Embed Size (px)
Citation preview

Journal of Affective Disorders 133 (2011) 120–127
Contents lists available at ScienceDirect
Journal of Affective Disorders
j ourna l homepage: www.e lsev ie r.com/ locate / j ad
Research report
The insular cortex and the neuroanatomy of major depression
Reiner Sprengelmeyer a,⁎, J. Douglas Steele b, Benson Mwangi b, Poornima Kumar c,David Christmas b, Maarten Milders d, Keith Matthews b
a University of St Andrews, School of Psychology, St Andrews, UKb University of Dundee, Division of Medical Sciences, Dundee, UKc University of Oxford, Department of Psychiatry, Oxford, UKd University of Aberdeen, School of Psychology, Aberdeen, UK
a r t i c l e i n f o
⁎ Corresponding author at: University of St AndrewsSt Mary's Quadrangle, St Andrews KY16 9JP, UK.
E-mail address: [email protected] (R. Sprengelme
0165-0327/$ – see front matter © 2011 Elsevier B.V.doi:10.1016/j.jad.2011.04.004
a b s t r a c t
Article history:Received 20 February 2011Received in revised form 7 April 2011Accepted 7 April 2011Available online 29 April 2011
Background: The neuroanatomical substrate underlying Major Depressive Disorder (MDD) isincompletely understood. Recent reports have implicated the insular cortex.
Methods: Two cohorts of participants with MDD were tested. In the first MDD cohort, we usedstandardised facial expression recognition tasks. In the second cohort, we focused on facialdisgust recognition, a function associated with the insular cortex. T1 weighted MR imaging wasused in the second cohort to test the hypothesis of abnormal insular volume being associatedwith impaired disgust recognition.
Results: Disgust recognition was particularly impaired in both cohorts. In the second cohort, themagnitude of the disgust recognition deficit correlated with reduced insula grey matter volume.Exploring the idea of insula involvement in MDD further, we identified the insular cortex and theanterior cingulate cortex as key neural correlates of core symptoms, in that scores of 3 clinical scales(the Beck Depression Inventory, the Hamilton Depression Rating Scale, and the Snaith–HamiltonPleasure Scale) correlated with grey matter volume in these structures.
Limitations: MDD participants were clinically representative of specialist and academicpsychiatric practice in the UK and presented with robust primary diagnoses; we did notexclude common co-morbidities such as anxiety and personality disorders.
Conclusions:Wepropose that cognitive and emotional functions assumed to be associated withthe insula are adversely affected in patients with MDD and that this may, therefore, representthe substrate for some core clinical features of MDD. Further exploration of the involvement ofthe insular cortex in MDD is warranted.
© 2011 Elsevier B.V. All rights reserved.
Keywords:MDDMajor Depressive DisorderDisgustInsular cortexVBM
1. Introduction
Major Depressive Disorder (MDD) can be conceptualised as aneuropsychological syndrome, in which the clinical picture ofMDD is considered as a cluster of symptoms associated withdisordered processing within a complex system of interlinked
, School of Psychology
yer).
All rights reserved.
,
neural circuits (Drevets et al., 2008; Mayberg, 2009). Dysfunc-tion within these circuits is thought to be associated with theavailability of specific neurotransmitters, neurotrophic or neu-roprotective substances and/or with structural changes to theneural substrate itself.
With the aim to identify candidate neural structuresforming part of the circuitry underlying MDD, we used anestablished structure–function relationship as a behaviouralmarker to infer the possible involvement of specific neuralsubstrates. First, we explored the capacity of a cohort ofparticipants with MDD to recognise facial expressions ofemotions, a) because this function is associated with a variety

121R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
of mainly limbic and paralimbic structures which are thoughtto be involved in MDD, b) because standardised tests areavailable to assess the integrity of emotion recognition, and c)because there is a reasonable consensus about the neuralsubstrate associated with recognition of at least some facialexpressions of emotion. Fear recognition is thought to beassociated with the amygdala (Adolphs et al., 1994) anddisgust recognition has been linked to the insular cortex(Hennenlotter et al., 2004; Kipps et al., 2007; Phillips et al.,1997; Sprengelmeyer et al., 1998;Wicker et al., 2003). Deficitsin disgust or fear recognition would therefore be indicative forthe possible involvement of these neural structures in MDD. Tominimise the probability of following up chance findings, wedecided a priori, to consider unpredicted results only asmeaningful if the same result showedup consistently in differenttest procedures or diagnostic tools assessing the same cognitiveor emotional function (Young et al., 1993).
We performed 2 studies — in the first study, we foundrecognition of facial expressions of disgust particularly impairedin participants withMDD, while recognition of fear did not differbetween groups. This finding suggested a probable dysfunctionof the insular cortex and not a substantial impairment ofamygdala function. In order to test the hypothesis of possibleinsula impairment in MDD, we attempted to replicate andextend our findings by looking at disgust recognition in adifferent group of participants with MDD and a different controlgroup. On this occasion, high resolutionMRI scanswere obtainedfrom both groups to allow estimates of possible grey mattervolume reduction in MDD and correlation statistics of perfor-mance on the facial expression recognition task with voxel-based volumetric measures. In addition, to generate furthertestable hypotheses about neural substrates associated with thecore symptoms of MDD, we correlated several standard clinicalmeasures of illness severitywith neuroimagingmeasures of greymatter volume.
2. Methods
Seventy-eight participants were recruited in total; 27participants withMDD, 51 controls. All testing was conductedin accordance with the Declaration of Helsinki — 2008, andrequired ethical approvals obtained from the relevant NHSresearch ethics committees. Study participants providedwritten consent.
In both studies, we have recruited clinically-referred partic-ipants with MDD. As a consequence, we have not only testedparticipantswhowere not only taking prescribed antidepressantmedication, but who had histories of exposure to a range ofpsychotropicmedications.We therefore cannot exclude an effectof previous or current medication to either the behavioural orstructural findings in the MDD participants. Similarly, althoughthe MDD participants were clinically representative of specialistand psychiatric practice in the UK and presented with a robustprimary diagnosis, we did not exclude common co-morbiditiessuch as anxiety and personality disorders.
