Embed Size (px)
Citation preview

1
PERTUMBUHAN, KEKUATAN TARIK DAN MULUR SERAT RAMI (Boehmeria nivea (L.) Gaudich) DENGAN PEMBERIAN ASAM
GIBERELAT (GA3) DAN VARIASI KETERSEDIAAN AIR
THE GROWTH, PULLING AND ELASTICITY STRENGTH OF HEMP FIBER Boehmeria nivea (L.) Gaudich WITH APPLICATION OF
GIBERELIC ACID GA3 AND WATER AVAILABILITY
Shafi Fauzi Rahman, Widya Mudyantini, M.Si., Dra. Endang Anggarwulan, M.Si. Jurusan Biologi FMIPA UNS Surakarta
Hemp plant Boehmeria nivea is an annual plant which is easy to grow and to reproduce in a tropical region. Hemp fiber has a higher strength than cotton fiber, so that it is not easily broken off. It provides less reduction than another fibers, the humidity of hemp fiber can achieve 12%, and hemp fiber has smooth characteristic, long lasting, and its glint is similar with the silk. This research used complete random design CRD using two factors that were GA3 with 3 concentration variations G, such as 0 ppm, 175 ppm, 200 ppm, and 3 water availability variations A), such as 50%, 75%, 100%. The treatment was given to the rhizome before it was planted and the water availability was given when the shoot was started to form. The measured parameters were parameter of growth and fiber quality. The conclusion of this research is that GA3 treatment has an influence toward the increase of shoot stem height, dry weight, wet weight, fiber pulling test fiber’s strength, but it has no influence in the change of shoot number, leaf number, and elasticity of the fiber. The water availability treatment has no influence toward the entire parameter. The interaction between GA3 and water availability has influence toward hemp B. nivea fiber elasticity. The giving of GA3 in the concentration of 200 ppm shows the best influence toward the entire parameter of growth and fiber quality observed except in wet weight and dry weight. Water availability treatment in SQ 100% shows good influence toward wet weight and dry weight, in SQ 75% it shows a good influence toward the elasticity of the fiber. Key words: Giberelat Acid GA3, water availability, Boehmeria nivea, growth,
pulling and elasticity test.

2
PENDAHULUAN
Indonesia memiliki industri serat yang terdiri dari industri serat alam, serat
buatan dan benang filamen; dan industri pemintalan serta pencelupan (spinning).
Saat ini Indonesia merupakan produsen serat buatan ke tujuh terbesar dunia yang
memasok 10% kebutuhan serat rayon dunia. Sekitar separuh dari produksi industri
pemintalan dikonsumsi di dalam negeri, dan sisanya di ekspor ke luar negeri
(Miranti, 2007). Penggunaan serat rami di Indonesia saat ini masih sebatas sebagai
campuran serat kapas pada industri tekstil dan produk tekstil (IT-PT). Panjang
serat rami disesuaikan dengan panjang serat kapas dengan cara dipotong-potong
terlebih dahulu, karena sebagai suplemen kebutuhannya belum begitu besar yaitu
sekitar 11 ton per tahun dan hampir seluruhnya dipenuhi dari impor asal China
(Deptan, 2007).
Tanaman rami (B. nivea) merupakan tanaman tahunan yang mudah
tumbuh dan berkembang baik di daerah tropis. Serat rami merupakan bahan yang
dapat diolah untuk kain fashion berkualitas tinggi dan bahan pembuatan selulosa
berkualitas tinggi (selulose α) (Tarmansyah, 2007). Serat rami mempunyai
kekuatan yang lebih tinggi dibanding dengan serat kapas, sehingga tidak mudah
putus. Kekurangan serat rami pada elastisitasnya yang lebih rendah dan kurang
fleksibel jika dibandingkan dengan serat kapas (Gossypium sp). Keunggulan
pakaian yang dibuat dari kain berbahan serat rami antara lain kemampuan
menyerap air tinggi dan mudah dicuci (Saroso dan Sastrosupadi, 2000; Brink dan
Escobin, 2003). Menurut Hill (1972), serat rami bersifat halus, tahan lama dan
kilatannya seperti sutera. Salah satu upaya untuk mengurangi ketergantungan pada
kapas adalah penggunaan serat alami yang berasal dari tanaman rami (B. nivea)
yang memiliki karakteristik mirip kapas dan dapat digunakan sebagai bahan baku
tekstil (Buxton dan Greenhalg, 1989).
Pertumbuhan dan perkembangan tanaman dikendalikan oleh zat pengatur
tumbuh, contohnya adalah Gibberelic acid (GA3) (Kastono, 2005). Menurut
Kusumo (1990), giberelin berperan dalam pembelahan sel dan mendukung
pembentukan RNA sehingga terjadi sintesis protein. Salah satu aspek yang sangat
penting dalam budidaya tanaman adalah air karena air berfungsi sebagai pelarut

3
hara tumbuhan di dalam tanah dan berperan dalam translokasi hara dan fotosintat
di dalam tubuh tumbuhan (Gardner et al., 1991). Tersedianya air dalam jumlah
yang tepat akan mendukung pertumbuhan tanaman, sebaliknya bila air terlalu
berlebih atau kurang, pertumbuhan tanaman juga akan terhambat sehingga hasil
panen yang didapatkan tidak optimal (Levitt, 1980).
Kekuatan tarik serat mengindikasikan besarnya kekuatan serat yang dapat
mendukung sebelum putus, kekuatan mulur serat didefinisikan sebagai
panjangnya serat yang dapat mulur sebelum putus (Lee, 1999 dalam Indrawan,
2007). Pemberian GA3 dan ketersediaan air yang tepat dapat meningkatkan
kualitas serat. Selulosa dan lignin sebagai penyusun dinding sel akan meningkat
jumlahnya seiring peningkatan jumlah floemnya karena pemberian GA3. Selulosa
merupakan penentu kualitas serat, sedangkan lignin menambah ketahanan serat.
Dengan adanya perlakuan GA3 dan ketersediaan air dalam penelitian ini
maka diharapkan dapat meningkatkan uji tarik yang cukup tinggi dan kemuluran
yang cukup baik, serta serat yang dihasilkan dari pemungutan hasil yang pertama
kali dari tanaman rami tersebut akan memiliki kualitas yang lebih tinggi, sehingga
dapat meningkatkan efisiensi waktu tunggu panen.
BAHAN DAN METODE
Bahan yang Digunakan
Bahan-bahan yang digunakan dalam penelitian ini adalah rhizoma rami;
air untuk mencuci dan menyiram media; media tanam yang berupa campuran
tanah, pasir, dan pupuk kandang; alkohol; GA3: 175 ppm, 200 ppm. Bahan untuk
uji tarik dan mulur berupa serat rami yang telah terpisahkan per helai.
Persiapan Media
Media dipersiapkan dengan mencampur tanah, pasir dan pupuk kandang
dengan perbandingan 1:1:1. Campuran media ditimbang untuk masing-masing
polibag ½ kg.
Persiapan dan penanaman Rimpang Rami
Dalam penelitian ini rhizoma rami dipilih yang seragam kemudian
dipotong-potong sepanjang 10 cm, dengan tiap-tiap rhizoma memiliki 1 mata

4
tunas. Potongan rhizoma tersebut kemudian ditanam pada media di dalam polibag
sedalam 5 cm dengan posisi agak miring kemudian disiram air.
Perlakuan Pemberian GA3
Pemberian GA3 dilakukan sekali sebelum penanaman. Masing-masing
rhizoma disemprot dengan hormon sebanyak 5 ml. Setelah penyemprotan,
tanaman langsung disimpan di tempat yang gelap dan tertutup sebelum ditanam
dalam polibag agar hormon tidak rusak terkena cahaya dan tidak menguap.
Penanaman dilakukan 2 hari setelah perlakuan (Mudyantini, 2008).
Penentuan kapasitas lapang
Campuran media tanam yang telah dikeringanginkan ditimbang seberat 1/2
kg dalam polibag yang telah dilubangi bagian bawahnya. Kemudian disiram
dengan air sampai air berhenti menetes dari lubang bawah polibag sehingga dapat
diketahui volume air yang digunakan untuk menyiram dan kapasitas lapangnya.
Kapasitas lapang dihitung dengan menggunakan rumus sebagai berikut:
KL = (Berat tanah + polibag + air) – (Berat tanah + polibag) (Patoni, 2000).
Pemeliharaan
Pemeliharaan dilakukan dengan penyiraman setiap 1 hari sekali dengan
berbagai variasi ketersediaan air yaitu 50%; 75%; 100% kapasitas lapang.
Pengamatan Pertumbuhan
Penghitungan jumlah tunas, tinggi batang tunas, dan jumlah daun
dilakukan setiap 1 minggu sekali dimulai hari ke-0 semenjak perlakuan selama 2
bulan. Berat basah tanaman diukur dengan penimbangan semua tunas yang
muncul pada masing-masing rhizoma pada akhir perlakuan. Berat kering tanaman
diukur dengan mengeringkan semua tunas yang muncul pada masing-masing
rhizoma dengan cara dikeringanginkan dibawah sinar matahari sampai kering
kemudian ditimbang.
Pengujian Kekuatan Tarik dan Mulur Serat
Serat rami dipisahkan perhelai dengan panjang ± 10 cm. Dibuat media
berupa kertas karton (kertas tebal) dengan panjang 10 cm, lebar 2 cm. Pada bagian
tengah kertas dilubangi berbentuk persegi panjang, dengan panjang 5 cm dan
lebar 1 cm. Serat yang telah dipisahkan perhelai ditempel ditengah media kertas

5
berlubang. Direkatkan ujung-ujung serat dengan isolatip dan lem agar menempel
pada media, kemudian diujikan di alat Tenso Lab. yang secara otomatis akan
menunjukkan angka kekuatan tarik dan mulurnya dalam nilai statistik
(Laboratorium Evaluasi Tekstil, 2008).
Analisis Data
Data kuantitatif yang diperoleh diuji dengan analisis sidik ragam (Anava)
untuk perlakuan awal/satu perlakuan; GA3, (Ancova) untuk perlakuan
berkelanjutan; variasi ketersediaan air, dan General Linier Model (GLM)
Univariate untuk 2 perlakuan; pada analisis kualitas serat. Untuk mengetahui beda
nyata di antara perlakuan, dilanjutkan dengan uji Duncans Multiple Range Test
(DMRT) pada taraf uji 5%.
HASIL DAN PEMBAHASAN
A. Pertumbuhan Tanaman
Jumlah Tunas
Hasil rerata jumlah tunas B. nivea dengan perlakuan GA3 disajikan pada
Tabel 1.
Tabel 1. Rerata jumlah tunas B. nivea dengan perlakuan GA3 berumur 32 hari setelah tanam.
Perlakuan GA3
Jumlah tunas
G0
G175
G200
4
2
1
Keterangan:
G = konsentrasi GA3(ppm), G0=0, G175=175, G200=200.

