Embed Size (px)
Citation preview

Biochem. J. (1967) 104, 716
The Cellulase of Trichoderma virideSEPARATION OF THE COMPONENTS INVOLVED IN THE SOLUBILIZATION OF COTTON
BY K. SELBY AND C. C. MAITLANDShirley Institute, Didebury, Marnche8ter 20
(Received 16 December 1966)
1. Culture filtrates from Trichoderma viride have been fractionated by gelfiltration on Sephadex G-75 followed by ion-exchange chromatography on DEAE-and SE-Sephadex. 2. The components essential for attack on cotton are a
carboxymethylcellulase, a cellobiase and a third (C1) component which has no
action on CM-cellulose, cellobiose or cotton. 3. These components, which togethercan completely convert cotton into water-soluble products, lose this ability whenseparated and regain it quantitatively when recombined in their originalproportions.
The study of the mechanism of action of theenzyme systems from cellulolytic micro-organismshas now progressed to a stage where interest centreson the type of enzyme that must be produced byorganisms which degrade highly ordered forms ofcellulose such as unmodified fibrous cotton. Thistype of enzyme (Cl-enzyme) was postulated byReese, Siu & Levinson (1950), who recognized thatit must be distinct from those 'cellulases' (Cs)capable of attacking only swollen or chemicallymodified forms of cellulose. Because of failure toproduce culture filtrates with significant activitytowards unmodified cotton most of the work in thefollowing decade was confined to studies of C.activity. More recently (Selby, Maitland &Thompson, 1963) culture filtrates have been pre-pared from Myrothecium vemrcaria capable ofextensive attack on cotton, provided that repeatedapplications of fresh quantities of filtrate are made.Extracts from Trichoderma viride, are even moreactive (Mandels & Reese, 1964) and have recentlybeen fractionated by these workers and by manyothers (Iwasaki, Hayashi & Funatsu, 1964; Shikata,1964; Niwa, Kawamura & Nisizawa, 1965; Ogawa &Toyama, 1965; Li, Flora & King, 1965). A Cl-typeenzyme was found to be the only componentcapable, on its own, of solubilizing cotton (Mandels& Reese, 1964; Flora, 1965) and 'crystallinehydrocellulose' (Li et al. 1965; Flora, 1965),although this activity was enhanced by synergismwith C,.components. This type of synergismmakes it imperative that enzyme components arerigorously purified if the mechanisms of theirindividual actions are to be examined. The presentpaper shows that, when highly purified, the Cl-component ofT. viride is no longer able to solubilizecotton; recombination with the C.-components (a
cellobiase and a carboxymethylcellulase), however,restores all the activity of the original culturefiltrate. Li et al. (1965) have stressed the desirabilityof thus accounting for the original activity against'highly organized (crystalline) cellulose', but it isnot clear from their published results that this wasdone.
MATERIALSOrgani8m. Trichoderma viride was strain 92 027 from the
Commonwealth Mycological Institute, Kew, Surrey. Thecotton flock on which it was grown, and the cotton yarnthat was used for measurements of cellulase activity, wereas described by Selby et al. (1963). Sephadex G-75, DEAE-Sephadex (A-50) and SE-(sulphoethyl-)Sephadex (C-50)were supplied by Pharmacia (G.B.) Ltd., London. Cello-biose was obtained from Thomas Kerfoot and Co. Ltd., Valeof Bardaley, Lancs. Yeast extract and mycological peptonewere supplied by Oxo Ltd., London. Malt extract wasobtained from Boots Pure Drug Co. Ltd., Nottingham.Distiller's solubles were from Hiram Walker and Sons,Dumbarton, Scotland.
METHODSGrowth of Trichoderma viride. The organism was grown
in 250ml. shake flasks (rotary shaker, 0l5in. eccentricity,140rev./min.) in 75ml. of a medium containing cotton (3%),NH4NO3 (0-06%), NaNO3 (0.38%), KH2PO4 (0.02%),K2HPO4 (0.015%), NaH2PO4 (0.2%), Na2HPO4 (0-15%),MgSO4,7H20 (0.03%), urea (0.03%), asparagine (0.05%),malt (0 02%), yeast (0.02%), mycological peptone (0 02%),and distiller's solubles (0.02%). In addition, each litre ofmedium containedtrace elements asfollows: Ca(NO3)2,4H20(1-2mg.), FeSO4,7H20 (0 054mg.), H3BO3 (006mg.),CuSO4,5H20 (0-2mg.), ZnSO4,7H20 (2.0mg.), MnC12,4H20(0-08mg.), CoCl2,6H20 (0-4mg.), (NH4)sMo7O24,4H20(0.04mg.). After incubation for 9 days at 25° the contentsof the flasks were filtered and the filtrate (culture filtrate)was protected from infection by adding 0O005mole ofNaN3/l.
716
CELLULASE OF TRICHODERMA VIRIDEPreparation of Sephadex G-75for column chromatography.
Dry Sephadex G-75 (bead form, 40-120u) was separated byair flotation in a vertical cylinder, the fine particles beingblown out of the top of the cylinder and collected. Theseparation was slow, and flow at any one rate was continuedfor 24hr. before increasing it to harvest the next largerfraction ofparticles. Particles of50-65,u diam. were swollenby suspension for 14 days in a culture filtrate from T. viridediluted fivefold with sodium phosphate eluting buffer(OO1M in total phosphate, 0-1M in NaCl and 0-005M inNaN3, pH6-3). They were then washed with, and re-suspended in, eluting buffer.
