Embed Size (px)
Citation preview

Traffic 2012 © 2012 John Wiley & Sons A/S
doi:10.1111/j.1600-0854.2012.01380.x
The Role of Conserved PEX3 Regions in PEX19-Bindingand Peroxisome Biogenesis
Friederike Schmidt1, Denise Dietrich1, Roy
Eylenstein2, Yvonne Groemping2, Thilo
Stehle1,3,∗ and Gabriele Dodt1,∗
1Interfaculty Institute of Biochemistry, University ofTubingen, D-72076, Tubingen, Germany2Max-Planck Institute of Developmental Biology,Department of Protein Evolution, D-72076, Tubingen,Germany3Department of Pediatrics, Vanderbilt University Schoolof Medicine, Nashville, TN 37232, USA*Corresponding authors: Gabriele Dodt, [email protected] and Thilo Stehle, [email protected]
The human peroxins PEX3 and PEX19 are essential
for peroxisome biogenesis. They mediate the import
of membrane proteins as well as the de novo forma-
tion of peroxisomes. PEX19 binds newly synthesized
peroxisomal membrane proteins post-translationally and
directs them to peroxisomes by engaging PEX3, a protein
anchored in the peroxisomal membrane. After protein
insertion into the lipid bilayer, PEX19 is released back to
the cytosol. Crystallographic analysis provided detailed
insights into the PEX3–PEX19 interaction and identi-
fied three highly conserved regions, the PEX19-binding
region, a hydrophobic groove and an acidic cluster, on the
surface of PEX3. Here, we used site-directed mutagene-
sis and biochemical and functional assays to determine
the role of these regions in PEX19-binding and peroxi-
some biogenesis. Mutations in the PEX19-binding region
reduce the affinity for PEX19 and destabilize PEX3. Fur-
thermore, we provide evidence for a crucial function of
the PEX3–PEX19 complex during de novo formation of
peroxisomes in peroxisome-deficient cells, pointing to
a dual function of the PEX3–PEX19 interaction in per-
oxisome biogenesis. The maturation of preperoxisomes
appears to require the hydrophobic groove near the base
of PEX3, presumably by its involvement in peroxisomal
membrane protein insertion, while the acidic cluster does
not appear to be functionally relevant.
Key words: affinity measurements, de novo peroxisome
formation, membrane protein import, peroxin, perox-
isomal membrane biogenesis, peroxisomal membrane
protein, preperoxisome, protein–protein interaction
Received 21 October 2011, revised and accepted for pub-
lication 18 May 2012, uncorrected manuscript published
online 24 May 2012
Peroxisomes are essential for a multitude of metabolicpathways, such as the β-oxidation of very long andbranched chained fatty acids, plasmalogen biosynthesis,
or the degradation of H2O2 (1,2). Defects in peroxisomebiogenesis can be linked to mutations in PEX genesand form a group of inherited diseases, the peroxisomebiogenesis disorders (3–7). Loss of one of the threeperoxins PEX3, PEX16 or PEX19 results in severe cellularphenotypes in which cells are not able to developdetectable peroxisomal membrane structures, but canbe complemented by reintroduction of the correspondingcDNA (8–12). The phenomenon of peroxisomal de novoformation has been intensively investigated in yeast.In PEX3-deficient cells, newly synthesized PEX3 isfirst located in concentrated foci on the endoplasmicreticulum (ER) before new peroxisomes are formed inthe presence of PEX19 (13–15). While this ER-dependentroute has been found to occur in peroxisome-deficientcells, peroxisomes can multiply by growth and divisionin wild-type cells (16). Recently, it was proposed thatER-dependent peroxisome biogenesis is not restrictedto de novo synthesis in mutant cells, but serves asthe main pathway in peroxisome biogenesis in all cells(17,18). However, it is not known whether these findingscan be generalized to mammalian cells. Although it hasbeen described that newly synthesized PEX16 or PEX3travels via the ER in PEX16- or PEX3-deficient cells,respectively (19,20); it is unclear whether and to whatextent this pathway is actually used when peroxisomesare present (21).
Post-translational import of peroxisomal membrane pro-teins (PMPs) is initiated by the cytosolic receptor PEX19(22), which binds to the membrane peroxisomal targetingsequence (mPTS) (23) of PMPs with its C-terminal domain(24). In addition, PEX19 acts as a chaperone for newly syn-thesized PMPs by shielding hydrophobic patches from theaqueous environment (25,26). The N-terminus of PEX19is sufficient and necessary to localize cargo-loaded PEX19to peroxisomes (25,27,28), where it interacts with PEX3(29–31). The cytosolic C-terminal domain of PEX3 formsan α-helical bundle and mediates high affinity bindingto PEX19 (32–34). PEX3 is anchored in the peroxisomalmembrane with its first 34 N-terminal amino acid residues,which also harbor the information for its targeting to perox-isomes (35,36). The mechanism of membrane insertion ofPMPs and the role of ATP in PMP import remain to be elu-cidated. Whereas one study detected an ATP-dependenttargeting of PEX19 to peroxisomes (22), other findingsindicate a negligible role of ATP during the whole process(37,38). In addition to ATP hydrolysis, the thermodynam-ically favored embedding of transmembrane segmentsinto the lipid bilayer might present a driving force for PMPinsertion.
www.traffic.dk 1

Schmidt et al.
The crystal structure of the cytosolic domain of PEX3 incomplex with a PEX19-derived peptide (33,34) identifiedseveral conserved surface regions: the region that bindsthe PEX19 peptide, as well as a hydrophobic groove anda patch of acidic residues near the base of the protein.We introduced structure-guided surface mutations intothese conserved regions in order to probe their functionalrelevance. Some of the PEX3 mutants were expressedin Escherichia coli, purified and analyzed with respectto their folding characteristics and their ability to bindPEX19 in vitro. In addition, full-length mutant proteinswere analyzed in vivo to define their localization in normalfibroblasts and their functionality in de novo peroxisomeformation in PEX3-deficient cells. Our findings provideinsights into the likely function of the analyzed areas, andallow for the formulation of a more detailed model forprotein import into the peroxisomal membrane.
Results
The soluble domain of human PEX3, which wascrystallized in complex with a peptide derived fromits interaction partner PEX19, folds into an elongated,α-helical bundle (33). At the peroxisome surface, the longN-terminal helix would be connected to a transmembrane-spanning segment that is not part of the crystallizedprotein (33,34). The predominantly hydrophobic PEX19-binding region is located at the top of the helical bundle,distant from the membrane anchor. Analysis of the proteinsurface identified three highly conserved regions withinPEX3: the binding region for PEX19 at the top of the helicalbundle, a hydrophobic groove near the base of the proteinand a cluster of acidic residues that also lies near the baseof PEX3, but on a different face of the protein (Figure 1)(33). Reasoning that conserved regions are likely to play arole in peroxisome biogenesis and binding to peroxisomalor other proteins, we mutated single amino acids in allthree areas (Figure 1). The first set of three mutations,L93N, A323Q and K324A, is located in the binding regionfor PEX19. L93 mediates hydrophobic interactions withPEX19 (L26), and mutation to asparagine would eliminatethese interactions while keeping the approximate size ofthe side chain. In addition, L93 might contribute slightlyto the structural integrity of PEX3 by connecting helicesα2 and α3. The methyl group of A323 forms van derWaals interactions with several hydrophobic amino acidsof PEX19 (L21, L22, A25). Its replacement with glutamineshould at least partly occupy the PEX19-binding groove,and introduce steric interference. The K324 side chaininteracts with PEX19 primarily via a hydrogen bond anda salt bridge. Furthermore, the carbon atoms of the sidechain are engaged in hydrophobic contacts with Leu21 andAla25 of the PEX19-derived peptide. Mutation to alaninewould eliminate these interactions. A complementarymutational analysis of the PEX19-helix interacting withthis hydrophobic-binding region revealed the importanceof L18, L21, L22 and F29 of PEX19 for PEX3 bindingand peroxisome biogenesis (34). The second set of
Figure 1: Distribution of conserved amino acids on the
surface of sPEX3. Surface representations of sPEX3 with thebound PEX19-derived peptide depicted as an orange ribbon(PDB entry 3MK4, reviewed in 33) differ by 180◦C along theindicated vertical axis. Conservation analysis of surface exposedresidues revealed three conserved regions within sPEX3, whichare highlighted in shades of green: The PEX19-binding regionat the top of the helical bundle, where amino acids L93, A323and K324 are located; a hydrophobic groove that includes aminoacids R53, H57, M72, I140 and L165; and an acidic patch thatis delineated by residues E266 and D275. The color code is thesame as used in Ref (33). Mutations in all three regions wereinserted into sPEX3 for in vitro analysis and into PEX3-myc orPEX3-YFP for in vivo experiments.
mutations probes the role of the highly conserved regionthat stretches along one side of PEX3 and defines ahydrophobic groove. Two amino acids (I140, L165) liedirectly in this groove whereas the other amino acids(R53, H57, M72) are located at its border. R53 and H57flank the hydrophobic groove toward the base of PEX3and were each changed to an alanine to abolish thepolar characteristics of the original amino acids. M72is located at the top of the hydrophobic groove andwas mutated to a lysine, thus keeping a large sidechain, but introducing a charge. Residues I140 and L165were each mutated to an asparagine, a more hydrophilicresidue of a similar size, in order to change the physicalproperties of the conserved hydrophobic groove. A doublemutant (I140N*L165N) was also generated to furtherreduce the hydrophobicity in this area. Furthermore,I140 and L165 were mutated to tyrosine and arginineresidues, respectively, to block the groove with largerside chains. The third set of mutations targeted theacidic cluster of PEX3. Here, residues E266 and D275were individually mutated to alanine to probe the putativefunction of this cluster by reducing its overall electrostaticpotential.
Mutations do not influence the overall fold
and the stability of recombinant sPEX3
Selected mutations were introduced into an E. coliexpression vector coding for human PEX341-373 witha cysteine to serine exchange at position 235. Thecorresponding protein (PEX341-373 C235S) is hereafterreferred to as sPEX3 and is identical to the proteincrystallized earlier (33). Six sPEX3 mutants (L93N,
2 Traffic 2012