6
Dari hasil analisis sidik ragam (Anava) diketahui bahwa perlakuan GA3
tidak berpengaruh terhadap perubahan jumlah tunas yang muncul. Jumlah tunas
tertinggi B. nivea pada Tabel 1 terdapat pada perlakuan G0 (kontrol) dengan rerata
sebesar 4 buah, sedangkan jumlah hasil tunas terendah diperoleh pada perlakuan
G175 dan G200 yaitu 2 dan 1 buah. Hasil ini lebih rendah jika dibandingkan dengan
kontrol. Hal ini menunjukkan bahwa setiap tumbuhan membutuhkan konsentrasi
yang sesuai untuk pertumbuhannya. Konsentrasi yang tidak sesuai tidak akan
memacu pertumbuhan, tetapi justru menghambat pertumbuhan. Menurut
penelitian Rahman, et al., (2006) pemberian GA3 pada konsentrasi 250 ppm
mendorong pertumbuhan Allium sativum 31,67%, sedangkan pada konsentrasi
500 ppm hanya 10,00%.
Adanya pengaruh yang tidak signifikan tersebut diduga disebabkan karena
jumlah tunas yang muncul ditentukan oleh jumlah mata tunas yang sudah ada
pada rhizoma. Jumlah mata tunas pada setiap potongan rhizoma ditentukan oleh
jarak antar ruas-ruas pada rhizoma dan merupakan faktor internal dari tanaman
rami itu sendiri. Oleh karena itu, antara potongan rhizoma yang satu dengan yang
lain dengan panjang yang sama dapat mempunyai jumlah mata tunas yang
berbeda, meskipun mata tunas yang sudah muncul telah dipotong sebelum
perlakuan. Selain itu, menurut Wahid (1990) dalam Hidayanto dkk., (2003),
kandungan karbohidrat yang terdapat pada bahan stek, yaitu rhizoma, merupakan
faktor utama untuk perkembangan primordia tunas dan akar.
Tabel 2. Rerata jumlah tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan air berumur 2 bulan setelah tanam.
Ketersediaan Air Perlakuan GA3 A1 A2 A3
Rerata
G0 - - 5,67 5,67 G1 2,67 5,33 6,00 4,67 G2 3,33 3,00 4,33 3,56 Rerata 3,00 4,17 5,33
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.

7
Dari hasil analisis sidik ragam (Ancova) pada Tabel 2 diketahui bahwa
pada perlakuan GA3 tidak memberikan pengaruh yang signifikan begitu juga
dengan perlakuan ketersediaan air juga tidak memberikan pengaruh yang
signifikan terhadap jumlah tunas B. nivea. Rerata jumlah tunas tertinggi terdapat
pada perlakuan G1A3 dan G0A3 (kontrol) yang memiliki jumlah tunas sebesar 6
buah, sedangkan rerata terendah terdapat pada perlakuan G1A1, G2A1 dan G2A2
yang memiliki jumlah tunas sebesar 3 buah. Rerata peningkatan jumlah tunas
setiap 1 minggu sekali disajikan dalam Tabel 3.
Tabel 3. Rerata peningkatan jumlah tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan air setiap 1 minggu sekali (cm).
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200 Ketersediaan Air (%), A1=50, A2=75, A3=100
Jumlah tunas dihitung setiap 1 minggu sekali. Tabel peningkatan jumlah
tunas B. nivea menunjukkan pertumbuhan yang meningkat setiap minggunya.
Mulai minggu ke-7 ditunjukkan adanya penurunan pada G1A2, G2A1 dan G2A3,
pada minggu ke-8 ditunjukkan adanya peningkatan pada G2A2. Perbandingan
peningkatan jumlah tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan
air ditunjukkan pada Gambar 1.
Rerata jumlah tunas minggu ke - Perlakuan GA3 dan
Air 1 2 3 4 5 6 7 8 9
G0A3 G1A1 G1A2 G1A3 G2A1 G2A2 G2A3
0 1 0 0 0 0 0
1 4 6 6 3 4
10
3 3 6 6 2 4 9
4 2 5 5 2 3 8
4 2 4 4 2 2 5
6 2 4 3 3 2 5
6 3 6 6 4 2 5
6 3 5 6 3 2 5
6 3 5 6 3 3 4

8
Gambar 1. Rerata peningkatan jumlah tunas B. nivea dengan perlakuan GA3 dan
variasi ketersediaan air setiap 1 minggu sekali. Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Konsentrasi GA3 yang diberikan dalam penelitian ini meliputi 0 ppm, 175
ppm dan 200 ppm. Dari ketiga perlakuan jumlah tunas tertinggi B. nivea di
hasilkan pada konsentrasi 0 ppm, sedangkan jumlah tunas terendah pada
konsentrasi 200 ppm. Hal ini menunjukkan bahwa setiap tumbuhan membutuhkan
konsentrasi yang sesuai untuk pertumbuhannya. Konsentrasi yang tidak sesuai
tidak akan memacu pertumbuhan, tetapi justru menghambat pertumbuhan.
Menurut Wareing dan Phillips (1981), pemberian senyawa IAA pada
konsentrasi yang optimum akan menyebabkan pembelahan sel yang bersifat
meristematis, sehingga akan menyebabkan jumlah tunas lebih cepat keluar.
Namun, pemberian GA3 pada rhizoma rami tidak dapat mempengaruhi perubahan
jumlah tunas yang dihasilkan karena tidak dapat meningkatkan jumlah mata tunas
yang ada pada rhizoma. GA3 lebih dominan dalam merangsang pemanjangan sel
dan IAA lebih dominan dalam merangsang pembesaran sel (Davies, 1995).
Variasi ketersediaan air yang diberikan pada penelitian ini meliputi 50%,
75% dan 100% KL. Jumlah tunas tertinggi B. nivea dihasilkan pada perlakuan
100% KL, sedangkan jumlah tunas terendah B. nivea pada perlakuan 50% KL
(Tabel 2).

9
Ketersediaan air yang semakin meningkat menyebabkan semakin
meningkatnya jumlah tunas tanaman, dan bila ketersediaan air menurun maka
jumlah tunas akan menurun. Fitter dan Hay (1998) menyatakan bahwa air
berpengaruh terhadap pertumbuhan sel, semakin menurun ketersediaan air maka
tekanan turgor akan menurun, hal ini menyebabkan menurunnya laju
pertumbuhan.
Cekaman air menyebabkan perubahan macam dan jumlah senyawa
karbohidrat di dalam tanaman. Pada tanaman yang mengalami cekaman air terjadi
penurunan tepung dan peningkatan kadar gula. Penelitian Kramer (1977) dalam
Islami dan Wani (1995), menunjukkan bahwa penurunan kadar tepung tidak selalu
diikuti kenaikan kadar gula. Bahkan pada tanaman buncis (Phaseolus sp) dan
tanaman tomat (Lycopersicon sp) cekaman air yang terjadi berkelanjutan
menurunkan kadar tepung, gula dan karbohidrat total pada buncis (Phaseolus sp)
dan tomat (Lycopersicon sp). Adanya pengaruh cekaman air terhadap
metabolisme karbohidrat dan nitrogen, dapat menghambat pembentukan auksin
pada tanaman yang menderita cekaman air. Kegiatan tersebut diikuti oleh
penurunan transpor auksin ke kambium sehingga terjadi modifikasi aktivitas
kambium. Cekaman air juga menyebabkan penurunan aktivitas sitokinin dan
penyediaan giberelin ke batang (Islami dan Wani, 1995). Menurut Mullet dan
Whitsitt (1996), efek utama dari kekurangan air adalah laju pertumbuhan batang
yang lebih rendah akibat akumulasi asam absisat (ABA).

10
Panjang Tunas
Hasil rerata panjang tunas tanaman rami dengan perlakuan GA3 disajikan
pada Tabel 4.
Tabel 4. Rerata panjang tunas B. nivea dengan perlakuan GA3 berumur 32 hari setelah tanam (cm).
Perlakuan GA3
Panjang tunas (cm)
G0
G175
G200
6,12a
18,13ab
22,92b
Keterangan: G= konsentrasi GA3(ppm), G0=0, G175=175, G200=200. Angka yang diikuti huruf sama pada kolom yang sama
menunjukkan tidak ada beda nyata pada uji DMRT pada taraf 5%.
Dari hasil analisis sidik ragam (Anava) menunjukkan bahwa perlakuan
GA3 berpengaruh nyata terhadap panjang tunas tanaman B. nivea. Rerata panjang
tunas selalu meningkat seiring dengan meningkatnya konsentrasi GA3.
Pertumbuhan panjang tunas tersebut antara lain dipercepat oleh penggunaan
hormon GA3 yang sesuai. Hasil ini sesuai dengan pendapat Sumiasri dan Priadi
(2003) yang menyatakan bahwa pada pertumbuhan stek cabang sungkai
(Peronema canescens Jack) pada konsentrasi GA3 5 mg/l optimum meningkatkan
tinggi tunas sungkai.
Berdasarkan hasil penelitian ini rerata panjang tunas tertinggi diperoleh
pada perlakuan G200 yaitu 22,92 cm dan rerata panjang tunas terendah adalah pada
G0 (kontrol) yaitu 6,12 cm. Hal ini menunjukkan bahwa setiap tumbuhan
membutuhkan konsentrasi GA3 yang sesuai untuk pertumbuhannya. Konsentrasi
GA3 yang tidak sesuai tidak akan memacu pertumbuhan, tetapi justru dapat
menghambat pertumbuhan. Hal ini sesuai dengan Salisbury dan Ross (1995) yang
menyatakan bahwa zat tumbuh aktif pada konsentrasi rendah dan sampai batas
tertentu akan merangsang pertumbuhan. Menurut Gul (2006) bahwa pemberian

