Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 267, No. 6, Issue of February 25, pp. 3976-3962,1992 Printed in U. S. A.
A Novel 40-kDa Membrane-associated EF-Hand Calcium-binding Protein in Plasmodium falciparum*
(Received for publication, October 8,1991)
David J. RawlingsS and David C. Kaslowg From the Malaria Section, Laboratory of Parasitic Diseases, National Institutes of Allergy and Infectious Diseases, National Institutes of Health, Bethesha,.Marylund 20892
A 40-kDa sexual stage radiolabeled surface protein of Plasmodium falciparum, Pfs40, was previously identified as a potential target antigen of transmission blocking immunity by an immunogenetic approach. Synthetic oligonucleotide “guessmers,” based on micro- sequenced tryptic peptides of Pfs40 purified by two- dimensional gel electrophoresis, were used to clone the full length cDNA and genomic DNA encoding Pfs40. The deduced amino acid sequence predicted an integral membrane protein containing five EF-hand calcium- binding domains. The biological activity of one or more of these domains was confirmed by binding of 4sCa to both native and recombinant Pfs40. Antisera to recom- binant Pfs40 immunoprecipitated the native radiola- beled 40-kDa surface protein. The predicted noncyto- solic membrane-associated localization of Pfs40 is unique within the EF-hand calcium-binding protein superfamily.
Calcium ions play a critical role in a wide variety of cellular processes and cellular interactions. The ability to store cal- cium within intracellular membranous organelles is consid- ered a major development in cellular evolution of multicellular organisms (Koch, 1990). Calcium mobilization and conse- quent calcium-modulated structural changes are critical for the function of diverse groups of intracellular, cytosolic, or membrane-associated proteins (Cheung, 1980; Klee, 1988), and for at least one group of cell surface proteins (Takeichi, 1991). The role of calcium in the development of several stages of the unicellular eukaryotic parasite that causes hu- man malaria, Plasmodium falciparum, has recently been iden- tified (Tanabe, 1990); however, other than calmodulin (Rob- son and Jennings, 1991), the molecules involved in calcium- mediated signalling, intracellular calcium storage, and extra- cellular calcium-dependent events are still ill defined.
The human malaria parasite is transmitted to its mamma- lian host after undergoing sexual reproduction in the mosquito midgut. At least one of the stages of sexual development,
* This work was supported by the United Nations Developmental Program/World Bank/World Health Organization Special Pro- gramme and Training in Tropical Diseases. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequeme(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) M77834.
$ Current address: Div. of Rheumatology, Children’s Hospital of Los Angeles, 4650 Sunset Blvd., Los Angeles, CA 90054.
5 To whom correspondence should be addressed Bldg. 4, Room B1-37, National Institutes of Health, Bethesda, MD 20892. Tel.: 301- 496-3655; Fax: 301-480-3807.
exflagellation, in which male microgametocytes transform into mature motile microgametes, requires mobilization of calcium (Kawamoto et al., 1990). This and later stages in both sexes have been studied as potential targets of transmission blocking antibodies. The immunological mechanisms whereby the mammalian host suppresses transmission of sexual stage parasites to the mosquito vector has been termed transmission blocking immunity. Humoral-mediated suppression of trans- mission occurs when gametes emerge from within erythro- cytes in the mosquito midgut, and are exposed to antibodies ingested with the bloodmeal. Antibodies directed against sur- face molecules of gametes can block infectivity of these par- asites to the mosquito by interfering with fertilization or post- fertilization development (Carter et al., 1988).
Several transmission blocking immunity candidate mole- cules have been identified by the ability of monoclonal anti- bodies to block infectivity of sexual stage parasites fed to mosquitoes in vitro (Rener et al., 1983; Vermeulen et al., 1985; Quakyi et al., 1987). Because of the poor immunogenicity of a 40-kDa radiolabeled surface protein of sexual stage parasites (Good et al., 1988), Pfs40,’ this molecule has recently also been suggested to be a potential target antigen of transmission blocking immunity (Carter et al., 1988; Kaslow, 1990). In this study we report the characterization of Pfs40 and present evidence that Pfs40 is a novel EF-hand calcium-binding pro- tein present in sexual stage parasites.
EXPERIMENTAL PROCEDURES
Parasites-Gametocytes of P. falciparum were obtained by in vitro culture using a modification of a previously described method (Ifediba and Vanderberg, 1981). The parasite isolates used included 7G8 (Burkot et al., 1984) and the 3D7 clone of Amsterdam airport isolate NF54 (Walliker et al., 1987). Purification of macrogametes of P. falciparum and induction of gametogenesis was as previously de- scribed (Quakyi et al., 1987).
Immunizution-Polyclonal antisera to Pfs40 in BlO.Br, AKR, or C3H mice (Jackson Laboratories; Frederick Cancer Institute) was produced by intraperitoneal immunization every 2 weeks for up to three immunizations with 1 X lo7 P. falciparum purified newly emerged extracellular 3D7 gametes/zygotes in phosphate-buffered saline. For each mouse, the initial immunization was with complete Freund’s adjuvailt, and subsequent immunizations were with incom- plete Freund’s adjuvant. C3H sera 268 was collected 3 days after a single intrasplenic immunization of 1 x lo7 gametes in PBS given 2 weeks following the initial intraperitoneal immunization as described above.
Immunological Studies-Western blot analysis, surface radioiodi- nation, metabolic labeling, immunoprecipitation (Good et al., 19881,
‘The abbreviations used are: Pfs40, 40-kDa sexual stage radio- labeled surface protein of Plasmodium falciparum; SDS-PAGE, so- dium dodecyl sulfate-polyacrylamide gel electrophoresis; CHAPS, 3-[(3-chloroamidopropyl)dimethylammonio]-1-propanesu1fonic acid; HPLC, high performance liquid chromatography; EGTA, [ethylene- bis(oxyethylenenitrilo)]tetraacetic acid MBP, maltose-binding pro- tein; BDGE, two-dimensional gel electrophoresis.
3976

Novel EF-Hand Calcium-binding Membrane Protein i n Malaria 3977
SDS-PAGE of gametes of P. falciparum (Quakyi et al., 1987), and development of monoclonal antibody IIC5B10 (Rener et al., 1983) were as previously described. Preparation of rabbit anti-gamete polyclonal antisera 128 and 129 are described elsewhere (Quakyi et al., 1987).
