Embed Size (px)
Citation preview

Review article
The relationship between spotted fever group Rickettsiaeand Ixodid ticks
Cristina SOCOLOVSCHI, Oleg MEDIANNIKOV, Didier RAOULT, Philippe PAROLA*
Unite de Recherche en Maladies Infectieuses et Tropicales Emergentes (URMITE), UMR CNRS-IRD 6236, WHOCollaborative Center for Rickettsial diseases and other arthropod-borne bacterial diseases,
Faculte de Medecine, 27 Bd Jean Moulin, 13385 Marseille Cedex 5, France
(Received 9 November 2008; accepted 9 April 2009)
Abstract – Spotted fever group Rickettsiae are predominantly transmitted by ticks. Rickettsiae havedeveloped many strategies to adapt to different environmental conditions, including those within theirarthropod vectors and vertebrate hosts. The tick-Rickettsiae relationship has been a point of interest formany researchers, with most studies concentrating on the role of ticks as vectors. Unfortunately, lessattention has been directed towards the relationship of Rickettsiae with tick cells, tissues, and organs. Thisreview summarizes our current understanding of the mechanisms involved in the relationship between ticksand Rickettsiae and provides an update on the recent methodological improvements that have allowed forcomprehensive studies at the molecular level.
ticks / spotted fever group Rickettsiae / vector / reservoir
Table of contents
1. Introduction........................................................................................................................................... 21.1. Primary infection of ticks with Rickettsiae ................................................................................. 2
1.1.1. Gut barrier and initial contact with tick cells ................................................................. 51.1.2. Hemolymph...................................................................................................................... 51.1.3. Salivary glands................................................................................................................. 71.1.4. Ovaries ............................................................................................................................. 7
1.2. Ticks as a life-long reservoir of Rickettsiae................................................................................ 72. How to live together............................................................................................................................. 9
2.1 Rickettsial challenges for ticks..................................................................................................... 92.2. Tick challenges for Rickettsiae.................................................................................................... 11
3. Interference ........................................................................................................................................... 124. Methods of research into tick-Rickettsiae interactions ........................................................................ 13
4.1. Experimental models ................................................................................................................... 134.2. Role of molecular tools in understanding tick-Rickettsiae interactions ..................................... 15
5. Conclusion and perspectives ................................................................................................................ 16
* Corresponding author: [email protected]
Vet. Res. (2009) 40:34DOI: 10.1051/vetres/2009017
� INRA, EDP Sciences, 2009
www.vetres.org
Article published by EDP Sciences

1. INTRODUCTION
Rickettsiae are Gram-negative, obligate intra-cellular bacteria in the family Rickettsiaceaeand order Rickettsiales. The spotted fever group(SFG) unites a phylogenetically well-definedclade of Rickettsiae that are distinct from otherspecies and that have a life cycle involvingarthropods, mainly ticks [63]. The SFGincludes a number of pathogenic organisms thatcause so-called tick-borne (TB) rickettsioses inhumans. Among them are Rickettsia rickettsii(Rocky Mountain spotted fever, RMSF),R. conorii conorii (Mediterranean spotted fever,MSF), R. conorii israelensis (Israeli spottedfever), R. conorii caspia (Astrakhan spottedfever), R. conorii indica (Indian tick typhusRickettsia), R. africae (African tick-bite fever,ATBF), R. heilongjiangensis (Far-eastern tick-borne rickettsiosis), R. australis (Queenslandtick typhus), R. slovaca (Tick-borne lympha-denopathy and Dermacentor-borne necrosiserythema lymphadenopathy, TIBOLA/DEBO-NEL), R. sibirica sibirica (North Asian ticktyphus or Siberian tick typhus), R. sibiricamongolitimonae (Lymphangitis-associated rick-ettsiosis), R. honei (Flinders Island spottedfever), R. japonica (Japanese or Orientalspotted fever), R. parkeri, R. aeschlimannii,R. massiliae, and R. raoultii. Numerous Rickett-siae are regularly associated with ticks and havebeen called symbionts (literally ‘‘livingtogether’’), microsymbionts, or endosymbionts(living in endocellular symbiosis) by entomolo-gists, ecologists, or endocytobiologists. How-ever, their potential for pathogenicity is stillunknown [62].
The ecology of SFG Rickettsiae has not beendefinitively elucidated. Some SFG Rickettsiaeare thought to circulate in enzootic or epizooticcycles between wild vertebrates and arthropodvectors [50, 89]. Ticks are usually thought tobe the main reservoir and vectors of SFG Rick-ettsiae in nature, due to the ability of Rickettsiaeto survive perpetually in ticks and to be trans-mitted transstadially and transovarially. How-ever, this has been demonstrated only for afew tick-borne Rickettsiae (Tab. I). Humansare only occasional hosts for ticks and rarelyplay a role in the subsequent transmission of
bacteria. Therefore, induced human rickettsios-es should be viewed as an accidental ecosystemchange for the Rickettsiae, and the humanshould be viewed as a ‘‘dead end’’ host, whichplays no role in the maintenance of these bacte-ria in nature.
Ticks are currently considered to be secondonly to mosquitoes as vectors of human infec-tious diseases worldwide. All of the nearly900 known species of ticks require blood fortheir development and reproduction, and theyparasitize every class of vertebrates in almostevery region of the world. Two families ofticks are of medical significance: Ixodidae (hardticks) and Argasidae (soft ticks). To date, mostticks infected with SFG Rickettsiae belong tothe Ixodidae family. Ixodid ticks feed oncewithin each stage but for a relatively long per-iod (several days), during which the tickremains strongly attached to the host. Thisblood-feeding may involve a great variety ofvertebrates that occupy very diverse habitats.Because the tick’s bite is usually painless, tickattachment may go unnoticed for several days,consequently enhancing the vector potential ofticks [62].
The tick-Rickettsiae relationship was a focusof interest for many pioneering rickettsiologists,with most of the early studies concentrating onthe role of ticks as vectors. At the beginning ofthe 20th century, the wood tick Dermacentorandersoni was found to be involved in thetransmission of R. rickettsii [77]. In the 1930s,the role of Rhipicephalus sanguineus, thebrown dog tick, was demonstrated in the trans-mission of R. conorii (Fig. 1) [10].
This review highlights the relationshipbetween ticks and Rickettsiae, with regards tothe features of Rickettsiae adaptation to ticksand transmission to the ticks’ progeny. Thedevelopment of genomic tools and the benefitsor deleterious effects of rickettsial infection,especially in terms of gene expression modifica-tion, will also be discussed.
1.1. Primary infection of ticks with Rickettsiae
The initial infection of ticks with Rickettsiaecan occur via the gut when bacteria-freeticks feed on rickettsemic hosts (Fig. 2). This
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 2 of 20 (page number not for citation purpose)

Figure 1. Rhipicephalus sanguineus (brown dog tick), the main vector of MSF and occasional vector ofRMSF. From left to right: female, male, nymph, larva, and egg. Bar scale, 1 mm.
Table I. The prevalence of infected ticks in nature and the study of vertical and transovarian transmission ofSFG Rickettsiae (transovarial transmission rate (TOT): proportion of infected females giving rise to at leastone positive egg or larva).
Rickettsia Tick species Infection rate (%) TOT (%)
R. conorii Rh. sanguineus 0–1.4 100*R. rickettsii D. andersoni 0.26–1.5 100*
D. variabilis 0.0143–1.3 30–40R. africae A. hebraeum 20–30 100
A. variegatum 27–100 YesR. massiliae Rh. turanicus 0.7–50 100R. slovaca D. marginatus 7.2–40.6 100R. rhipicephali Dermacentor sp. 1.26–1.32 38–100R. sibirica D. nuttalli 12 100R. bellii Amblyomma sp. 1.4–17.4 NS
I. loricatus 60.9 100Dermacentor sp. 1.3–2.2 NS
R. helvetica I. ricinus 0.6–46.45 100R. peacockii D. andersoni 66 73.3R. monacensis I. ricinus 2.4–52.9 NSR. aeschlimannii H. marginatum marginatum 1.8–57.9 YesR. amblyommii Amblyomma sp. 3.7–23.6 YesR. raoultii D. reticulatus 5.6–23 NS
D. marginatus 22.5–83.3 86.4–100
NS: Not studied.* TOT in naturally infected ticks, but no or low transmission in laboratory-infected ticks. TOT forR. conorii was studied only for the fifth generation. The duration of the infection in the ticks is unknown.
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 3 of 20

requires sufficient blood levels of Rickettsiaein free-living vertebrates, which may act asreservoirs for Rickettsiae [74]. For example,R. rickettsii was first isolated from small mam-mals known as meadow voles (Microtus penn-sylvanicus) [50, 91]. A decade later, R. rickettsiiwas isolated from eight other species ofmammals, including a pine vole (Pitymys pine-torum), a white-footed mouse (Peromyscusleucopus), a cotton rat (Sigmodon hispidus),cottontail rabbits (Sylvilagus floridanus), anopossum (Didelphis marsupialis virginiana),chipmunks (Eutamias amoenus), a snowshoehare (Lepus americanus), and golden-mantledground squirrels (Spermophilus lateralis tesco-rum) [50]. The more blood the tick ingests,the greater the level of bacteria in the blood-stream, and the longer the tick remains attached,the higher the probability of infection byRickettsiae. However, the relative importance
of this mode of infection in nature is unknownfor most tick-borne Rickettsiae.
Ticks may also acquire Rickettsiae byco-feeding which occurs when several ticksfeed next to each other on the same host. In thiscase, the direct spread of bacteria from aninfected tick to an uninfected tick can occurduring feeding at closely situated bite sites, asdemonstrated with R. rickettsii and D. ander-soni [65]. Co-feeding and/or sexual transmis-sion of R. massiliae was demonstrated in amale Rh. turanicus feeding on a rabbit withRh. sanguineus females [49]. However, femalesthat had been infected by sexual transmission/co-feeding did not transmit the bacteria transov-arially. The transmission of R. rickettsii frominfected male ticks to non-infected female tickshas also been described, but this process is unli-kely to significantly propagate rickettsial infec-tion in tick lineages, since venereally infected
Figure 2. Life cycle of Amblyomma variegatum, modified from [83].
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 4 of 20 (page number not for citation purpose)