11
hormon GA3 300 ppm pada Araucaria heterophylla, berpengaruh pada tinggi
tanaman yang maksimum. Aisyah (2004) juga menyatakan bahwa pemberian GA3
pada Allium cepa dengan perendaman cenderung semakin meningkatkan tinggi
tanaman seiring dengan meningkatnya konsentrasi GA3 hingga 10 ppm. Namun
pada konsentrasi di bawah maupun di atasnya justru semakin rendah.
Respon yang khas pada tanaman dengan perlakuan GA3 adalah terjadinya
pemanjangan batang akibat adanya aktivitas kambium di internodus; sehingga
tanaman menjadi lebih tinggi dari pada tanaman normal. Pemanjangan batang
selain di pengaruhi oleh aktivitas kambium juga disebabkan oleh peningkatan
mitosis di daerah meristem subapikal batang, sehingga jumlah sel pada masing-
masing internodus meningkat. Peningkatan jumlah sel menyebabkan pertumbuhan
batang lebih cepat, sehingga dihasilkan batang yang lebih panjang. Respon ini
pada batang biasanya hanya berupa peningkatan panjang internodus, dan
umumnya tidak meningkatkan jumlah internodus yang terbentuk (Wareing dan
Phillips, 1981).
Tabel 5. Rerata panjang tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan air berumur 2 bulan setelah tanam (cm).
Ketersediaan Air Perlakuan GA3 A1 A2 A3
Rerata
G0 - - 7,48 7,48 G1 16,35 9,87 4,89 10,37 G2 8,04 10,99 10,97 10,00 Rerata 12,20 10,43 7,78
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Dari hasil analisis sidik ragam (Ancova) pada Tabel 5 diketahui bahwa
pada perlakuan GA3 tidak memberikan pengaruh yang signifikan begitu juga
dengan perlakuan ketersediaan air juga tidak memberikan pengaruh yang
signifikan terhadap panjang tunas B. nivea. Panjang tunas tertinggi terdapat pada
perlakuan G1A1 yang memiliki rerata sebesar 16,35 cm, sedangkan rerata terendah
terdapat pada perlakuan G1A3 yang memiliki panjang tunas sebesar 4,89 cm. Hasil
12
ini lebih rendah jika dibandingkan dengan G0A3 dengan rata-rata panjang tunas
sebesar 7,48 cm. Hal ini diduga berkaitan erat dengan proses pemanjangan sel
yang menurun akibat adanya cekaman air.
Variasi ketersediaan air yang diberikan pada penelitian ini meliputi 50%,
75% dan 100% KL. Panjang tunas tertinggi B. nivea dihasilkan pada perlakuan
50% KL, sedangkan panjang tunas terendah B. nivea pada perlakuan 100% KL.
Pertumbuhan sel merupakan fungsi tanaman yang sensitif terhadap kekurangan
air. Nilai potensial air jaringan meristem pada siang hari sering menyebabkan
penurunan tekanan turgor dibawah yang dibutuhkan untuk pengembangan sel. Hal
ini menyebabkan pengurangan sintesis protein, dinding sel dan pengembangan sel
yang berakibat pertumbuhan yang lebih kecil (Gardner, et al., 1991). Berdasarkan
penelitian Dewi (1993) terhadap dua kultivar tanaman kedelai (Glycine max (L.))
Merry Willis dan Lompo Batang, setelah berumur 47 hari pada stress air yang
paling besar menyebabkan tinggi tanaman mengalami penurunan hampir 50% dan
diameter batang mengalami pengurangan 47,7% untuk Willis dan 42,14% untuk
Lompo Batang. Menurut Anggarwulan dkk., (2008) bahwa perlakuan ketersediaan
air 60% memberikan pertumbuhan kimpul (Xanthosoma sagittifolium) yang
terbaik pada semua taraf naungan. Rerata peningkatan panjang tunas setiap 1
minggu sekali disajikan dalam Tabel 6.
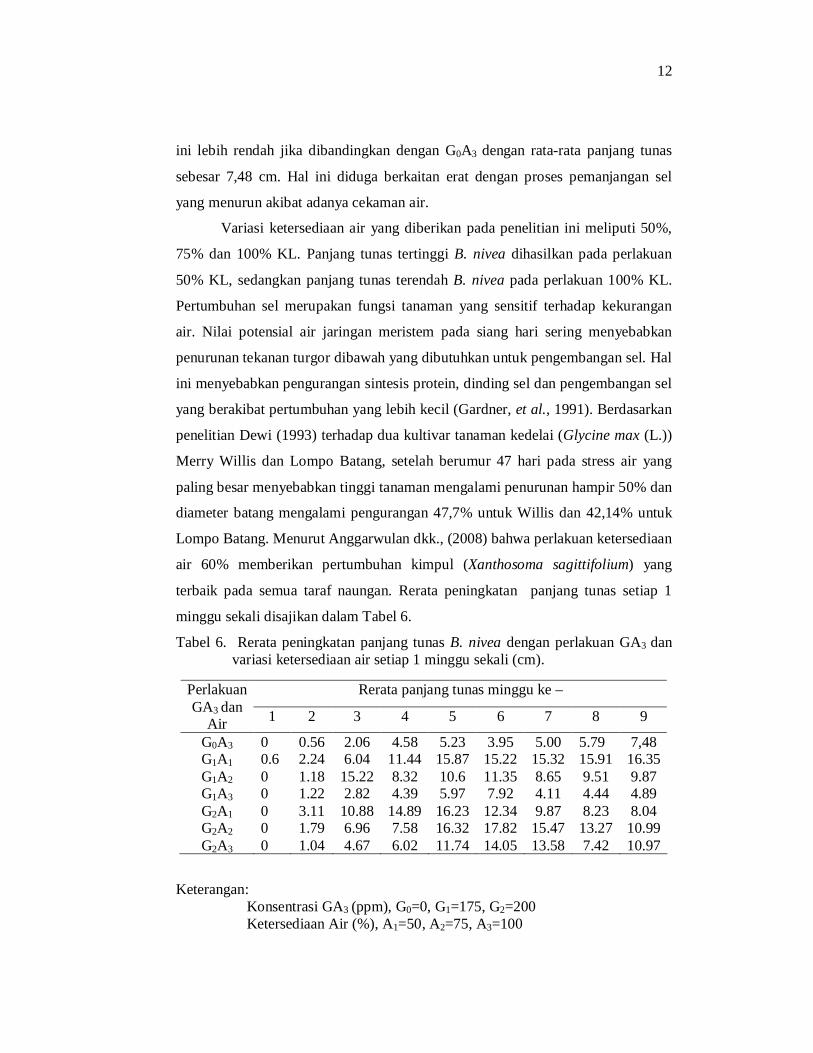
Tabel 6. Rerata peningkatan panjang tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan air setiap 1 minggu sekali (cm).
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200 Ketersediaan Air (%), A1=50, A2=75, A3=100
Rerata panjang tunas minggu ke – Perlakuan GA3 dan
Air 1 2 3 4 5 6 7 8 9
G0A3 0 0.56 2.06 4.58 5.23 3.95 5.00 5.79 7,48 G1A1 0.6 2.24 6.04 11.44 15.87 15.22 15.32 15.91 16.35 G1A2 0 1.18 15.22 8.32 10.6 11.35 8.65 9.51 9.87 G1A3 0 1.22 2.82 4.39 5.97 7.92 4.11 4.44 4.89 G2A1 0 3.11 10.88 14.89 16.23 12.34 9.87 8.23 8.04 G2A2 0 1.79 6.96 7.58 16.32 17.82 15.47 13.27 10.99 G2A3 0 1.04 4.67 6.02 11.74 14.05 13.58 7.42 10.97
13
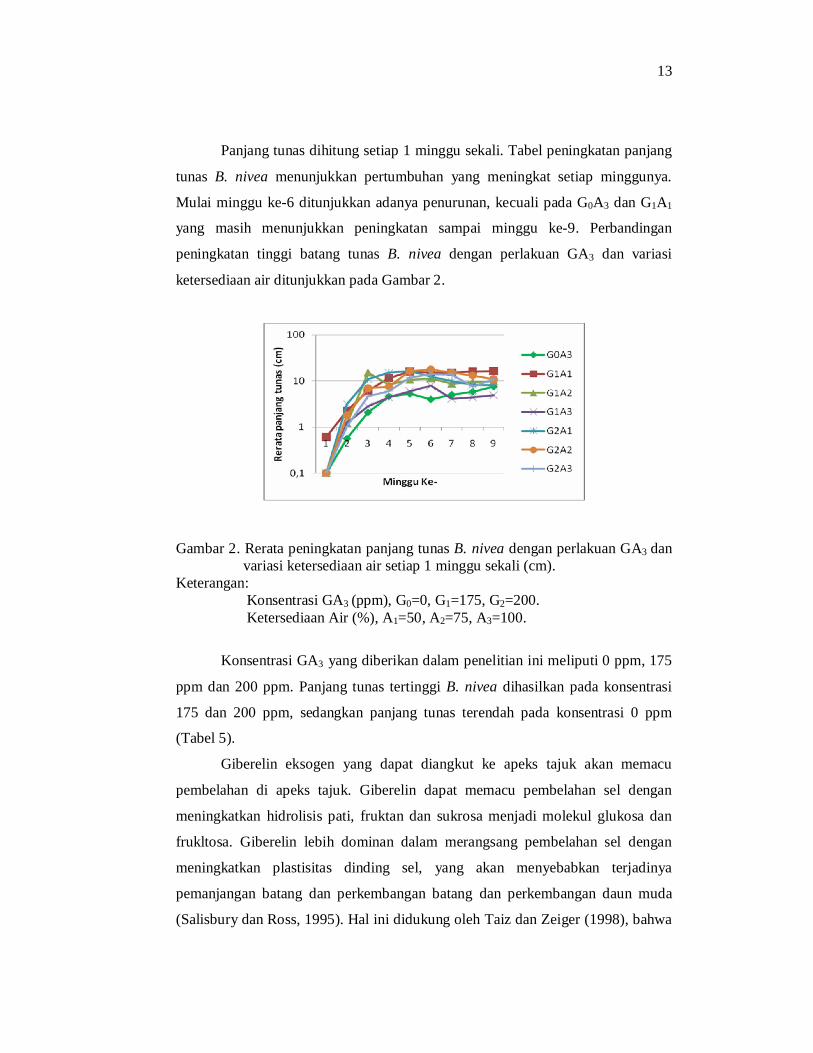
Panjang tunas dihitung setiap 1 minggu sekali. Tabel peningkatan panjang
tunas B. nivea menunjukkan pertumbuhan yang meningkat setiap minggunya.
Mulai minggu ke-6 ditunjukkan adanya penurunan, kecuali pada G0A3 dan G1A1
yang masih menunjukkan peningkatan sampai minggu ke-9. Perbandingan
peningkatan tinggi batang tunas B. nivea dengan perlakuan GA3 dan variasi
ketersediaan air ditunjukkan pada Gambar 2.
Gambar 2. Rerata peningkatan panjang tunas B. nivea dengan perlakuan GA3 dan variasi ketersediaan air setiap 1 minggu sekali (cm).
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Konsentrasi GA3 yang diberikan dalam penelitian ini meliputi 0 ppm, 175
ppm dan 200 ppm. Panjang tunas tertinggi B. nivea dihasilkan pada konsentrasi
175 dan 200 ppm, sedangkan panjang tunas terendah pada konsentrasi 0 ppm
(Tabel 5).
Giberelin eksogen yang dapat diangkut ke apeks tajuk akan memacu
pembelahan di apeks tajuk. Giberelin dapat memacu pembelahan sel dengan
meningkatkan hidrolisis pati, fruktan dan sukrosa menjadi molekul glukosa dan
frukltosa. Giberelin lebih dominan dalam merangsang pembelahan sel dengan
meningkatkan plastisitas dinding sel, yang akan menyebabkan terjadinya
pemanjangan batang dan perkembangan batang dan perkembangan daun muda
(Salisbury dan Ross, 1995). Hal ini didukung oleh Taiz dan Zeiger (1998), bahwa