2.1. Study 1
2.1.1. ParticipantsTen clinically referred participants with chronic (N2 years)
MDD (3 male, 7 female) and 30 control participants without
histories of MDD (9 male, 21 female) were tested. All MDDparticipants were assessed by an experienced psychiatrist andmet criteria for a diagnosis of DSM-IV MDE. Mean age ofparticipants with MDD was 49.7 (SD 9.2) years and their IQ asmeasured with the NART — National Adult Reading Test(Nelson, 1982) was 122.6 (SD 5.3). On the BDI-II — BeckDepression Inventory (Beck et al., 1996) and the (17 item)HDRS17 — Hamilton Depression Rating Scale (Hamilton, 1960)the group's mean scores were 34.9 (SD 8.8) and 22.5 (SD 5.0)respectively. All MDD participants were receiving antidepres-sant medication. Medication as total dose per day was:Phenelzine (60 mg), Sertraline (200 mg), Isocarboxazid(10 mg), Venlafaxine (75–375 mg), Trimipramine (150 mg),Amitriptyline (350 mg), Citalopram (20 mg), Tranylcypromine(30 mg). Additional combination/augmentation strategies were:Lithium carbonate (600–800 mg), Quetiapine (400–750 mg),Buproprion (300 mg), Mirtazapine (45 mg), Valproate Sodium(500 mg), L-Tryptophan (2–4 g), and Chlorpromazine (75 mg).Further, a total of 3 participants were taking benzodiazepines, ata Diazepam dose-equivalent of 10–30 mg/day. Two participantswere taking hypnotics (Zolpidem and Zopiclone). Mean age ofthe control participants was 49.9 (14.1) years, and their IQ was121.3 (SD 3.5). Student t-tests showed no significant differencesbetween groups for age (t=0.10, p=.92) and estimated pre-morbid intelligence (t=−0.92, p=.36).
2.1.2. Neuropsychological methodsParticipant's consent was obtained according to the
Declaration of Helsinki, and all testing was conducted withResearch Ethics Committee approval. Two separate teststaken from the Facial Expressions of Emotion: Stimuli and Tests(FEEST) were used to assess the ability to recognise facialexpressions of basic emotions (Young et al., 2002).
The Ekman 60 Faces test contained photographs of thefaces of 10 people from the Ekman and Friesen series (Ekmanand Friesen, 1976). For each face, there were poses corre-sponding to each of six basic emotions (happiness, surprise,fear, sadness, disgust, and anger), giving a total of 60photographs (10 for each emotion). These were presentedin random order. The maximum score is 10 for each of the 6emotion categories.
The Emotion Hexagon test used photographic-qualitycontinua of morphed images of an individual's face from theEkman and Friesen series, which were prepared by blendingbetween prototype expressions. The test set consisted of 30stimuli, comprising 5 morphed images for each of 6 emotioncontinua: happiness–surprise, surprise–fear, fear–sadness,sadness–disgust, disgust–anger, and anger–happiness, inproportions 90/10 (e.g., 90% fear+10% surprise), 70/30(70% fear+30% surprise), 50/50 (50% fear+50% surprise),30/70 (30% fear+70% surprise), and 10/90 (10% fear+70%surprise). The test involved a practice block of 30 trials,followed by 5 test blocks. In each block of trials the 30 imageswere presented once each, in random order. For the purposesof scoring, the responses to the 50/50 morphed images and tothe practice block trials were excluded, leaving a maximumscore of 20 for each of the 6 emotion categories (the 4 imagesmost similar to each specific emotional expression, repeatedacross 5 blocks of test trials). In both tests, faces werepresented one at a time for 5 s each, and the participant wasasked to decide which of the emotion names (happiness,

122 R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
surprise, fear, sadness, disgust, and anger) best described thefacial expression shown. The names of these six emotionsremained visible on the computer screen throughout the test,with the order in which the emotion names shown on thescreen randomised each time the test was given. Full detailsof the procedure for each test can be found in the manualaccompanying the FEEST.
2.2. Study 2
2.2.1. ParticipantsSeventeen participants with MDD (8 male, 9 female) and
21 control participants without histories of MDD (9 male, 12female) who were also free of psychiatric and neurologicdisorder took part in this study. All MDD participants wereassessed by an experienced psychiatrist and met criteria for adiagnosis of DSM-IV MDE. The mean age of participants withMDD was 45.6 (SD 12.3) years and their IQ as measured withthe NART — National Adult Reading Test (Nelson, 1982) was111.6 (SD 8.4). On the BDI-II — Beck Depression Inventory(Beck et al., 1996) and the HDRS17 — Hamilton DepressionRating Scale (Hamilton, 1960), the MDD group's mean scoreswere 22.9 (SD 8.2) and 23.2 (SD 4.3) respectively. On theSHAPS— Snaith–Hamilton Pleasure Scale (Snaith et al., 1995),where a low score indicates anhedonia, the group's meanscore was 35.0 (SD 6.7). All MDD participants were receivingantidepressant medication. Medication as total dose per daywas: Imipramine 200 mg, Phenelzine 45–90 mg, Trazodone300 mg, Mirtazapine 30–60 mg, Venlafaxine 150–225 mg,Amitryptyline 200 mg, Escitalopram 15 mg, Citalopram20 mg, Fluoxetine 40 mg, Sertraline 25–150 mg, and Lithium600–800 mg. Mean age of the control participants was 42.0(12.9) years and their estimated IQ was 113.9 (SD 8.2). The BDI-II score of control participants was 3.1 (SD 2.9) and the SHAPSscore was 51.7 (SD 4.3). Student t-tests showed no significantdifferences between groups with respect to age (t=0.97,p=.34) or estimated pre-morbid intelligence (t=−0.92,p=.37). As expected, the BDI-II scores were significantly higherin the MDD group compared to controls (t=8.86, pb .001), andthe SHAPS scores significantly lower in the MDD groupcompared to controls (t=8.23, pb .001).
