Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 257, No. 1. lasue of January 10. PP 477-4M. 1982 Printed m U S. A.
Lipid Effects on the Binding Properties of a Reconstituted Insulin Receptor*
(Received for publication, June 4, 1981)
Robert J. Gould, Barry H. Ginsberg, and Arthur A. Spector From the Departments of Biochemistry and Internal Medicine and the Diabetes-Endocrinology Research Center, The University of Ioula a n d Veterans Administration Medical Center, Iowa City, Iowa 52242
The turkey erythrocyte membrane insulin receptor was solubilized and reconstituted into vesicles com- posed of either soy or dimyristoyl phosphatidylcholine. Reconstitution with soy phosphatidylcholine provided a lipid environment containing 43% unsaturated fatty acids, as compared with 82% saturated fatty acids in the dimyristoyl phosphatidylcholine preparation. After reconstitution, both species of vesicles were isolated from a 2 to 30% continuous sucrose gradient at a density of 1.071 g/ml. Scatchard analysis of binding data ob- tained at 15 "C revealed that the reconstituted receptor had a greater affinity for ['251]iodoinsulin in the satu- rated lipid environment (Re = 0.167 1z"'; K1 = 2.18 m-') than in the unsaturated lipid environment ( K e = 0.0162 n"'; K1 = 0.479 IL"'). Low affinity binding also was increased in the saturated vesicles. These increases were paralleled by a reduction in the number of avail- able insulin binding sites in the saturated lipid environ- ment. There was no difference, however, in the relative affinity of the reconstituted receptor preparations for insulin or proinsulin. Electron microscopy and gel fil- tration indicated that the binding differences are not due to differences in vesicle size. They also are not due to differences in the orientation of the receptor within the lipid bilayer, for its sensitivity to trypsin digestion was similar in both types of vesicles. Solubilization studies with 1% 8-octylglucoside indicated, however, that the dimyristoyl phosphatidylcholine vesicles in- corporated a slightly lesser amount of insulin receptor. Similar results were also observed at 37 "C. These results suggest that the membrane lipid environment, especially the degree of unsaturation of the phospho- lipid fatty acyl chains, can influence the binding prop- erties of the insulin receptor.
The insulin receptor is a plasma membrane integral protein and, as such, is embedded within the lipid bilayer (Jarrett and Smith, 1974; Kahn, 1976). Because of the intimate contact with membrane lipids, it is possible that the structural and physical properties of adjacent phospholipids may affect the binding characteristics of the receptor. A number of studies have indirectly suggested that the physical state of the mem- brane lipids can influence insulin binding. For example, the dissociation of ['2sI]iodoinsulin from the insulin receptor of the IM-9 lymphoblast exhibits a decrease in activation energy at -21 "C (De Meyts et al., 1976) which corresponds to the
* This work was supported by research grants AM25295 and AM28516 from the National Institutes of Health. The costs of publi- cation of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertise- ment" in accordance wit% 18 U.S.C. Section 1734 solely to indicate this fact.
transition temperature of the cell membrane. A similar effect was observed with Friend erythroleukemia cells, where a concomitant decrease in the affinity of the receptor occurred at temperatures above this transition temperature (Ginsberg, 1981). Adipocytes also show a decrease in binding with in- creasing temperature, but this is due to a decrease in receptor number rather than a change in the affinit,y (Amatruda and Finch, 1979). These temperature-dependent binding altera- tions have been interpreted to result from a change in the physical properties of the membrane, such as fluidity (Gins- berg, 1981; Amatruda and Finch, 1979).
To further investigate this possibility, insulin binding was studied with Friend erythroleukemia cell preparations having altered membrane fatty acyl compositions (Ginsberg et al., 1979: Ginsberg et al., 1981). Increases in the degree of unsat- uration of the membrane phospholipids were associated with both an increase in the number of available insulin binding sites and a decrease in the affinity of these sites. Since the comparisons were all made a t 15 "C, the binding differences appear to be a response of the insulin receptor to a modifica- tion of the membrane lipid environment and not to a temper- ature-induced conformational change in the receptor itself. In support of this interpretation, the function of other plasma membrane proteins are known to be affected by lipid modifi- cations, including amino acid transport, glucose transport, and adenylate cyclase activity (Kaduce et al., 1977; Im et al., 1979; Im and Spector, 1980; Melchior and Czech, 1979; Engelhard et al., 1978). These proteins all mediate potentially insulin- sensitive processes. However, the possibility that the observed differences in insulin binding in Friend erythroleukemia cells may be due to some factor other than a direct effect of the membrane lipid modifications could not be excluded.
In the present study, we have utilized a solubilized and reconstituted turkey erythrocyte membrane insulin receptor preparation in an attempt to obtain more conclusive evidence as to whether the membrane lipid composition can directly influence insulin binding. This reconstituted system exhibits many of the characteristic properties of the membrane-bound insulin receptor (Gould et al., 1979). Our results demonstrate that the phospholipid fatty acyl composition can directly affect the binding properties of t,he insulin receptor. When incorporated into a more saturated lipid environment, the receptor has a higher affinity for insulin, but the insulin binding capacity is reduced.
MATERIALS AND METHODS
Solubilization and Reconstitution of Receptors-Turkey eryth- rocyte membranes were isolated as previously described (Ginsberg et al., 1976a). To solubilize the insulin receptor, 2.9 mg/ml of membrane protein were stirred with 1% P-octylglucoside at room temperature for 15 min. The buffer solution used throughout these studies con- tained 30 mM NaCI, 10 mM glucose, 1 rrrm EDTA and 85 mM Tris- HCI, pH 7.8 (Buffer I). After sedimentation a t 104,000 X g for 1 h a t
477
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

478 Lipid Effects on Insulin Binding
4 "C, the supernatant solution was collected and mixed with a 20-fold excess of a lipid mixture dissolved in 2% /3-octylglucoside. The unsat- urated lipid mixture contained soy phosphatidylcho1ine:bovine brain phosphatidylserine, 4:1 (w/w), while the saturated lipid mixture con- tained dimyristoyl phosphatidy1choline:bovine brain phosphatidylser- ine. 4:1 (w/w). The resulting solution was chromatographed over a Sephadex G-50 column equilibrated a t a temperature greater than the phase transition of the major phospholipid species in the mixture (Brunner et al., 1976; Gould et al., 1979). The void volume of the column was collected, dialyzed for 48 h, and centrifuged at 104,000 X g for 1 h a t 4 "C. The pellet was resuspended in a small volume of Buffer I and sedimented to equilibrium in a 2 to 30% continuous sucrose gradient. This separated the reconstituted material into two vesicle populations. One did not enter the gradient and consisted mainly of phospholipid. The other, which sedimented at a density of -1.071 g/ml and contained the insulin binding activity (Gould et al., 1979), was used for all further experimentation. Protein that was not associated with the lipid sedimented at the bottom of the gradient (Gould et al., 1979).
