Embed Size (px)
Citation preview

Hum Genet (1991) 86:509-514
�9 Springer-Verlag 1991
The human aldose reductase gene maps to chromosome region 7q35
Alexander Graham, Paul Heath, John E. N. Morten, and Alexander F.Markham
Biotechnology Department, ICI Pharmaceuticals, Alderley Park, Macclesfield SK10 4TG, UK
Received July 5, 1990
Summary. The human aldose reductase (AR) gene has been mapped to chromosome 7 using the polymerase chain reaction to specifically amplify the human A R se- quence in hamster/human hybrid D N A and also in mouse/ human monochromosome hybrids. The assignment to chromosome 7 was confirmed by in situ hybridisation to human metaphase chromosomes using a novel, rapid hybridisation, method giving a regional localisation at 7q35.
Introduction
Human aldose reductase (AR; EC 1.1.1.21; also refer- red to as "low Km" aldehyde reductase) catalyses the first reaction in the polyol pathway of D-glucose metabo- lism. This is an N A D P H - d e p e n d e n t reduction of glucose to sorbitol. Aldose reductase is implicated in the de- velopment of a variety of diabetic complications, and in- hibitors of this enzyme have been developed and tested in patients for the prevention of such complications. Sev- eral groups have recently isolated the c D N A for human aldose reductase (Bohren et al. 1989; Chung and La Mendola 1989; Graham et al. 1989; Grundmann et al. 1990). We have isolated genomic cosmid clones for aldose reductase and have characterised the gene coding for the enzyme (Graham et al. 1991). We have also screened a human genomic yeast artificial chromosome (YAC) library (Anand et al. 1990) and isolated a Y A C clone containing the complete aldose reductase gene on a frag- ment of 180 kb (Elvin et al. 1990).
Here we report on the regional localisation of A R by the use of the polymerase chain reaction (PCR) to speci- fically amplify the human A R sequence in somatic cell hybrids. The polymerase chain reaction has been used by several laboratories for the mapping of human genes in somatic cell hybrids (Abbot t et al. 1989; Dionne et al. 1990).
Offprint requests to: A. Graham
Concurrently, and independently, we have mapped this gene using in situ hybridisation. We have used an A R YAC, an A R cosmid and A R intronic sequences (PCR products) for the in situ hybridisation.
Materials and methods
Somatic cell hybrids and PCR
The human-hamster somatic cell panel of PCRable TM DNAs was obtained from Bios Corporation (New Haven, Conn.). The pro- duction of human mouse monochromosome hybrids has been pre- viously described (McNeill and Brown 1980; Saxon et al. 1985) and the DNA was isolated by standard methods. Because the hy- brids are not diploid we used 100ng per reaction for hybrid DNA and 50 ng per reaction of whole genomic DNA from human, ham- ster or mouse.
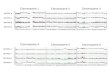
Oligonucleotide amplimers for PCR screening of the somat- ic cell hybrids were: human AR intron 5 (Graham et al. 1991); 5'd(GTGAGCTI'CCCACAGGCTCATGCTCCTGTG) and 5'd (GATAGAAAGACAGTCCAGGTCACACCATCA), which gives a PCR product of 320bp; human AR intron 6 (Graham et al. 1991), 5'd(CAGCCACITCAGGTGTI'GCTGACCGTCCAC) and 5' d (AAGGACGTGCTAGAACTCCACAGGTGATCA), which gives a PCR product of 510bp; and the human cystic fibrosis gene exon 13 (Riordan et al. 1989), 5'd(CTGTGTCTGTAAACTGAT GGCTAAC) and 5'd(ATCCAGTI'CAGTCAAGTITGCCTG), which gives a PCR product of 652bp. PCR (Saiki et al. 1985) was performed with 50 ng of whole genomic DNA (human, hamster or mouse DNA) and with 100 ng of DNA from the somatic cell hy- brids using 50 pmol of each amplimer pair, 10 mM Tris HCI pH 8.3 (at 25~ 50raM KC1, 1.2ram MgC12, 0.01% (w/v) gelatin and 200 [aM each of dATP, dCTP, dGTP and dTTP in a final volume of 100 I J1. Samples were boiled for 5 rain, then two units of Taq DNA polymerase (Perkin-Elmer Cetus, Emeryville, Calif.) were added and the samples overlaid with 50 gl mineral oil. Thirty-three cycles of amplification were performed at 66~ for 2 rain, 72~ for 2 rain and 92~ for 2min using an automated Techne PHC-1 Thermal Cycler. Aliquots of 18gl were removed and analysed on a 1.2% agarose gel and visualised with ethidium bromide. The results shown in Fig. 1 were obtained by mixing 18 lal of separate amplifi- cation reactions performed with AR intron 5, AR intron 6 and CF exon 13 amplimers and then drying each mixture down to 20gl, followed by agarose electrophoresis.

