Embed Size (px)
Citation preview

Review Article
The Neurobehavioral Teratology of Retinoids:A 50-Year History
Jane Adams*
Department of Psychology, University of Massachusetts, Boston, Massachusetts
Received 1 May 2010; Revised 24 June 2010; Accepted 7 July 2010
This review of the central nervous system (CNS) and behavioral teratology of the retinoids over the last 50 years is acommemorative retrospective organized by decade to show the prominent research focus within each period andthe most salient findings. In the 1960s, research focused on the gross CNS malformations associated with exposureand the delineation of dose-response and stage-specific responses in rodent models. Relevant scientific events beforeand during the 1960s are also discussed to provide the zeitgeist in which the field of neurobehavioral teratologyemerged in the 1970s. During this period, studies demonstrated that adverse effects on postnatal behavior could beproduced in animals exposed to doses of vitamin A lower than those that were teratogenic or impacted growth.Work during the 1980s showed an overrepresentation of behavioral studies focused on the reliability of screeningmethods, while the marked effects of human exposure were illustrated in children born to women treated with iso-tretinoin during pregnancy. The human catastrophe invigorated research during the 1990s, a period when techno-logical advances allowed more elegant examinations of the developing CNS, of biochemical, cellular, and moleculardevelopmental events and regulatory actions, and of the effects of direct genetic manipulations. Likewise, researchin the 1990s reflected a reinvigoration of research in neurobehavioral teratology evinced in studies that used animalmodels to try to better understand human vulnerability. These foci continued in the 2000–2010 period while exami-nations of the role of retinoids in brain development and lifelong functioning became increasingly sophisticated andbroader in scope. This review of the work on retinoids also provides a lens on the more general ontogeny of the fieldof neurobehavioral teratology. Birth Defects Research (Part A) 88:895–905, 2010. � 2010 Wiley-Liss, Inc.
Key words: retinoic acid teratology; hypervitaminosis A; all-trans-retinoic acid; isotretinoin; neurobehavioralteratology; prenatal brain development; neural activity of retinoids
INTRODUCTION
At the time of the founding of the Teratology Societyin 1960, much descriptive information had been gath-ered through carefully and systematically designedstudies of a variety of compounds. Indeed, the princi-ples of the field had been proposed (Wilson, 1959). Atthat time, however, the field of neurobehavioral teratol-ogy did not exist. This review focuses on the role of theretinoids within the field of neurobehavioral teratology.The retinoids hold a central place in the history of bothclassic teratology and neurobehavioral teratology. Inthis paper, the focus will be on the major issues withinthe field of neurobehavioral teratology that have beenaddressed as it has evolved across the last five decades.A primary goal is to commemorate past accomplish-
ments by presenting organizing themes that directedand defined the field during each decade, while alsoshowing how the field has changed over time withrespect to issues and approaches. It is necessary to beginwith a delimiting definition of neurobehavioral teratol-ogy as the science that deals with the abnormal develop-ment of the structure and the behavioral functions ofthe CNS resulting from exposure to exogenous agentsduring prenatal development. This definition has the
*Correspondence to: Jane Adams, Department of Psychology, 100 MorrisseyBlvd, University of Massachusetts–Boston, Boston, MA 02125.E-mail: [email protected] online 23 September 2010 in Wiley Online Library (wileyonlinelibrary.com).DOI: 10.1002/bdra.20721
Birth Defects Research (Part A): Clinical and Molecular Teratology 88:895–905 (2010)
� 2010 Wiley-Liss, Inc. Birth Defects Research (Part A) 88:895�905 (2010)

advantage of incorporating both structure and behav-ioral functioning within the framework of study and ofdelimiting the time of insult to the prenatal period, thusbeing in keeping with the timeframe of classic teratol-ogy. Two major disadvantages, however, are the failureto capture the fact that development of the CNS extendsbeyond birth and undergoes a second major wave dur-ing adolescence (Dahl, 2004; Sisk and Foster, 2004;Spear, 2007) and the fact that the CNS undergoesdynamic changes associated with neurogenesis, synapticremodeling, and regulation of signaling molecules andreceptors throughout life (Zhao et al., 2008; Deng et al.,2010; Shruster et al., 2010). Putting these caveats aside,this review will focus on the effects of exposure to reti-noids during the prenatal period and therefore will takean approach that aligns the purview of classic and neu-robehavioral teratology most closely. For readers seek-ing a broader understanding of the field of neurobeha-vioral teratology and/or retinoid-specific neurobehavio-ral teratology, detailed information can be found inreviews by Butcher (1985), Vorhees (1986a, 1986b), Nel-son (1990), Adams (1993), and Adams and Holson(1998).
There is an abundance of studies of the retinoids inrelation to the effects of both deficits and overactivity ofretinoid signaling. By necessity, in this paper I willfocus on excess conditions originally defined by studiesof hypervitaminosis A and later by studies of the excessadministration of specific retinoid compounds. This wasa decision of convenience based on the particularresearch focus of my past work and the impossible chal-lenge of doing both. To be comfortable excluding thesignificant work on vitamin A deficiency, however, I amcompelled to make a few points in that regard. First,studies of vitamin A deficiency have a critical and sub-stantial history within the field of classic teratology,although they have received lesser attention in neurobe-havioral teratology (Kalter, 1968; Clagett-Dame andDeLuca, 2002). Although early work emphasized hydro-cephalus as an outcome, newer studies demonstratethat abnormalities of the hindbrain and selected cranialnerves are also a result of gestational vitamin A defi-ciency. Second, from an applied point of view withrespect to maternal and child health, vitamin A defi-ciency represents a much larger problem. According tothe World Health Organization, vitamin A deficiencythreatens maternal and child health in more than half ofthe countries of the world, particularly low-incomeareas in Africa and Southeast Asia (WHO, 2010). Itranks as the number one cause of preventable blindnessworldwide. Given the continuum of expression of de-velopmental and behavioral deficits, this characteriza-tion likely underrepresents the spectrum of adverseeffects. The explosion of research in the last decade onthe role of retinoids in CNS functioning also brings con-cern about additional cognitive and psychiatric sequelaeassociated with both deficiency and excess during de-velopment and throughout life. These will be addressedin the final section of this review.
The present review will first outline the impressiveknowledge that was established before 1960: knowledgethat set the stage for the contributions of later years.Then the primary issues and accomplishments of each
subsequent decade will be presented. Finally, currentand future issues will be addressed.