Binding Analysis-["51~iodoinsulin was prepared by a modifica- tion of the chloramine T method (Ginsberg et al., 1976a). The specific activity of the ["JI]iodoinsulin preparation varied from 150 to 200 Ci/ g. Insulin binding activity was measured in 0.5 ml of Buffer I contain- ing 1 mg/ml of bovine serum albumin and -20 PM [""Iliodoinsulin. Nonspecific or nonsaturable binding was defined as the [""Iliodoin- sulin which bound in the presence of 2 pg/ml of porcine insulin. All of the data has been corrected for this nonspecific binding (less than IOc& of tracer binding). Bound hormone was separated by precipitation with 12.5% polyethylene glycol, with 3 PM bovine y-globulin as carrier (Gould et al., 1979). After standing for 5 to 10 min a t room tempera- ture, the precipitate was collected on 0.45-nm cellulose acetate filters (Scheicher and Schiell, Keene, NH) using a Bio-Rad filter manifold under a vacuum pressure of 25 inches or mercury. Radioactivity retained by the filters was determined with a Beckman Gamma 8000 or a Micromedic 4/200 scintillation counter a t counting efficiencies of 75%.
Binding was measured by incubating the receptor with tracer amounts of ["51]iodoinsulin plus increasing amounts of unlabeled porcine insulin. The data were plotted by the method of Scatchard, and the binding parameters were calculated by a computerized anal- ysis that fits the competition curve to a fourth order polynomial.' This program generates a Scatchard plot of the fitted curve and analyzes the data by both the negative cooperativity (De Meyts and Roth, 1975) and independent classes of sites (Thakur and Rodbard, 1979) models. The data is visually presented on a Tectronix 4006 graphics terminal.
In order to determine the relative affinity of the reconstituted receptor, the preparations were incubated with 20 to 25 PM of [""I] iodoinsulin and increasing amounts of unlabeled hormone. Porcine insulin was obtained from Elanco (Indianapolis, IN), proinsulin was a gift of Dr. Ronald Chance (Eli Lilly, Indianapolis, IN), and desoc- tapeptide insulin was a gift from Dr. Joseph Walder (University of Iowa). Protein concentration was determined by a modification of the method of Lowry, with bovine serum albumin as the standard (Pe- tersen, 1977).
Lipid Analysis-The fatty acid composition of the reconstituted material was determined by gas-liquid chromatography. After addi- tion of 20 ml of chloroformmethanol, 2:1 (v/v) and 4 ml of 0.04 N HCI in 0.9% NaCI, the chloroform phase which contained the phospholip- ids was isolated. Lipids were saponified in ethanolic KOH and meth- ylated with 14% BF:, in methanol as previously described (McGee and Spector, 1975). The fatty acid methyl esters were extracted into heptane and chromatographed on a glass column (1.9 m X 2 mm) containing 10% SP-2330 on 100 to 120 mesh Chromasorb W/AW (Supelco, Bellefonte, PA). A Hewlett I'ackard 5710A gas chromato- graph equipped with a flame ionization detector and a 3380A integra- tor was employed. N2 served as a carrier gas, the flow rate being 60 ml/min. Fatty acid methyl ester standards, obtained from Applied Science Laboratories (State College, PA) or Supelco, were utilized to identify the various peaks.
Phospholipids were measured colorimetrically with ammonium molybdate (Haheja et al., 1973) or with malachite green after digestion with perchloric acid (Chalvardjian and Rudnicki, 1970).
Electron Microscopy-The reconstituted receptor preparations were negatively stained with phosphotungstic acid, pH 7.0. on Form-
I B. H. Ginsberg, R. J. Spanheimer, and M. Granner, submitted for ""
publication.
var and carbon-coated copper grids (400 mesh). Samples were viewed on a Hitachi H-600 electron microscope under an accelerating voltage of 50 kV.
RESULTS
Characterization of the Reconstituted Vesicles-Most of the physical and biochemical characteristics of the insulin receptors reconstituted into vesicles with either dimyristoyl phosphotidylcholine or soy phosphatidylcholine plus phos- phatidylserine were the same. The reconstituted vesicles were separated from unreconstituted protein and lipid by sucrose gradient centrifugation (Gould et al., 1979) and banded at a density of 1.071 g/ml. Therefore, the ratio of lipid to protein was similar in both reconstituted vesicle preparations. As seen in Fig. 1, reconstitution with either dimyristoyl phosphatidyl-
4
I FIG. 1. Electron microscopy of reconstituted vesicles. Solu-
bilized turkev erythrocyte membrane protein was reconstituted with dim.yristoyl phosphatid.vlcholine ( A ) or soy phosphatidylcholine ( B ) plus phosphatidylserine. After sucrose gradient centrifugation, the preparations were negatively stained with 2% phosphotungstic acid. The bar indicates 0.5 pm.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Lipid Effects on Insulin Binding 479
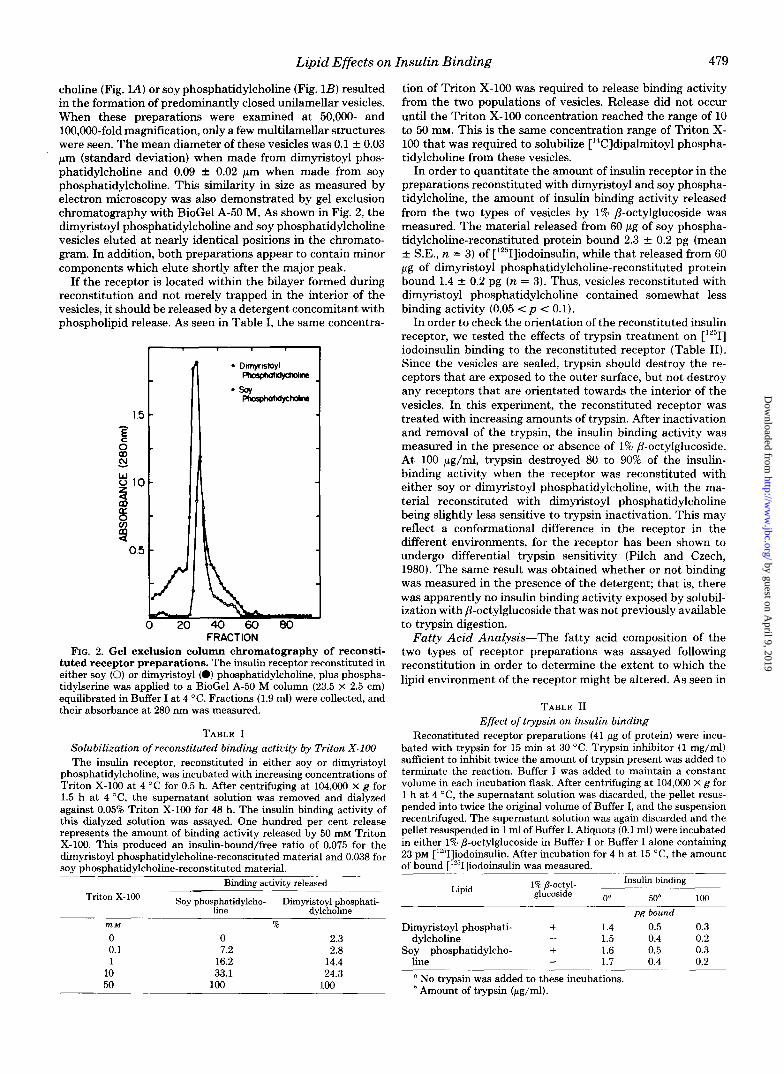
choline (Fig. lA) or soy phosphatidylcholine (Fig. 1B) resulted in the formation of predominantly closed unilamellar vesicles. When these preparations were examined at 50,000- and 100,000-fold magnification, only a few multilamellar structures were seen. The mean diameter of these vesicles was 0.1 2 0.03 pm (standard deviation) when made from dimyristoyl phos- phatidylcholine and 0.09 & 0.02 pm when made from soy phosphatidylcholine. This similarity in size as measured by electron microscopy was also demonstrated by gel exclusion chromatography with BioGel A-50 M. As shown in Fig. 2, the dimyristoyl phosphatidylcholine and soy phosphatidylcholine vesicles eluted at nearly identical positions in the chromato- gram. In addition, both preparations appear to contain minor components which elute shortly after the major peak.