Vertical tubes of precision-bore glass (2.5 cm. x 60cm.)were fitted with column ends designed to give minimal hold-up of liquid. A series cf closely-spaced V-shaped parallelgrooves, with a single diametral V-shaped groove at rightangles, was cut across the wider face of a silicone-rubberbung, chosen so that its wider end fitted the column reason-ably tightly. A -mm. hypodermic tube was pushedthrough the centre of the bung so that one end was flushwith the bottom of the groove and the other could take a
narrow-bore vinyl delivery tube. The cut face of the bungwas covered with a circle (2.5cm. diam., 0-75mm. thick) ofsintered polythene (pore size 50,u), which was madewettable by treatment with a solution of n-octanol inethanol.One of these ends was inserted into the bottom of the
column and the suspension of Sephadex G-75 was siphonedin, to avoid air bubbles, and allowed to settle under gravityaided by a slow percolation of eluting buffer. A second endwas then carefully introduced at the top of the colujmn andpushed down until it reached the upper surface of the bed,with the outlet at the bottom of the column closed to avoidcompression of the gel. In use the flow rate was 10ml./hr.and a flow of buffer was maintained at all times. For use incascade two similar columns were connected with narrow-
bore vinyl tubing.Calibration of Sephadex G-75 columns. The relation
between elution volume and the logarithm of molecularweight for a number of reference materials on the two
columns in cascade was established (cf Selby & Maitland,1965).
Jon-exchange chromatography. Columns (1-8cm. precisionbore, with ends as described above) were packed withparticles of SE- or DEAE-Sephadex of 15S-600,u diam.(fragmented gel, as supplied, with the fines removed).Buffer gradients were applied by means of a nine-chambervariable-gradient mixer (Peterson & Rowland, 1961).Measurement of carboxymethylcellukee activity. The rapid
method described by Selby & Maitland (1965) was used.Measurement of cellobiaee activity. Cellobiose (0-667mg.1
ml.) was dissolved in succinate buffer solution (pH4-5,O-OlM in Na+ and 0-005M in NaN3). A portion (100p4.) ofthe enzyme solution to be assayed, or of an appropriatedilution of it, was incubated with the cellobiose solution(3 ml.) for 3 hr. at 400. The increase in reducing power was
measured by the alkaline iodine method (Selby, 1965), theaddition of the carbonate-bicarbonate buffer terminatingthe enzymic reaction. The reducing power of the addedenzyme sample was allowed for by a blank determination.The percentage conversion of cellobiose into glucose was
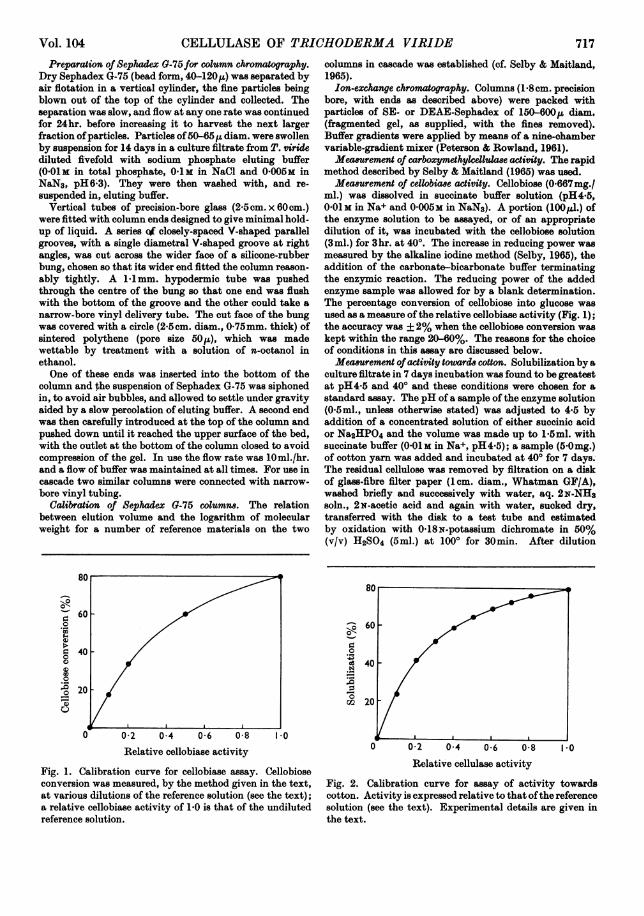
used as a measure ofthe relative cellobiase activity (Fig. 1);the accuracy was +2% when the cellobiose conversion waskept within the range 20-60%. The reasons for the choiceof conditions in this assay are discussed below.Measurement of activity towards cotton. Solubilization bya
culture filtrate in 7 days incubation was found to be greatestat pH4-5 and 40° and these conditions were chosen for a
standard assay. The pH of a sample of the enzyme solution(0-5ml., unless otherwise stated) was adjusted to 4-5 byaddition of a concentrated solution of either succinic acidor Na2HPO4 and the volume was made up to 1- ml. withsuccinate buffer (0-01 M in Na+, pH4-5); a sample (5-Omg.)of cotton yarn was added and incubated at 400 for 7 days.The residual cellulose was removed by filtration on a diskof glass-fibre filter paper (1cm. diam., Whatman GFJA),washed briefly and successively with water, aq. 2u-NH3soln., 2iN-acetic acid and again with water, sucked dry,transferred with the disk to a test tube and estimatedby oxidation with 0-18N-potaasium dichromate in 50%(v/v) H2SO4 (5ml.) at 1000 for 30min. After dilution
>0!'0
Go
C)00
4)
0
.._0
41)
0
0._
Ca
.._I-0
0
Relative cellobiase activity
Fig. 1. Calibration curve for cellobiase assay. Cellobioseconversion was measured, by the method given in the text,at various dilutions of the reference solution (see the text);a relative cellobiase activity of 1-0 is that of the undilutedreference solution.