14
GA3 mempunyai peran dalam mendukung pembelahan sel, pembentangan sel,
aktivitas kambium, pembentukan RNA, dan sintesis protein yang menyebabkan
peningkatan tinggi batang.
Peningkatan kecepatan pertumbuhan dan tinggi tanaman akibat pengaruh
GA3 dijelaskan dengan peran fisiologis zat tumbuh ini yang mempercepat
tumbuhnya tanaman dengan mendukung perkembangan dinding sel dan
merangsang pemanjangan sel karena adanya hidrolisis pati yang mendukung
terbentuknya enzim amilase, yang pada akhirnya bisa mempercepat
perkembangan sel (Wattimena, 1998). Menurut Wuryaningsih dan Sutater (1993)
bahwa pemberian GA3 25 ppm menunjukkan beda nyata terhadap tinggi batang
dan pembungaan yang lebih cepat. Hal tersebut sesuai dengan penelitian Sanjaya
(1991) bahwa pemberian GA3 dapat meningkatkan tinggi tanaman dan
berpengaruh nyata terhadap panjang tangkai bunga krisan pada konsentrasi
optimum 25 ppm yang diberikan dua kali yaitu pada umur 6 dan 8 minggu setelah
tanam.
Menurut Van Oberbeek (1966) dalam Weaver (1982), penggunaan GA3
akan mendukung pembentukan enzim proteolitik yang akan membebaskan
triptophan sebagai prekursor dari auksin. Hal ini berarti bahwa kehadiran giberelin
tersebut akan meningkatkan kandungan auksin. Mekanisme lain menyebutkan
bahwa giberelin akan menstimulasi pemanjangan sel karena adanya hidrolisis pati
yang dihasilkan dari giberelin akan mendukung terbentuknya -amilase. Sebagai
akibat dari proses tersebut, maka konsentrasi gula meningkat yang mengakibatkan
tekanan osmotik di dalam sel menjadi naik, sehingga ada kecenderungan sel
tersebut berkembang (Weaver, 1972 dalam Abidin, 1990).
Ketersediaan air yang terlalu melimpah dalam tanah menyebabkan
anoksia/berkurangnya oksigen pada daerah sekitar akar, hal ini dapat mengganggu
penyerapan hara dari tanah oleh akar tanaman (Pezehski, 1994). Menurut Suyana
dan Widijanto (2002), keberadaan air yang terlalu berlimpah dalam tanah juga
dapat berakibat tercucinya hara dalam tanah sehingga kesuburan tanah berkurang.

15
Pencucian unsur hara dari permukaan komplek adsorpsi dan larutan tanah oleh air
bersifat memiskinkan tanah.
Jumlah Daun
Hasil rerata jumlah daun tanaman rami dengan perlakuan GA3 disajikan
pada Tabel 7.
Tabel 7. Rerata jumlah daun B. nivea dengan perlakuan GA3 berumur 32 hari setelah tanam.
Perlakuan GA3
Jumlah daun
G0
G175
G200
7
5
3
Keterangan: G = konsentrasi GA3(ppm), G0=0, G175=175, G200=200.
Hasil analisis sidik ragam (Anava) menunjukkan bahwa perlakuan GA3
tidak berpengaruh nyata terhadap jumlah daun. Rerata jumlah daun tertinggi
terdapat pada perlakuan G0 (kontrol) yaitu sebanyak 7 buah, sedangkan nilai
terendah pada perlakuan G175 dan G200 yang mempunyai rerata jumlah daun 5 dan
3 buah. Hasil tersebut lebih rendah jika dibandingkan dengan kontrol. Hal ini
menunjukkan setiap tumbuhan membutuhkan konsentrasi yang sesuai untuk
pertumbuhannya. Konsentrasi yang tidak sesuai tidak akan memacu pertumbuhan,
tetapi justru menghambat pertumbuhan. Adanya peristiwa pengguguran daun juga
berpengaruh terhadap jumlah daun. Daun-daun yang sudah tua dan tidak aktif lagi
melakukan fotosintesis akan menjadi layu kemudian gugur, yang nantinya akan
mengurangi jumlah daun keseluruhan. Pada penelitian yang telah dilakukan
Aisyah (2004) melalui perendaman umbi bibit Allium cepa dengan GA3
menyatakan bahwa pada konsentrasi yang tidak sesuai tidak meningkatkan jumlah
daun bahkan cenderung menghambat karena semua hasil di bawah kontrol. Hal
tersebut dimungkinkan karena adanya persaingan unsur hara maupun hasil kerja
giberelin dengan organ reproduktif lain maupun adanya faktor genetik atau

16
lingkungan lain yang kurang sesuai. Gardner et al. (1991), menyebutkan bahwa
jumlah dan ukuran daun dipengaruhi oleh faktor genetik dan lingkungan. Posisi
daun pada tanaman dikendalikan oleh faktor genetik, posisi daun ini
mempengaruhi laju pertumbuhan daun.
GA3 diketahui dapat memacu pertumbuhan seluruh tanaman, termasuk
daun dan akar. GA3 yang diberikan dengan cara apapun di tempat yang dapat
mengangkutnya ke ujung tajuk, maka akan terjadi peningkatan pembelahan sel
dan pertumbuhan sel yang mengarah kepada pemanjangan batang dan (pada
beberapa spesies) perkembangan daun muda (Salisbury dan Ross, 1995).
Pengaruh GA3 terhadap proses pembentukan daun sesuai hasil penelitian
Anwarudin dkk., (1996), bahwa GA3 tidak mempengaruhi proses pembentukan
daun terhadap pertumbuhan manggis.
Tabel 8. Rerata jumlah daun B. nivea dengan perlakuan GA3 dan variasi ketersediaan air berumur 2 bulan setelah tanam.
Ketersediaan Air Perlakuan
GA3 A1 A2 A3 Rerata
G0 - - 29,33 29,33 G1 5,00 19,33 17,33 13,89 G2 10,33 12,33 17,00 13,22 Rerata 7,67 15,83 21,22
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200 Ketersediaan Air (%), A1=50, A2=75, A3=100
Dari hasil analisis sidik ragam (Ancova) pada Tabel 8 diketahui bahwa
perlakuan GA3 dan variasi ketersediaan air tidak memberikan pengaruh yang
signifikan terhadap jumlah daun B. nivea. Rerata jumlah daun tertinggi terdapat
pada perlakuan G0A3 (kontrol) yang memiliki jumlah daun sebesar 29 buah,
sedangkan rerata terendah terdapat pada perlakuan G1A1 yang memiliki jumlah
daun sebesar 5 buah. Rerata peningkatan jumlah daun setiap 1 minggu sekali
disajikan dalam Tabel 9.

17
Tabel 9. Rerata peningkatan jumlah daun B. nivea dengan perlakuan GA3 dan variasi ketersediaan air setiap 1 minggu sekali.
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Penghitungan jumlah daun dilakukan setiap 1 minggu sekali. Tabel
peningkatan jumlah daun B. nivea menunjukkan pertumbuhan yang meningkat
setiap minggunya. Mulai minggu ke-7 ditunjukkan adanya penurunan, kecuali
pada G0A3 yang masih menunjukkan peningkatan sampai minggu ke-9.
Perbandingan peningkatan jumlah daun B. nivea dengan perlakuan GA3 dan
variasi ketersediaan air ditunjukkan pada Gambar 3.
Gambar 3. Rerata peningkatan jumlah daun B. nivea dengan perlakuan GA3 dan variasi ketersediaan air setiap 1 minggu sekali.
Keterangan: Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Rerata jumlah daun minggu ke – Perlakuan GA3 dan
Air 1 2 3 4 5 6 7 8 9
G0A3 0 6 14 19 20 28 29 26 29 G1A1 3 23 24 23 20 22 18 9 5 G1A2 0 19 28 32 33 34 40 23 19 G1A3 0 26 37 35 27 25 23 21 17 G2A1 0 12 16 19 19 21 21 13 10 G2A2 0 15 21 21 21 22 17 11 12 G2A3 2 39 38 48 29 28 28 23 17

18
Daun sebagai alat fotosintesis akan dapat berperan secara optimal jika
didukung oleh ketersediaan air, cahaya dan unsur-unsur hara yang cukup. Air dan
hara akan diserap oleh akar. Auksin berperan dalam pembelahan sel dan diikuti
dengan pembesaran sel akan menghasilkan primordial daun yang berkembang
(Salisbury dan Ross, 1995; Loveless, 1991). Salah satu sifat GA3 adalah
mendukung pembentukan enzim proteolitik yang membebaskan triptofan sebagai
prekursor auksin sehingga kandungan kadar auksin akan meningkat (Abidin,
1990). Konsentrasi GA3 yang diberikan dalam penelitian ini meliputi 0 ppm, 175
ppm dan 200 ppm. Jumlah daun tertinggi B. nivea dihasilkan pada konsentrasi 0
ppm, sedangkan jumlah daun B. nivea terendah pada konsentrasi 200 ppm
(Tabel 8).
Jumlah daun B. nivea tertinggi dihasilkan pada perlakuan 100% KL yaitu
29 helai. Hal ini dikarenakan pada kondisi tersebut tumbuhan memiliki
ketersediaan air yang cukup selain itu peningkatan jumlah cabang juga akan
meningkatkan jumlah daun. Ketersediaan air yang cukup akan mendukung
peningkatan luas daun sehingga berhubungan dengan tingkat produksi tanaman
(Sulistyaningsih dkk., 1994).
Jumlah daun yang terendah pada perlakuan 50% KL yaitu 5 helai. Pada
kondisi ini terjadi kehilangan air (transpirasi) yang tidak diimbangi dengan
ketersediaan air yang cukup sehingga menghambat pertumbuhan tanaman.
Kecepatan absorbsi yang tidak dapat mengimbangi kehilangan air melalui proses
transpirasi akan menyebabkan cekaman air (Islami dan Wani, 1995). Menurut
Fitter dan Hay (1998), air berpengaruh terhadap pertumbuhan sel, semakin
menurun ketersediaan air maka tekanan turgor juga akan menurun. Hal ini
menyebabkan menurunnya laju pertumbuhan yaitu jumlah daun yang dihasilkan
rendah.

