Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 257. No 21, Issue of November 10. pp. 13018-13027, 1982 Prrnted in U S A .
Hormonal Regulation of the Conformation of the Ovalbumin Gene in Chick Oviduct Chromatin*
(Received for publication, February 8, 1982)
Kerry S. Bloom+ and John N. Anderson From the Department of Biological Sciences, Purdue Uniuersity, West Lafayette, Indiana 47907
We have examined the effects of steroid hormones on the chromatin sensitivity of- the ovalbumin gene to micrococcal nuclease and have attempted to define the importance of the nucleosome core, higher order chro- matin folding, and transcription in the maintenance of the nuclease-sensitive conformation of the ovalbumin chromatin. Solution hybridization studies demon- strated that the sensitivity of the ovalbumin gene in oviduct nuclei to micrococcal nuclease paralleled the hormone-dependent transcription of the ovalbumin gene in the immature chick. Blot hybridization analysis also revealed a hormone-dependent change in this chromatin region since ovalbumin DNA fragments from nuclease-treated hen and estrogen-stimulated chick oviduct nuclei exhibited nucleosomal repeat pat- terns that were less discrete than those observed for the ovalbumin specific fragments from liver and hor- mone-withdrawn oviducts. This transcription-related conformation was not the result of enhanced sensitivity of the ovalbumin-containing nucleosomal cores since the bulk of the nucleosomes associated with the oval- bumin chromatin were not preferentially cleaved inter- nally by micrococcal nuclease. Rather, an analysis of the fragmentation of the ovalbumin chromatin as a function of digestion extent suggested a mechanism in which the heightened sensitivity resulted from the col- lective expansion of the nuclease cutting sites in the linker regions of the ovalbumin chromatin because the gene was in an unfolded conformation. The transcrip- tion-specific conformation was not merely a conse- quence of RNA synthesis per se since the selective sensitivity of the gene was unaffected by treatment of oviduct nuclei with a-amanitin, actinomycin D, or RNase. In addition, the presence of the transcriptional complex on the ovalbumin chromatin was presumably not required for selective nuclease recognition since preferential cleavage was observed under conditions expected to deplete oviduct nuclei of template-bound RNA polymerase and nascent RNA chains. These re- sults are consistent with a model in which the ex- pressed ovalbumin gene is in an unfolded polynucleo- soma1 structure whose formation is related to tran- scriptional activity but not dependent on the transcrip- tional process.
The chicken oviduct is one model system for the study of the mechanisms by which steroid hormones regulate the
* This work was supported by Grant R01-CA25799-02 from the National Cancer Institute. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$Supported by a National Institutes of Health Predoctoral Traineeship.
transcription of specific genes (1-3). The hormonal regulation of ovalbumin, the major egg white protein, has received con- siderable attention since a variety of steroid hormones induce the synthesis of this protein primarily by stimulating the transcription of the ovalbumin gene in oviduct chromatin. A complete understanding of steroid hormone action in this system, therefore, requires a description of the chromatin environment of the ovalbumin gene during the hormone in- duction process.
Steroid hormones elicit rapid alterations in the organization of chromatin in a variety of vertebrate target cells. These structural changes, which include nuclear swelling and alter- ations in chromatin thermal denaturation patterns, circular dichroism spectrum, and polylysine, actinomycin, and RNA polymerase binding (4-lo), may be a reflection of alterations in the structure of specific chromosomal loci. Cytological analysis of the polytene chromosomes in insect larvae has indeed shown that ecdysteroid hormones induce puffing at a defined subset of chromosomal loci and these specific puffed loci have been associated with the synthesis of specific mRNA and protein products (11-13). Although there is no cytological or electron microscopic evidence relating the structure of the ovalbumin gene in chromatin to its transcriptional activity, nuclease probes can be used to explore certain features of the chromatin conformation of the ovalbumin gene. Previous studies have shown that the ovalbumin gene in hen oviduct nuclei is preferentially attacked by DNase I (14-18), micro- coccal nuclease (19-21), and a nuclease endogenous to oviduct nuclei (22). We have also demonstrated that the transcrip- tional inactivation of the ovalbumin gene in chick oviduct chromatin following the discontinuation of estrogen injection is accompanied by a progressive decrease in the sensitivity of ovalbumin chromatin to micrococcal nuclease (23). In this report, we provide additional evidence for a dynamic relation- ship between the hormone-induced expression of this gene and its chromatin sensitivity to micrococcal nuclease. We also attempted to define the importance of transcription, the tran- scriptional complex, and the nucleosome core in the genera- tion and maintenance of the nuclease-sensitive conformation. Finally, the fragmentation of the ovalbumin chromatin was examined as a function of digestion extent with the hope of providing information on the nature of the packaging of this gene in chromatin. The results of this study are consistent with a model in which the expressed ovalbumin gene is in an unfolded polynucleosomal structure whose formation is re- lated to heightened transcriptional activity but not dependent on the transcriptional process.
EXPERIMENTAL PROCEDURES
Preparation and Digestion of Nuclei-Four-day-old female chicks were injected with 2.5 mg/day of diethylstilbestrol (10 mg/ml of sesame oil) for 14 days (primary stimulation) and then withdrawn from the hormone for 4-5 days. Withdrawn chicks received a single
13018

Ovalbumin Chromatin 13019
injection of diethylstilbestrol (4 mg) or progesterone (4 mg) 12 h before they were killed (secondary stimulation). All laying hens were injected with a mixture of diethylstilbestrol (25 mg) and progesterone (25 mg) 12 h before they were killed. Freshly prepared oviduct, liver, and erythrocyte nuclei (19, 23, 24) were washed in digestion buffer (0.35 M sucrose, 10 nm Tris-HC1 (pH 7.4), 25 mM KC1, 5 nm MgCL, 0.5 mM CaC12, unless otherwise indicated), centrifuged at 10,OOO X g for 5 min, and resuspended in the same buffer at a concentration of 40 units/ml. Digestion with micrococcal nuclease (Worthington) and pancreatic DNase I (Worthington) was carried out at 37 "C as described in the figure legends. Nuclear estrogen receptors were quantified by the [3H]estradiol exchange assay in aliquots of oviduct nuclear suspensions (23, 25).
Preparation and Analysis of Nucleic Acids-For preparation of RNA, portions of the chick oviducts were excised, frozen in liquid nitrogen, and stored at -70 "C. Total RNA was extracted from a 25,000 X g supernatant fraction prepared from oviducts as described by Rhoads et al. (26). Any DNA remaining in the preparation was removed by DNase I digestion (50 pg/ml) in 0.1 M NaAc (pH 5), 5 mM MgC12 for 20 min at 25 "C. The DNase I had been passed over a CAMP-Sepharose column as described by Brison and Chambon (27) to remove small amounts of contaminating RNase; DNase was re- moved by chloroform/isoamyl alcohol (24:l) extraction.