0-2 0-4 0-6 0-8
Relative cellulase activity
Fig. 2. Calibration curve for assay of activity towardscotton. Activity is expressed relative to that ofthe referencesolution (see the text). Experimental details are given inthe text.
Vol. 104 717
K. SELBY AND C. C. MAITLANDwith water (15ml.) the excess of dichromate was titratedwith 01 N-ferrous ammonium sulphate, phenylanthranilicacid being used as an internal indicator. Although theoxidation ofcotton under these conditions is stoicheiometric(1-Oml. of O-lN-ferrous ammonium sulphate=0-675mg. ofdry cotton), in practice, each series of determinationsincluded a blank sample of unattacked cotton (5.0mg.), sothat frequent re-standardization of the reagents was un-necessary. Enzyme samples from gel filtration usuallycontained 01 m-NaCl but this had a negligible effect oncellulase activity. The reference solution (see below) atseveral dilutions gave the solubilizations shown in thecalibration curve (Fig. 2). Cellulase activity was mostaccurately measured by this method when the solubilizationwas between 20% and 80%.
RESULTS
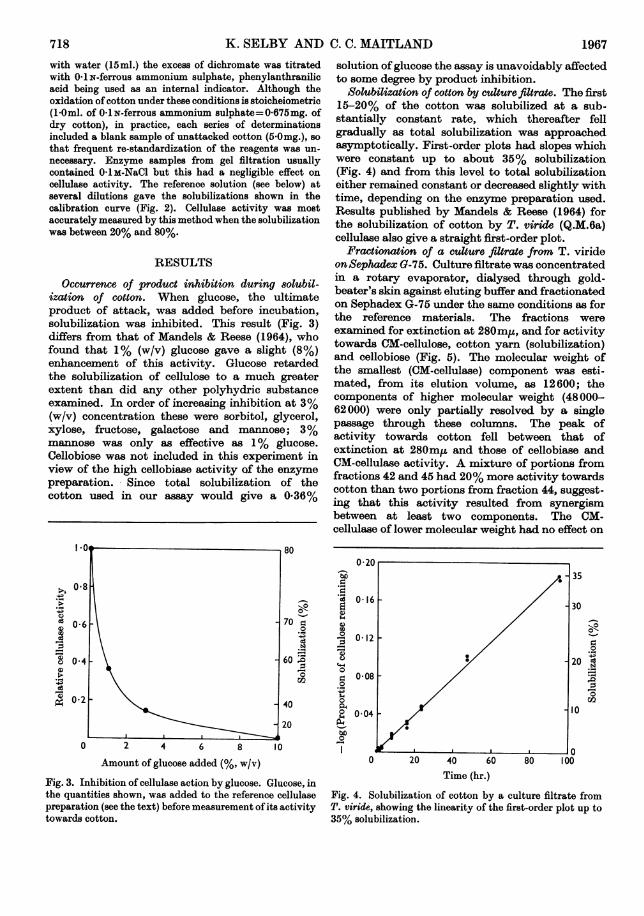
Occurrence of product inhibition during 8olubil-ization of cotton. When glucose, the ultimateproduct of attack, was added before incubation,solubilization was inhibited. This result (Fig. 3)differs from that of Mandels & Reese (1964), whofound that 1% (w/v) glucose gave a slight (8%)enhancement of this activity. Glucose retardedthe solubilization of cellulose to a much greaterextent than did any other polyhydric substanceexamined. In order of increasing inhibition at 3%(w/v) concentration these were sorbitol, glycerol,xylose, fructose, galactose and mannose; 3%mannose was only as effective as 1% glucose.Cellobiose was not included in this experiment inview of the high cellobiase activity of the enzymepreparation. Since total solubilization of thecotton used in our assay would give a 0-36%
solution ofglucose the assay is unavoidably affectedto some degree by product inhibition.
Solubilization of cotton by culture filtrate. The first15-20% of the cotton was solubilized at a sub-stantially constant rate, which thereafter fellgradually as total solubilization was approachedasymptotically. First-order plots had slopes whichwere constant up to about 35% solubilization(Fig. 4) and from this level to total solubilizationeither remained constant or decreased slightly withtime, depending on the enzyme preparation used.Results published by Mandels & Reese (1964) forthe solubilization of cotton by T. viride (Q.M.6a)cellulase also give a straight first-order plot.