19
Berat Basah
Hasil rerata berat basah tanaman rami dari penelitian ini disajikan dalam
Tabel 10.
Tabel 10. Rata-rata berat basah B. nivea dengan perlakuan GA3 dan variasi ketersediaan air berumur 2 bulan setelah tanam (gr).
Ketersediaan Air Perlakuan
GA3 A1 A2 A3 Rerata
G0 - - 24,54 24,54b G1 5,84 9,56 5,48 6,96a G2 5,40 6,40 6,80 6,19a Rerata 5,61 7,98 12,27
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
* Angka yang diikuti huruf sama pada baris dan kolom yang sama menunjukkan tidak ada beda nyata pada uji DMRT pada taraf 5%.
Dari hasil analisis sidik ragam (Ancova) pada Tabel 10 diketahui bahwa
pada perlakuan GA3 memberikan pengaruh yang nyata terhadap berat basah
B. nivea, dengan nilai signifikansi sebesar 0,00. Pada perlakuan ketersediaan air
tidak memberikan pengaruh yang signifikan terhadap berat basah B. nivea. Pada
Tabel 10 hasil tertinggi ditunjukkan pada perlakuan kontrol (G0A3) sebesar 24,54
gr, sedangkan untuk perlakuan yang lain lebih rendah jika dibandingkan dengan
kontrol. Hasil terendah ditunjukkan pada perlakuan G2A1 sebesar 5,40 gr. Hal ini
menunjukkan bahwa setiap tumbuhan membutuhkan konsentrasi yang sesuai
untuk pertumbuhannya. Konsentrasi yang tidak sesuai tidak akan memacu
pertumbuhan bahkan bisa menghambat. Perbandingan berat basah B. nivea
dengan perlakuan GA3 dan variasi ketersediaan air ditunjukkan pada Gambar 4.

20
Gambar 4. Berat basah B. nivea dengan perlakuan GA3 dan variasi ketersediaan
air berumur 2 bulan setelah tanam. Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Pada Tabel 10 (Gambar 4) menunjukkan bahwa berat basah tertinggi
B. nivea dihasilkan pada GA3 konsentrasi 0 ppm, sedangkan berat basah terendah
B. nivea pada GA3 konsentrasi 200 ppm. Untuk ketersediaan air yang diberikan
dalam penelitian ini, berat basah tertimggi B. nivea dihasilkan pada perlakuan
100% KL dan berat basah terendah B. nivea pada perlakuan 50% KL. Berat basah
tumbuhan dipengaruhi oleh kadar air dalam jaringan. Adanya pembesaran sel
mengakibatkan ukuran sel yang baru lebih besar dari sel induk. Pertambahan
ukuran sel menghasilkan pertambahan ukuran jaringan, organ dan akhirnya
meningkatkan ukuran tubuh tanaman secara keseluruhan maupun berat tanaman.
Peningkatan pembelahan sel menghasilkan jumlah sel yang lebih banyak. Jumlah
sel yang meningkat, termasuk di dalam jaringan pada daun, memungkinkan
terjadinya peningkatan fotosintesis penghasil karbohidrat, yang dapat
mempengaruhi berat tanaman (Wareing dan Phillip, 1981; Salisbury Ross, 1995).
Air merupakan komponen yang penting bagi pertumbuhan tanaman.
Pertumbuhan sendiri berfungsi sebagai proses yang mengolah masukan substrat
yang sesuai untuk menghasilkan produk pertumbuhan. Substrat yang berupa
bahan organik dan unsur lain yang diserap tanaman dari lingkungan seperti
karbondioksida, unsur hara, air dan cahaya matahari diolah menjadi bahan organik

21
yang dapat diukur dengan penambahan bobot keseluruhan tanaman (Sitompul dan
Guritno, 1995).
Cekaman air akan mengakibatkan perbanyakan dan perbesaran sel
menjadi terhambat. Hal ini berkaitan dengan pengaruh tekanan turgor sel. Selain
itu, kekurangan air akan mengakibatkan metabolisme sel terganggu termasuk
proses fotosistesis. Fotosintesis akan terhambat jika air yang merupakan bahan
utama hanya tersedia dalam jumlah sedikit, sehingga hasil dari fotosintesis juga
akan berkurang. Fotosintat yang dihasilkan akan terhambat pula dalam
peredarannya ke seluruh bagian tumbuhan, yang dapat menurunkan berat tanaman
(Harjadi dan Yahya, 1988). Peningkatan jumlah cabang akan meningkatkan
jumlah daun. Jumlah daun yang meningkat menyebabkan berat basah tumbuhan
juga meningkat. Menurut Kusumo (1990), spesies yang perkembangan daunnya
berlangsung cepat dan banyak akan semakin meningkatkan laju fotosintesis yang
kemudian menghasilkan peningkatan keseluruhan tanaman.
Berat Kering
Hasil rerata berat kering tanaman rami dari penelitian ini disajikan dalam
Tabel 11.
Tabel 11. Rata-rata berat kering B. nivea dengan perlakuan GA3 dan variasi ketersediaan air berumur 2 bulan setelah tanam (gr).
Ketersediaan Air Perlakuan GA3 A1 A2 A3
Rerata
G0 - - 5,46 5,46b G1 2,22 2,50 1,86 2,19a G2 1,88 2,19 2,20 2,09a Rerata 2,05 2,34 3,18
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
* Angka yang diikuti huruf sama pada baris dan kolom yang sama menunjukkan tidak ada beda nyata pada uji DMRT pada taraf 5%.

22
Dari hasil analisis sidik ragam (Ancova) pada Tabel 11 diketahui bahwa
pada perlakuan GA3 memberikan pengaruh yang nyata terhadap berat basah B.
nivea, dengan nilai signifikansi sebesar 0,00. Pada perlakuan ketersediaan air
tidak memberikan pengaruh yang signifikan terhadap berat basah B. nivea.
Berat kering mencerminkan akumulasi senyawa organik yang berhasil
disintesis tanaman dari senyawa anorganik, terutama air dan CO2. Tumbuhan
dapat memanfaatkan intensitas sinar matahari secara baik sehingga meningkatkan
pembentukan karbohidrat yang digunakan untuk pertumbuhan. Ketersediaan air
yang melimpah dan unsur hara yang diserap akan memberi kontribusi terhadap
pertambahan berat kering tumbuhan. Pada Tabel 11 hasil tertinggi ditunjukkan
pada perlakuan kontrol (G0A3) sebesar 5,46 gr, sedangkan untuk perlakuan yang
lain lebih rendah jika dibandingkan dengan kontrol. Hasil terendah ditunjukkan
pada perlakuan G1A3 sebesar 1,86 gr. Hal ini menunjukkan bahwa setiap
tumbuhan membutuhkan konsentrasi yang sesuai untuk pertumbuhannya.
Konsentrasi yang tidak sesuai tidak akan memacu pertumbuhan bahkan bisa
menghambat.
Peningkatan berat kering terjadi sebagai akibat bertambahnya protoplasma
yang terjadi karena ukuran dan jumlah sel bertambah. Pertambahan protoplasma
berlangsung melalui perubahan air, karbondioksida dan garam anorganik menjadi
bahan hidup. Proses ini meliputi fotosintesis, absorbsi dan metabolisme yang
menghasilkan karbohidrat sehingga meningkatkan berat kering tanaman (Harjadi,
1993; Lakitan, 1996). Perbandingan berat kering B. nivea dengan perlakuan GA3
dan variasi ketersediaan air ditunjukkan pada Gambar 5.

