Embed Size (px)
Citation preview

486 Biophysical Journal Volume 101 July 2011 486–493
Synchrotron X-Ray Microdiffraction Reveals Intrinsic Structural Featuresof Amyloid Deposits In Situ
Fatma Briki,†* Jerome Verine,‡ Jean Doucet,†Philippe Benas,§Barbara Fayard,†Marc Delpech,{Gilles Grateau,{
and Madeleine Ries-Kautt{†Laboratoire de Physique des solides, Centre Universitaire de Paris Sud, Orsay, France; ‡APHP, Hopital Saint-Louis, Laboratoire dePathologie, Paris, France; §Laboratoire de Cristallographie et RMN Biologiques, Universite Paris Descartes, Centre National de la RechercheScientifique, Faculte des Sciences Pharmaceutiques et Biologiques, Paris, France; and {Institut Cochin, Universite Paris Descartes, CentreNational de la Recherche Scientifique, Paris, France
ABSTRACT Amyloidoses are increasingly recognized as a major public health concern in Western countries. All amyloidosesshare common morphological, structural, and tinctorial properties. These consist of staining by specific dyes, a fibrillar aspect inelectron microscopy and a typical cross-b folding in x-ray diffraction patterns. Most studies that aim at deciphering the amyloidstructure rely on fibers generated in vitro or extracted from tissues using protocols that may modify their intrinsic structure.Therefore, the fine details of the in situ architecture of the deposits remain unknown. Here, we present to our knowledge the firstdata obtained on ex vivo human renal tissue sections using x-ray microdiffraction. The typical cross-b features from fixedparaffin-embedded samples are similar to those formed in vitro or extracted from tissues. Moreover, the fiber orientationmaps obtained across glomerular sections reveal an intrinsic texture that is correlated with the glomerulus morphology. Theseresults are of the highest importance to understanding the formation of amyloid deposits and are thus expected to trigger newincentives for tissue investigation. Moreover, the access to intrinsic structural parameters such as fiber size and orientation usingsynchrotron x-ray microdiffraction, could provide valuable information concerning in situ mechanisms and deposit formation withpotential benefits for diagnostic and therapeutic purposes.
INTRODUCTION
Amyloidoses constitute a heterogeneous group of acquiredor inherited diseases characterized by the presence ofamyloid deposits, mainly in the extracellular space, consist-ing of insoluble aggregated misfolded protein in the form offibrils (1–3). To date, 27 different human proteins are knownto have the ability to switch from a soluble structural form toinsoluble fibrillar aggregates. Several additional compo-nents are ubiquitously present in all amyloid deposits,including glycosaminoglycans (GAG), apolipoprotein E(ApoE), and serum amyloid P-component (SAP). The fibrilprecursor proteins are the basis of the modern nomenclatureof amyloidoses (4). Among these, serum amyloid A protein(SAA) is the precursor protein responsible for amyloid-associated (AA) amyloidosis, which sometimes complicatesdisorders that result from various chronic inflammations (5).The kidney is the organ most frequently affected in AAamyloidosis, which induces a progressive deterioration ofrenal function (6). Renal biopsy is widely used for the diag-nosis, with immunohistochemical techniques that revealthe AA protein accumulation as amorphous matter insidevarious kidney compartments. Such deposits generallyprevail in glomeruli and are also frequently present invessels, and more rarely in interstitium and along the tubularbasement membranes (7).
Submitted December 1, 2010, and accepted for publication May 27, 2011.
*Correspondence: [email protected]
Editor: Lois Pollack.
� 2011 by the Biophysical Society
0006-3495/11/07/0486/8 $2.00
Amyloid deposits are identified by a characteristicfibrillar signature in electron microscopy images, a typicalcross-b x-ray diffraction pattern, and histological stainingreactions, with a strong affinity for the Congo red dyewith apple green birefringence under polarized light.Although not fully understood, the fibrillar nature of theamyloid substance and its molecular configuration in theb-folded layer is supposed to be responsible for these char-acteristic tinctorial properties.
During the last two decades, major advances have beenmade in the field of amyloid structure characterization,partly thanks to technical developments. Recently, atomicforce microscopy observation of scrapie-infected cells re-vealed a growth process of amyloid-like fibrils on the cellsurface (8). Imaging the cell surface allowed characteriza-tion of the fibril distribution and their characteristic lateraland length dimensions, which were shown to be in agree-ment with in vitro structures. However, as noted by theauthors, the observed structures may result from a cellularresponse to the scrapie infection by changing membranecomposition or enhancing dendrite outgrowth, for example.Therefore, a characterization of the folded structure withinthe fibers is necessary to achieve an unambiguous demon-stration of the cross-b structure and the amyloid nature ofthe deposits. At the molecular scale, using a combinationof data from various in vitro experiments and molecularsimulation procedures, many sophisticated structure modelswere built (9,10). Spectroscopic analyses, such as circulardichroism or infrared spectroscopy of fibrils, generated
doi: 10.1016/j.bpj.2011.05.055

Structure Mapping of Amyloids In Situ 487
in vitro or extracted from tissues all lead to the same conclu-sion about the high b-sheet content of the fibrils. The partic-ular insoluble form of this cross-b-secondary-structure-richcore is an admitted basic common structural characteristicof amyloid protein fibrils (11). The lack of solubility pre-vents the use of the classical high-resolution techniques ofstructure determination involving protein solutions, suchas NMR or crystallography.