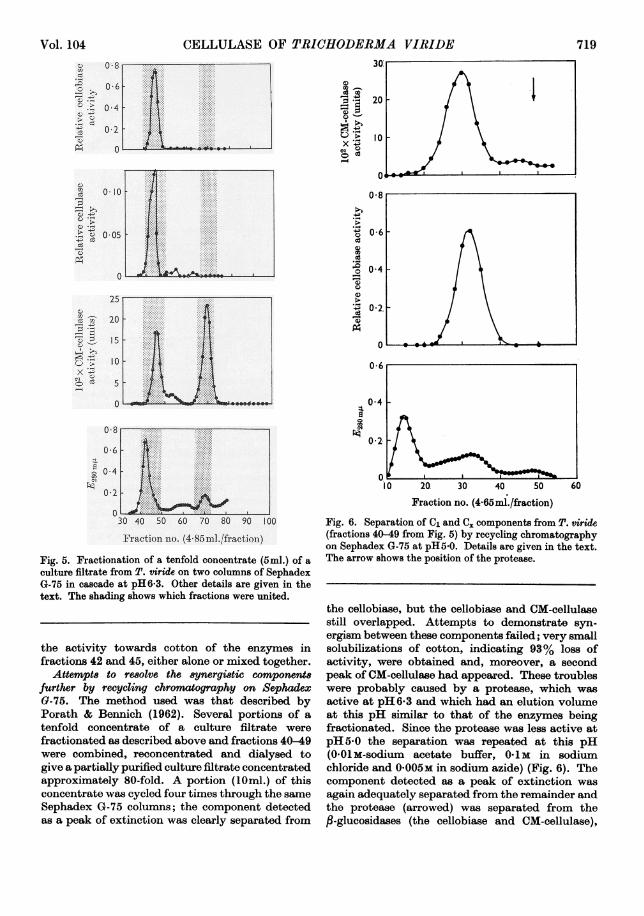
Fractionation of a culture filtrate from T. virideonSephadex G-75. Culture filtrate was concentratedin a rotary evaporator, dialysed through gold-beater's skin against eluting buffer and fractionatedon Sephadex G-75 under the same conditions as forthe reference materials. The fractions wereexamined for extinction at 280m,u, and for activitytowards CM-cellulose, cotton yarn (solubilization)and cellobiose (Fig. 5). The molecular weight ofthe smallest (CM-cellulase) component was esti-mated, from its elution volume, as 12600; thecomponents of higher molecular weight (48 000-62 000) were only partially resolved by a singlepassage through these columns. The peak ofactivity towards cotton fell between that ofextinction at 280mp, and those of cellobiase andCM-cellulase activity. A mixture of portions fromfractions 42 and 45 had 20% more activity towardscotton than two portions from fraction 44, suggest-ing that this activity resulted from synergismbetween at least two components. The CM-cellulase of lower molecular weight had no effect on
2 4 6 8
Amount of glucose added (%, w/v)
-70
O a
CS
._
60 C)
0 v
20 $I
Fig. 3. Inhibition of cellulase action by glucose. Glucose, inthe quantities shown, was added to the reference cellulasepreparation (see the text) before measurement of its activitytowards cotton.
40 60Time (hr.)
0-4-
coN1._
b;
0
Go2
Fig. 4. Solubilization of cotton by a culture filtrate fromT. viride, showing the linearity of the first-order plot up to35% solubilization.
PC
O.-
-a
C;Ca
C)
C.
718 1967
CELLULASE OF TRICHODERMA VIRIDE
*:.e L0.0
30 40 50 60 70 80 20 100
Fraction nlo. ([email protected])Fig. 5. Fractionation of a tenfold concentrate (5m1.) of aculture filtrate from T. viride on two columns of SephadexG-75 in cascade at pH6*3. Other details are given in thetext. The shading shows which fractions were united.
the activity towards cotton of the enzymes infractions 42 and 45, either alone or mixed together.
Attempt to re8olve the 8ynergi8tic component8further by recycling chromatography on SephadexG-75. The method used was that described byPorath & Bennich (1962). Several portions of a
tenfold concentrate of a culture filtrate were
fractionated as described above and fractions 40-49were combined, reconcentrated and dialysed togive a partially purified culture filtrate concentratedapproximately 80-fold. A portion (lOml.) of thisconcentrate was cycled four times through the sameSephadex G-75 columns; the component detectedas a peak of extinction was clearly separated from
_
-
0 ._acz -4
-4
U +
._
c;
c)
I._
sL
0 6
0-44
02
10 20 30 40 50
Fraction no. (4-65ml./fraction)60
Fig. 6. Separation of C1 and C. components from T. viride(fractions 40-49 from Fig. 5) by recycling chromatographyon Sephadex G-75 at pH5-0. Details are given in the text.The arrow shows the position of the protease.
the cellobiase, but the cellobiase and CM-cellulasestill overlapped. Attempts to demonstrate syn-ergism between these components failed; very smallsolubilizations of cotton, indicating 93% loss ofactivity, were obtained and, moreover, a secondpeak of CM-cellulase had appeared. These troubleswere probably caused by a protease, which wasactive at pH 6-3 and which had an elution volumeat this pH similar to that of the enzymes beingfractionated. Since the protease was less active atpH5-0 the separation was repeated at this pH(0801m-sodium acetate buffer, 0-1 M in sodiumchloride and 0-005M in sodium azide) (Fig. 6). Thecomponent detected as a peak of extinction wasagain adequately separated from the remainder andthe protease (arrowed) was separated from thefl-glucosidases (the cellobiase and CM-cellulase),
Vol. 104 719
T- 'TV%&

K. SELBY AND C. C. MAITLAND
which had changed their order of emergence butwere still not separated from each other. Maximumsynergism was obtained between fractions 14 and30, suggesting that the CM-cellulase was moreeffective than the cellobiase in synergizing with thecomponent detected by its extinction, which waspresumably Cl. The recovery of activity towardscotton (33%) was better at pH 5-0 than at pH 6-3.Removal of proteaae from partially fractionated
culture filtrate. After partly successful attempts toremove the protease by specific adsorption on solidproteins, it was found to be adsorbed on SE-Sephadex (C-50) at pH5-0 and, if necessary,recovered by elution at pH 7-0. A large sample ofculture filtrate was therefore purified preparatoryto further attempts to separate the synergisticcomponents.