23
Gambar 5. Berat kering B. nivea dengan perlakuan GA3 dan variasi ketersediaan
air berumur 2 bulan setelah tanam. Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
Pada Tabel 11 (Gambar 5) menunjukkan bahwa berat kering tertinggi
B. nivea dihasilkan pada GA3 konsentrasi 0 ppm, sedangkan berat kering terendah
B. nivea pada GA3 konsentrasi 200 ppm. Untuk ketersediaan air yang diberikan
dalam penelitian ini, berat kering tertimggi B. nivea dihasilkan pada perlakuan
100% KL dan berat kering terendah B. nivea pada perlakuan 50% KL.
Menurut Delvin dan Withan (1983) dalam Rahardjo dkk., (1999) berat
kering tanaman dapat dipakai sebagai petunjuk seberapa besar tanggapan tanaman
terhadap cekaman air, karena air merupakan faktor pembatas utama bagi
pertumbuhan tanaman. Gardner et al. (1991) menyebutkan kekurangan air yang
parah dapat menyebabkan penutupan stomata, sehingga mengurangi pengambilan
karbondioksida, akibatnya pertumbuhan terhambat dan produksi berat kering
berkurang.
Menurut Fitter dan Hay (1998), bahwa air memberi pengaruh terhadap
berat kering, hal ini berkaitan dengan proses metabolisme yaitu proses
fotosintesis. Berat kering total hasil panen tanaman budidaya merupakan akibat
dari penimbunan hasil bersih asimilasi CO2 sepanjang musim pertumbuhan.
Pemanfaatan hasil fotosintesis oleh tanaman antara lain untuk pembentukan
struktur tubuh dan cadangan makanan. Fotosintesis menambat CO2 untuk

24
produksi heksosa dan respirasi. Cekaman/stress air dapat mengurangi laju
fotosintesis yang lambat laun juga akan mengurangi sintesis/pembentukan struktur
tubuh dan cadangan makanan sehingga mengurangi berat kering. Meskipun air
merupakan salah satu bahan baku dalam proses fotosintesis, namun pengaruh dari
pengurangan air dalam daun terhadap kecepatan fotosintesis umumnya terjadi
secara tidak langsung. Pengaruh kadar air dalam tanah akan menyebabkan
pengurangan dalam kecepatan fotosintesis, hal ini disebabkan karena;
berkurangnya kapasitas difusi dari stomata karena stomata menutup, penurunan
hidrasi dari kloroplas dan bagian-bagian lain dari protoplasma sehingga
mengurangi efektifitas mekanisme fotosintesis, terjadi akumulasi gula sehingga
menghambat proses fotosintesis lebih lanjut (Haddy, 1987). Menurut Fitter dan
Hay (1998), menutupnya stomata, mengakibatkan difusi CO2 dari atmosfer ke
daun terhenti. Sebagai akibatnya fotosintesis tidak dapat terjadi, dan dalam jangka
panjang akan mengganggu proses-proses fisiologi lainnya sehingga pertumbuhan
tanaman terhambat.
B. Kualitas Serat
Kekuatan Tarik Serat
Hasil rerata kekuatan tarik serat tanaman rami dengan perlakuan GA3 dan
ketersediaan air disajikan pada Tabel 12.
Tabel 12. Rerata kekuatan tarik serat B. nivea dengan perlakuan GA3 dan ketersediaan air berumur 2 bulan setelah tanam (gr).
Ketersediaan Air Perlakuan
GA3 A1 A2 A3 Rerata
G0 - - 58 58a G1 102 148 82 110a G2 326 178 180 228b Rerata 214 163 106
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
* Angka yang diikuti huruf sama pada baris dan kolom yang sama menunjukkan tidak ada beda nyata pada uji DMRT pada taraf 5%.

25
Dari analisis sidik ragam General Linier Model (GLM) menunjukkan
bahwa perlakuan GA3 memberikan pengaruh yang nyata terhadap kekuatan tarik
serat, dengan nilai signifikansi sebesar 0,008. Perlakuan ketersediaan air, maupun
interaksi antara GA3 dan pemberian ketersediaan air tidak memberikan pengaruh
yang nyata terhadap kekuatan tarik serat. Perbandingan kekuatan tarik serat
B. nivea dengan perlakuan GA3 dan variasi ketersediaan air ditunjukkan pada
Gambar 6.
Gambar 6. Kekuatan tarik serat B. nivea dengan perlakuan GA3 dan perlakuan
ketersediaan air berumur 2 bulan setelah tanam. Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200 Ketersediaan Air (%), A1=50, A2=75, A3=100
Gambar 6 menunjukkan bahwa kekuatan tarik serat paling tinggi ada pada
perlakuan GA3 200 ppm dan ketersediaan air 50% (G2A1) yaitu sebesar 326,
sedangkan kekuatan tarik terendah ada pada perlakuan GA3 0 ppm dan
ketersediaan air 100% (G0A3/kontrol) yaitu sebesar 58. Pada Gambar 6 dapat
dilihat bahwa semakin tinggi konsentrasi pemberian GA3 maka hasil kekuatan
tarik serat akan semakin tinggi.
Penunjang kekuatan serat yaitu selulosa dan lignin. Semakin tinggi kadar
selulosa dan lignin, maka kekuatan serat yang dihasilkan akan semakin baik.
Tetapi selulosa sebagai penyusun utama dinding sel lebih berperan memberi
kekuatan pada serat. Salah satu sifat penting selulosa adalah kemampuannya
bertahan terhadap regangan karena kelenturannya. Lignin menambah ketahanan
dinding terhadap tekanan dan mencegah melipatnya mikrofibril selulosa. Arah

26
mikrofibril yang berbeda-beda merupakan faktor penting penentu kekuatan
dinding (Mudyantini dkk., 2006).
Menurut Salisbury dan Ross (1995) bahwa peningkatan GA3 endogen juga
dapat meningkatkan hidrolisis pati, fruktan, dan sukrosa menjadi molekul glukosa
dan fruktosa. Selulosa merupakan penggabungan unit-unit glukosa menjadi
senyawa makromolekul yang tidak larut dalam semua pelarut yang biasa
digunakan (Fengel dan Gerd, 1995). Menurut Abidin (1990) GA3 dapat
menghasilkan hidrolisa pati yang akan mendukung terbentuknya α-amilase.
Sebagai akibat dari proses tersebut, maka konsentrasi glukosa akan meningkat.
Kekurangan air akan mengakibatkan metabolisme sel terganggu termasuk
proses fotosintesis. Fotosintesis akan terhambat jika air yang merupakan bahan
utama hanya tersedia dalam jumlah sedikit, sehingga hasil dari fotosintesis juga
akan berkurang (Harjadi dan Yahya, 1988). Menurut Islami dan Wani (1995)
bahwa cekaman air menyebabkan perubahan macam dan jumlah senyawa
karbohidrat di dalam tanaman. Tanaman yang mengalami cekaman air terjadi
penurunan tepung dan peningkatan kadar gula.
Menurut Hamid (2001) dan Sjostrom (1995) menyebutkan bahwa
biosintesis selulosa dimulai dari glukosa. Maka dengan pemberian GA3 dan
variasi ketersediaan air kandungan glukosa dalam tanaman meningkat yang dapat
digunakan untuk bosintesis selulosa, sehingga kandungan selulosa dalam tanaman
juga ikut meningkat. Jalur sintesis selulosa dari glukosa dapat dilihat pada
Gambar 7.

27
Gambar 7. Jalur Biosintesis selulosa dari glukosa (Sjostrom, 1995).
Glukosil aktif (UDP-glukosa) merupakan prekursor dalam sintesis
selulosa. UDP-glukosa dihasilkan dalam sitoplasma dari dua sumber: dari sukrosa
oleh sintase sukrosa (1) (reaksi dapat balik) dan dari glukosa oleh reaksi-reaksi
berurutan yang dikatalisis oleh heksokinase (2), fosfoglukomutase (3), dan UDP-
glukopirofosfosilase (4). Setelah penembusan ke dalam selaput plasma, maka
UDP-glukosa mentranfer sisa glukosilnya ke rantai pertumbuhan glukan
(selulosa) disertai dengan pelepasan UDP. Penggabungan ini dikatalisis oleh
tempat-tempat aktif pada subunit-subunit kompleks sintase selulosa yang
disimpan dalam selaput plasma. Rantai-rantai glukan yang berasal dari satu
kompleks diperkirakan terasosiasi dengan ikatan hidrogen membentuk
mikrofibril, yang ukurannya dapat bervariasi diantara tipe-tipe sel yang berbeda.
Ketika sintesis berlangsung orientasi mikrofibril dapat ditentukan dengan gerakan
kompleks dalam lapisan ganda cairan lipida. Gerakan seperti ini dapat diarahkan
oleh mikrotubula yang terdapat pada permukaan dalam selaput plasma
(Sjostrom, 1995).
Masuknya bahan-bahan tambahan ke dalam dinding sel dalam rangka
selulosa disebut inkrustasi (pengerakan). Proses inkrustasi yang paling penting
pada tumbuhan tinggi adalah lignifikasi, tetapi pada banyak sel, bahan-bahan lain

28
seperti suberin, tanin, kutin, lilin kuinin, dan bahan organik serta bahan mineral
lainnya dapat pula melapisi dinding sel (Fahn, 1991).
Menurut Neish (1968), Sarkanen (1971), Griseboch (1977), Gross (1977)
dan (1978) dalam Fengel dan Gerd (1995) menyebutkan bahwa biosintesis lignin
dimulai dari glukosa. Maka dengan pemberian GA3 dan variasi ketersediaan air
kandungan glukosa dalam tanaman meningkat yang dapat digunakan untuk
bosintesis lignin, sehingga kandungan lignin dalam tanaman juga ikut meningkat.
Jalur metabolik dari glukosa menjadi senyawa induk lignin dapat dilihat pada
Gambar 8.
Gambar 8. Jalur metabolik dari glukosa menjadi senyawa induk lignin (Fengel
dan Gerd, 1995).
Fenilalanin ammonia liase Tirosin amonialiase
Fenolase
Fenolase
Fenilalanin
Tirosin
CoA Ligase Reduktase
Dehidrogenase
Asam Sinamat
Asam p-koumarat p- Koumaril-CoA-ester
p-Koumaraldehida
Asam kafeat
Asam 5-hidroksi ferulat
Asam ferulat Feruloil-CoA ester
Asam prefenat Asam shikimat
Reduktase
Sinapoil CoA ester
Sinapildehida
Koniferildehida
p-Koumaral-alkohol
p-Koniferil alkohol
CoA Ligase
Dehidrogenase
Asam sinapat Reduktase CoA Ligase
Metiltransferase
Fenolase
Metiltransferase
Sinapil alkohol
Dehidrogenase
Glkukosa

29
Pembentukan makromolekul lignin oleh tumbuhan terdiri atas sistem
biologi, biokimia dan kimia yang kompleks. Banyak studi dengan karbon radioktif
menegaskan bahwa p-hidroksisinamil alkohol, p-koumaril-alkohol, p-koniferil
alkohol dan sinapil alkohol merupakan senyawa induk (prekursor) primer dan
merupakan unit pembentuk semua senyawa lignin (Fengel dan Gerd, 1995).
Gambar 8 di atas memberikan gambaran secara umum langkah-langkah
utama pembentukan senyawa induk lignin. Biosintesis lignin dimulai dari glukosa
yang diperoleh dari fotosintesis. Ia diubah menjadi asam shikimat, senyawa antara
dari yang disebut jalur shikimat. Dua asam amino aromatik L-fenilalanin dan
L-tirosin dibentuk berdasarkan animasi reduktif melalui asam prefenat sebagai
senyawa-senyawa akhir dari jalur tersebut. Zat-zat tersebut merupakan zat-zat
awal (kelompok asam amino) untuk metabolisme enzimatik fenil propanoid (jalur
asam sinamat) yang menghasilkan tiga sinamil alkohol melalui turunan asam
sinamat teraktifasi. Asam-asam amino dideaminasi oleh deaminase (fenilalanin
ammonia liase dan tirosin aminoliase) menjadi asam sinamat yang sesuai.
Langkah-langkah utama lebih lanjut adalah hidroksilasi (oleh fenolase/
hidroksilase) menghasilkan asam p-koumarat, asam kafeat, asam ferulat, asam
5-hidroksi-ferulat, dan asam sinapat. Sinamil alkohol (p-koumaril-alkohol,
p-koniferil alkohol dan sinapil alkohol) akhirnya dibentuk oleh aktifasi enzimatik
(CoA ligase) dan reduksi (NADP reduktase, NADP hidrogenase) asam-asam yang
sesuai melalui koenzim-A trioster (p-koumaril-CoA-ester, Feruloil-CoA-ester dan
Sinapoil-CoA-ester) dan aldehida (p-kumaraldehida, koniferal dehida dan
sinapildehida) (Fengel dan Gerd, 1995). P-kumaril alkohol, koniferil alkohol dan
sinapil alkohol merupakan senyawa induk (precursor) primer dan merupakan unit
pembentuk semua lignin (Fengel dan Gerd, 1995; Robinson, 1995). Jalur
metabolik senyawa induk lignin menjadi lignin dapat dilihat pada Gambar 9.