2.2.2. Neuropsychological methodsA complementary facial recognition task was used with
computer-manipulated photographs of the faces of 4 differ-ent people taken from the FEEST (Young et al., 2002). For thistest there were neutral expressions and poses correspondingto the basic emotions of happiness, fear, sadness, disgust, andanger at 4 levels of intensity (20%, 40%, 60% and 80%). Eachpicture displaying an emotional expression was shown 4times on a computer screen, resulting in a maximum score of16 for each basic emotion. In addition, 20 pictures displayinga neutral facial expression were shown. This experimentaldesign was developed to allow calculation of signal detectionmeasures (results are reported in detail elsewhere). Thedependent variable for this experimental task was number ofbehavioural errors. The presentation time for each facialexpression was 500 ms and thereafter, participants had todecide which of the labels (happiness, fear, sadness, disgust,anger, or neutral) best described the facial expression shownby pressing one of six pre-specified keys. Following Corwin
(Corwin, 1994), the ‘discrimination accuracy’ (dL) for disgustwas calculated as
dL = lnHR � 1−FARð Þ1−HRð Þ FAR
� �
where the ‘False Alarm Rate’, FAR=(Total no. of false alarms+0.5)/(Total no. of distracters+1) and the ‘Hit Rate’, HR=(Totalnumber of hits+0.5)/(Total number of targets+1). Higher dLvalues indicate a better ability to discriminate targets (facialexpressions of disgust) from distracters (other facial expressionsincluding no emotional expression). For each subject thisresulted in five dL values for each distracter emotion and neutralfacial expression.
2.2.3. MR imaging data acquisitionA 1.5T GE Medical Systems Signa scanner was used with
T1 acquisition parameters: TR 20 ms, TE 6 ms, flip angle 35°,124 contiguous 1.6 mm axial slices of 256×256 voxels withan in-plane resolution of 0.938 mm2.
2.2.4. MR imaging data processing and voxel basedmorphometry (VBM)
T1 weighted MR images were visually inspected forartefacts and then segmented into grey matter maps usingstatistical parametric mapping (SPM5) (Friston et al., 2007).This process comprises normalisation to Montreal Neurolog-ical Institute (MNI) stereotactic space with modulation tocontrol rescaling effects. The segmented grey matter imageswere then smoothed with an 8 mm full-width half maximumGaussian kernel. A random effects design was implementedwith a two group t-test being used to test the null hypothesisof no difference in grey matter density between the twogroups. A linear regression was used to test the nullhypothesis of no increase in grey matter volume with betterperformance on the disgust recognition task for only theMDDgroup. Results from the clinical rating scales (BDI-II, HDRS17,SHAPS) were treated in the same way. The threshold ofsignificance was defined as pb .05 corrected at a whole braincluster level using a Monte Carlo Method (Ross and Slotnick,2008; Walther et al., 2009). Specifically, this calculation wasdone using 10,000 iterations modelling each voxel as anormally distributed random number (mean of zero, unitvariance), with type I error assumed equal to the individualvoxel significance value in a volume defined by the T1 imagedimensions, with spatial correlation simulated by smoothingwith an 8 mm full-width half-maximum Gaussian kernel. Theprobability of observing successively larger cluster sizes wascalculated based on the Monte Carlo cluster size distribution,and a cluster extent threshold of 144 voxels identified, suchthat the probability of observing this or larger clusters waspb .05 across the whole brain (Ross and Slotnick, 2008;Walther et al., 2009).
3. Results
3.1. Study 1
For the Ekman 60 Faces test a repeated measures ANOVAwith emotion (happiness, surprise, fear, sadness, disgust, andanger) as a within-subject factor, and group (MDD vs. control

123R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
participants) as a between-subjects factor showed a signifi-cant effect of emotion (F(5,190)=16.07, pb .001). No signifi-cant group effect was seen (F(1,38)=2.19, p=.15), but therewas a significant emotion×group interaction (F(5,190)=3.06,pb .05). To explore the interaction, we performed t-tests,which showed no significant differences for all emotions(t≥−1.03 ≤0.49, p≥ .31) except disgust (t=3.61, pb .01).Results are given in Fig. 1A.
For the Emotion Hexagon test a repeated measures ANOVAshowed a significant effect of emotion (F(5,190)=6.03, pb .001),a significant group effect (F(1,38)=6.46, pb .05), and asignificant emotion×group interaction (F(5,190)=2.37,pb .05). To explore the interaction, we performed t-tests,which showed no significant differences for happiness,surprise and fear (t≥0.09≤0.40, p≥ .31). Recognition ofsadness (t=2.60, pb .05), disgust (t=2.29, pb .05) and anger(t=2.76, pb .01), however, was impaired in MDD. Results aregiven in Fig. 1B.
3.2. Study 2
3.2.1. Behavioural resultsTo explore a hypothesised abnormality of disgust dis-
crimination accuracy further, in light of the results from study1, the five disgust dL values were averaged for each
Fig. 1. A) Performance of patients with MDD and control participants on theEkman 60 Faces test, and B) on the Emotion Hexagon test. The consistentdeficit in disgust recognition across tests in participants with MDD impliesinvolvement of the insular cortex.
Fig. 2. A) Decreased regional grey matter volume in participants with MDDcompared with healthy control participants. B) Correlation of grey mattervolume reduction with discrimination accuracy for disgust (dL) in partici-pants with MDD. Abbreviations: A–H=amygdala–hippocampal complex.
participant (MDD 2.4 SD 1.1, controls 3.4 SD 1.2) and a twogroup t-test used to test the null hypothesis of no difference.For the MDD group, the average discrimination accuracy dL
score for disgust was significantly reduced (t=−2.26,p=.03). Averaged dL scores for all other basic emotionswere not significant. These results will be reported in detailelsewhere.