Therefore, electron microscopy and x-ray fiber diffractionare the most widely used techniques. They are often handledin a complementary way giving access to information at thefiber and molecular scale, respectively. Electron microscopywas the first technique to demonstrate the fibrillar nature ofthe deposits and is thus generally used to characterize thefiber morphology (12).
As reported throughout the literature, x-ray fiber diffrac-tion reveals a cross-b secondary structure of amyloid mate-rials (13). Current studies using this technique mainly dealwith the detection of cross-b folding in amyloid structuresformed in vitro using various peptides or proteins. Thesestudies often aim to demonstrate the molecule propensityto form amyloid deposits. It has been suggested from variousin vitro studies (14) that different proteins have the ability toform amyloids under appropriate conditions (15). These allshare a common secondary-structure core, as evidenced byx-ray fiber diffraction (16). This molecular feature is nowa-days the unique common information extractable from allfiber diffraction data. Additional data, such as high-resolu-tion molecular structure, have recently been obtained onsmall designed amino acid sequences, but not on the longerones, which are implied in amyloidosis (17).
Although very useful for molecular structure determina-tion, the modes of preparation of such in vitro samples,which sometimes lead to highly ordered diffraction patterns,can nevertheless induce some biases on the fibers and theirorganization with respect to their natural state. Indeed,fibrillar peptides very likely interact with other tissue com-ponents during the fibrillation process. This state can bedifferent from in vitro conditions in which the peptidesundergo various purification and conditioning treatments(18). Up to now, the so-called in vivo amyloid structuralstudies have been carried out on amyloid material, whichwas extracted from the tissue and then purified. As withthe fibers generated in vitro, they stain with thioflavin andCongo red in a similar manner. However, comparativestudies of ultrastructural shapes of in situ isolated fibersusing electron microscopy have concluded that a differenceexists between their fine structure depending on the fixationused: cryofixation or glutaraldehyde fixation (18). Anotherstudy on AA amyloid isolated from a tissue used the classicalPras method (19), which consists of repeated washings, firstwith saline water and then with distilled water. It was shownthat a macrofibril scaffold on which fibrils were formed wasdisrupted. The fibrils, whose diameter was initially 1–3 nmwhen associated with the microfibril scaffold, were shown
to reorganize into structures resembling those observed infibrils prepared in vitro. These observations raise questionsabout which fine structure reflects the actual in vivo state.The best answer would be given by in situ structure studies.However, to date, no technical approach that we know ofhas made it possible to acquire such information. Indeed,only a very few infrared studies have been carried out onin situ material using procalcitonin (20) and Ab amyloids(21). All authors concluded that there was an increasedcontent of the characteristic b-sheet amyloid signature inareas of tissue containing these deposits. Surprisingly,neither circular dichroism nor x-ray diffraction experimentshave been performed with in situ fibers.
Here, we present the first study that we know of carriedout on paraffin-embedded and frozen human renal tissuesections, with the aim of revealing the structural featuresof amyloid fibrils in deposits without the potential for struc-ture modification induced by extraction. To this end, weused synchrotron-based microdiffraction, which providesaccess to micrometer-sized spatial resolution. This allowedus to detect variations in the molecular structure of the fibersalong the tissue at the micron scale. In this way, fiber orien-tation could be followed along a glomerulus, and the varia-tion inside and around the whole glomerulus unit could bemonitored.
We further demonstrate that the in situ fibers in amyloiddeposits are partly folded into cross-b sheets, exhibitinga diffraction feature at 4.7 A as fibers formed in vitro.This validates the structural data from extracted fibers andstrengthens the use of this diffraction signal as a main crite-rion of fibril formation in vitro. We provide the experimentalconditions for detecting the characteristic 4.7-A reflectioninside tissue cuts. Working on the tissue allows accessingintrinsic information about the molecules, and we therebyshow a correlation between the position inside the glomer-ulus and the partial orientation of the amyloid fibers.
