Embed Size (px)
Citation preview

Neuroscience 282 (2014) 198–216
REVIEW
THE ANTERO-POSTERIOR HETEROGENEITY OF THE VENTRALTEGMENTAL AREA
M. J. SANCHEZ-CATALAN, a,b J. KAUFLING, c,d
F. GEORGES, c,d P. VEINANTE a,b AND M. BARROT a,b*
a Institut des Neurosciences Cellulaires et Integratives,
Centre National de la Recherche Scientifique, Strasbourg, France
bUniversite de Strasbourg, Strasbourg, France
cCentre National de la Recherche Scientifique, Interdisciplinary
Institute for Neuroscience, UMR 5297, Bordeaux, France
dUniversite de Bordeaux, Interdisciplinary Institute for
Neuroscience, UMR 5297, Bordeaux, France
Abstract—The ventral tegmental area (VTA) is a brain region
processing salient sensory and emotional information, con-
trolling motivated behaviors, natural or drug-related reward,
reward-related learning, mood, and participating in their
associated psychopathologies. Mostly studied for its dopa-
mine neurons, the VTA also includes functionally important
GABA and glutamate cell populations. Behavioral evidence
supports the presence of functional differences between
the anterior VTA (aVTA) and the posterior VTA (pVTA), which
is the topicof this review.Thisantero-posterior heterogeneity
concerns locomotor activity, conditioned place preference
and intracranial self-administration, and can be seen in
response to ethanol, acetaldehyde, salsolinol, opioids
includingmorphine, cholinergic agonists including nicotine,
cocaine, cannabinoids and after local manipulation of GABA
and serotonin receptors. It has also been observed after viral-
mediated manipulation of GluR1, phospholipase Cc (PLCc)and cAMP response element binding protein (CREB) expres-
sion, with impact on reward and aversion-related responses,
on anxiety and depression-related behaviors and on pain
sensitivity. In this review, the substrates potentially underly-
ing these aVTA/pVTA differences are discussed, including
theVTAsub-nuclei and the heterogeneity in connectivity, cell
types and molecular characteristics. We also review the role
of the tail of the VTA (tVTA), or rostromedial tegmental
nucleus (RMTg), which may also participate to the observed
antero-posterior heterogeneity of the VTA. This region, partly
located within the pVTA, is an inhibitory control center for
dopamine activity. It controls VTA and substantia nigra dopa-
mine cells, thus exerting a major influence on basal ganglia
http://dx.doi.org/10.1016/j.neuroscience.2014.09.0250306-4522/� 2014 IBRO. Published by Elsevier Ltd. All rights reserved.
*Correspondence to: M. Barrot, Institut des Neurosciences Cellul-aires et Integratives, CNRS UPR3212, 5 rue Blaise Pascal, 67084Strasbourg, France. Tel: +33-388-456-633.
E-mail address: [email protected] (M. Barrot).Abbreviations: aVTA, anterior VTA; CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione; CREB, cAMP response element bindingprotein; EM-1, endomorphin-1; NMDA, N-methyl-D-aspartate; PLCc,phospholipase Cc; pVTA, posterior VTA; RMTg, rostromedialtegmental nucleus; tVTA, tail of the VTA; VTA, ventral tegmental area.
198
functions. This review highlights the need for a more
comprehensive analysis of VTA heterogeneity.
This article is part of a Special Issue entitled: Ventral
Tegmentum & Dopamine. � 2014 IBRO. Published by
Elsevier Ltd. All rights reserved.
Key words: dopamine, ventral tegmental area, tVTA,
behavior, drugs of abuse.
Contents
Introduction 198
VTA antero-posterior functional heterogeneity 199
GABA transmission 199
Ethanol, acetaldehyde and salsolinol 199
Ethanol behavioral studies 199
From ethanol to acetaldehyde and salsolinol 201
Neurochemical studies 202
Electrophysiological studies 202
Opioids 203
Cholinergic system 204
Cocaine 204
Other drugs 205
Affect and pain 205
Neuroanatomy of the VTA 205
Definition of the VTA 205
The VTA and the A10 dopamine cell group 206
Cytoarchitectonic subdivisions of the VTA 206
BASES for the aVTA/pVTA differences 207
The tVTA 209
Detecting the tVTA 209
tVTA control of dopamine cells 210
tVTA and behavior 210
Conclusion 211
Acknowledgments 211
References 211
INTRODUCTION
The ventral tegmental area (VTA) is studied for its
implication in a wide range of functions including the
processing of salient sensory and emotional information,
the control of motivated behavior, natural or drug-related
reward, reward-related learning, mood, and their
associated psychopathologies (Nestler and Carlezon,
2006; Fields et al., 2007; Grace et al., 2007; Bromberg-
Martin et al., 2010; Hong, 2013; Creed et al., 2014;
Gillies et al., 2014; Ikemoto and Bonci, 2014; Meye and

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 199
Adan, 2014; Nikulina et al., 2014; Overton et al., 2014;
Walsh and Han, 2014). While most work related to the
dopamine cells of the VTA, recent attention has also been
given to the GABA and glutamate cell populations
(Roeper, 2013; Creed et al., 2014; Morales and Root,
2014). Beyond this cellular heterogeneity, behavioral evi-
dence has accumulated since the late nineties supporting
the presence of a major antero-posterior heterogeneity
within the VTA (Ikemoto, 2007). The functional difference
between the anterior VTA (aVTA) and the posterior VTA
(pVTA) is particularly supported by studies of the locomo-
tor, rewarding and reinforcing properties of various drugs
of abuse. However, the substrate underlying such aVTA/
pVTA differences remains elusive, with hypotheses based
on neuroanatomy, connectivity and cellular and molecular
heterogeneity. Moreover, in the past decade, an inhibitory
control center for midbrain dopamine cells was identified
and named the tail of the VTA (tVTA) or rostromedial teg-
mental nucleus (RMTg). The tVTA is partly located within
the pVTA and thus should also be considered when
studying the antero-posterior heterogeneity of the VTA.
In this review, we will first describe the behavioral and
physiological evidence supporting the antero-posterior
heterogeneity of the VTA. Indeed, historically, the first
data highlighting the importance of the VTA antero-
posterior functional heterogeneity came from
experiments of behavioral pharmacology. Most of these
studies did a direct side-by-side comparison of intra
aVTA and pVTA drug injections. We will then provide
information on the possible bases for such
heterogeneity, including the presence of subnuclei within
the VTA and the presence of potential differences in
connectivity, in cell types and in molecular cell
characteristics. It is indeed important to also consider
the available information on VTA anatomical
heterogeneity, even though no direct link has been
established yet between the antero-posterior functional
heterogeneity and the precise VTA subnuclei. Last, we
will summarize the present knowledge on the tVTA, a
structure with its most rostral portion within the pVTA
and extending caudally beyond the VTA, that exerts a
major control over the activity of mesencephalic
dopamine cells. The tVTA may have a critical role in
basal ganglia functions. While published work has not
directly compared the aVTA, pVTA and tVTA, some
evidence suggests that the latter structure might be
mediating some of the functions that were previously
attributed to the pVTA.
VTA ANTERO-POSTERIOR FUNCTIONALHETEROGENEITY
GABA transmission
A third of a century ago, it was observed that injections of
GABA modulators in the VTA had different effects on
locomotor activity depending on the injection site (Arnt
and Scheel-Kruger, 1979) (Table 1). Agonists of GABAA
receptors increased locomotor activity when delivered in
the pVTA but not in the aVTA, while GABAA antagonists
increased activity when delivered in the aVTA but not
the pVTA. In the late nineties, Ikemoto et al. observed that
Wistar rats self-administered antagonists of the GABAA
receptor, such as picrotoxin and bicuculline, into the
aVTA but not into the pVTA (Ikemoto et al., 1997b;
Ikemoto, 2005), whereas they self-administered the
GABAA agonist muscimol into the pVTA but not into the
aVTA (Ikemoto et al., 1998). These data (Table 1) high-
lighted the presence of a prominent functional heteroge-
neity at the level of the GABAergic transmission along
the antero-posterior axis of the VTA. However, it may
be challenging to control for the anatomical selectivity of
local injections, due to the diffusion of injected com-
pounds. Thus, the rewarding effects of GABAA antagonist
in the aVTA were later proposed to be associated with the
supramammillary nucleus, a hypothalamic area anterior
to the VTA and that plays also a role in reward
(Ikemoto, 2005, 2010). Nevertheless, differences in the
consequences of pVTA and aVTA manipulations
remained valid, and these first functional data on the ante-
ro-posterior heterogeneity of the VTA opened the path to
other studies, in particular in the field of ethanol action and
alcoholism (Table 1). It should be noted that, for a long
time, the ‘‘aVTA’’ and the ‘‘pVTA’’ were functionally com-
pared without the frontier between them being anatomi-
cally defined. A study on the response to cocaine
showed that the aVTA/pVTA limit in rats was around
�5.5 mm from the bregma, which neuroanatomically cor-
responds to the position of the interpeduncular nucleus
below the VTA ((Olson et al., 2005), see cocaine section
below).
Ethanol, acetaldehyde and salsolinol
Ethanol behavioral studies. Manipulations of the VTA
can modify the ethanol intake in rats (Hodge et al., 1993;
Katner et al., 1997). While rats directly self-administer eth-
anol into the VTA, this reinforcing property displays a
neuroanatomical selectivity (Fig. 1, Table 1). Indeed, rats
self-administer ethanol into the pVTA but not into the
aVTA (Rodd-Henricks et al., 2000, 2003; Rodd et al.,
2004b, 2005d; Ding et al., 2014), and the infusion of etha-
nol into the pVTA also increases the rat locomotor activity
(Sanchez-Catalan et al., 2009). Considering data obtained
in a strain of alcohol-preferring rats, the preferential sensi-
tivity of the pVTA may be relevant to the vulnerability to
alcohol. Alcohol-preferring rats self-infuse lower doses of
ethanol into the pVTA than Wistar rats (Rodd et al.,
2004a), and the dose of ethanol eliciting self-infusion in
the pVTA is even lower after chronic ethanol drinking
(Rodd et al., 2005b,c).
Ethanol can act through several ion channels and
neurotransmitter systems (Morikawa and Morrisett,
2010), a major mediator of its action being the GABAergic
system. In this context, the alcohol intake (Melon and
Boehm, 2011) and the conditioned place preference to
ethanol (Bechtholt and Cunningham, 2005) may be
decreased by a manipulation of GABAA or GABAB recep-
tors in the pVTA respectively. However another study was
also supportive of an influence of aVTA GABAA receptors
on ethanol intake (Nowak et al., 1998). Despite the pVTA
selectivity for ethanol self-administration, it should be
noted that the oral ethanol intake and the locomotor effects
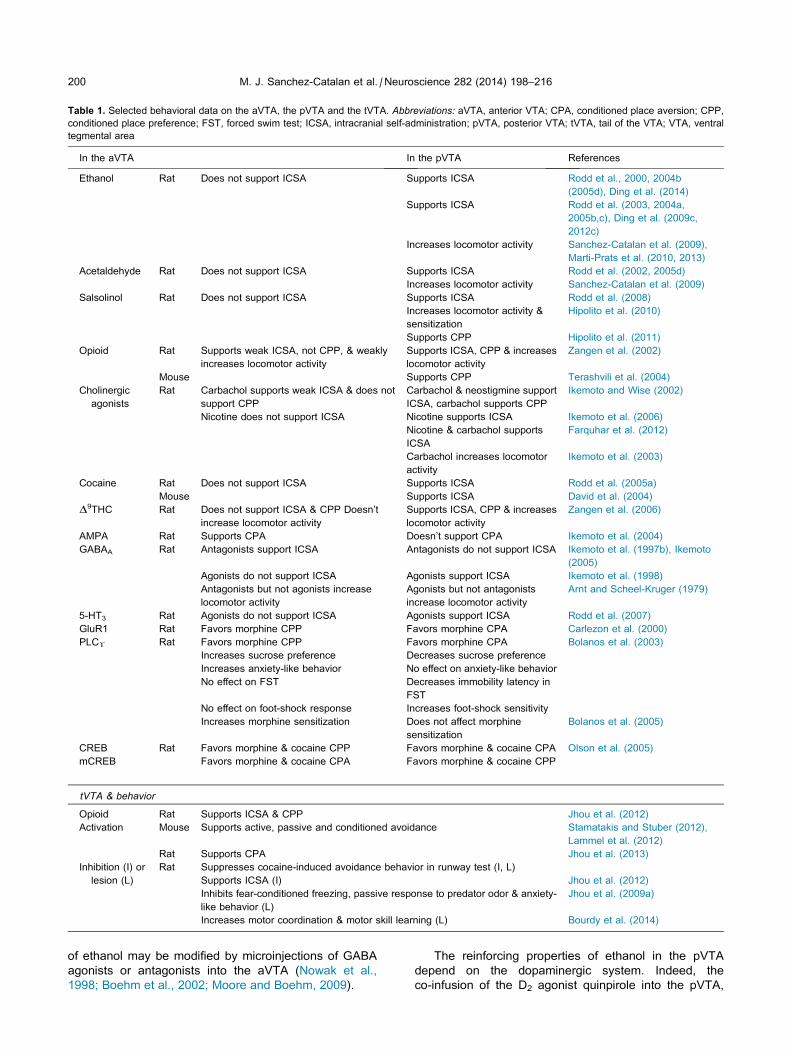
Table 1. Selected behavioral data on the aVTA, the pVTA and the tVTA. Abbreviations: aVTA, anterior VTA; CPA, conditioned place aversion; CPP,
conditioned place preference; FST, forced swim test; ICSA, intracranial self-administration; pVTA, posterior VTA; tVTA, tail of the VTA; VTA, ventral
tegmental area
In the aVTA In the pVTA References
Ethanol Rat Does not support ICSA Supports ICSA Rodd et al., 2000, 2004b
(2005d), Ding et al. (2014)
Supports ICSA Rodd et al. (2003, 2004a,
2005b,c), Ding et al. (2009c,
2012c)
Increases locomotor activity Sanchez-Catalan et al. (2009),
Marti-Prats et al. (2010, 2013)
Acetaldehyde Rat Does not support ICSA Supports ICSA Rodd et al. (2002, 2005d)
Increases locomotor activity Sanchez-Catalan et al. (2009)
Salsolinol Rat Does not support ICSA Supports ICSA Rodd et al. (2008)
Increases locomotor activity &
sensitization
Hipolito et al. (2010)
Supports CPP Hipolito et al. (2011)
Opioid Rat Supports weak ICSA, not CPP, & weakly
increases locomotor activity
Supports ICSA, CPP & increases
locomotor activity
Zangen et al. (2002)
Mouse Supports CPP Terashvili et al. (2004)
Cholinergic
agonists
Rat Carbachol supports weak ICSA & does not
support CPP
Carbachol & neostigmine support
ICSA, carbachol supports CPP
Ikemoto and Wise (2002)
Nicotine does not support ICSA Nicotine supports ICSA Ikemoto et al. (2006)
Nicotine & carbachol supports
ICSA
Farquhar et al. (2012)
Carbachol increases locomotor
activity
Ikemoto et al. (2003)
Cocaine Rat Does not support ICSA Supports ICSA Rodd et al. (2005a)
Mouse Supports ICSA David et al. (2004)
D9THC Rat Does not support ICSA & CPP Doesn’t
increase locomotor activity
Supports ICSA, CPP & increases
locomotor activity
Zangen et al. (2006)
AMPA Rat Supports CPA Doesn’t support CPA Ikemoto et al. (2004)
GABAA Rat Antagonists support ICSA Antagonists do not support ICSA Ikemoto et al. (1997b), Ikemoto
(2005)
Agonists do not support ICSA Agonists support ICSA Ikemoto et al. (1998)
Antagonists but not agonists increase
locomotor activity
Agonists but not antagonists
increase locomotor activity
Arnt and Scheel-Kruger (1979)
5-HT3 Rat Agonists do not support ICSA Agonists support ICSA Rodd et al. (2007)
GluR1 Rat Favors morphine CPP Favors morphine CPA Carlezon et al. (2000)
PLC! Rat Favors morphine CPP Favors morphine CPA Bolanos et al. (2003)
Increases sucrose preference Decreases sucrose preference
Increases anxiety-like behavior No effect on anxiety-like behavior
No effect on FST Decreases immobility latency in
FST
No effect on foot-shock response Increases foot-shock sensitivity
Increases morphine sensitization Does not affect morphine
sensitization
Bolanos et al. (2005)
CREB Rat Favors morphine & cocaine CPP Favors morphine & cocaine CPA Olson et al. (2005)
mCREB Favors morphine & cocaine CPA Favors morphine & cocaine CPP
tVTA & behavior
Opioid Rat Supports ICSA & CPP Jhou et al. (2012)
Activation Mouse Supports active, passive and conditioned avoidance Stamatakis and Stuber (2012),
Lammel et al. (2012)
Rat Supports CPA Jhou et al. (2013)
Inhibition (I) or
lesion (L)
Rat Suppresses cocaine-induced avoidance behavior in runway test (I, L)
Supports ICSA (I) Jhou et al. (2012)
Inhibits fear-conditioned freezing, passive response to predator odor & anxiety-
like behavior (L)
Jhou et al. (2009a)
Increases motor coordination & motor skill learning (L) Bourdy et al. (2014)
200 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
of ethanol may be modified by microinjections of GABA
agonists or antagonists into the aVTA (Nowak et al.,
1998; Boehm et al., 2002; Moore and Boehm, 2009).
The reinforcing properties of ethanol in the pVTA
depend on the dopaminergic system. Indeed, the
co-infusion of the D2 agonist quinpirole into the pVTA,