females do not appear to transmit the bacteriatransovarially [29]. However, more data areneeded to confirm rickettsial transfer duringcopulation.
1.1.1. Gut barrier and initial contactwith tick cells
The host blood ingested by a tick flowsthrough the canal formed by the cheliceraeand the hypostome, through the pharyngealcavity, and through a short esophagus into themid-gut and its diverticula [72]. Digestioninvolves the lysis of erythrocytes within thegut lumen, ingestion of hemolysate by thedigestive cells, and intracellular digestion ofprotein and lipids [54]. Here, the first contactwith tick cells occurs. Rickettsiae interact withyet unknown cellular surface receptors andescape the tick’s immune responses, whichprobably differ from those in vertebrates. Allof the ticks tested at 72 h post-experimentalfeeding were infected with R. montanensis[18]. Recently, electron microscopy showed atleast two forms of R. peacockii in the cytosolof infected cells. One is present in the cytosolof tick cells, and the second within the auto-phagolysosomes [38]. However, it is not knownwhether these findings also apply to other tick/SFG Rickettsiae interactions. A phospholipasebelonging to the putative phospholipase D(PLD) protein superfamily might be criticalfor the internalization and intracellular life ofRickettsiae. In rickettsial pathogenicity, PLD ishypothesized to exert functions attributed tophospholipase A2 (PLA2): to mediate entryinto the host cell, to escape from the phago-some, and to facilitate injury to host cells[76]. SFG Rickettsiae also exploit the host cellactin cytoskeleton to promote motility andcell-to-cell spreading [31, 90]. A rickettsial pro-tein called RickA promotes the nucleation ofactin monomers via the Arp2/3 complex. Thesefactors are expressed on the bacterial surfacebut lack signal sequences, and therefore, theirmode of secretion is unknown [28, 36]. Thedifferential actin-based motility of R. raoultiiand R. conorii observed in L929 and Vero cellssuggests that the expression of RickA is
not a sufficient condition to promote actinpolymerization in vivo and that another factorapart from RickA may be involved in this pro-cess [7]. Preliminary results assessed by differ-ential-display PCR have shown modificationsin tick gene expression in Rickettsia-infectedD. variabilis ticks [45]. In particular, the expres-sion levels of tubulin alpha-chain and V-ATPaseassociated with clathrin-coated vesicles wereup-regulated. V-ATPase is known to facilitateprotein sorting, receptor-mediated endocytosisby the cell, and the entry of a number of enve-lope viruses and bacterial toxins, including theinfluenza virus and anthrax toxin [33].
The tick mid-gut epithelial cells supporthighly replicative Rickettsiae without alteringthe host cell ultrastructure. After crossing thedigestive tract barrier, the bacteria penetrate intothe body cavity of arthropods and survive andmultiply there for a long time, virtually forthe entire life of the vector, as has been shownfor ‘‘Candidatus Rickettsia tarasevichiae’’ innaturally infected I. persulcatus ticks [69].The role of the mid-gut in rickettsial infectionis vital to successful rickettsial disseminationvia both saliva secretion and tick feces. Theexcreta during feeding consist of black hematinand other undigested residues from the mid-gut[87]. The purpose of excretion during feeding isto remove liquids, which are required for pro-ducing more saliva, and also to retain the lipidsfrom cell membranes. R. slovaca was identifiedin the feces of naturally infected D. marginatus,and successfully isolated. R. massiliae andR. conorii have also been detected in the fecesof infected ticks, using molecular tools andimmunofluorescence assays [49, 73, 84]. How-ever, the role of feces as an efficient source ofinfection is unknown, although the weak trans-mission of R. rickettsii to guinea pigs was foundto occur through this route [65, 72].
1.1.2. Hemolymph
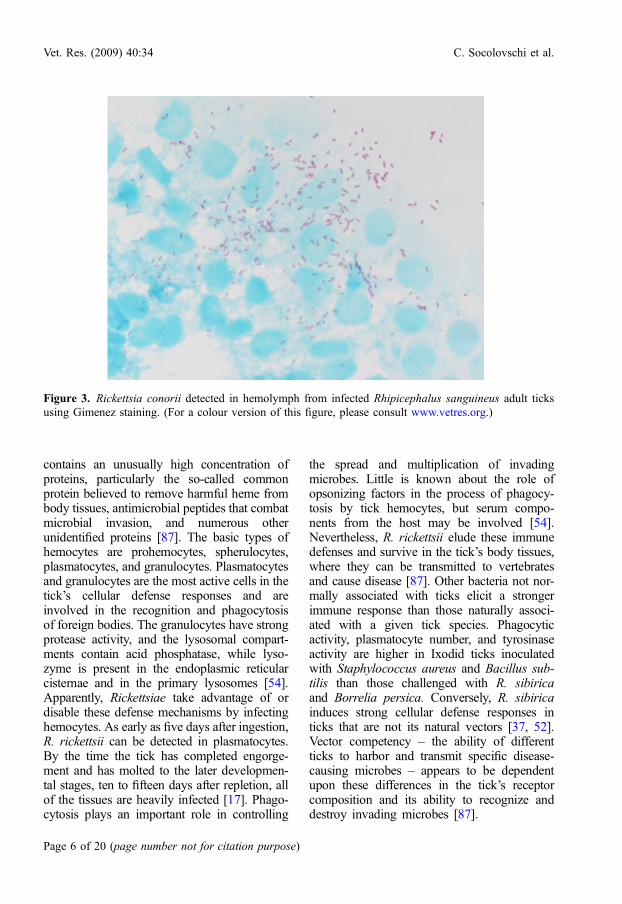
Rickettsiae that escape from the mid-gutinvade hemocytes, gaining access to virtuallyall tissues and organs and causing a gene-ralized infection, as shown for R. conorii inRh. sanguineus [79] (Fig. 3). The hemolymph
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 5 of 20
contains an unusually high concentration ofproteins, particularly the so-called commonprotein believed to remove harmful heme frombody tissues, antimicrobial peptides that combatmicrobial invasion, and numerous otherunidentified proteins [87]. The basic types ofhemocytes are prohemocytes, spherulocytes,plasmatocytes, and granulocytes. Plasmatocytesand granulocytes are the most active cells in thetick’s cellular defense responses and areinvolved in the recognition and phagocytosisof foreign bodies. The granulocytes have strongprotease activity, and the lysosomal compart-ments contain acid phosphatase, while lyso-zyme is present in the endoplasmic reticularcisternae and in the primary lysosomes [54].Apparently, Rickettsiae take advantage of ordisable these defense mechanisms by infectinghemocytes. As early as five days after ingestion,R. rickettsii can be detected in plasmatocytes.By the time the tick has completed engorge-ment and has molted to the later developmen-tal stages, ten to fifteen days after repletion, allof the tissues are heavily infected [17]. Phago-cytosis plays an important role in controlling
the spread and multiplication of invadingmicrobes. Little is known about the role ofopsonizing factors in the process of phagocy-tosis by tick hemocytes, but serum compo-nents from the host may be involved [54].Nevertheless, R. rickettsii elude these immunedefenses and survive in the tick’s body tissues,where they can be transmitted to vertebratesand cause disease [87]. Other bacteria not nor-mally associated with ticks elicit a strongerimmune response than those naturally associ-ated with a given tick species. Phagocyticactivity, plasmatocyte number, and tyrosinaseactivity are higher in Ixodid ticks inoculatedwith Staphylococcus aureus and Bacillus sub-tilis than those challenged with R. sibiricaand Borrelia persica. Conversely, R. sibiricainduces strong cellular defense responses inticks that are not its natural vectors [37, 52].Vector competency – the ability of differentticks to harbor and transmit specific disease-causing microbes – appears to be dependentupon these differences in the tick’s receptorcomposition and its ability to recognize anddestroy invading microbes [87].
Figure 3. Rickettsia conorii detected in hemolymph from infected Rhipicephalus sanguineus adult ticksusing Gimenez staining. (For a colour version of this figure, please consult www.vetres.org.)
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 6 of 20 (page number not for citation purpose)