3.2.2. Magnetic resonance imaging results
3.2.2.1. Regional grey matter alterations in MDD versus controlparticipants. Compared to healthy controls, participants withMDD showed significant decreases in regional grey mattervolume in the amygdala, hippocampal and parahippocampalregions (Brodmann area BA 36) and the ventral (BA 20) andmedial temporal lobes (BA 9, BA 21) as well as in insularcortex (BA 13). See Fig. 2A and Table 1 for results.
3.2.2.2. Correlation of grey matter volume with discriminationaccuracy (dL) for disgust. Having established a significantreduction in insular cortex grey matter volume in MDD, wetested whether discrimination accuracy (dL) for disgust wasassociated with insular cortex volume. Across all MDDparticipants, the average discrimination accuracy (dL) fordisgust was positively correlated with grey matter volume inthe right and left anterior insular cortex, (BA 13) and medialprefrontal gyrus (BA 10). See Fig. 2B and Table 2 for results.
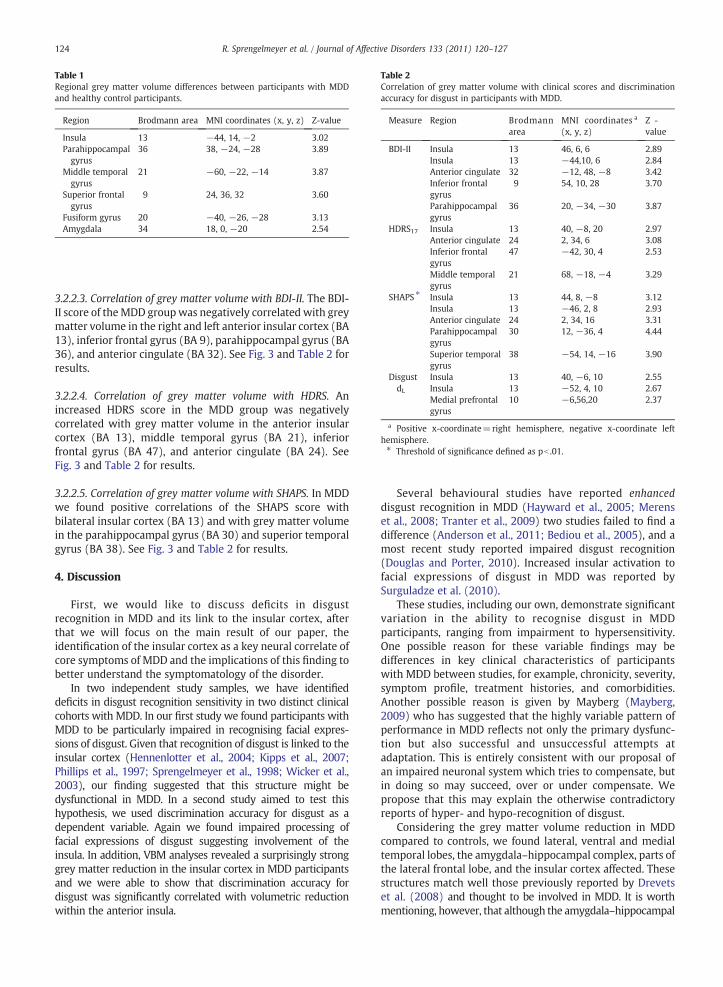
Table 1Regional grey matter volume differences between participants with MDDand healthy control participants.
Region Brodmann area MNI coordinates (x, y, z) Z-value
Insula 13 −44, 14, −2 3.02Parahippocampalgyrus
36 38, −24, −28 3.89
Middle temporalgyrus
21 −60, −22, −14 3.87
Superior frontalgyrus
9 24, 36, 32 3.60
Fusiform gyrus 20 −40, −26, −28 3.13Amygdala 34 18, 0, −20 2.54
Table 2Correlation of grey matter volume with clinical scores and discriminationaccuracy for disgust in participants with MDD.
Measure Region Brodmannarea
MNI coordinates a
(x, y, z)Z -value
BDI-II Insula 13 46, 6, 6 2.89Insula 13 −44,10, 6 2.84Anterior cingulate 32 −12, 48, −8 3.42Inferior frontalgyrus
9 54, 10, 28 3.70
Parahippocampalgyrus
36 20, −34, −30 3.87
HDRS17 Insula 13 40, −8, 20 2.97Anterior cingulate 24 2, 34, 6 3.08Inferior frontalgyrus
47 −42, 30, 4 2.53
Middle temporalgyrus
21 68, −18, −4 3.29
SHAPS ⁎ Insula 13 44, 8, −8 3.12Insula 13 −46, 2, 8 2.93Anterior cingulate 24 2, 34, 16 3.31Parahippocampalgyrus
30 12, −36, 4 4.44
Superior temporalgyrus
38 −54, 14, −16 3.90
DisgustdL
Insula 13 40, −6, 10 2.55Insula 13 −52, 4, 10 2.67Medial prefrontalgyrus
10 −6,56,20 2.37
a Positive x-coordinate=right hemisphere, negative x-coordinate lefthemisphere.⁎ Threshold of significance defined as pb .01.
124 R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
3.2.2.3. Correlation of grey matter volume with BDI-II. The BDI-II score of theMDD groupwas negatively correlated with greymatter volume in the right and left anterior insular cortex (BA13), inferior frontal gyrus (BA 9), parahippocampal gyrus (BA36), and anterior cingulate (BA 32). See Fig. 3 and Table 2 forresults.
3.2.2.4. Correlation of grey matter volume with HDRS. Anincreased HDRS score in the MDD group was negativelycorrelated with grey matter volume in the anterior insularcortex (BA 13), middle temporal gyrus (BA 21), inferiorfrontal gyrus (BA 47), and anterior cingulate (BA 24). SeeFig. 3 and Table 2 for results.
3.2.2.5. Correlation of grey matter volume with SHAPS. In MDDwe found positive correlations of the SHAPS score withbilateral insular cortex (BA 13) and with grey matter volumein the parahippocampal gyrus (BA 30) and superior temporalgyrus (BA 38). See Fig. 3 and Table 2 for results.