Culture filtrate (11.) was concentrated tenfold,dialysed against eluting buffer (pH6-3) andfractionated (in two parts; 60ml.) on a wide column(5.0cm. x 30cm.) of Sephadex G-75. The fractionswere batched to include all components that wouldhave appeared in fractions 38-51 in Fig. 5. Thiscrude fractlonation removed materials of lowmolecular weight, including the CM-cellulase thatappears between fractions 66 and 72 in Fig. 5. In asimilar fractionation it had been shown that activityagainst cotton was not lost; in fact, equivalentdilutions of the cellulase before and after removalof the material of low molecular weight gavesolubilizations of 58.0% and 56.8% respectively.The batched fractions were concentrated to 100ml.,dialysed against sodium succinate buffer (0.01M inNa+ and 0-005m in sodium azide, pH 5 0) and passedthrough a column (1cm. x 8cm.) of Sephadex C-50equilibrated with the same buffer, followed by 10ml.of the buffer. The resulting protease-free partiallypurified culture filtrate was used for the furtherfractionation studies described below and is calledthe 'reference solution'. Its activities towardscellobiose and cotton can be seen in the calibrationcurves (Figs. 1 and 2); its aM-cellulase activity was58 x 10-2 unit.Further fractionation of partially purified culture
filtrate by ion-exchange chromatography. In view ofthe small differences between the elution volumesof the synergistic components on Sephadex G-75,the behaviour of the system on ion-exchange formsof Sephadex was studied. On DEAE-Sephadexneither cellobiase nor CM-cellulase was stronglyadsorbed from 0-01 M buffer solutions with pHvalues below 7-0, and at higher pH values activitywas lost. However, the Cl-component was adsorbedfrom buffer solutions with pH values between 5-5and 6-5 and could be recovered without loss byelution at pH3-7. When the Ci-component wasobtained by elution in a pH gradient (Fig. 7), ithad little action on cotton and neither cellobiase
I
j
1_
'-4
SX '
-e
.
I-0
so403020I00
0.
0 _
0D4
0-
4-0
m3 8 A
3-6
0 10. 20 30 40 50 60Fraction no. (3 2ml.Ifaotion)
Fig. 7. Separation of the Cl- and C.-components fromT. viride by chromatography on a DEAE-Sephadex column(1-8cm. x 12-5 cm.; sodium succinate buffers 0 01 m in Na+and 0-005m in NaN3). A sample (lOml.) of the referencesolution (see text) slightly diluted (CM-cellulase activity51.5 x 10-2 unit, relative cellobiase activity 088) wasapplied to the column at pH535, followed by a steep pHgradient (30ml. each of buffer solutions at pHB535 and 4 6).The C.-component had emerged before the pH of the eluatebegan to change. When the pH had fallen to about 5-0, thegradualpH gradient shown in the Figure was applied [buffersolutions at pH4.6, 3-8, then four at 3-6 (30ml. in eachcompartment)] to elute the Cl-component. The rate offlow was lOml./hr. The shading shows which fractions wereunited.
nor CM-cellulase activity; it is believed to be themost homogeneous specimen of a Cl-component yetreported.The cellobiase activities of dilutions of the
batched fractions (4-10) were compared with thoseof the reference solution at corresponding dilutionschosen to allow for the fact that, in chromatographyand batching, the Cs-components had been diluted2-55 times; there had been negligible loss ofactivity.The CM-cellulase activity in fractions 4-10 was23 x 10-2 unit, which also represents completerecovery. The Cl-component had been diluted1-6 times by chromatography and was therefore 1-8times as dilute as in the reference solution. Beforeassessing the effect of recombination a sample(1.8ml.) of the fractionated Ci-component wasdiluted to 2-55ml. so that it had the same concentra-tion relative to the reference solution as C.. The
Cx IpHcx ,
720 1967
A% I 9 Pk ^l.

CELTULASE OF TRICHODERMA VIRIDETable 1. Activitime of the Ci- and CS fractione from Trichoderma viride towards cotton
Component(s)(0 5ml. ofeach component,total volume made up to Solubilization1-5 ml. with buffer solution) (%)
Reference solution diluted 59.02-55 timesCi
Cx
C1+Cs
2*03*8
59.5
Relativecellulaseactivity
0395
<0-010-020 405
Recovery ofcellulase activity
(%)100
1
5102
activities towards cotton of these diluted com-
ponents were compared with that of the referencesolution, diluted 2-55 times (Table 1) to confirm thatno activity had been lost during separation of theCi- and C,.components.