MATERIAL AND METHODS
Sample preparation
Renal tissue samples of an individual with a systemic AA amyloidosis
secondary to unknown hereditary disease were used for this study. They
were obtained from a radical nephrectomy performed before renal trans-
plantation. The presence of amyloid was established by the appearance of
an apple green birefringence from alkaline Congo red staining under polar-
ized light. AA amyloidosis was demonstrated by positive immunofluores-
cence staining with mouse anti-human Amyloid A monoclonal antibody
(clone mc1; 1:10 dilution; DakoCytomation, Glostrup, Denmark). The
samples were either frozen into liquid nitrogen or fixed in 4% buffered
paraformaldehyde and embedded in paraffin according to the routine tissue
processing for pathological examination.
For an initial examination of the amyloid deposit morphology, the renal
tissue specimens were cut at 5 mm thickness and stained with Congo red.
After verification, serial sections were cut from paraffin-embedded speci-
mens 50 mm thick and frozen specimens 40 mm thick and inserted between
two mica muscovite plates (cleaving planes [001]) for x-ray diffraction
analyses.
Biophysical Journal 101(2) 486–493

488 Briki et al.
X-ray microdiffraction mapping experiment
The experiments were performed at the European Synchrotron Radiation
Facility (Grenoble, France) at the microfocus beamline ID13 (22). The
high-intensity monochromatic beam (wavelength l ¼ 0.976 A), obtained
with an undulator source and a Si(111) double-crystal monochromator,
was focused with an ellipsoidal mirror (focal spot 20 (h) � 40 (v) mm2)
and then size-limited down to a 2-mm-diameter circular section by a colli-
mator placed in the focal plane. A guard aperture (Pt-Ir, 10 mm in diameter)
reduced diffuse scattering from the collimator exit. Samples were mounted
with the cut perpendicular to the x-ray beam on a computer-controlled
gantry coupled with a microscope, which permitted sample positioning
with 0.1-mm resolution.
Experiments were carried out using a sample-detector distance of
149 mm. The distance calibration was done with mica Bragg spots whose
first-order spacing is 4.48 A. Thanks to the small size of the beam stop,
two-dimensional x-ray scattering patterns were acquired from 0.008 to
0.4 A�1. After some trials to determine the best experimental parameters,
each pattern was recorded with an exposure time of 4 s on a MAR-CCD
camera (16-bit readout; 130-mm entrance window; 2048 � 2048 pixels;
pixel size of 64.46 � 64.46 mm2). No radiation damage effects on the scat-
tering features were detected for this exposure time, in agreement with
recent observations on radiation effects on fibrous proteins (23).
Various zones along the whole tissue section were sampled randomly by
mapping with large step sizes. When the patterns presented a significant
signal, a more detailed map was acquired all around with step size of
a few micrometers.
X-ray microdiffraction data treatment
The x-ray diffraction data were examined using the Fit2D software (24).
The patterns were first rapidly analyzed by visual inspection to make
a selection of the most interesting zones. A selection based on two criteria
was used: the overall intensity of the pattern and the presence of a reflection
at 4.7 A. We then focused our analysis on one glomerulus to follow the
variability of the diffraction patterns over the whole glomerulus unit and
all the surrounding tissue.
It is well known from birefringence in polarized light, that the signal
obtained from paraffin-embedded tissue contains a component from the
paraffin overlaid with that of the fibrous proteins. In a similar way, diffrac-
tion patterns exhibited signals arising from paraffin embedding and also
from the mica sheets used as sample holders. Therefore, the data-processing
step involved subtraction of a scattering pattern of the mica plates and of the
paraffin. However, it was not possible to completely remove the parasitic
signal arising from these materials. Therefore these two signals will be
present with a more or less intense contribution in the data presented
here. No further data treatment was applied to minimize any distortion of
the weak scattering features from the tissue.
For several selected patterns, the radial profiles were obtained over
limited angular sectors to avoid intense parasitic reflections. This was
achieved by circular integration within the same angle around the
c
b
da d
Biophysical Journal 101(2) 486–493
equator and around the meridian (encompassing the 4.7-A extent of
reflection) when the pattern was anisotropic, and over 360� when it
was isotropic.
RESULTS
Histological aspect of renal AA amyloidosis
The histological examination of the renal parenchyma re-vealed an abundant renal amyloid load, predominantly inglomeruli. All glomeruli indeed showed a nearly completeobliteration of glomerular tuft by amyloid deposits. Renalvessels were also involved in the amyloid deposits. Thus,arterioles frequently showed a complete replacement ofthe vessel walls by amyloid. Some amyloid deposits werealso noted in interstitium and along tubular basementmembranes. All these amyloid deposits were Congo-red-positive (Fig. 1 a) and strongly stained with mouse anti-human amyloid A monoclonal antibody (data not shown).