2DGE Protein Purification"3D7 gametes were extracted in two- dimensional gel electrophoresis (2DGE) sample buffer containing 8 M urea, 4% CHAPS, 4% pH 4-6 Ampholines (Bio-Rad), and protease inhibitors (200 p~ L-1-tosylamido-2-phenylethyl chloromethyl ke- tone, l p ~ pepstatin, 1 p M leupeptin, 100 p M PMSF, 0.1 p M aprotinin, 0.5 mM EDTA), vortexed briefly, placed on ice for 10 min, sonicated for 30 s (maximum setting, Tekmar Sonic Disrupter), and stored at -70 "C. For separation of radioiodinated proteins, approximately 10,000 cpm of Triton X-100 extracted surface radioiodinated parasites was added to 5 volumes of the 2DGE sample buffer. 2DGE was performed as previously described with minor modifications (Hoch- strasser et al., 1988). Extracted protein from 5 X lo7 gametes/zygotes was separated in an isoelectric focusing (IEF) gel containing a mixture of 80% pH 4-6 and 20% pH 3.5-10 ampholines run in capillary tubes (Mini-Protean I1 2D-cell, Bio-Rad) for 2200 V-h. In microsequencing preparative gels, the region from pH 4.7 to 5.5 from the IEF gel was run on a 9% polyacrylamide second dimension slab gel (Bio-Rad Mini Protein 11) at 30 mA/gel for 1 h. Gels for analysis of protein migration patterns or immunoprecipitations were Coomassie stained, fixed, dried, and autoradiographed. For microsequencing, gels were soaked in transfer buffer (25 mM Tris, pH 8.0, 200 mM glycine, 10% MeOH) for 10 min and the protein electroblotted onto 0.2-pm diameter nitrocellulose (Schleicher and Schuell) in a Transblot cell (Bio-Rad) a t 100 V for 1.5 h at 4 "C. The nitrocellulose was stained for 1 min in 0.1% Ponceau S, 1% aqueous acetic acid, and destained 1-2 min in 1% aqueous acetic acid (Aebersold et al., 1987) in high pressure liquid chromatography (HPLC) grade water (Baxter). Stained Pfs40 spots were carefully cut from the filter, placed in Eppendorf tubes contain- ing HPLC water, pH 7.0, vortexed briefly, and stored moist at -20 "C.
For NH2-terminal sequencing, second dimensional slab gels were run using the Jovin 3328.IV buffer system, protein electroblotted onto polyvinylidene difluoride membrane (Immobilon-P, Millipore) in CAPS buffer, and membranes were Coomassie stained as previously described (Moos et al., 1988).
Protein Microsequencing-A total of 26 spots (0.75-mm2 surface area containing approximately 100 pmol of protein) were pooled and digested overnight at 37 "C with trypsin after blocking of membrane with polyvinylpyrrolidone. Enzymatic cleavage fragments (40 pmol by amino acid analysis) were separated by narrow-bore reverse phase HPLC as previously described (Aebersold et al., 1987). Peptide se- quence analysis, from tryptic peptides collected in fractions 50, 51, and 55 from HPLC, was performed on an automated gas-phase sequenator. Tryptic digestion, HPLC separation, and gas-phase se- quencing were performed by Dr. w . Lane (Harvard Microchem, Cambridge, MA).
NHz-terminal sequencing of electroblotted protein and in situ generated CNBr peptides was performed as previously described (Matsudaira, 1987; Urdal et al., 1984) by Dr. J. Colligan (Biological Resources Branch, National Institute of Allergy and Infectious Dis- eases).
Isolation of Pfs40 Genomic and cDNA Clones-Total cellular RNA from stage I11 to IV 3D7 P. falciparum gametocytes (RNAzol, Biotext) was used to construct an oligo-(dT) primed size selected BstXI- linkered cDNA library of approximately 1-2 X lo6 recombinants in plasmid vectors pcDNAl and pcDNA2 (Invitrogen). Two antisense oligonucleotide "guessmer" probes (4050A3:5'-TTTAAAATCTTC T/AGGATATCTT/AATT/AACATCT/ACCATAATC-3'; 4055A2: 5"GCATCA/GAAA/GAAATCATCC/TAAIGCCATTTCA TCATCTTTTTT-3') of one-sixteenth and one-eighth degeneracy, respectively, based on tryptic peptide amino acid sequences from HPLC fractions 50 and 55 using P. falciparum codon bias (Hyde et ai., 1989) and/or inosine in positions of degeneracy were synthesized using standard procedures (Applied Biosystems, Model 391 oligosyn- thesizer). 5' Termini-labeled oligonucleotides were used to screen approximately 4 X 10' colonies from each cDNA library and from an amplified mung bean nuclease genomic DNA library of P. falciparum clone 7G8 in pUC13 (Kaslow and Hill, 1990). Filters were hybridized at 55 "C overnight in 6 X SSC, 200 pg ml" of heparin sulfate, 5 X Denhardt's, 0.5% SDS, and 20 mM sodium phosphate, pH 6.8, and washed three times at room temperature in 6 X SSC (1 M sodium chloride, 0.1 M sodium citrate, pH 7.4), 0.5% SDS with a final 4-min wash at 55 "C in the same solution. Six pcDNAl, nine pcDNA2, and five genomic clones hybridizing to both oligonucleotide probes were
isolated. 100% of both strands of a 1.9-kilobase cDNA clone (pc2- 11.1) and a 1.8-kilobase genomic clone (MB-8.1) were sequenced by the dideoxynucleotide terminator method (Chen and Seeburg, 1985). Universal forward and reverse sequencing primers (Sequenase, U. S. Biochemical) and the oligonucleotide guessmer probes described above were used to obtain initial sequence from genomic clone MB- 8.1. Subsequent sequence of all clones was obtained using synthetic oligonucleotides. Sequence analysis was carried out using program software from PC/Gene (Intelligenetics, Mt. View, CAI. The Swiss- prot and PIR protein data bases were searched for homology with the Pfs40 sequence (PC/Gene; NIH Convex system, Blastp).