1.1.3. Salivary glands
Salivary gland secretions facilitate tickfeeding and are important vehicles for the trans-mission of tick-borne pathogens to the verte-brate host. In naturally infected D. marginatus,R. slovacamultiplied in all of the type I-III aciniand also in the duct cells. An ultrastructuralstudy showed that in the salivary glands of ticksexperimentally infected with R. conorii, growthoccurred preferentially in central, peripheral,and interstitial acinar cells [79]. Therefore, thesalivary glands play an active role in Rickettsiapropagation. They are not simply organs inwhich Rickettsiae are collected before theirrelease into the host via saliva. The presence ofa large number of R. honei-containing nucleo-plasmic vacuoles of various sizeswas an unusualfinding in the salivary glands andmid-gut epithe-lium of the reptilian tick Aponomma hydrosauri,since the presence of such vacuoles is usuallyconsidered to be characteristic of a typhus groupRickettsia [92]. R. rickettsii can present distinctultrastructural forms in the salivary glands,depending on the physiological state of theinfected tick (starved or fully engorged). Thisphenomenon, known as ‘‘reactivation’’, mayreflect an adaptation of the pathogen to the vec-tor’s physiological rhythm [30].
1.1.4. Ovaries
Rickettsiae probably invade the oocytes dur-ing active oogenesis following the nymphal andadult blood meals (Fig. 4). Oviposition occursafter the completion of blood-feeding. Ultra-structural examination of ovaries infected withR. honei revealed that each oocyte and immatureegg examined was infected [92]. Ticks experi-mentally infected as adults with R. rickettsiiand R. montanensis had ovarial tissues with asmany as 2.5 · 107 Rickettsiae following ovipo-sition [56]. Rickettsial development in the ovar-ian interstitial cells of nymphal ticks, and lateralso within the oogonia and oocytes, leads totransovarian transmission [16, 17]. Recently,Gimenez staining and electron microscopyrevealed the presence of R. conorii in the sali-vary glands and ovaries of naturally infectedRh. sanguineus ticks (Fig. 3) [84].
1.2. Ticks as a life-long reservoir of Rickettsiae
Many species of the genus Rickettsia areconsidered to be vertically transmitted symbi-onts of invertebrates. It has been suggested thatRickettsiae were initially symbionts of inverte-brates that secondarily became vertebrate patho-gens [64]. Ticks and SFG Rickettsiae couldtherefore represent only one branch of a possi-bly larger group of evolutionary associationsbetween Rickettsiales and arthropods.
Transstadial (TS, from one life stage to thenext) and transovarial (TOT) transmission ofdifferent rickettsial species has been reportedin many tick species (Tab. I). Since Ixodid ticksfeed only once during each stage of theirindividual development, TS transmission isnecessary for rickettsial survival in ticks. Tran-sovarial transmission can be defined by twospecific infection rates: (i) the transovarialinfection rate, which is the percentage offemales that pass microorganisms to their prog-eny, and (ii) the filial infection rate (FIR), whichis the percentage of infected progeny derivedfrom an infected female. Both TS and TOTtransmission of Rickettsiae in ticks have beendemonstrated for human pathogens such asR. ricketsii [13], R. slovaca [73], R. sibirica[67], R. africae [35, 85], R. parkeri [26],R. massiliae [49], and R. conorii [84] (Tab. I).The infection rates were obtained by allowingnaturally infected ticks to feed on healthy labo-ratory animals or by studying experimentallyinfected ticks under laboratory conditions.However, the prevalence of ticks infected bythe same bacterium may vary significantlyand be either low (R. conorii – Rh. sanguineus,usually < 1%) or high (R. africae – A. variega-tum, up to 100%). When Rickettsiae are effi-ciently transmitted by TS and by TOT in agiven tick species, this tick species may serveas a reservoir of the bacteria, and the distribu-tion of rickettsial disease should be identicalto that of its tick host [62]. However, the dogbrown tick Rh. sanguineus occurs in manyregions where R. conorii does not.
Many questions still remain unanswered:Why are there so many differences in the distri-bution of infected ticks in nature? Which factorscontrol the prevalence of infected ticks in
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 7 of 20

nature? Is there some specificity between ticksand Rickettsiae? For instance, some Rickettsiae,such as R. rickettsii, may be associated with dif-ferent tick vectors belonging to different genera.This contrasts with other Rickettsiae, such asR. conorii, which appear to be associatedmainly with one tick vector, Rh. sanguineus.Between these extremes, there are some Rickett-siae that are associated with several tick specieswithin the same genus, such as R. africae andR. slovaca, which are associated with variousAmblyomma spp. and Dermacentor spp.,respectively (Tab. I) [62, 63]. Rickettsiae prob-ably possess some specificity in their abilityto enter the cells of a given tick species. Forexample, R. peacockii multiplied in cell linesobtained from the hard ticks D. andersoni,
D. albipictus, I. scapularis, and I. ricinus, butthese bacteria were non-permissive in a cell lineobtained from the tick Amblyomma america-num [38]. The original sources of the tick cellsin these assays are not always clear, and itmight be difficult to interpret whether the failureof certain Rickettsiae to invade is due to nonper-missiveness at a species level or whether thetick cell line used is more characteristic of atissue type that is rarely invaded.
Ricketts’ hypothesis that the agent of SFG ismaintained in nature by the establishment ofnew populations of infected ticks has gained arenewed significance. The probability of newpopulations of ticks becoming infected withRickettsiae is difficult to precisely calculate,but a rough estimate can be obtained based on
Figure 4. Rickettsia conorii detected in ovaries from infected Rhipicephalus sanguineus adult ticks usingelectron microscopy.
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 8 of 20 (page number not for citation purpose)

the assumed life span of susceptible mammals,the antibody prevalence in mammals, the aver-age number of days of peak rickettsemia ininfected animals, and the number of days ofinfectious feeding on rickettsemic animalsrequired to establish generalized infections inticks [50].
2. HOW TO LIVE TOGETHER
2.1. Rickettsial challenges for ticks
Little is known about the consequences ofthe presence of Rickettsiae in host ticks. Oneshould not conclude that Rickettsiae and theirtick hosts have developed a perfect symbioticrelationship or that the infection of various tickspecies with Rickettsiae is always systemic, per-manent, and mutually beneficial, despite theirlong-term relationship.
Rickettsiae that infect various invertebratespecies and that have no known pathogenicityfor vertebrate hosts have been shown to inducedifferent effects on their hosts, including male-killing in beetles (Coleoptera), reduced fecun-dity and weight in true bugs (Hemiptera),parthenogenesis in wasps (Hymenoptera), andlarger body size in leeches (Hirudinida).Therefore, it is interesting to note that the tickAmblyomma rotundum, one of the few strictlyparthenogenetic tick species, has been foundto be 100% infected with R. bellii [39]. The firstand only male discovered in this species wasrecently reported, but it was not determinedwhether it harbored R. bellii [40].
Harmful effects in laboratory-infected tickshave been reported for pathogenic Rickettsiaeas well as for Rickettsiae of unknown pathoge-nicity in humans. Burgdorfer [16] reported thatin the fifth generation of ticks experimentallyinfected with R. rickettsii, close to 50% of thedepleted females died within 1 to 2 weeks,while the surviving females oviposited poorlyand only a few eggs hatched. However, it isnot known whether this effect was due torickettsial infection or to any other factors. Itshould also be noted that 100% FIR wasobserved among viable ticks through twelvegenerations of infected D. andersoni, despite
the high mortality among infected ticks [16].A subsequent study confirmed the adverseeffects of the highly virulent R. rickettsii infec-tion on tick development/oviposition. The studyreported decreased fecundity in ticks naturallyinfected with R. montanensis, R. bellii, andR. rhipicephali, but not for other speciessuch as R. peacockii [56]. Interestingly, somelimited cytopathological effects (mitochondrialchanges, membrane breakage, and general lossof ground substance) in the salivary glandsand the ovarian tissues of Rh. sanguineusinfected with R. rhipicephali were noted,although feeding and oviposition were notaffected [28]. Santos et al. [79] used intracelo-mic inoculation of R. conorii to show a negativeeffect on Rh. sanguineus nymphs, includingdeath during molting or soon after hatching intoadult instars, while the remaining 50% ofinfected adults exhibited severe malformations.The inoculation method used may have led to adecrease in tick survival, but the use of controlgroups suggested that R. conorii itself, not theinoculation method, was responsible for theeffect on the survival of its tick vector. Later,a high mortality in Rh. sanguineus ticksinfected with R. conorii was reported using dif-ferent methods of inoculation, including the useof bacteremic rabbits [48]. Possible reasons forthis reduction in fitness included the geographicorigin of the ticks, which came from Thailandwhere R. conorii has not been reported, or theirassociation with the pathogen load acquiredduring laboratory experiments. However, simi-lar experiments have been performed usingticks from southern France, and it was con-cluded that the lethal effect of R. conorii onRh. sanguineus ticks is unrelated to the geo-graphical origin of the ticks [86]. Even withthese findings, it is not known how viabilityin field conditions can be extrapolated fromthe results from laboratory-infected ticks, sinceexperimental models do not reproduce real-lifesituations. Recently, thirty engorged femaleRh. sanguineus ticks were collected from sevendogs owned by patients who had contractedMSF in Algeria during 2006 [84]. One femalewas found to be infected by R. conorii, thecausative agent of MSF. The larvae and allsubsequent stages of this infected female tick
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 9 of 20