Fig. 1. Schematic of the behavioral action of ethanol, acetaldehyde and salsolinol in the aVTA and the pVTA. The aVTA/pVTA limit in rats is around
�5.5 mm from the bregma, which neuroanatomically corresponds to the presence of the interpeduncular nucleus below the VTA. Various receptors
can influence the behavioral consequences of the pVTA microinjections. Abbreviations: 5-HT, 5-hydroxytryptamine; aVTA, anterior ventral
tegmental area; CPP, conditioned place preference; ICSA, intracranial self-administration; IP, interpeduncular nucleus; pVTA, posterior ventral
tegmental area.
M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 201
which would inhibit the dopamine system by stimulating
local autoreceptors (Ford, 2014), prevents the local self-
administration of ethanol (Rodd et al., 2004b, 2005d).
The self-administration can then be reinstated by the
co-infusion into the pVTA of the D2 antagonist sulpiride
(Rodd et al., 2004b). Moreover, at terminal fields, the
blockade of D1 and D2 dopamine receptors in the nucleus
accumbens shell, the ventral pallidum or the medial pre-
frontal cortex, but not in the nucleus accumbens core,
reduces the pVTA ethanol self-administration (Ding
et al., 2014). This role of the dopaminergic system is
not limited to the self-administration of ethanol into the
pVTA. Indeed, the oral intake of ethanol and ethanol
seeking behavior are both reduced by the pVTA microin-
jection of a D2 agonist (Nowak et al., 2000; Hauser et al.,
2011). Pre-treatment with the D1 antagonist SCH23390 in
the pVTA also decreases ethanol intake, but does not
alter ethanol-seeking (Czachowski et al., 2012). Interest-
ingly, while ethanol is not directly self-administered in
the aVTA, the local manipulation of the dopaminergic sys-
tem can influence some ethanol-related behaviors. Thus,
the aVTA microinjection of D2 agonists reduces ethanol
intake but not ethanol-seeking behavior (Nowak et al.,
2000; Hauser et al., 2011).
The pVTA self-administration of ethanol is also locally
influenced by other transmitter systems, including the
opioid, the 5-HT, the cholinergic (nicotinic), the
glutamatergic and the cannabinoid systems, reflecting
the complex regulation of VTA activity. The endogenous
opioid system is intimately associated with ethanol
addiction (Gianoulakis, 2009), and opioid antagonists are
even used as a treatment for alcoholism (Spanagel and
Kiefer, 2008). The intra-pVTA administration of an opioid
antagonist prevents the locomotor-activating effects of
intra-pVTA ethanol in rats (Sanchez-Catalan et al.,
2009), and decreases ethanol-induced conditioned place
preference in mice (Bechtholt and Cunningham, 2005).
Various intra-pVTA 5-HT3 antagonists can also reduce
the pVTA ethanol self-administration (Rodd-Henricks
et al., 2003; Rodd et al., 2005d), the pVTA self-administra-
tion of a mixture of ethanol and cocaine (Ding et al., 2012c)
and the oral self-administration of ethanol (Rodd et al.,
2010), while the 5-HT3 antagonists had no effect on oral
ethanol self-administration when delivered into the aVTA
(Rodd et al., 2010). In fact, rats will even self-administer
a 5-HT3 agonist in the pVTA but not in the aVTA (Rodd
et al., 2007). The pVTA ethanol self-administration is also
attenuated by the co-infusion of a 5HT2A antagonist
(R96544), but not by a 5HT1B antagonist (GR55562)
(Ding et al., 2009c). An intra-pVTA microinjection of nico-
tine can favor ethanol seeking, which is prevented by a
nicotinic (mecamylamine) or 5-HT3 receptor antagonist
(zacopride) (Hauser et al., 2014). An intra-pVTA adminis-
tration of the glutamate antagonist CNQX (6-cyano-7-
nitroquinoxaline-2,3-dione) also reduces ethanol-seeking,
but without affecting ethanol intake (Czachowski et al.,
2012). Last, the local cannabinoid system also influences
ethanol action, since the administration of a CB1 agonist
(WIN 55–212) into the pVTA, but not into the aVTA, alters
the time-course of binge-like ethanol intake in mice
(Linsenbardt and Boehm, 2009). These various interac-
tions may also participate in a cross-vulnerability between
alcohol and other drugs of abuse.
In addition, the hyperpolarization-activated cyclic
nucleotide–gated (HCN) ion channels have also been
proposed as a molecular target of ethanol (Brodie and
Appel, 1998; Okamoto et al., 2006), and their overexpres-
sion in the pVTA increases voluntary ethanol intake in rats
(Rivera-Meza et al., 2014).
From ethanol to acetaldehyde and salsolinol. Signifi-
cant evidence implicates the first metabolite of ethanol,
acetaldehyde, in the mechanisms underlying the
psychopharmacological effects of ethanol (Correa et al.,
2012). In fact, it has been demonstrated that acetalde-
hyde has rewarding properties per se (Correa et al.,
2012). The relevance of the role of acetaldehyde in the
VTA is further supported by the local presence of the
enzymatic machinery necessary to metabolize ethanol
(Moreno et al., 1995; Sanchez-Catalan et al., 2008).
Rats self-administer acetaldehyde into the pVTA
(Fig. 1) but not the aVTA, and the co-administration of
the D2/3 agonist quinpirole is able to block this intra-
pVTA acetaldehyde self-administration (Rodd-Henricks
et al., 2002; Rodd et al., 2005d). Acetaldehyde microin-
jection into the pVTA also increases locomotor activity

202 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
in rats, which is prevented by an intra-pVTA pre-treatment
with a l-opioid receptor antagonist (Sanchez-Catalan
et al., 2009). The critical role of acetaldehyde in pVTA eth-
anol action is demonstrated by the pharmacological use
of an acetaldehyde sequestering agent, D-penicillamine.
Indeed, the locomotor activity induced by an intra-pVTA
ethanol administration can be reduced by D-penicillamine
(Marti-Prats et al., 2010). Moreover, a low dose of intra-
pVTA ethanol can even induce motor depressant action
after pre-treatment with D-penicillamine or with sodium
azide, a catalase inhibitor blocking the transformation of
ethanol into acetaldehyde. Interestingly, this depressant
effect can be prevented by the local microinjection of
the GABAA antagonist bicuculline (Marti-Prats et al.,
2013). Similarly, pre-treatment with the aldehyde dehy-
drogenase inhibitor cyanamide, which reduces the acetal-
dehyde degradation, stimulates motor activity in response
to the intra-pVTA administration of a normally non-
effective dose of ethanol (Marti-Prats et al., 2013). These
studies suggest that acetaldehyde is involved in the
activating effects of ethanol in the pVTA, while the non-
metabolized ethanol would display a motor depressant
action through GABAA receptors (Marti-Prats et al.,
2013). The use of sequestering-agents of acetaldehyde
proves to be a useful pharmacological tool to assess the
role of this metabolite in ethanol effects. The recent dem-
onstration that the intra-pVTA administration of D-penicil-
lamine can block relapse in a preclinical model of alcohol
deprivation (Orrico et al., 2013) also suggests a therapeu-
tic potential for these compounds.
The acetaldehyde is also a highly reactive compound.
In the presence of dopamine, a condensation reaction
leads to the production of salsolinol (1-methyl-6,7-
dihidroxy-1,2,3,4-tetrahydroisoquinoline) (Fig. 1), which
has been implicated in some of the neurobiological
effects of ethanol (Hipolito et al., 2012). Thus, a salsolinol
microinjection into the pVTA is sufficient to increase the
locomotor activity of rats in a dose-dependent manner,
this action being blocked by an intra-pVTA pre-treatment
with the opioid antagonist naltrexone or with the selective
l-opioid antagonist b-funaltrexamine (Hipolito et al.,
2010). When repeated, the intra-pVTA administration of
salsolinol leads to locomotor sensitization (Hipolito et al.,
2010) and induces conditioned place preference
(Hipolito et al., 2011). Furthermore, rats self-administer
salsolinol directly into the pVTA, but not into the aVTA,
this self-administration being impaired by pre-treatment
with the D2 agonist quinpirole or the 5-HT3 antagonist
ICS205–930 (Rodd et al., 2008). The reinforcing proper-
ties of salsolinol are thus similar to those of acetaldehyde
and of ethanol itself but they can be seen at much lower
doses (Rodd-Henricks et al., 2000, 2002, 2003; Rodd
et al., 2004b, 2005d; Sanchez-Catalan et al., 2009), high-
lighting salsolinol as an important and likely candidate to
mediate the neurobiological effects of ethanol. (Table 1)
Neurochemical studies. While there is agreement that
ethanol administration into the aVTA does not influence
dopamine levels in the nucleus accumbens (Ericson
et al., 2008) (more specifically in the nucleus accumbens
shell (Ding et al., 2009b)), there are discrepancies
concerning the consequences of pVTA administration,
some authors observing increased dopamine levels in
the nucleus accumbens shell (Melis et al., 2007; Ding
et al., 2009b), whereas others did not observe changes
when sampling the transition zone between the core
and the shell (Ericson et al., 2008). The ethanol injection
into the pVTA but not the aVTA was also proposed to
increase extracellular dopamine levels in the ventral palli-
dum and in the medial prefrontal cortex (Ding et al.,
2011). When repeated, the infusion of ethanol into the
pVTA sensitizes the dopaminergic response, further
increasing extracellular dopamine levels in the nucleus
accumbens shell (Ding et al., 2009b); and cross-sensitiza-
tion between drugs of abuse can also be observed, as
repeated pVTA administration of nicotine increases the
effect of a pVTA ethanol administration on the nucleus
accumbens shell dopamine levels (Ding et al., 2012b).
In line with an interaction between ethanol action and
the cholinergic system, the antagonism of nicotinic recep-
tors by mecamylamine in the aVTA blocks the dopamine
level increase in the nucleus accumbens after intra-
accumbens ethanol perfusion (Ericson et al., 2008).
Interestingly, acetaldehyde and salsolinol
administered into the pVTA increase the extracellular
dopamine levels in the nucleus accumbens shell (Melis
et al., 2007; Hipolito et al., 2011; Deehan et al., 2013), sup-
porting the idea that the properties of these compounds are
similar to the ones of ethanol. The stimulation of the dopa-
minergic transmission by an intra-pVTA salsolinol adminis-
tration can be blocked by the pVTA pre-treatment with the
l-opioid antagonist b-funaltrexamine, which parallels the
behavioral action of salsolinol (Hipolito et al., 2011).
The pVTA dopamine levels are also associated with
spontaneous preference for alcohol. Indeed, the basal
pVTA dopamine levels are lower in a strain of alcohol-
preferring rats compared to their Wistar controls (Liu
et al., 2006). However, when exposed to alcohol drinking,
the alcohol preferring rats display increased basal pVTA
dopamine levels (Engleman et al., 2011). This could sug-
gest that the alcohol intake in these rats may be driven by
compensatory mechanisms in order to reach a higher
steady-state of dopaminergic activity. Such a hypothesis
is attractive and would be in agreement with a ‘‘self-med-
ication’’ hypothesis, suggesting that alcohol helps these
animals reach an optimized ‘‘hedonic set-point’’. Explor-
ing this hypothesis will require further research.
Finally, ethanol may not only recruit the dopamine
system. Acute and repeated systemic ethanol
administration as well as chronic ethanol drinking
increases the extracellular levels of glutamate in the
pVTA (Ding et al., 2012a, 2013).
Electrophysiological studies. Despite behavioral
evidence, electrophysiological studies do not usually
differentiate between the anterior and the pVTA
dopamine neurons. Some recent studies, however,
assessed the effects of ethanol and its derivates on
dopamine neurons of the aVTA and the pVTA. These
dopamine neurons have similar electrophysiological
features, although different responses to ethanol (Guan
et al., 2012). In ex vivo brain slices, ethanol inhibits the