30
Gambar 9. Jalur metabolik senyawa induk lignin menjadi lignin (Robinson, 1995;
Sjostrom, 1995).
Menurut Biemelt et al (2004) bahwa, pemberian GA3 meningkatkan
biosintesis lignin dan menstimulasi formasi xilem pada tembakau transgenik.
Xinjun Li et al (2003) juga menyatakan bahwa pemberian GA3 selama
pembungaan dan induksi anakan mampu meningkatkan kandungan lignin pada
Myrica rubra. Mudyantini (2008) juga menyatakan bahwa pemberian GA3
meningkatkan kandungan lignin pada B. nivea.
Lignin

31
Kekuatan Mulur Serat
Hasil rerata kekuatan mulur serat tanaman rami dengan perlakuan GA3 dan
ketersediaan air disajikan pada Tabel 13.
Tabel 13. Rerata kekuatan mulur serat B. nivea dengan perlakuan GA3 dan ketersediaan air berumur 2 bulan setelah tanam (%).
Ketersediaan Air Perlakuan
GA3 A1 A2 A3 Rerata
G0 - - 1,88 1,88 G1 0,94 2,09 1,26 1,43 G2 1,74 1,66 2,23 1,88 Rerata 1,34a 1,87b 1,79ab
Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200. Ketersediaan Air (%), A1=50, A2=75, A3=100.
* Angka yang diikuti huruf sama pada baris dan kolom yang sama menunjukkan tidak ada beda nyata pada uji DMRT pada taraf 5%.
Dari analisis sidik ragam General Linier Model (GLM) menunjukkan
bahwa perlakuan GA3 dan ketersediaan air tidak memberikan pengaruh yang nyata
terhadap kekuatan mulur serat, sedangkan interaksi antara GA3 dan ketersediaan
air memberikan pengaruh yang nyata terhadap kekuatan mulur serat dengan nilai
signifikansi sebesar 0,017. Perbandingan kekuatan mulur serat B. nivea dengan
perlakuan GA3 dan variasi ketersediaan air ditunjukkan pada Gambar 10.

32
Gambar 10. Kekuatan mulur serat B. nivea dengan perlakuan GA3 dan perlakuan
ketersediaan air berumur 2 bulan setelah tanam. Keterangan:
Konsentrasi GA3 (ppm), G0=0, G1=175, G2=200 Ketersediaan Air (%), A1=50, A2=75, A3=100
Gambar 10 menunjukkan bahwa kekuatan mulur serat paling tinggi ada
pada perlakuan GA3 200 ppm dan ketersediaan air 100% (G2A3) yaitu sebesar
2,23%, sedangkan kekuatan mulur terendah ada pada perlakuan GA3 175 ppm dan
ketersediaan air 50% (G1A1) yaitu sebesar 0,94%. Hal ini menunjukkan bahwa
setiap tumbuhan membutuhkan konsentrasi yang sesuai untuk pertumbuhannya.
Salah satu sifat penting selulosa adalah kemampuannya bertahan terhadap
regangan karena kelenturannya. Lignin menambah ketahanan dinding terhadap
tekanan dan mencegah melipatnya mikrofibril selulosa. Arah mikrofibril yang
berbeda-beda merupakan faktor penting penentu kekuatan dinding. Sifat mekanik
yang luar biasa dari selulosa ialah kekuatan meregang, sedangkan di bawah
tekanan kompresif, fibril-fibril selulosa itu membengkok. Sifat fisik dinding sel
diantaranya adalah regangan, kekuatan, ketahanan terhadap tekanan,
mengembang, dan sifat permeabilitas ditentukan oleh perbedaan komposisi dan
struktur lamela yang bertambah terus selama proses pembentukan dinding.
Perbedaan struktur dapat disebabkan karena perbedaan arah dan kerapatan
mikrofibril selulosa, perbedaan kandungan lignin dan lain-lain (Fahn, 1991).
Pemberian GA3 dan variasi ketersediaan air dapat meningkatkan
kandungan glukosa dalam tanaman maka kandungan selulosa juga akan
meningkat. Abidin (1990) mengemukakan bahwa GA3 dapat menghasilkan

33
hidrolisa pati yang akan mendukung terbentuknya α-amilase. Sebagai akibat dari
proses tersebut, maka konsentrasi glukosa akan meningkat. Menurut Mudyantini
(2008) bahwa perlakuan GA3 dapat meningkatkan kandungan glukosa dalam
tanaman yang dapat meningkatkan kandungan selulosa. Perlakuan GA3 yang
paling optimal dalam meningkatkan selulosa pada B. nivea yaitu pada konsentrasi
200 ppm dengan nilai kandungan selulosa sebesar 26,33 % b/b.
Kekurangan air dapat menghambat laju fotosintesa, karena turgiditas sel
penjaga stomata akan menurun (Haryati, 2003). Menurut Harjadi dan Yahya
(1988) bahwa fotosintesis akan terhambat jika air yang merupakan bahan utama
hanya tersedia dalam jumlah sedikit, sehingga hasil dari fotosintesis juga akan
berkurang. Menurut Islami dan Wani (1995) bahwa cekaman air menyebabkan
perubahan macam dan jumlah senyawa karbohidrat di dalam tanaman. Tanaman
yang mengalami cekaman air terjadi penurunan tepung dan peningkatan kadar
gula. Menurut Lee (1999) dalam Indrawan (2007), faktor yang berpengaruh
terhadap kekuatan tarik mulur serat adalah kelembaban. Semakin besar
kelembaban semakin besar pula kekuatan mulur serat dan sebaliknya akan
cenderung menurunkan kekuatan tarik.
KESIMPULAN
1. Perlakuan GA3 berpengaruh meningkatkan pertumbuhan B. nivea pada
panjang tunas dengan konsentrasi 200 ppm, tetapi menurun pada berat
basah dan berat kering. Perlakuan GA3 tidak berpengaruh pada jumlah
tunas dan daun. Sedangkan perlakuan ketersediaan air, maupun interaksi
antara GA3 dan ketersediaan air tidak berpengaruh terhadap semua
parameter pertumbuhan B. nivea.
2. Perlakuan GA3 berpengaruh meningkatkan terhadap kekuatan tarik serat
pada konsentrasi 200 ppm, tetapi tidak berpengaruh terhadap kekuatan
mulur serat. Perlakuan ketersediaan air tidak berpengaruh terhadap semua
parameter serat, sedangkan interaksi antara GA3 dan ketersediaan air
berpengaruh terhadap kekuatan mulur serat B. nivea dengan KL 75%.
DAFTAR PUSTAKA

34
Abidin, Z. 1990. Dasar-dasar Pengetahuan tentang Zat Pengatur Tumbuh. CV.
Angkasa. Bandung. Aisyah, W. 2004. “Pengaruh Perendaman Umbi Bibit dalam Larutan Giberelin
terhadap Pertumbuhan dan Kandungan Allicin Umbi Bawang Merah Allium cepa var. ascalonicum (L) Back”. Skripsi. Fakultas Biologi. UGM. Yogyakarta.
Anggarwulan, E., Solichatun dan W. Mudyantini. 2008. “Karakter Fisiologi
Kimpul (Xanthosoma sagittifolium (L.) Schott) pada Variasi Naungan dan Ketersediaan Air”. Biodiversitas. 9(4): 264-268.
Anwarudin, M.J; N.L.P. Indriyani; S. Hadiati dan E. Masyah. 1996. “Pengaruh
Konsentrasi Giberelin dan Lama Perendaman Terhadap Perkecambahan dan Pertumbuhan Biji Manggis”. J. Host. 6(1): 1-5.
Biemelt, S., H. Tschiersch and U. Sonnewald. 2004. Impact of Altered Giberelin
Metabolisme on Biomass Accumulation, Lignin Biosintesis, and Photosyntesis in Transgenic Tobacco Plants. Plant Physiol 135(1) : 254-256.
Brink, M. and R.P. Escobin.(Eds).2003.Plant Resurces of South-East Asia No.17.
Fibre Plants. Prosea Foundation. Bogor. Indonesia. Pp.86-91. Buxton, A. and P. Greenhalg. 1989. Ramie, Short live Curiosity or fibre. The
future Textile Outlook international, May, 1989. The Economist Intellegence Unit, London. (5): 52-71.
Davies, J.P. 1995. Plant Hormone: Their Nature, Occurrence and Function). In
P.J. Davies (edt) : Plant Hormones : Phisiology, Biochemistry, and Moleculer Biology. Kluwer Academic Publisher. Boston.
Deptan. 2007. Tanaman Rami, Emas Putih yang Terpendam.
http://www.ditjenbun.deptan.go.id [24 Juli 2008]. Dewi, A.P. 1993. Pengaruh Stress Air terhadap Perkembangan Dua Kultivar
Tanaman Kedelai (Glycine max (L.) Merry Wilis dan Lompo Batang. http://digilib.bi.itb.ac.id/go.php?id=jbptitbbi-gdl-s1-1993-anugerahpa-841 [2 Mei 2009].
Fahn, A. 1991. Anatomi Tumbuhan. (diterjemahkan oleh Ahmad Soediarto dkk.).
Universitas Gadjah Mada Press, Yogyakarta. Fengel, D and W. Gerd. 1995. Kayu, Kimia, Ultrastruktur, Reaksi-Reaksi.
(diterjemahkan oleh Hardjono S). Gadjah Mada University Press, Yogyakarta.