The in situ amyloid molecular fibers displaythe same 4.7-A ring as those formed in vitro
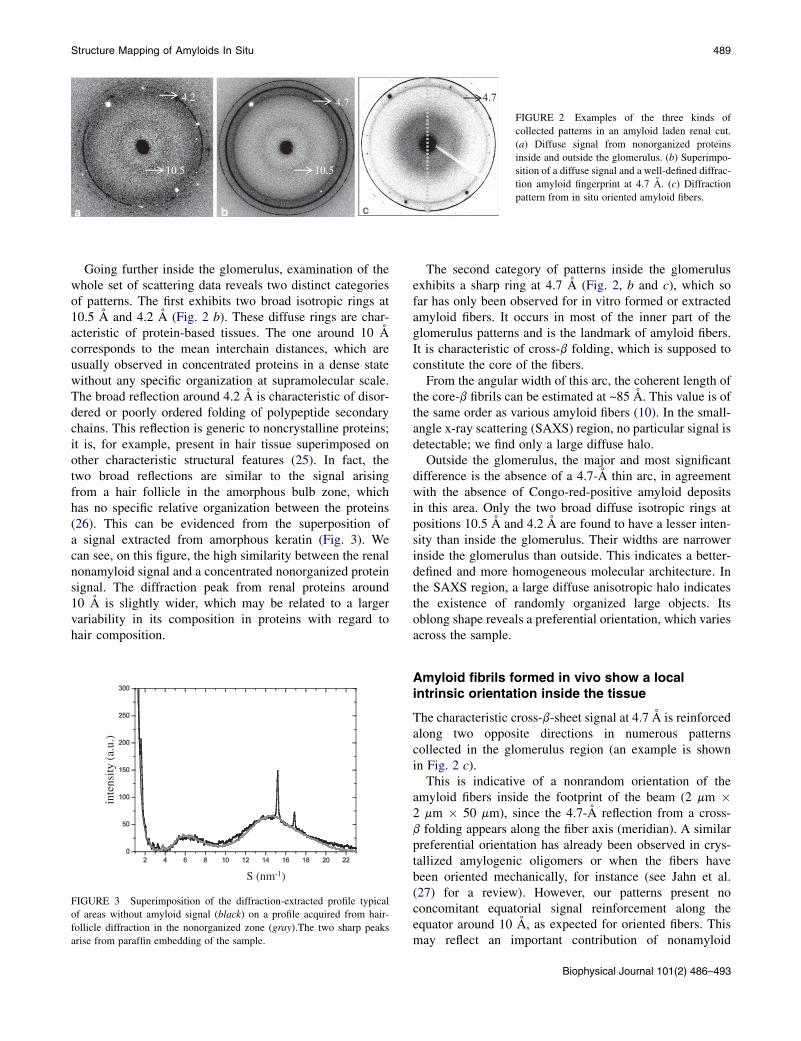
Fig. 1 b shows a nonstained image of a renal slice 50 mmthick. The zone indicated inside the rectangle was mappedby x-ray beam and at each point a whole diffraction patternwas recorded and mapped in Fig. 2 c.
The scan of the sample allowed us to distinguish threetypes of diffraction patterns, depending on the location ofthe tissue section: those with no 4.7-A reflection, fromoutside the glomerulus (Fig. 2 a); those with a thin reflectionat 4.7 A, from inside the glomerulus (Fig. 2 b.); or evensome with a thin anisotropic arc (Fig. 2 c). The intensityof this peak was roughly estimated by the relative heightof the peak after background subtraction. Indeed, as thesample was embedded in paraffin, as is customary in anato-mopathological conditioning, the paraffin diffraction peakswere present at a higher intensity than the protein peaks.In such conditions, signal processing to remove the paraffinand extract the 4.7-A intensity was not possible withouta large disturbance of the values. The intensity distributionobtained in this way is shown in Fig. 1 d, where one cansee the higher amyloid concentration inside the glomerulus,indicated by the higher signal intensity.
FIGURE 1 (a) Massive renal AA amyloidosis
clearly predominant in glomeruli (Congo red stain-
ing, original magnification 100�). (b and c) An
x-ray diffraction zone is approximately indicated
(b) with the whole produced diffraction patterns
(c). (d) Amyloid fiber distribution based on the
diffraction intensity of the 4.7-A reflection arising
from cross-b folded molecules within the kidney
tissue.
4.74.2 4.7
10.510.5
ca b
FIGURE 2 Examples of the three kinds of
collected patterns in an amyloid laden renal cut.
(a) Diffuse signal from nonorganized proteins
inside and outside the glomerulus. (b) Superimpo-
sition of a diffuse signal and a well-defined diffrac-
tion amyloid fingerprint at 4.7 A. (c) Diffraction
pattern from in situ oriented amyloid fibers.