Fractionation of the Cs-component. The view thatthe cellobiase and CM-cellulase activities of C. were
due to different enzymes, suggested by their partialseparation on Sephadex G-75, was strengthened bythe results of selective destruction. At pH2-5 and400 99.7% of the cellobiase activity of C. was
destroyed in 18hr., and 54% of the original CM-cellulase activity remained. Conversely, after 18hr.at pH75 and 40°, 99.6% ofthe CM-cellulaseactivitywas destroyed and 51% of the cellobiase activityremained. However, these treatments also reducedthe ability of the two enzymes to act synergisticallywith the Cl-component; whereas intact C., added toC1, gave 69% solubilization of cotton, the heat-treated cellobiase and CM-cellulase together, addedto C1, gave only 32% solubilization, although theirconcentrations had been chosen to give un-
diminished cellobiase and CM-cellulase activities.Li et al. (1965) have described a method for
separating the exoglucanase (cellobiase?) andendoglucanase of T. viride, in which the endo-glucanase was adsorbed on alkali-swollen cellulosefrom 0-O0M-phosphate buffer at pH5-5 and 20 andwas then eluted with water. When this method wasapplied to our C.-fraction the cellobiase, which wasnot adsorbed, was quantitatively recovered freefrom CM-cellulase, but elution of the adsorbed CM-cellulase, whether with water, 1% CM-cellulose,aq. 30% ethanol or 0O01m-EDTA, was slow andincomplete.A better method of separation was devised with
SE-Sephadex, with a long column to compensatefor weak retention. The behaviour of these ,B-glucosidases on SE-Sephadex was studied in theregion pH4-5-55. pH proved to be critical in thisseparation; too low apH led to loss of cellobiase butat higher pH values the separation was inadequate.The best compromise was achieved at pH5 1 (Fig. 8)and the recoveries of both CM-cellulase (fractions12-20) and cellobiase (fractions 23-29) were 96%.
12
; 6 bC *X *
-I
1v
8 cM.cSUul
0 3
0*2
_
II
*1
I0-1
- 10 20 30a 40Fraotion no. (3-2mL/fiaton)
Fig. 8. Separation, on a SE-Sephadex column (1.8cm.x63-5cm.; sodium succinate buffer, pH5B1, 001m in Na+ and0 005m in NaNg), of the CM-cellulase and cellobiase froma sample (3.2ml., CM-cellulase activity 45*0 x 10-2 unit,relative cellobiase activity 0.63) of C. from T. viride. Thissample was obtained from a fractionation of Cl and C.similar to that shown in Fig. 7, from which, to avoidexcessive dilution, two fractions only (corresponding tofractions 6 and 7 in Fig. 7) were taken from the peak of C.,giving a somewhat altered ratio of CM-cellulase to cellobiaseactivity. The rate of flow was 10ml./hr. The shadingshows which fractions were united.
These small elution volumes (column volume= 50fractions) show that this separation depended on
exclusion by ionic repulsion and not on retentionby ionic attraction. The CM-cellulase had no
activity on cellobiose, and the cellobiase had very
little on CM-cellulose.Behavior of the separated componfentM on
Sephadex G-75. Since they had unchanged elutionvolumes when chromatographed on the calibrated
Vol. 104 721
I
1.Oi

K. SELBY AND C. C. MAITLAND
Table 2. Solubilization of cotton by component8 of Trichoderma viride cellula8e, alone and in combination
Component(s)(0.5ml. of each component,total volume made up to1-5ml. with buffer solution,
pH4-5)Reference solution diluted4 times
C1+CxClCM-cellulaseCellobiaseCM-cellulase+ cellobiaseC1+ CM-cellulaseCl+ cellobiaseCl+ cellobiase+ CM-cellulase
Recovery ofRelative cellulase
Solubilization cellulase activity(%) activity (%)46-0 0-24 100
47-71.03*20*81*8
21-014-047*2
0-25<0-01
0-01<0-01<0-010.0850-050-25
10414
<12
3520104
pair of columns of Sephadex G-75, each of theseparated components had survived the wholepurification procedure without detectable changeof molecular weight.Synergism between component8. The cellobiase
and CM-cellulase had been somewhat diluted duringseparation on SE-Sephadex and were thereforeconcentrated and dialysed against buffer (pH4-5)to give activities corresponding to a fourfolddilution of the reference solution (CM-cellulase=14-5 x 10-2 unit, cellobiose conversion= 40%). Thesolution containing Cl was diluted to a similarequivalent concentration (1.8ml. of solution +2*2ml. of pH4-5 buffer) as was that containing C.(2.55ml. of solution+ 1-45ml. of buffer). Theactivities towards cotton of combinations of thesesolutions are shown in Table 2. On CM-cellulose orcellobiose there was no synergism between any ofthe three components. Activity towards cotton,lost by separation ofthe components, was recoveredcompletely on recombining all three in the propor-tions present in the reference solution. There waslittle synergismbetween CM-cellulase and cellobiase.Neither component alone could restore all thelost activity of C1 and the CM-cellulase was moreeffective than the cellobiase, as had been tentativelydeduced from the results of recycling chromato-graphy on Sephadex G-75 at pH50.
Effect of dialy8i8 on the aolubilization of cotton byC, and C.. Each enzyme preparation (0.5ml.),diluted with succinate buffer solution (1Oml.,OO1M in Na+ and 0-005M in sodium azide, pH4-5),was applied to a sample (5mg.) of cotton andcontinuously dialysed against the same buffer(150ml.) during incubation for 7 days at 400 in aglass cell (1-8cm. diam.), closed at the bottom withgoldbeaters' skin. The activity of C. was notenhanced but that of Ci was increased by 50%(solubilization rose from 5.6% to 8.0%). Thiseffect, though small, is reproducible and has beenrepeated with other preparations of Cl.