510
In situ hybridisation on metaphase chromosomes
Normal male PHA-stimulated lymphocytes were cultured in RPMI 1640, 10% fetal calf serum for 72 h. BrdU (200 gg/ml) was added for 17 h, the cells were washed three times in phosphate-buffered saline and cultured for a further 4 h in fresh medium supplemented with 10-5M thymidine. One hour before harvest Colcemid (0.1 gg/ ml) was added. The origins of the AR-YAC (4FA1) and AR cos- mid probes (AR-C) have been described elsewhere (Elvin et al. 1990; Graham et al. 1991). Probes for intron 8 and intron 9 of AR were generated by PCR. The oligonucleotide amplimers were for human AR intron 8,5'd(TGCACTGGAGCTCTGTCAGCTACC) and 5 'd(TATATGGAACGATGGAAACAG), which gives a PCR product of 1470bp, and AR intron 9,5'd(GTTGGCCTGGCCTGAA GTAACTAC) and 5'd(GGCCAGACACAGTGGCTCACGCC), which gives a PCR product of 1868 bp. Introns 8 and 9 are 1952 bp and 2 460 bp respectively (Graham et al. 1991). PCR was performed as described above except that for intron 8 the annealing tempera- ture was lowered to 55~ The probes (100 ng of each) were label- led with Biotin-ll-UTP (BRL, Gaithersburg, Md.) using an Amer- sham (UK) Multiprime DNA labelling system. For the YAC and cosmid probes, repeat sequences were quenched by re-annealing to human placental DNA (Elvin et al. 1990). In brief, following the labelling reactions the probes were phenol/chloroform extracted, ethanol precipitated and resuspended in H20. Prior to the rean- nealing reaction, we calculated the amount of DNA required to drive the reaction to Cot250 over 90 rain at 65~ Sheared human placental DNA was added and, after 10rain denaturation, SSC was added to a final concentration of 5 x SSC.
Slides were dehydrated through an alcohol series and incu- bated for 1 h at 37~ with 100 gg/ml RNAase in 2 x SSC under a coverslip in moist conditions. Following a PBS wash they were pas- sed through a fresh alcohol series.
Hybridisation was at 42~ overnight in moist conditions. 10 ng probe per slide was applied in a solution containing 50% formamide, 10% dextran sulphate, 2 • SSC, 0.1 mM EDTA, 0.05 mM Tris pH 7.6, 100 gg/ml denatured salmon sperm DNA. The mix was cov- ered with a coverslip, sealed with Cow Gum and denatured at 80~ for 10 rain prior to the overnight incubation. Following the re- moval of the coverslip, slides were washed twice in 2 x SSC at room temp for 30min, washed once in 0.1 x SSC at 42~ for 30 rain and finally for 15 min in 2 x SSC at 42~
The slides were coated with 100 gl DAKO streptavidin diluted 1:10000 in TBS (50raM Tris pH 7.6, 150raM NaC1, 2raM MgCI2, 0.1% BSA) for 30rain. Slides were washed in TBS then coated with 100gl DAKO biotinylated alkaline phosphatase (1/1000 in TBS) for 30 rain. Slides were washed again in TBS and the process repeated. The signal was observed by rinsing the slides in veronal acetate buffer and staining with fast red for lb . [Veronal acetate buffer: 30 mM sodium acetate, 30 mM sodium barbitone, 100 mM NaCI, 50raM MgCI2 pH 9.2. Fast Red stain: 150rag fast red, 150 mg naphthol ASBI phosphate (predissolved in 1 ml dimethylform- amide), and 72 mg levamisole in 300 ml veronal acetate buffer].
Slides were counterstained with 12.5% Giemsa in 0.3 M di- sodium hydrogen phosphate pH 10.4 for 1 rain (Aghamahammadi and Savage 1989). Hence the signal distribution could be visualised by direct microscopic observation. ?~2 analysis used the relative chromosome lengths given in ISCN 1985, Appendix 2D.