Preparation, fractionation, and electrophoretic analysis of DNA has been described in detail (19, 22, 23). The preparation and char- acterization of ovalbumin and globin mRNAs and their respective ['HIcDNAs has previously been reported (19). [3'PlcDNAs were prepared as described above except 10 pM [3'P]dCTP (500 Ci/mmol for the experiments in Tables I1 and I11 and Figs. 2, 3, 4, 5, and 7 or 3000 Ci/mmol for the experiment in Fig. 6 (right panel)) replaced [3H]dCTP in the reaction mixture. For synthesis of full length oval- bumin cDNA (Fig. 6, left panel), 500 p~ [32P]dCTP (25 Ci/mmol) was employed. Greater than 70% of the reaction products from this synthesis ran as a single band of about 1850 nucleotides on a 1.870 alkaline agarose gel.
Solution hybridizations were performed as described previously (19, 23) and outlined in the figure legends. Transfer of DNA from agarose gels to nitrocellulos,. fdters (Schleicher & Schuell, BA 85) and hybridization of the "P-labeled probes to the immobilized DNA were performed using the procedure of Southern (28) according to condi- tions described elsewhere (22). In the experiments described in Fig. 4, the following revisions of this basic procedure were employed to favor the detection of low molecular weight ovalbumin and globin specific sequences. 1) The gels were not stained prior to transfer; 2) after transfer, the fdters were immediately heated in vacuo at 80 "C for 2- 3 h; and 3) the hybridization temperature, the hybridization time, and the total period of rinses were reduced to 64 "C, 10 h and 4 h, respectively. Transfer of DNA from 7 M urea, 6% polyacrylamide gels to nitrocellulose fdters (Schleicher & Schuell, BA 83) was performed immediately after electrophoresis as described by Thomas (29). Filters were heated at 80 "C, hybridized at 50 "C for 3 days in 0.5 M NaC1,O.l M NaP (pH 7.0), 6 mM EDTA, 1% sodium dodecyl sulfate, and ["PI
TABLE I Effect of steroid hormones on ovalbumin mRNA, nuclear estrogen
receptors, and cell composition in the chick oviduct Immature chicks were injected with hormones as described in the
text and detailed under "Experimental Procedures." RNA prepared from these oviducts (0.2 or 2 mg/ml) or purified ovalbumin mRNA (0.2 pg/ml) was hybridized to [3H]ovalbumin cDNA for 0-19 h. The ROt1/2 values from these data were used to obtain the percentage of ovalbumin mRNA present in each RNA sample. The Rotlr2 value of pure ovalbumin mRNA was 5 X Nuclear estrogen receptors and tubular gland cells were quantified as described previously (23, 25).
Primary Estrogen- ary estro- pro- Second. Second-
withdrawn gen-stim- gester-
(12 hj $,tgy lated (4-5 days) dated one-stim-
Ovalbumin mRNA (% of 0.5 0.003 0.5 0.26
Nuclear estrogen recep- 2300 200 1140 ND" total RNA sequences)
tors (receptor sites/nu- cleus)
% tubular gland cells 77 76 ND 74 ND, not determined.
ESTROGEN ESTROGEN STIMULATED WITHDRAWN
SECONDARY SECONDARV ESTROGEN PROGESTERONE
STIMULATED STIMULATED
.
1 2 3 4 1 2 3 4
DNA SIZE CLASS
1 2 3 4
F I G . 1. Effect of estrogen and progesterone on the distribu- tion of ovalbumin and globin genes in different size DNA fragments. Immature chicks were injected with hormones as de- scribed under "Experimental Procedures." The oviduct nuclei were incubated with micrococcal nuclease (50 units/ml) for 1 min at, 37 "C and the DNA was separated into four molecular weight size classes (19,23). The size classes 1-4, corresponded to DNA fragment lengths of 100-200, 200-800, 800-1200, and >1200 base pairs, respectively. In all groups, t.he size classes 1, 2, 3, and 4 contained 2-3, 2-3, 4-6, and 87-91% of the DNA, respectively. The two larger molecular weight classes of DNA were sonicated and the DNA from all size classes was then subjected to base hydrolysis to remove RNA. DNA (0.2,0.5, and 1 mg/ml) from each size class was annealed to 2 X IO4 cpm/ml of ["HI ovalbumin cDNA for 24 h. A standard curve, constructed by the annealing of nondigested oviduct DNA (0.1-12 mg/ml) to ovalbumin cDNA (2 X lo4 cpm/ml), was used for the determination of the relative concentration of ovalbumin sequences in each fraction as described previously (23). The value 1 on the ordinate corresponds to the concentration of ovalbumin sequences in undigested DNA. The data represent the results from 10 separate experiments with 15-19 oviducts/experiment.
ovalbumin or [32P]globin cDNAs and washed. The dry nitrocellulose filters from all experiments were exposed to Kodak XAR-5 film backed by intensifying screens at -80 "C for the times indicated.
RESULTS
Effect of Steroid Hormones on the Nuclease Sensitivity of Ovalbumin Chromatin in the Chick Oviduct-Table I shows the levels of ovalbumin mRNA, nuclear estrogen receptor, and tubular gland cells in the chick oviducts used in the present studies. In chicks injected daily with estrogen for 14 days (primary estrogen stimulated), 70-80% of the oviduct cell mass was composed of ovalbumin-producing tubular gland cells and about 0.5% of the total cytoplasmic RNA content within each gland cell was ovalbumin mRNA. At 4 days after estrogen administration was discontinued (estrogen-with- drawn), tubular gland cells were retained while the level of ovalbumin mRNA and nuclear estrogen receptor declined 200- and 10-fold, respectively. Administration of estrogen or progesterone to withdrawn chicks for 12 h (secondary stim- ulation) resulted in a 100-200-fold increase in ovalbumin mRNA in pre-existing tubular gland cells. These hormone- dependent changes in ovalbumin mRNA levels, which have been reported previously by a number of laboratories (1-3), have been shown to be accompanied by corresponding changes in the rate of synthesis of ovalbumin-specific RNA sequences (30-34). Fig. 1 shows that these hormone-induced alterations in ovalbumin gene expression are accompanied by changes in the sensitivity of ovalbumin chromatin to micrococcal nu- clease. In these experiments, chick oviduct nuclei were incu-