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 203
dopamine neurons of the aVTA whereas it stimulates the
ones of the pVTA (Melis et al., 2007, 2013; Guan et al.,
2012). This latter effect is due to a reduction of the GAB-
Aergic influence on pVTA dopamine neurons (Guan et al.,
2012), which thus favors the disinhibition of dopamine
cells (Johnson and North, 1992). Likewise, voluntary eth-
anol intake increases the number of active dopamine neu-
rons in the pVTA of alcohol-preferring rats (Morzorati
et al., 2010). As this action of alcohol is associated with
a reduced pVTA sensitivity of N-methyl-D-aspartate
(NMDA) receptors in the pVTA, it is more likely due to a
disinhibitory mechanism rather than to the recruitment of
excitatory inputs (Fitzgerald et al., 2012).
Behavioral and neurochemical studies have shown
that acetaldehyde and salsolinol have stimulating
properties into the pVTA (Rodd-Henricks et al., 2002;
Rodd et al., 2008; Sanchez-Catalan et al., 2009; Hipolito
et al., 2010, 2011). Accordingly, it has been observed that
they enhance the activity of the pVTA dopamine neurons
(Melis et al., 2007; Xie et al., 2012; Xie and Ye, 2012).
Moreover, the acetaldehyde derived from ethanol has
been proposed to be responsible for the activating effect
of ethanol on dopamine cells (Melis et al., 2013). Salsolin-
ol was proposed to enhance dopamine activity through an
opioid-dependent mechanism (Xie et al., 2012), however
an action on VTA glutamate inputs was also suggested
(Xie and Ye, 2012).
Opioids
Opioids have both dopamine-dependent and dopamine-
independent activating and rewarding properties. In the
Fig. 2. Schematic of the manipulations of the aVTA and the pVTA sustaini
preference (CPP) and stimulating locomotor activity (activity), local elec
consequences of viral-mediated local expression of GluR1, PLCc, CREB an
bregma, which neuroanatomically corresponds to the presence of the
hydroxytryptamine; aVTA, anterior ventral tegmental area; CPA, conditioned
test; IP, interpeduncular nucleus; pVTA, posterior ventral tegmental area.
VTA, the opioids can recruit dopamine cells through a
disinhibitory mechanism (Johnson and North, 1992;
Jalabert et al., 2011). While some authors observed no
significant correlation between the VTA antero-posterior
placement of morphine injection and the resulting place
conditioning (De Jaeger et al., 2013), data from other
groups support the idea that opioids can have different
behavioral consequences depending on the antero-
posterior level within the VTA (Zangen et al., 2002;
Jhou et al., 2012). Indeed, endomorphin-1 (EM-1) induces
intracranial self-administration, conditioned place prefer-
ence and increases locomotor activity when administered
into the pVTA (Zangen et al., 2002; Terashvili et al., 2004)
(Fig. 2, Table 1), whereas it displays weaker intracranial
self-administration and stimulating effects and fails to
induce conditioned place preference when delivered into
the aVTA (Zangen et al., 2002).
Similar to ethanol studies, this aVTA/pVTA dichotomy
associated with local microinjections does not mean that
the aVTA cannot influence systemic drug response.
Indeed the reversible inactivation of the aVTA by
lidocaine decreases morphine-induced conditioned place
preference (Moaddab et al., 2009). Moreover, the inacti-
vation of the aVTA, but not of the pVTA, by GABA ago-
nists impairs heroin-conditioned immunomodulation
(Hutson et al., 2014). Last, the inactivation of AMPA
receptors by CNQX delivery into the aVTA blocks the
morphine-induced place conditioning and the heroin
self-administration, although it does not modulate those
behaviors (but it modulates the motor-activating effects)
when the AMPA antagonist is administered into the pVTA
(Shabat-Simon et al., 2008).
ng intracranial self-administration (ICSA), inducing conditioned place
trophysiological response of dopamine cells (DA), and behavioral
d mCREB. The aVTA/pVTA limit in rats is around �5.5 mm from the
interpeduncular nucleus below the VTA. Abbreviations: 5-HT, 5-
place aversion; D9THC, D9tetrahydrocannabinol; FST, forced swim

204 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
A set of studies using viral vector-mediated expression
of various proteins in the aVTAor in the pVTAare important
to mention here. Thus, the overexpression of the GluR1
subunit of the AMPA receptor (Carlezon et al., 2000), of
the phospholipase Cc (PLCc) which is implicated in the
intracellular cascade activated by neurotrophic factors
(Bolanos et al., 2003) or of the transcription factor cAMP
response element binding protein (CREB) (Olson et al.,
2005)modulates in oppositeways the rewarding properties
of a low dose of morphine depending whether this overex-
pression is performed in the aVTA or in the pVTA. When
GluR1, PLCc or CREB are overexpressed in the aVTA,
the rewarding properties ofmorphine as evaluated by place
conditioning are enhanced, while the same dose of mor-
phine becomes aversive if these proteins are overexpres-
sed in the pVTA (Carlezon et al., 2000; Bolanos et al.,
2003; Olson et al., 2005). These data are further reinforced
by the fact that the aVTA or the pVTA overexpression of
mCREB, a dominant negative form of CREB acting as an
antagonist, leads to consequences that are opposite to
the ones of CREB overexpression (Olson et al., 2005),
i.e. an aversion to a low dose of morphine if overexpressed
in aVTA and an enhancement of morphine reward if over-
expressed in the pVTA. The opposite action of the aVTA
and the pVTA manipulations on morphine place condition-
ing was particularly striking after the PLCc overexpression
(Bolanos et al., 2003); furthermore, the PLCc overexpres-
sion in the aVTA, but not in the pVTA, also enhanced
morphine-induced locomotor sensitization (Bolanos et al.,
2005). As neurotrophic factors in the VTA have been
related to the effects of drug of abuse (Nikulina et al.,
2014), the PLCc data suggest that the action of neurotro-
phic factors might also depend on the considered antero-
posterior level within the VTA.
Cholinergic system
The cholinergic system is a key modulator of the VTA
dopamine cells (Faure et al., 2014), but the behavioral
effect of cholinergic agents may vary depending on the
VTA subregions. Thus, nicotine, the cholinergic agonist
carbachol and the acetylcholinesterase inhibitor neostig-
mine support intracranial self-administration in the pVTA
of rats (Ikemoto and Wise, 2002; Ikemoto et al., 2003,
2006; Farquhar et al., 2012) (Fig. 2, Table 1), whereas
the intracranial self-administration of carbachol is weaker
(Ikemoto and Wise, 2002) and the intracranial self-admin-
istration of nicotine does not occur (Ikemoto et al., 2006)
into the aVTA.
The pVTA self-administration of carbachol is
prevented by a muscarinic (scopolamine), a nicotinic
(dihydro-b-erythroidine) or a D1 receptor (SCH23390)
antagonist (Ikemoto and Wise, 2002), and the pVTA
self-administration of nicotine by a nicotinic antagonist
(mecamylamine) and a D2 agonist (quinpirole) (Ikemoto
et al., 2006). Furthermore, the administration of carbachol
into the pVTA also induces locomotor activity, conditioned
place preference and c-Fos expression in several brain
regions (Ikemoto and Wise, 2002; Ikemoto et al., 2003;
Schifirnet et al., 2014). Some discrepancies are however
present concerning the ability of carbachol to induce con-
ditioned place preference into the aVTA (Ikemoto and
Wise, 2002; Schifirnet et al., 2014), which may be related
to the definition of ‘‘aVTA’’. Indeed, a recent study pro-
posed the presence of an aVTA (supporting carbachol
conditioned place preference), a mid-VTA (lacking such
conditioned response) and a pVTA (supporting carbachol
conditioned place preference) along the VTA antero-pos-
terior axis (Schifirnet et al., 2014). However, it may be dif-
ficult to discriminate aVTA from lateral hypothalamus
effects with very rostral injection sites.
The microinjection of a mGlu2/3 receptor agonist
(LY379268) or mGlu5 receptor antagonist (2-methyl-6-
(phenylethynyl)pyridine, MPEP) into the pVTA
decreases nicotine intravenous self-administration in
Wistar rats (Liechti et al., 2007; D’Souza and Markou,
2011), further highlighting the role of the pVTA in the rein-
forcing properties of nicotine.
Ex vivo electrophysiological studies have also
suggested that nicotine preferentially activates
dopamine neurons of the pVTA, whereas nicotine effect
is weaker on neurons from the aVTA, which is likely due
to a different expression of nicotinic receptors (Li et al.,
2011; Zhao-Shea et al., 2011).
Overall, these studies provide evidence of an
activating effect of cholinergic agonists in the VTA, and
most importantly within the pVTA.
Cocaine
Rodents self-administer cocaine into the pVTA (David
et al., 2004; Rodd et al., 2005a) (Fig. 2, Table 1), but
not in the aVTA (Rodd et al., 2005a). This pVTA self-
administration of cocaine can be blocked by systemic
pre-treatment with a D1 (SCH23390) or a 5HT1B
(GR127935) antagonist, by the co-infusion of a 5HT3
(ICS205–930) antagonist, or by the co-infusion of the
D2/3 agonist quinpirole (David et al., 2004; Rodd et al.,
2005a).
While the pVTA preferentially supports intracranial
cocaine self-administration, both the aVTA and the
pVTA can influence systemic cocaine effects. For
example, the inhibition of the aVTA (but not of the
pVTA) by muscimol increases the motivation for cocaine
intake (as shown by higher breaking-point in self-
administration), and also decreases the number of
cocaine self-infusions under a FR1 fixed ratio,
suggesting an enhanced rewarding effect of cocaine
(Lee et al., 2007). Moreover, cocaine place conditioning
can also be suppressed by the microinjection of a
l-opioid agonist into the aVTA, but not into the pVTA,
while microinjections into the pVTA, but not the aVTA,
suppress cocaine-induced locomotor activity (Soderman
and Unterwald, 2008). These data could suggest a possi-
ble dissociation between the pathways underlying the
rewarding and the locomotor effects of cocaine. However,
the dopaminergic lesion of the medial pVTA (sparing the
parabrachial pigmented nucleus of the VTA) (Ouachikh
et al., 2013), but not of the aVTA (Ouachikh et al.,
2014), impairs cocaine-induced conditioned place prefer-
ence. The respective roles of the aVTA and the pVTA in
cocaine responses are thus complex, which may also
reflect the complexity of the feed-forward and feed-back
circuitry of the ventral tegmentum and basal ganglia.

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 205
Similarly to morphine place conditioning, the aVTA
and the pVTA manipulations of the transcription factor
CREB have opposite actions on cocaine place
conditioning (Olson et al., 2005). A viral-mediated overex-
pression of CREB in the rat aVTA favors place preference
for a threshold dose of cocaine, while the same dose
becomes aversive if CREB is overexpressed in the pVTA;
and these effects are mirrored after the expression of the
dominant negative mCREB (Olson et al., 2005). This
study was particularly important as it also searched for
the limit between the aVTA and the pVTA in rats. For this,
the viral vectors for CREB or mCREB expression were
injected at different antero-posterior levels within the
VTA, and the response to cocaine was tested in the place
conditioning paradigm. An inflexion point was observed
between the rewarding and aversive effects of cocaine,
around �5.5 mm from the bregma, which neuroanatomi-
cally correspond to the location of the interpeduncular
nucleus below the VTA. This transition point is similar to
the one that was more recently observed between the
lack or presence of conditioned place preference after
intra-VTA carbachol administration (Schifirnet et al.,
2014).
Other drugs
An opposite modulation of brain stimulation reward has
been observed after blocking NMDA and AMPA
receptors in the VTA. The changes in reward threshold
positively correlated with the VTA antero-posterior
position of a NMDA antagonist injection, while it
negatively correlated with the antero-posterior position
of an AMPA antagonist injection (Ducrot et al., 2013).
The influence of AMPA antagonism was thus stronger in
the aVTA. Interestingly, intra-aVTA infusions of AMPA
itself were also reported to induce conditioned place
avoidance, while intra-pVTA AMPA infusions had no
effect (Ikemoto et al., 2004).
D9tetrahydrocannabinol can induce locomotor activity
and conditioned place preference and can support
intracranial self-administration when delivered into the
pVTA but not into the aVTA (Zangen et al., 2006)
(Table 1).
Affect and pain
Recent attention has been given to the heterogeneity of
the VTA cells (Roeper, 2013; Morales and Root, 2014)
and of the projection pathways originating from the VTA
(Lammel et al., 2014; Walsh and Han, 2014), particularly
concerning reward, aversion and depression. Similarly,
the aVTA/pVTA differences are not limited to the effect
of rewarding or locomotor activating substances (Figs. 1
and 2, Table 1). For example, the overexpression of
PLCc in the aVTA, but not in the pVTA, increased the
anxiety-like behavior in an elevated plus-maze, while the
pVTA overexpression, but not the aVTA one, decreased
the latency to immobility in the forced swim test
(Bolanos et al., 2003). These data partially dissociated
the influence on anxiety and depression-related behav-
iors, which may be further supported by the fact that the
aVTA overexpression of PLCc increased sucrose
preference, while the overexpression in the pVTA blunted
it. These findings should also be considered in the context
of VTA neurotrophic factors and mood disorders (Nikulina
et al., 2014; Walsh and Han, 2014), as PLCc participates
to the intracellular signaling cascade activated by these
factors.
Last, some data also report differences in the control
of pain-related responses along the antero-posterior
extent of the VTA. Thus, pVTA PLCc overexpression in
rats lowered the foot-shock thresholds inducing a jump
response, while an aVTA overexpression did not affect it
(Bolanos et al., 2003). Experiments on tail shocks’ thresh-
olds inducing vocalizations also showed aVTA/pVTA dif-
ferences in the control of such responses. The
microinjection of the cholinergic agonist carbachol into
the pVTA strongly increased the shock intensity neces-
sary to elicit a vocalization (Schifirnet et al., 2014), which
was prevented by either nicotinic (mecamylamine) or
muscarinic (atropine) antagonists. No influence was
observed at more anterior levels (designed in this article
as ‘‘midVTA’’), and an increase in the shock intensity elic-
iting vocalization was again observed more rostrally
(designed in this article as ‘‘aVTA’’) but in this case only
mecamylamine prevented it.
NEUROANATOMY OF THE VTA
All the behavioral findings reported above provide
convincing evidences for the functional antero-posterior
heterogeneity of the VTA. This raises the question of
the morphofunctional substrate of such heterogeneity.
Among the parameters to consider, the neuroanatomy
of the VTA (for review: (Yetnikoff et al., 2014)) is a critical
one.
Definition of the VTA
The first occurrence of the VTA in the literature is due to
Tsai in 1925 (Tsai, 1925). In its description of the opos-
sum brain, he identified with Nissl and Golgi staining a
region lateral to the interpeduncular nucleus as the trigo-
num interpeduncular. This region included the mamillary
peduncle, the medial lemniscus and the nucleus tegmenti
ventralis (presently VTA). This latter nucleus constituted
the medial part of the trigonum interpeduncular and it
spanned from the cerebral peduncle to the ventral tip of
the substantia nigra.
Before Tsai’s study, the fusiform aspect of the cells of
the nucleus tegmenti ventralis had led it to be considered
as a part of the substantia nigra (Kosaka and Hiraiwa,
1915; Castaldi, 1923). Accordingly, it has also been des-
ignated as the nucleus niger suboculomotorius (Hassler,
1937). However, the small size of the cells as well as
the close proximity to the mamillo- and the olfacto-
tegmentalis tracti suggested specific anatomical and
functional features for the nucleus tegmenti ventralis;
but whether the nucleus tegmentis ventralis belonged or
not to the substantia nigra remained an issue that was
long discussed. This debate was also due to the cytoar-
chitectonic heterogeneity of the region, and to the diffi-
culty to clearly differentiate it from the substantia nigra
compact part in primates as compared to rodents. In the
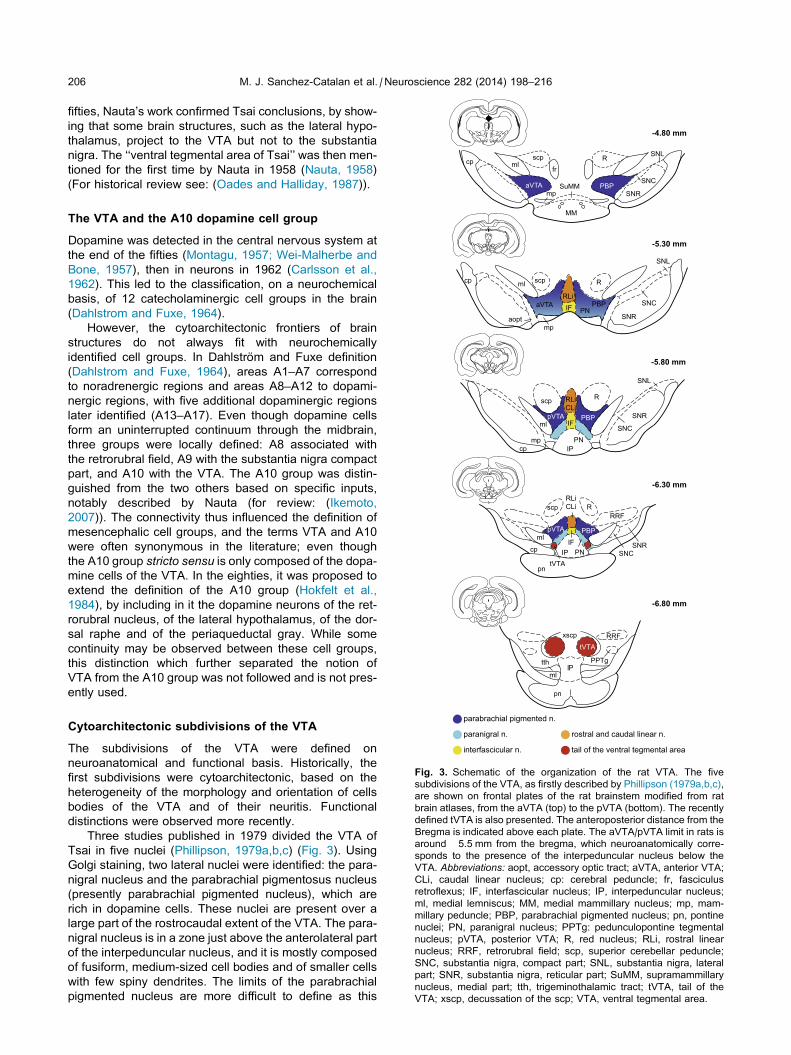
-4.80 mm
Rfr
SuMM
scpmlcp
aVTA
SNL
SNC
SNRmpPBP
206 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
fifties, Nauta’s work confirmed Tsai conclusions, by show-
ing that some brain structures, such as the lateral hypo-
thalamus, project to the VTA but not to the substantia
nigra. The ‘‘ventral tegmental area of Tsai’’ was then men-
tioned for the first time by Nauta in 1958 (Nauta, 1958)
(For historical review see: (Oades and Halliday, 1987)).
-5.30 mm
-5.80 mm
-6.30 mm
-6.80 mm
scp
aoptIF
RLi
PN
R
PBP
mlcp
mp
aVTA
SNR
SNC
SNL
Rscp
ml
cp IP
pVTA PBP
PN
IF
RLiCLi
mp
SNR
SNC
SNL
RRF
tVTA
Rscp
SNRSNC
mlcp
pn
PNIPIF
pVTA PBP
RLiCLi
tVTA
IP
pn
RRFxscp
PPTgtth
ml
MM
The VTA and the A10 dopamine cell groupDopamine was detected in the central nervous system at
the end of the fifties (Montagu, 1957; Wei-Malherbe and
Bone, 1957), then in neurons in 1962 (Carlsson et al.,
1962). This led to the classification, on a neurochemical
basis, of 12 catecholaminergic cell groups in the brain
(Dahlstrom and Fuxe, 1964).
However, the cytoarchitectonic frontiers of brain
structures do not always fit with neurochemically
identified cell groups. In Dahlstrom and Fuxe definition
(Dahlstrom and Fuxe, 1964), areas A1–A7 correspond
to noradrenergic regions and areas A8–A12 to dopami-
nergic regions, with five additional dopaminergic regions
later identified (A13–A17). Even though dopamine cells
form an uninterrupted continuum through the midbrain,
three groups were locally defined: A8 associated with
the retrorubral field, A9 with the substantia nigra compact
part, and A10 with the VTA. The A10 group was distin-
guished from the two others based on specific inputs,
notably described by Nauta (for review: (Ikemoto,
2007)). The connectivity thus influenced the definition of
mesencephalic cell groups, and the terms VTA and A10
were often synonymous in the literature; even though
the A10 group stricto sensu is only composed of the dopa-
mine cells of the VTA. In the eighties, it was proposed to
extend the definition of the A10 group (Hokfelt et al.,
1984), by including in it the dopamine neurons of the ret-
rorubral nucleus, of the lateral hypothalamus, of the dor-
sal raphe and of the periaqueductal gray. While some
continuity may be observed between these cell groups,
this distinction which further separated the notion of
VTA from the A10 group was not followed and is not pres-
ently used.
parabrachial pigmented n.
paranigral n.
interfascicular n.
rostral and caudal linear n.
tail of the ventral tegmental area
Fig. 3. Schematic of the organization of the rat VTA. The five
subdivisions of the VTA, as firstly described by Phillipson (1979a,b,c),
are shown on frontal plates of the rat brainstem modified from rat
brain atlases, from the aVTA (top) to the pVTA (bottom). The recently
defined tVTA is also presented. The anteroposterior distance from the
Bregma is indicated above each plate. The aVTA/pVTA limit in rats is
around �5.5 mm from the bregma, which neuroanatomically corre-
sponds to the presence of the interpeduncular nucleus below the
VTA. Abbreviations: aopt, accessory optic tract; aVTA, anterior VTA;
CLi, caudal linear nucleus; cp: cerebral peduncle; fr, fasciculus
retroflexus; IF, interfascicular nucleus; IP, interpeduncular nucleus;
ml, medial lemniscus; MM, medial mammillary nucleus; mp, mam-
millary peduncle; PBP, parabrachial pigmented nucleus; pn, pontine
nuclei; PN, paranigral nucleus; PPTg: pedunculopontine tegmental
nucleus; pVTA, posterior VTA; R, red nucleus; RLi, rostral linear
nucleus; RRF, retrorubral field; scp, superior cerebellar peduncle;
SNC, substantia nigra, compact part; SNL, substantia nigra, lateral
part; SNR, substantia nigra, reticular part; SuMM, supramammillary
nucleus, medial part; tth, trigeminothalamic tract; tVTA, tail of the
VTA; xscp, decussation of the scp; VTA, ventral tegmental area.
Cytoarchitectonic subdivisions of the VTA
The subdivisions of the VTA were defined on
neuroanatomical and functional basis. Historically, the
first subdivisions were cytoarchitectonic, based on the
heterogeneity of the morphology and orientation of cells
bodies of the VTA and of their neuritis. Functional
distinctions were observed more recently.
Three studies published in 1979 divided the VTA of
Tsai in five nuclei (Phillipson, 1979a,b,c) (Fig. 3). Using
Golgi staining, two lateral nuclei were identified: the para-
nigral nucleus and the parabrachial pigmentosus nucleus
(presently parabrachial pigmented nucleus), which are
rich in dopamine cells. These nuclei are present over a
large part of the rostrocaudal extent of the VTA. The para-
nigral nucleus is in a zone just above the anterolateral part
of the interpeduncular nucleus, and it is mostly composed
of fusiform, medium-sized cell bodies and of smaller cells
with few spiny dendrites. The limits of the parabrachial
pigmented nucleus are more difficult to define as this