Results and discussion
The resul ts o b t a i n e d by P C R using the A R in t ron 5, A R in t ron 6 and C F exon 13 p r imers are shown in Fig. 1 and the resul ts of the ass ignment of the A R gene are sum- mar i s ed in Tab le 1. E a c h o f the ampl i f i ca t ion reac t ions was ini t ial ly p e r f o r m e d and ana lysed s epa ra t e ly (da ta no t shown) . F r o m the resul ts , the h u m a n D N A ( U P O O 4 and 6167) was specif ical ly ampl i f i ed to give the expec t ed
p roduc t s of 320bp ( A R in t ron 5), 5 1 0 b p ( A R in t ron 6) and 652 bp (CF exon 13). The A R in t ron 5 and in t ron 6 ampl imer s give p roduc t s wi th an A R cosmid c lone which we have i so la ted , whe reas the C F exon 13 amp l imer s do not give a p r o d u c t wi th this cosmid (see Fig. 1). M i n o r bands of the same sizes were seen with hamster ( C H O 1 0 4 ) and mouse (MEK79) cell lines under condit ions of higher Mg 2+ concen t ra t ions or lower annea l ing t e m p e r a t u r e s . H o w e v e r , wi th the cond i t ions desc r ibed above , the hu-
Fig. 1. PCR of human AR (introns 5 and 6) and CF exon 13 on so- matic cell hybrids and their parent cell lines. PCR using oligonu- cleotides specific for AR intron 5 (320bp product), AR intron 6 (510bp product) and CF exon 13 (652bp product) on a panel of hamster/human and mouse/human hybrids. PCRs 1-25 correspond to hamster/human hybrids (Bios Corporation) and 26-33 corre- spond to mouse/human monochromosome hybrids (see Table 1 for cell-line designation). Control PCRs with human DNA (UPO04 and 6167), hamster DNA (CHOI04) and mouse DNA (MEK79) are also shown. An AR cosmid DNA encoding the complete AR gene was also used as a template for PCR (50pg per reaction). Separate PCRs were performed with each amplimer pair and then equal volumes of each reaction were mixed and subjected to aga- rose gel electrophoresis (size marker d~x174 DNA cut with HaeIII, 1.5 gg per track, BRL)

5 1 1
, ~ ~ ~ o *-~
o .~ o~ . t~
m ~
+ ~ + ~
�9
+
~ . . ~ ,..~
I l l l l + + l + I I + l [ I +
+
+ I
I I
I I
I +
I +
I +
I
I
I
+
I + l l l
I + l l l
I I + l l
I I I +
~ +
I +
I
I
I
+
+
I I
I I I
~ + ~
I + +
~ +
I + +
I I I -I-
I I I +
I I I
I I I
+ + ~
I I -I-
+ ~ - +
I + I
I I I
I I I
+ I
I I
+
[
I
I
+
I
+
- I - +
I I
I
I
+ I
I + I
I
I
+
I
I
I
I
I
I
I
+
I
I
I
-I- I
[ I
-I-
I
+
I
I
I
+
I
+ +
I +
+ +
+ +
I +
+ +
I +
I +
+ l +
~ l +
I 1 +
I + +
I I I +
I 1 + 1 +
i l l ~ +
+ I l l +
+ I
[ I
I l l I 1 +
l [ + + l l l
I + l § §
I l l + l l +
i + i I + i i + + I I I I I +
o-~ + ~ + + + + ~ + + ~ + + ~ + + ~ + + + + + ~ +
i i + i i r i i i i i I I l I I i I ~ I I I +
+ I I i ~ i I I + I I i [ i i i [ i i i i + i +
i i i i [ i i i i i i i [ I I + [ i I l I i i +
I i I I i ~ I I I I i i I I + I i i I I I i + +
I I I I I I I I I r + i I I I I l i I i i i i §
c q
o o
k O
e Q
r
<
+ l I
+ § I
+ l
+
+
+
+
+
+
+ +
+
+ I +
+ I
+ + l +
+ [ i
+ I I
+ I +
+ + +
+
+
+
+
+
+ +
+ i + + 0
~ ; + l + ~ + + ~

5 1 2
a
| P 5 q )
~ �9 ~ i " . : . : : ' . : . . . : ! ! : : : : . . . . . . : : : . : : . . . . . . . : . . i
6 ~ 7 ( 8 t 9 ( 10 I 11 ( 12 )
I p q )P q I p q t IJ q )P' q IP' q (P q)P'qiPq~P'qI P I q IP'ql b , 13 , 14 , 15 ( 16 i 1 7 ( i s ( 19 ) 2 0 { 21 / 2 2 # x ) Y i
! . !
C
~ .,
: ,d ,~-~-~ '~e& : : . . . . . : . . . . . : .