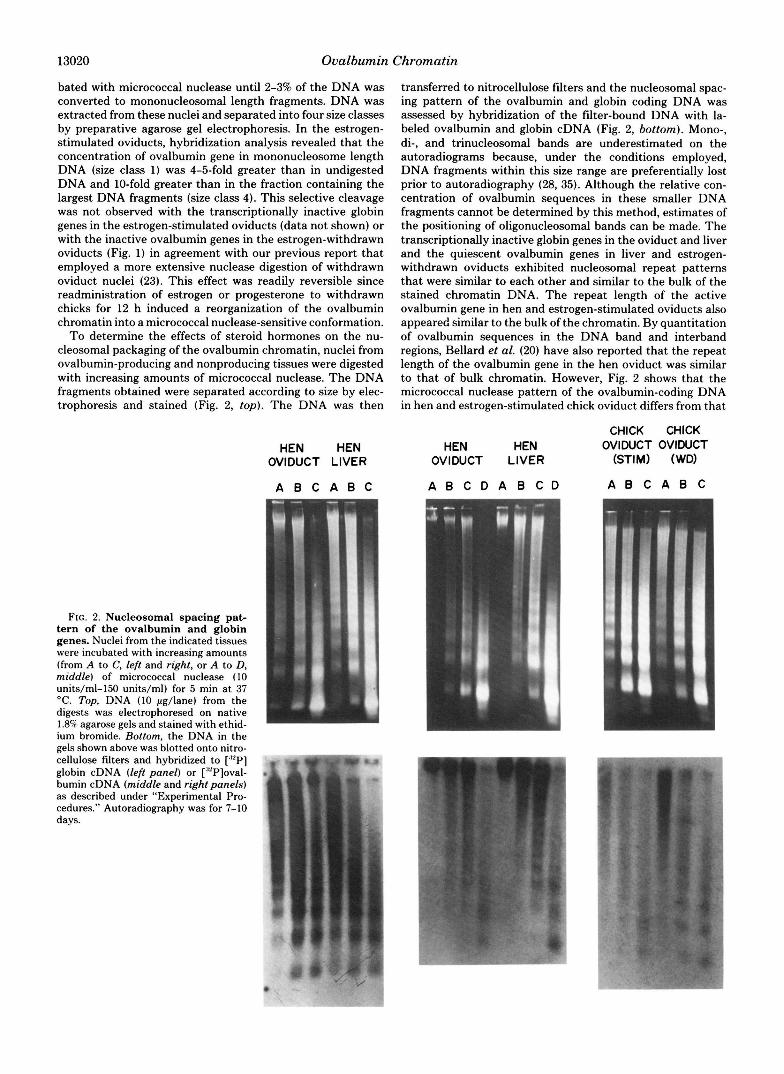
13020 Ovalbumin Chromatin
bated with micrococcal nuclease until 2-3% of the DNA was converted to mononucleosomal length fragments. DNA was extracted from these nuclei and separated into four size classes by preparative agarose gel electrophoresis. In the estrogen- stimulated oviducts, hybridization analysis revealed that the concentration of ovalbumin gene in mononucleosome length DNA (size class 1) was 4-5-fold greater than in undigested DNA and 10-fold greater than in the fraction containing the largest DNA fragments (size class 4). This selective cleavage was not observed with the transcriptionally inactive globin genes in the estrogen-stimulated oviducts (data not shown) or with the inactive ovalbumin genes in the estrogen-withdrawn oviducts (Fig. 1) in agreement with our previous report that employed a more extensive nuclease digestion of withdrawn oviduct nuclei (23). This effect was readily reversible since readministration of estrogen or progesterone to withdrawn chicks for 12 h induced a reorganization of the ovalbumin chromatin into a micrococcal nuclease-sensitive conformation.
To determine the effects of steroid hormones on the nu- cleosomal packaging of the ovalbumin chromatin, nuclei from ovalbumin-producing and nonproducing tissues were digested with increasing amounts of micrococcal nuclease. The DNA fragments obtained were separated according to size by elec- trophoresis and stained (Fig. 2, top). The DNA was then
HEN HEN OVIDUCT LIVER
A B C A B C
FIG. 2. Nucleosomal spacing pat- tern of the ovalbumin and globin genes. Nuclei from the indicated tissues were incubated with increasing amounts (from A to C, left and right, or A to D, middle) of micrococcal nuclease (10 units/ml-150 units/ml) for 5 min at 37 “C. Top, DNA (10 pg/lane) from the digests was electrophoresed on native 1.8% agarose gels and stained with ethid- ium bromide. Bottom, the DNA in the gels shown above was blotted onto nitro- cellulose filters and hybridized to [:97 globin cDNA (left panel) or [:“P]oval- bumin cDNA (middle and right panels) as described under “Experimental Pro- cedures.” Autoradiography was for 7-10 days.
transferred to nitrocellulose filters and the nucleosomal spac- ing pattern of the ovalbumin and globin coding DNA was assessed by hybridization of the filter-bound DNA with la- beled ovalbumin and globin cDNA (Fig. 2, bottom). Mono-, di-, and trinucleosomal bands are underestimated on the autoradiograms because, under the conditions employed, DNA fragments within this size range are preferentially lost prior to autoradiography (28, 35). Although the relative con- centration of ovalbumin sequences in these smaller DNA fragments cannot be determined by this method, estimates of the positioning of oligonucleosomal bands can be made. The transcriptionally inactive globin genes in the oviduct and liver and the quiescent ovalbumin genes in liver and estrogen- withdrawn oviducts exhibited nucleosomal repeat patterns that were similar to each other and similar to the bulk of the stained chromatin DNA. The repeat length of the active ovalbumin gene in hen and estrogen-stimulated oviducts also appeared similar to the bulk of the chromatin. By quantitation of ovalbumin sequences in the DNA band and interband regions, Bellard et al. (20) have also reported that the repeat length of the ovalbumin gene in the hen oviduct was similar to that of bulk chromatin. However, Fig. 2 shows that the micrococcal nuclease pattern of the ovalbumin-coding DNA in hen and estrogen-stimulated chick oviduct differs from that
HEN HEN OVIDUCT LIVER
A B C D A B C D
CHICK CHICK OVIDUCT OVIDUCT
(STIM) (WD)
A B C A B C

Ovalbumin Chromatin 13021
of bulk chromatin and inactive genes in that the hybridization pattern is smeared and the oligonucleosomal bands are less distinct. The blurring of the nuclease digestion patterns, which is especially apparent in the more minimal digestions, has been consistently observed in 8 experiments similar to those shown in Fig. 2.
Transcription and the Nuclease-sensitive Conformation of the Ovalbumin Chromatin-The experiments in Figs. 1 and 2 show that the micrococcal nuclease-sensitive feature of the ovalbumin chromatin is dependent on steroid hormones and suggest that this conformation is dynamically related to the expression of this gene. It is possible, therefore, that selective nuclease recognition of the ovalbumin chromatin is merely a consequence of the active transcription of the ovalbumin gene. This possibility is unlikely, however, since ribonucleotide tri- phosphates, which are required for the synthesis of ovalbumin mRNA sequences in isolated oviduct nuclei (30-34), are not present in the standard digestion buffer used in our studies. In addition, treatment of hen oviduct nuclei with a-amanitin or actinomycin D prior to and during micrococcal nuclease digestion was without effect on the selective cleavage of the ovalbumin chromatin (Table 11, B and C). The concentrations of a-amanitin and actinomycin D used in these studies are greater than those required to completely inhibit the synthesis of ovalbumin mRNA sequences in isolated hen oviduct nuclei (33, 34). Treatment of hen oviduct nuclei with pancreatic RNase prior to nuclease digestion (Table 11, D) was also without significant effect on the selective cleavage of the ovalbumin gene in oviduct chromatin. The selective cleavage of the ovalbumin gene was not observed, however, when deproteinized hen oviduct DNA was used as a substrate (Table 11, E) which shows that the packaging of the ovalbumin gene into oviduct chromatin is required for the preferential nuclease sensitivity.