Northern and Southern Blot and Pulsed Field Gel Analysis-Total cellular parasite RNA was electrophoresed through a 1% agarose/ formaldehyde gel and transferred to nylon membrane (Genescreen Plus, Du Pont-New England Nuclear) as described with minor mod- ifications (Sambrook et al., 1982). Pulse field gel electrophoresis was performed as described (Wellems et al., 1987). Single and multiple restriction enzyme digests of parasite genomic DNA were fractionated in a 1% Tris/acetate/EDTA-agarose gel and transferred to nylon membrane in NH4Ac buffer (Rigaud et al., 1987) after acid depuri- nation. Probes were radiolabeled using the random hexamer primer method (Feinberg and Vogelstein, 1984) and used for Southern and Northern blot analysis. Filters were hybridized in 1% SDS, 1 M NaCl, 10 mM Tris, pH 7.4, with heparin sulfate (Singh and Jones, 1984) at 55 "C overnight and then washed three times at room temperature for 5 min in 2 X SSC, 0.5% SDS with one or two final washes for 30 min at 55 "C in the same solution. Additional 30-min washes with 0.2 X SSC, 0.5% SDS were performed for higher stringency analysis.
Expression of Recombinant Protein and Immunizations-The cod- ing region of Pfs40 (amino acids 27-374) without the putative signal sequence was amplified from MB-8.1 (with 40Nterm-SmaI: GTCCC GGGGATAACATGAAATACAAT and M13 universal forward primers) using the polymerase chain reaction (Saiki et al., 1988); cloned into the StuI and EcoRI restriction sites of the maltose-binding protein (MBP) prokaryotic expression vector pIH902 (gift of P. Riggs, New England Biolabs) and the ligation transformed into E. coli, BRL 3133 (protein degradation minus, K12 strain; gift of J . Jessee, BRL). A single recombinant colony pIH902-40.1, confirmed by nucleotide sequencing, was grown to mid-log phase in enriched L broth (Super- broth, Quality Biological), induced with 0.3 mM isopropyl-1-thio$- D-galactopyranoside, and harvested after 3-6 h growth at 37 "C. Pelleted cells from 100 ml of culture were resuspended in 10 ml of lysis buffer (10 mM Tris, pH 7.5, 0.5 M NaC1, 0.25% Tween, 10 mM EDTA, 10 mM EGTA, 10 mM 2-mercaptoethanol) containingprotease inhibitors (2 pg/ml of pepstatin, 1 pg/ml of leupeptin, and 100 p M phenylmethylsulfonyl fluoride), freeze thawed three times in liquid nitrogen, sonicated, and pelleted. The pellet was diluted 10-fold in column buffer (10 mM Tris, pH 8.0, 0.5 M NaCl) and incubated overnight at 4 "C with 10 ml of amylose resin (New England Biolabs). Purified fusion protein was eluted with 10 mM maltose in column buffer with an average yield of 20 mg liter" by Bradford assay. Alternatively, bound fusion protein as cleaved in situ on the amylose column with Factor Xa (1 unit/20 pg of fusion protein) in FXa buffer (10 mM Tris, pH 8.0, 100 mM NaC1, 2 mM CaC12) for 24-48 h at 4 "C and collected after centrifugation at 2000 rpm in the same buffer.
Anti-recombinant sera was produced from CAFl (Jackson Labs) and C3H mice (Frederick Cancer Institute) immunized intraperito- neally with 50-100 pg of MBP-Pfs40 fusion protein or purified FXa cleaved fusion protein in monophosphyl lipid A and trehalose dimy- colate adjuvant (Ribi Immunochem) at days 0, 14,35, and 56.
DGFISLNE) and CBD3 (C)DKDKDGKLSINE) were synthesized Peptide Synthesis and Immunization-Peptides CBD2 (C)IDSDK
using 9-fluorenylmethyloxycarbonyl chemistry (J. Colligan, BRB, National Institute of Allergy and Infectious Disease). Peptides were conjugated through NH2-terminal sulfhydryl linkage to cationized- bovine serum albumin and purified as recommended (Pierce Chemical Co., Imject Activated Supercarrier). Rabbit antipeptide sera was produced by subcutaneous immunization of New Zealand White rabbits using 50-100 pg of conjugate in monophosphyl lipid A, tre- halose dimycolate, and cell wall skeleton adjuvant (Ribi Immuno- chem).
Calcium Binding Studies-Native or recombinant proteins sepa- rated by 2DGE or SDS-PAGE were electroblotted to nitrocellulose. Detection of calcium binding by 45Ca autoradiography was performed as described (Maruyama et al., 1984). Briefly, membranes were washed in calcium-binding buffer (10 mM imidazole-HC1, pH 6.8, 60 mM KCl, 5 mM MgCl), incubated in 1 pCi ml" of 45Ca in the same buffer, washed with distilled water, dried, and autoradiographed.
3978 Novel EF-Hand Calcium-binding Membrane Protein in Malaria
RESULTS
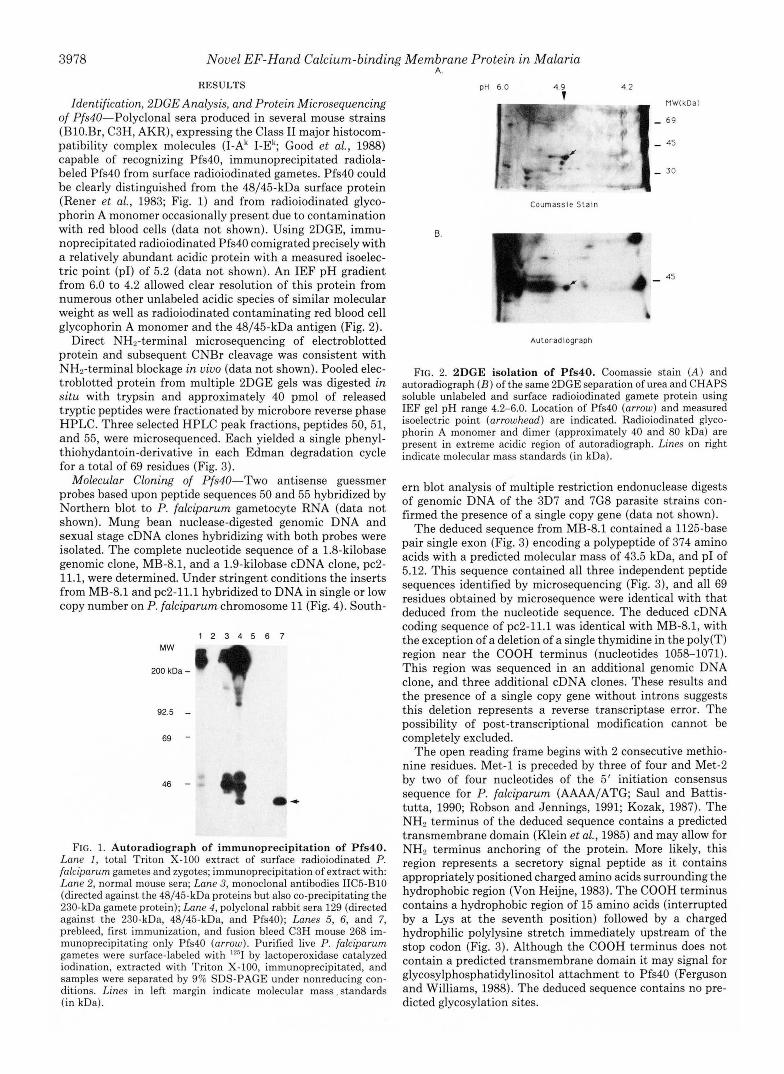
Identification, BDGE Analysis, and Protein Microsequencing of Pfs40"Polyclonal sera produced in several mouse strains (BlO.Br, C3H, AKR), expressing the Class I1 major histocom- patibility complex molecules (I-Ak I-Ek; Good et al., 1988) capable of recognizing Pfs40, immunoprecipitated radiola- beled Pfs40 from surface radioiodinated gametes. Pfs40 could be clearly distinguished from the 48/45-kDa surface protein (Rener et al., 1983; Fig. 1) and from radioiodinated glyco- phorin A monomer occasionally present due to contamination with red blood cells (data not shown). Using 2DGE, immu- noprecipitated radioiodinated Pfs40 comigrated precisely with a relatively abundant acidic protein with a measured isoelec- tric point (PI) of 5.2 (data not shown). An IEF pH gradient from 6.0 to 4.2 allowed clear resolution of this protein from numerous other unlabeled acidic species of similar molecular weight as well as radioiodinated contaminating red blood cell glycophorin A monomer and the 48/45-kDa antigen (Fig. 2).