Functions of Conserved Regions in PEX3
K324A, E266A, I140N, L165N and the double mutantI140N*L165N) were expressed in E. coli and purified tohomogeneity using an established protocol (33). To testwhether the mutations affect the overall fold of sPEX3,all mutants were subjected to circular dichroism (CD)spectroscopy. sPEX3 and the six mutants show similarspectra, with clear α-helical features represented by twominima of ellipticity at 208 and 220 nm (Figure S1A).This indicates that the mutagenesis did not alter theoverall fold of the protein. However, CD spectroscopycan only reveal larger changes in the folding of aprotein, and is not sensitive enough to detect smallerstructural rearrangements or local unfolding. Therefore,the stabilities of the mutants were also analyzed usinga thermal denaturation assay, in which a CD-signal wasrecorded at 208 nm while the sample was simultaneouslyheated to 95◦C. The melting point of sPEX3 wasdetermined as 43◦C. As all mutants melt at similar oreven higher temperatures (Figure S1B), we conclude thatintroduction of the mutations has no adverse effect oneither fold or stability of sPEX3.
Mutations in the PEX19-binding region reduce the
affinity of sPEX3 for PEX19
The binding affinities of sPEX3 and the sPEX3 mutantsfor full-length PEX19 were determined using isothermaltitration calorimetry (ITC). sPEX3 binds PEX19 withnanomolar affinity (Kd = 14 nM, Figure 2A, E), consistentwith previous findings (32,33). The mutations I140N,L165N and E266A do not result in significant changesof affinity, as the Kd values lie in the range of 20 nM
for all three proteins (Figures 2D and S2A–C, Table 1).However, the mutations located in the PEX19-bindingregion influence the PEX3–PEX19 interaction properties.The K324A mutation lowers the affinity for PEX19 about20-fold (Kd = 270 nM). This decrease is in agreement withthe expected loss of one salt bridge, one hydrogen bondand the reduction of hydrophobic interactions to L21 andA25 upon complex formation. This would reflect the 100-fold reduction of the Kd to 346 nM for the Pex19 L21Amutant compared to wild type as determined by Satoet al. (34) with surface plasmon resonance analysis. Thebinding constant is still high enough to detect the complexformation between the K324A mutant and PEX19 insolution (Figure S2D). With a Kd value of 3.7 μM, the L93Nmutant is severely impaired in PEX19-binding (Figure 2B,E). As the sPEX3 L93N mutant was shown to be correctlyfolded and stable in vitro (Figure S1), the major structuralproperties of sPEX3 should still be conserved in themutant protein. The Kd value corresponds to a 250-foldaffinity reduction compared to sPEX3. The observationthat the complex of the sPEX3 L93N mutant andPEX19 cannot be detected by gel filtration experiments(Figure S2D) confirms these results. As described abovePEX3 L93 mediates hydrophobic interactions with PEX19(L26). When the L26A mutation was complementarilyintroduced into PEX19 and investigated by surface plasminresonance analysis, the reduction in PEX3 binding (10-fold) was smaller (34) as suggested by our results. The
thermodynamic parameters of the interaction of sPEX3mutants with PEX19 are listed in Table 1.
The N-terminus of PEX3 is not involved in high
affinity PEX19-binding
To exclude that the N-terminus of PEX3, which isabsent in sPEX3 and the mutant proteins, influencesthe interaction with PEX19, we tested full-length PEX3for the ability to bind PEX19 by coimmunoprecipitation(CoIP). All PEX3 mutants used in ITC measurements(L93N, K324A, E266A, I140N, L165N) and the additionalmutants A323Q (PEX19-binding region), R53A, H57A,M72K, I140Y, L165R, I140N*L165N (hydrophobic groove)and D275A (acidic cluster) were introduced into themammalian expression vector PEX3-myc, which encodeshuman full-length PEX3 with a C-terminal myc-tag. PEX19and all PEX3-myc mutants were first transcribed andtranslated separately in vitro, then mixed, incubated for1 h and subjected to CoIP using beads coupled to α-mycantibodies. The translation products, the supernatantsand the elution fractions were separated by SDS–PAGEprior to autoradiographic detection of the 35S-methionine-labeled proteins (Figure 2F, G). Full-length PEX3-myc andthe mutants in the hydrophobic groove (R53A, H57A,M72K, I140N, I140Y, L165N, L165R, I140N*L165N) andin the acidic cluster (E266A and D275A) are able to interactwith PEX19, as indicated by its detection in the eluate.PEX3-myc L93N and A323Q are not able to precipitatePEX19, as the corresponding PEX19-band is absent in theeluate (Figure 2F, lane 3, G, lane 8). In case of PEX3-mycK324A, PEX19 can be detected in the eluated fraction(lane 4); however, the corresponding band is weakercompared to PEX3-myc. The negative control (Figure 2F,lane 2, G, lane 2), where PEX19 was mock treated in theabsence of PEX3-myc, excludes any unspecific binding ofPEX19 to the beads. Thus, it is likely that the mutationsin the PEX19-binding pocket (L93N and A323Q) severelyimpair PEX19 binding and that the N-terminus of PEX3 isnot involved in a high affinity interaction with PEX19.
PEX3 mutants localize correctly to peroxisomes
in human fibroblasts
Selected plasmids coding for a subset of PEX3-mycmutants (L93N, K324A, I140N, L165N, I140N*L165N,E266A, D275A) were transfected in human fibroblastcells, and the expressed proteins were tested for per-oxisomal localization using indirect immunofluorescencemicroscopy. PEX3-myc and all mutants showed an unam-biguous peroxisomal staining pattern and colocalized withthe peroxisomal membrane protein PEX14 (Figures 3Aand S3A). However, the expression rates differ signif-icantly among some of the mutants (Figure 3B). Theamount of PEX3-expressing cells for the mutations E266A,D275A, I140N and the double mutation I140N*L165N liesin the same range as quantified for PEX3-myc (9–12%of total cells). With an expression rate of 3.5%, a three-fold reduction was observed for the PEX3-myc L165Nmutant. Interestingly, the two mutations located in the
Traffic 2012 3

Schmidt et al.
A
F G
D E
B C
Figure 2: Analysis of complex formation between different sPEX3 mutants and PEX19 using ITC and CoIP experiments. RawITC-data for titrations of sPEX3 with PEX19 (A), sPEX3 L93N with PEX19 (B), sPEX3 K324A with PEX19 (C) and sPEX3 L165N withPEX19 (D) are shown. E) Integrated heat values for the different ITC experiments showing fits to a single binding isotherm for therespective titrations. The data for affinity measurements of the mutants E266A and I140N with PEX19 are displayed in Figure S2. F)and G) Autoradiography of CoIP experiments using full-length PEX3-myc and full-length PEX19. PEX3-myc, the corresponding mutantsand PEX19 were obtained by an in vitro TNT assay, mixed and subjected to coimmunoprecipitations using α-myc antibodies coupledto magnetic beads. The translation products, the supernatants and the eluates are shown separately for each PEX3-myc mutant. AllPEX3-myc mutants except PEX3-myc L93N and A323Q precipitate PEX19. Note that the corresponding PEX19-bands are lacking in theeluated fractions (F, lane 3 and G, lane 8) for PEX3-myc L93N and A323Q.
PEX19-binding region, L93N and K324A, have decreasedexpression rates of 2.7 and 6.4%, respectively, comparedto PEX3-myc. As all PEX3-myc mutants are transcribed atthe mRNA levels in amounts that are 120–250-fold higherthan the mRNA of endogenous PEX3 (Figure S3B), it islikely that the PEX3-myc mutants were also present at theprotein level. The two PEX3 mutants, L93N and K324A,that are impaired in binding to PEX19 might be less stablein a cellular environment, resulting in a lower expressionrate compared to PEX3-myc. To further investigate thispossibility in vitro, we analyzed the stability of recombi-nant sPEX3 in the presence of a PEX19-derived peptidecomprising residues 14–33 (PEX19Pep), which previouslyhas been shown to bind sPEX3 with high affinity (33).
Thermal denaturation curves for sPEX3 alone and in com-bination with PEX19Pep were determined by recording theCD-signal at 208 nm (Figure 4A). The melting point of thesPEX3–PEX19Pep complex is significantly shifted towardhigher temperatures, indicating a higher thermal stabilityof almost 10◦C upon peptide-binding. Thus, PEX19-bindingstabilizes PEX3 in vitro.
The post-translational import of newly synthesized
PEX3 into peroxisomes is independent of PEX19
As the PEX3 L93N and K324A mutants impaired inPEX19-binding still localized correctly to peroxisomes,we investigated whether newly synthesized PEX3 isimported into peroxisomes independently of PEX19.
4 Traffic 2012