THE STATE OF THE SCIENCE PRIOR TO 1960:ESTABLISHING TERATOLOGY AS A
DISCIPLINE
Kalter has attributed the start of experimental mamma-lian teratology to the 1930s and noted remarkable pro-gress in the intervening years leading up to the 1960s:‘‘Thirty years have passed since then. Unthought-of sci-entific progress has occurred and already seems com-monplace. The field of experimental mammalian teratol-ogy has won a modest place; and this recognition is dueto the work of a small group of people who erected a setof guiding principles and thereby created a new biologicdiscipline’’ (Kalter, 1968).Studies of vitamin A deficiency played an important
role during those 30 years of remarkable progress, whileresearch on the effects of vitamin A excess was minimal.It is important to recognize that all of this work was con-ducted as basic science studies, and it was beautifullyand systematically crafted, despite technical limitationsof the day. It is also important to recognize that the zeit-geist of this pre-thalidomide period was that the placentaprotected the fetus and that although the development ofnonmammalian species was vulnerable to environmentalinfluences, this was not the case for mammals, and cer-tainly not humans (Murakami and Kameyama, 1965; Kal-ter, 1968).Prior to 1960, extensive research using radiation, try-
pan blue, hypoxia, corticosteroids, uracil, salicylic acid,and a variety of nutrient deficiencies including vitamin Aidentified most of the major scientific issues that are withus today, and provided descriptive data that allowed themajor principles of the field to be put forth in later years.Among the issues that this breadth of early studies(Kalter, 1968) examined were the following: (1) the influ-ence of gestational stage on type and incidence of majormalformations, (2) the influence of dose amount andadministration by single, multiple, or continuous treat-ment, (3) influences of route, (4) the influence of vehicle,(5) agent-specific effects, (6) the influence of genotype atlevel of species and strain differences, (7) the morphogen-esis of certain malformations and their change in mani-festation over time (exencephaly to anencephaly), (8) thedifferential morphogenesis of malformations induced byone agent versus another, (9) morphologic distinctionsbetween spontaneous and induced malformations, (10)the influence of season on malformation rates, (11) theinfluence of uterine position on malformation rates, and(12) the role of maternal age. Thus, the primary issueswere the effects of stage, the malformations relevant tospecific stages, agent and administration effects, other ex-ogenous influences, and the influence of genotype. Therichness of these questions and the care, innovation, andlabor intensity inherent in the experimental designsneeded to address them are impressive and provide aninspiring tribute to the creativity and dedication of thefounders of classic teratology.With respect to the neural aspects of the neurobehavio-
ral teratology of hypervitaminosis A, the earliest researchcan be attributed to the foundational knowledge aboutmalformations of the CNS that emerged from early work
896 ADAMS
Birth Defects Research (Part A) 88:895–905 (2010)

on the effects of hypervitaminosis A by Cohlan (1953,1954) and Giroud and Martinet (1955). These investiga-tors demonstrated that hypervitaminosis A induced anarray of malformations in rats that included exencephaly,hydrocephalus, and spina bifida. This work laid the foun-dation for a flurry of activity in studies of excess vitaminA that emerged in the 1960s
THE 1960s: CNS MALFORMATIONS INDUCEDBY MATERNAL HYPERVITAMINOSIS A
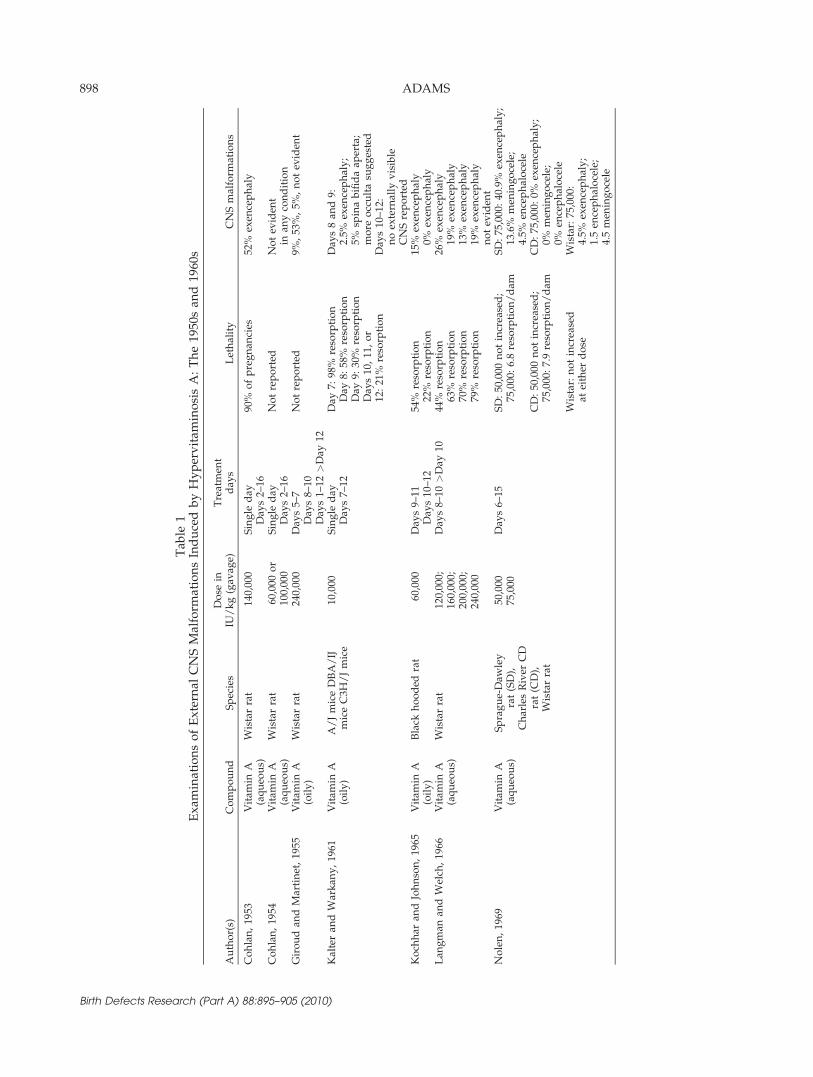
Table 1 shows studies from the 1950s and 1960s thatfocused on the CNS malformations induced by excessvitamin A following oral treatment. Kalter and Warkany(1961) opened the decade with research examining theeffects of hypervitaminosis A across three inbred strainsof mice. With respect to inter- and intralitter variability,Kalter and Warkany stressed the importance of usinginbred strains to best examine genetic sources of varia-tion and indicated the availability of 200 inbred strains. Itis perhaps worth noting that today’s market providesover 475 inbred strains of mice and over 200 inbredstrains of rats, not to mention the many individually gen-erated transgenic and knockout mice (MGI et al., 2010).Although Kalter and Warkany (1961) failed to demon-strate marked strain differences, they contributed veryimportant work indicating the stage and dose-specificmanifestation of malformations as well as species differ-ences between rats and mice. CNS malformations follow-ing treatment on days 8 or 9 (and not day 11) werereported with respect to the externally visible malforma-tions, exencephaly and spina bifida, and microscopicevaluations of sectioned tissue were also described in ele-gant detail. Kalter and Warkany described a ‘‘crowdedbrain’’ defect: ‘‘One other defect of the CNS seen in sec-tioned material can probably best be called ‘crowdedbrain.’ In this defect, the brain appeared too large for thecranium, with decrease or obliteration of the basal, peri-cerebellar, and perimedullary cisternas and reduction insize or complete closure of the rostral region of the aque-duct.’’ This may indicate early recognition of disruptionof the anterior-posterior patterning of CNS developmentas noted in later studies.
In 1969 Nolen focused on the effects of hypervitamino-sis A on days 6–15 of gestation in three strains of rats.He reported striking differences across strains in the ratesof resorptions and malformations as well as the types ofmalformations. With respect to the CNS, Nolen reporteddifferences in rates of exencephaly, encephalocele, andmeningocele, thus also indicating changes in terminol-ogy. Additional studies of the decade are also shown inTable 1. The work of the period clearly demonstratedthat excess vitamin A induced CNS malformations pri-marily between days 8 and 10 in the rat, and illustratedstrain as well as species differences in the expression ofgross CNS malformations.
Kalter and Warkany (1961) emphasized two importantissues that were just beginning to be recognized: (1) dif-ferent teratogens administered at the same developmen-tal stage can produce different malformations and (2) thepresence of litter-to-litter as well as intralitter variabilityin the severity and frequency of malformations. Discus-sion by Murakami and Kameyama (1965) on the firstissue also illustrated the growing recognition of the needto overturn the then predominant view that vulnerability
specific to stage determined the resulting malformationsas opposed to compound specific action. In Kalter’s clas-sic book published in 1968, he confirmed the manifesta-tion of agent-specific effects and stated that ‘‘all means ofcreating embryonic deviations are not equivalent . . . in alarge measure, each agent or class of agents probablycauses a specific defect or combination of defects.’’Although much of the research focused on stage and
dose relationships, attention to mechanisms was also evi-dent in a study examining how the variability in malfor-mations might be modulated through the influence ofhormones. Cohlan and Stone (1961) examined influencesof growth hormone, parathyroidectomy, thymectomy, ad-renalectomy, cortisone, or insulin administration on theCNS malformations following excess vitamin A. In thecontext of reliable induction of CNS malformations, thesemanipulations did not demonstrate any potentiating orameliorating effects at the doses used, but the work isimportant as representative of an early attempt to under-stand physiologic influences.With a body of work clearly demonstrating that hyper-
vitaminosis A reliably induced gross CNS malformations,the foundation was set for the introduction in the 1970sof investigations of behavioral effects at doses compatiblewith survival. In the remainder of this paper, attentionwill focus on the evolution of neurobehavioral teratologythrough the lens of research on retinoid compounds.First, however, it is critical to describe the broader con-text at the initiation of experimental studies in this incipi-ent field.