Direct NH2-terminal microsequencing of electroblotted protein and subsequent CNBr cleavage was consistent with NH2-terminal blockage in uiuo (data not shown). Pooled elec- troblotted protein from multiple 2DGE gels was digested in situ with trypsin and approximately 40 pmol of released tryptic peptides were fractionated by microbore reverse phase HPLC. Three selected HPLC peak fractions, peptides 50,51, and 55, were microsequenced. Each yielded a single phenyl- thiohydantoin-derivative in each Edman degradation cycle for a total of 69 residues (Fig. 3).
Molecular Cloning of Pfs40"Two antisense guessmer probes based upon peptide sequences 50 and 55 hybridized by Northern blot to P. falciparum gametocyte RNA (data not shown). Mung bean nuclease-digested genomic DNA and sexual stage cDNA clones hybridizing with both probes were isolated. The complete nucleotide sequence of a 1.8-kilobase genomic clone, MB-8.1, and a 1.9-kilobase cDNA clone, pc2- 11.1, were determined. Under stringent conditions the inserts from MB-8.1 and pc2-11.1 hybridized to DNA in single or low copy number on P. falciparum chromosome 11 (Fig. 4). South-
1 2 3 4 5 6 7
200 MW kDa - 8 9 i b ..
92.5 -
69 -
46 - "
FIG. 1. Autoradiograph of immunoprecipitation of Pfs40. Lane I , total Triton X-100 extract of surface radioiodinated P. falciparum gametes and zygotes; immunoprecipitation of extract with: Lane 2, normal mouse sera; Lane 3, monoclonal antibodies IIC5-Bl0 (directed against the 48/45-kDa proteins but also co-precipitating the 230-kDa gamete protein); Lane 4, polyclonal rabbit sera 129 (directed against the 230-kDa, 48/45-kDa, and Pfs40); Lanes 5, 6, and 7, prebleed, first immunization, and fusion bleed C3H mouse 268 im- munoprecipitating only Pfs40 (arrow). Purified live P. falciparum gametes were surface-labeled with by lactoperoxidase catalyzed iodination, extracted with Triton X-100, immunoprecipitated, and samples were separated by 9% SDS-PAGE under nonreducing con- ditions. Lines in left margin indicate molecular mass .standards (in kDa).
A.
pH 6.0 4.9 4 2
Coumassle Staln
I - 45
b
Autoradlograph
FIG. 2. BDGE isolation of Pfs40. Coomassie stain ( A ) and autoradiograph ( B ) of the same 2DGE separation of urea and CHAPS soluble unlabeled and surface radioiodinated gamete protein using IEF gel pH range 4.2-6.0. Location of Pfs40 (arrow) and measured isoelectric point (arrowhead) are indicated. Radioiodinated glyco- phorin A monomer and dimer (approximately 40 and 80 kDa) are present in extreme acidic region of autoradiograph. Lines on right indicate molecular mass standards (in kDa).
ern blot analysis of multiple restriction endonuclease digests of genomic DNA of the 3D7 and 7G8 parasite strains con- firmed the presence of a single copy gene (data not shown).
The deduced sequence from MB-8.1 contained a 1125-base pair single exon (Fig. 3) encoding a polypeptide of 374 amino acids with a predicted molecular mass of 43.5 kDa, and PI of 5.12. This sequence contained all three independent peptide sequences identified by microsequencing (Fig. 3), and all 69 residues obtained by microsequence were identical with that deduced from the nucleotide sequence. The deduced cDNA coding sequence of pc2-11.1 was identical with MB-8.1, with the exception of a deletion of a single thymidine in the poly(T) region near the COOH terminus (nucleotides 1058-1071). This region was sequenced in an additional genomic DNA clone, and three additional cDNA clones. These results and the presence of a single copy gene without introns suggests this deletion represents a reverse transcriptase error. The possibility of post-transcriptional modification cannot be completely excluded.
The open reading frame begins with 2 consecutive methio- nine residues. Met-1 is preceded by three of four and Met-2 by two of four nucleotides of the 5' initiation consensus sequence for P. falciparum (AAAA/ATG; Saul and Battis- tutta, 1990; Robson and Jennings, 1991; Kozak, 1987). The NH, terminus of the deduced sequence contains a predicted transmembrane domain (Klein et al., 1985) and may allow for NH, terminus anchoring of the protein. More likely, this region represents a secretory signal peptide as it contains appropriately positioned charged amino acids surrounding the hydrophobic region (Von Heijne, 1983). The COOH terminus contains a hydrophobic region of 15 amino acids (interrupted by a Lys at the seventh position) followed by a charged hydrophilic polylysine stretch immediately upstream of the stop codon (Fig. 3). Although the COOH terminus does not contain a predicted transmembrane domain it may signal for glycosylphosphatidylinositol attachment to Pfs40 (Ferguson and Williams, 1988). The deduced sequence contains no pre- dicted glycosylation sites.