Functions of Conserved Regions in PEX3
Table 1: Thermodynamic parameters for the interaction ofsPEX3 proteins and full-length PEX19
Protein Kd (nM) �H (kcal/mol) Molar ratio
sPEX3a 14.8 −9.3 0.89sPEX3 L93N 3750 −10.4 0.90sPEX3 K324A 270.3 −8.0 1.03sPEX3 E266A 17.9 −9.4 0.94sPEX3 I140N 18.3 −7.8 1.25sPEX3 L165N 20.9 −5.6 0.87
aPEX341-373 C235S. All experiments were carried out at 25◦C anddata were fit to a single binding isotherm.
Normal human fibroblasts were treated either with siRNAagainst PEX19 (25) or with control siRNA. Immunoblottingagainst PEX19 revealed a severe reduction of theendogenous PEX19 protein level of 83% compared tocontrol cells (Figure 4B). Interestingly, PEX3 was alsoreduced in these PEX19-knockdown cells and in PEX19-deficient cells (Figure 4B), indicating that PEX3 is indeedmore stable in the presence of PEX19. The siRNA-treated cells were subsequently transfected either withPEX3-myc or PMP70-myc. In both cases, YFP-PTS1 wascotransfected for normalization of transfection efficiency.Colocalization of PEX3-myc and PMP70-myc with theperoxisomal matrix marker YFP-PTS1 revealed a distinctperoxisomal staining in control cells as well as inPEX19-knockdown cells. PEX3-myc and PMP70-mycwere exclusively detected in peroxisomes and were notmislocalized to other organelles (Figure 5A). However,PMP70-myc was less frequently detected in cells treatedwith PEX19 siRNA, likely due to an import defect andsubsequent degradation. These cells showed a 1.5-fold reduction in the ratio of PMP70-myc to YFP-PTS1transfection rates compared to control cells (Figure 5B).However, this PEX19 reduction does not interfere withPEX3-myc import, as PEX3 was localized to peroxisomesto the same extent in control and PEX19-knockdowncells (Figure 5B). These findings support a discrete importpathway for PEX3 to peroxisomes, which is independentof PEX19.
The PEX3 mutants differ in their efficiency
to complement PEX3-deficient human fibroblasts
As PEX3-deficient cells lack any detectable peroxisomes,PMPs are either mistargeted to other organelles, to thecytosol, or degraded (8,9). For example, the peroxisomalmembrane protein PEX14 is mislocalized to mitochondriain �PEX3T cells (Figure 6A, reviewed in 8), and analysisof PEX14 localization therefore can be used as an assay tostudy PEX3 function. A characteristic feature of functionalPEX3 is the ability to induce formation of new peroxisomesin these PEX3-deficient cells. In complemented cells,PEX14 is detected in the restored peroxisomes. Todetermine the functionality of the different PEX3-mycmutants, �PEX3T cells were transfected with thecorresponding plasmids and analyzed for restoration ofperoxisomes 7 days after transfection. The amount of
complemented cells for the PEX3-myc mutants I140Nand E266A did not differ from the complementationlevel of PEX3-myc (4–5% of total cells), whereas themutant D275A lowered the complementation rate byabout 50% (Figure 6B). The import competence of thesenew peroxisomes was validated by immunofluorescencestaining against the matrix protein catalase, whichcolocalizes with the peroxisomal membrane markerPEX14 (Figure S4). The PEX3-myc mutant K324A showeda significantly reduced complementation rate (2% oftotal cells, Figure 6B). However, distinct peroxisomesthat are import-competent for catalase were detected(Figure 6A). This reduced complementation efficiency maybe explained by the reduced PEX19-binding affinity. Incase of the L93N mutation, the complementation abilityis lost, showing the typical mitochondrial mistargeting ofPEX14 (Figure 6A) in �PEX3T cells (8). This observationis consistent with the severe impact of the PEX3L93N mutation on PEX19-binding affinity. Interestingly,a defect in complementing PEX3-deficient human cellswas also observed for the mutant L165N and thedouble mutant I140N*L165N (about 1% of total cells,Figure 6B). Quantitative analysis of complementation ratesfor PEX3-myc and all PEX3-myc mutants is shown inFigure 6B. The in vitro studies described above, usedsPEX3, which carries a cysteine to serine mutation atposition 235. The same mutation was now introduced intothe PEX3-myc plasmid and then transfected into �PEX3Tcells. As PEX3-myc C235S is able to complement �PEX3Tcells to the same extent as PEX3-myc (4.2%, FigureS4), we can exclude any negative impact of the C235Smutation on essential functions of PEX3.
Maturation of preperoxisomal vesicles
The overall complementation rate, which was determined7 days after transfection, does not reflect the influenceof PEX3 on the different steps necessary to create newperoxisomes. We observed that cotransfection of PEX3-YFP and PEX16-myc plasmids into �PEX3T cells leads tothe formation of vesicular structures within 24 h. Thesevesicular structures develop into mature peroxisomeswithin the next few days. In order to investigate the veryearly steps of peroxisome biogenesis in more detail, theappearance of vesicular structures was analyzed after 24 husing antibodies against the peroxisomal proteins PEX14,PEX13 and PMP70. These vesicles do not contain theperoxisomal membrane marker proteins PEX14, PEX13or PMP70, but exhibit a positive staining for PEX3-YFPand PEX16 (Figure 7, day 1). Here, these structures arenow defined as preperoxisomes. The development ofthese structures was also monitored at day 2. At thistime the preperoxisomes import PEX14, PEX13 (Figure 7,day 2), and later also other membrane proteins such asPMP70 (days 2–3) (Figure 7, day 2). This is followedby the translocation of matrix proteins, finally leading tofully functional peroxisomes within 3–7 days, as shownby the peroxisomal catalase staining (Figure 7, day 5).The general time course for the complementation of�PEX3T cells has been described previously (8,39). In
Traffic 2012 5

Schmidt et al.
A
B
Figure 3: Peroxisomal localization of different PEX3-myc mutants in human fibroblasts. Normal fibroblasts (GM5756T) weretransfected with the corresponding PEX3-myc plasmids. A) Cellular localization of the expressed mutants was detected using α-mycantibodies 2 days after transfection. Peroxisomes were visualized using α-PEX14 antibodies. All PEX3-myc mutants reveal a peroxisomaldistribution pattern validated by colocalization with PEX14. The pictures for PEX3-myc and for the representative mutants L93N, K324Aand L165N are shown here, the mutants E266A, D275A, I140N and I140N*L165N are displayed in Figure S3. Scale bar: 10 μm. B)Expression rates, determined by the amount of cells that display a myc-positive peroxisomal staining, are quantified for the individualPEX3-myc mutants. For each of the three independent experiments, at least 500 cells were counted for calculation. Values wererepresented as means with SDs.
6 Traffic 2012

Functions of Conserved Regions in PEX3
A
B
Figure 4: PEX3 is stabilized upon PEX19-binding. A) Thermaldenaturation curves for sPEX3 (black) and sPEX3+PEX19Pep (red).Protein solutions were used at a concentration of 12 μM. Meltingcurves were recorded with a scan rate of 1◦C/min at 208 nmto detect α-helical protein content. The temperature was raisedstepwise to 95◦C. The inflection point indicates the meltingtemperature of the protein, which is clearly shifted to highertemperatures in the presence of the PEX19-derived peptide.B) Human fibroblasts (GM5756T) were electroporated twiceeither with siRNA against endogenous PEX19 or with controlsiRNA. To control the efficiency of PEX19 knockdown, cells wereharvested 48 h after the second electroporation. Equal amountsof total protein were separated by SDS–PAGE and analyzedby immunoblotting using antibodies against PEX19 and PEX3and against tubulin-β and tubulin-α as protein loading controls.Quantification reveals an 83% reduction of PEX19 protein andan 87% reduction of PEX3 protein in PEX19-knockdown cellscompared to control cells.
the experiment presented here, an artificial enhancementof preperoxisome formation by additional expressionof PEX16-myc was applied. Without overexpression ofPEX16, the PEX3-labeled structures are barely detectableafter 24 h (data not shown). Interestingly, the expressionof PEX16-myc is high after 24 h but vanishes below thedetection level in the following days (Figure 7, day 2).
Binding to PEX19 is important for the formation
of preperoxisomes
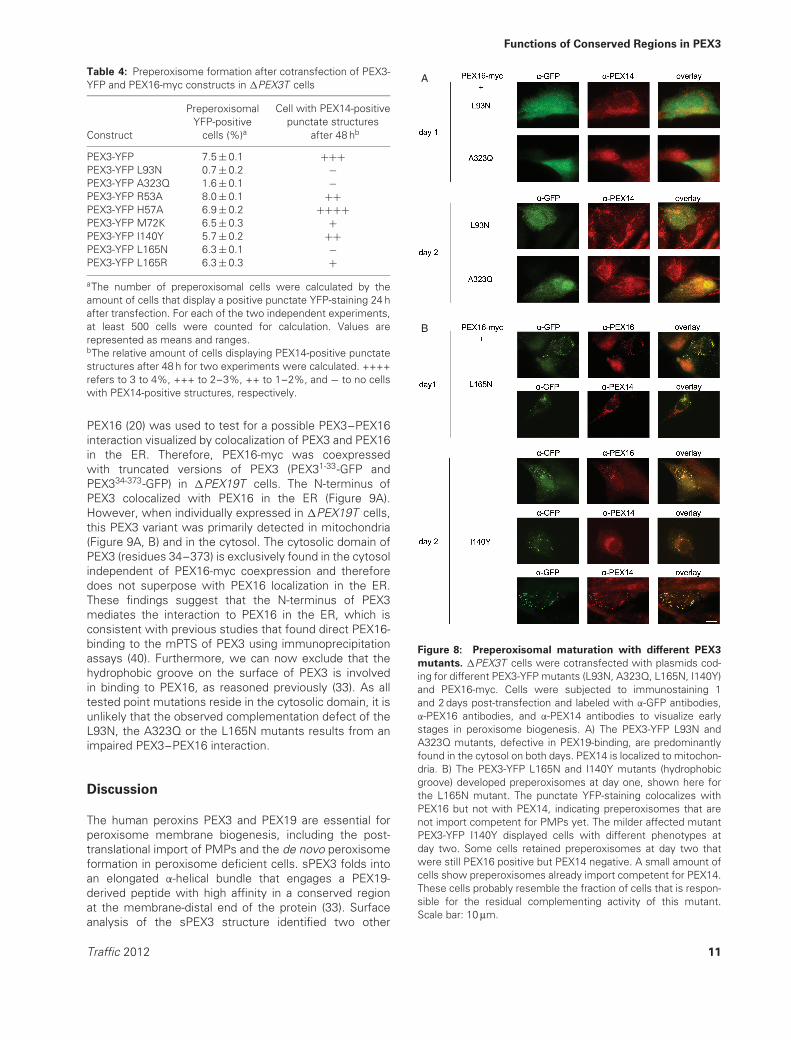
One known function of PEX3 in peroxisome biogenesis isthe binding to PEX19. To further investigate this specificrole in the formation of preperoxisomes, two mutationslocated in the PEX19-binding pocket (L93N, A323Q), werealso introduced into the PEX3-YFP variant. These PEX3-YFP mutants are highly expressed in normal fibroblastcells (Table 2) and localize correctly to peroxisomes.However, it is noteworthy that PEX3-YFP L93N exhibitsa partial cytosolic staining in some cells that was notobvious using the lower expressing PEX3-myc L93Nprotein and which is only barely found for the PEX3-YFPA323Q mutant. The two PEX3-YFP mutants L93N andA323Q have lost the ability to complement PEX3-deficientcells (Table 3). This is likely due to their loss of PEX19-binding as detected in CoIP experiments (Figure 2F, G).The same preperoxisomal assay as described abovewas now carried out for these two PEX3-YFP mutantsand PEX16-myc. Both mutants, PEX3-YFP L93N andPEX3-YFP A323Q, lead to a severely reduced numberof preperoxisomal structures 1 day after transfection(Table 4) and were primarily detected in the cytosol(Figure 8A). In addition these mutants failed to generatePEX14-containing peroxisomes after 2 days, indicating acrucial function of the PEX3–PEX19 complex in the earlystages of peroxisomal de novo biogenesis.
PMPs are imported post-translationally into
preperoxisomes
To probe the function of conserved residues within thehydrophobic groove in the early steps of peroxisomebiogenesis, several PEX3-YFP mutants (R53A, H57A,M72K, I140Y, L165N, L165R) were tested for their abilityto generate preperoxisomes using the assay describedabove. All six PEX3-YFP mutants are expressed andlocalize correctly to peroxisomes in normal cells (Table 2).Furthermore, the mutants R53A, M72K, I140Y andL165R are impaired in complementing �PEX3T cells withcomplementation rates in the range of 50% compared tonormal PEX3-YFP (Table 3). The H57A mutation has almostno influence on the restoration of peroxisomes in �PEX3Tcells. The L165N mutant exhibits a severely reducedcomplementation rate, which is lowered about 85%compared to normal PEX3-YFP (Table 3). This result alsoconfirms our findings obtained for the PEX3-myc L165Nconstruct in complementation experiments (Figure 6B).Surprisingly, all these PEX3-YFP mutants are able togenerate preperoxisomal structures to almost the sameextent as normal PEX3-YFP after 24 h (Table 4). As forPEX3-YFP, these preperoxisomes already contain PEX3and PEX16, but are negative for other PMPs like PEX14(Figure 8B, day 1). Thus, the hydrophobic groove of PEX3is likely not involved in the very early stages of de novoperoxisome biogenesis.
The scenario is different 2 days after transfection: forthe PEX3 H57A mutant, cells begin to exhibit a punctatestaining positive for PEX14 similar to normal PEX3 (see
Traffic 2012 7