BEYOND THE RETINOIDS: THE SCIENTIFICCONTEXT IN WHICH STUDIES OF
BEHAVIORAL TERATOLOGY BEGAN
The critical historical events that are identified in mostreviews as having fertilized the growth of classic teratol-ogy and seeded the birth of neurobehavioral teratologyare knowledge of the relationships between exposure torubella, thalidomide, methymercury, and alcohol and con-sequent malformations in the newborn. Each of these com-pounds was identified as producing effects on CNS func-tioning as well. Gregg (1941) reported that rubella pro-duced blindness and deafness among its outcomes, andlater work described mental retardation (Warkany, 1971).Lenz (1961) described the effects of thalidomide andincluded sensory dysfunction on the list. Minamata dis-ease (Matsumoto et al., 1965) associated methylmercuryexposure with infant characteristics resembling cerebralpalsy with associated mental retardation (Nishimura,1979). Early research on the effects of alcohol during preg-nancy (Lemoine et al., 1968; Ulleland, 1972; Jones andSmith, 1973) all described CNS functional disturbances ashallmark to the syndrome of effects. It was customary torecognize effects on the CNS as among the sequelae thatdescribed human malformation syndromes associatedwith exposure to these agents, and the full syndrome wasdeemed necessary to define adverse outcomes followingexposure. In 1966 FDA introduced guidelines to screen forreproductive and developmental effects on fertility,embryotoxicity, teratogenicity, intrauterine growth, partu-rition, lactation, and early postnatal development (Hoar,1984). The three-segment design did not include measure-ment of CNS functional integrity, however. Shortly after
897CNS AND BEHAVIORAL TERATOLOGY OF RETINOIDS
Birth Defects Research (Part A) 88:895–905 (2010)
Tab
le1
Exam
inationsofExternal
CNSMalform
ationsInducedbyHypervitam
inosisA:The1950san
d1960s
Author(s)
Compound
Species
Dose
inIU
/kg(gav
age)
Treatmen
tday
sLethality
CNSmalform
ations
Cohlan,19
53Vitam
inA
(aqueo
us)
Wistarrat
140,00
0Single
day
Day
s2–
1690
%ofpregnan
cies
52%
exen
cephaly
Cohlan,19
54Vitam
inA
(aqueo
us)
Wistarrat
60,000
or
100,00
0Single
day
Day
s2–
16Notreported
Notev
iden
tin
anycondition
Giroudan
dMartinet,19
55Vitam
inA
(oily)
Wistarrat
240,00
0Day
s5–
7Day
s8–
10Day
s1–
12>Day
12
Notreported
9%,53
%,5%
,notev
iden
t
Kalteran
dW
arkan
y,19
61Vitam
inA
(oily)
A/JmiceDBA/IJ
miceC3H
/Jmice
10,000
Single
day
Day
s7–
12Day
7:98
%resorption
Day
8:58%
resorption
Day
9:30%
resorption
Day
s10
,11
,or
12:21
%resorption
Day
s8an
d9:
2.5%
exen
cephaly;
5%sp
inabifidaap
erta;
more
occultasu
ggested
Day
s10
–12:
noexternally
visible
CNSreported
Koch
har
andJohnson,19
65Vitam
inA
(oily)
Black
hooded
rat
60,000
Day
s9–
11Day
s10
–12
54%
resorption
22%
resorption
15%
exen
cephaly
0%exen
cephaly
Lan
gman
andW
elch
,19
66Vitam
inA
(aqueo
us)
Wistarrat
120,00
0;16
0,00
0;20
0,00
0;24
0,00
0
Day
s8–
10>Day
1044%
resorption
63%
resorption
70%
resorption
79%
resorption
26%
exen
cephaly
19%
exen
cephaly
13%
exen
cephaly
19%
exen
cephaly
notev
iden
tNolen,19
69Vitam
inA
(aqueo
us)
Sprague-Daw
ley
rat(SD),
CharlesRiver
CD
rat(C
D),
Wistarrat
50,000
75,000
Day
s6–
15SD:50
,000
notincreased;
75,000
:6.8resorption/dam
SD:75
,000
:40
.9%
exen
cephaly;
13.6%
men
ingocele;
4.5%
encephalocele
CD:50,000
notincreased;
75,000
:7.9resorption/dam
CD:75
,000
:0%
exen
cephaly;
0%men
ingocele;
0%en
cephalocele
Wistar:notincreased
ateither
dose
Wistar:75
,000
:4.5%
exen
cephaly;
1.5en
cephalocele;
4.5men
ingocele
898 ADAMS
Birth Defects Research (Part A) 88:895–905 (2010)

the passage of these guidelines, experimental behavioralteratology began in the 1970s The emergent fieldmet much resistance. Scientists within the field of classicexperimental teratology had no training in the behavioralsciences, and most did not understand or find it crediblethat animal studies could be used to model humanbehavior. Hoar (1984) commented that the screeningguidelines were criticized with respect to their ability todetect human hazard with respect to physical malforma-tions, while ‘‘the continuing controversy of the rele-vance of animal data to humans was never more stri-dent than in the area of behavioral teratology, and thisnihilism has significantly delayed both the developmentand understanding of these important measures ofaltered central nervous system function.’’ Behavioral ter-atology began at a time when screening guidelines werein place that identified doses associated with teratoge-nicity as well as maternal toxicity and assumed thatlower doses were ‘‘safe.’’ In introducing the scientificfield of study, early behavioral teratologists ferventlysought to create and demonstrate a field with regulatoryand scientific significance. The early researchers set outto demonstrate that certain compounds at doses belowthose that caused maternal toxicity or teratogenicitycould nevertheless have adverse effects on CNS devel-opment that had functional consequences.
THE 1970s: BEHAVIORAL STUDIES BEGIN
With respect to research on the retinoids, one of thefirst studies that examined the behavioral consequencesof prenatal exposure to hypervitaminosis A was con-ducted by Butcher and colleagues (1972). They demon-strated that ‘‘excess vitamin A administered in quantitiestoo low to yield gross malformations of the central nerv-ous system produces behavioral impairments in the off-spring.’’ This statement is somewhat of a mantra thatcaptures the intent (and the battle) of the early studies.
Table 2 shows the characteristics of studies conductedon the effects of hypervitaminosis A on behaviors meas-ured during this early decade. It was typical to measurebody weight and developmental milestones such as
reflex development, and to observationally measuremotor activity and simple forms of learning such aslearning to escape from water mazes, to avoid shock, orto acquire food. Some measures were quite simplistic bytoday’s standards. A convergence of evidence, however,clearly demonstrated the detection of behavioral effectsat subteratogenic doses. As a field, Behavioral Teratologyemerged in this decade based on research primarily onvitamin A and on alcohol, methylmercury, and phenyt-oin, all recognized as compounds with effects on thehuman CNS. Among researchers in the field, attentionfocused on teratology’s most salient relationships: dose-response and stage-response relationships. Additionally,the behavioral scientists were interested in developing orapplying methods known to index the many categoriesof behavior (motor development, sensory developmentand ability, activity level, reactivity to stimulation, learn-ing and memory abilities) and in determining the rela-tionships between underlying structures and behavioraldeficits. The 1980s became a decade focused, in part, onthese issues.The simple and robust empirical studies of the 1970s
provoked strong reactions. Prior and concurrent researchon gross CNS malformations had examined the abnor-malities as present or absent, and when absent, normalityof CNS development had been accepted as evidence ofsafety within the regulatory environment. The data frombehavioral teratology studies met with skepticism andcriticism that challenged the reliability and validity of themethodologies used and questioned research interpreta-tions. This redirected the focus of much of the laterresearch from structure-function relationships to the reli-ability and validity of the behavioral methods used: afocus that has been consistently addressed to the present(Makris et al., 2009). In 1979 the results of the first intra-and interlaboratory comparison study were published(Butcher et al., 1979). This study demonstrated the consis-tency in the data from a maze learning task that was runin three cooperating laboratories. For researchers whoworked as regulatory scientists, attention to the reliabilityof methods became a dominant focus of the next decadeas behavioral scientists fought for recognition of the im-
Table 2Behavioral Teratology Studies of Hypervitaminosis A in the 1970s
Authors SpeciesDose in
IU/kg (gavage)aTreatment
daysBehaviorevaluated
Effectspresent
Butcher et al., 1972 Sprague-Dawley rat 100,000 Days 8–10 Water maze YesHutchings et al., 1973 Wistar rat 240,000 Days 13–14 Operant learning YesHutchings and Gaston, 1974 Wistar rat 360,000 Days 16–17 Operant learning YesVorhees, 1974 Fischer 344 rat 10,000
25,00040,000
Days 8–10 Open field activity,shock avoidance learning
No, Yes,all groups
Vorhees et al., 1978 SpragueDawley rat
80,000 Days 5–7Days 8–10Days 11–13Days 14–16Days 17–19
T maze activity,open field activity,Biel maze
Yes, 11–13Yes, 8–10Yes, 8–10, 11–13,and 17–19
Vorhees et al., 1979 SpragueDawley rat
40,000 Days 7–20 Motor development YesOpen field activity YesBiel maze YesActive avoidance learning YesPassive avoidance learning NoRotorod motor skill Yes
aIn all studies, an oily solution of vitamin A was used.