Although the results discussed above suggest that the nu- clease-sensitive feature of the ovalbumin chromatin is not dependent on RNA synthesis per se, they do not exclude the
TABLE I1 Effect of a-amanitin, actinomycin D, and ribonuclease on the
distribution of ovalbumin gene sequences in different size DNA fragments
Hen oviduct nuclei were incubated at 4 "C for 20 min in digestion buffer (0.35 M sucrose, 25 mM KC1, 100 mM NaCI, 5 mM MgC12, 0.5 m~ CaCb, 10 mM Tris-HC1, pH 7.4) alone (A) or in digestion buffer containing 5 pg/ml of a-amanitin (B), 40 pg/ml of actinomycin D (C), or 100 pg/ml of pancreatic RNase (D). The nuclei were transferred to a 37 "C water bath for an additional 10 min and digested for 5 min by addition of micrococcal nuclease (75 units/ml). Deproteinized oviduct DNA (1 mg/ml buffer) was incubated with micrococcal nuclease (2 units/ml) for 5 min at 37 'C (E). The relatively high levels of cations in this buffer and in the buffers used in the experiments described in Table I11 and Figs. 3 and 4 served to inhibit endogenous nuclease activity (22). The fragmented DNA was separated into four molecular weight size classes and DNA (0.06-8 mg/ml) from each size class and unfractionated nondigested oviduct DNA (U) was annealed to 4 x lo6 cpm/ml of [32P]ovalbumin cDNA for 20 h. The relative concen- tration of ovalbumin sequences in size classes 1-4 were calculated from the Cot1/* values for these hybridizations (19). The value 1.0 for unfractionated DNA (U) corresponded to Cotll2 values that ranged from 2.5-3.2 X 10'. In experiments A-E, about 2% of the total DNA was in the smallest molecular weight size class.
~~
Relative concentration of ovalbumin gene in the following DNA size classes
U 1 2 3 4
Treatment
A. None 1.0 11.4 4.4 1.4 0.5 B. a-Amanitin 1.0 13.0 4.0 1.2 0.4 C . Actinomycin D 1.0 8.8 3.7 1.3 0.5 D. Ribonuclease 1.0 9.0 3.5 1.5 0.5 E. Deproteinization 1.0 1.0 1.0 1.0 1.0
TABLE I11 Effect of ribonucleotide triphosphates on the distribution of ovalbumin gene sequences in different size DNA fragments
Hen oviduct nuclei were incubated at 37 "C for 15 min in buffer alone (2 mM 2-mercaptoethanol, 3 m~ MgCL, 3 mM MnC12, 100 mM KCI, 0.25 M sucrose, 10 mM Tris-HC1, pH 7.8) (A), in buffer containing 0.25 m~ concentration each of ATP, CTP, GTP, and UTP (B), and in buffer containing the four triphosphates and 5 pg/ml of a-amanitin (C). The nuclei were sedimented (5000 X g, 5 min), suspended in digestion buffer containing 50 m~ KCl, and digested for 5 min at 37 "C with micrococcal nuclease (75 units/ml). The relative concen- tration of the ovalbumin sequence in the four size classes were determined as described in the legend to Table 11. The Cot1/2 values for unfractionated DNA (U) ranged from 2.6-3.1 X lo3. In experiments A-C, 2-376 of the total DNA was in the smallest size class. The results are expressed as the means f S.E. of the means from three separate
u 1 2 3 4
A. Buffer 1.0 9.6 5.0 1.7 0.5
B. Buffer + ATP, CTP, 1.0 7.5 4.3 1.5 0.6
C. Buffer + ATP, CTP, 1.0 7.1 4.0 1.4 0.6
kO.l f1.2 f0.7 f0.2 f0.04
GTP, UTP kO.l f0.6 k0.4 fO.l f0.04
GTP, UTP + a-amani- fO.l f0.7 f0.4 kO.2 f0.03 tin
possibility that the configuration depends on a component of the ovalbumin transcriptional apparatus such as RNA polym- erase which remains bound to the chromatin template in the absence of RNA chain elongation (36, 37). Previous studies have shown that the elongation of ovalbumin nascent RNA chains is essentially complete in oviduct nuclei incubated for 5 min at 37 "C in an appropriate transcription buffer contain- ing ribonucleotide triphosphates (33, 34). Since initiation of transcription of the ovalbumin gene does not occur in isolated nuclei (30-34), incubation of oviduct nuclei at 37 "C with the triphosphates for periods longer than 5 min should result in the disappearance of RNA polymerase and nascent RNA chains from the ovalbumin chromatin template. To examine the effects of this treatment on the nuclease sensitivity of the ovalbumin chromatin, hen oviduct nuclei were incubated for 15 min at 37 "C in a transcription buffer alone, in buffer containing the 4 ribonucleotide triphosphates, and in buffer containing the triphosphates and a-amanitin. Following in- cubation, the nuclei were digested with micrococcal nuclease, the fragmented DNA was separated into 4 size classes, and the concentration of the ovalbumin sequence in each size class was determined (Table 111). The ovalbumin chromatin was selectively cleaved by the nuclease in all three groups of nuclei indicating that the nuclease-sensitive feature is retained in nuclei that should lack the template-associated RNA polym- erase and nascent RNA. The experiment presented in Fig. 3 is also consistent with this view. Oviduct nuclei were incubated in the presence and absence of the ribonucleotide triphos- phates for 20 min at 37 "C. The nuclei were then digested with low levels of micrococcal nuclease and the purified DNA was restricted with HindIII. The DNA was electrophoresed on agarose gels, transferred to nitrocellulose filters, and hybrid- ized to ovalbumin and globin cDNAs. With increasing levels of micrococcal nuclease, the DNA fragment containing the ovalbumin sequence was preferentially reduced in both groups of nuclei as compared to the bands containing the globin genes. The ovalbumin gene fragment shown in the figure contains 4.8 kilobase pairs of DNA and corresponds to about 60% of the transcriptional unit of the gene at the 3' end (38).
To visualize the effects of ribonucleotide triphosphate treat- ment on the nucleosomal spacing pattern of the ovalbumin

13022 Ovalbumin Chromatin
OVALBUMIN GLOBIN
1 2 3 4 1 2 3 4
FIG. 3. Effect of ribonucleotide triphosphates on the selec- tive cleavage of the ovalbumin gene. Hen oviduct nuclei were incubated in the absence ( A ) and presence ( B ) of the four ribonucle- otide triphosphates at 37 “C for 20 min, sedimented, and digested with micrococcal nuclease (lanes 1-4, 0. 0.5, 2, and 5 units/ml, respectively) as described in the legend of Table 111. The purified DNA was restricted with HindIII, electrophoresed on 1.1% agarose gels (5 pg/lane), transferred to filters, and hybridized to [:”P]ovalbu- min or [”P]globin cDNAs. The length of the fragments that reacted with the globin cDNA ranged from 5.2 to 8.8 kb, whereas the oval- bumin fragment was 4.8 kb. Only the portions of the autoradiograms that contained bands are presented in the figure. Autoradiography was for 4 days.
chromatin, nuclei were preincubated as described above and digested with nuclease, and the ovalbumin and globin specific sequences in the fragmented DNA were detected by transfer- hybridization analysis (Fig. 4). In this experiment, several revisions of the basic DNA transfer and hybridization proce- dures were made with the hope of favoring the transfer and retention of small DNA fragments on the nitrocellulose filters (see “Experimental Procedures”). The blurring of the oligo- nucleosomal length DNA fragments derived from the oval- bumin but not globin chromatins was apparent in the DNAs derived from control (A) , triphosphate ( B ) , and triphosphate plus a-amanitin (C) pretreated nuclei. The loss of the discrete nucleosomal ladder of the ovalbumin DNA was also evident from other exposures of the autoradiogram shown in the figure (not shown). Inspection of Fig. 4 also reveals that the relative intensities of the DNA spots corresponding to the smaller ovalbumin-containing fragments are enhanced as compared with the globin specific fragments in all three groups in agreement with the solution hybridization data presented in Table 111.