Schmidt et al.
A
B
Figure 5: Newly synthesized
PEX3 is imported into peroxi-
somes independently of PEX19.
Human fibroblasts (GM5756T)were electroporated twice eitherwith siRNA against PEX19 orwith control siRNA. A) The siRNAtreated fibroblasts were trans-fected transiently with plasmidDNA coding for PEX3-myc orPMP70-myc and were processedfor indirect immunofluorescence atthe indicated time points post-transfection using α-myc antibod-ies. In both cases, YFP-PTS1was cotransfected to normalizefor cell transfection efficiency.PMP70-myc and PEX3-myc wereexclusively detected in peroxi-somes validated by colocalizationwith YFP-PTS1, scale bar: 10 μm. B)The ratios of transfection rates forPEX3-myc or PMP70-myc to YFP-PTS1 are displayed as a diagram,showing a peroxisomal importdefect in the case of PMP70-myc,but not for PEX3-myc in PEX19-knockdown cells. Please note thatthe time frame is probably too shortto observe an influence of PEX19-knockdown on YFP-PTS1 import.Means of three independent exper-iments are presented with SDs; atleast 1000 cells were counted foreach experiment.
Figure 7 and Table 4). In contrast, the remaining PEX3-YFP hydrophobic groove mutants have only a reducednumber of cells, which show a positive punctate PEX14staining at day 2 (Table 4, Figure 8B). These mutants canbe grouped by severity of this phenotype, with PEX3-YFPL165N, L165R and M72K being strongly impaired in further
development of preperoxisomes and PEX3-YFP I140Y andR53A exhibiting a milder phenotype (Table 4). For thesePEX3 mutants, it is striking that some preperoxisomespersist at day 2 that contain PEX16, but have not yetimported PEX14 (Figure 8B, day 2). If no preperoxisomesare present in a cell, the PEX3 mutants are found in the
8 Traffic 2012

Functions of Conserved Regions in PEX3
A
B
Figure 6: Complementation of
PEX3-deficient human fibroblasts.
�PEX3T cells were transfectedwith plasmids coding for differentPEX3-myc mutants. For comparison,untransfected cells are displayed inthe first lane. A) The correspond-ing complementation efficiency wasdetermined 7 days after transfec-tion using α-PEX14 antibodies todetect newly formed peroxisomes.The import competence of theseorganelles was tested with α-catalaseantibodies. The mutation L93N abol-ishes the ability of PEX3 to com-plement �PEX3T cells showing amitochondrial localization of PEX14.The other PEX3 mutants are stillable to complement �PEX3T cells.�PEX3T transfected with PEX3-mycincluding the mutations L93N, K324Aand L165N are shown here exem-plarily, the mutants E266A, D275A,I140N and I140N*L165N are dis-played in Figure S4. Scale bar: 10 μm.The image for the L165N mutantwas acquired using a z-stack over-lay. B) Complementation rates for allmutants were calculated by count-ing cells that exhibit PEX14-positiveperoxisomal structures. In each ofthe three independent experiments atleast 500 cells were counted for quan-tification. Values were represented asmeans with SDs.
Traffic 2012 9

Schmidt et al.
Figure 7: Stages of complementation of ΔPEX3T cells.
�PEX3T cells were cotransfected with plasmids coding forPEX3-YFP and PEX16-myc. Day 1: Cells were subjected toimmunostaining 24 h post-transfection and labeled with α-GFPantibodies, α-PEX16 antibodies, α-PEX14 antibodies, α-PEX13and α-PMP70 antibodies to visualize early stages in peroxisomebiogenesis. The punctate YFP-staining colocalizes with PEX16 butnot with PEX14, PEX13 or PMP70 indicating preperoxisomes thatare not import competent for PMPs yet. In all cells, endogenousPEX14 is localized to mitochondria. Day 2: The preperoxisomesare negative for PEX16 but contain PEX14 and PEX13. Only veryfew cells are already positive for PMP70 import. On day 5, theperoxisomes are import-competent for catalase. Scale bar: 10 μm.
Table 2: Expression rates of PEX3-YFP constructs in normalfibroblasts
Construct YFP-positive cells (%)a
PEX3-YFP 26.7 ± 1.3PEX3-YFP L93N 16.3 ± 1.4PEX3-YFP A323Q 21.3 ± 1.6PEX3-YFP R53A 28.5 ± 2.1PEX3-YFP H57A 27.8 ± 0.2PEX3-YFP M72K 27.1 ± 0.3PEX3-YFP I140Y 27.8 ± 2.6PEX3-YFP L165N 29.5 ± 1.6PEX3-YFP L165R 28.1 ± 0.8
aExpression rates were calculated by the amount of cells thatdisplay a YFP-positive staining 24 h after transfection of thecorresponding constructs. For each of the two independentexperiments, at least 500 cells were counted for calculation.Values are represented as means and ranges.
Table 3: Complementation rates of PEX3-YFP constructs in�PEX3T cells
Construct Peroxisomal PEX14-positive cells (%)a
PEX3-YFP 7.0 ± 0.9PEX3-YFP L93N No complementationPEX3-YFP A323Q No complementationPEX3-YFP R53A 2.6 ± 0.3PEX3-YFP H57A 6.2 ± 0.1PEX3-YFP M72K 3.1 ± 0.3PEX3-YFP I140Y 3.4 ± 0.3PEX3-YFP L165N 0.8 ± 0.2PEX3-YFP L165R 2.8 ± 0.7
aComplementation rates were calculated by the amount of cellsthat display a positive peroxisomal PEX14-staining 5 days aftertransfection of the corresponding constructs. For each of the twoindependent experiments, at least 500 cells were counted forcalculation. Values are represented as means and ranges.
cytosol, the ER or sometimes in mitochondria. It thereforeseems that the maturation step from preperoxisomes(PEX16 positive) to peroxisomes (PEX14 positive) isblocked or delayed. The PEX3 mutants are still able tofurther develop a low number of cells with PEX14-positivestructures that probably represent the small amountof complemented and import-competent cells detectedafter 5–7 days (Table 3). The reduced number of cellscontaining new peroxisomes with the PEX3-YFP mutantsR53A, M72K, I140Y, L165N and L165R might be dueto the degradation of import-defective preperoxisomes.These findings indicate that the hydrophobic groove mightparticipate in the post-translational import of PMPs intopreperoxisomes.
The N-terminus of PEX3 colocalizes with PEX16
in the ER
In addition to PEX19, only one other peroxin, PEX16, hasbeen shown to interact with PEX3 in mammalian cells(20,40). The observation that transfection of PEX16 intoPEX19-deficient cells leads to a partial ER-localization of
10 Traffic 2012
Functions of Conserved Regions in PEX3
Table 4: Preperoxisome formation after cotransfection of PEX3-YFP and PEX16-myc constructs in �PEX3T cells
Construct
PreperoxisomalYFP-positive
cells (%)a
Cell with PEX14-positivepunctate structures
after 48 hb
PEX3-YFP 7.5 ± 0.1 +++PEX3-YFP L93N 0.7 ± 0.2 −PEX3-YFP A323Q 1.6 ± 0.1 −PEX3-YFP R53A 8.0 ± 0.1 ++PEX3-YFP H57A 6.9 ± 0.2 ++++PEX3-YFP M72K 6.5 ± 0.3 +PEX3-YFP I140Y 5.7 ± 0.2 ++PEX3-YFP L165N 6.3 ± 0.1 −PEX3-YFP L165R 6.3 ± 0.3 +aThe number of preperoxisomal cells were calculated by theamount of cells that display a positive punctate YFP-staining 24 hafter transfection. For each of the two independent experiments,at least 500 cells were counted for calculation. Values arerepresented as means and ranges.bThe relative amount of cells displaying PEX14-positive punctatestructures after 48 h for two experiments were calculated. ++++refers to 3 to 4%, +++ to 2–3%, ++ to 1–2%, and − to no cellswith PEX14-positive structures, respectively.
PEX16 (20) was used to test for a possible PEX3–PEX16interaction visualized by colocalization of PEX3 and PEX16in the ER. Therefore, PEX16-myc was coexpressedwith truncated versions of PEX3 (PEX31-33-GFP andPEX334-373-GFP) in �PEX19T cells. The N-terminus ofPEX3 colocalized with PEX16 in the ER (Figure 9A).However, when individually expressed in �PEX19T cells,this PEX3 variant was primarily detected in mitochondria(Figure 9A, B) and in the cytosol. The cytosolic domain ofPEX3 (residues 34–373) is exclusively found in the cytosolindependent of PEX16-myc coexpression and thereforedoes not superpose with PEX16 localization in the ER.These findings suggest that the N-terminus of PEX3mediates the interaction to PEX16 in the ER, which isconsistent with previous studies that found direct PEX16-binding to the mPTS of PEX3 using immunoprecipitationassays (40). Furthermore, we can now exclude that thehydrophobic groove on the surface of PEX3 is involvedin binding to PEX16, as reasoned previously (33). As alltested point mutations reside in the cytosolic domain, it isunlikely that the observed complementation defect of theL93N, the A323Q or the L165N mutants results from animpaired PEX3–PEX16 interaction.
Discussion
The human peroxins PEX3 and PEX19 are essential forperoxisome membrane biogenesis, including the post-translational import of PMPs and the de novo peroxisomeformation in peroxisome deficient cells. sPEX3 folds intoan elongated α-helical bundle that engages a PEX19-derived peptide with high affinity in a conserved regionat the membrane-distal end of the protein (33). Surfaceanalysis of the sPEX3 structure identified two other
A
B
Figure 8: Preperoxisomal maturation with different PEX3
mutants. �PEX3T cells were cotransfected with plasmids cod-ing for different PEX3-YFP mutants (L93N, A323Q, L165N, I140Y)and PEX16-myc. Cells were subjected to immunostaining 1and 2 days post-transfection and labeled with α-GFP antibodies,α-PEX16 antibodies, and α-PEX14 antibodies to visualize earlystages in peroxisome biogenesis. A) The PEX3-YFP L93N andA323Q mutants, defective in PEX19-binding, are predominantlyfound in the cytosol on both days. PEX14 is localized to mitochon-dria. B) The PEX3-YFP L165N and I140Y mutants (hydrophobicgroove) developed preperoxisomes at day one, shown here forthe L165N mutant. The punctate YFP-staining colocalizes withPEX16 but not with PEX14, indicating preperoxisomes that arenot import competent for PMPs yet. The milder affected mutantPEX3-YFP I140Y displayed cells with different phenotypes atday two. Some cells retained preperoxisomes at day two thatwere still PEX16 positive but PEX14 negative. A small amount ofcells show preperoxisomes already import competent for PEX14.These cells probably resemble the fraction of cells that is respon-sible for the residual complementing activity of this mutant.Scale bar: 10 μm.
Traffic 2012 11