899CNS AND BEHAVIORAL TERATOLOGY OF RETINOIDS
Birth Defects Research (Part A) 88:895–905 (2010)

portance of the research for human health protection. Forbasic scientists, the skepticism and criticism among thebiologic scientists within the field of teratology wasattributed to a misunderstanding of behavioral science asa discipline adherent to exceptionally rigorous methodo-logical design and detail. Indeed, the natural and desira-ble variation in behavior acted historically to promulgatethe need for rigorous experimental methodologies anddesigns that allow behavioral researchers to detect clearsignals in the midst of the expected variation inherent innormal behavior. Whereas anatomic and physiologic end-points have a narrow range of variation that does notdemand elegant control over a multitude of environmen-tal variables, behavior, if measured correctly and withappropriate sensitivity to its expected considerable rangeof variation, demands control over many variables. Thisfundamental difference across disciplines continues tocontribute to a misunderstanding that appropriately con-ducted behavioral measurements are reliable, particularlywhen reliability is not apparent in replication studiesdesigned, conducted, or interpreted by inappropriatelytrained experimenters (Tyl et al., 2008). This disciplinaryconflict, the rapid emergence of research activity inbehavioral teratology in the 1970s, and the need for acentralized forum contributed to the formation of theBehavioral Teratology Society in 1977. The BehavioralTeratology Society, now known as the NeurobehavioralTeratology Society, has been a sister society of the Tera-tology Society since that time and has continued to holdits annual meetings in conjunction with those of the Tera-tology Society.
While the behavioral research on hypervitaminosis Awas beginning, the 1970s evinced little activity withrespect to the CNS teratology of retinoids. Examinationsof other malformations and effects across species werequite active and revealed the importance of structure-ac-tivity relationships across species and strains. All-trans-retinoic acid, a naturally occurring congener, was identi-fied as the putative agent of teratogenesis (Kochhar et al.,1984). In the interest of improved quantification of dosesin light of quality control concerns associated with thevarious sources of vitamin A, and the species-specific dif-ferences in pharmacokinetics, researchers began to utilizeall-trans-retinoic acid as the administered compound interatology studies. Nolen (1972) reported that a singledose of 10 mg/kg all-trans-retinoic acid on gestation day9 in Charles River Sprague Dawley CD rats increasedresorption rates and caused a low incidence of exence-phaly. He also reported cleft palate, eye defects, and skel-etal defects in the animals.
THE 1980s: A BURST OF EXPERIMENTAL ANDCLINICAL ACTIVITY
During the 1980s, vitamin A studies as well as behav-ioral teratology research in general saw a burst of activ-ity. The research on vitamin A was primarily focused onits use as a positive control in development of methodsand methodological evaluation studies. The one excep-tion was an extensive study by Nolen (1986). In this mas-sive undertaking, Nolen conducted three experimentsthat examined the effects of dose and stage at treatmenton body weight and multiple measures of behavior,including reflex development, learning, and activity. InExperiment 3, he also examined amphetamine-stimulated
activity. In the first experiment, he examined the effectsof 5 mg/kg all-trans-retinoic acid administered toSprague Dawley rats on gestation days 8–10, 11–13, or14–16. He identified days 11–13 as a period of vulnerabil-ity for reduced postnatal survival, reporting lethality fol-lowing dosing at that stage. During the prior interval,days 8–10, this subteratogenic dose had no effect onbody weight but did alter reflex development and activ-ity. Treatment on days 14–16 reduced weight and alteredreflex development, activity, and learning performance.In Experiment 2, he administered 2.5 or 5 mg/kg ondays 11–13 or 14–16 and reported behavioral effects atthe lower dose following both stages of administration.The higher dose on days 11–13 reduced survival, whileat days 14–16 it did not affect survival or weight but didalter reflex development, activity, and learning perform-ance. Experiment 3 examined the effects of 2, 4, or 6 mg/kg administered on days 14–16. No adverse effects wereseen at the lowest dose. The 4 mg/kg dose did not affectoffspring body weight but did alter reflex development,activity, and amphetamine-challenged activity. The high-est dose reduced the weight of males and altered allbehaviors examined: reflex development, two differentmeasures of activity, amphetamine-challenged activity,and two different measures of learning. This importantwork demonstrated that 5 mg/kg doses caused behav-ioral disruption following treatment on gestations days8–10 and were lethal when given on gestation days 11–13, when a 2.5 mg/kg dose disrupts postnatal behavior.On gestation days 14–16, doses of 2.5 up to 6 mg/kg pro-duced marked behavioral alterations.In 1993 Adams synthesized this work and other rat
neurobehavioral studies from the 1970s and 1980s thatused treatment with retinol or all-trans-retinoic acid toexamine structure-activity relationships and basic princi-ples within neurobehavioral teratology. The evidenceindicated that postnatal growth and behavioral altera-tions were altered by lower doses than malformation orpostnatal death following treatments by retinol, all-trans-retinoic acid, and 13-cis-retinoic acid. Collectively, gesta-tional days 9–13 were more sensitive to treatment effectson postnatal endpoints, with days 11–13 being particu-larly sensitive (effects occurred at lower doses). Compar-ing across endpoints of death, CNS malformation,growth alteration, and behavioral dysfunction, Adams(1993) demonstrated that behavioral effects were pro-duced by retinol and all-trans-retinoic acid at doses lowerthan those necessary to produce effects on growth. Withrespect to structure-activity relationships, consistent dif-ferences were seen across the retinoid compoundsregarding the most sensitive endpoints and gestationalstages. A four- to sixfold potency differential was seenbetween retinol and all-trans-retinoic acid for each of theoutcomes of death, CNS malformation, growth reduction,and behavioral dysfunction. At that time, Kochhar andcolleagues (1984) had shown that retinol had one-fourththe potency of all-trans-retinoic acid in the teratogenicproduction of cleft palate and limb defects in mice. Thedata were interpreted as arguing for a common agentthat disrupted various developmental processes relevantto these diverse endpoints.Most of the other studies of the 1980s employed vita-
min A or all-trans-retinoic acid as a positive test com-pound to produce behavioral changes that could be eval-uated in studies focused on reliability of methods across
900 ADAMS
Birth Defects Research (Part A) 88:895–905 (2010)

replicates and within and across laboratories. These stud-ies sought to demonstrate that behavior could be meas-ured reliably and reproducibly, therefore deserving aplace in drug and chemical screening to protect humanhealth. Makris (2009) has recently reviewed the history ofscreening in developmental neurotoxicology anddescribed multiple collaborative efforts undertaken byU.S., Japanese, and European investigators. She con-cludes that the international scientific peer-reviewedwork attests to the reproducibility, reliability, and sensi-tivity of the methods contained in the EPA’s develop-mental neurotoxicity testing guidelines. It is hoped thatthis now represents the informed, consensus-based scien-tific judgment for work performed with appropriateattention to variables of relevance to reliable measure-ment.
Early criticisms of behavioral teratology as an experi-mental science were twofold, however, with concernsabout methodological reliability voiced alongside equallystrong concerns about the ability of rodent behavior toserve as a model for human behavior. In the 1980s, theintroduction of 13-cis-retinoic acid as the active ingredi-ent of the drug Accutane unfortunately permitted thisissue to be addressed with respect to the retinoids. Thepharmaceutical use of this and other synthetic retinoidsalso stimulated a great deal of research that examinedthe differential teratogenic actions of natural and syn-thetic retinoids across species.