Structure of Ovalbumin Chromatin at the Nucleosome Level-The blurring of ovalbumin-specific oligonucleosomal patterns in the DNA from ovalbumin-producing cells (Fig. 2) is indicative of an altered nucleosomal organization of the transcribed ovalbumin chromatin. These altered oligonucleo- somal patterns, as well as the preferential nuclease digestion of the ovalbumin chromatin (19-21, 23, Fig. l), could result from a loss or selective nuclease cleavage of the nucleosomal cores associated with the ovalbumin gene in chromatin. Al- though previous studies have shown that the ovalbumin DNA in oviduct chromatin is not preferentially destroyed upon extensive micrococcal nuclease digestion (14, 15, 19, 20, 39), they do not necessarily prove that the ovalbumin gene is associated with nucleosomal cores containing 140 base pairs of DNA that are resistant to preferential internal cleavage by micrococcal nuclease. In addition, pancreatic DNase I, in contrast to micrococcal nuclease, selectively destroys ovalbu- min-containing chromatin in the oviduct (14-18), but it is not known if DNase I produces a 10-11 base periodicity in this chromatin DNA as it does in the bulk of the nuclear sequences (40). We have therefore investigated the micrococcal nuclease and DNase I cutting sites introduced at the nucleosomal level in transcribed ovalbumin chromatin. First, nuclei from hen
oviduct and liver were digested with micrococcal nuclease until more than 85% of the DNA was converted to fragments of less than 200 base pairs. The DNA was denatured and separated into monomer length (140-160 bases) and submon- omer length (less than 140 bases) fragments by electrophoresis on alkaline agarose gels. Titration of these DNAs with the ovalbumin cDNA (Fig. 5) revealed that the concentration of the ovalbumin gene in monomer and submonomer length DNA fragments from both oviduct and liver was the same as the concentration of this sequence in nondigested oviduct DNA. To further study the length of the ovalbumin DNA protected from digestion by micrococcal nuclease, DNA frag- ments produced by extensive micrococcal nuclease digestion of nuclei from ovalbumin producing and nonproducing tissues were hybridized to full length ovalbumin cDNA. The resulting hybrids were treated with S1 nuclease, electrophoresed on a polyacrylamide gel, and detected autoradiographically (Fig. 6, left panel). Visual inspection of the autoradiogram reveals that the length of the protected monomeric and submono- meric fragments is the same regardless of whether the oval- bumin gene is in the transcriptionally active (Fig. 6, leftpanel, lanes A , C, and D) or inactive (lanes B and E ) state.
To visualize the fragments of the ovalbumin and globin genes produced by DNase I, hen oviduct nuclei were digested with DNase I and the fragmented DNA was denatured and electrophoresed on urea-polyacrylamide gels. The DNA was then transferred immediately to nitrocellulose filters and hy- bridized to ovalbumin or globin cDNA (Fig. 6, right panel). Estimates of the size of the bands on the autoradiograms reveal that both the ovalbumin (lanes A-C) and globin gene (lanes E-G) fragments occur in the same 10-11 nucleotide repeating pattern as the bulk of the oviduct DNA. Fig. 6 also shows that the ovalbumin (lane D) and globin (lane H) sequences are converted primarily to single fragments of mon- onucleosomal core length by extensive micrococcal nuclease digestion. The results presented in Figs. 5 and 6 suggest, therefore, that the bulk of the active ovalbumin coding DNA is packaged into nucleosomes and that these nucleosomes are
TOTAL DNA GLOBIN OVALBUMIN
A B C A B C A B C
FIG. 4. Effect of ribonucleotide triphosphates on the nucleo- somal spacing pattern of the ovalbumin and globin genes. Hen oviduct nuclei were incubated at 37 “C for 15 min in buffer alone (A) , in buffer containing the four ribonucleotide triphosphates ( B ) , and in buffer containing the triphosphates and a-amanitin ( C ) . The nuclei were digested with micrococcal nuclease (50 units/ml) as described in the legend of Table 111 and the fragmented DNA electrophorsed on 1.8% agarose gels (10 pg/lane). The DNA in the gels was stained (left panel) or transferred to filters and hybridized to [:‘2I’]globin cDNA (middle panel) or [““Plovalbumin cDNA (right panel) as described under “Experimental Procedures.” Autoradiography was for 3 weeks.

Ovalbumin Chromatin 13023 7 0
60
50
2 40 a m > I
* 30
20
FIG. 5. Concentration of the ovalbumin gene in mononucleo- soma1 and submononucleosomal length DNA fragments fol- lowing extensive micrococcal nuclease digestion. Hen oviduct and liver nuclei were incubated with micrococcal nuclease (400 units/ ml, 15 min) until about 85% of the DNA was less than 200 nucleotides in length. The DNA was denatured and separated into mononucleo- some length (140-200 nucleotides) and subnucleosome length (<I40 nucleotides) fragments by preparative electrophoresis on a 1.86 al- kaline agarose gel. Nondigested oviduct DNA (A), mononucleosomal
not preferentially susceptible to internal cleavage by micro- coccal nuclease. In addition, to the extent that the nuclease cutting patterns reflect the configuration of the DNA in the nucleosome, these results indicate that the nucleosomal core configuration along the ovalbumin coding DNA is not dra- matically altered by transcription. This conclusion, of course, as well as others derived from nuclease digestion experiments, assumes that histones are not redistributed along the DNA during the digestion process.
Previous studies have shown that the ovalbumin gene in hen oviduct nuclei is destroyed by micrococcal nuclease at about the same rate as the bulk of the nucleosome covered sequences (14, 15, 19, 20, 39) and the results shown in Figs. 5 and 6 are in complete agreement with these observations. In contrast to these findings, it has recently been reported that the incubation of hen oviduct nuclei with micrococcal nuclease in 10 m~ Tris, pH 7.9,O.l mM CaClp, 1.5% sucrose for up to 90 min at 37 "C resulted in the preferential loss of ovalbumin gene sequences (21). As shown in Fig. 7, however, incubation of hen oviduct nuclei in this buffer for 90 min in the absence of micrococcal nuclease also results in the selective destruction of the ovalbumin gene. Approximately 70% of the ovalbumin sequences were destroyed selectively during this incubation as compared to the bulk of the oviduct sequences and as compared to the globin genes. The preferential loss of hybri- dizable ovalbumin sequences during this prolonged nuclear incubation resulted from endogenous nuclease activity asso- ciated with the oviduct nuclei that produced a 10-11 base periodicity at the terminal stages of chromatin digestion (Ref. 22 and Fig. 7, inset). The breakdown of DNA in the absence of exogenous nucleases should therefore be monitored during
length DNA from oviduct (0) and liver (O), and subnucleosomal length DNA from oviduct and liver (0) was annealed to [:',PI ovalbumin cDNA as described in legend of Table 11. The inset shows samples of the mononucleosomal length (C and D) and subnucleoso- mal length (A and B ) DNA from oviduct (A and C) and liver ( B and D) electrophoresed on a denaturing 7 M urea, 10% polyacrylamide slab gel. The mononucleosomal fraction contains DNA fragments of
Hue I11 restriction fragments of @X174 (not shown). primarily 140-160 nucleotides in length as determined by reference to
nuclear incubations to ensure that the endogenous nuclease is not fragmenting the chromatin.