Novel EF-Hand Calcium-binding Membrane Protein in Malaria 3979
l I I t 1 1 I I 1 1 1 l f I 1 g 1 1 ~ l l ~ ~ 1 1 1 ~ l l 1 ~ 1 l 1 1 ~ ~ 1 1 1 1 ~ ~ I 1 1 1 1 t 1 t l 1 l 1 1 1 l ~ 5 ~ ~ ~ ~ . A A A T A A A T T T G T A C A A G T T G T T A T G C T T C A T A T G T G T A A T A T T T T T G T T A 5 7 W K .. !...N _ _ _ _ I...! _ _ _ _ n."L _ _ _ _ 5....c _... F .". 1 .". c....v .... !...F .." 5....L..
c C A T A A A A A T G T G G T A A G A T C A G G T G A T A A C A T G A A A T A C A A T G A T A T G A A A G G A T T A G A T G A T C T T A G T A A G T T A A A T G A T G A T C A G G T G A A G G A T A T T T T A G G A T T A A A A A T A G A T G G A 1 7 7
4 BclI 2 0 . . M . . K N V V R S G D N M K Y N D M K G L D D L S K L N D D O V K D l L G L K I D G
6 O A K E R I E U L F H L I D K N N D K E I T E E E L N T W S S F L G C A A A A G A A ~ G A A T A G A A A A A T T A T T T C A T T T G A T A G A T A A A A A C A A T G A T A A A G A A A T A A C A G A A G A A G A A T T A A A T A C T T G G T ~ T A G C T T T T T A ~ A A A A T G A A A T A T T T T T A A A ~ A K N E I F L K d 2 9 7
Domun 1
G T A C A A G C C G A A A T G G G A C A A A T A G A T T C T G A C A A G G A T G G G T T C A T A T C T T T A A A C G A A T T A A A T G A T G C A T T T G C A C A A A A T T T A G A ~ G C T A A A G A A G T T G A A A A A C A T T C A G A GGA 4 1 7
Domain 2
M"l I
1 0 0 V O A E M G O I D S D K D G F I S L N E L N D A F A O N L D A K E V E K H S E ~
T T A T T A A A A A G A T T T C A A A T T G T T G A T A A A G A T A A A G A T G G G A A A T T A A G T A T T A A T G A A G T T G G A T T A T T G A T T G A T C C T A T G A A G A T G A A G A A T T A A A A G A A T T A G A A A T A A A T G A A 5 3 7
Domu" 3 I I O L L C R F O I V D K D K D G K L S I N E V G L L I D P M U D E E L K E L ~
A T T C T T G A A C A C C A T G A T G T A A A T A A A G A T G G A A A A A T A T C A T T A G A T G A A T T T A A A C A A A C A A G A T C T G A T G A A G T T A G G T G T T A A A A A A G A T G A T G A A A T G G C T C T T G A T G A T T T T 6 5 7 1 LIII
I B O I L E H ~ ~ V N K D G K X S L D E F K O T R S D E Dom.in 4 '' ' ' & l 1 d ? 5 5 ~ E' ' ' I. ' '
A A T T T ~ T T T G A T G C T A A T A A A G A T G G A T T T A T A G A T A A A G A A G A A A T T A T T A A A G T T T A T T T T G A T C C T C T C A T G A A A G T G G A G C A A T C A A T G T A A A C G A A A T A A A G G A A A A T A T ~ T T T 7 7 7 2 2 0 N F F D I N K O G F T D K E E I I K V Y F D P A H E S G A I N V N E I K E N I F
D o m m 5 Psptna. SI
G A A G G T A A A A A A A T T A C T T A T G A T T T A T G G A A T G A A A A A G C A C T T A A A A T T G C T G T A A C A T C A T T A A C T G A T T A T G G T G A T G T T A T A A G A T A T C C A G A A G A T T T T A A A C T A G A T A T A G G A 1 9 7 1 Emw
2 6 0 E G N K I T Y D L W N E K A L ~ 1 I V T T L 1 D D ~ D 1 , d ~ 5 0 V I R I P E D F K L D I G
AAAAATGTTATCTTACCAACAGCTCGTAGTAGAGCATTCGAAGATGATGATATGGATGCTGATACACAGAAGATGATAAAGATGAAGCTGACGATGCATCCCAACAAAAATCCCCAGCAA l o t 7 3 0 0 K N V I L P T A R S R A F E D D D M D A D T O K M l K M K L T M ~ P N K N P 0 0
T T G A T G A A T T A T A G G G T A T A T A T A T A T T T A l A T A A A C A T A T T T T T T T T T T T T T T A A C A T A A A G G G A A A C A A A A A A A A A A A A A A A ~ A A T T T C G T A C A C C C C A C A A A T ~ A l 8 l l * l ~ ~ ~ C ~ 1 $ 1 3 7 3 4 0 L M N Y R ..V _ _ _ _ Y .___ I _ _ _ _ Y _ _ _ _ L... X. K .. U...l...E...F _ _ _ _ E...F....N...l.. K G N K K K K K I( F R T P H K
1 g . . 1 . 1 . 1 1 l . 1 t 1 1 1 1 1 g 1 . 1 . 1 1 . l l 1 1 1 . 1 g . g S S S . . . l . 1 . 1 . 1 . l , g , ~ l * l ,
FIG. 3. Nucleotide sequence and deduced amino acid sequence of genomic DNA clone MB-8.1. The 1125 base pairs predicted coding region of the nucleotide sequence and underlying deduced amino acid sequence of Pfs40 are indicated (in upper case). Inverse print highlights two possible start codons; asterisk (*), indicates stop codon; dashed lines, hydrophobic regions; right arrow, start of recombinant portion of MBP-rPfs40 fusion protein; down arrows, restriction sites; boxes, putative calcium-binding domains; and shaded amino acids, tryptic peptide sequences determined by microsequencing. Amino acids are numbered on left and nucleotides on right.