The Introduction of Accutane
Accutane, 13-cis-retinoic acid (isotretinoin), entered theU.S. market in 1982 for the treatment of severe cysticacne. In 1985, Lammer et al. reported the findings of thefirst prospective study that clearly defined its high terato-genic potency. Risks associated with human exposure tothe therapeutic dose during embryogenesis werereported to be 40% for spontaneous abortion, 4–5% forperinatal mortality, 16% for premature birth, and 25% formajor malformation. The characteristic pattern of malfor-mations included craniofacial, cardiac, thymic, and CNSstructures. Major malformations of the CNS were themost frequently occurring category of malformation andwere largely defined by hindbrain abnormalities repre-sented by cerebellar hypoplasia, agenesis of the vermis,abnormalities of the pons and medulla, and malforma-tion-associated hydrocephalus, although cortical andother abnormalities were also reported (Lammer et al.,1985, 1988). The most striking craniofacial features wereear malformations and cranial motor nerve (oculomotorand facial) abnormalities, which were typically asymmet-ric. Longitudinal follow-up of the children, both withand without major malformations, indicated that at5 years of age, 47% of the children had mental abilityscores in the borderline to mentally retarded range onthe Stanford-Binet IV test of intelligence (Adams andLammer, 1991, 1993). This striking teratogenic potencywas not anticipated, because earlier as well as laterresearch documented that in rodent models, isotretinoinhad lower teratogenic potency than all-trans-retinoic acidand other retinoids (Kochhar et al., 1984; Kochhar andPenner, 1987; Kraft et al., 1987; Kochhar et al., 1988; Kluget al., 1989). Notably, however, isotretinoin is teratogenicin humans at the therapeutic doses of 0.5–1.5 mg/kg/day that are employed in its treatment regimen. In a Ter-
atology Society Position Paper on the use of retinoidsduring pregnancy (1987), the lowest teratogenic doses(mg/kg/day) of isotretinoin were reported as 0.4 forhumans, 5.0 for subhuman primates, 150.0 for the rat (re-vised to 75.0 in Anonymous, 1987), 100.0 for the mouse,25.0 for the hamster, and 10.0 for the rabbit. These dis-parate potencies across species are in stark contrast to therelative compression across species for all-trans-retinoicacid, which were reflected in the lowest teratogenic dosesreported as 7.5, 0.4–2.0, 4, 12.5, and 2–10 across subhu-man primates, rat, mouse, hamster, and rabbit, respec-tively. Considerable research was later directed at under-standing the maternal, placental, and fetal metabolicevents underlying these differences (Nau, 2001). Whileinitially proposed to be based largely on species differen-ces in the extent of isomerization of the parent com-pound to all-trans-retinoic acid, Nau concluded that thiswas but one of several critical factors.With respect to issues relevant to screening in the reg-
ulatory context, the retinoid-induced CNS malformationsand behavioral, particularly learning-related, impair-ments in humans provided ample validation that effectsin rodents could indeed be relevant to humans. Simulta-neously, however, the metabolic and teratogenic differen-ces challenged simple formulas for the determination ofsafe levels of human exposure and demanded attentionto risk estimation based on data from several species. Ofparamount importance in the regulatory context, how-ever, was the challenge to regulatory intent provoked bythe presence of a drug used to treat a non-life-threateningcondition that was demonstrated to be a human terato-gen. Although a new drug might be precluded fromcoming to market if teratogenicity was of prior knownconcern, once marketed and known to be teratogenic, theappropriate action was less clear and considerably morecomplicated. Opinions differed as to whether such adrug should be pulled from the market such as wouldoccur if a cardiac arrhythmia or other adult life-threaten-ing effect were observed. This issue, the consequent regu-latory activities, and the current occurrences of isotreti-noin-exposed pregnancies are important issues of whichthe reader should be aware; they are, however, beyondthe scope of this paper (Mitchell et al., 1995; Honeinet al., 2001; Perlman et al., 2001; Brinker et al., 2005; Rob-ertson et al., 2005).With respect to validation of principles of neurobeha-
vioral teratology that were established based on experi-mental findings in animal models, knowledge of theeffects of embryonic exposure to isotretinoin on child-hood behavior had considerable value. Our resultsclearly showed that while intellectual deficits accompa-nied frank brain malformation and other physical mani-festations of isotretinoin embryopathy, intellectual deficitscould also be induced in children without physical signsof abnormal development (Adams and Lammer, 1991,1993). This fact irrefutably argues that efforts to safe-guard the human population by identifying the lowestadverse effect levels of chemical exposures during preg-nancy and early development must include measurementof effects on postnatal behavioral development.The burst of activity in the 1980s provided further
foundation for the growth and elaboration of neurobeha-vioral teratology in the 1990s. Although attention to thereliability and reproducibility of methods continued tomeet the goals of implementation of screening require-
901CNS AND BEHAVIORAL TERATOLOGY OF RETINOIDS
Birth Defects Research (Part A) 88:895–905 (2010)

ments (Makris et al., 2009), the 1990s also ushered instudies directed at using rodents to model what had nowbeen described in humans, and to better understand thepotency of exposure effects, as well as the relationshipsbetween effects in brain and on behavior. A colossalamount of research activity explored the nuclear actionsof retinoids and their prominence as morphogens in ver-tebrate development. During the 1990s, these studiesextended our scientific vocabularies to include sonichedgehog, HOX genes, and knockouts as commonplaceterms. Highlights of selected studies are discussed.
THE 1990s: MORE IN-DEPTHUNDERSTANDING OF MECHANISMS AND
MANIFESTATIONS OF RETINOIDNEUROBEHAVIORAL TERATOGENICITY
Research aimed at illuminating the factors that contrib-uted to species differences in the teratogenic activity ofdifferent retinoid congeners occurred at the same time asa substantial amount of basic scientific research focusedon the role of retinoids as controllers or contributors toneural differentiation, and even larger amounts ofresearch on the role of retinoids in other aspects of verte-brate development. During the 1990s, PubMed indicates1748 papers on the role of retinoids in vertebrate devel-opment and 463 on retinoic acid teratology. In this bodyof work are studies on insects, amphibians, and mam-mals using methods from whole embryo culture toexaminations of teratogenicity in non-human primates.Here I will present the highlights of the work on the roleof retinoids in CNS development and provide moredetailed information on the neurobehavioral teratologystudies of the period.
The Role of Retinoids in Brain Development
The demonstration of CNS abnormalities followinghuman exposure to 13-cis-retinoic acid stimulated a greatdeal of research on the role of retinoids in brain develop-ment. During the 1990s insights and breakthroughs inunderstanding occurred. Some of the principal undertak-ings and advancements of relevance to teratology (Mar-shall et al., 1996; Durston et al., 1998; Maden et al., 1999)included the following: (1) delineation of the biologicconversion of retinol to various metabolites and the iden-tification of the critical enzymes; (2) investigations of thedistribution of enzymes and retinol isomers in embryonicneural tissues; (3) detection of retinoid receptors in thedeveloping neural plate; (4) descriptions of contributoryroles of retinoids in neural patterning via regulation ofhox genes relevant to the development of the anteriorhindbrain and critical to the development of the posteriorhindbrain; (5) identification of retinoic acid receptorligands of relevance to axial patterning; (6) demonstra-tion that certain RAR and Hoxa-1 knockouts had rhom-bencephalic abnormalities; (7) demonstrations that hind-brain abnormalities also occurred when active retinoidligands were depleted; and (8) indication that a retinoidgradient regulated the development of the hindbrainalong an anterior-posterior axis. These huge leaps for-ward represented major contributions from embryology,genetics, and teratology in the 1990s decade of the brain.
Neurobehavioral Teratology in the 1990s
The onset of the human isotretinoin tragedy provokedstudies of teratogenicity in non-human primates, invigo-rated experimental studies on the behavioral effects ofexposure to all-trans-retinoic acid, the most potent terato-gen within the rodent system, and motivated rodent be-havioral studies of isotretinoin. In a study of isotretinointeratogenicity conducted in cynomolgus monkeys(Hummler et al., 1990), several features similar to thehuman syndrome were reported, including ear, thymic,and cardiovascular abnormalities following a dose of 2.5mg/kg prior to and during early organogenesis (gesta-tion days 10–27). This laboratory (Makori et al., 1998)also reported effects on hindbrain and craniofacial devel-opment following a similar treatment regimen.Research in rats was undertaken to examine periods of
sensitivity, to examine structure-activity relationshipsacross several endpoints, and to further examine behav-ioral changes following prenatal exposures. The vulnera-ble period for postnatal lethality described for all-trans-retinoic acid following a 5 mg/kg dose on days 11–13 inSprague-Dawley rats (Nolen, 1986) was also reported fol-lowing exposure to 125 or 150 mg/kg of 13-cis-retinoicacid to Wistar rats on gestation days 11–13 (Eskessenet al., 1990). Holson et al. (1997a, 1997b, 1999) furtherexamined the effects of exposure to all-trans-retinoic acidin Sprague Dawley rats on postnatal survival, weight,and behavior and replicated the day 11–13 period of sen-sitivity as compared to earlier or later treatment. Theyshowed dose-related effects on postnatal survival ofSprague-Dawley rats treated with 2.5, 5.0, or 10 mg/kgon days 11–13 with reductions in postnatal survival of38, 90, or 100%, respectively, in association with noeffects on litter size at birth. Holson et al. (1997c)reported severe effects on the early development of suck-ling reflexes and breathing patterns prior to the death ofpups exposed to the 10 mg/kg dose. Effects on bodyweight and behavior were seen at the lower dose of 2.5mg/kg dose in studies that examined activity and learn-ing by Holson and colleagues (Holson et al., 1999). Inthese rats, effects on daytime running wheel activity andamphetamine-challenged open field activity were seenfollowing treatment on gestation days 11–13 and days14–16. However, neither the learning ability of water-deprived rats in a complex spatial maze (dry) nor audi-tory startle responding were affected. At this dose andstage, Nolen (1986) had reported disrupted learning abil-ities measured under shock avoidance and water escapeconditions. Holson et al. (1997a, 1997b, 1999) also exam-ined regional brain weights in eight-to-nine brain regionsand reported reduced whole brain weight and cerebellarweight as well as alterations in certain medullary nuclei.These effects show a nice parallel to the human CNSabnormalities and effects on learning would be expected.It is possible that their detection may require the addedchallenge of shock or swimming stress. The effects on ac-tivity, and particularly reduced responding to ampheta-mine-stimulated activity, were interpreted as suggestiveof alterations in the mesolimbic dopamine system conse-quent to gestational treatment on days 11–13 or 14–16.Jensh et al. (1991) showed that a 100 but not a 50 mg/
kg dose of 13-cis-retinoic acid administered on days 11–13 affected behavior without impacting survival or body
902 ADAMS
Birth Defects Research (Part A) 88:895–905 (2010)

weight. These investigators reported effects on two activ-ity measures as well as on avoidance learning, again sug-gesting added stress as a possible requisite for demon-strating effects on learning. Across studies on all-trans-and 13-cis-retinoic acid, treatment on days 11–13 provedto be a sensitive period for the induction of postnatallethality and, at lower doses, for behavioral dysfunction.