Kinetic Analysis of the Fragmentation of the Ovalbumin Chromatin by Nuclease-Although previous reports have shown that the transcribed ovalbumin chromatin is selectively cleaved by micrococcal nuclease (19-21), a kinetic study of the preferential fragmentation process had not been performed. Since such an analysis could provide information on the specific chromatin feature recognized by this enzyme, we examined the concentration of the ovalbumin gene in different size DNA fragments at various extents of nuclease digestion (Fig. 8). Hen oviduct nuclei were incubated with micrococcal nuclease until 1,2,3,4, and 5% of the DNA was converted to monomer length fragments. DNA from several of these diges- tions was separated into the four size classes and quantified (Fig. 8, top panel). The concentration of ovalbumin-specific DNA in each size class (middle panel) was determined by solution hybridization analysis and the amount of ovalbumin sequences in each size class (bottom panel) was calculated from the data provided in the top and middle panels of the figure. Enrichment of the ovalbumin gene sequences in the smaller DNA fragments (size classes 1-3) was greatest when the quantity of this DNA was at a minimum. When 1% of the DNA had been converted to monomer length fragments, the concentration of the ovalbumin gene in this size class (size class 1) was 15-20-fold greater than in undigested DNA and 30-50-fold greater than in the fraction containing the largest DNA fragments. About 60-70% of the ovalbumin sequences and 10% of the total DNA were recovered from this minimal digestion in the three smaller size classes. As the amount of DNA in size classes 1-3 increased with increasing digestion

13024 Ovalbumin Chromatin
A B C D E
* I60 1 4 0
OVALBUMIN
A B C D
n-
z 5
- 6 .
GLOBIN
A B C D
4
t
I. FIG. 6. Length of the ovalbumin coding DNA in DNA from
micrococcal nuclease and DNase I-digested nuclei. Left panel, nuclei from hen oviduct ( A ) , hen liver ( B ) , primary estrogen-stimu- lated chick oviducts (C) , estrogen withdrawn oviducts (D), and sec- ondary estrogen-stimulated oviducts ( E ) were incubated with micro- coccal nuclease (450 units/ml; 15 min) until about 90% of the DNA was less than 200 nucleotides in length. The purified DNA (2 mg/ml) was annealed to full length [3*P]ovalbumin cDNA (1 x cpm/ml) for 8 h a t 68 “C and the mixture was treated with S1 nuclease to destroy single-stranded sequences. The samples were then chloro-
form-extracted, ethanol-precipitated, redissolved in 5 mM EDTA, and electrophoresed on a 8% native polyacrylamide gel. Autoradiography was for 3 days. Right panel, oviduct nuclei were incubated for 5 min with 5 pg/ml, 10 pg/ml, and 25 pg/ml of DNase I (lanes A, B, and C ) or for 10 min with 600 units/ml of micrococcal nuclease (lane D). DNA isolated from these nuclei was electrophoresed on 7 M urea, 6 9 polyacrvlamide gels, transferred to nitrocellulose filters, and hybrid- ized to [“2P]ovalbumin or [“‘Plglobin cDNAs as described under “Experimental Procedures.”
A
1 Id 0‘
m u FIG. 7. Selective loss of ovalbumin gene sequences following
prolonged incubation of hen oviduct nuclei. Hen oviduct nuclei were suspended in digestion buffer (10 mM Tris, pH 7.9,O.l rn CaC12, 1.5% sucrose) and immediately deproteinized (0) or incubated for 90 min a t 37 “C before deproteinization (0). DNA prepared from these nuclei was annealed to [”P]ovalbumin (left panel) or [”Plglobin
’<I B
6 3
0 m
4 0
t I
* 30.
1 i i io.
Eq Col
(right panel) cDNAs for quantitation of ovalbumin and globin genes as described in the legend of Table 11. The inset (left panel) shows samples of the DNA before incubation (left) and after incubation (right) electrophoresed on a denaturing 7 M urea, 10% polyacrylamide gel.

Ovalbumin Chromatin 13025
'"1 4
I.
I
i 3
I .
1
3
I
I 2 3 4 fl 1.
1 2 3 4
nm 18
2 3 4
4.
I 2 3 4
m 5.
E- 0 - 18- Ru
FIG. 8. Quantitation of the ovalbumin gene sequences in different sue DNA fragments with increasing micrococcal nuclease digestion. Hen oviduct nuclei were digested with micro- coccal nuclease (5-50 units/&, 1.5-10 min) until about 1, 2, 3, 4, or 5% of the DNA was converted to mononucleosomal length fragments (abscissa). These percentages were estimated by optical density scans of negatives of 1.8% agarose gels. The digested DNA was separated into four size classes, quantified by A2m absorption (top panel) and the relative concentration of the ovalbumin gene sequence in each size class (middle panel) determined as described in the legend of Fig. 1. The percentage of the ovalbumin gene sequence in each size class (bottom panel) is the product of the percentage of the total DNA (top panel) and the relative concentration of the gene (middle panel) in each size class. The data represents the results from 11 separate experiments with 2-4 oviducts/experiment.
(top panel), there was a progressive decrease in the concen- tration of ovalbumin sequences in each of these fractions (middle panel) but the total amount of ovalbumin DNA recovered from each fraction remained relatively constant (bottom panel). When 5% of the DNA was converted to mononucleosomal length fragments, this DNA was enriched only 4-5-fold in the ovalbumin sequence. In this digestion, the three smaller size classes stiU contained 60-70% of the oval- bumin sequence but 4-5 times the amount of DNA as com- pared to the minimal 1% digestion. These results suggest that the preferential cleavage of the ovalbumin gene occurred very
early in the digestion whereas the nuclease cuts produced a t later times were relatively nonselective with regard to the transcribed ovalbumin sequence.