DISCUSSION
Determination of relative cellobia8e activitiem. Theassay for cellobiase departed from ideal in two ways,which led to the use of non-absolute units. First,the substrate concentration (1.9mM) was below thatneeded to saturate the enzyme. Li et al. (1965)report for their exoglucanase acting on cellobiose aKm 2-2mm and apparent inhibition by the substrateat concentrations above 0-05M; we have found,measuring conversions of between 5% and 15%,that the rate of conversion shows a sharp maximumat cellobiose concentrations near 5mM. It seemedpreferable to work at a lower concentration,althoughthe rate would be influenced by availabilityof substrate, rather than at a higher concentrationwhere inhibition predominates. Secondly, sub-stantial degrees of substrate conversion weremeasured; at lower degrees of conversion thepossible effects of product inhibition would belessened and the conversion would be more nearlyproportional to enzyme concentration but themeasurement of conversion by this method, or byany other depending on reducing power, wouldbecome less accurate because of the initial reducingpower of the cellobiose.
Solubilization of cotton by culture filtrate. In theassay of activity towards cotton high levels ofconversion were employed in case the earliest stagesof cellulase attack were on some particularlysusceptible part of the cotton structure but, in viewof the nearly first-order course of the solubilizationreaction (see below), this fear may have been un-founded. Comparisons between the attack of thecellulase of Myrothecium verrucaria and of mineralacid on cotton have already been made (Selby,1961); others can now be made between acid andthe cellulase of T. viride. Attack by mineral acidfollows first-order kinetics except in the initialstages when a part of the cotton (about 10%)is solubilized at a rate much higher than the
1967722

CELLULASE OF TRICHODERMA VIRIDE
remainder. This reactive part has been termedamorphous, though in the fringe-micellar theorythe amorphous part was often considered to be ashigh as 40% to account for other properties of thecotton (Howsmon & Sisson, 1954; but for a morerecent theory see Warwicker, Jeffries, Colbran &Robinson, 1966). A supposed analogy of itsstructure to that ofcertain reprecipitated cellulosesthat are very susceptible to cellulases may have ledseveral workers (Norkrans, 1950; Walseth, 1952;Reese, Segal & Tripp, 1957) to believe that these'amorphous' regions must also be the first to beattacked by cellulases. The absence of any markedfall in rate during the early stages of solubilizationby the cellulase of T. viride (Fig. 4) shows that theseacid-labile regions are not especially susceptible tothese enzymes. This may be because the enzymemolecules are confined by their size to largeropenings in the cotton structure and so, for example,may be able to act only on the surfaces of theelementary fibrils of which cotton is composed. Inthe solubilization of a solid substrate by superficialerosion, first-order kinetics would not be expectedunless the erodible area were, throughout theprocess, proportional to the amount of substrateremaining. Foster & Wardrop (1951) and Sharples(1957) have shown that this requirement is fulfilledby a model in which the erodible dimension of thesubstrate particles is exponentially distributed (inthe statistical sense) and have so explained thefirst-order erosion of cotton by acid. The first-ordererosion by cellulases may be explained by a similarmodel but, if our present views on cellulase actionare correct, the erodible dimension of the fibrils istheir width, whereas Sharples (1957) suggests thatthe dimension erodible by acids is the length offibrils lying between the acid-susceptible regions.
Properties of the separated components. The Cl-component is a glycoprotein with a carbohydrate:protein ratio approx. 1: 1 [carbohydrate determinedas equivalent glucose by orcinol method (Weimer &Moshin, 1952), protein by micro-Kjeldahl digestionfollowed by determination of ammonia on anamino acid analyser]. Its molecular weight,estimated from its behaviour during gel filtration, isapprox. 61000; it is heat-labile, losing 50% of itspotential activity towards cotton on heating insolution at pH4-5 and 1000 for about 2-5min. It ishomogeneous on Sephadex G-75, DEAE-Sephadexand SE-Sephadex and gives a single protein bandby disk electrophoresis at pH 6-7 (method of Davis,1964, as modified by Broome, 1963). The lack ofactivity in the purified Cl-component raised thepossibility that its role when acting synergisticallywith the C.-components might be a non-specificprotein effect. However, attempts to enhance theactivity of the mixed C.-components towardscotton by adding serum albumin (0-7mg./ml.)
failed completely; the already low activity of the CQpreparation was further reduced.
Synergistic action of two enzymes in solubilizinga solid substrate may be of two kinds. It may beeither on the substrate, when the first enzymemakes the substrate susceptible to solubilization bythe second, or in solution, when the first solubilizesthe substrate but a second is needed to removeinhibitory products formed. To test for synergismon the substrate, Ci and C. were applied to cottoneither simultaneously or in sequence (Table 3).Clearly neither component can complete its actionin the absence of the other. The limited synergismfound in the sequential treatments could have beendue to incomplete removal of one component fromthe cotton before application of the other. Becausecontinuous dialysis enhanced the solubilization ofcotton by C1 but not by C., it seems likely that thesynergism may be in solution, C1 being stronglyinhibited by a soluble, dialysable product which C.is able to remove.The most significant feature of these results is
that the Cl-component of T. viride, when freedfrom the C.-components, has little or no action oncellobiose, CM-cellulose or cotton. Mandels & Reese(1964) have reported a similar separation of aculture filtrate from T. viride on DEAE-Sephadex,but without prior removal of protease or CM.cellulase of low molecular weight, and the relativepositions of their f,-, A- and C-components weresimilar to those of our cellobiase, CM-cellulase andCi-component respectively. The major differencewas that their C-component had lost less activitytowards cotton than had our Ci-component, so thatthe synergistic effect was less marked. Flora (1965)also separated a Cl-component from T. viride, byadsorption on Avicel, but found that it still degradedcotton; he obtained only a threefold enhancementof activity on recombining it with his C.-fraction.