4. Discussion
First, we would like to discuss deficits in disgustrecognition in MDD and its link to the insular cortex, afterthat we will focus on the main result of our paper, theidentification of the insular cortex as a key neural correlate ofcore symptoms of MDD and the implications of this finding tobetter understand the symptomatology of the disorder.
In two independent study samples, we have identifieddeficits in disgust recognition sensitivity in two distinct clinicalcohorts with MDD. In our first study we found participants withMDD to be particularly impaired in recognising facial expres-sions of disgust. Given that recognition of disgust is linked to theinsular cortex (Hennenlotter et al., 2004; Kipps et al., 2007;Phillips et al., 1997; Sprengelmeyer et al., 1998; Wicker et al.,2003), our finding suggested that this structure might bedysfunctional in MDD. In a second study aimed to test thishypothesis, we used discrimination accuracy for disgust as adependent variable. Again we found impaired processing offacial expressions of disgust suggesting involvement of theinsula. In addition, VBM analyses revealed a surprisingly stronggrey matter reduction in the insular cortex in MDD participantsand we were able to show that discrimination accuracy fordisgust was significantly correlated with volumetric reductionwithin the anterior insula.
Several behavioural studies have reported enhanceddisgust recognition in MDD (Hayward et al., 2005; Merenset al., 2008; Tranter et al., 2009) two studies failed to find adifference (Anderson et al., 2011; Bediou et al., 2005), and amost recent study reported impaired disgust recognition(Douglas and Porter, 2010). Increased insular activation tofacial expressions of disgust in MDD was reported bySurguladze et al. (2010).
These studies, including our own, demonstrate significantvariation in the ability to recognise disgust in MDDparticipants, ranging from impairment to hypersensitivity.One possible reason for these variable findings may bedifferences in key clinical characteristics of participantswith MDD between studies, for example, chronicity, severity,symptom profile, treatment histories, and comorbidities.Another possible reason is given by Mayberg (Mayberg,2009) who has suggested that the highly variable pattern ofperformance in MDD reflects not only the primary dysfunc-tion but also successful and unsuccessful attempts atadaptation. This is entirely consistent with our proposal ofan impaired neuronal system which tries to compensate, butin doing so may succeed, over or under compensate. Wepropose that this may explain the otherwise contradictoryreports of hyper- and hypo-recognition of disgust.
Considering the grey matter volume reduction in MDDcompared to controls, we found lateral, ventral and medialtemporal lobes, the amygdala–hippocampal complex, parts ofthe lateral frontal lobe, and the insular cortex affected. Thesestructures match well those previously reported by Drevetset al. (2008) and thought to be involved in MDD. It is worthmentioning, however, that although the amygdala–hippocampal
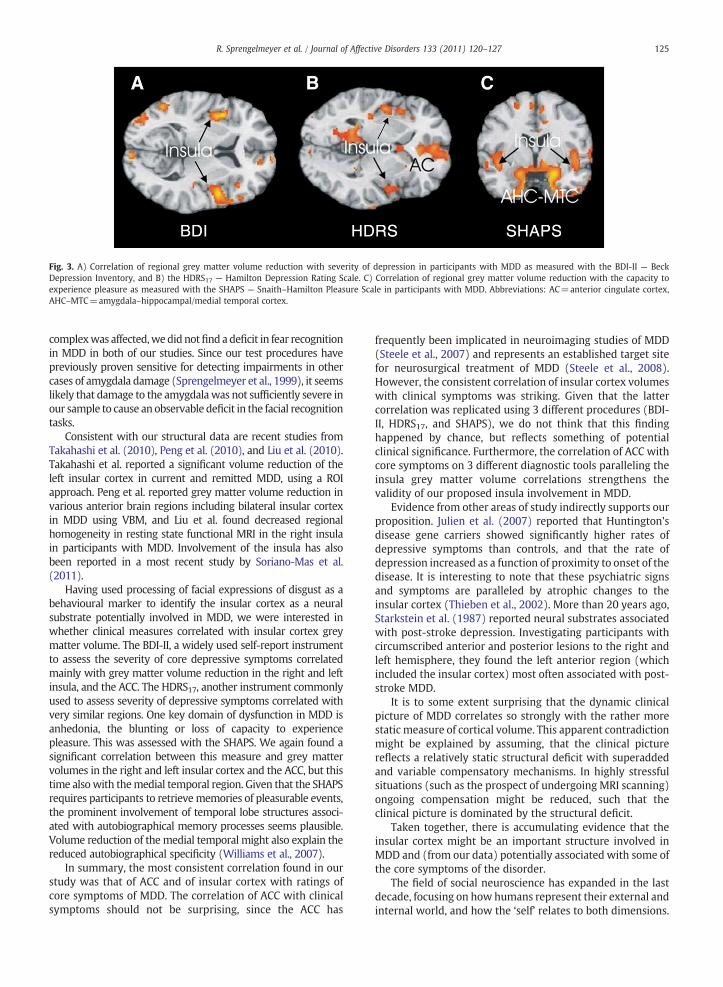
Fig. 3. A) Correlation of regional grey matter volume reduction with severity of depression in participants with MDD as measured with the BDI-II — BeckDepression Inventory, and B) the HDRS17 — Hamilton Depression Rating Scale. C) Correlation of regional grey matter volume reduction with the capacity toexperience pleasure as measured with the SHAPS — Snaith–Hamilton Pleasure Scale in participants with MDD. Abbreviations: AC=anterior cingulate cortex,AHC–MTC=amygdala–hippocampal/medial temporal cortex.
125R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
complexwas affected,wedid notfind a deficit in fear recognitionin MDD in both of our studies. Since our test procedures havepreviously proven sensitive for detecting impairments in othercases of amygdala damage (Sprengelmeyer et al., 1999), it seemslikely that damage to the amygdalawas not sufficiently severe inour sample to cause an observable deficit in the facial recognitiontasks.