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 207
subregion of the VTA is found dorsally and/or dorsolater-
ally to the paranigral nucleus depending on the antero-
posterior level. It also constitutes the lateral limit between
the VTA and the substantia nigra. One of the types of cell
bodies is in fact similar to the fusiform neurons found on
the dorsal tiers of the substantia nigra compact part.
Other neurons of the parabrachial pigmented nucleus
have a medium-sized globular cell body with numerous
radial dendrites. Three median nuclei have been pro-
posed in the VTA: the interfascicular, the rostral linear
and the caudal linear nuclei. The interfascicular nucleus
extends over the whole antero-posterior extent of the
VTA and it is composed of round and densely packed
cells, smaller than in the rest of the VTA. Overlying it in
its anterior part is the rostral linear nucleus, containing
the largest cells of the VTA, and in its posterior part, the
caudal linear nucleus.
This architectonic parcellation of the VTA into five sub-
regions still remains valid, but other subdivisions have
also been proposed (for review of VTA subdivisions,
see: (Halliday and Tork, 1986; Oades and Halliday,
1987; Fallon and Loughlin, 1995; Ikemoto, 2007)). In this
context, the recent work from Ikemoto is particularly sig-
nificant. Based solely on the cytoarchitectonic dimension
of the VTA, four lateral nuclei were proposed (Ikemoto,
2007). These include the classical paranigral nucleus
and parabrachial pigmented nucleus, rich in dopamine
cells (Phillipson, 1979a,b,c; Halliday and Tork, 1986;
Oades and Halliday, 1987), as well as two additional
nuclei: the parafascicular retroflexus area, and the tVTA
(Perrotti et al., 2005; Kaufling et al., 2009; Jhou et al.,
2009b). In this latter case, only the rostral third of what
is now known as the tVTA/RMTg was really described.
The parafascicular retroflexus area is the most anterior
part of the VTA, with a low density of dopamine cells, con-
tinuous with those of the lateral hypothalamus and of the
supramamillary nucleus. This area is thus composed of
the most anterior part of the paranigral nucleus and of
the parabrachial pigmented nucleus. These two nuclei
are indeed difficult to differentiate at this anterior level,
while they are clearly distinguished in the mid third of
the VTA. Then, in the posterior part of the VTA, another
group of cells localized dorsolaterally to the interpeduncu-
lar nucleus can be seen, based on a Nissl staining. This
region was previously included in the paranigral nucleus,
but data from the last decade converge to designate it as
a distinct region: the tVTA or RMTg (Perrotti et al., 2005;
Kaufling et al., 2009; Jhou et al., 2009a,b; Bourdy and
Barrot, 2012).
This recent analysis of VTA subregions (Ikemoto,
2007) also highlights a classical problem in the definition
of the VTA: do the median nuclei (the interfascicular,
the rostral linear and the caudal linear nuclei) really
belong to the VTA or should they be associated with other
structures such as the raphe nuclei? Different authors
(Swanson, 1982; Kalivas, 1993; Ikemoto, 2007) would
consider that these three median nuclei are not part of
the VTA. Thus, while the VTA and the A10 dopamine
group are often used as synonyms, the VTA would only
include the lateral nuclei (the paranigral nucleus and the
parabrachial pigmented nucleus), while the A10 group of
cell would also include the three median nuclei (Kalivas,
1993). Ikemoto also designated the rostral linear nucleus
and the caudal linear nucleus as raphe nuclei, which is
also the case since the 4th edition (1998) of the Paxinos
and Watson atlas of the rat brain (Paxinos and Watson,
1998, 2007). Last, it is to be noted that in the recent ver-
sions of the rat and of the mouse brain atlases (Paxinos
and Watson, 2007; Franklin and Paxinos, 2008) a parain-
terfascicular nucleus has also been identified in the mid to
posterior third of the VTA. The parainterfascicular nucleus
is wedged between the paranigral nucleus and the par-
abrachial pigmented nucleus. This name has then been
reused by different groups (over 35 references can be
presently retrieved under Google Scholar) in rats
(Colussi-Mas and Schenk, 2008; Nair-Roberts et al.,
2008; Wang and Morales, 2008; Brischoux et al., 2009;
Zhang et al., 2014) and in other species (Reyes et al.,
2012; Cavalcanti et al., 2014; Schweimer et al., 2014).
Accordingly, a recent cytoarchitectonic and chemoarchi-
tectonic reanalysis of the mouse midbrain dopamine cell
groups has been done (Fu et al., 2012).
Despite the obvious existence of different anatomical
subdivisions within the VTA, the question and the study
of their respective functions has not yet really been
addressed. This is likely due to the difficulty in
selectively manipulating these groups of cells, due to
their relatively small size, proximity and mostly shared
neurochemistry. Recent progress has allowed the study
of selected VTA cell populations based on their
neurochemistry and/or connectivity (for example
mesolimbic vs. mesocortical) (Roeper, 2013; Lammel
et al., 2014; Walsh and Han, 2014), leading to major sci-
entific advances. However, these studies did not differen-
tiate yet between VTA subnuclei. The search for specific
molecular markers of these sub-regions would allow
major progress in the field, by making possible to perform
promoter-driven selective targeting of VTA subregions.
BASES FOR THE aVTA/pVTA DIFFERENCES
The aVTA and the pVTA cannot be considered as
separate brain structures per se. The functional
differences that exist between them are thus likely to be
based on the preferential targeting of different VTA
subnuclei (see above), on differences in connectivity, on
differences in the proportion of the various cell types
(dopamine, GABA, glutamate, neuropeptides. . .), and on
the differential expression of membrane receptors and
channels.
Different afferents and/or efferents for the aVTA and
the pVTA are likely to contribute to the functional
differences. The inputs/outputs of the VTA are well
described (Yetnikoff et al., 2014), even though detailed
differences between VTA subnuclei or along the VTA
antero-posterior axis are yet to be completed. A widely
studied projection of the VTA dopamine cells is the ventral
striatum, with the nucleus accumbens constituting the
densest efferent target of VTA cells. This VTA-ventral stri-
atum projection displays a preferential, but not exclusive,
posteromedio-anterolateral topography (Phillipson and
Griffiths, 1985; Heimer et al., 1991; Berendse et al.,

208 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
1992; Haber and Fudge, 1997; Haber et al., 2000; Hasue
and Shammah-Lagnado, 2002; Ikemoto, 2007, 2010) that
could partly explain why the pVTA is a preferential sub-
strate for the intracranial self-administration of various
substances. The aVTA and lateral portions of the VTA
preferentially innervates the lateral part of the ventral stri-
atum, formed by the lateral olfactory tubercle and the lat-
eral parts of the shell and of the core of the nucleus
accumbens, whereas the pVTA and the medial portions
of the VTA preferentially projects to the medial part of
the ventral striatum, formed by the medial olfactory tuber-
cle and the medial shell of the nucleus accumbens
(Ungerstedt, 1971; Heimer et al., 1991, 1997; Berendse
et al., 1992; Usuda et al., 1998; Hasue and Shammah-
Lagnado, 2002; Zhou et al., 2003; Ikemoto, 2007,
2010). In the mouse, the antero-posterior analysis of the
VTA dopamine cells projecting to various forebrain
regions also showed a lack of projection from the cau-
dal-most part of the VTA to the lateral shell (Lammel
et al., 2008). A topographical organization is also
observed for the VTA projections to the dorsal striatum,
which mostly arise in the lateral part of the VTA and pref-
erentially the lateral aVTA (Ikemoto, 2007). Similarly, the
striato-pallidal projections to the VTA also display a topo-
graphical organization, with an antero-posterior gradient
and an inverted dorso-lateral gradient (for review see:
(Yetnikoff et al., 2014)). The relation between the VTA
and the prefrontal cortex still deserves a more detailed
analysis. The meso-accumbens and meso-cortical
pathways are mostly segregated (Deniau et al., 1980;
Fallon, 1981; Lammel et al., 2008). In the rat, meso-corti-
cal cells are distributed throughout the VTA (Deniau et al.,
1980), but non-dopamine meso-cortical cells (TH-nega-
tive projecting to the cingulate cortex) appear to be more
frequently observed in the aVTA (Swanson, 1982). In the
mouse, dopamine meso-cortical projections to the prelim-
bic and infralimbic cortices preferentially arise from the
medial pVTA, and non-dopamine ones from the aVTA
and from cells in the caudal-most part of the VTA (see
supplemental data in Lammel et al., 2008). The neuro-
chemical nature of VTA cell types may thus be important
to consider. Moreover, it would be useful to determine
whether the prelimbic, infralimbic and cingulate compo-
nents of the meso-cortical projections arise or not from
the same cells, and/or display a topographical organiza-
tion within the VTA. Differences in the serotonergic
(Herve et al., 1987) inputs have also been proposed, as
well as for cholinergic inputs from the laterodorsal teg-
mental nucleus that differently target meso-accumbens
and meso-cortical neurons (Omelchenko and Sesack,
2005). It is also noteworthy that some aspects of the
pVTA strongly innervate the aVTA, and that the paranigral
nucleus project to the interfascicular nucleus (Ferreira
et al., 2008), highlighting the existence of intra-VTA cir-
cuitries. However, there are few studies aimed at provid-
ing a precise analysis of input heterogeneity within the
VTA; and a comprehensive re-assessment of the VTA
subnuclei connectivity would be likely useful to the field.
The VTA contains a relatively large number of
neurons, between 10,000 and 20,000 dopamine
neurons unilaterally in the rat according to different
studies (Swanson, 1982; German and Manaye, 1993;
Nair-Roberts et al., 2008), constituting around 2/3 of the
VTA neuronal population (Swanson, 1982; German and
Manaye, 1993; Harris and Nestler, 1996; Nair-Roberts
et al., 2008). GABA cells and glutamate cells (Morales
and Root, 2014; Yetnikoff et al., 2014), including gluta-
mate-dopamine cells, are also present in the VTA. Thus,
a differential distribution of the VTA cell types along the
antero-posterior axis could be an important factor under-
lying the functional differences observed between the
aVTA and the pVTA after local manipulations. The distri-
bution of dopamine cells indeed differs along the antero-
posterior and medio-lateral axes (Swanson, 1982;
Fallon and Loughlin, 1995; Ikemoto, 2007; Nair-Roberts
et al., 2008). While the dopamine neurons are present
throughout the rostro-caudal extent of the VTA, they are
more prevalent in the pVTA. Tyrosine hydroxylase-immu-
noreactive cells are particularly dense in the antero-lateral
part of the pVTA, in the paranigral and the parabrachial
pigmented nuclei. This density progressively decreases
rostrally and caudally to this region. A decreasing latero-
medial gradient is also present; however, another region
rich in dopamine neurons is also observed in the midline
nuclei, corresponding to the interfascicular and the caudal
linear nuclei. These midline cell bodies are much smaller
but more densely packed than the lateral VTA neurons.
While the pVTA is richer in dopamine cells, the aVTA is
relatively richer in GABA neurons (Olson et al., 2005;
Ikemoto, 2007; Olson and Nestler, 2007) and the VTA
glutamate (VGluT2-positive) neurons are also more pres-
ent within the anterior midline nuclei even though that are
found in each VTA subnucleus (Morales and Root, 2014).
Different antero-posterior gradients between neuronal cell
types could partly explain differences in responses after
local drug administration.
It has also been proposed that neurons of the aVTA
and of the pVTA may express different types of
receptors or different subunits of a same receptor.
Inactivating GABAA receptors in aVTA is for example
reinforcing, while activating these receptors is
reinforcing in the pVTA (Ikemoto et al., 1997b,1998).
The perfusion of GABAA antagonists into the aVTA also
increases the extracellular levels of dopamine in the
nucleus accumbens, suggesting that GABAA receptors
tonically inhibit the aVTA dopamine neurons projecting
to the nucleus accumbens (Ikemoto et al., 1997a), and
more specifically to the nucleus accumbens shell (Ding
et al., 2009a). Thus, GABAA receptors might be differently
localized in the aVTA and in the pVTA, being on dopa-
mine neurons in the aVTA but on inhibitory interneurons
in the pVTA. However, such differences in the location
of the GABA receptors has not been demonstrated, and
another explanation could be that intra-pVTA GABA
agents could also modulate nearby tVTA GABA neurons
that are known to control the activity of midbrain dopa-
mine cells (Barrot et al., 2012; Bourdy and Barrot,
2012). The pVTA dopamine neurons projecting to the
nucleus accumbens seem to be under tonic inhibition
through D2 receptors, since the pVTA administration of
the D2 antagonist sulpiride increases the extracellular
dopamine levels in nucleus accumbens (shell and core),