were placed on a New Zealand White rabbit(Oryctolagus cuniculus) that was used as thehost for the ticks’ blood meals, and specimensof all stages of the following generations weretested by PCR. Vertical transmission of R. con-orii in naturally infected Rh. sanguineus tickswas demonstrated over five generations. Fur-thermore, the TOT rate was 100%, and theFIR was up to 99% for the fourth generationof infected ticks. R. conorii were also detectedin the ovaries of infected ticks, lending supportto the mechanism of transmission found ininfected Rh. sanguineus [84]. More investiga-tions on Rh. sanguineus-R. conoriii interactionsare needed to understand the discrepancybetween the efficient vertical transmission ofthe agent in naturally infected ticks and thelow prevalence in nature.
The role of external factors in the tick-Rickettsiae relationship deserves specific atten-tion. The stress conditions encountered byRickettsiae within the tick include starvationand temperature shifts. Interestingly, femaleD. andersoni ticks infected with R. rickettsiiand incubated at 4 �C demonstrated lower mor-tality than infected ticks held at 21 �C [56].Similarly, we have recently compared the fit-ness and survival of several stages of Rh. san-guineus that were either uninfected or infectedwith R. conorii. Interestingly, engorged nymphsinfected with R. conorii and exposed to a lowtemperature (4 �C) for one month exhibitedan absence of molting and had a higher mortal-ity when transferred to 25 �C, in comparison touninfected ticks. Since Rh. sanguineus over-winter as engorged nymphs, these preliminaryresults suggest that a low proportion of infectedticks would survive the winter1.
Once ingested, Rickettsiae appear in differ-ent organs due to the high degree of adaptationof these microorganisms to their vector [72].Engorged ticks infected with R. slovaca containmore Rickettsiae than starved infected ticks.The highest concentrations of Rickettsiae wereobserved in the hemolymph and hemocytes,followed by the cells of the ‘‘fat’’ body or tra-cheal complex, intestine, ovaries, synganglion,and salivary glands; limited infestation was
observed in the cells of Gene’s organ [73].The ultrastructure of Rickettsiae in naturallyinfected ticks is similar to that of all membersof the SFG Rickettsiae [21, 28, 68, 92]. Infectedtick cells do not apparently exhibit any cyto-pathic effects. However, in the case of certaintick-Rickettsiae associations, such as R. peac-okii in D. andersoni, Rickettsiae are apparentlyunable to invade tick hemocytes or salivarygland tissues. Therefore, R. peacockii may notbe transmitted to vertebrates at all, but mayremain strictly as symbionts of the ticks andmay be transmitted only vertically [4].
Despite the absence of any evident cyto-pathic effect of Rickettsiae on tick cells, theinfluence of bacterial infection on the tickorganism is widespread. It affects multipleorgans and systems, and its impact may berevealed by measuring differences in oxygenuptake and CO2 elimination, as shown inH. asiaticum ticks infected with R. sibirica,and by measuring changes in amino acid com-position in the same tick species [3]. Recentstudies have shown an increased expression ofantimicrobial peptides or induced phagocytosis[51]. Antimicrobial gene expression in ticks islocalized in the hemolymph, hemocytes, mid-gut, and fat body, illustrating the immunocom-petence of many tissues that Rickettsiaepresumably invade once acquired by a tick[18, 41]. Antimicrobial gene expression pat-terns of D. variabilis ticks challenged withR. montanensis show increases in defensin-1(vsnA1), defensin-2, and lysozyme, suggestingthat antimicrobial genes play a role during theacquisition-invasion stages of Rickettsiae in atick [18].
The molecular mechanisms of the interac-tions between Rickettsiae and D. variabilis tickshave been studied using molecular techniquesincluding differential display [45], expressionlibrary screening [46], subtractive hybridization[51], and sequence cloning [82]. These multi-faceted approaches have led to the identificationof several tick-derived molecules that aresuspected in the initiation of tick infectionand in rickettsial transmission (Tab. II). Thedifferentially expressed gene products, whichwere classified according to their putative func-tions, include receptor and adhesion molecules,1 Socolovschi C., unpublished data.
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 10 of 20 (page number not for citation purpose)

stress response proteins, and immune responseproteins [45, 51]. The up-regulation of a num-ber of these molecules in the Rickettsia-infectedtissues may be correlated with the reactivationand massive replication of Rickettsiae withinthe ovaries [30]. However, down-regulation ofall of these molecules was observed in themid-gut, an organ not directly associated withvertical transmission.
2.2. Tick challenges for Rickettsiae
The influence of the host on various proper-ties of SFG Rickettsiae is not only related to the
fundamental characteristics of the tick, but alsoto other factors, such as vertebrate hosts andenvironmental conditions, which certainly exerttheir influence on Rickettsiae through their vec-tors. In ticks, pathogens experience drastic fluc-tuations in temperature, hemolymph osmoticpressure (values of 350 mosmol/L may increaseto > 450 in unfed ticks), pH (varying from 6.8in the gut to 9.5 in saliva), O2 and CO2 tensionin tissues (17 to 18% O2 and 3% CO2 in activefeeding ticks versus O2 levels as low as 6% inunfed ticks), and nutrient flow [42, 52, 55]. Atemperature increase and an initiation ofengorgement are signals that have been shown
Table II. Activity and predicted function of novel tick genes identified from uninfected and Rickettsia-infected D. variabilis using molecular techniques.
Predicted function Putative identification Expression duringrickettsial infection
Adhesion or invasion Mucin-like protein - - -Clathrin adaptor protein +++Tetraspanin +++Protein inhibitor of signaltransducer and activator of transcription 1/3
- - -
ATPase of clathrin-coated vesicles +++Catenin +++
Tick immune andstress response
Ferritin +++a-dehydrogenase reductase +++Glutathione S-transferase +++Nucleosome assembly protein +++Cyclin A2 protein +++Cu2+-transporting ATPase +++Tubulin a chain +++Defensin -/+Lysozyme +Serine protease NSProphenoloxidase-activating factor +++
Tick-host interactions a-2 macroglobulin +++Salivary glue precursor +++IgE-dependent histamine release factor +++ENA vasodilator +++Calreticulin - - -Histamine release factor +++
Unknown Probable elongation factor +++Similar to Drosophila melanogasterCG17525
+++
Glycine-rich protein +++
NS: Not studied.
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 11 of 20

to activate multiplication. Reactivation may bea universal adaptation of tick-borne agents tothe long periods of metabolic inactivity in theiracarine hosts, but it remains poorly understood[54]. By becoming dormant during the longtransstadial phase and during host-seeking, theagent does not utilize scarce stored resourcesand reduces any effect on fitness. Once the tickattaches, the change in temperature and physiol-ogy of the tick host induces the agent to emergefrom dormancy and attain infectivity. In nature,stress conditions encountered by Rickettsiaewithin the tick include starvation and tempera-ture shifts. In the laboratory, R. rickettsii inD. andersoni ticks lose their virulence for gui-nea pigs when the ticks are subjected to physi-ological stress such as low environmentaltemperature or starvation. However, subsequentexposure of these same ticks to 37 �C for 24 to48 h or the acquisition of a blood meal mayrestore the original virulence of the bacteria.The number of plaque-forming units per dropof hemolymph is almost 100-fold greater forpartially engorged ticks than for starved ticks[93]. Reacquisition of infectivity correlates withthe reappearance of the microcapsular and slimlayers of the rickettsial outer surface [30] thatmay be involved in actin polymerization andrickettsial mobility in tick cells [53, 81]. Theelectron-lucent ‘‘halos’’ of Rickettsiae inengorged ticks were also noted for R. slovacain D. marginatus [21] and R. rhipicephali inRh. sanguineus [28].
During tick blood-feeding,Rickettsiaeundergovarious physiological changes and proliferateintensively [88]. The changes induced by abloodmeal in the tick activate the energymetab-olism of Rickettsiae, involving coenzyme Nico-tinamide adenine dinucleotide, coenzyme A,ATP, glutathione, andglutamate oxidization.Vir-ulent R. rickettsii could be rendered avirulentwith para-aminobenzoic acid [25]. However,the stress adaptation in some Gram-negativebacteria, also known as the stringentresponse, has been shown to be mediated bythe nucleotide guanosine-3,5(bis)pyrophos-phate(ppGpp), which is modulated by spoTgenes [78]. This phenomenon could play a rolein the adaptation of Rickettsiae to ticks andmay be involved in the process of reactivation.
It has also been hypothesized that changes inouter surface proteins occur during alternatinginfection in ticks and in mammals [78], sincethe expression of R. massiliae rOmpAwas lowerduring the larval stage while the expression ofrOmpB did not change with temperature orbetween life stagesof infectedRh. turanicus ticks[60].
3. INTERFERENCE
Ticks are candidate hosts for the demonstra-tion of interference, defined as the ‘‘inhibition(partial or complete) of rickettsial replicationby another Rickettsia’’, due to their possibleexposure to multiple rickettsial species whenfeeding on multiple hosts [64]. In the 1980s,Burgdorfer et al. [15] demonstrated that ticksinfected with R. peacockii were refractory toinfection with and maintenance of R. rickettsii.The interference phenomenon was also testedunder laboratory conditions in which the block-age of transovarial transmission of R. rickettsiiwas observed in ticks infected with eitherR. montanensis or R. rhipicephali [12]. Thesestudies corroborate with findings indicating thatR. rickettsii occurs with a lower frequency inDermacentor ticks, as compared to other Rick-ettsiae. A recent study of interspecies competi-tion between different Rickettsiae in the sametick, using cohorts of R. montanensis-infectedand R. rhipicephali-infected D. variabilis, havedemonstrated similar inhibitory effects betweenRickettsiae: infected ticks exposed to other rick-ettsial species by capillary feeding were incapa-ble of maintaining both rickettsial speciestransovarially. It was suggested that rickettsialinfection of tick ovaries may alter the patternof molecular expression in the oocytes, thusresulting in interference or blocking of thesecond infection [44]. These data indicate thatticks are not able to maintain two differentspecies of Rickettsia via transovarial transmis-sion. It was speculated that competitionbetween Rickettsiae for establishing successfultick infection facilitates a single rickettsialinfection.Thesedataalso support theobservationthat ticks collected from various geographic
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 12 of 20 (page number not for citation purpose)

