simulans, whose males are more affected bysensory deprivation .
5. The behaviour of hybrid flies is brieflydiscussed .
REFERENCESBastock, M. & Manning, A . (1955) . The courtship of
Drosophila melanogaster. Behaviour, 8, 85-111 .Begg, M. & Packmann, E. (1951). Antennae and mating
behaviour in Drosophila melanogaster. Nature,(Land.), 168, 953 .
Dethier, V. G . (1955) . The physiology and histology ofthe contact chemoreceptors of the blowfly . Quart.Rev . Biol., 30, 348-371 .
Koopman, K. F. (1950) . Natural selection for repro-ductive isolation between Drosophila pseudo-obscura and Drosophila persimilis . Evolution, 4,135-148 .
Manning, A. (In press) . The sexual behaviour of twosibling Drosophila species. Behaviour .
Maynard Smith, J . (1958). The effects of temperature andof egg-laying on the longevity of Drosophilasubobscura. J. exp . Biol., 35, 832-842 .
Mayr, E . (1946). Experiments on sexual isolation inDrosophila. VII. The nature of the isolatingmechanisms between Drosophila pseudoobscuraand Drosophila persimilis. Proc . nat . Acad. Sci.(Wash.), 32,128-137 .
Mayr, E . (1950) . The role of the antennae in the matingbehaviour of female Drosophila. Evolution, 4,149-154 .
Mayr, E. & Dobzhansky, T . (1945) . Experiments onsexual isolation in Drosophila. IV. Modificationof the degree of isolation between D. pseudo-obscura and D. persimilis and sexual preferencein D. prosaltans . Proc . nat. Acad. Sci . (Wash .),31, 75-82 .
Patterson, J . T. & Stone, W. S. (1952) . Evolution In thegenus Drosophila. New York: Macmillan .
Spieth, H. T . (1952) . Mating behaviour within the genusDrosophila (Diptera) . Bull. amer . Mus. nat .Hist ., 99, 395-474 .
Sturtevant, A. H . (1920) . Genetic studies on Drosophilasimulans . I. Introduction. Hybrids with Drosophilanielanogaster . Genetics, 5, 488-500.
Accepted for Publication 17th February, 1959,





