Structure Mapping of Amyloids In Situ 489
Going further inside the glomerulus, examination of thewhole set of scattering data reveals two distinct categoriesof patterns. The first exhibits two broad isotropic rings at10.5 A and 4.2 A (Fig. 2 b). These diffuse rings are char-acteristic of protein-based tissues. The one around 10 Acorresponds to the mean interchain distances, which areusually observed in concentrated proteins in a dense statewithout any specific organization at supramolecular scale.The broad reflection around 4.2 A is characteristic of disor-dered or poorly ordered folding of polypeptide secondarychains. This reflection is generic to noncrystalline proteins;it is, for example, present in hair tissue superimposed onother characteristic structural features (25). In fact, thetwo broad reflections are similar to the signal arisingfrom a hair follicle in the amorphous bulb zone, whichhas no specific relative organization between the proteins(26). This can be evidenced from the superposition ofa signal extracted from amorphous keratin (Fig. 3). Wecan see, on this figure, the high similarity between the renalnonamyloid signal and a concentrated nonorganized proteinsignal. The diffraction peak from renal proteins around10 A is slightly wider, which may be related to a largervariability in its composition in proteins with regard tohair composition.
u.)
inte
nsity
(a.
S (nm-1)
FIGURE 3 Superimposition of the diffraction-extracted profile typical
of areas without amyloid signal (black) on a profile acquired from hair-
follicle diffraction in the nonorganized zone (gray).The two sharp peaks
arise from paraffin embedding of the sample.
The second category of patterns inside the glomerulusexhibits a sharp ring at 4.7 A (Fig. 2, b and c), which sofar has only been observed for in vitro formed or extractedamyloid fibers. It occurs in most of the inner part of theglomerulus patterns and is the landmark of amyloid fibers.It is characteristic of cross-b folding, which is supposed toconstitute the core of the fibers.
From the angular width of this arc, the coherent length ofthe core-b fibrils can be estimated at ~85 A. This value is ofthe same order as various amyloid fibers (10). In the small-angle x-ray scattering (SAXS) region, no particular signal isdetectable; we find only a large diffuse halo.
Outside the glomerulus, the major and most significantdifference is the absence of a 4.7-A thin arc, in agreementwith the absence of Congo-red-positive amyloid depositsin this area. Only the two broad diffuse isotropic rings atpositions 10.5 A and 4.2 A are found to have a lesser inten-sity than inside the glomerulus. Their widths are narrowerinside the glomerulus than outside. This indicates a better-defined and more homogeneous molecular architecture. Inthe SAXS region, a large diffuse anisotropic halo indicatesthe existence of randomly organized large objects. Itsoblong shape reveals a preferential orientation, which variesacross the sample.
Amyloid fibrils formed in vivo show a localintrinsic orientation inside the tissue
The characteristic cross-b-sheet signal at 4.7 A is reinforcedalong two opposite directions in numerous patternscollected in the glomerulus region (an example is shownin Fig. 2 c).
This is indicative of a nonrandom orientation of theamyloid fibers inside the footprint of the beam (2 mm �2 mm � 50 mm), since the 4.7-A reflection from a cross-b folding appears along the fiber axis (meridian). A similarpreferential orientation has already been observed in crys-tallized amylogenic oligomers or when the fibers havebeen oriented mechanically, for instance (see Jahn et al.(27) for a review). However, our patterns present noconcomitant equatorial signal reinforcement along theequator around 10 A, as expected for oriented fibers. Thismay reflect an important contribution of nonamyloid
Biophysical Journal 101(2) 486–493

490 Briki et al.
proteins to the diffracted intensity at 10 A, preventing thedetection of any small contribution of amyloid material.This effect may be reinforced by a poorly ordered lateralpacking of the molecules in the fibrils. This point is dis-cussed in detail in the Discussion section.
The anisotropic orientation seems to be globallylinked to the glomerulus morphology
To investigate this anisotropy further, we collected x-raymicrodiffraction patterns of a renal tissue section in scan-ning mode with sufficient resolution to allow establishmentof a possible correlation with histological components.A large glomerulus zone and its surrounding (300 �140 mm2) was scanned with a resolution of 7 � 7 mm2
(Fig. 1 c). A surprising observation was that the meandirection of the intensity reinforcement of the 4.7-A reflec-tion is roughly the same over the whole glomerulus, whichmeans that the amyloid fibers have a common averagepreferential orientation inside the glomerulus section.This result strongly supports a relationship between histo-logical zones and the produced scattering features. Unfor-tunately, the experimental conditions did not allow us toestablish a histological identification at the same scale;moreover, the degree of orientation is not high enough(see Fig. 4) to allow quantification of the degree of orien-tation. In Fig. 4, one can see that the difference betweenthe meridian and the equatorial intensities is too small tobe used for this purpose (Fig. 4, inset). However, the sensi-tivity of this diffraction technique allows one to evidencefibrous protein orientation on the two-dimensional pattern(Fig. 2 c).
Fig. 4 also shows that the equator and meridian intensitiesat 10.5 A are identical.