DISCUSSION
Central to the understanding of eukaryotic gene regulation is the question of whether chromatin structural modifications are involved in transcriptional control or result from the transcription machinery. I t is clear that a chromosomal con- formation recognized during extensive digestion of oviduct nuclei with pancreatic DNase I is not a consequence of the transcription process since this conformation is independent of transcriptional rate (17,18,41), encompasses the ovalbumin gene and flanking inactive sequences (17, 18), and is retained by ovalbumin chromatin after estrogen administration has been discontinued (42). In contrast, our previous studies have shown that the feature of the ovalbumin coding chromatin recognized by micrococcal nuclease is gradually lost during the inactivation of the gene by estrogen withdrawal (23) . The results in Fig. 1 c o n f i i this finding and, in addition, show that ovalbumin chromatin is reorganized into a sensitive con- formation within 12 h after administration of estrogen or progesterone. The observation that the rapidly transcribed ovalbumin gene in hen oviduct nuclei is cleaved selectively by this enzyme whereas transcriptionally inert flanking se- quences and the modestly transcribed X and Y genes are not,' provides additional support for the proposal that the feature of chromatin recognized by micrococcal nuclease is directly related to transcriptional activity. The enhanced nuclease sensitivity, however, is not merely secondary to transcription at the ovalbumin locus since it was observed in the absence of ribonucleotide triphosphates and in the presence of actino- mycin D and a-amanitin (Table 11). In addition, the presence of the transcription complex on the ovalbumin chromatin is presumably not required for selective nuclease recognition since preferential cleavage, as revealed by three different criteria, was observed under conditions predicted to deplete oviduct nuclei of template-bound RNA polymerase and nas- cent RNA chains (Table I11 and Figs. 3 and 4). It must be pointed out, however, that this interpretation assumes that:l) complete elongation of ovalbumin RNAs occurred in the presence of the triphosphates in the experiments described in Table I11 and Figs. 3 and 4; 2) the RNA polymerases and ovalbumin mRNA precursor dissociated from the template after termination of transcription; and 3) initiation of tran- scription of the ovalbumin gene did not occur in the isolated nuclei. Although we feel these assumptions are correct be- cause of previous studies by others (30-34, 43), we did not experimentally test their validity in this report. With these qualifications in mind, the observations described above pro- vide evidence that the acquisition of the micrococcal nuclease- sensitive feature on the ovalbumin chromatin may play a role in regulating the transcriptional activation of the ovalbumin gene.
Analysis of the nucleosomal spacing patterns of specific genes in the DNA from micrococcal nuclease-digested chro- matin has revealed an inverse relationship between distinct oligonucleosomal length fragments and transcriptional activ- ity. Inactive genes exhibit DNA repeat patterns which are at least as discrete as those of the bulk of the chromatin DNA (35, 44, 45). Similarly, discrete nuclease digestion patterns have been reported for genes that are transcribed at relatively low rates. These include the active thymidine kinase genes in cultured mouse cells (46), genes coding for poly(A) RNA in
' J. N. Anderson, J. N. Vanderbilt, G. M. Lawson, M.-J. Tsai, and B. W. O'Malley, manuscript submitted for publication.

13026 Ovalbumin Chromatin
mouse liver (47), and the copia and heat shock genes in Drosophila in the absence of heat stimulation (35,44,45). On the other hand, there is a progressive loss in the resolution of the oligonucleosomal bands of the heat shock genes upon heightened transcriptional activity following heat treatment and a complete disruption of the ladder has been reported to occur in the maximally induced state (35,44,45). The results reported in Figs. 2 and 4 are also indicative of some alteration in the nucleosomal packaging of the ovalbumin chromatin in ovalbumin-producing cells since the smearing of oligonucleo- somal patterns derived from the ovalbumin chromatin was greatest when the gene was in the active state. It is unlikely that this change in the oviduct system resulted from a loss of ovalbumin-containing nucleosomes upon transcriptional acti- vation since the ovalbumin gene is packaged into nucleosomes regardless of its transcriptional status (14, 15, 39; Figs. 5 and 6). Because the ovalbumin DNA within the nucleosomal core is apparently not selectively attacked by micrococcal nuclease (Figs. 5 and 6), the blurring of ovalbumin-containing oligo- nucleosomal bands, as well as the preferential release of small ovalbumin gene fragments (19-21, Figs. 1 and 8) , probably results from an expansion of the nuclease cutting sites in the linker regions of the ovalbumin chromatin upon transcrip- tional activation.
The analysis of the sequential nuclease digestion of the ovalbumin chromatin in Fig. 8 shows that the quantity of ovalbumin sequences in different size DNA fragments re- mained relatively constant throughout the course of the diges- tion even though the amount of DNA in the three smaller size classes increased 4-5-fold. Thus, it is unlikely that each linker region of the ovalbumin chromatin is intrinsically mod- ified in such a way as to enhance its nuclease sensitivity since the ovalbumin-containing oligo- and polynucleosomes pro- duced early in the digestion are not subsequently processed to mononucleosomal length fragments in a selective manner. It is conceivable that only a small fraction of the nucleosomal linker regions of ovalbumin chromatin are in an altered nu- clease-sensitive conformation. Accordingly, the cleavage of these specific sites early in the digestion could produce a defined distribution of ovalbumin fragments that would re- main stable during subsequent digestion. Although this scen- ario cannot be discarded at this time, hypersensitive nuclease cutting sites have not been detected in the chromatin coding regions of a variety of expressed genes (48-51). Similarly, we have been unable to detect specific micrococcal nuclease cutting sites in the ovalbumin coding chromatin by the map- ping procedure of Wu (48) (data not shown). However, a limited number of hypersensitive nuclease sites distributed randomly with respect to the DNA coding sequence would not have been detected by this procedure. Perhaps a more attractive model to explain the data in Fig. 8 is that all of the linker regions of the intact ovalbumin chromatin are collec- tively sensitive to micrococcal nuclease because the gene is in an extended conformation as has been observed by micro- scopic analysis of active chromatin in a variety of systems (11-13, 52). A limited number of selective nuclease cleavages within an extended conformation might be expected to disrupt the higher order folding thereby producing oligo- and poly- nucleosomes which would no longer be a target for preferential nuclease digestion.
The sequential digestion of chromatin containing the estro- gen receptor in rat uterine nuclei (53) is similar to that shown in Fig. 8 for the ovalbumin chromatin in the oviduct in that the receptor-containing nucleosomes are preferentially excised during very early digestion and the selectivity of the digestion is lost upon more extensive nuclease treatment. The steroid receptor protein, therefore, may also be associated with ex-
tended regions of the chromatin. The observation that the ecdysone-receptor complex is associated with ecdysone-in- duced puffs in Drosophila salivary gland chromosomes pro- vides direct evidence for this view (54). The steroid receptors may therefore represent a structural type of regulatory protein whose function is to unfold specific chromatin regions. The observation that the receptor-progesterone complex binds to multiple sites on the ovalbumin and other egg white protein genes is consistent with such a structural role (55). However, a relationship between the binding of the receptor-steroid complex to specific chromatin or DNA sites and the hormone- dependent changes in the ovalbumin chromatin reported in this study remains to be established.
Acknowledgments-We thank Bonnie Germain for her skillful technical assistance and the anonymous referee for critical comments and helpful suggestions during the preparation of this manuscript.