If the receptor is located within the bilayer formed during reconstitution and not merely trapped in the interior of the vesicles, it should be released by a detergent concomitant with phospholipid release. As seen in Table I, the same concentra-
1.5
0 QD (u
s - g 10 Q v)
m a 0 5
0 2 0 4 0 6 0 8 0 FRACTION
FIG. 2. Gel exclusion column chromatography of reconsti- tuted receptor preparations. The insulin receptor reconstituted in either soy (0) or dimyristoyl (0) phosphatidylcholine, plus phospha- tidylserine was applied to a BioGel A-50 M column (23.5 X 2.5 cm) equilibrated in Buffer I a t 4 "C. Fractions (1.9 m l ) were collected, and their absorbance at 280 nm was measured.
TABLE I Solubilization of reconstituted binding activity by Triton X-100 The insulin receptor, reconstituted in either soy or dimyristoyl
phosphatidylcholine, was incubated with increasing concentrations of Triton X-100 at 4 "C for 0.5 h. After centrifuging at 104,000 X g for 1.5 h at 4 "C, the supernatant solution was removed and dialyzed against 0.05% Triton X-100 for 48 h. The insulin binding activity of this dialyzed solution was assayed. One hundred per cent release represents the amount of binding activity released by 50 m~ Triton X-100. This produced an insulin-bound/free ratio of 0.075 for the dimyristoyl phosphatidylcholine-reconstituted material and 0.038 for soy phosphatidylcholine-reconstituted material.
Binding activity released
Triton x-100 Soy phosphatidylcho- Dimyristoyl phosphati- line dylcholine
mM
0 0.1 1 10 50
% 0 2.3 7.2 2.8
16.2 14.4 33.1 24.3
100 100
tion of Triton X-100 was required to release binding activity from the two populations of vesicles. Release did not occur until the Triton X-100 concentration reached the range of 10 to 50 m. This is the same concentration range of Triton X- 100 that was required to solubilize ['4C]dipalmitoyl phospha- tidylcholine from these vesicles.
In order to quantitate the amount of insulin receptor in the preparations reconstituted with dimyristoyl and soy phospha- tidylcholine, the amount of insulin binding activity released from the two types of vesicles by 1% P-octylglucoside was measured. The material released from 60 pg of soy phospha- tidylcholine-reconstituted protein bound 2.3 k 0.2 pg (mean f. S.E., n = 3) of ['251]iodoinsulin, while that released from 60 pg of dimyristoyl phosphatidylcholine-reconstituted protein bound 1.4 f 0.2 pg (n = 3). Thus, vesicles reconstituted with dimyristoyl phosphatidylcholine contained somewhat less binding activity (0.05 < p < 0.1).
In order to check the orientation of the reconstituted insulin receptor, we tested the effects of trypsin treatment on [1251] iodoinsulin binding to the reconstituted receptor (Table 11). Since the vesicles are sealed, trypsin should destroy the re- ceptors that are exposed to the outer surface, but not destroy any receptors that are orientated towards the interior of the vesicles. In this experiment, the reconstituted receptor was treated with increasing amounts of trypsin. After inactivation and removal of the trypsin, the insulin binding activity was measured in the presence or absence of 1% /I-octylglucoside. At 100 pg/ml, trypsin destroyed 80 to 90% of the insulin- binding activity when the receptor was reconstituted with either soy or dimyristoyl phosphatidylcholine, with the ma- terial reconstituted with dimyristoyl phosphatidylcholine being slightly less sensitive to trypsin inactivation. This may reflect a conformational difference in the receptor in the different environments, for the receptor has been shown to undergo differential trypsin sensitivity (Pilch and Czech, 1980). The same result was obtained whether or not binding was measured in the presence of the detergent; that is, there was apparently no insulin binding activity exposed by solubil- ization with P-octylglucoside that was not previously available to trypsin digestion.
Fatty Acid Analysis-The fatty acid composition of the two types of receptor preparations was assayed following reconstitution in order to determine the extent to which the lipid environment of the receptor might be altered. As seen in
TABLE I1 Effect of trypsin on insulin binding
Reconstituted receptor preparations (41 pg of protein) were incu- bated with trypsin for 15 min at 30 "C. Trypsin inhibitor (1 mg/ml) sufficient to inhibit twice the amount of trypsin present was added to terminate the reaction. Buffer I was added to maintain a constant volume in each incubation flask. After centrifuging at 104,000 X g for 1 h at 4 "C, the supernatant solution was dscarded, the pellet resus- pended into twice the original volume of Buffer I, and the suspension recentrifuged. The supernatant solution was again discarded and the pellet resuspended in 1 ml of Buffer I. Aliquots (0.1 ml) were incubated in either 1% P-octylglucoside in Buffer I or Buffer I alone containing 23 PM ['251]iodoinsulin. After incubation for 4 h at 15 "C, the amount of bound r'2511iodoinsulin was measured.
Lipid 1% p-octy1- Insulin binding glucoside oo 50* 100
p g bound
Dimyristoyl phosphati- + 1.4 0.5 0.3 dylcholine - 1.5 0.4 0.2
Soy phosphatidylcho- + 1.6 0.5 0.3 line - 1.7 0.4 0.2
a No trypsin was added to these incubations. Amount of trypsin (pg/ml).