)
120
1404.7
10
20
30
40
4.7
inte
nsity
(a.u
.)
60
80
100
10.5
5 10 15 20-10
0
S (nm-1)5 10 15 20
40
FIGURE 4 Two extracted profiles from an oriented pattern from the
glomerulus. Integration over orthogonal sectors of 10� angular width along
the meridian (gray) and the equator (black). The small shoulder (black
arrow) arises from the sample supporting material, and the two sharp
intense peaks come from paraffin embedding. (Inset) Representation of
the difference between the meridian and equatorial profiles.
Biophysical Journal 101(2) 486–493
DISCUSSION
The molecular structure is independent of samplepreparation conditions
Various in vitro studies on amyloid fibrils revealed the exis-tence of different molecular architectures from the sameprecursor protein, depending on physical or chemical treat-ments. For example, heating the prion-like Ure2p fibrils at60�C changed the molecular organization within the fibrils(9). Our data are identical for the frozen and paraffin-embedded tissues. Therefore, the treatment for paraffinembedding, especially the heating to 60�C, has no effecton the fibril molecular organization. Indeed, the two char-acteristic reflections of cross-b structure for paraffin-embedded samples are present at the same positions inpatterns of frozen samples (data not shown). Paraffinembedding therefore appears harmless compared with cry-ofixation or glutaraldehyde chemical preparation. Never-theless, previous ultrastructural analyses of specimenstreated with those procedures resulted in a helical or fila-mentous form, depending on the pretreatment (28). Inthis study, the structural similarity of the two sampletreatments can be explained by the presence of thewhole-deposit components, which may contribute to thestructure’s stability.
Similarities and differences between in vitroor extracted amyloid fibers and ex vivo ones
A cross-b core structure evidenced by the meridional 4.7-Afeature has already been shown to be common to all knownamyloids, as much for in vitro generated fibers as for thoseextracted from a tissue. To the best of our knowledge, theonly case where a cross-b structure was observed withminimal tissue treatment is for transthyretin (TTR), whichis present in the vitreous fluid and throughout other organsof familial amyloid polyneuropathy patients. The structuresof the TTR fibrils from the vitreous fluid (without Prasmethod extraction) and those extracted from kidney by thePras method are identical (29). No data about in situcross-b molecular structure have been reported so far.Therefore, the results presented here show, for the firsttime that we know of, that the molecular structures ofamyloid material deposited in vivo inside a tissue sharewith in vitro or extracted amyloid fibers the common char-acteristic cross-b core structure. This is very important invalidating the data obtained for in vitro samples, whichare used to develop models based on the cross-b core struc-ture. Indeed, a plethora of data from various proteins thathave been coerced into making fibrils in the test tube haveused this figure to support the conclusion that amyloid fibrilswere actually produced.
On the other hand, differences between in vitro or ex-tracted oriented fibers and in situ ones are apparent bylooking at the equatorial 10.5-A scattering region. We

Structure Mapping of Amyloids In Situ 491
observe a nearly isotropic ring in the patterns, with anoriented peak at 4.7 A. This is better illustrated when look-ing at the projections in Fig. 4. This reflection is sometimesslightly more intense around the equator, whereas it is morepronounced and anisotropic for fibers formed in vitro or ex-tracted. The most likely explanation is that in vitro andextracted amyloid fibers are in general purified systems inwhich the cross-b core structures can self-assemble later-ally in a rather compact and regular way, unlike thein situ cross-b core structures, which are associated withother components forming a complex aggregation scaffold.The scaffold of the AA components may prevent lateraldirect association between amyloid fibrils, thus leading tothe absence or weakening of a specific amyloid signalaround 10 A. The scattering features from the cross-b core structures are thus superimposed against those ofthe other components, and are consequently less visiblethan the signal of pure cross-b core structures. For instance,one can assume a contribution from AA components suchas SAP, which are known to have a mostly b-sheetsecondary structure. This assumption is reinforced byprevious studies on tissue-extracted AA amyloids, whichself-assemble again after disruption of the macrofibril scaf-fold (18).
Moreover, the fine-structural examination of fibrils usingultrastructural techniques has suggested that in situ AAproteins are made of 10- to 30-A-diameter helical filamentsthat are aggregated into a microfibril-like structure, able tocontribute to the diffuse 10 A signal.
Another likely explanation, which is not in contradic-tion to the previous one, consists in assuming that the pre-cursor proteins could also preserve their native secondaryfolds and associate longitudinally, giving rise to the cross-b signal without the necessity of interactions for stabi-lizing their lateral structure. This is supported byin vitro studies of extended b-structured peptides, whichshow that long fibers can be formed without any lateralpacking (10), giving rise to a very weak 10-A feature.Indeed, it is likely that the absence of a well-defined pro-tofibrillar structure leads to an absence of this 10-Areflection.