The extraordinary progress made during the 1990s ledto the characterization of multiple exposure, metabolic,ligand, and receptor activities relevant to retinoid terato-genicity and behavioral teratogenicity, as well as to therole of retinoids in normal development. Techniquesevolved from the observational and microscopic charac-terizations of gross CNS malformations during the 1970sand 1980s to detailed descriptions of tissues, tracts, andmolecular and cellular substrates. The areas in whichadvancements have been described have continued to beexplored since 2000 with definitive findings in many cat-egories. In the early twenty-first century, studies of therole of retinoids in neural differentiation and patterninghave also broadened in focus to include examinations ofinfluences on CNS functioning. Research has continuedin neurobehavioral teratology, focusing on influences onmotor system development and function. These are thestudies that will be presented as representative of workbeginning in 2000 as neurobehavioral teratology crossedinto the new millennium.
FROM 2000 TO THE PRESENT:CONSIDERATIONS OF ADDITIONAL TYPES
AND PERIODS OF VULNERABILITYStudies within Neurobehavioral Teratology
Within the specific neurobehavioral teratology arena,research by Holson and colleagues continued (2000,2001), and several studies were reported by Coluccia andcolleagues (2008a, 2008b, 2009). Holson et al. (2000)sought to better understand the cause of postnatal deathin animals exposed to all-trans-retinoic acid on gesta-tional days 11–13. Apparent difficulties in breathing andabsence of milk in the stomach of dead pups had beenreported by Holson et al. (1997c). In this study (Holsonet al., 2000), milk was infused into the mouths of neona-tal rats that had been exposed prenatally to 10 mg/kgall-trans-retinoic acid and to unexposed controls. Certainactivational behaviors were seen by both groups indicat-ing responsivity to the milk, but the exposed pupsappeared unable to swallow. This was interpreted as dif-ficulty in the motor control of swallowing.
Holson et al. (2001) examined the effects of 2.5 or 10mg/kg of all-trans-retinoic acid administered on gesta-tional days 11–13 or 14–16 to Sprague-Dawley rats and toLong-Evans (hooded) rats. They reported that bothstrains showed similar responses to exposure, althoughcertain responses were greater in hooded than in SpragueDawley rats. As in prior work, increased mortality wasseen following treatment in both periods, and hoodedrats showed a stronger effect on days 11–13 than did theSprague-Dawley rats. Reduced body weight was seenneonatally and at 35 days of age following both treat-ments, but this did not account for reduced cerebellarweight that was seen following both treatments, with astronger effect in hooded rats. Interestingly, gestationaltreatment on days 11–13 was associated with greatereffects on the vermis, and treatment on gestational days
14–16 was associated with greater effects on the weightof the cerebellar hemispheres. This study did not exam-ine behavioral sequaelae other than effects on the devel-opment of the righting reflex. Righting reflex wasdelayed following treatment on gestational days 11–13but not 14–16, and the magnitude was larger in thehooded rats. Thus, the two strains showed similarresponses to treatment, but the hooded rats appeared tobe somewhat more sensitive.Coluccia and colleagues (2008a, 2008b, 2009) examined
the effects of exposure to 2.5 mg/kg all-trans-retinoic acidfollowing treatment of Sprague-Dawley rats on gestationaldays 11–13. This design therefore followed that of Holsonand colleagues, and authors stated the goal of furtherchallenging motor performance and motor learningthrough testing under more demanding rotarod condi-tions involving higher rotation speeds. Collucia et al.(2008a) examined rotarod performance, motor strength viagrip strength, vertical and horizontal open field activity,and morphology of the cerebellum observed throughstained cryostat sections. The investigators replicated Hol-son et al.’s (1997a, 1997b, 1999) findings of increased post-natal lethality (38% death) in the absence of effects on lit-ter size at birth. Significant delays in the maturation ofearly reflexes were seen, such as delayed righting reflexdevelopment, thus also replicating this finding. Decreasedlocomotor activity and decreased emergence from the cen-ter of the open field at 90 days of age were significantcompared to the controls, but no effects on vertical rear-ing were obtained. Rotarod performance was impairedwith respect to decreased latency to fall and reducedimprovement (motor learning) across four test sessions.No differences in motor strength were reported, thus sup-porting interpretations based on motor coordination andlearning deficits. Evaluations of the cerebellum at post-natal ages of 1, 3, 8, or 40 were conducted. Characteristicsat postnatal day 1 included reductions in the size of thecerebellar vermis coupled with larger reductions in lateralsections involving folia IX–X. At day 3, differences in thesize of folia IX–X were also seen with reductions of 21.5%compared to controls. Cerebellar differences were no lon-ger significant after the first postnatal week. The differen-ces in locomotor activity, motor coordination, and motorlearning were therefore manifest at ages when morpho-logic effects on the cerebellum were no longer evident.Coluccia and colleagues (2008b) examined the effects of2.5 mg/kg all-trans-retinoic acid administered on days 8–10 using a similar behavioral testing battery (cerebellarmorphology was not examined). The treatment resulted ina mortality rate of 7.8% and delayed the righting and neg-ative geotaxis reflexes as well as altered open field activityand rotarod performance and learning. Coluccia et al.(2009) presented results for a similarly designed study inwhich treatment with 2.5 mg/kg all-trans-retinoic acidoccurred on gestational days 14–16 (cerebellar morphol-ogy was not examined). Here they reported impairedopen field activity and impaired rotarod performance aswell as learning. The authors interpreted their findings inall three studies as consistent with the effects of alteredcerebellar development and postulated effects on circuitryinvolving cortico-cerebellar connections.Studies on the Role of Retinoids in CNS Functioning.
Work outside of the field but integral to its interpretationof data and development of hypotheses for future workhas explored the role of retinoids in other psychological
903CNS AND BEHAVIORAL TERATOLOGY OF RETINOIDS
Birth Defects Research (Part A) 88:895–905 (2010)

and behavioral processes. This has been motivated, inpart, by anecdotal and scientific reports of depressionand possible suicidal behavior among isotretinoin users(Marqueling and Zane, 2007). Those studies remain con-troversial, are tangential to this review, and will not becovered. However, the emerging understanding of therole of retinoids in emotional and psychological function-ing as well as in cognitive ability and adult neuroplastic-ity are worthy of consideration. Central points emergentfrom reviews of this literature are the following: (1) reti-noic acid receptors show distinctive expression patternsin specific regions of the adult brain such as the hippo-campus, thalamus, and pons for RARa, and the striatum,hypothalamus, and medulla for RARb (Tafti and Ghyse-linck, 2007); (2) brain areas high in retinoic acid signalingand receptors overlap with areas of relevance to stressand depression (Bremmer and McCaffery, 2008); (3) reti-noid signaling is important in the developmental controlof the differentiation of striatal neurons and their neuro-chemistry (Wang and Liu, 2005); (4) retinoic acid is im-portant in the control and regulation of dopamine neuro-transmission (Tafti and Ghyselinck, 2007; Bremmer andMcCaffery, 2008); (5) Studies in Rarb- and Rxrg-null mu-tant mice show deficiencies in the animals’ spatial learn-ing and memory similar to that of vitamin A–deficientrats (Tafti and Ghyselinck, 2007); (6) vitamin A deficiencyinterferes with long-term potentiation and the establish-ment of new memories, and deficits can be reversed byretinoic acid administration (Tafti and Ghyselinck, 2007;Olson and Mello, 2010); (7) age-related changes in reti-noic acid signaling are associated with cognitive declines,and improvements follow retinoic acid treatment (Taftiand Ghyselinck, 2007); and (8) retinoic acid modulatesneurogenesis, neuronal survival, and synaptic plasticityand helps maintain neuronal plasticity and cognitivefunctioning in adulthood (Olson and Mello, 2010). Recog-nition of these vital roles of retinoids in brain functioninghave led multiple investigators to begin studies or putforth hypotheses about the importance of retinoids inaffective disorders (Bremmer and McCaffery, 2008), inAlzheimer’s disease and other neurodegenerative dis-eases of relevance to cognition (van Neerven et al., 2008),in schizophrenia (Maynard et al., 2001; Palha and Good-man, 2006), and in autism while also arguing for treat-ment approaches associated with retinoid action or inhi-bition of action.