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
480 Lipid Effects on Insulin Binding
Table III, the predominant fatty acid after reconstitution with soy phosphatidylcholine is linoleic acid (18:2), which com- prises 35% of the total fatty acids. This is consistent with the fact that 52% of the fatty acids of the soy phosphatidylcholine and phosphatidylserine mixture is linoleic acid. By contrast, 66% of the total fatty acids in the preparation reconstituted with dimyristoyl phosphatidylcholine was myristic acid (14:0), and only 4% was linoleic acid. These differences result in a lipid environment that contains 43% polyunsaturated and 30% saturated fatty acids in the soy phosphatidylcholine-reconsti- tuted preparation, as opposed to 5% polyunsaturated and 82% saturated fatty acids in the dimyristoyl phosphatidylcholine reconstituted preparation.
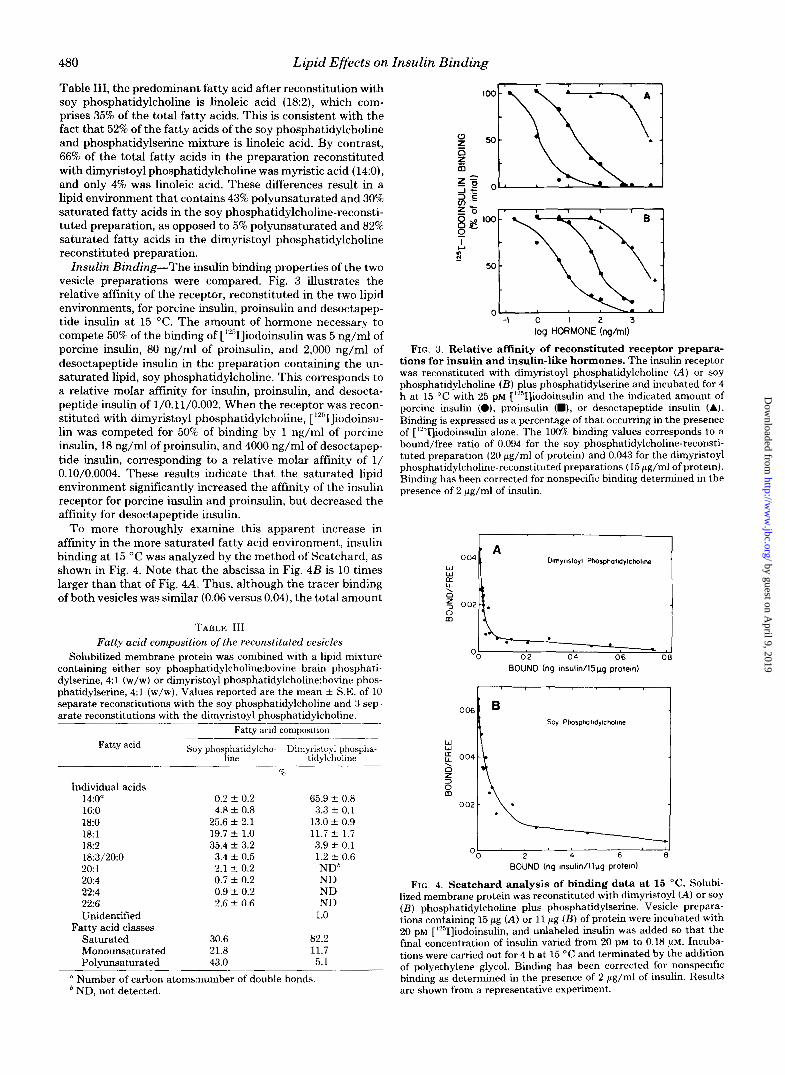
Insulin Binding-The insulin binding properties of the two vesicle preparations were compared. Fig. 3 dustrates the relative affinity of the receptor, reconstituted in the two lipid environments, for porcine insulin, proinsulin and desoctapep- tide insulin at 15 "C. The amount of hormone necessary to compete 50% of the binding of ['"I]iodoinsulin was 5 ng/ml of porcine insulin, 80 n g / d of proinsulin, and 2,000 ng/ml of desoctapeptide insulin in the preparation containing the un- saturated lipid, soy phosphatidylcholine. This corresponds to a relative molar affinity for insulin, proinsulin, and desocta- peptide insulin of 1/0.11/0.002. When the receptor was recon- stituted with dimyristoyl phosphatidylcholine, ['251]iodoinsu- lin was competed for 50% of binding by 1 ng/ml of porcine insulin, 18 ng/ml of proinsulin, and 4000 ng/ml of desoctapep- tide insulin, corresponding to a relative molar affinity of 1/ 0.10/0.0004. These results indicate that the saturated lipid environment significantly increased the affinity of the insulin receptor for porcine insulin and proinsulin, but decreased the affinity for desoctapeptide insulin.
To more thoroughly examine this apparent increase in affinity in the more saturated fatty acid environment, insulin binding at 15 "C was analyzed by the method of Scatchard, as shown in Fig. 4. Note that the abscissa in Fig. 4B is 10 times larger than that of Fig. 4-4. Thus, although the tracer binding of both vesicles was similar (0.06 versus 0.04), the total amount
TABLE I11 Fatty acid composition of the reconstituted vesicles
Solubilized membrane protein was combined with a lipid mixture containing either soy phosphatidylcho1ine:bovine brain phosphati- dylserine, 4:l (w/w) or dimyristoyl phosphatidy1choline:bovine phos- phatidylserine, 4:l (w/w). Values reported are the mean f S.E. of 10 separate reconstitutions with the soy phosphatidylcholine and 3 sep- arate reconstitutions with the dimyristoyl phosphatidylcholine.
~- Fatty acid composition
Fatty acid Soy phosphatidylcho- Dimyristoyl phospha-
" _ _ _ _ _ ~ line tidylcholine "c
~. ~
Individual acids 14:O" 0.2 & 0.2 65.9 f 0.8 16:O 4.8 f 0.8 3.3 f 0.1 18:O 25.6 f 2.1 13.0 f 0.9 18:l 19.7 f 1.0 11.7 f 1.7 182 35.4 f 3.2 3.9 f 0.1 18:3/20:0 3.4 f 0.5 1.2 % 0.6 20 1 2.1 f 0.2 NDh 20:4 0.7 f 0.2 ND 22:4 0.9 rt_ 0.2 ND 2 2 6 2.6 -t 0.6 ND Unidentified 1 .o Saturated 30.6 82.2 Monounsaturated 21.8 11.7 Polyunsaturated 43.0 5.1
Fatty acid classes
" Number of carbon atomxnumber of double bonds. * ND, not detected.
. ~~
" I
0 I H N -
l o o -
50-
0 '
I 1
l o o -
50 :,-Y$; 0 - 1 0 1 2 3
log HORMONE (ng/rnl)
FIG. 3. Relative affinity of reconstituted receptor prepara- tions for insulin and insulin-like hormones. The insulin receptor was reconstituted with dimyristoyl phosphatidylcholine (A) or soy phosphatidylcholine ( B ) plus phosphatidylserine and incubated for 4 h at 15 "C with 25 PM [lYSI]iodoinsulin and the indicated amount of porcine insulin (O), proinsulin (m), or desoctapeptide insulin (A). Binding is expressed as a percentage of that occurring in the presence of [1251]iodoinsulin alone. The 100% binding values corresponds to a bound/free ratio of 0.094 for the soy phosphatidylcholine-reconsti- tuted preparation (20 gg/ml of protein) and 0.043 for the dimyristoyl phosphatidylcholine-reconstituted preparations (15 pg/ml of protein). Binding has been corrected for nonspecific binding determined in the presence of 2 pg/ml of insulin.