13 ' 14 ~ 15, I 10 ) 17 I 1 8 I 19 ] 2 0 ] 21 I 2 2 1 X I y [
d
t 1 I a " [ " a + ~ ~ [ P s q
, . - . , ,
~ . ~ _ ~ : : : : : ' " �9 . : . . i - . . . . : �9 .i : : . : i . . . : . :
6 7 I 8 I 9 I 10 I 11 I 12 I
Fig . 2. D i a g r a m of t h e s igna l d i s t r i b u t i o n in h u m a n m e t a p h a s e c h r o m o s o m e s h y b r i d i s e d w i t h a A R - Y A C D N A ( 4 F A 1 ) , b A R - c o s m i d D N A ( A R - C ) , e A R i n t r o n 8 P C R p r o d u c t a n d d A R i n t r o n 9 P C R p r o d u c t

Table 2. Distribution of signals in human metaphase chromosomes hybridised with AR probes
513
Probe No. of Total Chromosome 7 Z: meta- no. of phases signals No. of %
signals
7q32-qter 7q35
No. of % No. of % signals signals
AR-YAC (4FA1) 32 152 22 14.5 AR cosmid (AR-C) 51 240 54 22.5 AR intron 8 (PCR) 22 157 27 17.0 AR intron 9 (PCR) 42 247 61 24.6
24.5 15 9.8 14 9.2 147.0 35 14.5 26 10.8 45.0 14 8.9 11 7.0
200.0 30 12.0 18 7.3
man DNA was specifically amplified and no background bands were visible. We have repeated the PCR reactions with each amplimer pair and confirmed that the same re- sults are obtained. The gene for human cystic fibrosis is known to map to the long arm of chromosome 7 (7q22-31) and so we confirmed the localisation of this to chromo- some 7 as a control for the assignment of human AR. The results shown in Fig. 1 were obtained by mixing equal volumes from each of the reactions performed with separate amplimer pairs and, as can be seen, the hy- brids which generate AR intron 5 and AR intron 6 pro- ducts have also resulted in the amplification of the CF exon 13 product. This strongly suggests that the A R gene is located on chromosome 7.
The hybrid panel (Bios Corporation) has been cer- tified "PCRable" by the manufacturers. We have con- firmed this and checked that the monochromosome panel DNAs are PCRable by using amplimers specific for [3-hexosaminidase, which have been shown to give products with human, hamster and mouse D N A (Abbott et al. 1989). All of the hybrid DNAs could be amplified using this set of amplimers and gave the expected 123-bp product. We have also performed PCR chromosome as- signment with tissue plasminogen activator (t-PA) which is known to be found on chromosome 8 (Yang-Feng et al. 1985). The amplimers were 5 ' d ( C G C A T G A C T T T G G T G G G C A T C A T C ) and 5 ' d ( C T A A T G C A A T C C G T C T T T C C T G A A ) . We have confirmed that all the cell hybrids shown in Table 1 which contain a chromosome 8 give a PCR product of 537bp (data not shown). The localisation of the AR gene was confirmed by in situ hybridisation, using AR YAC DNA (4FA1), AR cosmid D N A (AR-C) and two AR intron PCR products (introns 8 and 9), to human metaphase chromosomes. The re- sults of the in situ hybridisations using these probes are summarised in Fig. 2 and Table 2. With each probe a sig- nificant signal was observed on chromosome 7, the major- ity of these signals being localised at band 7q35. A sec- ondary peak, accounting for 6% of the total signal, was seen with the intron 9 probe at 7p21; this was not seen with the other probes.
Intron 9 PCR product contains a complete Alu repeat sequence, which shows highest homology (74%) with human ornithine decarboxylase Alu repeat III (position 6123-6410) (Van Steeg et al. 1989). This PCR product was not pre-associated with human placental D N A prior to in situ hybridisation, and this may be the reason for signals in locations other than 7q35.
The use of alkaline phosphatase-fast red staining has not, to our knowledge, been reported for chromosomal in situ hybridisation, though it is used for in situ hybridi- sation with tissues. Before using this method to localise an unmapped gene we established its suitability by map- ping a gene whose location is known. Because we had isolated a series of overlapping YAC clones from the cystic fibrosis (CF) locus in this laboratory, we used one of these CF-YAC clones (14DC12), which contains a 475-kb fragment, as a probe for in situ hybridisation. This probe resulted in a signal at 7@2-31, the region known to be the location of the CF gene (Dean et al. 1985; Duncan et al. 1988). We had previously used tritiated 14DC12 as a probe to map the gene by standard in situ methods. The in situ technique described here is suitable for probes which vary in size from YAC clones (180- and 475-kb fragments), cosmids (40-kb fragment) to relative- ly small probes (1 500 bp).