Schmidt et al.
A
B
Figure 9: The N-terminus of PEX3 colocalizes with PEX16
to the ER in PEX19-deficient cells. �PEX19T cells weretransfected with plasmids coding for different GFP-tagged PEX3variants. Additionally, either the control vector pcDNA3.1Zeoor PEX16-myc was cotransfected. A) Cells were analyzed byimmunofluorescence microscopy 24 h after transfection. PEX3localization was detected using α-GFP antibodies; PEX16 wasvisualized with α-myc antibodies, respectively. In the absenceof PEX16-myc, the N-terminus of PEX3 is mainly found inmitochondria. In the presence of PEX16-myc that is partiallylocalized in the ER, this PEX3 variant exhibits a partial ER staining.The C-terminal domain of PEX3 (PEX334-373) is exclusively found inthe cytosol independently of PEX16-myc coexpression; scale bar:10 μm. B) The distribution of the different PEX3-GFP constructswithin the cell in the absence or presence of PEX16-myc wasevaluated and plotted as a diagram. For each of the threeindependent experiments at least 500 cells were counted. Valueswere represented as means with SDs.
regions with high levels of conservation in additionto the PEX19-binding region: a hydrophobic grooveand a cluster of acidic residues. It is tempting tospeculate that the observed evolutionary conservationtranslates into specific functions of these three regionsin peroxisome biogenesis. We have therefore probedthe putative functions of all three regions using site-directed mutagenesis, in vitro binding assays andin vivo localization experiments. We find that mutationsin the PEX19-binding groove destabilize PEX3 at thecellular level, but do only partially interfere with PEX3targeting to the peroxisomal membrane. In addition, theinteraction between these two peroxins is essential forpreperoxisome formation. A set of mutations in thehydrophobic groove does not influence PEX19-bindung
and preperoxisome formation, but is selectively impairedin later stages of peroxisome biogenesis, most likely inpost-translational membrane protein import. The E266Amutation in the cluster of acidic residues did not affectPEX19-binding. In addition, the E266A variant is notimpaired in complementation efficiency, indicating thatthe glutamate at position 266 plays a negligible role inperoxisome biogenesis.
The surface of PEX3 has one major PEX19-binding
site
On the basis of the observation that full-length PEX19binds PEX3 with a higher affinity than an N-terminalPEX19-derived peptide, a second PEX19-binding sitewas proposed to exist in PEX3 (33). Due to theirconservation, the hydrophobic groove and the acidiccluster were thought to be possible candidates for sucha binding site (33). However, as mutations in eitherregion (I140N, L165N, E266A) result in affinities forPEX19 that are comparable to sPEX3 (Kd = 15–20 nM)this possibility can now be excluded, at least for non-farnesylated PEX19 used in this study. Additional CoIPexperiments support these findings, as PEX3 mutantsin the hydrophobic groove (R53A, H57A, M72K, I140Y,I140N, L165N, L165R, I140N*L165N) or in the acidiccluster (E266A, D275A) are still able to precipitate PEX19.Furthermore, ITC measurements demonstrate that PEX3mutations located in the PEX19-binding groove (L93N andK324A) reduce the affinity for PEX19 about 250- and20-fold, respectively. However, the PEX3 protein usedfor Kd-value determination did not include the N-terminaltransmembrane region. This region was proposed earlierto form a possible additional PEX19-binding site using pull-down assays with different splice variants of PEX19 (41).In our CoIP studies, two full-length PEX3 mutants in thePEX19-binding pocket (L93N, A323Q) did not bind PEX19at all, whereas the PEX3 K324A mutant shows a severereduction in PEX19-binding. Thus, an interaction of theN-terminal transmembrane region of PEX3 with PEX19 isunlikely. However, the method used here and in previousstudies (as reviewed in 31) is not suitable to detect lowaffinity binding of the N-terminal first 34 amino acids ofPEX3 and PEX19.
PEX3 is stabilized upon PEX19-binding
Pulse-chase experiments in CHO cells demonstratedthat the presence of PEX19 extends the half-life ofPEX3 (40). Our in vitro findings support this observationas binding to a PEX19-derived peptide leads to asignificant increase in PEX3 stability. The interactionwith PEX19 shields hydrophobic PEX3 residues from theaqueous environment (33). By inserting into the PEX3groove, the PEX19-derived peptide also crosslinks thewalls of the groove, limiting their thermal mobility. Acomparable effect in terms of temperature stability hasbeen observed in major histocompatibility complexes,which also become more stable upon peptide-bindingto a groove that is similar in length to the one present
12 Traffic 2012

Functions of Conserved Regions in PEX3
in sPEX3 (42,43). Remarkably, the mutations in thePEX19-binding region (L93N, K324A) exhibit a reducedcellular expression level. As both mutants are properlyfolded and stable in vitro, a destabilizing effect resultingfrom the amino acids changes within PEX3 can beexcluded. However, additional factors such as proteasesor chaperones that might influence protein integrity andstability in vivo have to be considered. As the mRNAlevels of these PEX3-myc mutants in fibroblasts are200-fold higher than that of endogenous PEX3, thereduced expression levels are unlikely to be linkedto mRNA instability. Furthermore, the third mutant inthe PEX19-binding pocket, PEX3-YFP A323Q, is alsoexpressed at slightly lower levels in normal fibroblastscompared to normal PEX3-YFP. The diminished PEX19-binding may lower the stability of PEX3, and the moreunstable protein may undergo subsequent degradation.Our findings that the PEX3 protein level is reducedin PEX19-knockdown and �PEX19T cells support thisscenario.
PEX3 targeting to peroxisomes
Recent siRNA experiments in HEK293 cells implicatePEX19 as a targeting receptor not only for PMPs, butalso for PEX3 (40). We have employed a similar genesilencing approach using fibroblast cells and find thatin this case, post-translational PEX3 import into existingperoxisomes is independent of PEX19. In our case thePEX19 protein level is at least reduced by 83% andthe import of PMP70 is impaired, while the PEX3-mycimport seems unaffected. Our observation that PEX3-myc is targeted to peroxisomes independently of PEX19is further supported by a dominant peroxisomal stainingpattern in normal fibroblasts for the three PEX3 proteinsimpaired in PEX19-binding (L93N, A323Q, K324A). Theadditional slight cytosolic staining observed for thehighly expressed PEX3-YFP L93N might indicate a sofar unexplained PEX19-dependent influence on PEX3-targeting.
A dual function for the PEX3–PEX19 complex
in peroxisome biogenesis
Our studies also reveal a strikingly reduced complemen-tation level for PEX3 variants carrying mutations in thePEX19-binding groove. PEX3 mutants L93N and A323Qhave lost the ability to complement PEX3-deficient humanfibroblasts, while the corresponding ability of the K324Amutant is lowered by 60% (Figure 6 and Table 3). Theseresults agree with our in vitro PEX19 affinity studies for theL93N, A323Q and K324A mutants (Figure 2 and Table 1).A similar loss in complementing PEX19-deficient cells hasbeen described for a PEX19 mutant in which F29, a residueinvolved in PEX3 binding, is replaced with an alanine (34).In that case, the mutation led to a 300-fold decrease inaffinity compared to original PEX19 (32), consistent withour observation that a 250-fold affinity reduction for theinteraction of the PEX3 L93N mutant with PEX19 is alsosufficient to prevent complementation. In addition, Sato
(34) could show a similar severe effect of the PEX19 L22Amutant in complementing PEX19-deficient cells. Further-more, the L93N and the A323Q mutants are not able toform preperoxisomes to the same extent as PEX3. Thissuggests a crucial function of the PEX3-PEX19 complexalready in the very first steps of peroxisome membranebiogenesis similar as described for yeast cells (13,15,44)and not only in the direct import of proteins into theperoxisomal membrane.
The hydrophobic groove is likely to participate
in PMP import
The PEX3 mutations R53A, M72K, I140Y, L165N, L165Rand I140N*L165N interfere with the reconstitution ofperoxisomes in �PEX3T cells, whereas the H57Amutant is not affected. As all seven mutants areexpressed in normal and PEX3-deficient cells, the reducedcomplementation efficiency is not a result of low proteinexpression levels. Moreover, the seven PEX3 mutantsare still able to generate preperoxisomes at day 1 toa similar extent as normal PEX3. However, in six ofthe mutants these preperoxisomes do not fully developinto functional peroxisomes. The preperoxisomes fail toimport membrane proteins such as PEX14 or PEX13,and the PEX3 proteins are localized to the cytosol orto mitochondria. In contrast to normal PEX3, the PEX3mutants I140Y and R53A retain a reduced number ofpreperoxisomes that are still positive for PEX16, butnegative for PEX14 at day 2. The disappearance of PEX16and the appearance of PMPs such as PEX14 as observedfor normal PEX3 on day 2 therefore appears to be acritical switch for the maturation of preperoxisomes intofunctional peroxisomes. It is likely that the hydrophobicgroove of PEX3 participates in these very early steps ofperoxisome biogenesis. Residue L165 is located at thetop of the hydrophobic groove (Figure 1). Mutation to anasparagine could interfere with PMP import because thehydrophobic character of the groove would be reduced.However, if this leucine is mutated to an arginine, theguanidinium group would project far from the surface, butcould conceivably face to one side, and the remaining,hydrophobic part of the arginine side chain could thensubstitute for the leucine side chain, thus providing ahydrophobic surface that would still allow for some PMPimport. A similar scenario could apply to the M72Kmutation: The lysine side chain is flexible and chargedat its terminus, and would be expected to interferewith binding of the PMP to the hydrophobic groove.However, as most of the lysine side chain is hydrophobic,it might not completely block PMP binding but stillallow for limited favorable interactions. The reducedcomplementation ability for the PEX3 I140Y mutant islikely a result of the large tyrosine ring that would projectinto the hydrophobic groove and render it at least partiallyinaccessible to PMP binding. The arginine at position53 is located at the base of PEX3 in the elongationof the hydrophobic groove (Figure 1). It is temptingto speculate that the reduced complementation abilityof the R53A mutant might be linked to a loss of a
Traffic 2012 13