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 209
but also locally into the pVTA (Ding et al., 2009a). It has
also been shown that the activation of 5HT3 receptors
has a stronger action on dopamine levels in the pVTA
than in the aVTA (Liu et al., 2006). The aVTA/pVTA differ-
ences in behavior, or in changes in extracellular dopa-
mine levels or dopamine cell activity, that are observed
after local manipulation of GABA, dopamine, serotonin,
cannabinoid, opioid and nicotinic receptors (see the
‘‘VTA antero-posterior functional heterogeneity’’ section)
might reflect some differences in the distribution of these
receptors, but specific analyses are still needed to evalu-
ate this.
THE tVTA
In the past decade, a new mesopontine region associated
to the VTA has been defined: the tVTA or RMTg (Perrotti
et al., 2005; Kaufling et al., 2009; Jhou et al., 2009a,b;
Barrot and Thome, 2011; Bourdy and Barrot, 2012). As
the tVTA is partially embedded in the paranigral nucleus
of the VTA and exerts a major control on the activity of
dopamine cells, it is likely a new actor to consider when
studying and/or interpreting functional antero-posterior
differences of the VTA.
Detecting the tVTA
Experimentally, the easiest way to visualize the tVTA in
rats is to expose them acutely or chronically, either
through injections or self-administration procedures, to a
psychostimulant drug, such as cocaine, amphetamines
(D-amphetamine, methamphetamine, or (±)-3,4-methylen-
edioxymethamphetamine (MDMA)) or even modafinil,
and to process the midbrain for immunohistochemistry
against either cFos or FosB/DFosB (Scammell et al.,
2000; Perrotti et al., 2005; Geisler et al., 2008; Jhou
et al., 2009a,b; Kaufling et al., 2009, 2010a,b; Rotllant
et al., 2010; Zahm et al., 2010; Lecca et al., 2011; Matsui
and Williams, 2011; Cornish et al., 2012; Lavezzi et al.,
2012). Within the posterior part of the VTA, a cluster of
Fos-positive nuclei can be observed after exposure to
any of these drugs. This cluster is located dorso-laterally
tVTA
Fig. 4. Schematic of the consequences of tVTA manipulations. Abbreviatconditioned place preference; DA, dopamine; FC, fear conditioning; ICSA,
posterior VTA; tVTA, tail of the VTA; VTA, ventral tegmental area.
to the interpeduncular nucleus, in a subpart of the region
that is designated as the paranigral nucleus in the rat brain
atlas (Paxinos and Watson, 2007) (Figs. 3 and 4). Both
immunohistochemistry against the GABA synthesizing
enzyme, glutamic acid decarboxylase (GAD67) (Olson
andNestler, 2007), andNissl staining (Ikemoto, 2007) sup-
port the idea that this cluster of cells should be differenti-
ated from the surrounding paranigral nucleus. This highly
localized Fos staining extends caudally, beyond borders
of the VTA as defined by 6th edition of the rat brain atlas
(Paxinos and Watson, 2007). In fact, some difference is
even present between this continuous cell group and the
posterior-most drawings of the ‘‘VTA’’ in this atlas. Indeed,
the Fos-stained cell group shifts dorsally along its antero-
posterior axis (Kaufling et al., 2009, 2010b) (Figs. 3 and
4), following the position of the superior cerebellar pedun-
cle fibers, laterally to the median raphe nuclei. At some
point it is even partly embeddedwithin the superior cerebel-
lar peduncle. At this level, it might correspond to the ‘‘inter-
stitial nucleus of the decussation of the superior cerebellar
peduncle’’ newly proposed in the plates 89–92 of the rat
brain atlas (Paxinos andWatson, 2007). This mesopontine
group of cells, starting within the pVTA and spanning over
1 mm along the antero-posterior axis in rats, corresponds
to the tVTA or RMTg (Perrotti et al., 2005; Kaufling et al.,
2009; Jhou et al., 2009a,b; Lavezzi and Zahm, 2011;
Barrot et al., 2012; Bourdy and Barrot, 2012).
The psychostimulant-induced Fos staining allows an
easy visualization of the tVTA because surrounding
structures are unstained and because almost no staining
is present in the tVTA of control animals. It is to be noted
that the Fos induction in the tVTA is selectively observed
after exposure to arousing psychostimulant drugs, but it is
not present after exposure to various other drugs. Indeed,
the opiate morphine, ethanol, the benzodiazepine
diazepam, the cannabinoid D9-tetrahydrocannabinol, the
NMDA antagonists and dissociative drugs ketamine and
phencyclidine (PCP), the antidepressants reboxetine,
nortriptyline, venlafaxine, and fluoxetine, the 5-HT
releaser dexfenfluramine, and the anticonvulsants
valproic acid and gabapentin, do not induce Fos in the
tVTA (Perrotti et al., 2005; Kaufling et al., 2010b). A lack
ions: aVTA, anterior VTA; CPA, conditioned place aversion; CPP,
intracranial self-administration; IP, interpeduncular nucleus; pVTA,

210 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
of tVTA Fos expression is also observed after exposure to
stress (Perrotti et al., 2005), to the exception of electric
foot-shocks that induce a tVTA Fos response (Jhou et al.,
2009a; Brown and Shepard, 2013).
Based on connectivity, the tVTA has also been
observed in monkeys (Hong et al., 2011) and in mice
(Stamatakis and Stuber, 2012; Wasserman et al., 2013).
However, the borders of this structure are not yet been
clearly defined in these species, and the psychostimu-
lant-induced Fos that is often used in rats might be less
pertinent to visualize the tVTA in mice (unpublished
observations).
tVTA control of dopamine cells
The tVTA is mostly composed of GABA neurons (Perrotti
et al., 2005; Olson and Nestler, 2007; Kaufling et al.,
2009; Jhou et al., 2009b; Balcita-Pedicino et al., 2011),
with a notable expression of the l-opioid receptor (Jhou
et al., 2009b; Jalabert et al., 2011). Afferents of the tVTA
mostly arise in brain regions that also project to the VTA
(Kaufling et al., 2009; Jhou et al., 2009b); even though
inputs to the VTA and to the tVTA likely originate from dif-
ferent cell populations within these brain regions. For
example, the tVTA inputs from the dorsal raphe and the
locus cœruleus are non-aminergic and those from the lat-
eral hypothalamus are mostly non-orexinergic (Kaufling
et al., 2009), and the VTA and tVTA inputs from the lateral
habenula preferentially arise from its medial and lateral
subdivisions respectively (Goncalves et al., 2012). Pres-
ently, the most studied input to the tVTA remains the lat-
eral habenula (Herkenham and Nauta, 1979; Jhou et al.,
2009a, 2013; Brinschwitz et al., 2010; Balcita-Pedicino
et al., 2011; Hong et al., 2011; Goncalves et al., 2012;
Lammel et al., 2012; Stamatakis and Stuber, 2012;
Good et al., 2013; Stamatakis et al., 2013). The output
from the tVTA targets relatively few forebrain regions.
One of the main forebrain outputs is the lateral hypothal-
amus. The efferents of the tVTA are more prominently
directed toward the brainstem, in particular toward the
dopamine cell areas (the VTA, the substantia nigra com-
pact part, and to a lesser extent the retrorubral field)
(Ferreira et al., 2008; Jhou et al., 2009a,b; Kaufling
et al., 2010a; Balcita-Pedicino et al., 2011; Bourdy and
Barrot, 2012). This connectivity and the fact that tVTA
fibers do form synapses with dopamine cells of the VTA
(Balcita-Pedicino et al., 2011) and of the substantia nigra
compact part (Bourdy et al., 2014) and the results of elec-
trophysiological analyses support the hypothesis that the
tVTA may be an inhibitory control center for dopamine cell
activity (Hong et al., 2011; Jalabert et al., 2011, 2012;
Lecca et al., 2011; Matsui and Williams, 2011; Bourdy
et al., 2014).
The stimulation of the tVTA inhibits midbrain
dopamine cells, while its inhibition increases their firing
(Hong et al., 2011; Jalabert et al., 2011; Lecca et al.,
2011, 2012; Matsui and Williams, 2011; Melis et al.,
2013; Bourdy et al., 2014), which indicates the presence
of both phasic and tonic controls. This inhibitory control
can also be recruited by the stimulation of the lateral
habenula inputs to the tVTA (Hong et al., 2011; Lammel
et al., 2012; Stamatakis and Stuber, 2012), and a
VTA-lateral habenula-tVTA feedback loop has been
recently proposed (Good et al., 2013; Jhou et al., 2013;
Stamatakis et al., 2013).
Both morphine and the l-opioid agonist, DAMGO, can
inhibit the tVTA cells (Lecca et al., 2011; Matsui and
Williams, 2011; Matsui et al., 2014). Moreover, the
l-opioid receptors expressed by the tVTA GABA cells
and their terminals within the VTA are critical to the acute
recruitment of dopamine neurons by opiates (Jalabert
et al., 2011; Matsui and Williams, 2011; Matsui et al.,
2014). This led to the proposal that the classic disinhibi-
tion model for acute opiate action on dopamine cells
should be updated (Johnson and North, 1992; Barrot
et al., 2012; Bourdy and Barrot, 2012). The inhibitory
action of a cannabinoid agonist on tVTA neurons (Lecca
et al., 2011., 2012) supports the idea that the role of the
tVTA in disinhibition models could also be extended to
cannabinoids.
It remains to be explored whether the tVTA exerts a
differential inhibitory control on the aVTA and on the
pVTA, which could also participate to VTA antero-
posterior functional heterogeneity. Moreover, no
information is available yet on developmental aspects
of the tVTA, either concerning the origin of the tVTA
cells or the development of the tVTA inhibitory
innervations of the VTA and of the substantia nigra.
Such information would be useful to the field, for
example to appreciate whether the tVTA participates to
the known heterogeneity in dopamine neuron activity
across age or across individuals (Marinelli and
McCutcheon, 2014).
tVTA and behavior
The above data are supportive of a particular role of the
tVTA in the response to opiates. This role is behaviorally
confirmed by the fact that rats preferentially self-
administer EM-1 and develop conditioned place
preference to this opioid when it is microinjected into
the tVTA (Jhou et al., 2012) but not into surrounding
regions. The local administration of EM-1 likely inhibits
tVTA GABA cells. Accordingly, the inhibition of the tVTA
by local muscimol administration also sustains self-
administration (Jhou et al., 2012). For both EM-1 and
muscimol, this reinforcing effect decreases with the dis-
tance of the cannula from the tVTA core. These findings
led to reconsideration of previous reports of intra-pVTA
muscimol (Ikemoto et al., 1998) and EM-1 (Zangen
et al., 2002; Terashvili et al., 2004) reinforcing and
rewarding properties (see section on ‘‘VTA antero-pos-
terior functional heterogeneity’’), and suggest that these
properties might in fact have been due to an inhibition
of what has been designed as the tVTA a few years
later. The question is open as to whether other findings
previously associated with the manipulation of the pVTA
should be revisited by taking the tVTA into consideration
(Fig. 4, Table 1).
The tVTA has also been associated with the coding of
errors in reward prediction (Hong et al., 2011), but the
largest set of behavioral data in relation to the tVTA con-
cerns aversive and avoidance-related responses. Lesion
of the tVTA inhibits fear-conditioned freezing, passive