All these assumptions are in agreement with electronmicroscopy images of different samples of variousamyloids, which showed that sections from AA-containingsamples produced diffuse images. Those images are difficultto interpret and cannot provide the number of protofilamentsin the fiber section, whereas this was easily done for othertypes of ex vivo amyloid samples (28).
It can be summarized from our x-ray patterns that onlythe association during the axial growth seems similar tothat observed for in vitro preparations, unlike the lateralpacking. This indicates the specificity of the interactionsbetween some amino acids that initiate the self-assem-bling into cross-b on large coherent domains as long as~85 A.
Morphology-related parameters frommicrodiffraction: implications for fibrillogenesisin a tissue
It follows from this study that fiber orientation can bemeasured with a 2-mm precision at the x-ray beam size reso-lution. Our results show that in the whole glomerulus, fibersare preferentially oriented in the direction parallel to theaxis that joins the afferent and efferent arterioles to the prox-imal convoluted tubule (Fig. 1 b, dashed arrow).This direc-tion is indicated on the diffraction pattern with the dashedarrow joining the two intensity maxima in Fig. 2 c, althoughthe orientation is in fact continuous over 360�. This aniso-tropic and uniform orientation is somewhat surprising giventhe heterogeneity of the glomerulus. Indeed, to obtainin vitro equivalent quality of orientation, various sampleconstraints are necessary, such as shear constraints, mechan-ical stretching, or intense magnetic field application.
The common average orientation of the fibrils in a givenarea could have been detected with birefringence analyses(30). However, two main points should be reported here.First, birefringence studies are always carried out afterCongo red staining, and it is the dye intercalation that likelyfollows fibril orientation that is detected. This might intro-duce artifacts that prevent a possible correlation of themeasured birefringence signal with structural information.In our diffraction study, the measured parameter is the directfiber orientation without any extrinsic staining. This mayallow us to measure well-characterized parameter valuessuch as the orientation angle and its width and the numberof oriented molecules. Moreover, these parameters can bedirectly correlated to an exact position in a structural tissueunit such as the glomerulus in this case, with a resolutiondefined by the operator as the mapping step size. On anotherhand, observations have been made about fibril orientationat the electron microscopy level, showing a correlationbetween fibril shape and the surrounding cells or connectivetissues (31). This is in agreement with the correlation estab-lished between fibril average orientation and glomerulusmorphology.
An explanation for the origin of the observed commonpreferential orientation could be due to a physicochemicallydriven growth process of the amyloid deposit, but it couldalso result from a specific interaction of the fibers with givensubglomerulus components, the surrounding tissue applyingan effect similar to that of mechanical constraints.
To date, data about invivofiber growth arevery limited and
are based only on the assembly of in vitro amyloidlike fibrils
using amyloidogenic peptides. Their kinetics is consistent
with a nucleation process. Accordingly, monitoring the accu-
mulation of amyloid in animal models is consistent with the
nucleation phenomenon that follows a sigmoidal shape. The
first scenario would imply starting from a unique fiber acting
as a seed, which would afterward impose its orientation
during the whole-deposit growth. The existence of a unique
Biophysical Journal 101(2) 486–493

492 Briki et al.
seed seems not realistic, however; several seeds probablyappear simultaneously in the glomerulus. The uniform orien-tation, which is present in the whole glomerulus, thereforesuggests another scheme for fiber deposition.
Our suggestion is an alternative scenario where fibergrowth is governed by tissue-specific physical or chemicalfactors and takes place when an adequate precursor concen-tration is present. The interaction with subglomeruluscomponents is much more likely. Indeed, the direction ofthe average orientation of the fibers on the section corre-sponds to the axis of the glomerulus, which joins the afferentand efferent arterioles to the proximal convoluted tubule.Looking at a histological image, one can see that the capil-lary tuft of the glomerulus is formed by a series of sectionsof the capillary that are mainly directed along the axis of theglomerulus and that form loops between two sections.Therefore, the amyloid fibers would be deposited alongthe glomerular basement membrane (with the fiber axislying on the membrane), and an anisotropic angular distri-bution of the fibers would be expected.
Another hypothesis may involve a growth steered bymolecules other than the amyloid protein, and which maybe involved in the fiber polymerization process, leading totheir intrinsic alignment. The tissue can then be seen asa composite material of normally folded proteins reinforcedby a well-organized distribution of fibrous molecules soakedinto the amorphous matrix; this would lead to the excep-tional resistance of the deposits to various treatments suchas proteolysis. To understand fibrillogenesis more fully, itwould be useful to complete this work with a detailed anal-ysis of the in vivo driving conditions of these structuresmonitoring the 4.7-A reflection, following the process atvarious stages of the pathology with animal models, andto track down the zones where the deposits are initiated.