REFERENCES 1. Palmiter, R. D. (1975) Cell 4, 189-197 2. Schimke, R. T., McKnight, G. S., Shapiro, D. J., Sullivan, D., and
Palacios, R. (1975) Recent Prog. Horm. Res. 31, 175-211 3. O’Malley, B. W . , Roop, D. R., Lai, E. C., Nordstrom, J. L.,
Catterall, J. F., Swaneck, G. E., Colbert, D. A,, Tsai, M.-J., Dugaiczyk, A,, and Wood, S. L. C. (1979) Recent Prog. Horm. Res. 35, 1-46
4. Mainwaring, W . I. P., and Jones, D. M. (1975) J . Steroid Biochem. 6,475-481
5. Schwartz, R. J., Tsai, M.J . , Tsai, S. Y., and O’Malley, B. W . (1975) J. Biol. Chem. 250, 5175-5182
6. Tsai, S. Y., Tsai, M.-J., Schwartz, R. J., Kalimi, M., Clark, J. H., and O’Malley, B. W. (1975) Proc. Natl. Acad. Sci. U. S. A. 72, 4228-4232
7. Kalimi, M., Tsai, S . Y., Tsai, M.-J., Clark, J. H., and O’Malley, B. W . (1976) J. Biol. Chem. 251, 516-523
8. Loor, R. M., Hu, A.-L., and Wang, T. Y. (1977) Biochim. Biophys. Acta 477, 312-321
9. Snow, L. D., Eriksson, H., Hardm, J. W . , Chan, L., Jackson, R. L., Clark, J. H., and Means, A. R. (1978) J. Steroid Biochem. 9, 1017-1026
10. Clark, J. H., and Peck, E. J. Jr. (1979) Monogr. Endocrinol. 14,
11. Clever, U. (1966) Am. 2001. 6,33-41 12. Ashburner, M., Chihara, C., Meltzer, P., and Richards, G. (1973)
13. Ashburner, M., and Bonner, J. J. (1979) Cell 17, 241-254 14. Garel, A., and Axel, R. (1976) Proc. Natl. Acad. Sci. U. S. A. 73,
15. Garel, A., and Axel, R. (1978) Cold Spring Harbor Symp. Quant. Biol. 42, 701-708
16. Palmiter, R. D., Mulvihill, E. R., McKnight, G. S., and Senear, A. W . (1978) Cold Spring Harbor Symp. Quant. Biol. 42,639-648
17. Lawson, G. M., Tsai, M.-J., and O’Malley, B. W . (1980) Biochem- istry 19,4403-4411
18. Lawson, G. M., Knoll, B. J., March, C. J., Woo, S. L. C., Tsai, M- J., and O’Malley, B. W . (1982) J. Biol. Chem. 257, 1501-1507
19. Bloom, K. S., and Anderson, J. N. (1978) Cell 15, 141-150 20. Bellard, M., Gannon, F., and Chambon, P. (1978) Cold Spring
21. Senear, A. W . , and Palmiter, R. D. (1981) J. Biol. Chern. 256,
22. Vanderbilt, J. N., Bloom, K. S., and Anderson, J. N. (1982) J.
23. Bloom, K. S., and Anderson, J. N. (1979) J. Biol. Chem. 254,
24. Bloom, K. S., and Anderson, J. N. (1978) J. Biol. Chem. 253,
25. Anderson, J. N., Clark, J. H., and Peck, E. J. (1972) Biochem. J .
70-97
Cold Spring Harbor Symp. Quant. Biol. 38,655-662
3966-3970
Harbor Symp. Quant. Biol. 42, 779-791
1191-1198
Biol. Chem. 257, 13009-13017
10532-10539
4446-4450
126, 561-567 26. Rhoads. R. E.. McKnight, G. S., and Schimke, R. T. (1973) J.
Biol. Chem. 248,2031-2039 27. Brison, 0.. and Chambon, P. (1976) Anal. Biochem. 75,402-409 28. Southern, E. M. (1975) J . Mol. Biol. 98, 503-517 29. Thomas, P. S. (1980) Proc. Natl. Acad. Sci. U. S. A . 77, 5201-
5205

Ovalbumin Chromatin 13027
30. Roop, D. R., Nordstrom, J . L., Tsai, S. Y., Tsai, M.-J., and O’Malley, B. W. (1978) Cell 15,671-685
31. Schiitz, G., Nguyen-Huu, M. C., Giesecke, K., Hynes, N. E., Groner, B., Wurtz, T., and Sippel, A. E. (1978) Cold Spring Harbor Symp. Quant. Biol. 42,617-625
32. McKnight, G. S., and Palmiter, R. D. (1979) J. Biol. Chem. 254, 9050-9058
33. Swaneck, G. E., Nordstrom, J. L., Kreuzaler, F., Tsai, M.-J., and O’Malley, B. W. (1979) Proc. Natl. Acad. Sci. U. S. A . 76, 1049-1053
34. Swaneck, G. E., Kreuzaler, F., Tsai, M.-J., and O’Malley, B. W. (1979) Biochem. Biophys. Res. Commun. 88, 1412-1418
35. Wu, C., Wong, Y.-C., and Elgin, S. C. R. (1979) Cell 16, 807-814 36. Tsai, M.-J., and Saunders, G. F. (1973) Proc. Natl. Acad. Sci. U.
37. Sass, H. (1982) Cell 28, 269-278 38. Woo, S. L. C., Beattie, W. G., Catterall, J . F., Dugaiczyk, A,,
Staden, R., Brownlee, G. G., and O’Malley, B. W. (1981) Bio- chemistry 20, 6437-6446
39. Lacy, E., and Axel, R. (1975) Proc. Natl. Acad. Sci. U. S. A. 72, 3978-3982
40. Noll, M. (1978) Cold Spring Harbor Symp. Quant. Biol. 42, 77-
S. A. 70,2072-2076
87
S. A . 74,4867-4871
D., (1980) J. Cell Biol. 87, 142-151
41. Garel, Z., Zolan, M., and Axel, R. (1977) Proc. Natl. Acad. Scz. U.
42. Shepherd, J. H., Mulvihill, E. R., Thomas, P. S., and Palmiter, R.
43. Yang, V. W., and Flint, S. J. (1979) J. Virol. 32, 394-403 44. Levy, A., and Noll, M. (1981) Nature 289, 198-203 45. Levinger, L., and Varshavsky, A. (1982) Cell 28, 375-385 46. Camerini-Otero, R. D., and Zasloff, M. A. (1980) Proc. Natl.
Acad. Sci. U. S. A . 77,5079-5083 47. Gottesfeld, J. M., and Melton, D. A. (1978) Nature 273, 317-319 48. Wu, C. (1980) Nature 286, 854-860 49. Storb, U., Arp, B., and Wilson, R. (1981) Nature 294,90-92 50. Weintraub, H., Larsen, A., and Groudine, M. (1981) Cell 24,333-
51. Elgin, S. C. R. (1981) Cell 27, 413-415 52. Pruitt, S. C., and Grainer, R. M. (1981) Cell 23, 711-720 53. Senior, M. B., and Frankel, F. R. (1978) Cell 14,857-863 54. Gronemeyer, H., and Pongs, 0. (1980) Proc. Natl. Acad. Sci. U.
55. Mulvihill, E. R., LePennec, J-P., and Chambon, P. (1982) Cell 24,
344
S. A . 77,2108-2112
621-632





