regions are often infected with only one SFGRickettsia species [2].
Nevertheless, preliminary studies showedthat R. bellii, which is not an SFG Rickettsia,can coexist with other Rickettsiae in ticks inthe wild [9]. Blanc et al. [9] demonstrated thatR. massiliae recently acquired by lateral genetransfer the tra region, presumably involved inpilus formation and conjugal DNA transferfrom a species related to R. bellii. Thus, the pat-tern of identifiable horizontal gene transfer inRickettsiae validates the ‘‘intracellular arena’’hypothesis [11], which stipulates that geneticmaterial can move in and out of communitiesof obligate intracellular bacteria that co-infectthe same intracellular host environment. Furtheranalysis of the genomic sequences identifiedadditional candidates for lateral gene transferbetween Rickettsiae. Moreover, it was demon-strated that R. bellii evidently share commonorigin genes with chlamydial intracellular bac-teria residing in amoebas [59]. Therefore, thepossibility of a close interaction between SFGRickettsiae and Rickettsiae from other rickettsialgroups, as well as with other organisms, is quiterealistic and might be beneficial for all partici-pants. For example, I. scapularis can harborboth a rickettsial endosymbiont that is not trans-mitted and the Lyme disease spirocheteBorrelia burgdorferi, and several reports havedemonstrated that Ixodes ticks can harborB. burgdorferi and the human granulocyticagent Anaplasma phagocytophilum [54].
4. METHODS OF RESEARCH INTO TICK-RICKETTSIAE INTERACTIONS
The first study on tick-Rickettsiae interac-tions was carried out by Ricketts during hisinvestigation of RMSF in western Montana,in which he demonstrated that D. andersoniwas the principal vector of R. rickettsii [77].Subsequently, naturally infected ticks wereused to investigate such interactions, includingthe vertical transmission of R. conorii inRh. sanguineus [84], of R. africae in A. hebra-eum [35] or in A. variegatum [85], and ofR. massiliae in Rh. turanicus [49]. They werealso used to study the impact of Rickettsiae
on their host’s physiology and reproduction[94]. One of the challenges of using wild-caught ticks is the collection of sufficient num-bers of infected ticks (because the prevalence ofinfection in nature may be low) and their main-tenance in a laboratory environment. Herein,we present the main methods of creating exper-imental models with laboratory-infected ticks.
4.1. Experimental models
Historically, Rickettsiae-infected ticks havebeen most commonly produced by allowingticks to feed on rickettsemic animals, such asguinea pigs. This has been performed withA. americanum and R. parkeri; D. andersoniand R. rickettsii; and Rh. sanguineus andR. conorii [17, 26, 48, 56].
The capillary tube feeding (CTF) systemoffers a method of exposing ticks to pathogenswithout the use of infected hosts and providesan artificial system in which the compositionof the tick meals could be modified for experi-mental purposes. The CTF system, used ini-tially as a feeding system for soft ticks, wasadapted to infect Ixodid ticks with rickettsialorganisms [71]. This method was used to studythe following: (i) the transmission of R. conoriiin Rh. sanguineus and of R. montanensis andR. rhipicephali in D. variabilis; (ii) the visuali-zation of R. monacensis in I. scapularis; and(iii) the antimicrobial gene expression profilesof R. montanensis [5, 18, 43, 48]. The capillaryfeeding method allows researchers to quantifythe volume of solution ingested by ticks andto confirm the dissemination of Rickettsiae fromthe gut of orally infected ticks to other tissues.Matsumoto et al. [48] have used immersion ofengorged nymphs with one cut leg, two cutlegs, or a cut cuticle in a solution containingR. conorii, to infect R. sanguineus ticks. Thismethod was recently used to infect I. ricinuswith B. burgdorferi for an assay monitoringthe dynamics of infection within the tick hostafter feeding [23].
In the early 20th century, partially engorgedfemale ticks were used as laboratory subjectsfor various microbiological studies, using, forexample, the intracelomic inoculation method[75]. Recently, uninfected engorged nymphs
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 13 of 20

were inoculated intracelomically with a rickett-sial suspension to study the infection processof R. conorii in the salivary glands ofRh. sanguineus ticks [79]. This model was usedfor cultivation not only of Rickettsiae, but alsofor arboviruses and microsporidia.
The feeding of ticks with blood throughanimal-derived or artificial membranes hasbeen documented since 1956, when Boophilusmicroplus larvae were cultivated on the cellmembrane of an embryonated hen egg [66].Subsequently, this membrane technique wasmodified to accommodate different species ofticks, and the latest improvement was theintroduction of the elastic characteristic of theskin into the membrane structure [36]. Feed-ing assays could be used for studies on thedynamics of pathogen transmission – fromthe nutrient medium to the tick, from the tickto the medium, and between infected anduninfected ticks feeding in the same feedingunit – without having to take into account par-asite-host-pathogen interactions.
The successful isolation and propagation ofseveral tick-borne pathogens in tick cell lineshave resulted in a useful model for studying
interactions between tick cells and Rickettsiae.Over forty cell lines are currently available fromthirteen Ixodid and one Argasid tick species.Most of the currently available tick cell lineswere established from embryonic cells, usingsimple methodology, with no attempt to selectparticular tissue types [8]. The tick cell linesused in SFG Rickettsiae studies are listed inTable III. These tick cell lines were used forthe isolation and propagation of Rickettsiae,such as the isolation of SFG Rickettsiae bycultivation with mid-gut tissues from A. ameri-canum [53]. Recently, the ISE6 cell line (Ixodesscapularis ticks) was used to isolate previouslyuncultivated strains of R. felis from catfleas [70]. R. peacockii, an endosymbiont ofD. andersoni that seems to interfere with thetransmission of R. rickettsii, was found to causechronic infection in the D. andersoni cell lineDAE100 [80]. Tick cell lines are also essentialfor studies on genomics, proteomics, andgenetic manipulation. With the availability ofgenomics tools, tick cell lines will become anincreasingly important support for tick andtick-borne disease research in vivo, oncegenetic transformation and gene silencing
Table III. Tick cell lines used for isolation and propagation of SFG Rickettsiae.
Rickettsia Tick species used for tick cell lines Tick cell lines used
R. rickettsii Ixodes scapularis (embryo) IDE2, IDE8, ISE6Dermacentor albipictus (embryo) DALBE3
R. peacockii Dermacentor andersoni (embryo) DAE100, DAE3, DAE15Ixodes scapularis (embryo) ISE6, IDE 12, IDE2, IDE8Boophilus microplus BME26Dermacentor variabilis DVE1Ixodes ricinus IRE 111Carios capensis CCE3
R. monacensis Ixodes scapularis ISE6Ixodes ricinus IRE11Dermacentor andersoni (embryo) DAE100Ixodes scapularis IDE8
R. helvetica Ixodes ricinus IRE11R. montanensis Ixodes scapularis (embryo) IDE2
Dermacentor albipictus (embryo) DALBE3R. felis Ixodes scapularis ISE6Rickettsia spp. (SFG) Rhipicephalus appendiculatus RAE25
Ixodes scapularis IDE2, IDE8Carios capensis CCE3, CCE2
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 14 of 20 (page number not for citation purpose)