Consistent with our structural data are recent studies fromTakahashi et al. (2010), Peng et al. (2010), and Liu et al. (2010).Takahashi et al. reported a significant volume reduction of theleft insular cortex in current and remitted MDD, using a ROIapproach. Peng et al. reported grey matter volume reduction invarious anterior brain regions including bilateral insular cortexin MDD using VBM, and Liu et al. found decreased regionalhomogeneity in resting state functional MRI in the right insulain participants with MDD. Involvement of the insula has alsobeen reported in a most recent study by Soriano-Mas et al.(2011).
Having used processing of facial expressions of disgust as abehavioural marker to identify the insular cortex as a neuralsubstrate potentially involved in MDD, we were interested inwhether clinical measures correlated with insular cortex greymatter volume. The BDI-II, a widely used self-report instrumentto assess the severity of core depressive symptoms correlatedmainly with grey matter volume reduction in the right and leftinsula, and the ACC. The HDRS17, another instrument commonlyused to assess severity of depressive symptoms correlated withvery similar regions. One key domain of dysfunction in MDD isanhedonia, the blunting or loss of capacity to experiencepleasure. This was assessed with the SHAPS. We again found asignificant correlation between this measure and grey mattervolumes in the right and left insular cortex and the ACC, but thistime alsowith themedial temporal region. Given that the SHAPSrequires participants to retrievememories of pleasurable events,the prominent involvement of temporal lobe structures associ-ated with autobiographical memory processes seems plausible.Volume reduction of themedial temporal might also explain thereduced autobiographical specificity (Williams et al., 2007).
In summary, the most consistent correlation found in ourstudy was that of ACC and of insular cortex with ratings ofcore symptoms of MDD. The correlation of ACC with clinicalsymptoms should not be surprising, since the ACC has
frequently been implicated in neuroimaging studies of MDD(Steele et al., 2007) and represents an established target sitefor neurosurgical treatment of MDD (Steele et al., 2008).However, the consistent correlation of insular cortex volumeswith clinical symptoms was striking. Given that the lattercorrelation was replicated using 3 different procedures (BDI-II, HDRS17, and SHAPS), we do not think that this findinghappened by chance, but reflects something of potentialclinical significance. Furthermore, the correlation of ACC withcore symptoms on 3 different diagnostic tools paralleling theinsula grey matter volume correlations strengthens thevalidity of our proposed insula involvement in MDD.
Evidence from other areas of study indirectly supports ourproposition. Julien et al. (2007) reported that Huntington'sdisease gene carriers showed significantly higher rates ofdepressive symptoms than controls, and that the rate ofdepression increased as a function of proximity to onset of thedisease. It is interesting to note that these psychiatric signsand symptoms are paralleled by atrophic changes to theinsular cortex (Thieben et al., 2002). More than 20 years ago,Starkstein et al. (1987) reported neural substrates associatedwith post-stroke depression. Investigating participants withcircumscribed anterior and posterior lesions to the right andleft hemisphere, they found the left anterior region (whichincluded the insular cortex) most often associated with post-stroke MDD.
It is to some extent surprising that the dynamic clinicalpicture of MDD correlates so strongly with the rather morestatic measure of cortical volume. This apparent contradictionmight be explained by assuming, that the clinical picturereflects a relatively static structural deficit with superaddedand variable compensatory mechanisms. In highly stressfulsituations (such as the prospect of undergoing MRI scanning)ongoing compensation might be reduced, such that theclinical picture is dominated by the structural deficit.
Taken together, there is accumulating evidence that theinsular cortex might be an important structure involved inMDD and (from our data) potentially associated with some ofthe core symptoms of the disorder.
The field of social neuroscience has expanded in the lastdecade, focusing on how humans represent their external andinternal world, and how the ‘self’ relates to both dimensions.

126 R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
Many studies address domains and functions, which appearto be affected inMDD, andmany of these studies highlight theinsular cortex as a neural substrate potentially associatedwith these functions. Functional imaging studies of self-related processes found increased insula activation associatedwith thinking about oneself (Modinos et al., 2009), self-assurance in situations of personal set back, mistake or failure(Longe et al., 2010), frustration (Abler et al., 2005), joy(Takahashi et al., 2008), attending to pleasant music (Koelschet al., 2006), and affective touch (Olausson et al., 2002) as oneaspect of an intimate social interaction. There is furtherevidence that our sense of affiliation may depend on theintegrity of the insula. Gobbini et al. (2004) reportedactivation in right and left insular cortex to personallyfamiliar people, and Bartels and Zeki (2004) found increasedinsula activation when participants viewed pictures of theirown children. Similar results came from Leibenluft et al.(2004) who found increased activation of the insular cortexto own compared to familiar children, and to familiar childrencompared to unfamiliar ones. Decoding visually and vocallydisplayed social signals (Johnstone et al., 2006; Kipps et al.,2007; Sprengelmeyer et al., 1998) as well as empathicfeelings are associated with the insular cortex (Hennenlotteret al., 2005; Singer et al., 2004; Wicker et al., 2003). Theresponse to unfair offers (Sanfey et al., 2003) and feelingsocially rejected (Eisenberger et al., 2003) has been linked tothe insular cortex as well as sexual pleasure (Komisaruk et al.,2004).
If future studies substantiate this putative link betweeninsular cortex functioning and core symptoms of MDD, itwould be possible to define some aspects of the highlyvariable clinical picture of MDD more precisely in terms ofimpaired cognitive and emotional processes.
Role of funding sourceThe study “The insular cortex and the neuroanatomy of major
depression” received no funding.
Conflict of interestThere is no conflict of interest.
Acknowledgements
We would like to thank Andy Young, Dave Perrett, JeremyHall, and Verity Brown for helpful comments and discussions.
References
Abler, B., Walter, H., Erk, S., 2005. Neural correlates of frustration.Neuroreport 16, 669–672.
Adolphs, R., Tranel, D., Damasio, H., Damasio, A., 1994. Impaired recognitionof emotion in facial expressions following bilateral damage to the humanamygdala. Nature 372, 669–672.