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 211
(but not active) response to a predator odor, and the anx-
iety-like behavior in an elevated plus-maze (Jhou et al.,
2009a). It also suppresses cocaine-induced avoidance
behavior in a runway operant paradigm (Jhou et al.,
2013). Reciprocally, the pharmacological stimulation of
the tVTA by AMPA is sufficient to induce a conditioned
place aversion (Jhou et al., 2013), and the stimulation of
the lateral habenula terminals in a midbrain region that
includes the tVTA induces active, passive and condi-
tioned avoidance of the stimulation (Lammel et al.,
2012; Stamatakis and Stuber, 2012).
It should also be considered that the tVTA control not
only the VTA, but also the substantia nigra compact part.
In this context, a lesion of the tVTA chronically stimulates
the nigrostriatal pathway and increases the motor
coordination performances and the motor learning in a
rotarod task (Bourdy et al., 2014). These last data high-
light the critical influence that the tVTA has on basal gan-
glia circuitry and function.
CONCLUSION
There is strong evidence for a functional antero-posterior
heterogeneity of the VTA. However, the morphofunctional
substrate(s) for this heterogeneity is(are) not clearly
identified yet. Differences related to the sub-regions of
the VTA, including the recently defined tVTA, and how
they may be differently inserted into larger brain
circuitries is likely critical. The neurochemical and
molecular heterogeneity of VTA cell populations are also
important to be considered. Important progress,
benefiting from technical advances, has recently been
made in relation to some aspects of VTA heterogeneity.
Many questions remain unanswered. Detailed re-
assessments of the VTA cytoarchitecture across
species, of the definition of its sub-nuclei, of their cell
composition and of their connectivity and of the
molecular heterogeneity of these cells, are required.
These, together with the search for specific molecular
markers that would also allow designing transgenic
tools, are among challenges that need to be addressed
in order to progress in our understanding of the
physiological and pathophysiological influences of the
VTA.
Acknowledgments—Supported by the Centre National de la
Recherche Scientifique, Universite de Strasbourg, Universite
Bordeaux Segalen, and by the Agence Nationale de la Recher-
che (ANR-11-bsv4-002). We thank Pr. Paul Bolam for his com-
ments on the manuscript.
REFERENCES
Arnt J, Scheel-Kruger J (1979) GABA in the ventral tegmental area:
differential regional effects on locomotion, aggression and food
intake after microinjection of GABA agonists and antagonists. Life
Sci 25:1351–1360.
Balcita-Pedicino JJ, Omelchenko N, Bell R, Sesack SR (2011) The
inhibitory influence of the lateral habenula on midbrain dopamine
cells: ultrastructural evidence for indirect mediation via the
rostromedial mesopontine tegmental nucleus. J Comp Neurol
519:1143–1164.
Barrot M, Thome J (2011) Discovering a new anatomical structure in
the brain: implications for neuropsychiatry and therapy. World J
Biol Psychiatry 12(S1):19–22.
Barrot M, Sesack SR, Georges F, Pistis M, Hong S, Jhou TC (2012)
Braking dopamine systems: a new GABA master structure for
mesolimbic and nigrostriatal functions. J Neurosci
32:14094–14101.
Bechtholt AJ, Cunningham CL (2005) Ethanol-induced conditioned
place preference is expressed through a ventral tegmental area
dependent mechanism. Behav Neurosci 119:213–223.
Berendse HW, Groenewegen HJ, Lohman AH (1992) Compartmental
distribution of ventral striatal neurons projecting to the
mesencephalon in the rat. J Neurosci 12:2079–2103.
Boehm 2nd SL, Piercy MM, Bergstrom HC, Phillips TJ (2002) Ventral
tegmental area region governs GABA(B) receptor modulation of
ethanol-stimulated activity in mice. Neuroscience 115:185–200.
Bolanos CA, Perrotti LI, Edwards S, Eisch AJ, Barrot M, Olson VG,
Russell DS, Neve RL, Nestler EJ (2003) Phospholipase Cgamma
in distinct regions of the ventral tegmental area differentially
modulates mood-related behaviors. J Neurosci 23:7569–7576.
Bolanos CA, Neve RL, Nestler EJ (2005) Phospholipase C gamma in
distinct regions of the ventral tegmental area differentially
regulates morphine-induced locomotor activity. Synapse
56:166–169.
Bourdy R, Barrot M (2012) A new control center for dopaminergic
systems: pulling the VTA by the tail. Trends Neurosci
35:681–690.
Bourdy R, Sanchez-Catalan MJ, Kaufling J, Balcita-Pedicino JJ,
Freund-Mercier MJ, Veinante P, Sesack SR, Georges F, Barrot
M (2014) Control of the nigrostriatal dopamine neuron activity and
motor function by the tail of the ventral tegmental area.
Neuropsychopharmacology. http://dx.doi.org/10.1038/npp.2014.
129.
Brinschwitz K, Dittgen A, Madai VI, Lommel R, Geisler S, Veh RW
(2010) Glutamatergic axons from the lateral habenula mainly
terminate on gabaergic neurons of the ventral midbrain.
Neuroscience 168:463–476.
Brischoux F, Chakraborty S, Brierley DI, Ungless MA (2009) Phasic
excitation of dopamine neurons in ventral VTA by noxious stimuli.
Proc Natl Acad Sci U S A 106:4894–4899.
Brodie MS, Appel SB (1998) The effects of ethanol on dopaminergic
neurons of the ventral tegmental area studied with intracellular
recording in brain slices. Alcohol Clin Exp Res 22:236–244.
Bromberg-Martin ES, Matsumoto M, Hikosaka O (2010) Dopamine in
motivational control: rewarding, aversive, and alerting. Neuron
68:815–834.
Brown PL, Shepard PD (2013) Lesions of the fasciculus retroflexus
alter footshock-induced cFos expression in the mesopontine
rostromedial tegmental area of rats. PLoS One 8:e60678.
Carlezon Jr WA, Haile CN, Coppersmith R, Hayashi Y, Malinow R,
Neve RL, Nestler EJ (2000) Distinct sites of opiate reward and
aversion within the midbrain identified using a herpes simplex
virus vector expressing GluR1. J Neurosci 20:RC62.
Carlsson A, Falck B, Hillarp NA (1962) Cellular localization of brain
monoamines. Acta Physiol Scand Suppl 56:1–28.
Castaldi L (1923)Studdi sulla strutturae sulosviluppodelmesencefalo. I
ricerche in Cavia cobaya. Arch Ital Embrio 20:23–225.
Cavalcanti JR, Soares JG, Oliveira FG, Guzen FP, Pontes AL, Sousa
TB, Cavalcante JS, Nascimento Jr ES, Cavalcante JC, Costa MS
(2014) A cytoarchitectonic and TH-immunohistochemistry
characterization of the dopamine cell groups in the substantia
nigra, ventral tegmental area and retrorubral field in the rock cavy
(Kerodon rupestris). J Chem Neuroanat 55:58–66.
Colussi-Mas J, Schenk S (2008) Acute and sensitized response to
3,4-methylenedioxymethamphetamine in rats: different behavioral
profiles reflected in different patterns of Fos expression. Eur J
Neurosci 28:1895–1910.
Cornish JL, Hunt GE, Robins L, McGregor IS (2012) Regional c-Fos
and FosB/DeltaFosB expression associated with chronic
methamphetamine self-administration and methamphetamine-
seeking behavior in rats. Neuroscience 206:100–114.

212 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
Correa M, Salamone JD, Segovia KN, Pardo M, Longoni R, Spina L,
Peana AT, Vinci S, Acquas E (2012) Piecing together the puzzle
of acetaldehyde as a neuroactive agent. Neurosci Biobehav Rev
36:404–430.
Creed MC, Ntamati NR, Tan KR (2014) VTA GABA neurons
modulate specific learning behaviors through the control of
dopamine and cholinergic systems. Front Behav Neurosci 8:8.
Czachowski CL, Delory MJ, Pope JD (2012) Behavioral and
neurotransmitter specific roles for the ventral tegmental area in
reinforcer-seeking and intake. Alcohol Clin Exp Res
36:1659–1668.
D’Souza MS, Markou A (2011) Metabotropic glutamate receptor 5
antagonist 2-methyl-6-(phenylethynyl)pyridine (MPEP)
microinfusions into the nucleus accumbens shell or ventral
tegmental area attenuate the reinforcing effects of nicotine in
rats. Neuropharmacology 61:1399–1405.
Dahlstrom A, Fuxe K (1964) Localization of monoamines in the lower
brain stem. Experientia 20:398–399.
David V, Segu L, Buhot MC, Ichaye M, Cazala P (2004) Rewarding
effects elicited by cocaine microinjections into the ventral
tegmental area of C57BL/6 mice: involvement of dopamine D1
and serotonin1B receptors. Psychopharmacology (Berl)
174:367–375.
De Jaeger X, Bishop SF, Ahmad T, Lyons D, Ng GA, Laviolette SR
(2013) The effects of AMPA receptor blockade in the prelimbic
cortex on systemic and ventral tegmental area opiate reward
sensitivity. Psychopharmacology (Berl) 225:687–695.
Deehan Jr GA, Engleman EA, Ding ZM, McBride WJ, Rodd ZA
(2013) Microinjections of acetaldehyde or salsolinol into the
posterior ventral tegmental area increase dopamine release in
the nucleus accumbens shell. Alcohol Clin Exp Res 37:722–729.
Deniau JM, Thierry AM, Feger J (1980) Electrophysiological
identification of mesencephalic ventromedial tegmental (VMT)
neurons projecting to the frontal cortex, septum and nucleus
accumbens. Brain Res 189:315–326.
Ding ZM, Liu W, Engleman EA, Rodd ZA, McBride WJ (2009a)
Differential effects of dopamine D2 and GABA(A) receptor
antagonists on dopamine neurons between the anterior and
posterior ventral tegmental area of female Wistar rats. Pharmacol
Biochem Behav 92:404–412.
Ding ZM, Rodd ZA, Engleman EA, McBride WJ (2009b) Sensitization
of ventral tegmental area dopamine neurons to the stimulating
effects of ethanol. Alcohol Clin Exp Res 33:1571–1581.
Ding ZM, Toalston JE, Oster SM, McBride WJ, Rodd ZA (2009c)
Involvement of local serotonin-2A but not serotonin-1B receptors
in the reinforcing effects of ethanol within the posterior ventral
tegmental area of female Wistar rats. Psychopharmacology (Berl)
204:381–390.
Ding ZM, Oster SM, Hall SR, Engleman EA, Hauser SR, McBride WJ,
Rodd ZA (2011) The stimulating effects of ethanol on ventral
tegmental area dopamine neurons projecting to the ventral
pallidum and medial prefrontal cortex in female Wistar rats:
regional difference and involvement of serotonin-3 receptors.
Psychopharmacology (Berl) 216:245–255.
Ding ZM, Engleman EA, Rodd ZA, McBride WJ (2012a) Ethanol
increases glutamate neurotransmission in the posterior ventral
tegmental area of female Wistar rats. Alcohol Clin Exp Res
36:633–640.
Ding ZM, Katner SN, Rodd ZA, Truitt W, Hauser SR, Deehan Jr GA,
Engleman EA, McBride WJ (2012b) Repeated exposure of the
posterior ventral tegmental area to nicotine increases the
sensitivity of local dopamine neurons to the stimulating effects
of ethanol. Alcohol 46:217–223.
Ding ZM, Oster SM, Hauser SR, Toalston JE, Bell RL, McBride WJ,
Rodd ZA (2012c) Synergistic self-administration of ethanol and
cocaine directly into the posterior ventral tegmental area:
involvement of serotonin-3 receptors. J Pharmacol Exp Ther
340:202–209.
Ding ZM, Rodd ZA, Engleman EA, Bailey JA, Lahiri DK, McBride WJ
(2013) Alcohol drinking and deprivation alter basal extracellular
glutamate concentrations and clearance in the mesolimbic system
of alcohol-preferring (P) rats. Addict Biol 18:297–306.
Ding ZM, Ingraham CM, Rodd ZA, McBride WJ (2014) The
reinforcing effects of ethanol within the posterior ventral
tegmental area depend on dopamine neurotransmission to
forebrain cortico-limbic systems. Addict Biol. http://dx.doi.org/
10.1111/adb.12138.
Ducrot C, Fortier E, Bouchard C, Rompre PP (2013) Opposite
modulation of brain stimulation reward by NMDA and AMPA
receptors in the ventral tegmental area. Front Syst Neurosci 7:57.
Engleman EA, Keen EJ, Tilford SS, Thielen RJ, Morzorati SL (2011)
Ethanol drinking reduces extracellular dopamine levels in the
posterior ventral tegmental area of nondependent alcohol-
preferring rats. Alcohol 45:549–557.
Ericson M, Lof E, Stomberg R, Chau P, Soderpalm B (2008) Nicotinic
acetylcholine receptors in the anterior, but not posterior, ventral
tegmental area mediate ethanol-induced elevation of accumbal
dopamine levels. J Pharmacol Exp Ther 326:76–82.
Fallon JH (1981) Collateralization of monoamine neurons:
mesotelencephalic dopamine projections to caudate, septum,
and frontal cortex. J Neurosci 1:1361–1368.
Fallon JH, Loughlin SE (1995) Substantia nigra. In: Paxinos G, editor.
The rat nervous system, 2nd edition. San Diego: Academic Press.
Farquhar MJ, Latimer MP, Winn P (2012) Nicotine self-administered
directly into the VTA by rats is weakly reinforcing but has strong
reinforcement enhancing properties. Psychopharmacology (Berl)
220:43–54.
Faure P, Tolu S, Valverde S, Naude J (2014) Role of nicotinic
acetylcholine receptors in regulating dopamine neuron activity.
Neuroscience 282:86–100.
Ferreira JG, Del-Fava F, Hasue RH, Shammah-Lagnado SJ (2008)
Organization of ventral tegmental area projections to the ventral
tegmental area-nigral complex in the rat. Neuroscience
153:196–213.
Fields HL, Hjelmstad GO, Margolis EB, Nicola SM (2007) Ventral
tegmental area neurons in learned appetitive behavior and
positive reinforcement. Annu Rev Neurosci 30:289–316.
Fitzgerald GJ, Liu H, Morzorati SL (2012) Decreased sensitivity of
NMDA receptors on dopaminergic neurons from the posterior
ventral tegmental area following chronic nondependent alcohol
consumption. Alcohol Clin Exp Res 36:1710–1719.
Ford CP (2014) The role of D2-autoreceptors in regulating dopamine
neuron activity and transmission. Neuroscience 282:13–22.
Franklin KBJ, Paxinos G (2008) Mouse in brain stereotaxic
coordinates, 3rd edition. San Diego, CA: Academic Press.
Fu Y, Yuan Y, Halliday G, Rusznak Z, Watson C, Paxinos G (2012) A
cytoarchitectonic and chemoarchitectonic analysis of the
dopamine cell groups in the substantia nigra, ventral tegmental
area, and retrorubral field in the mouse. Brain Struct Funct
217:591–612.
Geisler S, Marinelli M, Degarmo B, Becker ML, Freiman AJ, Beales
M, Meredith GE, Zahm DS (2008) Prominent activation of
brainstem and pallidal afferents of the ventral tegmental area by
cocaine. Neuropsychopharmacology 33:2688–2700.
German DC, Manaye KF (1993) Midbrain dopaminergic neurons
(nuclei A8, A9, and A10): three-dimensional reconstruction in the
rat. J Comp Neurol 331:297–309.
Gianoulakis C (2009) Endogenous opioids and addiction to alcohol
and other drugs of abuse. Curr Top Med Chem 9:999–1015.
Gillies GE, Virdee K, McArthur S, Dalley JW (2014) Sex-dependent
diversity in ventral tegmental dopaminergic neurons and
developmental programing: a molecular, cellular and behavioral
analysis. Neuroscience 282:69–85.
Goncalves L, Sego C, Metzger M (2012) Differential projections from
the lateral habenula to the rostromedial tegmental nucleus and
ventral tegmental area in the rat. J Comp Neurol 520:1278–1300.
Good CH, Wang H, Chen YH, Mejias-Aponte CA, Hoffman AF,
Lupica CR (2013) Dopamine D-4 receptor excitation of lateral
habenula neurons via multiple cellular mechanisms. J Neurosci
33:16853–16864.