Schmidt et al.
Figure 10: Model for PMP import into peroxisomes. Rightpanel: Overall surface representation of PEX3 (gray) withthe hydrophobic groove highlighted in shades of green. Thestructures of the N- and C-terminal domains of PEX19 are depictedas cartoon (orange), whereas the flexible part in between is shownas a solid line. The PEX19 C-terminal helix bundle refers to PDBentry 2WL8 (24). PEX3 is anchored in the peroxisomal membranewith its N-terminus (light blue) that mediates the interaction toPEX16 (red). PEX19 binds to the PMP with its C-terminal domain.The N-terminal helix of PEX19 directs the cargo-loaded PEX19-PMP-complex to peroxisomes by binding to PEX3. Left panel:Zoomed view of the hydrophobic groove rotated by 90◦ alongthe indicated horizontal axis. A hydrophobic helix of a putativeinteraction partner is modeled as a blue cylinder that fits well intothe shape of the hydrophobic groove.
long-range electrostatic interaction that involves the R53guanidinium group. In contrast, the PEX3 H57A proteinfeatures a more conservative mutation, perhaps explainingwhy this mutant is not affected in complementationefficiency of �PEX3T cells. Taken together, the analysisof the mutants establishes the hydrophobic groove ofPEX3 as a key contact point for a hydrophobic ligand.While we cannot exclude that this ligand is an unknownprotein, we think it more likely that the PMP itself orthe farnesyl anchor of PEX19 might serve as ligands. Itis perhaps unlikely that mutation of one single residuecan totally abolish interactions with PMPs. However,the significant effects observed with several mutationsclearly delineate an interaction surface that covers mostof the hydrophobic groove. The shape and curvature ofthe groove would be well suited for the accommodationof a single hydrophobic helix contributed by a ligand(Figure 10).
A more detailed model of PMP import
Our results lead to a more informative model of PMPimport by providing support for a specific role of PEX3during PMP insertion (Figure 10). In this model, PEX19directs newly synthesized PMPs in an import-competentform to the peroxisomal membrane. Here, PEX3, which isanchored to the peroxisomal membrane via its N-terminalregion, is able to bind to PEX19 using its cytosolic domain.It is tempting to speculate that structural rearrangementswithin the PEX19–PMP complex upon binding to PEX3allow access to previously covered hydrophobic areas of
the PMP. Such rearrangements might explain why onlyternary complexes comprising PEX3, PEX19 and a PMPhave been described so far (22,37). PEX3 likely adoptsan active role in PMP import by offering the hydrophobicgroove as a docking site for the polypeptide chain thathas to be inserted into the peroxisomal membrane.However, the insertion and the folding process of thePMP into the peroxisomal membrane are currently notwell understood.
Taken together, our data enhance an understandingof peroxisome biogenesis and provide evidence for adual role of the PEX3–PEX19 complex in de novoformation of peroxisomes and in post-translational importof PMPs for the investigated mammalian system.Moreover, engagement of PEX19 leads to significantstabilization of PEX3 in solution. However, interactionwith PEX19 is not essential for PEX3 targeting toperoxisomes, which implies a PEX19-independent post-translational import of PEX3. These findings supportthe coexistence of de novo formation of peroxisomesin peroxisome-deficient cells and the post-translationalimport of membrane proteins in existing peroxisomes thatproliferate and divide independently of de novo formation(18,21,45,46).
Materials and Methods
PlasmidsThe plasmids containing the coding sequence for human sPEX3(HsPEX341-373 (C235S) in pET32a) and for human PEX19 (full-lengthHsPEX19 in pColdI) used for expression in E. coli have been describedpreviously (33). For expression studies in human fibroblasts full-lengthHsPEX3 encoding a C-terminal myc-tag and two truncated versionswith a C-terminal GFP-tag (PEX31-33-GFP and PEX334-373-GFP) (35), or aC-terminal YFP-tag (PEX3-YFP1-373) (30) have been used. The PEX16-mycconstruct was generated by amplifying the ORF with the forward (5′-CCGGTACCAGGAATTCACCATGGAGAAGCTGCGGCTCC-3’, GD119) andreverse primers (5′-CGGAGATCTGCCCCAACTGTAGAAGTAG-3′). The PCRproduct was cloned into the Acc65I and BamHI site of pcDNA3.1Zeo-myc followed by the coding sequence of the c-myc epitope. The plasmidPMP70-myc in pcDNA3 encoding the HsPMP70 (ABCD3) with a C-terminalmyc-epitope is a generous gift of Stephen Gould. YFP-PTS1 refers topEYFP-Peroxi (Clontech).
Site-directed mutagenesisPrimers used for inserting point mutations in sPEX3, PEX3-myc orPEX3-YFP are listed in Table S1. sPEX3 (sPEX3 in pET32a), PEX3-myc (PEX3-myc in pcDNA3.1zeo) and PEX3-YFP (PEX3 in pEYFP-N1)were used as template DNA for PCR amplification. The PCR productswere digested overnight with DpnI (Fermentas) at 37◦C to removeparental methylated DNA. A 2 μL of each PCR sample was transformedinto E. coli DH5α cells and plasmid-containing clones were selectedvia antibiotic resistance on LB-agar plates. Several overnight culturesof LB-medium containing the corresponding antibiotic were inoculatedwith individual clones. DNA preparation was carried out with mi-Plasmid Miniprep Kit (Metabion) according to manufacturer’s instructions.DNA was sent for sequencing (MWG Operon) to test for successfulmutagenesis.
14 Traffic 2012

Functions of Conserved Regions in PEX3
Protein expression and purificationPEX19 full-length protein, sPEX3 and the sPEX3 mutants were expressedand purified as described previously (33). Protein concentrations weredetermined by measurements of absorption at 280 nm using a NanoDropND-1000 (PeqLab). Analytical size exclusion chromatography was carriedout with a SuperdexTM 200 PC 3.2/30 (GE Healthcare) in buffer A(10 mM Na2HPO4, 1.8 mM KH2PO4, 140 mM NaCl, 2.7 mM KCl, 0.5 mM
tris-2-carboxyethyl-phosphine, pH 7.2).
CD analysisCD spectra were recorded with a Jasco J-720 spectropolarimeter. Allpurified proteins were used at concentrations of 10–15 μM in buffer B(5 mM Na2HPO4, 0.9 mM KH2PO4, 70 mM NaCl, 1.35 mM KCl, 0.25 mM tris-2-carboxyethyl-phosphine, pH 7.2). The spectra were recorded eight timesand corrected for the buffer spectrum. Thermal denaturation curves wererecorded on a Jasco J-810 spectropolarimeter. The proteins were dena-tured with a scan rate of 1◦C/min from 20 to 95◦C measuring the CD signalat 208 nm. The purified proteins were used at concentrations of 10–15 μM
in buffer C (10 mM Na2HPO4, 1.8 mM KH2PO4, 5 mM NaCl, pH 7.4).
Affinity measurements using ITCBinding studies between full-length PEX19 and different mutants of sPEX3were carried out with a VP-ITC calorimeter (GE Healthcare). Six PEX3proteins (sPEX3, K324A, E266A, I140N, L165N, I140N*L165N) werepresent in approximately 10 μM concentrations, whereas sPEX3 L93Nwas used at 25 μM concentration. PEX19 was injected stepwise at an8- to 12-fold higher concentration (Table S2). All ITC experiments wereperformed once at 25◦C in buffer A. Data were fitted by least-squaresprocedures according to a one binding site model using Microcal Origin®
version 7.0.
In vitro transcription/translation and CoIPIn vitro transcription and translation experiments were carried out withthe TNT® Coupled Reticulocyte Lysate Systems (Promega) according tothe manufacturer’s instructions. The translation products were labeledwith 35S-methionine (37 TBq/mmol, Hartmann Analytik) for detection.The empty vector pcDNA3.1zeo was used as a negative control. A10 μL of the translated PEX19 was mixed with 10 μL of PEX3-myc,with the corresponding PEX3-myc mutants or with the negative control.The remaining translation products were mixed with SDS-loading bufferand heated to 80◦C for 5 min. The protein mixtures were incubated at30◦C for 1 h, and then subjected to CoIP experiments. For this, 50 μLof Dynabeads® M-280 sheep α-mouse IgG (Invitrogen) was preparedaccording to the manufacturer’s instructions using buffer D (20 mM
HEPES, 110 mM KAc, 5 mM NaAc, 2 mM MgAc, 1 mM EDTA, pH 7.3)and loaded with mouse α-myc antibodies (Cell Signalling, 1:200) in thepresence of 0.1% (w/v) BSA overnight at 4◦C. After washing with bufferE [buffer D with 0.5% (v/v) Triton-X-100], the beads were resuspended in130 μL buffer F [buffer E with protease inhibitor cocktail (Sigma, 1:200)]and the translation mixtures were added. The samples were incubatedunder slight rotation for 2 h at 4◦C and then placed on a magnet toseparate the magnetic beads from the solvent. The supernatant wasremoved, immediately mixed with SDS-loading buffer and heated to80◦C for 5 min. The beads were washed with buffer F, resuspendedin 25 μL SDS-loading buffer and heated to 80◦C for 5 min. The tubeswere then again placed onto the magnet to obtain the eluate. Thesamples were analyzed by10% SDS–PAGE. The gels were incubatedin 0.5 M Na-salicylate (AppliChem) for 20 min, dried onto Whatman paperand exposed to an autoradiography film (BioMax MR film, Kodak) for17–20 h.
Cell culture and transient transfection of human
fibroblastsThe transformed human fibroblasts (GM5657T) were provided by StephenGould (Baltimore). The PEX3-deficient human fibroblasts (47) were a
kind gift by Barbara Paton. The PEX19-deficient cells were obtainedfrom Ron Wanders (Amsterdam). These fibroblasts were transformedwith pRSV-SV40T as described (48) and referred to as �PEX3T and�PEX19T fibroblasts, respectively. Cells were cultured in Dulbecco’smodified Eagle’s medium (DMEM) containing 10% fetal calf serum, 2 mM
glutamine and 0.1 mM (50 mg/L) gentamicin at 37◦C and 8.5% CO2. Ingeneral, cells were seeded onto cover slips 24 h before transfection,whereas �PEX3T cells used for complementation studies were grownin 25 cm2 culture flasks. The cells were transfected transiently withthe corresponding plasmid DNA using jetPEI (PeqLab) according to themanufacturer’s instruction and prepared for indirect immunofluorescencemicroscopy 1 day (GM5756T, �PEX19T , �PEX3T ), 2 days (GM5756T),5 days (�PEX3T ) or 7 days (�PEX3T ) after transfection, respectively, asindicated in the figure legends.
Indirect immunofluorescence microscopyAfter washing three times with Dulbecco’s-PBS (D-PBS, Gibco), cells oncover slips were fixed with formaldehyde (3% in D-PBS) for 20 min.The fixed cells were permeabilized with Triton-X-100 (1% in D-PBS)for 5 min and washed three times with D-PBS. Incubation with primaryantibodies was carried out for 30 min. After extensive washing with D-PBS,cells were incubated with secondary antibodies for 10 min. Following 10additional washing steps with D-PBS, cells were embedded in Mowiol(100 mg/mL, Calbiochem) containing 2.5 mg/mL n-propylgallate (Fluka)prior to analysis by fluorescence microscopy. All steps were carried outat room temperature. Mouse primary antibodies against the C-terminalmyc-tag were purchased from Cell Signalling and diluted 1:200. Rabbitpolyclonal antibodies detecting PEX14 were raised against the first N-terminal 133 amino acids of human PEX14 and were used at a 1:400dilution (49). Monoclonal α-AFP mouse antibodies (3E6) detecting all GFPvariants (1:400 dilution) were purchased from QBiogene. The polyclonalrabbit antibodies against HsPEX16 (JH290) and the sheep antibodiesagainst PMP70 were a kind gift of Stephen Gould and used at a 1:100dilution. Catalase antibodies produced in sheep were obtained frombinding site and used at a 1:100 dilution. Antibodies against PEX13(rabbit) were kindly provided by Marc Fransen (50) and used at a1:250 dilution. Corresponding secondary antibodies were either donkeyor goat IgGs conjugated with AlexaFluor-596 or AlexaFluor-488 (MolecularProbes, Invitrogen). Immunofluorescence images were acquired using aZeiss Axiovert 200 M fluorescence microscope equipped with an AxioPlanApochromat 63×/1.4 oil objective, an AxioPlan Neofluar 100×/1.3 oilobjective and an AxioCam MRm camera in combination with AxioVision4.7.2 software.
Knockdown of endogenous PEX19 with siRNAHuman skin fibroblasts (GM5756T) were electroporated twice eitherwith 1 nmol (20 μM) siRNA against endogenous PEX19 (sense 5′-GAGAUCGCCAGGAGACACUTT-3′); reviewed in 25 or with1 nmol (20 μM)Negative Control siRNA (Qiagen) at an interval of 24 h. For siRNAtransfection cells were resuspended in DMEM and treated with 230 V for25 milliseconds. Later, knockdown cells and control cells were transfectedtransiently either with PEX3-myc or PMP70-myc using jetPEI (PeqLab)24 h after the second electroporation. In both cases, YFP-PTS1 wascotransfected in order to form the ratio of transfection rates. Indirectimmunofluorescence staining was performed 6, 12 and 24 h after plasmidtransfection.
ImmunoblottingThe siRNA-transfected fibroblasts were harvested for immunoblotting 48 hafter the second electroporation. A 20 μg of total protein was separatedon a 12% SDS–PAGE gel and transferred onto a polivinylidene fluoridemembrane (GE healthcare) using a semidry blotting system (Biorad). Blotswere blocked for 2 h with 10% (w/v) non-fat dry milk. The membraneswere decorated for 2 h with polyclonal rabbit α-PEX19 antibodies raisedagainst the recombinant full-length PEX19 (see Protein expression andpurification) at a dilution of 1:10 000 in PBS-ST (10 mM Na2HPO4, 1.8 mM
Traffic 2012 15