techniques using RNA interference becomeroutine [8].
4.2. Role of molecular tools in understandingtick-Rickettsiae interactions
The genome of SFG Rickettsiae is highlyconserved. Complete genome sequences canbe found in the public domain for severalSFG Rickettsiae: R. conorii [57], R. rickettsii[22], R. sibirica [47], R. massiliae [9], andR. africae [24]. Genome sequences will soonbe available for other Rickettsiae, includingR. slovaca, R. helvetica, R. raoultii, R. parkeri,R. australis, and R. rhipicephali. The small gen-omes of Rickettsiae have arisen through a recentand ongoing genome degradation process, withmany pseudo-genes and a high proportion ofnon-coding DNA [1]. Genomic data revealedmarked similarities between the various species,including the loss of genes encoding enzymesfor sugar metabolism and for lipid, nucleotide,and amino acid synthesis; this loss is responsi-ble for the inability to cultivate Rickettsiae incell-free media. Each genome exhibits specificfeatures, reflecting a large diversity in the para-sitic and infectious strategies of Rickettsiae.
SFG Rickettsiae associated with ticks havedeveloped a molecular mechanism to synchro-nize their replication with the physiology oftheir tick hosts. Molecular mechanisms impli-cated in the adaptation of SFG Rickettsiae todifferent host conditions and in the reactivationof virulence are unknown. Therefore, genesfound in multiple copies may outline specificadaptations. Among these genes is spoT, forwhich five copies were identified. SpoT genesare regulators of the global cellular metabolismor ‘‘stringent’’ response to starvation andstress and enhance cell survival [57]. InR. conorii, preliminary experiments showedthat environmental stress conditions are accom-panied by variable spoT1 transcription, aphenomenon that could intervene in the adapta-tion of these bacteria to unfed ticks and in reac-tivation [78].
The completion of genomic sequences ofnumerous tick-transmitted bacterial speciesin the families Anaplasmataceae and Rickettsi-aceae allows for comparative genomic
approaches to detect genes and pathwaysunique to tick-transmitted species. Importantly,comparative approaches are unbiased to thelocation or function of a protein and will detectsurface proteins, regulators, and transportersthat may be required for replication in a tick,as well as novel enzymes and proteins ofunknown function. To illustrate this approach,Brayton et al. [12] compared the genomes ofthree tick-transmitted pathogens (A. marginale,E. ruminantium, and R. conorii) with thegenome of W. pipientis, a non-tick-transmittedbacterium. The majority of the genes hadPFAM matches (a large collection of proteinmultiple sequence alignments and profile-hidden Markov models) [7], but the gene namesor functions could not be definitively assigned.Some genes included sequences for a conservedcell-surface protein, and several genes coded fornucleotide-processing enzymes such as tRNApseudouridine 55 synthase, GTP cyclohydrolaseI, cytidylate kinase, and exo- deoxyribonuclease.
The recent discovery of pRF in R. felis, anSFG Rickettsia associated with fleas, usingwhole genome sequencing [58] has put intoquestion the long-held belief that plasmids arenot present in Rickettsiae. Baldridge et al. [6]have shown that plasmids are present in severalSFG Rickettsiae, including R. amblyommii,R. massiliae, R. peacockii, R. helvetica, Candid-atus R. hoogstraalii, and R. monacensis. Theseauthors also demonstrated the loss of plasmidsduring serial cultures of R. peacockii, which ismaintained by TOT in the tick host and is nota vertebrate pathogen, suggesting a possiblerole for plasmids in adaptation to the tick host.The location of genes encoding a-Hsps, in addi-tion to membrane transport proteins, cell sur-face antigens, and unique rickettsial proteinsof unknown function, on a plasmid that maybe present in multiple copies per cell mightfacilitate enhanced transcription and expressionof genes involved in adaptation to changes inhost physiology [6].
The tick genome also provides an unparalleledresource for studying not only tick biology, butalso tick-host-pathogen relationships. Informa-tion on expressed sequence tags is available forseveral tick species, including Rh. appendicula-tus, Rh. microplus, A. variegatum, I. scapularis,
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 15 of 20

I. ricinus, I. pacificus, and Hyalomma anatoli-cum [34]. Some of these tick genes have beenused to create a repository of clustered andauto-annotated data in the form of species-specific gene indices. So far, only the genomeof I. scapularis (2.15 billion bp) has beensequenced [32]. It is estimated that the genomeof Rhipicephalus (previously Boophilus)microplus is 7.1 billion bp in length (over twiceas long as the human genome), and its sequenc-ing is in progress [27]. A. americanum isanother metastriate tick whose genome size(1 billion bp) and organization have been deter-mined [61]. The genome size and organizationof these three tick species (I. scapularis,Rh. microplus, and A. americanum) are distinctfrom those of other arthropods, due to a greaterproportion of moderately repetitive DNA.
The single genomic technique that has hadthe greatest impact on tick research is RNA inter-ference (RNAi). RNAi is a nucleic-acid-basedreverse genetic approach that involves the dis-ruption of gene expression to determine genefunction or its effect on a metabolic pathway.RNAi has been used to study gene function,the characterization of the tick-pathogen inter-face, and the screening and characterization oftick protective antigens [19, 20]. The applica-tions of RNAi to tick research will contributeto the development of vaccines to control tickinfestations and the transmission of tick-bornepathogens. Genomics tools do not act as a substi-tute for established methods in the study of ticksand tick-borne pathogens, but rather comple-ment them. A major goal will be the analysisof the complete genetic information of allRickettsiae, in order to study global gene expres-sion of many genes (transcriptomics), knowingthat these microorganisms most probably adaptto different conditions in the environment orduring interaction with the host by changingtheir gene expression program, and to character-ize all expressed proteins in an organism(proteomics).
5. CONCLUSION AND PERSPECTIVES
Sixteen new tick-transmitted rickettsioseshave been described over the last twenty years,
whereas only four had been characterized priorto 1984. This dramatic increase in the numberof recognized rickettsial infections is a resultof the broad use of cell culture systems andthe development of molecular tools for theidentification of Rickettsiae from human sam-ples and ticks [63]. Continuing investigationson tick-Rickettsiae interactions may provide abetter understanding of the factors influencingthe emergence and distribution of arthropod-transmitted pathogens and their co-evolution.It is evident that in the context of the relation-ship between Rickettsiae and their vectors,many questions remain unanswered. Someunclear issues need further elucidation, suchas the fitness of infected and non-infected ticks,the nature of the tick-Rickettsia relationship, theimpact of extrinsic factors on infected ticks,dual infection with different agents and hori-zontal gene transfer, the time and duration ofco-evolution, and the benefits for Rickettsiaein becoming pathogenic for vertebrates. Muchwork is still needed to define the molecularbasis of interactions between Rickettsiae andtick cells. The use of modern detection and iso-lation methods, including electron microscopy,histochemistry, and tissue cultures from vectors,as well as the use of genomic tools and analysisof the transcriptome and proteome of Rickett-siae, will certainly accelerate our understandingof these pathogens.
REFERENCES
[1] Andersson S.G.E., Zomorodipour A., AnderssonJ.O., Sicheritz-Ponten T., Alsmark U.C.M., PodowskiR.M., et al., The genome sequence of Rickettsiaprowazekii and the origin of mitochondria, Nature(1998) 396:133–140.
[2] Azad A.F., Beard C.B., Rickettsial pathogens andtheir arthropod vectors, Emerg. Infect. Dis. (1998)4:179–186.
[3] Balashov I.S., Daiter A., Stanyukovich A.K., Theeffect of infestation with the Rickettsia Coxiellaburneti and Dermatoxenus sibiricus on the contentof free amino acids in the tick Hyalomma asiaticum,Parazitologiia (1969) 3:281–285.
[4] Baldridge G.D., Burkhardt N.Y., Simser J.A.,Kurtti T.J., Munderloh U.G., Sequence and expression
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 16 of 20 (page number not for citation purpose)

analysis of the ompA gene of Rickettsia peacockii, anendosymbiont of the Rocky Mountain wood tick,Dermacentor andersoni, Appl. Environ. Microbiol.(2004) 70:6628–6636.
[5] Baldridge G.D., Kurtti T.J., Burkhardt N., Bald-ridge A.S., Nelson C.M., Oliva A.S., Munderloh U.G.,Infection of Ixodes scapularis ticks with Rickettsiamonacensis expressing green fluorescent protein: Amodel system, J. Invertebr. Pathol. (2007) 94:163–174.
[6] Baldridge G.D., Burkhardt N.Y., Felsheim R.F.,Kurtti T.J., Munderloh U.G., Plasmids of the pRM/pRF family occur in diverse Rickettsia species, Appl.Environ. Microbiol. (2008) 74:645–652.
[7] Bateman A., Birney E., Durbin R., Eddy S.R.,Howe K.L., Sonnhammer E.L., The Pfam proteinfamilies database, Nucleic Acids Res. (2000) 28:263–266.
[8] Bell-Sakyi L., Zweygarth E., Blouin E.F., GouldE.A., Jongejan F., Tick cell lines: tools for tick andtick-borne disease research, Trends Parasitol. (2007)23:450–457.
[9] Blanc G., Ogata H., Robert C., Audic S., ClaverieJ.M., Raoult D., Lateral gene transfer between obligateintracellular bacteria: evidence from the Rickettsiamassiliae genome, Genome Res. (2007) 17:1657–1664.
[10] Blanc J.L., Caminopetros J., Epidemiologicaland experimental studies on Boutonneuse fever doneat the Pasteur Institute in Athens, Arch. Inst. PasteurTunis (1932) 20:343–394.
[11] Bordenstein S.R., Wernegreen J.J., Bacterio-phage flux in endosymbionts (Wolbachia): infectionfrequency, lateral transfer, and recombination rates,Mol. Biol. Evol. (2004) 21:1981–1991.
[12] Brayton K.A., Kappmeyer L.S., Herndon D.R.,Dark M.J., Tibbals D.L., Palmer G.H., et al., Completegenome sequencing of Anaplasma marginale revealsthat the surface is skewed to two superfamilies of outermembrane proteins, Proc. Natl. Acad. Sci. USA (2005)102:844–849.
[13] Burgdorfer W., Brinton L.P., Mechanisms oftransovarial infection of spotted fever Rickettsiae inticks, Ann. N.Y. Acad. Sci. (1975) 266:61–72.
[14] Burgdorfer W., Aeschlimann A., Peter O., HayesS.F., Philip R.N., Ixodes ricinus: vector of a hithertoundescribed spotted fever group agent in Switzerland,Acta Trop. (1979) 36:357–367.
[15] Burgdorfer W., Hayes S.F., Mavros A.J., Non-pathogenic Rickettsiae in Dermacentor andersoni: alimiting factor for the distribution of Rickettsiarickettsii, in: Brugdorfer W., Anacker R.L. (Eds.),
Rickettsiae and rickettsial diseases, New York, Aca-demic Press, 1981, 585–594.
[16] Burgdorfer W., Transovarial transmission: aneffective ecological means for the survival of tick-borne spotted fever group Rickettsiae, Ann. N.Y.Acad. Sci. (1985) 265–271.
[17] Burgdorfer W., Ecological and epidemiologicalconsiderations of Rocky Mountain spotted fever andscrub typhus, in: Walker D.H. (Ed.), Biology ofrickettsial disease, Boca Raton, Florida, CRC Press,1988, pp. 33–50.
[18] Ceraul S.M., Dreher-Lesnick S.M., Gillespie J.J.,RahmanM.S., AzadA.F., New tick defensin isoform andantimicrobial gene expression in response to Rickettsiamontanensis challenge, Infect. Immun. (2007) 75:1973–1983.
[19] De la Funte J., Almazan C., Blouin E.F., NaranjoV., Kocan K.M., RNA interference screening in ticksfor identification of protective antigens, Parasitol. Res.(2005) 96:137–141.
[20] De la Fuente J., Kocan K.M., Almazan C.,Blouin E.F., RNA interference for the study andgenetic manipulation of ticks, Trends Parasitol. (2007)23:427–433.
[21] Diehl P.A., Rehacek J., Bazlikova M., Theultrastructure of Rickettsia slovaca in naturallyinfected females of the tick Dermacentor marginatus,Ann. Parasitol. Hum. Comp. (1980) 55:259–270.
[22] Ellison D.W., Clark T.R., Sturdevant D.E.,Virtaneva K., Porcella S.F., Hackstadt T., Genomiccomparison of virulent Rickettsia rickettsii SheilaSmith and avirulent Rickettsia rickettsii Iowa, Infect.Immun. (2008) 76:542–550.
[23] Fiserova L., Cerna K., Horka H., Kopecky J.,Two ways of experimental infection of Ixodes ricinusticks (Acari: Ixodidae) with spirochetes of Borreliaburgdorferi sensu lato complex, Folia Parasitol.(Praha) (2008) 55:150–154.
[24] Fournier P.E., El Karkouri K., Leroy Q., RobertC., Giumelli B., Renesto P., et al., Analysis of theRickettsia africae genome reveals that virulenceacquisition in Rickettsia species may be explained bygenome reduction, BMC Genomics (2009) 10:166.
[25] Gilford J.H., Price W.H., Virulent-avirulent con-versions of Rickettsia Rickettsii in vitro, Proc. Natl.Acad. Sci. USA (1955) 41:870–873.
[26] Goddard J., Experimental infection of lone starticks, Amblyomma americanum (L.), with Rickettsiaparkeri and exposure of Guinea pigs to the agent,J. Med. Entomol. (2003) 40:686–689.
[27] Guerrero F.D., Nene V.M., George J.E., BarkerS.C., Willadsen P., Sequencing a new target genome:
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 17 of 20