A. 8.
FIG. 4. Pulsed field gel electrophoresis blot analysis. DNA from 3D7 strain of P. falciparurn was fractionated by high resolution pulsed field gel electrophoresis to separate chromosomes 10-14 and stained with ethidium bromide ( A ) . A Southern blot of the gel was hybridized overnight with random primed MB 8.1 insert and the autoradiograph after a 24-h exposure a t -70 "C is shown ( B ) . Line identifies location of chromosome 11. No hybridization was present with smaller chromosomes (data not shown).
Pfs40 is a Calcium-binding Protein-A striking feature of the deduced amino acid sequence are five predicted EF-hand calcium-binding domains based upon homology with the cal- cium-binding domains of members of the EF-hand calcium- modulated protein superfamily (Figs. 3 and 5A; Tufty and Kretsinger, 1975; Bairoch and Cox, 1990). The structure of the EF-hand domain, determined crystallographically, con- sists of a 12-residue loop flanked by two perpendicular a- helices. The oxygen containing residues which coordinate calcium-binding, denoted as X, Y, Z, -X, and -Z, are present in the loop and a 29-residue consensus sequence containing 16 conserved positions has been used to predict this helix- loop-helix structure (Fig. 5A; Tufty and Kretsinger, 1975). Domains 1 through 5 (Dl-5) of Pfs40 have identity with this consensus sequence ranging between 14/16 residues (D2) and 11/16 residues (D5) (Fig. 5A) . With only a single exception (replacement of the Gly with Lys in position 15 of Dl), the consensus residues within the predicted calcium binding loops of all five Pfs40 domains are identical.
The structure of EF-hand calcium-binding domains has also been evaluated using protein hydropathy profiles where the helix-loop-helix corresponds to a hydropathy pattern of hydrophobicity (peak)-hydrophilicity (valley)-hydrophobicity
(peak) (Kobayashi et al., 1984). The calcium-binding domains of Pfs40 also follow this pattern (Fig. 5B). D l and D3 have the most marked conformity.
Identity of the calcium-binding domains was present in representatives of nearly all members of the EF-hand protein superfamily; however, the identity was limited to the calcium- binding loop regions of the EF-hand domains, and not else- where. There was no overall homology with these or any other protein in the data base. Notably, there was no homology, outside the calcium-binding loops, with the calmodulin gene of P. falciparum (Robson and Jennings, 1991).
To determine if native Pfs40 has calcium-binding activity, 45Ca autoradiography of total soluble gamete/zygote protein separated using 2DGE was performed (Fig. 6). Pfs40 was the only calcium-binding protein present in the pH range of 6.0 to 4.2. SDS-PAGE separation and 45Ca autoradiography of total gamete protein also revealed a single predominant cal- cium-binding protein of approximately 40 kDa (data not shown). 45Ca autoradiography also clearly indicated that re- combinant Pfs40 (rPfs40) binds calcium when expressed as a MBP fusion protein (Fig. 7A) and after cleavage of the fusion protein with Factor Xa (data not shown).
Identity of the Cloned Gene-Antisera raised against rPfs40 was used for immunoprecipitation of Triton X-100 extract of surface radioiodinated P. falciparum gametes. After four im- munizations, sera raised in CAFl (F1 progeny of BalbC x CJ), C3H, and AKR mice (expressing the previously docu- mented responsive Class I1 alleles) immunoprecipitated native Pfs40 (Fig. 8). Antisera to peptides based on the calcium- binding loops of D2 and D3 (peptides CBD2 and CBD3) recognized rPfs40, after cleavage of the fusion protein with Factor Xa, by immunoblot (Fig. 7B). Peptide-CBD2 antisera immunoprecipitated native Pfs40 and recognized a 40-kDa protein by immunoblot of SDS-PAGE separated gamete pro- tein (data not shown).
DISCUSSION
Pfs40, a 40-kDa radioiodinable surface protein in P. falci- parum gametes and zygotes was first identified as a potential target of transmission blocking immunity by an immunoge- netic approach. Carter et al. (1988) interpreted the limited

3980 Novel EF-Hand Calcium-binding Membrane Protein in Malaria
FIG. 5. Calcium-binding domains of Pfs40. A , EF-hand consensus indi- cating location of calcium-binding loop, (r-helices, and conserved residues (underlined); n, nonpolar residues (L, I, V, F, or M); asterisk (*), any residue; X, Y, Z, -X, -Z, calcium chelating residues (D, E, N, S, or T); I , hydrophobic residue (I , L or V). 0 1 - 0 5 , putative calcium- binding domains of Pfs40 with conserved residues underlined. E , hydrophobicity plot (Kyte and Doolittle, 1982; calcu- lated using a span of 9 amino acids) and overlying diagram of Pfs40 coding se- quence indicating calcium-binding do- mains (dark blocks), hydrophobic 5’ and 3’ regions (gray blocks), polylysine re- gion (striped block).
A. Uasic
Ponceau-S stain
U.
A
CONSENSUS Dl D2 D3 04 D5
d e l l x 1303
H e l i x
~ n * * n n * * n X * X * Z G * L - X * * - Z n * * n n ~ * n R L E K L E H L L Q K N N P K E I T E E E L N T W S S F L Q Y Q R E M G Q L Q S Q K P G F I S L N E L N D A E A Q N G L L K R E Q I Y P K Q K Q G K L S I N E Y G L L I D P M E L N E I L E H H P V N K P G K I S L D E E K Q T R S D E M A L D D E N F E P A N K Q G F I R K E E I I K Y Y F D P
3 .
Dl D2 D3 5 ’
D4 D5
# I I I I ” 3’
- 4 9 -I I -5. . , , , , . . , . . , . . . , , , . . , , , . . , . , . , , , . . . . . . . . , . . . . 1 . . . I
1 7.’ 149’ 219’ 20e’ 3s.’