Schmidt et al.
KH2PO4, 140 mM NaCl, 2.7 mM KCl, 0.02% (w/v) SDS, 0.1% (v/v) Triton-X-100), polyclonal rabbit α-PEX3.1 antibodies (raised against PEX326-373
C235S purified as reviewed in 33) at a dilution of 1:1000 in PBS-ST,α-Tubulin-β I + II antibodies (monoclonal mouse, Sigma) at a dilution of1:2000 in TBS-T (100 mM Tris, 100 mM NaCl, pH 7.4, 0.1% (v/v) Tween) orα-Tubulin-α antibodies (monoclonal mouse, Sigma) at a dilution of 1:2000in TBS-T. After washing, the membranes were incubated for 1 h eitherwith horseradish peroxidase-conjugated goat α-mouse IgGs or horseradishperoxidase-conjugated goat α-rabbit IgGs at a 1:15 000 dilution (Sigma). Theblots were washed and developed using enhanced chemiluminescenceWestern Blotting substrate (Thermo Scientific). All steps were carried outat room temperature.
Quantification of PEX3 mRNA levelsHuman skin fibroblasts were transfected with PEX3-myc or themutated PEX3-myc variants with jetPEI and harvested 24 h aftertransfection. Total RNA was extracted (RNA spin Kit, PrepEase, USB)and subjected to cDNA synthesis (iScript, BioRad) according to themanufacturer’s instruction. Quantitative real-time PCR was performedusing Maxima SYBR Green/ROX qPCR Master Mix (Fermentas) andprimers for PEX3 (forward 5′-GGCTGAGTTCTTTCGACCTACTG-3′, reverse5′-TCACTGCAAACTGAATGGATCTG-3′) or Glyceraldehyde 3-phosphatedehydrogenase (GAPDH, forward 5′-CATCAAGAAGGTGGTGAAGCAG-3′,reverse 5′-CAAAGTGGTCGTTGAGGGGCAATG-3′). The PCR reactionswere carried out in the StepOne Plus Real-time PCR System (AppliedBiosystems) under following conditions: 50◦C for 2 min, 95◦C for 10 min,40 cycles at 95◦C for 15 seconds and 60◦C for 1 min followed by meltingcurve analysis. Data were collected at the 60◦C step. The mRNA levels ofPEX3-myc and PEX3-myc mutants compared to endogenous PEX3 werecalculated via the 2−��ct method (51).
Acknowledgments
We thank Karin Steiger for technical assistance, our student HannaLeins for assistance in cell culture, Petra Krensel for establishing siRNAtechnology and Katrin Bagner and Birgit Gurke for providing PEX16plasmids. We are grateful to Stephen Gould, Ron Wanders, Barbara Patonand Marc Fransen for providing cell lines, plasmids and antibodies. Theauthors declare no conflicts of interest.
Supporting Information
Additional Supporting Information may be found in the online version ofthis article:
Figure S1: Folding studies and stability analysis of different sPEX3
mutants. A) Circular dichroism (CD) spectroscopy of all sPEX3 mutantsshow an α-helical structural content, indicating no change in the overall foldof the protein compared to sPEX3. CD spectra were recorded with 12 μM
protein solutions and corrected with the buffer spectrum. B) Thermalstability analysis for sPEX3 mutants. Proteins were heated stepwiseto 95◦C with a scan rate of 1◦C/min. The CD signal at 208 nm wasrecorded during the whole denaturation process. Curves are displayedusing a smoothing factor of 5. The inflection point represents themelting temperature of the protein. For all mutants, this melting pointis somewhat higher than for sPEX3 (black), implicating no loss in stabilityupon introducing the mutations in sPEX3.
Figure S2: Affinity measurements of complex formation between
different sPEX3 mutants and PEX19. Raw data for binding of sPEX3I140N to PEX19 (A) and sPEX3 E266A to PEX19 (B). C) Integrated heatvalues for the different ITC experiments showing fits to a single bindingisotherm for the respective titrations. The thermodynamic parametersfor each sample are listed in Table 1. D) Analytical size exclusionchromatography for sPEX3 + PEX19, L93N + PEX19, K324A + PEX19 andPEX19 alone are shown. It is known, that PEX19 exists as a monomerin solution but elutes at higher molecular weight mass in gel filtration
(32). Here, PEX19 (molecular mass: 32.8 kDa) elutes at an apparentmolecular mass of 115 kDa due to its extended conformation. Thecomplex sPEX3 + PEX19 represents a molecular mass of 160 kDa,corresponding to the total of the two monomers building a 1:1 complex.The second peak in the profile of the complex can be assigned to anexcess of monomeric sPEX3 (molecular mass: 37.5 kDa). The elutionprofile for sPEX3 K324A + PEX19 overlays with the complex profile forsPEX3 + PEX19. Two single peaks, corresponding to the individual proteinsPEX19 and sPEX3 L93N, characterize the elution profile of a mixture ofsPEX3 L93N and PEX19.
Figure S3: Peroxisomal localization of different PEX3-myc mutants in
human fibroblasts. A) Normal fibroblasts (GM5756T) were transfectedwith plasmids coding for the different PEX3-myc mutants E266A, D275A,I140N and I140N*L165N. The cellular localization of the expressedproteins was detected with α-myc antibodies two days after transfection.Peroxisomes were visualized with α-PEX14 antibodies. These PEX3-myc mutants reveal a peroxisomal distribution within the cell validatedby colocalization with PEX14. Scale bar: 10 μm. B) mRNA levels ofendogenous PEX3 and the corresponding PEX3-myc mutants in relation toPEX3-myc, which was set to 1. Values represent the mean and range oftwo experiments.
Figure S4: Complementation of PEX3-deficient human fibroblasts.
�PEX3T cells were transfected with plasmids coding for the differentPEX3-myc mutants E266A, D275A, I140N, I140N*L165N and C235S.The ability for complementation of �PEX3T cells was determined 7 daysafter transfection using α-PEX14 antibodies to detect newly formedperoxisomes. The import competence of these organelles was tested withα-catalase antibodies. The PEX3-myc mutants are still able to complement�PEX3T cells, which is validated by the colocalization of PEX14 withcatalase-positive punctate structures. Scale bar: 10 μm.
Table S1: Primers for inserting point mutations in sPEX3, PEX3-myc andPEX3-YFP.
Table S2: Protein concentrations used for ITC experiments.
Please note: Wiley-Blackwell are not responsible for the content orfunctionality of any supporting materials supplied by the authors.Any queries (other than missing material) should be directed to thecorresponding author for the article.
References
1. Schrader M, Fahimi HD. The peroxisome: still a mysterious organelle.Histochem Cell Biol 2008;129:421–440.
2. Wanders RJ, Waterham HR. Biochemistry of mammalian peroxi-somes revisited. Annu Rev Biochem 2006;75:295–332.
3. Steinberg SJ, Dodt G, Raymond GV, Braverman NE, Moser AB,Moser HW. Peroxisome biogenesis disorders. Biochim Biophys Acta2006;1763:1733–1748.
4. Wanders RJ, Waterham HR. Peroxisomal disorders I: biochemistryand genetics of peroxisome biogenesis disorders. Clin Genet2005;67:107–133.
5. Wanders RJ, van Grunsven EG, Jansen GA. Lipid metabolism inperoxisomes: enzymology, functions and dysfunctions of the fattyacid alpha- and beta-oxidation systems in humans. Biochem SocTrans 2000;28:141–149.
6. Weller S, Gould SJ, Valle D. Peroxisome biogenesis disorders. AnnuRev Genomics Hum Genet 2003;4:165–211.
7. Sacksteder KA, Gould SJ. The genetics of peroxisome biogenesis.Annu Rev Genet 2000;34:623–652.
8. Muntau AC, Mayerhofer PU, Paton BC, Kammerer S, Roscher AA.Defective peroxisome membrane synthesis due to mutations inhuman PEX3 causes Zellweger syndrome, complementation groupG. Am J Hum Genet 2000;67:967–975.
9. Ghaedi K, Honsho M, Shimozawa N, Suzuki Y, Kondo N, Fujiki Y. PEX3is the causal gene responsible for peroxisome membrane assembly-defective Zellweger syndrome of complementation group G. Am JHum Genet 2000;67:976–981.
16 Traffic 2012