00 Oil 0 4 06 0
BOUND (ng 1nsulm/15kg proteln)
LL1
0 m
Oo4 002
O b ' ' ' 4 ' 6 ' 8 ' 2 BOUND (nq Insulln/llpg proteln)
FIG. 4. Scatchard analysis of binding data at 15 "C. Solubi- lized membrane protein was reconstituted with dimyristoyl (A) or soy ( B ) phosphatidylcholine plus phosphatidylserine. Vesicle prepara- tions containing 15 pg (A) or 11 gg ( B ) of protein were incubated with 20 PM [lYSI]iodoinsulin, and unlabeled insulin was added so that the final concentration of insulin varied from 20 PM to 0.18 pM. Incuba- tions were carried out for 4 h at 15 "C and terminated by the addition of polyethylene glycol. Binding has been corrected for nonspecific binding as determined in the presence of 2 gg/ml of insulin. Results are shown from a representative experiment.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Lipid Effects on Insulin Binding 48 1
TABLE IV
Binding model
Two-site model KI (nM- I )
KZ (nM") n I (pmol/mg of protein) n2 (pmol/mg of protein)
Negative-cooperativity model Re (nM-:) K , (nM- )
~~~~~~
Parameters from Scatchard analysis of binding data " ~
Binding parameters
15 "C 37 "C ~ _ _ _ ~ ~ ~ ~~
Saturated" - ---___
Unsaturated* Saturated ~~ ~~ ""
Unsaturatedh
2.18 f 0.90.'' 0.479 f 0.158 0.82 f 0.25 0.491 f 0.21 0.025 f 0.005' 0.0026 f 0.0011 0.0242 f 0.0059' 0.0048 f 0.0018
0.6 7 3 3 12 370 23 80
0.167 f 0.017' 0.0162 f 0.0019 0.0878 f 0.0086 0.F201 f 0.045 0.038 f O.Uh 0.0013 f 0.0002 0.0242 f 0.0059' 0.0048 f 0.0017
n'(pmo1jmg of protein) 13 ___ _ _ _ _ _ _ _ _ _ _ _ _ ~ ~
" 3 separate reconstitutions and determinations. ' 4 separate reconstitutions and determinations. ' Mean f S.E. d p < 0.1 uersus unsaturated at 15 "C. ' p < 0.01 uersus unsaturated system a t 15 "C. ' p < 0.025 uersus unsaturated system at 37 "C. # p < 0.001 uersus unsaturated system a t same temperature. h p < 0.05 uersus unsaturated system at 15 "C.
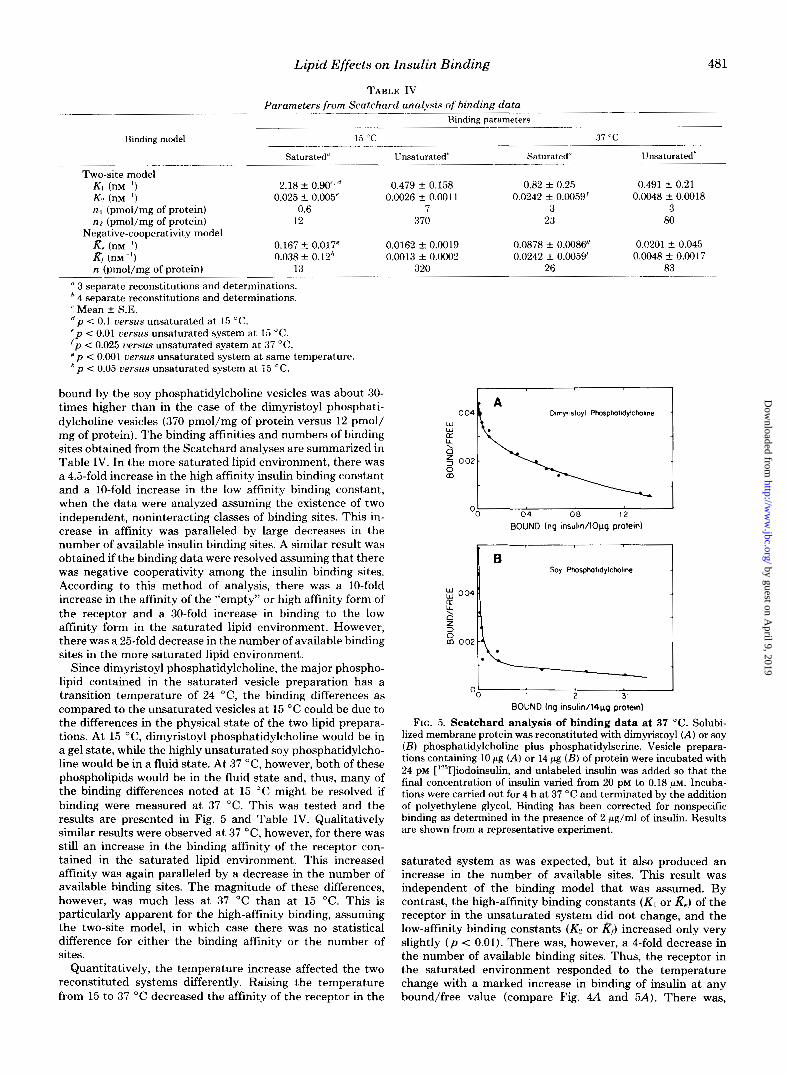
bound by the soy phosphatidylcholine vesicles was about 30- times higher than in the case of the dimyristoyl phosphati- dylcholine vesicles (370 pmol/mg of protein versus 12 pmol/ mg of protein). The binding affinities and numbers of binding sites obtained from the Scatchard analyses are summarized in Table IV. In the more saturated lipid environment, there was a 4.5-fold increase in the high affinity insulin binding constant and a 10-fold increase in the low affinity binding constant, when the data were analyzed assuming the existence of two independent, noninteracting classes of binding sites. This in- crease in affinity was paralleled by large decreases in the number of available insulin binding sites. A similar result was obtained if the binding data were resolved assuming that there was negative cooperativity among the insulin binding sites. According to this method of analysis, there was a 10-fold increase in the affinity of the "empty" or high affinity form of the receptor and a 30-fold increase in binding to the low affinity form in the saturated lipid environment. However, there was a 25-fold decrease in the number of available binding sites in the more saturated lipid environment.