THE FUTURE
We study the abnormal not just to identify and amelio-rate adverse effects but also to understand the normal.Can it be that the basic and applied knowledge obtainedwithin the field of neurobehavioral teratology not onlycan be used to prevent exposures and consequent behav-ioral and cognitive deficits, but also portends applicationto therapeutic use in the amelioration of neurologic andpsychiatric disease? Is it possible that a field that so farhas attended only minimally to the social and emotionalconsequences of prenatal exposure will see a preponder-ance of such studies in the near future? Will prenatal ex-posure and programming of regulatory systems emergeas a fundamental cause of adult disease? Is it possiblethat the tragedy associated with isotretinoin embryopathyhas spurred research that may bear on the facilitation of
healthy and robust cognition and psychological well-beingthroughout the lifespan? Addressing these questions willno doubt be focal points in the decades to come.
REFERENCES
Adams J. 1993. Structure-activity and dose-response relationships in theneural and behavioral teratogenesis of retinoids. Neurotoxicol Teratol15:193–202.
Adams J, Holson RR, editors. 1998. The neurobehavioral teratology ofVitamin A analogs. In: Handbook of Developmental Neurotoxicol-ogy. Academic Press: San Diego. pp. 631–642.
Adams J, Lammer E. 1991. Relationship between dysmorphology and neu-ropsychological function in children exposed to isotretinoin ‘‘in utero.’’In: Fujii T, Boer GJ, editors. Functional neuroteratology of short termexposure to drugs. Tokyo: Teikyo University Press. pp. 159–170.
Adams J, Lammer EJ. 1993. Neurobehavioral teratology of isotretinoin.Reprod Toxicol 7:175–177.
Anonymous. 1987. Teratology Society Position Paper: recommendationsfor vitamin A use during pregnancy. Teratology 35:269–275.
Bremmer JD, McCaffery P. 2008. The neurobiology of retinoic acid inaffective disorders. Prog Neuro-Psychoph 32:315–331.
Brinker A, Kornegay C, Nourjah P, et al. 2005. Trends in adherence to arevised risk management program designed to isotretinoin-exposeddecrease or eliminate pregnancies: evaluation of the AccutaneSMART program. Arch Dermatol 141:563–569.
Butcher RE. 1985. An historical perspective on behavioral teratology. Neu-robehav Toxicol Teratol 7:537–540.
Butcher RE, Brunner RL, Roth T, Kimmel CA. 1972. A learning impair-ment associated with maternal hypervitaminosis-A in rats. Life Sci I11:141–145.
Butcher RE, Hoar RM, Nolan GA, et al. 1979. Interlaboratory comparisonof behavioral testing. J Assoc Anal Chem 62:840–843.
Clagett-Dame M, DeLuca HF. 2002. The role of vitamin A in mammalianreproduction and embryonic development. Annu Rev Nutr 22:347–381.
Cohlan SQ. 1953. Excessive intake of vitamin A as a cause of congenitalanomalies in the rat. Science 117:535–536.
Cohlan SQ. 1954. Congenital anomalies in the rat produced by excessiveintake of vitamin A during pregnancy. Pediatrics 13:556–567.
Cohlan SQ, Stone SM. 1961. Observations on the effect of experimentalendocrine procedures on the teratogenic action of hypervitaminosisA in the rat. Biol Neonat 3:330–342.
Coluccia A, Belfiore D, Bizzoca A, et al. 2008a. Gestational all-trans reti-noic acid treatment in the rat: neurofunctional changes and cerebellarphenotype. Neurotoxicol Teratol 30:395–403.
Coluccia A, Borracci P, Belfiore D, et al. 2008b. Effects of early gestationalall-trans retinoic acid treatment on motor skills: a longitudinal study inthe offspring of Sprague-Dawley rats. Neurotoxicology 29:1107–1113.
Coluccia A, Borracci P, Belfiore D, et al. 2009. Late embryonic exposure toall-trans retinoic acid induces a pattern of motor deficits unrelated tothe developmental stage. Neurotoxicology 30:1120–1126.
Dahl RE. 2004. Adolescent brain development: a period of vulnerabilitiesand opportunities. Keynote address. Ann N Y Acad Sci 1021:1–22.
Deng W, Aimone JB, Gage FH, et al. 2010. New neurons and new memo-ries: how does adult hippocampal neurogenesis affect learning andmemory? Nat Rev Neurosci 11:339–350.
Durston AJ, van der Wees J, Pijnappel WWM, Godsave SF. 1998. Reti-noids and related signals in early development of the vertebrate cen-tral nervous system. In: Pederson RA, Schatten GP, editors. Currenttopics in developmental biology. San Diego, CA: Academic Press.pp. 111–175.
Eskessen MB, Jensh RP, Kochhar DM, et al. 1990. Postnatal behavioralsequelae of prenatal exposure to 13-cis retinoic acid. Teratology41:621–622.
Gaston J, Hutchings DE. 1974. The effects of vitamin A excess adminis-trated during the mid-fetal period on learning and development inrat offspring. Dev Psychobiology 7:225–233.
Gibbon J, Hutchings DE, Kaufman, MA. 1973. Maternal vitamin A excessduring the early fetal period: effects on learning and development inthe offspring. Dev Psychobiology 6:445–657.
Giroud PA, Martinet M. 1955. Hypervitaminoses A et anomalies chez lefoetus de rat. Rev Intern Vitaminol 26:10–18.
Gregg NM. 1941. Congenital cataract following German measles in themother. Aust N Z J Ophthalmol 19:267–276.
Hoar RM. 1984. Reproduction/teratology. Fundam Appl Toxicol 4(3 Pt2):S335–S340.
Holson RR, Adams J, Ferguson SA, et al. 1999. Gestational stage-specificeffects of retinoic acid exposure in the rat. Neurotoxicol Teratol21:393–402.
904 ADAMS
Birth Defects Research (Part A) 88:895–905 (2010)

Holson RR, Adams J, Ferguson SA, et al. 2000. Retinoic acid exposure ongestational days 11 to 13 impairs swallowing in rat offspring. Neuro-toxicol Teratol 22:541–545.
Holson RR, Cogan JE, Adams J, et al. 2001. Gestational retinoic acid expo-sure in the rat: effects of sex, strain and exposure period. Neurotoxi-col Teratol 23:147–156.
Holson RR, Gazzara RA, Ferguson SA, et al. 1997a. Behavioral effects oflow-dose gestational day 11–13 retinoic acid exposure. NeurotoxicolTeratol 19:355–362.
Holson RR, Gazzara RA, Ferguson SA, et al. 1997b. A behavioral andneuroanatomical investigation of the lethality caused by gestationalday 11–13 retinoic acid exposure. Neurotoxicol Teratol 19:347–353.
Holson RR, Gazzara RA, Ferguson SA, et al. 1997c. Gestational retinoicacid exposure: a sensitive period for effects on neonatal mortalityand cerebellar development. Neurotoxicol Teratol 19:335–346.
Honein MA, Paulozzi LJ, Erickson JD, et al. 2001. Continued occurrenceof Accutane-exposed pregnancies. Teratology 64:142–147.
Hummler H, Korte R, Hendrickx AG, et al. 1990. Induction of malforma-tions in the cynomolgus monkey with 13-cis retinoic acid. Teratology42:263–272.
Jensh RP, Kochhar DM, Till MK, et al. 1991. Effects of prenatal exposureof isotretinoin (13-cis retinoic acid) on neonatal vocalization. Teratol-ogy 43:497.
Jones KL, Smith DW. 1973. Recognition of the fetal alcohol syndrome inearly infancy. Lancet 302:999–1001.
Kalter H. 1968. Teratology of the central nervous system. Chicago: Uni-versity of Chicago Press.
Kalter H, Warkany J. 1961. Experimental production of congenital malfor-mations in strains of inbred mice by maternal treatment with hyper-vitaminosis A. Am J Pathol 38:1–21.