35
Fitter, A.H., R.K.M. Hay. 1998. Fisiologi Lingkungan Tanaman. (diterjemahkan
oleh Sri Andani dan E. D. Purbayanti). Gadjah Mada University Press, Yogyakarta.
Gardner, F.P., R.B. Pearce dan R.L. Mitcheli. 1991. Fisiologi Tanaman Budidaya.
(diterjemahkan oleh Herawati Susilo). UI Press, Jakarta. Gul, H., A.M. Khatak and N. Amin. 2006. Accelerating the Growth of Araucaria
heterophylla Seedlings Through Different Gibberellic Acid Concentrations and Nitrogen levels. Journal of Agricultural and Biological Science.1(2): 25-29.
Haddy, S. 1987. Biologi Pertanian. Rajawali Press, Jakarta . Hamid, A. 2001. Biokimia : Metabolisme Biomolekul. Penerbit Alfabeta.
Bandung. Harjadi, S.S. 1993. Pengantar Agronomi. Jakarta: Penerbit PT. Gramedia. Harjadi, S.S. dan S. Yahya. 1988. Fisiologi Stres Lingkungan. PAU Bioteknologi
IPB, Bogor. Haryati. 2003. Pengaruh Cekaman Air Terhadap Pertumbuhan dan Hasil
Tanaman. Program Studi Hasil Pertanian. Fakultas Pertanian. Universitas Sumatera Utara. Medan.
Hidayanto, M., S. Nurjanah., dan F. Yossita. 2003. “Pengaruh Panjang Stek Akar
dan Konsentrasi Natriumnitrofenol terhadap Pertumbuhan Stek Akar Sukun (Artocarpus communis F.)”. Jurnal Pengkajian dan Pengembangan Teknologi Pertanian. 6(2):154-160.
Hill, A.F. 1972. Economic Botany. Tata Mc. Graw Hill Publishing Company
Limited. New Delhi. Indrawan, M. 2007. ”Karakter Sutera Dari Ulat Jedung (Attacus atlas L.) Yang
Dipelihara Pada Tanaman Senggugu (Clerodendron Serratum Spieng)”. Laporan Penelitian Dosen Muda. Jurusan Biologi Fakultas MIPA UNS. Surakarta.
Islami, T. dan H.U. Wani. 1995. Hubungan Air dan Tanaman. IKIP Semarang
Press, Semarang. Kastono, D. 2005. “Pengaruh Jumlah Batang Bawah dan Kadar IAA terhadap
Pertumbuhan Bibit Durian Sambung Pucuk”. Agrivet. 9(1): 1-5.

36
Kusumo, S. 1990. Zat Pengatur Tumbuh Tanaman. Penerbit Yusa Guna. Bogor. Pp: 54-61.
Laboratorium Evaluasi Tekstil. 2008. Cara Kerja Pengoprasian Alat Uji Tenso
Lab./ Mesdan Lab.(Alat Uji Kekuatan Tarik dan Mulur). Jurusan Teknik Industri FTI-Universitas Islam Indonesia, Yogyakarta.
Lakitan, B. 1996. Fisiologi Pertumbuhan dan Perkembangan Tanaman. Jakarta:
Penerbit PT. Raja Grafindo Persada. Levitt, J. 1980. Responses of Plants to Environmental Stress Volume II. Academic
Press, New York. Loveless, A.R. 1991. Prinsip-Prinsip Biologi Tumbuhan Daerah Tropik
(diterjemahkan oleh Kusuma Kartawinata). Gramedia. Jakarta. Miranti, E. 2007. Mencermati Kinerja Tekstil Indonesia antara Potensi dan
Peluang. Jakarta. Mudyantini, W. 2008. “Pertumbuhan, Kandungan Selulosa, dan Lignin pada Rami
(Boehmeria nivea L. Gaudich) dengan Pemberian Asam Giberelat (GA3)”. Biodiversitas. 9(4): 269-274.
Mudyantini, W., Suratman, dan Sutarno. 2006. “Kandungan Selulosa dan Lignin
pada Rami (Boehmeria nivea L. Gaudich.) serta Limbah Hasil Perendaman Jerami dengan Perlakuan Bakteri Alkalofil”. Enviro. 7(1): 27-31.
Mullet, J.E and M.S. Whitsitt. 1996. Plant Cellular Responses to Water Deficit.
In: E. Belhassen (edt) Drought Tolerance in Higher Plants. Kluwer Academic Publisher, Netherland.
Patoni, 2000. “Pengaruh Cekaman Kekeringan terhadap Pertumbuhan, Hasil dan
Kandungan Vitamin C Buah Tanaman Tomat (Lycopersicum esculentum Mill.)”. Skripsi. Fakultas Biologi UGM. Yogyakarta.
Pezelshki, S.R. 1994. Plant Response to Flooding. In Wilkinson, R.E. (ed). In
Plant –Environment Interaction. Mercel Dekker, Inc., USA. Rahardjo, M., S.M.D. Rosita, R. Fathan, dan Sudiarto. 1999. “Pengaruh Cekaman
Air Terhadap Mutu Simplisia Pegagan (Centella asiatica L.)”. Jurnal Littri. 5(3): 92-97.
Rahman, M.H., M.H. Haque, M.A. Karim and M. Ahmed. 2006. Effects of
Gibberellic Acid (GA3) on Breaking Dormancy in Garlic (Allium sativum L.). Jounal of Agriculture and Biology. 8(1).

37
Robinson, T. 1995. Kandungan Organik Tumbuhan Tinggi (diterjemahkan oleh Kosasih Padmawinata). Penerbit ITB, Bandung.
Salisbury, F.B. and C.W. Ross. 1995. Fisiologi Tumbuhan Jilid 3 (diterjemahkan
oleh Dian Lukman). Penerbit ITB, Bandung. Sanjaya, L. 1991. Pengaruh asam giberelin terhadap pertumbuhan dan kualitas
bunga seruni. Prosiding Tanaman Hias. Cipanas, Sub Balithor Cipanas. Sitompul, S.M. dan B. Guritno. 1995. Analisis Pertumbuhan Tanaman. Gadjah
Mada University Press, Yogyakarta. Sjostrom, E. 1995. Kimia Kayu Dasar-dasar dan Penggunaan. Edisi Kedua.
(diterjemahkan oleh Hardjono S). Gadjah nada University Press, Yogyakarta.
Sulistyaningsih, Y.C., Dorli dan H. Akmal. 1994. “Studi Anatomi Daun Saccarum
sp. Sebagai Induk dalam Pemuliaan Tebu”. Hayati. 1(2): 61-65. Sumiasri, N dan D. Priadi. 2003. ”Pertumbuhan Stek Cabang Sungkai (Peronema
canescens Jack) dalam Media Cair”. Jurnal Nature. 6(1). Suyana, J. Dan H. Widijanto. 2002. Studi Kualitas Air dan Sumbangan Hara dari
Irigasi Sidorejo Jawa Tengah Pada Budidaya Padi Sawah. Sains Tanah. 2(1): 1-5.
Saroso, B. dan A. Sastrosupadi. 2000. “Tanaman Rami sebagai Bahan Tekstil,
Pulp, Pakan Ternak dan Obat”. Warta Penelitian dan Pengembangan Tanaman Industri. Pusat Penelitian dan Pengembangan Tanaman Perkebunan. Bogor.
Taiz, L. and E. Zeiger. 1998. Plant Physiologi. Massashusetts: Sinauer
Assosiattes, Inc. Tarmansyah, U.S. 2007. “Pemanfaatan Serat Rami untuk Pembuatan Selulosa”.
Tim Puslitbang Indhan Balitbang Dephan. Buletin Litbang Dephan.10(18): 1-12.
Wareing, P.F. and I.D.J. Phillips. 1981. Growth and Differentiation in Plants. 2nd
Edition. Pergamon Press. Toronto. Wattimena, G.A. 1998. Zat Pengatur Tumbuh Tanaman. Pusat Antar Universitas.
IPB. Bogor. pp: 12-15, 54-56.

38
Weaver, T.E., C.R. Stocking., M.G. Barbour., and T.L. Rost., 1982. Botany: An Introduction to Plant Biology. 6nd edition. University of California. California.
Wuryaningsih, S dan T. Sutater. 1993. Pengaruh zat pengatur tumbuh dan pupuk
N terhadap pertumbuhan dan produksi bunga krisan standar warna putih. Buletin Tanaman Hias. 1(1): 47-55.
Xingjun Li; Sanyu Li and JinXing Lin. 2003. Effect of GA3 Spraying on Lignin
and Auxin Contens and the Corellated Enzyme Activities In Bayberry (Myrica rubra Bieb.) During Flower-Bud Induction. Plant Science. 164(4): 549-556.

39
PERTUMBUHAN, KEKUATAN TARIK DAN MULUR SERAT RAMI
(Boehmeria nivea (L.) Gaudich) DENGAN PEMBERIAN ASAM
GIBERELAT (GA3) DAN VARIASI KETERSEDIAAN AIR
Naskah Publikasi
Skripsi
Untuk memenuhi sebagian persyaratan guna memperoleh gelar Sarjana Sains
Oleh : Shafi Fauzi Rahman
M0405057
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SEBELAS MARET
SURAKARTA
2009

40
PERSETUJUAN
Naskah Publikasi
SKRIPSI
PERTUMBUHAN, KEKUATAN TARIK DAN MULUR SERAT RAMI (Boehmeria nivea (L.) Gaudich) DENGAN PEMBERIAN ASAM
GIBERELAT (GA3) DAN VARIASI KETERSEDIAAN AIR
Oleh:
Shafi Fauzi Rahman
NIM. M0405057
Telah disetujui untuk dipublikasikan
Surakarta, ………………...
Menyetujui,
Pembimbing I
Widya Mudyantini, M.Si. NIP. 197305051999032001
Pembimbing II
Dra. Endang Anggarwulan, M.Si.
NIP. 195003201978032001
Mengetahui, Ketua Jurusan Biologi
Dra. Endang Anggarwulan, M.Si. NIP. 195003201978032001





