Acidic
4 ba binding FIG. 6. “Ca binding to 2DGE-isolated Pfs40. A , urea and
CHAPS solubilized protein from 2.5 X 10’ gametes separated by 2DGE was transferred to nitrocellulose membrane, washed in cal- cium-binding buffer, incubated in same buffer containing l WCi ml” of “Ca, washed in distilled water, and autoradiographed overnight; H , the same filter stained with Ponceau S. Location of Pfs40 (arrows) and molecular weight standards (lines) are indicated. Note that the molecular weight standards do not bind “Ta.
immune response to this protein among congenic, major his- tocompatability complex disparate, mouse strains (Good et al., 1988) as evidence that the molecule had a paucity of T cell epitopes as a consequence of immune selection. Recogni- tion of this molecule by only a limited number of hyperim- mune adults from an endemic region of Ghana is consistent with this interpretation.’ This limited immune response is similar to the limited immunogenicity of the two known transmission blocking immunity target surface antigens pres- ent on circulating gametocytes in infected humans (Good et al., 1988). To characterize the biological role(s) of Pfs40 and
I. A. Quakyi, personal communication.
A. 8.
1 2 a b a b
KW a b c d
200 - KW - 46 -
30 - 1 - 4-
FIG. 7. 4sCa binding and Western blot analysis of rPfs4O. A: Lane I , Coomassie stain of 10% SDS-PAGE separation of amylose resin purified: a, MBP control (monomer and dimer) and b, MBP- rPfs40 fusion protein; and Lane 2, ‘“Ca binding of identical gel after transfer to nitrocellulose membrane. Arrow, indicates position of rPfs40 fusion protein. R, Western blot analysis of Factor Xa cleaved purified MBP-rPfs40 fusion protein after separation by 9% SDS- PAGE and transfer to nitrocellulose using: a, anti-rPfs40 mouse sera; b, normal mouse sera; c, anti-peptide-CBDZ sera; and d, anti-peptide- CBD3 sera. Position of MBP portion (line) and rPfs40 portion (arrow) of FXa cleaved fusion protein are indicated.
the immunological response directed against the molecule, we have purified and microsequenced Pfs40, and cloned the gene encoding this protein.
Several lines of evidence indicate that we have cloned the gene encoding this 2DGE-isolated protein. First, and most importantly, the deduced amino acid sequence contains all three microsequenced tryptic peptides. Second, the sequence predicts a nonglycosylated polypeptide of 43-kDa with a PI of 5.12. Third, the 2DGE-isolatedprotein binds 45Ca as predicted by the presence of calcium-binding domains in the deduced amino acid sequence. Finally, sera raised against the recom- binant protein reacts with the 2DGE-isolated protein. Two lines of evidence indicate that the 2DGE-isolated and micro- sequenced protein is the radiolabeled 40-kDa surface protein immunoprecipitated by mice expressing I-Ak, I-Ek, Class I1 myosin heavy chain molecules (Good et al., 1988). First, the deduced amino acid sequence predicts an NHP-terminal secre- tory signal sequence, and a COOH-terminal hydrophobic do- main which may signal transfer to a glycosylphosphatidyl- inositol anchor. These data suggest that the predicted polypeptide is membrane associated. Second and most signif-

Novel EF-Hand Calcium-binding Membrane Protein in Malaria 3981 1 8 2 3 4 5 6
MW
200-
92 -
69 -
46 - 4
FIG. 8. Immunoprecipitation of native Pfs40 using anti- rPfs4O sera. Autoradiograph of total Triton X-100 extract of surface radioiodinated gametes and zygotes (lane 1 ) and immunoprecipita- tions of extract using: lane 2, monoclonal antibody IIC5-Bl0; lune 3, C3H mouse 268; lune 4, prebleed CAFl mouse 49; lane 5, CAFl 49 anti-rPfs40 fusion protein immune sera; and lane 6, CAFl 44 anti- MBP control immune sera. Position of Pfs40 (arrow); and molecular weight standards (in kDa) are indicated. Lune B is blank. Samples were separated by 10% SDS-PAGE under nonreducing conditions. For further details of antibody reagents refer to Fig. 1 and “Materials and Methods.”
icantly, anti-peptide and anti-recombinant sera immunopre- cipitate a 40-kDa radioiodinated protein from surface labeled Triton X-100 extracted P. falciparum gametes.
The most remarkable feature of the deduced amino acid sequence of Pfs40 is the five calcium-binding domains pre- dicted by homology with domains of the EF-hand calcium- modulated protein superfamily. Members of this superfamily, which include the ubiquitous calcium-binding protein, cal- modulin, are cytosolic proteins, consisting of groups of hom- ologs with two to eight copies of EF-hand calcium-binding domains (Moncrief et al., 1990). The calcium-mediated struc- tural changes in the EF-hand superfamily proteins are well described (Cheung, 1980). When these proteins bind calcium present in the cytoplasm of the stimulated cell, they are activated to function ultimately as enzymes, regulate other enzymes, or interact with structural or contractile proteins (Moncrief et al., 1990). In Pfs40 the calcium-binding loops within these domains are all highly conserved but there is no additional homology with these calcium-modulated proteins. ‘ T a autoradiography of native and recombinant protein con- firmed that Pfs40 is a calcium-binding protein and using that assay, Pfs40 was identified as the major calcium-binding protein in P. falciparum gametes.
The role Pfs40 plays in calcium-mediated events may relate t o critical cellular interactions between sexual stage parasites within the mosquito midgut. The homophilic binding inter- actions of the cadherin family is one well described example of calcium-dependent cellular adhesion orchestrated by cell surface calcium-binding proteins (Takeichi, 1991). Even a single amino acid substitution in a non EF-hand calcium- binding domain of uvomorulin (a member of the cadherin family) abolishes its adhesive function (Ozawa et al., 1990). Similarly, calcium modulated structural changes in Pfs40 may be responsible for molecular interactions necessary for cellular recognition or aggregation.
Whatever its role, Pfs40 is unique among EF-hand calcium- binding proteins, as it has a presumptive secretory signal sequence and, potentially, a glycosylphosphatidylinositol an- chor. These features along with its ability to be surface radiolabeled with ‘*‘I strongly suggest that Pfs40 is a noncy- tosolic membrane-associated polypeptide. While antibodies
elicited by bacterially expressed rPfs40 immunoprecipitate native protein, this sera primarily recognizes denahred pro- tein (data not shown). Expression in recombinant eukaryotic systems, currently in progress, may allow proper folding of the protein and production of high titer antibodies to native Pfs40. The definitive subcellular localization of Pfs40, in addition to evaluation of its predicted role as a target of transmission blocking immunity, must await production of antisera that primarily recognizes nondenatured parasite- produced protein.
The only other reported exception to the cytosolic localiza- tion of EF-hand containing calcium-binding proteins is in another parasite, Toxoplasma gondii, also of the subclass Coccidia. A 24-kDa calcium-binding protein (P24) with two EF-hand domains is produced by T. gondii and localizes to the dense granules of the invasive tachyzoite (Cesbron-De- lauw et al., 1989). P24 is present within the parasite vacuolar space after invasion of the host cell and evidence suggests it is secreted during invasion. P24 has no homology with Pfs40 with the exception of the calcium-binding loops. In Coccidia, novel subfamilies of membrane-targeted EF-hand calcium- binding proteins may have evolved to modulate subcellular or extracellular calcium-dependent events. Study of Pfs40, P24, and perhaps other EF-hand calcium-binding molecules pres- ent in other Coccidia parasites may enlarge the current view of the biological roles of the EF-hand domain superfamily.