Anderson, I.M., Shippen, C., Juhasz, G., Chase, D., Thomas, E., Downey, D., Toth,Z.G., Lloyd-Williams, K., Elliott, R., Deakin, J.F., 2011. State-dependentalteration in face emotion recognition in depression. Br. J. Psychiatry[Epub ahead of print].
Bartels, A., Zeki, S., 2004. The neural correlates of maternal and romantic love.Neuroimage 21, 1155–1166.
Beck, A., Steer, R., Brown, G., 1996. Manual for the Beck DepressionInventory-II. Psychological Corporation, San Antonio, TX.
Bediou, B., Krolak-Salmon, P., Saoud, M., Henaff, M.A., Burt, M., Dalery, J.,D'Amato, T., 2005. Facial expression and sex recognition in schizophreniaand depression. Can. J. Psychiatry 50, 525–533.
Corwin, J., 1994. On measuring discrimination and response bias: unequalnumbers of targets and distractors and two classes of distractors.Neuropsychology 8, 110–117.
Douglas, K.M., Porter, R.J., 2010. Recognition of disgusted facial expressions insevere depression. Br. J. Psychiatry 197, 156–157.
Drevets, W.C., Price, J.L., Furey, M.L., 2008. Brain structural and functionalabnormalities in mood disorders: implications for neurocircuitry modelsof depression. Brain Struct. Funct. 213, 93–118.
Eisenberger, N.I., Lieberman, M.D., Williams, K.D., 2003. Does rejection hurt?An FMRI study of social exclusion. Science 302, 290–292.
Ekman, P., Friesen, W.V., 1976. Pictures of Facial Affect. ConsultingPsychologist's Press, Palo Alto, CA.
Friston, K.J., Ashburner, J.T., Kiebel, S.J., Nichols, T.E., Penny, W.D., 2007.Statistical Parametric Mapping: The Analysis of Functional Brain Images.Academic Press, London.
Gobbini, M.I., Leibenluft, E., Santiago, N., Haxby, J.V., 2004. Social andemotional attachment in the neural representation of faces. Neuroimage22, 1628–1635.
Hamilton, M.A., 1960. A rating scale for depression. J. Neurol. Neurosurg.Psychiatry 23, 56–62.
Hayward, G., Goodwin, G.M., Cowen, P.J., Harmer, C.J., 2005. Low-dosetryptophan depletion in recovered depressed patients induces changesin cognitive processing without depressive symptoms. Biol. Psychiatry 57,517–524.
Hennenlotter, A., Schroeder, U., Erhard, P., Castrop, F., Haslinger, B., Stoecker,D., Lange, K.W., Ceballos-Baumann, A.O., 2005. A common neural basisfor receptive and expressive communication of pleasant facial affect.Neuroimage 26, 581–591.
Hennenlotter, A., Schroeder, U., Erhard, P., Haslinger, B., Stahl, R., Weindl, A.,von Einsiedel, H.G., Lange, K.W., Ceballos-Baumann, A.O., 2004. Neuralcorrelates associated with impaired disgust processing in pre-symp-tomatic Huntington's disease. Brain 127, 1446–1453.
Johnstone, T., van Reekum, C.M., Oakes, T.R., Davidson, R.J., 2006. The voice ofemotion: an FMRI study of neural responses to angry and happy vocalexpressions. Soc. Cogn. Affect. Neurosci. 1, 242–249.
Julien, C.L., Thompson, J.C., Wild, S., Yardumian, P., Snowden, J.S., Turner, G.,Craufurd, D., 2007. Psychiatric disorders in preclinical Huntington'sdisease. J. Neurol. Neurosurg. Psychiatry 939–943.
Kipps, C.M., Duggins, A.J., McCusker, E.A., Calder, A.J., 2007. Disgust andhappiness recognition correlate with anteroventral insula and amygdalavolume respectively in preclinical Huntington's disease. J. Cogn.Neurosci. 19, 1206–1217.
Koelsch, S., Fritz, T., Cramon, D., Muller, K., Friederici, A.D., 2006. Investigatingemotion with music: an fMRI study. Hum. Brain Mapp. 27, 239–250.
Komisaruk, B.R., Whipple, B., Crawford, A., Liu, W.C., Kalnin, A., Mosier, K.,2004. Brain activation during vaginocervical self-stimulation and orgasmin women with complete spinal cord injury: fMRI evidence of mediationby the vagus nerves. Brain Res. 1024, 77–88.
Leibenluft, E., Gobbini, M.I., Harrison, T., Haxby, J.V., 2004. Mothers' neuralactivation in response to pictures of their children and other children.Biol. Psychiatry 56, 225–232.
Liu, Z., Xu, C., Xu, Y., Wang, Y., Zhao, B., Lv, Y., Cao, X., Zhang, K., Du, C., 2010.Decreased regional homogeneity in insula and cerebellum: a resting-state fMRI study in patients with major depression and subjects at highrisk for major depression. Psychiatry Res 30, 211–215.
Longe, O., Maratos, F.A., Gilbert, P., Evans, G., Volker, F., Rockliff, H., Rippon, G.,2010. Having a wordwith yourself: neural correlates of self-criticism andself-reassurance. Neuroimage 49, 1849–1856.
Mayberg, H.S., 2009. Targeted electrode-based modulation of neural circuitsfor depression. J. Clin. Invest. 119, 717–725.
Merens, W., Booij, L., Haffmans, P.J., van der Does, A., 2008. The effects ofexperimentally lowered serotonin function on emotional informationprocessing and memory in remitted depressed patients. J. Psychophar-macol. 22, 653–662.
Modinos, G., Ormel, J., Aleman, A., 2009. Activation of anterior insula duringself-reflection. PLoS One 4, e4618.
Nelson, H.E., 1982. The National Adult Reading Test (NART): Test Manual.NFER-Nelson, Windsor, Berkshire.