the Boophilus microplus (Acari: Ixodidae) genomeproject, J. Med. Entomol. (2006) 43:9–16.
[28] Hayes S.F., Burgdorfer W., Ultrastructure ofRickettsia rhipicephali, a new member of the spottedfever group Rickettsiae in tissues of the host vectorRhipicephalus sanguineus, J. Bacteriol. (1979) 137:605–613.
[29] Hayes S.F., Burgdorfer W., Aeschlimann A.,Sexual transmission of spotted fever group Rickettsiaeby infected male ticks: detection of Rickettsiae inimmature spermatozoa of Ixodes ricinus, Infect.Immun. (1980) 27:638–642.
[30] Hayes S.F., Burgdorfer W., Reactivation ofRickettsia rickettsii in Dermacentor andersoni ticks:an ultrastructural analysis, Infect. Immun. (1982)37:779–785.
[31] Heinzen R.A., Hayes S.F., Peacock M.G.,Hackstad T., Directional actin polymerization associ-ated with spotted fever group Rickettsia infection ofVero cells, Infect. Immun. (1993) 61:1926–1935.
[32] Hill C.A., Wikel S.K., The Ixodes scapularisGenome Project: an opportunity for advancing tickresearch, Trends Parasitol. (2005) 21:151–153.
[33] Hinton A., Bond S., Forgac M., V-ATPasefunctions in normal and disease processes, PflugersArch. (2009) 457:589–598.
[34] Jongejan F., Nene V., de la Fuente J., Pain A.,Willadsen P., Advances in the genomics of ticks andtick-borne pathogens, Trends Parasitol. (2007) 23:391–396.
[35] Kelly P.J., Mason P.R., Transmission of a spottedfever group Rickettsiae by Amblyomma hebraeum(Acari : Ixodidae), J. Med. Entomol. (1991) 28:598–600.
[36] Krober T., Guerin P.M., An in vitro feedingassay to test acaricides for control of hard ticks, Pest.Manag. Sci. (2007) 63:17–22.
[37] Kryuchechnikov V.N., Protective responses ofIxodoidea hemocytes, in: Dusbabek F., BukvaV. (Eds.),Modern Acarology, vol. 1, Academia, Prague, 1991,pp. 331–334.
[38] Kurtti T.J., Simser J.A., Baldridge G.D., PalmerA.T., Munderloh U.G., Factors influencing in vitroinfectivity and growth of Rickettsia peacockii (Rick-ettsiales: Rickettsiaceae), an endosymbiont of theRocky Mountain wood tick, Dermacentor andersoni(Acari, Ixodidae), J. Invertebr. Pathol. (2005) 90:177–186.
[39] Labruna M.B., Whitworth T., Bouyer D.H.,McBride J.,CamargoL.M.,CamargoE.P., et al.,Rickettsiabellii and Rickettsia amblyommii in Amblyomma
ticks from the State of Rondonia, Western Amazon,Brazil, J. Med. Entomol. (2004) 41:1073–1081.
[40] Labruna M.B., Terrassini F.A., Camargo L.M.,First report of the male of Amblyomma rotundatum(Acari: Ixodidae) from a field-collected host, J. Med.Entomol. (2005) 42:945–947.
[41] Lai R., Lomas L.O., Jonczy J., Turner P.C., ReesH.H., Two novel non-cationic defensin-like antimi-crobial peptides from haemolymph of the female tick,Amblyomma hebraeum, Biochem. J. (2004) 379:681–685.
[42] Lighton J.R.B., Fielden L.J., Rechav Y., Discon-tinuous ventilation in a non-insect, the tick Ambly-omma marmoreum (Acari: Ixodidae): characterizationand metabolic modulation, J. Exp. Zool. (1993)180:229–245.
[43] Macaluso K.R., Sonenshine D.E., Ceraul S.M.,Azad A.F., Infection and transovarial transmission ofRickettsiae in Dermacentor variabilis ticks acquiredby artificial feeding, Vector Borne Zoonotic Dis.(2001) 1:45–53.
[44] Macaluso K.R., Sonenshine D.E., Ceraul S.M.,Azad A.F., Rickettsial infection in Dermacentorvariabilis (Acari: Ixodidae) inhibits transovarial trans-mission of a second Rickettsia, J. Med. Entomol.(2002) 39:809–813.
[45] Macaluso K.R., Mulenga A., Simser J.A., AzadA.F., Differential expression of genes in uninfectedand Rickettsia-infected Dermacentor variabilis ticksas assessed by differential-display PCR, Infect.Immun. (2003) 71:6165–6170.
[46] Macaluso K.R., Mulenga A., Simser J.A., AzadA.F., Interactions between Rickettsiae and Dermacen-tor variabilis ticks: analysis of gene expression, Ann.N.Y. Acad. Sci. (2003) 990:568–572.
[47] Malek J.A., Wierzbowski J.M., Tao W., BosakS.A., Saranga D.J., Doucette-Stamm L., et al., Proteininteraction mapping on a functional shotgun sequenceof Rickettsia sibirica, Nucleic Acids Res. (2004) 32:1059–1064.
[48] Matsumoto K., Brouqui B., Raoult D., Parola P.,Experimental infection models of ticks of the Rhipi-cephalus sanguineus group with Rickettsia conorii,Vector Borne Zoonotic Dis. (2005) 5:363–372.
[49] Matsumoto K., Ogawa M., Brouqui P., RaoultD., Parola P., Transmission of Rickettsia massiliae inthe tick, Rhipicephalus turanicus, Med. Vet. Entomol.(2005) 19:263–270.
[50] McDade J.E., Newhouse V.F., Natural history ofRickettsia rickettsii, Annu. Rev. Microbiol. (1986)40:287–309.
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 18 of 20 (page number not for citation purpose)