Acknowledgments-We thank Louis H. Miller for encoura, uement and support, David Keister for cult,uring parasites, Lynn Lambert and Carol Rugh for assistance with immunizations, and Kim William- son, Mohammed Shahabuddin, Gary Ward, and Michael Mowatt for critical discussions and reading of the manuscript.
REFERENCES Aebersold, R. H., Leavitt, J., Saavedra, R. A., Hood, L. E., and Kent,
Bairoch, A., and Cox, J. A. (1990) FEBS. Lett. 269,454-456 Burkot, T. R., Williams, J. L., and Schneider, I. (1984) Trans. R. SOC.
S. B. H. (1987) Proc. Natl. Acad. Sci. U. S. A. 8 4 , 6970-6974
Trop. Med. Hyg. 7 8 , 339-341 Carter. R.. Kumar. N.. Quakvi. I.. Good. M.. Mendis. K.. Graves, P.,
and Miller, L. (1988) Prog:Allergy 4 1 , 193-214 Cesbron-Delauw, M. F., Guy, B., Torpier, G., Pierce, R. J., Lenzen,
G., Cesbron, J. Y., Charif, H., Lepage, P., Darcy, F., Lecocq, J. P., and Capron, A. (1989) Proc. Natl. Acad. Sci. U. S. A. 8 6 , 7537- 7541
Chen, E. Y., and Seeburg, P. H. (1985) DNA (N.Y.) 4,165-170 Cheung, W. Y. (1980) Science 207,19-27 Feinberg, A. P., and Vogelstein, B. (1984) Anal. Biochem. 137, 266-
Fermson. M. A. J.. and Williams, A. F. (1988) Annu. Reu. Biochem. 267
57,285-320 Good. M. F.. Miller. L. H.. Kumar. S.. Quakvi. I. A.. Keister. D..
Adams, J. ’H., Moss, B., ’Berzofsky, J. A., and Carter, R. (1988) Science 242,574-577
Hochstrasser, D. F., Harrington, M. G., Hochstrasser, AX., Miller, M. J., and Merril, C. R. (1988) And. Riochem. 173, 424-435
Hyde, J. E., Kelly, S. L., Holloway, S. P., Snewin, V. A., and Sims, P. F. G. (1989) Mol. Biochem. Parasitol. 32, 247-262
Ifediba, T., and Vanderberg, J. P. (1981) Nature 294,364 Kaslow, D. C. (1990) Immunol. Lett. 25,83-86 Kaslow, D. C., and Hill, S. (1990) J. Biol. Chem. 265 , 12337-12341 Kawamoto, F., Alejo-Blanco, R., Fleck, S. L., Kawamoto, Y., and
Klee, C. B. (1988) Biochemistry 27,6645-6650 Klein, P., Kanehisa, M., and DeLisi, C. (1985) Biochem. Biophys.
Kobayashi, T., Takasaki, Y., Takagi, T., and Konishi, K. (1984) Eur.
Koch, G. L. (1990) BioEssays 12 , 527-531 Kozak, M. (1987) J. Mol. Biol. 196 , 947-950 Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132 Marusama, K., Mikawa, T., and Ebashi, S. (1984) J. Biochem. (Tokyo)
Sinden, R. E. (1990) Mol. Biochem. Purmitol. 42 , 101-108
Acta 815,468-476
J. Biochem. 144,401-408
95,-511-519 Matsudaira, P. (1987) J. Biol. Chem. 262,10035-10038

3982 Novel EF-Hand Calcium-binding Membrane Protein in Malaria
Moncrief, N. D., Kretsinger, R. H., and Goodman, M. (1990) J. Mol. Saul, A., and Battistutta, D. (1990) Mol. Biochern. Parasitol. 42 ,
Moos, M., Jr., Nguyen, N. Y., and Liu, T-Y. (1988) J. Biol. Chem. Singh, L., and Jones, K. W. (1984) Nucleic Acids Res. 12,5627-5637
Ozawa, M., Engel, J., and Kemler, R. (1990) Cell 6 3 , 1033-1038 Tanabe, K. (1990) Blood Cells 16,437-449 Quakyi, I. A., Carter, R., Rener, J., Kumar, N., Good, M. F., and Tufty, R. M., and Kretsinger, R. H. (1975) Science 187 , 167-169
Miller, L. H. (1987) J. Zmmunol. 139,4213-4217 Urdal, D. L., Mochizuki, D., Conlon, P. J., March, C. J., Remerowski, Rener, J., Graves, P. M., Carter, R., Williams, J. L., and Burkot, T. M. L., Eisenman, J., Ramthun, C., and Gillis, S. (1984) J. Chro-
R. (1983) J. Exp. Med. 158,976-981 matogr. 296 , 171-179 Rigaud, G., Grange, T., and Pictet, R. (1987) Nucleic Acids Res. 15 , Vermeulen, A. N., Ponnudurai, T., Beckers, P. J. A., Verhave, J-P.,
85 7 Smits, M. A., and Meuwissen, J. H. E. (1985) J . Exp. Med. 1 6 2 , Robson, K. J . H., and Jennings, M. J. (1991) Mol. Biochern. Parasitol. 1460-1476
Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Walliker, D., Quakyi, I. A., Wellems, T. E., McCutchan, T. F., Horn, G. T., Mullis, K. B., and Erlich, H. A. (1988) Science 239, Szarfman, A., London, W. T., Corcoran, L. M., Burkot, T. R., and
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1982) Molecular Wellems, T. E., Walliker, D., Smith, C. L., do Rosario, V. E., Maloy, Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory W. L., Howard, R. J., Carter, R., and McCutchan, T. F. (1987) Cell Press, Cold Spring Harbor, NY 49,633-642
Evol. 30,522-562 55-62
263,6005-6008 Takeichi, M. (1991) Science 251,1451-1455
4 6 , 19-34 Von Heijne, G. (1983) Eur. J. Biochem. 133 , 17-21
487-491 Carter, R. (1987) Science 236,1661-1666





