Olausson, H., Lamarre, Y., Backlund, H., Morin, C., Wallin, B.G., Starck, G.,Ekholm, S., Strigo, I., Worsley, K., Vallbo, A.B., Bushnell, M.C., 2002.Unmyelinated tactile afferents signal touch and project to insular cortex.Nat. Neurosci. 5, 900–904.
Peng, J., Liu, J., Nie, B., Li, Y., Shan, B., Wang, G., Li, K., 2010. Cerebral andcerebellar gray matter reduction in first-episode patients with majordepressive disorder: a voxel-based morphometry study. Eur. J. Radiol.[Epub ahead of print].
Phillips, M.L., Young, A.W., Senior, C., Brammer, M., Andrew, C., Calder, A.J.,Bullmore, E.T., Perrett, D.I., Rowland, D., Williams, S.C., Gray, J.A., David,A.S., 1997. A specific neural substrate for perceiving facial expressions ofdisgust. Nature 389, 495–498.

127R. Sprengelmeyer et al. / Journal of Affective Disorders 133 (2011) 120–127
Ross, R.S., Slotnick, S.D., 2008. The hippocampus is preferentially associatedwith memory for spatial context. J. Cogn. Neurosci. 20, 432–446.
Sanfey, A.G., Rilling, J.K., Aronson, J.A., Nystrom, L.E., Cohen, J.D., 2003. Theneural basis of economic decision-making in the ultimatum game.Science 300, 1755–1758.
Singer, T., Seymour, B., O'Doherty, J., Kaube, H., Dolan, R.J., Frith, C.D., 2004.Empathy for pain involves the affective but not sensory components ofpain. Science 303, 1157–1162.
Snaith, R.P., Hamilton, M., Morley, S., Humayan, A., Hargreaves, D., Trigwell,P., 1995. A scale for the assessment of hedonic tone the Snaith–HamiltonPleasure Scale. Br. J. Psychiatry 167, 99–103.
Soriano-Mas, C., Hernandez-Ribas, R., Pujol, J., Urretavizcaya, M., Deus, J.,Harrison, B.J., Ortiz, H., Lopez-Sola, M., Menchon, J.M., Cardoner, N., 2011.Cross-sectional and longitudinal assessment of structural brain alter-ations in melancholic depression. Biol. Psychiatry 69, 318–325.
Sprengelmeyer, R., Rausch, M., Eysel, U.T., Przuntek, H., 1998. Neuralstructures associated with recognition of facial expressions of basicemotions. Proc. Biol. Sci. 265, 1927–1931.
Sprengelmeyer, R., Young, A.W., Schroeder, U., Grossenbacher, P.G., Feder-lein, J., Buttner, T., Przuntek, H., 1999. Knowing no fear. Proc. Biol. Sci.266, 2451–2456.
Starkstein, S.E., Robinson, R.G., Price, T.R., 1987. Comparison of cortical andsubcortical lesions in the production of poststroke mood disorders. Brain110 (4), 1045–1059.
Steele, J.D., Christmas, D., Eljamel, M.S., Matthews, K., 2008. Anteriorcingulotomy for major depression: clinical outcome and relationship tolesion characteristics. Biol. Psychiatry 63, 670–677.
Steele, J.D., Currie, J., Lawrie, S.M., Reid, I., 2007. Prefrontal cortical functionalabnormality in major depressive disorder: a stereotactic meta-analysis.J. Affect. Disord. 101, 1–11.
Surguladze, S.A., El-Hage, W., Dalgleish, T., Radua, J., Gohier, B., Phillips, M.L.,2010. Depression is associated with increased sensitivity to signals of
disgust: a functional magnetic resonance imaging study. J. Psychiatr. Res.44, 894–902.
Takahashi, H., Matsuura, M., Koeda, M., Yahata, N., Suhara, T., Kato, M., Okubo, Y.,2008. Brain activations during judgments of positive self-consciousemotion and positive basic emotion: pride and joy. Cereb. Cortex 18,898–903.
Takahashi, T., Yücel, M., Lorenzetti, V., Tanino, R., Whittle, S., Suzuki, M.,Walterfang, M., Pantelis, C., Allen, N.B., 2010. Volumetric MRI study of theinsular cortex in individuals with current and past major depression.J. Affect. Disord. 121, 231–238.
Thieben, M.J., Duggins, A.J., Good, C.D., Gomes, L., Mahant, N., Richards, F.,McCusker, E., Frackowiak, R.S., 2002. The distribution of structuralneuropathology in pre-clinical Huntington's disease. Brain 125,1815–1828.
Tranter, R., Bell, D., Gutting, P., Harmer, C., Healy, D., Anderson, I.M., 2009. Theeffect of serotonergic and noradrenergic antidepressants on faceemotion processing in depressed patients. J. Affect. Disord. 118, 87–93.
Walther, K., Bendlin, B.B., Glisky, E.L., Trouard, T.P., Lisse, J.R., Posever, J.O.,Ryan, L., 2009. Anti-inflammatory drugs reduce age-related decreases inbrain volume in cognitively normal older adults. Neurobiol. Aging.
Wicker, B., Keysers, C., Plailly, J., Royet, J.P., Gallese, V., Rizzolatti, G., 2003.Both of us disgusted inMy insula: the common neural basis of seeing andfeeling disgust. Neuron 40, 655–664.
Williams, J.M., Barnhofer, T., Crane, C., Herman, D., Raes, F., Watkins, E.,Dalgleish, T., 2007. Autobiographical memory specificity and emotionaldisorder. Psychol. Bull. 133, 122–148.
Young, A.W., Newcombe, F., de Haan, E.H., Small, M., Hay, D.C., 1993. Faceperception after brain injury. Selective impairments affecting identityand expression. Brain 116 (Pt 4), 941–959.
Young, A.W., Perrett, D.I., Calder, A., Sprengelmeyer, R., Ekman, P., 2002.Facial Expressions of Emotion: Stimuli and Tests (FEEST). Thames ValleyTest Company, Bury St. Edmunds, Suffolk.