Since dimyristoyl phosphatidylcholine, the major phospho- lipid contained in the saturated vesicle preparation has a transition temperature of 24 "C, the binding differences as compared to the unsaturated vesicles at 15 "C could be due to the differences in the physical state of the two lipid prepara- tions. At 15 "C, dimyristoyl phosphatidylcholine would be in a gel state, while the highly unsaturated soy phosphatidylcho- line would be in a fluid state. At 37 "C, however, both of these phospholipids would be in the fluid state and, thus, many of the binding differences noted a t 15 "C might be resolved if binding were measured at 37 "C. This was tested and the results are presented in Fig. 5 and Table IV. Qualitatively similar results were observed at 37 "C, however, for there was still an increase in the binding affinity of the receptor con- tained in the saturated lipid environment. This increased affinity was again paralleled by a decrease in the number of available binding sites. The magnitude of these differences, however, was much less a t 37 "C than at 15 "C. This is particularly apparent for the high-affinity binding, assuming the two-site model, in which case there was no statistical difference for either the binding affinity or the number of sites.
Quantitatively, the temperature increase affected the two reconstituted systems differently. Raising the temperature from 15 to 37 "C decreased the affinity of the receptor in the
r I
0 04 k A Dlrnyrlsloyl Phosphalldylctmlme
1 04 08 12
BOUND (ng Insulln/lOpg protem)
Soy Ptmsphalldylchallne
I I 2 3
BOUND (ng 1nsulin/l4pg protem)
FIG. 5. Scatchard analysis of binding data at 37 "C. Solubi- lized membrane protein was reconstituted with dimyristoyl (A) or soy ( B ) phosphatidylcholine plus phosphatidylserine. Vesicle prepara- tions containing 10 pg (A) or 14 pg ( B ) of protein were incubated with 24 PM ['"I]iodoinsulin, and unlabeled insulin was added so that the final concentration of insulin varied from 20 PM to 0.18 PM. Incuba- tions were carried out for 4 h at 37 "C and terminated by the addition of polyethylene glycol. Binding has been corrected for nonspecific binding as determined in the presence of 2 pg/ml of insulin. Results are shown from a representative experiment.
saturated system as was expected, but it also produced an increase in the number of available sites. This result was independent of the binding model that was assumed. By contrast, the high-affinity binding constants (K1 or Re) of the receptor in the unsaturated system did not change, and the low-affinity binding constants (Kz or &) increased only very slightly ( p < 0.01). There was, however, a 4-fold decrease in the number of available binding sites. Thus, the receptor in the saturated environment responded to the temperature change with a marked increase in binding of insulin at any bound/free value (compare Fig. 4A and 5A). There was,
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
482 Lipid Effects on Insulin Binding
0- 2 3 4 5
TIME (hr) FIG. 6. Association of [125KJiodoinsulin with reconstituted re-
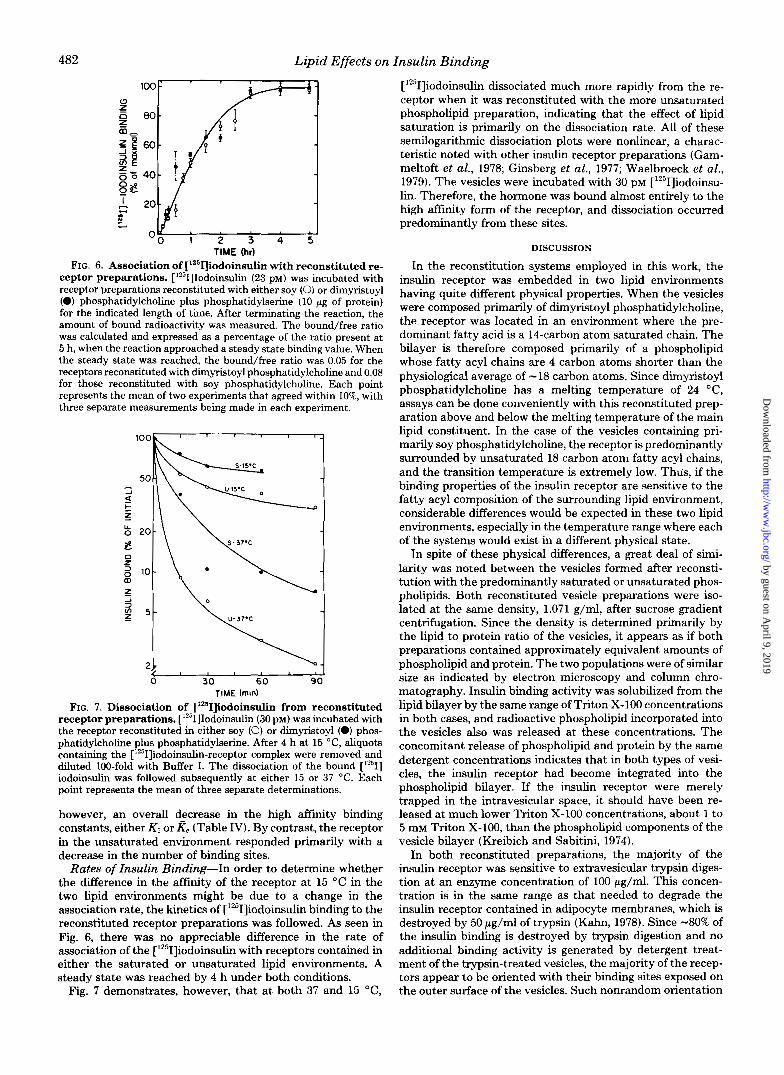
ceptor preparations. ['251]Iodoinsulin (23 p ~ ) was incubated with receptor preparations reconstituted with either soy (0) or dimyristoyl (0) phosphatidylcholine plus phosphatidylserine (10 pg of protein) for the indicated length of time. After terminating the reaction, the amount of bound radioactivity was measured. The bound/free ratio was calculated and expressed as a percentage of the ratio present at 5 h, when the reaction approached a steady state binding value. When the steady state was reached, the bound/free ratio was 0.05 for the receptors reconstituted with dimyristoyl phosphatidylcholine and 0.08 for those reconstituted with soy phosphatidylcholine. Each point represents the mean of two experiments that agreed within IO%, with three separate measurements being made in each experiment.
TIME (mm)
FIG. 7. Dissociation of ['26KJiodoinsulin from reconstituted receptor preparations. ['251]Iodoinsulin (30 PM) was incubated with the receptor reconstituted in either soy (0) or dimyristoyl (0) phos- phatidylcholine plus phosphatidylserine. After 4 h at 15 "C, aliquots containing the ['251]iodoinsulin-receptor complex were removed and diluted 100-fold with Buffer I. The dissociation of the bound [1251] iodoinsulin was followed subsequently at either 15 or 37 "C. Each point represents the mean of three separate determinations.
however, an overall decrease in the high affinity binding constants, either Kl or Re (Table IV). By contrast, the receptor in the unsaturated environment responded primarily with a decrease in the number of binding sites.