Functions of Conserved Regions in PEX3
10. South ST, Gould SJ. Peroxisome synthesis in the absence ofpreexisting peroxisomes. J Cell Biol 1999;144:255–266.
11. Matsuzono Y, Kinoshita N, Tamura S, Shimozawa N, Hamasaki M,Ghaedi K, Wanders RJ, Suzuki Y, Kondo N, Fujiki Y. Human PEX19:cDNA cloning by functional complementation, mutation analysis in apatient with Zellweger syndrome, and potential role in peroxisomalmembrane assembly. Proc Natl Acad Sci USA 1999;96:2116–2121.
12. Honsho M, Tamura S, Shimozawa N, Suzuki Y, Kondo N, Fujiki Y.Mutation in PEX16 is causal in the peroxisome-deficient Zellwegersyndrome of complementation group D. Am J Hum Genet 1998;63:1622–1630.
13. Lam SK, Yoda N, Schekman R. A vesicle carrier that mediatesperoxisome protein traffic from the endoplasmic reticulum. Proc NatlAcad Sci USA 2010;107:21523–21528.
14. Tam YY, Fagarasanu A, Fagarasanu M, Rachubinski RA. Pex3p initiatesthe formation of a preperoxisomal compartment from a subdomain ofthe endoplasmic reticulum in Saccharomyces cerevisiae. J Biol Chem2005;280:34933–34939.
15. Hoepfner D, Schildknegt D, Braakman I, Philippsen P, Tabak HF.Contribution of the endoplasmic reticulum to peroxisome formation.Cell 2005;122:85–95.
16. Motley AM, Ward GP, Hettema EH. Dnm1p-dependent peroxisomefission requires Caf4p, Mdv1p and Fis1p. J Cell Sci 2008;121:1633–1640.
17. van der Zand A, Braakman I, Tabak HF. Peroxisomal membraneproteins insert into the endoplasmic reticulum. Mol Biol Cell 2010;21:2057–2065.
18. Ma C, Agrawal G, Subramani S. Peroxisome assembly: matrix andmembrane protein biogenesis. J Cell Biol 2011;193:7–16.
19. Toro AA, Araya CA, Cordova GJ, Arredondo CA, Cardenas HG, MorenoRE, Venegas A, Koenig CS, Cancino J, Gonzalez A, Santos MJ.Pex3p-dependent peroxisomal biogenesis initiates in the endoplasmicreticulum of human fibroblasts. J Cell Biochem 2009;107:1083–1096.
20. Kim PK, Mullen RT, Schumann U, Lippincott-Schwartz J. The origin andmaintenance of mammalian peroxisomes involves a de novo PEX16-dependent pathway from the ER. J Cell Biol 2006;173:521–532.
21. Huybrechts SJ, Van Veldhoven PP, Brees C, Mannaerts GP, Los GV,Fransen M. Peroxisome dynamics in cultured mammalian cells. Traffic2009;10:1722–1733.
22. Matsuzono Y, Fujiki Y. In vitro transport of membrane proteins toperoxisomes by shuttling receptor Pex19p. J Biol Chem 2006;281:36–42.
23. Rottensteiner H, Kramer A, Lorenzen S, Stein K, Landgraf C, Volkmer-Engert R, Erdmann R. Peroxisomal membrane proteins containcommon Pex19p-binding sites that are an integral part of theirtargeting signals. Mol Biol Cell 2004;15:3406–3417.
24. Schueller N, Holton SJ, Fodor K, Milewski M, Konarev P, StanleyWA, Wolf J, Erdmann R, Schliebs W, Song YH, Wilmanns M.The peroxisomal receptor Pex19p forms a helical mPTS recognitiondomain. EMBO J 2010;29:2491–2500.
25. Jones JM, Morrell JC, Gould SJ. PEX19 is a predominantly cytosolicchaperone and import receptor for class 1 peroxisomal membraneproteins. J Cell Biol 2004;164:57–67.
26. Shibata H, Kashiwayama Y, Imanaka T, Kato H. Domain architectureand activity of human Pex19p, a chaperone-like protein for intracellulartrafficking of peroxisomal membrane proteins. J Biol Chem 2004;279:38486–38494.
27. Fransen M, Vastiau I, Brees C, Brys V, Mannaerts GP, Van VeldhovenPP. Analysis of human Pex19p’s domain structure by pentapeptidescanning mutagenesis. J Mol Biol 2005;346:1275–1286.
28. Matsuzono Y, Matsuzaki T, Fujiki Y. Functional domain mapping ofperoxin Pex19p: interaction with Pex3p is essential for function andtranslocation. J Cell Sci 2006;119:3539–3550.
29. Fang Y, Morrell JC, Jones JM, Gould SJ. PEX3 functions as a PEX19docking factor in the import of class I peroxisomal membrane proteins.J Cell Biol 2004;164:863–875.
30. Muntau AC, Roscher AA, Kunau WH, Dodt G. The interaction betweenhuman PEX3 and PEX19 characterized by fluorescence resonanceenergy transfer (FRET) analysis. Eur J Cell Biol 2003;82:333–342.
31. Fujiki Y, Matsuzono Y, Matsuzaki T, Fransen M. Import of peroxisomalmembrane proteins: the interplay of Pex3p- and Pex19p-mediatedinteractions. Biochim Biophys Acta 2006;1763:1639–1646.
32. Sato Y, Shibata H, Nakano H, Matsuzono Y, Kashiwayama Y,Kobayashi Y, Fujiki Y, Imanaka T, Kato H. Characterization of theinteraction between recombinant human peroxin Pex3p and Pex19p:identification of Trp-104 in Pex3p as a critical residue for theinteraction. J Biol Chem 2008;283:6136–6144.
33. Schmidt F, Treiber N, Zocher G, Bjelic S, Steinmetz MO, Kalbacher H,Stehle T, Dodt G. Insights into peroxisome function from the structureof PEX3 in complex with a soluble fragment of PEX19. J Biol Chem2010;285:25410–25417.
34. Sato Y, Shibata H, Nakatsu T, Nakano H, Kashiwayama Y, Imanaka T,Kato H. Structural basis for docking of peroxisomal membrane proteincarrier Pex19p onto its receptor Pex3p. EMBO J 2010;29:4083–4093.
35. Soukupova M, Sprenger C, Gorgas K, Kunau WH, Dodt G. Identificationand characterization of the human peroxin PEX3. Eur J Cell Biol 1999;78:357–374.
36. Kammerer S, Holzinger A, Welsch U, Roscher AA. Cloning andcharacterization of the gene encoding the human peroxisomalassembly protein Pex3p. FEBS Lett 1998;429:53–60.
37. Pinto MP, Grou CP, Alencastre IS, Oliveira ME, Sa-Miranda C, FransenM, Azevedo JE. The import competence of a peroxisomal membraneprotein is determined by Pex19p before the docking step. J Biol Chem2006;281:34492–34502.
38. Diestelkotter P, Just WW. In vitro insertion of the 22-kD peroxisomalmembrane protein into isolated rat liver peroxisomes. J Cell Biol1993;123:1717–1725.
39. Ghaedi K, Tamura S, Okumoto K, Matsuzono Y, Fujiki Y. The peroxinpex3p initiates membrane assembly in peroxisome biogenesis. MolBiol Cell 2000;11:2085–2102.
40. Matsuzaki T, Fujiki Y. The peroxisomal membrane protein importreceptor Pex3p is directly transported to peroxisomes by a novelPex19p- and Pex16p-dependent pathway. J Cell Biol 2008;183:1275–1286.
41. Mayerhofer PU, Kattenfeld T, Roscher AA, Muntau AC. Two splicevariants of human PEX19 exhibit distinct functions in peroxisomalassembly. Biochem Biophys Res Commun 2002;291:1180–1186.
42. Fahnestock ML, Tamir I, Narhi L, Bjorkman PJ. Thermal stabilitycomparison of purified empty and peptide-filled forms of a class IMHC molecule. Science 1992;258:1658–1662.
43. Bouvier M, Wiley DC. Structural characterization of a soluble andpartially folded class I major histocompatibility heavy chain/beta 2 mheterodimer. Nat Struct Biol 1998;5:377–384.
44. Agrawal G, Joshi S, Subramani S. Cell-free sorting of peroxisomalmembrane proteins from the endoplasmic reticulum. Proc Natl AcadSci USA 2011;108:9113–9118.
45. Delille HK, Agricola B, Guimaraes SC, Borta H, Luers GH, Fransen M,Schrader M. Pex11pbeta-mediated growth and division of mammalianperoxisomes follows a maturation pathway. J Cell Sci 2010;123:2750–2762.
46. Koch J, Pranjic K, Huber A, Ellinger A, Hartig A, Kragler F, Brocard C.PEX11 family members are membrane elongation factors that coordi-nate peroxisome proliferation and maintenance. J Cell Sci 2010;123:3389–3400.
47. Poulos A, Christodoulou J, Chow CW, Goldblatt J, Paton BC, Orii T,Suzuki Y, Shimozawa N. Peroxisomal assembly defects: clinical,pathologic, and biochemical findings in two patients in a newlyidentified complementation group. J Pediatr 1995;127:596–599.
48. Dodt G, Braverman N, Wong C, Moser A, Moser HW, Watkins P,Valle D, Gould SJ. Mutations in the PTS1 receptor gene, PXR1, definecomplementation group 2 of the peroxisome biogenesis disorders.Nat Genet 1995;9:115–125.
49. Will GK, Soukupova M, Hong X, Erdmann KS, Kiel JA, Dodt G, KunauWH, Erdmann R. Identification and characterization of the humanorthologue of yeast Pex14p. Mol Cell Biol 1999;19:2265–2277.
50. Fransen M, Wylin T, Brees C, Mannaerts GP, Van Veldhoven PP.Human pex19p binds peroxisomal integral membrane proteins atregions distinct from their sorting sequences. Mol Cell Biol 2001;21:4413–4424.
51. Livak KJ, Schmittgen TD. Analysis of relative gene expression datausing real-time quantitative PCR and the 2−��CT. Methods 2001;25:402–408.
Traffic 2012 17





