Table 3. Relative activities of Cl and C,, applied tocotton separately, together, and in sequence
These samples of Ci and C. were obtained by recyclingchromatography (Fig. 6, Ci comprised fractions 13, 14, 15;C. comprised fraction 29). C, (1-Oml.) and C. (0-5ml.) wereeither mixed together or diluted to 1-5ml. Each treatmentwas for 1 week; when two were applied in sequence, eachwas for 1 week with thorough washing of the cotton samplewith buffer solution (pH4-5) after the first. The activitiesare expressed relative to the mixture of C1 and C., whichgave a solubilization of 78%.
TreatmentCi+ C.CiC,C1 then C.C. then C1
Relative activity (%)100
821528
Vol. 104 723

724 K. SELBY AND C. C. MAITLAND 1967
The fact that the activity lost during the fractiona-tion procedure described here was regained com-pletely on recombining the fractions in their originalproportions (Table 2) proved that the Cl-component, though inactive alone, was essentialfor extensive solubilization of cotton, but itscontribution to the synergistic action of the wholecellulase system is still not understood.We are indebted to our colleague Mrs K. V. A. Thompson,
who supplied culture filtrates from T. viride, and to Dr N. J.King of Forest Products Research Laboratory, PrincesRisborough, who examined the Cl-component by diskelectrophoresis. We are grateful to Glaxo LaboratoriesLtd. for their sponsorship of this work.
REFERENCESBroome, J. (1963). Nature, Lond., 199, 179.Davis, B. J. (1964). Ann. N.Y. Acad. Sci. 121, 404.Flora, R. M. (1965). Ph.D. Thesis: Virginia Polytechnic
Institute; Ann Arbor, Mich.: University Microfilms(65-2041).
Foster, D. H. & Wardrop, A. B. (1951). Aust. J. 8Ci. Res.A, 4, 412.
Howsmon, J. A. & Sisson, W. A. (1954). In Cellulose andCellulose Derivatives, 2nd ed., p. 231. Ed. by Ott, E.,Spurlin, H. M. & Grafflin, M. W. New York: IntersciencePublishers Inc.
Iwasaki, T., Hayashi, K. & Funatsu, M. (1964). J. Biochem.,Tokyo, 55, 209.
Li, L. H., Flora, R. M. & King, K. W. (1965). Arch. Biochem.Biophys. 111, 439.
Mandels, M. & Reese, E. T. (1964). In Development8 inIndustrial Mycology, vol. 5, p. 5. Washington, D.C.:American Institute of Biological Sciences.
Niwa, T., Kawamura, K. & Nisizawa, K. (1965). Proc. 5thSymp. Cellulase and Related Enzyme8, p. 44. OsakaUniversity, Japan: Cellulase Association.
Norkrans, B. (1950). Physiol. Plant. 3, 75.Ogawa, K. & Toyama, N. (1965). Proc. 5th Symp. Cellulkeand Related Enzymes, p. 85. Osaka University, Japan:Cellulase Association.
Peterson, E. A. & Rowland, J. (1961). J. Chromat. 5, 330.Porath, J. & Bennich, H. (1962). Arch. Biochem. Biophys.
Suppl. 1, p. 152.Reese, E. T., Segal, L. & Tripp, V. W. (1957). Text.Res. (J.),
27, 626.Reese, E. T., Siu, R. G. H. & Levinson, H. S. (1950). J.Bad. 59, 485.
Selby, K. (1961). Biochem. J. 79, 562.Selby, K. (1965). J. chem. Soc. p. 1554.Selby, K. & Maitland, C. C. (1965). Biochem. J. 94, 578.Selby, K., Maitland, C. C. & Thompson, K. V. A. (1963).
Biochem. J. 88, 288.Sharples, A. (1957). Tranm. Faraday Soc. 53, 1003.Shikata, S. (1964). J. Biochem., Tokyo, 56, 1.Walseth, C. S. (1952). T.A.P.P.I. 35, 233.Warwicker, J. O., Jeffries, R., Colbran, R. L. & Robinson,
R. N. (1966). In Shirley Institute Pamphlet no. 93: AReview of the Literature on the Effect of Caustic Soda andother Swelling Agents on the Fine Structure of Cotton.Manchester: Cotton Silk and Man-Made Fibres ResearchAssociation.
Weimer, H. E. & Moshin, J. R. (1952). Amer. Rev. Tuberc.pulm. Dis. 68, 594.










![Trichoderma Viride FNCC 6013 terhadap Produksi Kitinase ...eprints.undip.ac.id/73018/13/Pengaruh_Penambahan... · digunakan sebagai substrat dalam berbagai aplikasi industri [1, 2]](https://img.dokumen.tips/doc/110x75/5e477b1adeabcf45b51311d3/trichoderma-viride-fncc-6013-terhadap-produksi-kitinase-digunakan-sebagai-substrat.jpg)


