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 213
Grace AA, Floresco SB, Goto Y, Lodge DJ (2007) Regulation of firing
of dopaminergic neurons and control of goal-directed behaviors.
Trends Neurosci 30:220–227.
Guan Y, Xiao C, Krnjevic K, Xie G, Zuo W, Ye JH (2012) GABAergic
actions mediate opposite ethanol effects on dopaminergic
neurons in the anterior and posterior ventral tegmental area. J
Pharmacol Exp Ther 341:33–42.
Haber SN, Fudge JL (1997) The primate substantia nigra and VTA:
integrative circuitry and function. Crit Rev Neurobiol 11:323–342.
Haber SN, Fudge JL, McFarland NR (2000) Striatonigrostriatal
pathways in primates form an ascending spiral from the shell to
the dorsolateral striatum. J Neurosci 20:2369–2382.
Halliday GM, Tork I (1986) Comparative anatomy of the ventromedial
mesencephalic tegmentum in the rat, cat, monkey and human. J
Comp Neurol 252:423–445.
Harris HW, Nestler EJ (1996) Immunohistochemical studies of
mesolimbic dopaminergic neurons in Fischer 344 and Lewis
rats. Brain Res 706:1–12.
Hassler R (1937) Zur pathologie der paralysis agitans und des
postenkephalitischer parkonsonismus. J Psychol Neurol
48:387–476.
Hasue RH, Shammah-Lagnado SJ (2002) Origin of the dopaminergic
innervation of the central extended amygdala and accumbens
shell: a combined retrograde tracing and immunohistochemical
study in the rat. J Comp Neurol 454:15–33.
Hauser SR, Ding ZM, Getachew B, Toalston JE, Oster SM, McBride
WJ, Rodd ZA (2011) The posterior ventral tegmental area
mediates alcohol-seeking behavior in alcohol-preferring rats. J
Pharmacol Exp Ther 336:857–865.
Hauser SR, Deehan Jr GA, Toalston JE, Bell RL, McBride WJ, Rodd
ZA (2014) Enhanced alcohol-seeking behavior by nicotine in the
posterior ventral tegmental area of female alcohol-preferring (P)
rats: modulation by serotonin-3 and nicotinic cholinergic
receptors. Psychopharmacology (Berl) 231:3745–3755.
Heimer L, Zahm DS, Churchill L, Kalivas PW, Wohltmann C (1991)
Specificity in the projection patterns of accumbal core and shell in
the rat. Neuroscience 41:89–125.
Heimer L, Alheid GF, de Olmos JS, Groenewegen HJ, Haber SN,
Harlan RE, Zahm DS (1997) The accumbens: beyond the core-
shell dichotomy. J Neuropsychiatry Clin Neurosci 9:354–381.
Herkenham M, Nauta WJH (1979) Efferent connections of the
habenular nuclei in the rat. J Comp Neurol 187:19–47.
Herve D, Pickel VM, Joh TH, Beaudet A (1987) Serotonin axon
terminals in the ventral tegmental area of the rat: fine structure
and synaptic input to dopaminergic neurons. Brain Res
435:71–83.
Hipolito L, Sanchez-Catalan MJ, Zornoza T, Polache A, Granero L
(2010) Locomotor stimulant effects of acute and repeated
intrategmental injections of salsolinol in rats: role of mu-opioid
receptors. Psychopharmacology (Berl) 209:1–11.
Hipolito L, Marti-Prats L, Sanchez-Catalan MJ, Polache A, Granero L
(2011) Induction of conditioned place preference and dopamine
release by salsolinol in posterior VTA of rats: involvement of mu-
opioid receptors. Neurochem Int 59:559–562.
Hipolito L, Sanchez-Catalan MJ, Marti-Prats L, Granero L, Polache A
(2012) Revisiting the controversial role of salsolinol in the
neurobiological effects of ethanol: old and new vistas. Neurosci
Biobehav Rev 36:362–378.
Hodge CW, Haraguchi M, Erickson H, Samson HH (1993) Ventral
tegmental microinjections of quinpirole decrease ethanol and
sucrose-reinforced responding. Alcohol Clin Exp Res
17:370–375.
Hokfelt T, Johansson O, Goldstein M (1984) Chemical anatomy of the
brain. Science 225:1326–1334.
Hong S, Jhou TC, Smith M, Saleem KS, Hikosaka O (2011) Negative
reward signals from the lateral habenula to dopamine neurons are
mediated by rostromedial tegmental nucleus in primates. J
Neurosci 31:11457–11471.
Hong S (2013) Dopamine system: manager of neural pathways. Front
Hum Neurosci 7:854.
Hutson LW, Szczytkowski JL, Saurer TB, Lebonville C, Fuchs RA,
Lysle DT (2014) Region-specific contribution of the ventral
tegmental area to heroin-induced conditioned
immunomodulation. Brain Behav Immun 38:118–124.
Ikemoto S, Kohl RR, McBride WJ (1997a) GABA(A) receptor
blockade in the anterior ventral tegmental area increases
extracellular levels of dopamine in the nucleus accumbens of
rats. J Neurochem 69:137–143.
Ikemoto S, Murphy JM, McBride WJ (1997b) Self-infusion of
GABA(A) antagonists directly into the ventral tegmental area
and adjacent regions. Behav Neurosci 111:369–380.
Ikemoto S, Murphy JM, McBride WJ (1998) Regional differences
within the rat ventral tegmental area for muscimol self-infusions.
Pharmacol Biochem Behav 61:87–92.
Ikemoto S, Wise RA (2002) Rewarding effects of the cholinergic
agents carbachol and neostigmine in the posterior ventral
tegmental area. J Neurosci 22:9895–9904.
Ikemoto S, Witkin BM, Morales M (2003) Rewarding injections of the
cholinergic agonist carbachol into the ventral tegmental area
induce locomotion and c-Fos expression in the retrosplenial area
and supramammillary nucleus. Brain Res 969:78–87.
Ikemoto S, Witkin BM, Zangen A, Wise RA (2004) Rewarding effects
of AMPA administration into the supramammillary or posterior
hypothalamic nuclei but not the ventral tegmental area. J Neurosci
24:5758–5765.
Ikemoto S (2005) The supramammillary nucleus mediates primary
reinforcement via GABA(A) receptors.
Neuropsychopharmacology 30:1088–1095.
Ikemoto S, Qin M, Liu ZH (2006) Primary reinforcing effects of
nicotine are triggered from multiple regions both inside and
outside the ventral tegmental area. J Neurosci 26:723–730.
Ikemoto S (2007) Dopamine reward circuitry: two projection systems
from the ventral midbrain to the nucleus accumbens-olfactory
tubercle complex. Brain Res Rev 56:27–78.
Ikemoto S (2010) Brain reward circuitry beyond the mesolimbic
dopamine system: a neurobiological theory. Neurosci Biobehav
Rev 35:129–150.
Ikemoto S, Bonci A (2014) Neurocircuitry of drug reward.
Neuropharmacology 76 Pt B:329–341.
Jalabert M, Bourdy R, Courtin J, Veinante P, Manzoni OJ, Barrot M,
Georges F (2011) Neuronal circuits underlying acute morphine
action on dopamine neurons. Proc Natl Acad Sci U S A
108:16446–16450.
Jhou TC, Fields HL, Baxter MG, Saper CB, Holland PC (2009a) The
rostromedial tegmental nucleus (RMTg), a GABAergic afferent to
midbrain dopamine neurons, encodes aversive stimuli and inhibits
motor responses. Neuron 61:786–800.
Jhou TC, Geisler S, Marinelli M, Degarmo BA, Zahm DS (2009b) The
mesopontine rostromedial tegmental nucleus: a structure targeted
by the lateral habenula that projects to the ventral tegmental area
of Tsai and substantia nigra compacta. J Comp Neurol
513:566–596.
Jhou TC, Xu SP, Lee MR, Gallen CL, Ikemoto S (2012) Mapping of
reinforcing and analgesic effects of the mu opioid agonist
endomorphin-1 in the ventral midbrain of the rat.
Psychopharmacology 224:303–312.
Jhou TC, Good CH, Rowley CS, Xu SP, Wang HK, Burnham NW,
Hoffman AF, Lupica CR, Ikemoto S (2013) Cocaine drives
aversive conditioning via delayed activation of dopamine-
responsive habenular and midbrain pathways. J Neurosci
33:7501–7512.
Johnson SW, North RA (1992) Opioids excite dopamine neurons by
hyperpolarization of local interneurons. J Neurosci 12:483–488.
Kalivas PW (1993) Neurotransmitter regulation of dopamine neurons
in the ventral tegmental area. Brain Res Brain Res Rev
18:75–113.
Katner SN, McBride WJ, Lumeng L, Li TK, Murphy JM (1997) Alcohol
intake of P rats is regulated by muscarinic receptors in the
pedunculopontine nucleus and VTA. Pharmacol Biochem Behav
58:497–504.

214 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M
(2009) Afferents to the GABAergic tail of the ventral tegmental
area in the rat. J Comp Neurol 513:597–621.
Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M
(2010a) Gamma-aminobutyric acid cells with cocaine-induced
DeltaFosB in the ventral tegmental area innervate mesolimbic
neurons. Biol Psychiatry 67:88–92.
Kaufling J, Waltisperger E, Bourdy R, Valera A, Veinante P, Freund-
Mercier MJ, Barrot M (2010b) Pharmacological recruitment of the
GABAergic tail of the ventral tegmental area by acute drug
exposure. Br J Pharmacol 161:1677–1691.
Kosaka K, Hiraiwa K (1915) Zur anatomie der sehnervenbahn und
ihrer zentren. Folia Neurobiol (leipzig) 9:367.
Lammel S, Hetzel A, Hackel O, Jones I, Liss B, Roeper J (2008)
Unique properties of mesoprefrontal neurons within a dual
mesocorticolimbic dopamine system. Neuron 57:760–773.
Lammel S, Lim BK, Ran C, Huang KW, Betley MJ, Tye KM,
Deisseroth K, Malenka RC (2012) Input-specific control of reward
and aversion in the ventral tegmental area. Nature 491:212–217.
Lammel S, Lim BK, Malenka RC (2014) Reward and aversion in a
heterogeneous midbrain dopamine system. Neuropharmacology
76 Pt B:351–359.
Lavezzi HN, Zahm DS (2011) The mesopontine rostromedial
tegmental nucleus: an integrative modulator of the reward
system. Basal Ganglia 1:191–200.
Lavezzi HN, Parsley KP, Zahm DS (2012) Mesopontine rostromedial
tegmental nucleus neurons projecting to the dorsal raphe and
pedunculopontine tegmental nucleus: psychostimulant-elicited
Fos expression and collateralization. Brain Struct Funct
217:719–734.
Lecca S, Melis M, Luchicchi A, Ennas MG, Castelli MP, Muntoni AL,
Pistis M (2011) Effects of drugs of abuse on putative rostromedial
tegmental neurons, inhibitory afferents to midbrain dopamine
cells. Neuropsychopharmacology 36:589–602.
Lecca S, Melis M, Luchicchi A, Muntoni AL, Pistis M (2012) Inhibitory
inputs from rostromedial tegmental neurons regulate
spontaneous activity of midbrain dopamine cells and their
responses to drugs of abuse. Neuropsychopharmacology
37:1164–1176.
Lee DY, Guttilla M, Fung KD, McFeron S, Yan J, Ranaldi R (2007)
Rostral-caudal differences in the effects of intra-VTA muscimol on
cocaine self-administration. Pharmacol Biochem Behav
86:542–549.
Li W, Doyon WM, Dani JA (2011) Acute in vivo nicotine administration
enhances synchrony among dopamine neurons. Biochem
Pharmacol 82:977–983.
Liechti ME, Lhuillier L, Kaupmann K, Markou A (2007) Metabotropic
glutamate 2/3 receptors in the ventral tegmental area and the
nucleus accumbens shell are involved in behaviors relating to
nicotine dependence. J Neurosci 27:9077–9085.
Linsenbardt DN, Boehm 2nd SL (2009) Agonism of the
endocannabinoid system modulates binge-like alcohol intake in
male C57BL/6J mice: involvement of the posterior ventral
tegmental area. Neuroscience 164:424–434.
Liu W, Thielen RJ, Rodd ZA, McBride WJ (2006) Activation of
serotonin-3 receptors increases dopamine release within the
ventral tegmental area of Wistar and alcohol-preferring (P) rats.
Alcohol 40:167–176.
Marinelli M, McCutcheon JE (2014) Heterogeneity of dopamine
neuron activity across traits and states. Neuroscience
282:176–197.
Marti-Prats L, Sanchez-Catalan MJ, Hipolito L, Orrico A, Zornoza
T, Polache A, Granero L (2010) Systemic administration of
D-penicillamine prevents the locomotor activation after intra-
VTA ethanol administration in rats. Neurosci Lett 483:
143–147.
Marti-Prats L, Sanchez-Catalan MJ, Orrico A, Zornoza T, Polache A,
Granero L (2013) Opposite motor responses elicited by ethanol in
the posterior VTA: the role of acetaldehyde and the non-
metabolized fraction of ethanol. Neuropharmacology 72:
204–214.
Matsui A, Williams JT (2011) Opioid-sensitive GABA inputs from
rostromedial tegmental nucleus synapse onto midbrain dopamine
neurons. J Neurosci 31:17729–17735.
Matsui A, Jarvie BC, Robinson BG, Hentges ST, Williams JT (2014)
Separate GABA afferents to dopamine neurons mediate acute
action of opioids, development of tolerance, and expression of
withdrawal. Neuron 82:1346–1356.
Melis M, Enrico P, Peana AT, Diana M (2007) Acetaldehyde
mediates alcohol activation of the mesolimbic dopamine system.
Eur J Neurosci 26:2824–2833.
Melis M, Carboni E, Caboni P, Acquas E (2013) Key role of salsolinol
in ethanol actions on dopamine neuronal activity of the posterior
ventral tegmental area. Addict Biol. http://dx.doi.org/10.1111/
adb.12097.
Melon LC, Boehm 2nd SL (2011) GABAA receptors in the posterior,
but not anterior, ventral tegmental area mediate Ro15-4513-
induced attenuation of binge-like ethanol consumption in C57BL/
6J female mice. Behav Brain Res 220:230–237.
Meye FJ, Adan RA (2014) Feelings about food: the ventral tegmental
area in food reward and emotional eating. Trends Pharmacol Sci
35:31–40.
Moaddab M, Haghparast A, Hassanpour-Ezatti M (2009) Effects of
reversible inactivation of the ventral tegmental area on the
acquisition and expression of morphine-induced conditioned
place preference in the rat. Behav Brain Res 198:466–471.
Montagu KA (1957) Catechol compounds in rat tissues and in brains
of different animals. Nature 180:244–245.
Moore EM, Boehm 2nd SL (2009) Site-specific microinjection of
baclofen into the anterior ventral tegmental area reduces binge-
like ethanol intake in male C57BL/6J mice. Behav Neurosci
123:555–563.
Morales M, Root DH (2014) Glutamate neurons within the midbrain
dopamine regions. Neuroscience 282:60–68.
Moreno S, Mugnaini E, Ceru MP (1995) Immunocytochemical
localization of catalase in the central nervous system of the rat.
J Histochem Cytochem 43:1253–1267.
Morikawa H, Morrisett RA (2010) Ethanol action on dopaminergic
neurons in the ventral tegmental area: interaction with intrinsic ion
channels and neurotransmitter inputs. Int Rev Neurobiol
91:235–288.
Morzorati SL, Marunde RL, Downey D (2010) Limited access to
ethanol increases the number of spontaneously active dopamine
neurons in the posterior ventral tegmental area of nondependent
P rats. Alcohol 44:257–264.
Nair-Roberts RG, Chatelain-Badie SD, Benson E, White-Cooper H,
Bolam JP, Ungless MA (2008) Stereological estimates of
dopaminergic, GABAergic and glutamatergic neurons in the
ventral tegmental area, substantia nigra and retrorubral field in
the rat. Neuroscience 152:1024–1031.
Nauta WJ (1958) Hippocampal projections and related neural
pathways to the midbrain in the cat. Brain 81:319–340.
Nestler EJ, Carlezon WA (2006) The mesolimbic dopamine reward
circuit in depression. Biol Psychiatry 59:1151–1159.
Nikulina EM, Johnston CE, Wang J, Hammer Jr RP (2014)
Neurotrophins in the ventral tegmental area: role in social
stress, mood disorders and drug abuse. Neuroscience
282:122–138.
Nowak KL, McBride WJ, Lumeng L, Li TK, Murphy JM (1998)
Blocking GABA(A) receptors in the anterior ventral tegmental area
attenuates ethanol intake of the alcohol-preferring P rat.
Psychopharmacology (Berl) 139:108–116.
Nowak KL, McBride WJ, Lumeng L, Li TK, Murphy JM (2000)
Involvement of dopamine D2 autoreceptors in the ventral
tegmental area on alcohol and saccharin intake of the alcohol-
preferring P rat. Alcohol Clin Exp Res 24:476–483.
Oades RD, Halliday GM (1987) Ventral tegmental (A10) system:
neurobiology. 1. Anatomy and connectivity. Brain Res
434:117–165.
Okamoto T, Harnett MT, Morikawa H (2006) Hyperpolarization-
activated cation current (Ih) is an ethanol target in midbrain
dopamine neurons of mice. J Neurophysiol 95:619–626.