Acknowledgements. We thank R. Anand, J. Riley and R. Butler for preparation of the human genome YAC library and P. Hedge for the human cosmid library. We would also thank Eric Stanbridge (University of California, Irvine) and his colleagues for the mono- chromosome hybrids.
References
Abbott C, West L, Povey S, Jeremiah S, Murad Z, DiScipio R, Fey G (1989) The gene for human complement component C9 mapped to chromosome 5 by polymerase chain reaction. Geno- mics 4: 606-609
Aghamahammadi SZ, Savage JRK (1989) A pulse BrdU method for SCE Murat Res 216: 259-266
Anand R, Riley JH, Butler R, Smith JC, Markham AF (1990) A 3.5 genome equivalent multi-access YAC library: construction, characterisation, screening and storage. Nucleic Acids Res 18 : 1951-1956
Bohren KM, Bullock B, Wermuth B, Gabbay KH (1989) The aldo-keto reductase superfamily. J Biol Chem 264: 9547-9551
Chung S, La Mendola J (1989) Cloning and sequence determina- tion of human placental aldose reductase gene, J Biol Chem 264:14775-14777
Dean M, Park M, LeBeau MM, Robins TS, Diaz MO, Rowley JD, Blair DG, Van de Woude GF (1985) The human met onco- gene is related to the tyrosine kinase oncogenes. Nature 318: 385-388
Dionne CA, Kaplan R, Seuanez H, O'Brien S J, Jaye M (1990) Chromosome assignment by PCR techniques: assignment of the oncogene FGF-5 to human chromosome 4. Biotechniques 8:190-194

514
Duncan AMV, Buchwald M, Tsui L-C (1988) In situ hybridisation of two cloned chromosome 7 sequences tightly linked to the cystic fibrosis locus. Cytogenet Cell Genet 49 : 309-310
Elvin P, Slynn G, Black D, Graham A, Butler R, Riley JH, Anand R, Markham AF (1990) Isolation of cDNA clones using yeast artificial chromosome probes. Nucleic Acids Res 18 : 3913-3917
Graham A, Hedge PJ, Powell S J, Riley J, Brown L, Gammack AJ, Carey F, Markham AF (1989) Nucleotide sequence of cDNA for human aldose reductase. Nucleic Acids Res 17 : 8368
Graham A, Brown L, Hedge P J, Gammack AJ, Markham AF (1991) Structure of the human aldose reductase gene. J Biol Chem (in press)
Grundmann U, Bohn H, Obermeier R, Amann E (1990) Cloning and prokaryotic expression of a biologically active human pla- cental aldose reductase. DNA Cell Biol 9 : 149-157
ISCN (1985) An international system for human cytogenetic nomen- clature. In: Harnden DG, Klinger HP (eds) Published in col- laboration with Cytogenet Cell Genet. Kargel, Basel
McNeill C, Brown R (1980) Genetic manipulation by means of microcell-mediated transfer of normal human chromosomes into recipient mouse cells. Proc Natl Acad Sci USA 77 : 5394- 5398
Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou J-L, Drumm
ML, Iannuzzi MC, Collins FS, Tsui L-C (1989) Identification of the cystic fibrosis gene: cloning and characterisation of com- plementary DNA. Science 245 : 1066-1073
Saiki RK, Scharf S, Faloona F, Mullis KB, Horn GT, Erlich HA, Arnheim N (1985) Enzymatic amplification of ~-globin geno- mic sequences and restriction site analysis for diagnosis of sickle cell anaemia. Science 230:1350-1354
Saxon PJ, Srivatsan ES, Leipzig GV, Sameshima JG, Stanbridge EJ (1985) Selective transfer of individual human chromosomes to recipient cells. Mol Cell Biol 5 : 140-146
Saxon PJ, Srivatsan ES, Stanbridge EJ (1986) Introduction of hu- man chromosome 11 via microeell transfer controls tumoro- genic expression of Hela cells. EMBO J 5 : 3461-3466
Van Steeg H, Van Oostrom CTM, Martens JWM, Van Kreyl CF, Schepens J, Wieringa B (1989) Nucleotide sequence of the human ornithine decarboxylase gene. Nucleic Acids Res 17: 8855-8856
Yang-Feng TL, Opdenakker G, Volckaert G, Francke U (1985) Mapping of the human tissue-type plasminogen activator (PLAT) gene to chromosome 8 (Sp12-q11.2). Cytogenet Cell Genet 40: 784