Klug S, Creech Kraft J, Wildi E, et al. 1989. Influence of 13-cis and all-trans retinoic acid on rat embryonic development in vitro: correlationwith isomerisation and drug transfer to the embryo. Arch Toxicol63:185–192.
Kochhar DM, Johnson EM. 1965. Morphological and autoradiographicstudies of cleft palate induced in embryos by maternal hypervitami-nosis A. J Embryol Exp Morphol 14:223–238.
Kochhar DM, Penner JD. 1987. Developmental effects of isotretinoin and4-oxo-isotretinoin: the role of metabolism in teratogenicity. Teratology36:67–75.
Kochhar DM, Penner JD, Satre MA, et al. 1988. Derivation of retinoic acidand metabolites from a teratogenic dose of retinol (vitamin A) inmice. Toxicol Appl Pharmacol 96:429–441.
Kochhar DM, Penner JD, Tellone CI, et al. 1984. Comparative teratogenicactivities of two retinoids: effects on palate and limb development.Teratog Carcinog Mutagen 4:377–387.
Kraft JC, Kochhar DM, Scott WJ, et al. 1987. Low teratogenicity of 13-cis-retinoic acid (isotretinoin) in the mouse corresponds to low embryoconcentrations during organogenesis: comparison to the all-trans iso-mer. Toxicol Appl Pharmacol 87:474–482.
Lammer EJ, Chen DT, Hoar RM, et al. 1985. Retinoic acid embryopathy.N Engl J Med 313:837–841.
Lammer EJ, Schunior A, Hayes AM, et al. 1988. Isotretinoin dose and ter-atogenicity. Lancet 2:503–504.
Langman J, Welch GW. 1966. Effect of vitamin A on development of thecentral nervous system. J Comp Neural 128:1–16.
Lemoine P, Harousseau H, Borteyru J, et al. 1968. The children of alcoholicparents: anomalies observed in 127 cases. Quest Med 25:476–482.
Lenz W. 1961. Kindliche Missbildungen nach Medikament: Einnahmewahrend der Graviditat? Dtsch Med Wochenschr 86:2555–2556.
Maden M, Gale E, Zile M, et al. 1999. Hindbrain respecification in the ret-inoid-deficient quail. Mech Dev 89:43–54.
Makori N, Peterson PE, Blankenship TN, et al. 1998. Effects of 13-cis-reti-noic acid on hindbrain and craniofacial morphogenesis in long-tailedmacaques (Macaca fascicularis). J Med Primatol 27:210–219.
Makris SL, Raffaele K, Allen S, et al. 2009. A retrospective performanceassessment of the developmental neurotoxicity study in support ofOECD test guideline 426. Environ Health Perspect 117:17–25.
Marqueling AL, Zane LT. 2007. Depression and suicidal behavior in acnepatients treated with isotretinoin: a systematic review. Semin CutanMed Surg 26:210–220.
Marshall H, Morrison A, Studer M, et al. 1996. Retinoids and Hox genes.FASEB J 10:969–978.
Matsumoto H, Koya G, Takeuchi T, et al. 1965. Fetal Minamata disease. Aneuropathological study of two cases of intrauterine intoxication by amethyl mercury compound. J Neuropathol Exp Neurol 24:563–574.
Maynard TM, Sikich L, Lieberman JA, et al. 2001. Neural development,cell-cell signaling, and the ‘‘two-hit’’ hypothesis of schizophrenia.Schizophr Bull 27:457–476.
MGI,Bult C, Kadin J, et al. 2010. The Mouse Genome Database: enhance-ments and updates. Nucleic Acids Res 38(Database Issue):D586–D592.
Mitchell AA, Van Bennekom CM, Louik C. 1995. A pregnancy-preventionprogram in women of childbearing age receiving isotretinoin. N EnglJ Med 333(2):1.
Murakami U, Kameyama Y. 1965. Malformations of the mouse fetuscaused by hypervitaminosis-a of the mother during pregnancy. ArchEnviron Health 10:732–741.
Nau H. 2001. Teratogenicity of isotretinoin revisited: species variationand the role of all-trans-retinoic acid. J Am Acad Dermatol 45:S183–S187.
Nelson BK. 1990. Origins of behavioral teratology and distinctionsbetween research on pharmaceutical agents and environmental/industrial chemicals. Neurotox Teratol 12:301–306.
Nishimura H. 1979. Detection of environmental teratogens: some thoughtson the correlation between human and experimental teratogenesis.In: Klingberg MA, editor. Contributions to epidemiology and biosta-tistics. pp. 78–90.
Nolen GA. 1969. Variations in teratogenic response to hypervitaminosis Ain three strains of the albino rat. Food Cosmet Toxicol 7:209–214.
Nolen GA. 1972. The effects of various levels of dietary protein on reti-noic acid-induced teratogenicity in rats. Teratology 5:143–152.
Nolen GA. 1986. The effects of prenatal retinoic acid on the viability andbehavior of the offspring. Neurobehav Toxicol Ter 8:643–654.
Olson CR, Mello CV. 2010. Significance of vitamin A to brain function,behavior and learning. Mol Nutr Food Res 54:489–495.
Palha JA, Goodman AB. 2006. Thyroid hormones and retinoids: a possiblelink between genes and environment in schizophrenia. Brain Res Rev51:61–71.
Perlman SE, Leach EE, Dominguez L, et al. 2001. ‘‘Be smart, be safe, besure’’: the revised pregnancy prevention program(SM) for women onisotretinoin. J Reprod Med 46:179–185.
Robertson J, Polifka JE, Avner M, et al. 2005. A survey of pregnantwomen using isotretinoin. Birth Defects Res A Clin Mol Teratol73:881–887.
Shruster A, Melamed E, Offen D. 2010. Neurogenesis in the aged and neuro-degenerative brain. Apoptosis. In press. Mar 26. [Epub ahead of print].
Sisk CL, Foster DL. 2004. The neural basis of puberty and adolescence.Nat Neurosci 7:1040–1047.
Spear LP. 2007. Assessment of adolescent neurotoxicity: rationale andmethodological considerations. Neurotoxicol Teratol 29:1–9.
Tafti M, Ghyselinck NB. 2007. Functional implication of the vitamin Asignaling pathway in the brain. Arch Neurol 64:1706–1711.
Teratology Society. 1987. Teratology Society Position Paper: recommenda-tions for vitamin A use during pregnancy. Teratology 35:269–275.
Tyl R, Crofton K, Moretto A, et al. 2008. Identification and interpretationof developmental neurotoxicity effects: a report from the ILSIResearch Foundation/Risk Science Institute expert working group onneurodevelopmental endpoints. Neurotoxicol Teratol 30:349–381.
Ulleland CN. 1972. The offspring of alcoholic mothers. Ann N Y Acad Sci197:167–169.
van Neerven S, Kampmann E, Mey J, et al. 2008. RAR/RXR and PPAR/RXR signaling in neurological and psychiatric diseases. Prog Neuro-biol 85:433–451.
Vorhees CV. 1974. Some behavioral effects of maternal hypervitaminosisA in rats. Teratology 10:269–274.
Vorhees CV, Brunner RL, McDaniel CR, Butcher RE. 1978. The relation-ship of gestational age to vitamin A induced postnatal dysfunction.Teratology 17:271–276.
Vorhees CV. 1986a. Origins of behavioral teratology. In: Riley EP, VorheesCV, editors. Handbook of behavioral teratology. New York: PlenumPress. pp. 3–22.
Vorhees CV. 1986b. Principles of behavioral teratology. In: Riley EP,Vorhees CV, editors. Handbook of behavioral teratology. New York:Plenum Press. pp. 23–48.
Vorhees CV, Brunner RL, Butcher RE, et al. 1979. Psychotropic drugs asbehavioral teratogens. Science 205:1220–1225.
Wang HF, Liu FC. 2005. Regulation of multiple dopamine signal trans-duction molecules by retinoids in the developing striatum. Neuro-science 134:97–105.
Warkany J. 1971. Congenital malformations: notes and comments. Chi-cago: Year Book Medical.
World Health Organization (WHO). 2010. Global prevalence of vitamin adeficiency in populations at risk. 1995–2005. Geneva, Switzerland:World Health Organization.
Wilson JG. 1959. Experimental studies on congenital malformations.J Chronic Dis 10:111–130.
Zhao C, Deng W, Gage FH, et al. 2008. Mechanisms and functional impli-cations of adult neurogenesis. Cell 132:645–660.
905CNS AND BEHAVIORAL TERATOLOGY OF RETINOIDS
Birth Defects Research (Part A) 88:895–905 (2010)





