Rates of Insulin Binding-In order to determine whether the difference in the affinity of the receptor at 15 "C in the two lipid environments might be due to a change in the association rate, the kinetics of [1251]iodoinsulin binding to the reconstituted receptor preparations was followed. As seen in Fig. 6, there was no appreciable difference in the rate of association of the ['251]iodoinsulin with receptors contained in either the saturated or unsaturated lipid environments. A steady state was reached by 4 h under both conditions.
Fig. 7 demonstrates, however, that at both 37 and 15 "C,
['251]iodoinsulin dissociated much more rapidly from the re- ceptor when it was reconstituted with the more unsaturated phospholipid preparation, indicating that the effect of lipid saturation is primarily on the dissociation rate. All of these semilogarithmic dissociation plots were nonlinear, a charac- teristic noted with other insulin receptor preparations (Gam- meltoft et al., 1978; Ginsberg et al., 1977; Waelbroeck et al., 1979). The vesicles were incubated with 30 PM [1251]iodoins~- lin. Therefore, the hormone was bound almost entirely to the high affinity form of the receptor, and dissociation occurred predominantly from these sites.
DISCUSSION
In the reconstitution systems employed in this work, the insulin receptor was embedded in two lipid environments having quite different physical properties. When the vesicles were composed primarily of dimyristoyl phosphatidylcholine, the receptor was located in an environment where the pre- dominant fatty acid is a 14-carbon atom saturated chain. The bilayer is therefore composed primarily of a phospholipid whose fatty acyl chains are 4 carbon atoms shorter than the physiological average of -18 carbon atoms. Since dimyristoyl phosphatidylcholine has a melting temperature of 24 "C, assays can be done conveniently with this reconstituted prep- aration above and below the melting temperature of the main lipid constituent. In the case of the vesicles containing pri- marily soy phosphatidylcholine, the receptor is predominantly surrounded by unsaturated 18 carbon atom fatty acyl chains, and the transition temperature is extremely low. Thus, if the binding properties of the insulin receptor are sensitive to the fatty acyl composition of the surrounding lipid environment, considerable differences would be expected in these two lipid environments, especially in the temperature range where each of the systems would exist in a different physical state.
In spite of these physical differences, a great deal of simi- larity was noted between the vesicles formed after reconsti- tution with the predominantly saturated or unsaturated phos- pholipids. Both reconstituted vesicle preparations were iso- lated at the same density, 1.071 g/ml, after sucrose gradient centrifugation. Since the density is determined primarily by the lipid to protein ratio of the vesicles, it appears as if both preparations contained approximately equivalent amounts of phospholipid and protein. The two populations were of similar size as indicated by electron microscopy and column chro- matography. Insulin binding activity was solubilized from the lipid bilayer by the same range of Triton X-100 concentrations in both cases, and radioactive phospholipid incorporated into the vesicles also was released at these concentrations. The concomitant release of phospholipid and protein by the same detergent concentrations indicates that in both types of vesi- cles, the insulin receptor had become integrated into the phospholipid bilayer. If the insulin receptor were merely trapped in the intravesicular space, it should have been re- leased at much lower Triton X-100 concentrations, about 1 to 5 mM Triton X-100, than the phospholipid components of the vesicle bilayer (Kreibich and Sabitini, 1974).
In both reconstituted preparations, the majority of the insulin receptor was sensitive to extravesicular trypsin diges- tion at an enzyme concentration of 100 pg/ml. This concen- tration is in the same range as that needed to degrade the insulin receptor contained in adipocyte membranes, which is destroyed by 50 p g / d of trypsin (Kahn, 1978). Since -80% of the insulin binding is destroyed by trypsin digestion and no additional binding activity is generated by detergent treat- ment of the trypsin-treated vesicles, the majority of the recep- tors appear to be oriented with their binding sites exposed on the outer surface of the vesicles. Such nonrandom orientation
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Lipid Effects on Insulin Binding 483
has also been observed for the acetylcholine receptor and the G protein of vesicular stomatitis virus when they are recon- stituted following solubilization with P-octylglucoside (Gon- zalez-Ros et al., 1980; Petri and Wagner, 1979). Therefore, reconstitution from P-octylglucoside results in an asymmetric orientation in the case of each of these proteins, with the binding sites of the receptors being exposed on the exterior surface of the vesicles.
Reconstitution apparently does not cause any major alter- ation in the relative affinity of the receptor for various insulin- like hormones, except for a slight decrease in the affinity of the receptor for desoctapeptide insulin in the more saturated lipid environment. As opposed to this finding, the thyrotropin receptor reportedly shows major alterations in specificity for thyrotropin and tetanus toxin, depending on whether the surrounding lipid is dipalmitoyl phosphatidylcholine or di- oleoyl phosphatidylcholine (Lee et al., 1978). The failure to see similar changes in specificity with the reconstituted insulin receptor suggests that such major binding effects do not result from lipid interactions with receptors in all cases.
The increase in affinity in the saturated lipid environment indicates, however, that the binding properties of the insulin receptor are sensitive to some extent to the surrounding structure of the membrane lipid bilayer. The saturated phos- pholipid environment increased the low affinity binding con- stant of the insulin receptor a t both 37 and 15 "C. At 37 "C, however, the high affinity binding constant did not differ in the two lipid environments (0.3 < p c 0.4). It would seem, then, that saturated lipids can influence the affinity of the receptor under some conditions. Part of the observed effect may be due to the 14-carbon atom chains that constitute 66% of the fatty acyl groups in the dimyristoyl phosphatidylcho- line-reconstituted vesicles. Even if this were the case, it still demonstrates that the membrane lipids can have a direct effect on the binding properties of the insulin receptor, whether it be through a change in fatty acyl saturation or chain length. Our findings suggest that the greater affinity is due to a decrease in the dissociation rate of ['251]iodoinsulin.
Secondly, the saturated lipid environment causes a very large decrease in the number of available insulin binding sites. Some of these sites reappear when the temperature is raised to 37 "C, suggesting that this occurs through a direct, physical effect of the membrane structure. The large decrease in the number of available binding sites when the receptor is con- tained in the more saturated environment cannot be due exclusively to a decrease in the content of receptor protein, for there was at most a 2-fold decrease in the amount of binding activity released by exposure to a 1% P-octylglucoside solution. Since the trypsin studies indicate that there is no appreciable difference in the orientation of the receptor in the two reconstituted preparations, the binding differences also cannot be attributed to a change in the relative proportion of receptor molecules accessible to the extravesicular fluid.
Because these studies were done with impure mixtures that are highly complex, the data do not lend themselves to a detailed analysis of the molecular mechanisms responsible for the binding differences. The subunit structure of the insulin receptor has been described in detail through the use of chemical cross-linking agents, and some progress is being made concerning how the subunit structure may play a role in the transmission of the biological signal (Massague et al., 1981; Seals and Czech, 1981). The receptor has only recently been purified as judged by immunological criteria, however, and it is still very difficult to obtain a purified preparation that retains binding activity (Harrison and Itin, 1980). More- over, it is still not possible to prepare purified material in the amounts required for reconstitution and binding studies. Be-
cause of these technological limitations, the receptor prepa- ration utilized for this work was purified only -10-fold, with respect to protein, relative to the turkey erythrocyte mem- brane. Although this introduces uncertainties regarding inter- pretation, there appears to be definite value in testing a relative crude reconstituted preparation. It has been suggested that the insulin receptor may interact with one or more regulatory proteins (Krupp and Livingston, 1979; Harmon et al., 1980). Therefore, the use of the impure membrane protein preparation lessens the possibility of removing a potentially important regulatory protein.