[51] Mulenga A., Macaluso K.R., Simser J.A., AzadA.F., Dynamics of Rickettsia-tick interactions: identi-fication and characterization of differentially expressedmRNAs in uninfected and infected Dermacentorvariabilis, Insect Mol. Biol. (2003) 12:185–193.
[52] Munderloh U.G., Kurtti T.J., Cellular andmolecular interrelationships between ticks and pro-karyotic tick-borne pathogens, Annu. Rev. Entomol.(1995) 40:221–243.
[53] Munderloh U.G., Hayes S.F., Cummings J.,Kurtti T.J., Microscopy of spotted fever Rickettsiamovement through tick cells, Microscopy and Micro-analysis (1998) 4:115–121.
[54] Munderloh U.G., Jauron S.D., Kurtti T.J., Thetick: a different kind of host for human pathogens, in:Goodman J.L., Dennis D.T., Sonenshine D. (Eds.),Tick borne diseases of humans, Washington, 2005, pp.37–64.
[55] Needham G.R., Teel P.D., Water balance by ticksbetween bloodmeals, in: Sauer J.R., Hair J.A. (Eds.),Morphology, physiology, and behavioral biology ofticks, Chichester, United Kingdom, 1986, pp. 100–151.
[56] Niebylski M.L., Peacock M.G., Schwan T.G.,Lethal effect of Rickettsia rickettsii on its tick vector(Dermatocentor andersoni), Appl. Environ. Micro-biol. (1999) 65:773–778.
[57] Ogata H., Audic S., Renesto-Audiffren P., Four-nier P.E., Barbe V., Samson D., et al., Mechanisms ofevolution in Rickettsia conorii and R. prowazekii,Science (2001) 293:2093–2098.
[58] Ogata H., Renesto P., Audic S., Robert C., BlancG., Fournier P.E., et al., The genome sequence ofRickettsia felis identifies the first putative conjugativeplasmid in an obligate intracellular parasite, PLoSBiol. (2005) 3:e248.
[59] Ogata H., La Scola B., Audic S., Renesto P.,Blanc G., Robert C., et al., Genome sequence ofRickettsia bellii illuminates the role of amoebae ingene exchanges between intracellular pathogens, PLoSGenet. (2006) 2:e76.
[60] Ogawa M., Matsumoto K., Parola P., Raoult D.,Brouqui P., Expression of rOmpA and rOmpB proteinin Rickettsia massiliae during the Rhipicephalusturanicus life cycle, Ann. N.Y. Acad. Sci. (2006)1078:352–356.
[61] Palmer M.J., Bantle J.A., Guo X., Fargo W.S.,Genome size and organization in the Ixodid tickAmblyomma americanum (L.), Insect Mol. Biol.(1994) 3:57–62.
[62] Parola P., Raoult D., Ticks and tickborne bacte-rial diseases in humans: an emerging infectious threat,Clin. Infect. Dis. (2001) 32:897–928.
[63] Parola P., Paddock C.D., Raoult D., Tick bornerickettsioses around the world: emerging diseaseschallenging old concepts, Clin. Microbiol. Rev.(2005) 18:719–756.
[64] Perlman S.J., Hunter M.S., Zchori-Fein E., Theemerging diversity of Rickettsia, Proc. Biol. Sci.(2006) 273:2097–2106.
[65] Philip C.B., Some epidemiological consider-ations in Rocky Mountain spotted fever, Public HealthRep. (1959) 74:595–600.
[66] Pierce A.E., Pierce M.H., A note on the culti-vation of Boophilus microplus (Canestrini, 1887)(Ixodidae: Acarina) on the embryonated hen egg,Aust. Vet. J. (1956) 32:144–146.
[67] Podboronov V.M., Pchelkina A.A., Characteris-tics of the transphase and transovarial transmission ofRickettsia sibirica by Ixodid and Argasid ticks, Med.Parazitol. (Mosk) (1989) 4:14–18 (in Russian).
[68] Popov V.L., Dyuisalieva R.G., Smirnova N.S.,Tarasevich I.V., Rybkin N.N., Ultrastructure of Rick-ettsia sibirica during interaction with the host cell,Acta Virol. (1986) 30:494–498.
[69] Popov V.L., Korenberg E.I., Nefedova V.V., HanV.C., Wen J.W., Kovalevskii Y.V., et al., Ultrastruc-tural evidence of the ehrlichial developmental cycle innaturally infected Ixodes persulcatus ticks in thecourse of coinfection with Rickettsia, Borrelia, and aflavivirus, Vector Borne Zoonotic Dis. (2007) 7:699–716.
[70] Pornwiroon W., Pourciau S.S., Foil L.D., Mac-aluso K.R., Rickettsia felis from cat fleas: isolation andculture in a tick-derived cell line, Appl. Environ.Microbiol. (2006) 72:5589–5595.
[71] Rau U., Hannoun C., The use of a capillary-tubetechnique for artificially feeding Argas reflexusreflexus ticks, Bull. World Health Organ. (1968) 39:332–333.
[72] Rehacek J., Development of animal viruses andRickettsiae in ticks and mites, Annu. Rev. Entomol.(1965) 10:1–24.
[73] Rehacek J., Rickettsia slovaca, the organism andits ecology, Acta Scientifica National Academy Sci-entifica Bohemoslovacae Brno. (1984) 18:1–50.
[74] Rehacek J., Ecological relationships betweenticks and Rickettsiae, Eur. J. Epidemiol. (1989) 5:407–413.
[75] Rehacek J., Sutakova G., Kocianova E., Use ofpartially engorged female ticks as laboratory animalsin microbiological research, Med. Vet. Entomol.(1994) 8:165–171.
Rickettsiae and ticks Vet. Res. (2009) 40:34
(page number not for citation purpose) Page 19 of 20

[76] Renesto P., Dehoux P., Gouin E., Touqui L.,Cossart P., Raoult D., Identification and characteriza-tion of a phospholipase D-superfamily gene in Rick-ettsiae, J. Infect. Dis. (2003) 188:1276–1283.
[77] Ricketts H.T., Some aspects of Rocky Moutainspotted fever as shown by recent investigations, Med.Rec. (1909) 16:843–855.
[78] Rovery C., Renesto P., Crapoulet N., MatsumotoK., Parola P., Ogata H., Raoult D., Transcriptionalresponse of Rickettsia conorii exposed to temperaturevariation and stress starvation, Res. Microbiol. (2005)156:211–218.
[79] Santos A.S., Bacellar F., Santos-Silva M., For-mosinho P., Gracio A.J., Franca S., Ultrastructuralstudy of the infection process of Rickettsia conorii inthe salivary glands of the vector tick Rhipicephalussanguineus, Vector Borne Zoonotic. Dis. (2002)2:165–177.
[80] Simser J., Palmer A.T., Munderloh U.G., KurttiT.J., Isolation of a spotted fever group rickettsia,Rickettsia peacockii, in a rocky mountain wood tick,Dermacentorn andersoni, cell line, Appl. Environ.Microbiol. (2001) 67:546–552.
[81] Simser J.A., Palmer A.T., Fingerle V., Wilske B.,Kurtti T.J., Munderloh U.G., Rickettsia monacensis sp.nov., a spotted fever group Rickettsia, from ticks(Ixodes ricinus) collected in a european city park,Appl. Environ. Microbiol. (2002) 68:4559–4566.
[82] Simser J.A., Macaluso K.R., Mulenga A., AzadA.F., Immune-responsive lysozymes from hemocytesof the American dog tick, Dermacentor variabilis andan embryonic cell line of the Rocky Mountain woodtick, D. andersoni, Insect Biochem. Mol. Biol. (2004)34:1235–1246.
[83] Socolovschi C., Doudier B., Pages F., Parola P.,Ticks and human tick-borne diseases in Africa, Med.Trop. (Mars.) (2008) 68:119–133 (in French).
[84] Socolovschi C., Bitam I., Raoult D., Parola P.,Transmission of Rickettsia conorii conorii in naturallyinfected Rhipicephalus sanguineus, Clin. Microbiol.Infect. (2009) May 7.
[85] Socolovschi C., Huynh T.P., Davoust B., GomezJ., Raoult D., Parola P., Transovarial and transstadialtransmission of Rickettsia africae in Amblyommavariegatum ticks, Clin. Microbiol. Infect. (2009)(in press).
[86] Socolovschi C., Matsumoto K., Brouqui B.,Raoult D., Parola P., Experimental infection of Rhipi-cephalus sanguineus with Rickettsia conorii conorii,Clin. Microbiol. Infect. (2009) (in press).
[87] Sonenshine D., The biology of tick vectors ofhuman disease, in: Goodman J.L., Dennis D.T.,Sonenshine D. (Eds.), Tick borne diseases of humans,Washington, ASM press, 2005, pp. 12–36.
[88] Spencer R.R., Parker R.R., Rocky mountainspotted fever: infectivity of fasting and recently fedticks, Public Health Rep. (1923) 38:333–339.
[89] Telford S.R., Parola P., Arthropods and Rickett-siae, in: Parola P., Raoult D. (Eds.), Rickettsialdiseases, New York, Infectious diseases and therapycollection, edited by B.A. Cuhna, Informa Healthcare,2007, pp. 27–36.
[90] Teysseire N., Chiche-Portiche C., Raoult D.,Intracellular movements of Rickettsia conorii andR. typhi based on actin polymerization, Res. Micro-biol. (1992) 143:821–829.
[91] Weiss E., History of Rickettsiology, in: WalkerD.H. (Ed.), Biology of rickettsial disease, Boca Raton,Florida, CRC Press,1988, pp. 15–32.
[92] Whitworth T., Popov V., Han V., Bouyer D.,Stenos J., Graves S., Ndip L., Walker D., Ultrastruc-tural and genetic evidence of a reptilian tick,Aponomma hydrosauri, as a host of Rickettsia honeiin Australia: possible transovarial transmission, Ann.N.Y. Acad. Sci. (2003) 990:67–74.
[93] Wike D.A., Burgdorfer W., Plaque formation intissue cultures by Rickettsia rickettsi isolated directlyfrom whole blood and tick hemolymph, Infect.Immun. (1972) 6:736–738.
[94] Zhong J., Jasinskas A., Barbour A.G., Antibiotictreatment of the tick vector Amblyomma americanumreduced reproductive fitness, PLoS ONE (2007)2:e405.
Vet. Res. (2009) 40:34 C. Socolovschi et al.
Page 20 of 20 (page number not for citation purpose)





