M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216 215
Olson VG, Zabetian CP, Bolanos CA, Edwards S, Barrot M, Eisch AJ,
Hughes T, Self DW, Neve RL, Nestler EJ (2005) Regulation of
drug reward by cAMP response element-binding protein:
evidence for two functionally distinct subregions of the ventral
tegmental area. J Neurosci 25:5553–5562.
Olson VG, Nestler EJ (2007) Topographical organization of
GABAergic neurons within the ventral tegmental area of the rat.
Synapse 61:87–95.
Omelchenko N, Sesack SR (2005) Laterodorsal tegmental
projections to identified cell populations in the rat ventral
tegmental area. J Comp Neurol 483:217–235.
Orrico A, Hipolito L, Sanchez-Catalan MJ, Marti-Prats L, Zornoza T,
Granero L, Polache A (2013) Efficacy of D-penicillamine, a
sequestering acetaldehyde agent, in the prevention of alcohol
relapse-like drinking in rats. Psychopharmacology (Berl)
228:563–575.
Ouachikh O, Dieb W, Durif F, Hafidi A (2013) Differential behavioral
reinforcement effects of dopamine receptor agonists in the rat with
bilateral lesion of the posterior ventral tegmental area. Behav
Brain Res 252:24–31.
Ouachikh O, Dieb W, Durif F, Hafidi A (2014) Anterior ventral
tegmental area dopaminergic neurons are not involved in the
motivational effects of bromocriptine, pramipexole and cocaine in
drug-free rats. Behav Brain Res 262:1–7.
Overton PG, Vautrelle N, Redgrave P (2014) Sensory regulation of
dopaminergic cell activity: phenomenology, circuitry and function.
Neuroscience 282:1–12.
Paxinos G, Watson C (1998) The rat brain in stereotaxic
coordinates. CA: San Diego.
Paxinos G, Watson C (2007) The rat brain in stereotaxic
coordinates. CA: San Diego.
Perrotti LI, Bolanos CA, Choi KH, Russo SJ, Edwards S, Ulery PG,
Wallace DL, Self DW, Nestler EJ, Barrot M (2005) DeltaFosB
accumulates in a GABAergic cell population in the posterior tail of
the ventral tegmental area after psychostimulant treatment. Eur J
Neurosci 21:2817–2824.
Phillipson OT (1979a) A Golgi study of the ventral tegmental area of
Tsai and interfascicular nucleus in the rat. J Comp Neurol
187:99–115.
Phillipson OT (1979b) The cytoarchitecture of the interfascicular
nucleus and ventral tegmental area of Tsai in the rat. J Comp
Neurol 187:85–98.
Phillipson OT (1979c) Afferent projections to the ventral tegmental
area of Tsai and interfascicular nucleus: a horseradish peroxidase
study in the rat. J Comp Neurol 187:117–143.
Phillipson OT, Griffiths AC (1985) The topographic order of inputs to
nucleus accumbens in the rat. Neuroscience 16:275–296.
Reyes S, Fu Y, Double K, Thompson L, Kirik D, Paxinos G, Halliday
GM (2012) GIRK2 expression in dopamine neurons of the
substantia nigra and ventral tegmental area. J Comp Neurol
520:2591–2607.
Rivera-Meza M, Quintanilla ME, Bustamante D, Delgado R,
Buscaglia M, Herrera-Marschitz M (2014) Overexpression of
hyperpolarization-activated cyclic nucleotide-gated channels into
the ventral tegmental area increases the rewarding effects of
ethanol in UChB drinking rats. Alcohol Clin Exp Res 38:911–920.
Rodd-Henricks ZA, McKinzie DL, Crile RS, Murphy JM, McBride WJ
(2000) Regional heterogeneity for the intracranial self-
administration of ethanol within the ventral tegmental area of
female Wistar rats. Psychopharmacology (Berl) 149:217–224.
Rodd-Henricks ZA, Melendez RI, Zaffaroni A, Goldstein A, McBride
WJ, Li TK (2002) The reinforcing effects of acetaldehyde in the
posterior ventral tegmental area of alcohol-preferring rats.
Pharmacol Biochem Behav 72:55–64.
Rodd-Henricks ZA, McKinzie DL, Melendez RI, Berry N, Murphy JM,
McBride WJ (2003) Effects of serotonin-3 receptor antagonists on
the intracranial self-administration of ethanol within the ventral
tegmental area of Wistar rats. Psychopharmacology (Berl)
165:252–259.
Rodd ZA, Bell RL, Melendez RI, Kuc KA, Lumeng L, Li TK, Murphy
JM, McBride WJ (2004a) Comparison of intracranial self-
administration of ethanol within the posterior ventral tegmental
area between alcohol-preferring and Wistar rats. Alcohol Clin Exp
Res 28:1212–1219.
Rodd ZA, Melendez RI, Bell RL, Kuc KA, Zhang Y, Murphy JM,
McBride WJ (2004b) Intracranial self-administration of ethanol
within the ventral tegmental area of male Wistar rats: evidence for
involvement of dopamine neurons. J Neurosci 24:1050–1057.
Rodd ZA, Bell RL, Kuc KA, Zhang Y, Murphy JM, McBride WJ
(2005a) Intracranial self-administration of cocaine within the
posterior ventral tegmental area of Wistar rats: evidence for
involvement of serotonin-3 receptors and dopamine neurons. J
Pharmacol Exp Ther 313:134–145.
Rodd ZA, Bell RL, McQueen VK, Davids MR, Hsu CC, Murphy JM, Li
TK, Lumeng L, McBride WJ (2005b) Prolonged increase in the
sensitivity of the posterior ventral tegmental area to the reinforcing
effects of ethanol following repeated exposure to cycles of ethanol
access and deprivation. J Pharmacol Exp Ther 315:648–657.
Rodd ZA, Bell RL, McQueen VK, Davids MR, Hsu CC, Murphy JM, Li
TK, Lumeng L, McBride WJ (2005c) Chronic ethanol drinking by
alcohol-preferring rats increases the sensitivity of the posterior
ventral tegmental area to the reinforcing effects of ethanol.
Alcohol Clin Exp Res 29:358–366.
Rodd ZA, Bell RL, Zhang Y, Murphy JM, Goldstein A, Zaffaroni A, Li
TK, McBride WJ (2005d) Regional heterogeneity for the
intracranial self-administration of ethanol and acetaldehyde
within the ventral tegmental area of alcohol-preferring (P) rats:
involvement of dopamine and serotonin.
Neuropsychopharmacology 30:330–338.
Rodd ZA, Gryszowka VE, Toalston JE, Oster SM, Ji D, Bell RL,
McBride WJ (2007) The reinforcing actions of a serotonin-3
receptor agonist within the ventral tegmental area: evidence for
subregional and genetic differences and involvement of dopamine
neurons. J Pharmacol Exp Ther 321:1003–1012.
Rodd ZA, Oster SM, Ding ZM, Toalston JE, Deehan G, Bell RL, Li TK,
McBride WJ (2008) The reinforcing properties of salsolinol in the
ventral tegmental area: evidence for regional heterogeneity and
the involvement of serotonin and dopamine. Alcohol Clin Exp Res
32:230–239.
Rodd ZA, Bell RL, Oster SM, Toalston JE, Pommer TJ, McBride WJ,
Murphy JM (2010) Serotonin-3 receptors in the posterior ventral
tegmental area regulate ethanol self-administration of alcohol-
preferring (P) rats. Alcohol 44:245–255.
Roeper J (2013) Dissecting the diversity of midbrain dopamine
neurons. Trends Neurosci 36:336–342.
Rotllant D, Marquez C, Nadal R, Armario A (2010) The brain pattern
of c-fos induction by two doses of amphetamine suggests different
brain processing pathways and minor contribution of behavioural
traits. Neuroscience 168:691–705.
Sanchez-Catalan MJ, Hipolito L, Guerri C, Granero L, Polache A
(2008) Distribution and differential induction of CYP2E1 by
ethanol and acetone in the mesocorticolimbic system of rat.
Alcohol Alcohol 43:401–407.
Sanchez-Catalan MJ, Hipolito L, Zornoza T, Polache A, Granero L
(2009) Motor stimulant effects of ethanol and acetaldehyde
injected into the posterior ventral tegmental area of rats: role of
opioid receptors. Psychopharmacology (Berl) 204:641–653.
Scammell TE, Estabrooke IV, McCarthy MT, Chemelli RM,
Yanagisawa M, Miller MS, Saper CB (2000) Hypothalamic
arousal regions are activated during modafinil-induced
wakefulness. J Neurosci 20:8620–8628.
Schifirnet E, Bowen SE, Borszcz GS (2014) Separating analgesia
from reward within the ventral tegmental area. Neuroscience
263:72–87.
Schweimer JV, Coullon GSL, Betts JF, Burnet PW, Engle SJ,
Brandon NJ, Harrison PJ, Sharp T (2014) Increased burst-firing of
ventral tegmental area dopaminergic neurons in d-amino acid
oxidase knockout mice in vivo. Eur J Neurosci. http://dx.doi.org/
10.1111/ejn.12667.
Shabat-Simon M, Levy D, Amir A, Rehavi M, Zangen A (2008)
Dissociation between rewarding and psychomotor effects of
opiates: differential roles for glutamate receptors within anterior

216 M. J. Sanchez-Catalan et al. / Neuroscience 282 (2014) 198–216
and posterior portions of the ventral tegmental area. J Neurosci
28:8406–8416.
Soderman AR, Unterwald EM (2008) Cocaine reward and
hyperactivity in the rat: sites of mu opioid receptor modulation.
Neuroscience 154:1506–1516.
Spanagel R, Kiefer F (2008) Drugs for relapse prevention of
alcoholism: ten years of progress. Trends Pharmacol Sci
29:109–115.
Stamatakis AM, Stuber GD (2012) Activation of lateral habenula
inputs to the ventral midbrain promotes behavioral avoidance. Nat
Neurosci 15:1105–1107.
Stamatakis AM, Jennings JH, Ung RL, Blair GA, Weinberg RJ, Neve
RL, Boyce F, Mattis J, Ramakrishnan C, Deisseroth K, Stuber GD
(2013) A unique population of ventral tegmental area neurons
inhibits the lateral habenula to promote reward. Neuron
80:1039–1053.
Swanson LW (1982) The projections of the ventral tegmental area
and adjacent regions: a combined fluorescent retrograde tracer
and immunofluorescence study in the rat. Brain Res Bull
9:321–353.
Terashvili M, Wu HE, Leitermann RJ, Hung KC, Clithero AD,
Schwasinger ET, Tseng LF (2004) Differential conditioned place
preference responses to endomorphin-1 and endomorphin-2
microinjected into the posterior nucleus accumbens shell and
ventral tegmental area in the rat. J Pharmacol Exp Ther
309:816–824.
Tsai CE (1925) The optic tracts and centers of the opossum,
didelphis virginiana. J Comp Neurol 39:173–219.
Ungerstedt U (1971) Stereotaxic mapping of the monoamine
pathways in the rat brain. Acta Physiol Scand Suppl 367:1–48.
Usuda I, Tanaka K, Chiba T (1998) Efferent projections of the nucleus
accumbens in the rat with special reference to subdivision of the
nucleus: biotinylated dextran amine study. Brain Res 797:73–93.
Walsh JJ, Han MH (2014) The heterogeneity of ventral tegmental
area neurons: projection functions in a mood-related context.
Neuroscience 282:101–108.
Wang HL, Morales M (2008) Corticotropin-releasing factor binding
protein within the ventral tegmental area is expressed in a subset
of dopaminergic neurons. J Comp Neurol 509:302–318.
Wasserman DI, Wang HG, Rashid AJ, Josselyn SA, Yeomans JS
(2013) Cholinergic control of morphine-induced locomotion in
rostromedial tegmental nucleus versus ventral tegmental area
sites. Eur J Neurosci 38:2774–2785.
Wei-Malherbe H, Bone AD (1957) Intracellular distribution of
catecholamines in the brain. Nature 180:1050–1051.
Xie G, Hipolito L, Zuo W, Polache A, Granero L, Krnjevic K, Ye JH
(2012) Salsolinol stimulates dopamine neurons in slices of
posterior ventral tegmental area indirectly by activating mu-
opioid receptors. J Pharmacol Exp Ther 341:43–50.
Xie G, Ye JH (2012) Salsolinol facilitates glutamatergic transmission
to dopamine neurons in the posterior ventral tegmental area of
rats. PLoS One 7:e36716.
Yetnikoff L, Lavezzi HN, Reichard RA, Zahm DS (2014) An update on
the connections of the ventral mesencephalic dopaminergic
complex. Neuroscience 282:1–12.
Zahm DS, Becker ML, Freiman AJ, Strauch S, Degarmo B, Geisler S,
Meredith GE, Marinelli M (2010) Fos after single and repeated
self-administration of cocaine and saline in the rat: emphasis on
the Basal forebrain and recalibration of expression.
Neuropsychopharmacology 35:445–463.
Zangen A, Ikemoto S, Zadina JE, Wise RA (2002) Rewarding and
psychomotor stimulant effects of endomorphin-1: anteroposterior
differences within the ventral tegmental area and lack of effect in
nucleus accumbens. J Neurosci 22:7225–7233.
Zangen A, Solinas M, Ikemoto S, Goldberg SR, Wise RA (2006) Two
brain sites for cannabinoid reward. J Neurosci 26:4901–4907.
Zhang D, Dragomir A, Akay YM, Akay M (2014) Nicotine exposure
increases the complexity of dopamine neurons in the
parainterfascicular nucleus (PIF) sub-region of VTA. J Neuroeng
Rehabil 11:103.
Zhao-Shea R, Liu L, Soll LG, Improgo MR, Meyers EE, McIntosh JM,
Grady SR, Marks MJ, Gardner PD, Tapper AR (2011) Nicotine-
mediated activation of dopaminergic neurons in distinct regions of
the ventral tegmental area. Neuropsychopharmacology
36:1021–1032.
Zhou L, Furuta T, Kaneko T (2003) Chemical organization of
projection neurons in the rat accumbens nucleus and olfactory
tubercle. Neuroscience 120:783–798.
(Accepted 10 September 2014)(Available online 18 September 2014)



















![Opioid stimulation in the ventral tegmental area ...cogprints.org/6311/1/VTA.pdf · tegmental area (VTA) on maternal responsiveness [76]. The VTA, like the medial preoptic area, is](https://img.dokumen.tips/doc/110x75/5f4a93971087b136eb4517e9/opioid-stimulation-in-the-ventral-tegmental-area-tegmental-area-vta-on-maternal.jpg)


![Concurrent androgenic stimulation of the ventral tegmental area … pdf... · 2008. 1. 3. · [9,19,29,30,37]. Moreover, electrically stimulating the me- dial forebrain bundle increased](https://img.dokumen.tips/doc/110x75/5fd58dd8a591e17404617104/concurrent-androgenic-stimulation-of-the-ventral-tegmental-area-pdf-2008-1.jpg)