The insulin receptor may exist in two forms, whose inter- conversion is mediated by insulin (Ginsberg et al., 1976b; Maturo and Hollenberg, 1978 Krupp and Livingston, 1978). According to this model, there is a 40 .& low affinity form of the receptor which can self-associate to a 72 A high affinity form. If the insulin receptor is contained in a saturated, and presumably less fluid lipid environment, there may be a greater tendency for it to associate, and thereby be present in the high affinity state. Such a mechanism would explain the increased affinity observed with the dimyristoyl phosphati- dylcholine system when the comparisons were made at 15 "C. Since self-association could reduce the number of available sites, the decrease observed in the saturated system also can be explained by such a process. In the unsaturated environ- ment or in the more saturated system a t 37 "C, the lipid environment would be more fluid and probably facilitate dissociation to the low affinity form. This also might lead to a greater accessibility of binding sites, accounting for their increased availability. Whereas the subunit association model is consistent with many of our findings, it does not explain the decreased number of binding sites observed when the receptor is reconstituted in the unsaturated system and assayed a t 37 "C, as compared to 15 "C.
In summary, these results demonstrate that the structure of the surrounding lipid environment can directly influence the properties of the insulin receptor. This conclusion is consistent with information obtained from cultured cell and isolated membrane studies (De Meyts et al., 1976; Ginsberg, 1981; Ginsberg et al., 1981) and provides a more direct dem- onstration of lipid sensitivity than was previously available. Although information regarding possible functwnal effects is not available, the findings suggest that localized changes in the membrane lipid microenvironment could play some role in modulating the interaction of insulin with cells.
REFERENCES Amatruda, J. M., and Finch, E. D. (1979) J. Biol. Chem. 254, 2619-
Brunner, J., Skrabal, P., and Hauser, H. (1976) Biochim. Biophys.
Chalvardjian, A., and Rudnicki, E. (1970) Anal. Biochem. 36, 225-
De Meyts, P., and Roth, J. (1975) Biochem. Biophys. Res. Commun.
De Meyts, P., Bianco, A. R., and Roth, J. (1976) J. Biol. Chem. 251,
Engelhard, V. H., Glaser, M., and Storm, D. R. (1978) Biochemistry
Gammeltoft, S., Kristensen, L. O., and Sestoft, L. (1978) J. Biol.
Ginsberg, B. (1982) in Biological Growth and Regulation (Yama-
Ginsberg, B. H., Kahn, C. R., and Roth, J. (1976a) Biochim. Biophys.
Ginsberg, B. H., Kahn, C. R., Roth, J., and De Meyts, P. (1976b)
Ginsberg, B. H., Kahn, S. R., and Roth, J. (1977) Endocrinology 100,
Ginsberg, B. H., Brown, T., Gould, R., and Spector, A. A. (1979) Clin.
2625
Acta 455,322-331
226
66, 1118-1126
1877-1888
17, 3191-3200
Chem. 253,8406-8413
moto, K., ed) Plenum Press, New York, in press
Acta 443, 227-242
Biochem. Biophys. Res. Commun. 73, 1068-1074
82-90
Res. 27, 655 (abstr.)
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

484 Lipid Effects on Insulin Binding
Ginsberg, B. H., Brown, T. J., Simon, I., and Spector, A. A. (1981)
Gonzalez-Ros, J. M., Paraschos, A,, and Martinez-Carrion, M. (1980)
Gould, R. J., Ginsberg, B. H., and Spector, A. A. (1979) Endocrin.
Harmon, J. T., Kahn, C . R., Kempner, E. S., and Schlegel, W. (1980)
Harrison, L. C., and Itin, A. (1980) J. Biol. Chem. 255, 12006-12072 Im, W. B., Deutchler, J. T., and Spector, A. A. (1979) Lipids 14,1003-
Im, W. B., and Spector, A. A. (1980) J. Biol. Chem. 255, 764-770 Jarrett, L., and Smith, R. (1974) J. Biol. Chem. 249, 7024-7031 Kaduce, T. L., Awad, A. B., Fontenelle, L. J., and Spector, A. A.
Kahn, C. R. (1976) J . Cell Biol. 70, 261-286 Kahn, C. R. (1978) Metabolism 27, 1893-1900 Kreibich, G., and Sabatini, D. D. (1974) Methods Enzymol. 31, 215-
Krupp, M. N., and Livingston, J. N. (1978) Proc. Natl. Acad. Sci. U.
Diabetes 30, 773-780
Proc. Natl. Acad. Sci. U. 5'. A . 77, 1796-1800
Res. Cvmmun. 6,279-290
J . Biol. Chem. 255, 3412-3419
1008
(1977) J. Biol. Chem. 252,6624-6630
225
S. A . 75, 2593-2597
L. D. (1978) Biochem. Biophys. Res. Commun. 83, 313-320 Lee, G., Consiglio, E., Habig, W., Dyer, S., Hardegree, C., and Kohn,
McGee, R., and Spector, A. A. (1975) J . Biol. Chem. 250,5419-5425 Massague, J., Pilch, P. F., and Czech, M. P. (1981) J. Biol. Chem.
Maturo, J. M., 111, and Hollenberg, M. D. (1978) Proc. Natl. Acad.
Melchior, D. L., and Czech, M. P. (1979) J. Biol. Chem. 254, 8744-
Petersen, G. L. (1977) Anal. Biochem. 83, 346-356 Petri, W. A,, Jr., and Wagner, R. R. (1979) J. Biol. Chem. 254,4313-
Pilch, P. A,, and Czech, M. P. (1980) Science 210, 1152-1153 Raheja, P. K., Kaur, C., Singh, A., and Bhatia, I. S. (1973) J. Lipid
Seals, J. R., and Czech, M. P. (1981) J. Biol. Chem. 256, 2894-2899 Thakur, A. K., and Rodbard, D. (1979) J. Theor. Biol. 80,383-403 Waelbroeck, M., Obberghen, E., and De Meyts, P. (1979) J . Biol.
256,3182-3190
Sci. U. S. A. 75, 3070-3074
8747
4316
Res. 14,695-697
Chem. 254, 7736-7740
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

R J Gould, B H Ginsberg and A A SpectorLipid effects on the binding properties of a reconstituted insulin receptor.
1982, 257:477-484.J. Biol. Chem.
http://www.jbc.org/content/257/1/477Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/257/1/477.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