X-ray microdiffraction as a structural imaging toolin amyloidosis
Amyloidosis is typically identified by the birefringence ofCongo red staining of a biopsy, whereas its typing relieson immunohistochemical tests. Congo red has been shownto react with any highly aggregated material, regardless ofits composition (32).
Although the staining process is easy and inexpensive, thechemical mechanisms are poorly understood, and results canvary widely depending on the protocol or the reagent quality.Moreover, the Congo red dye was shown to be nonspecific,as it induces circular dichroism even with native structures,as demonstrated on a variety of native proteins (33). Thisstudy showed that the Congo red must be used with cautionas a diagnostic tool for the presence of amyloid fibrils.
Our results presented here emphasize the high capabil-ities of x-ray diffraction as an imaging tool for ex vivosamples with no post treatment after renal tissue resection.This technique is quite sensitive to the presence of amyloid
Biophysical Journal 101(2) 486–493
fibers inside a tissue, as well as to fiber orientation. It is alsothe only approach we know of that unambiguously givesaccess to the molecular fingerprint of amyloid fibers, ableto overcome the weaknesses of other techniques, such asFTIR-microscopy. The FTIR technique has already beenused for Ab-peptide secondary-structure distribution (34)inside a tissue section, but it is not sensitive enough todistinguish cross-b structures from extended-b ones.
Experimental x-ray maps could give access to even moreinformation than the fiber distribution and orientationparameters, such as size and possibly basic structural char-acteristics. One could expect to have more variability in thestructures, depending on the precursor protein, the amyloid-laden tissue, or the local biological conditions. In vitrostudies show that a given peptide can give rise to a largevariety of cross-b fibers according to the experimentalconditions, which might be of a biological significance.Given the potential importance of the environmental effectson the formation of amyloid aggregates, in situ diffractionstudies of deposits formed in vivo may also provide infor-mation about this precise environment state.
A further interesting aspect of our work concerns theseadditional structural parameters becoming available to char-acterize amyloidoses. The current classification of amyloid-oses is based on fibril precursor proteins, which arenormally found in the plasma and usually without anybiochemical connection. However, differences at the supra-molecular level of the protofibril assembly are known toexist ex vivo (28). Our study opens the possibility of accessto new parameters that could characterize amyloid deposits,from the atomic or molecular level to the microscopic level:crystallite size from coherence length, possible lateral crys-tallinity parameters, quality of organization, and orientation.We shall then have means to characterize the variousamyloidoses using additional parameters, which becomesignificant in situ, whereas they might be influenced bythe sample preparation when measured in vitro.
CONCLUSION
We succeeded in recording in situ fiber x-ray microdiffrac-tion patterns from an amyloid-laden human kidney tissuespecimen, opening new perspectives for future investigationtools in the field of amyloidosis.
Our data provide evidence that the in situ fibers inamyloid deposits are partly folded into cross-b sheets asin fibers formed in vitro. Moreover, we prove that despiteits weak intensity, this structural signature is directlyobservable in tissue sections, opening new opportunitiesfor histological examination.
New pieces of information were added to the existingdata from studies carried out on in vitro systems. First, thelateral organization of the cross-b core objects is disrupted,probably because of other components that are associatedwith the amyloid molecules, unlike the longitudinal cross-b

Structure Mapping of Amyloids In Situ 493
ordering, which seems to be the same as for in vitro systems.Second, the amyloid fibers are more or less naturally alignedlocally within the beam footprint (2 � 2 � 50 mm3). Third,the mapping mode at a few micrometers resolution showedthat the alignment direction of the amyloid fibers is roughlythe same over the whole glomerulus section. The orientationof the fibers, which is usually used in vitro to improve thequality of the diffraction pictures and thus facilitate the struc-tural interpretation, is observed here as intrinsic to the tissue.This structural feature is certainly significant, because it canbe specific to the tissue type or to the precursor proteins andtheir fibrillation process.
The detailed analysis of thesevarious parameters in diversecontrolled conditions will be very important for advancingour understanding of the biological processes involved inthe mechanisms of the amyloid fiber formation and the exactconditions of their deposition. To our knowledge, theseeffects are not accessible by any other method in a conditionso close to in vivo and with molecular resolution. De-signing well-controlled experiments that will allow us toobtain similar information in vitro would be very helpful inimproving the understanding of fibrillogenesis conditionsand in following the effects of possible treatments on a tissue.
We thank Aurelien Gourrier for critical reading of the manuscript. We also
thank the European Sychrotron Radiation Facility staff of the ID13 micro-
focus beamline.
We are grateful for financial support from the European Community’s Sixth
Framework Program, EURAMY Project (LSHM-CT-2006-037525).
REFERENCES
1. Grateau, G., J. Verine, ., M. Ries. 2005. [Amyloidosis: a model ofmisfolded protein disorder]. Med. Sci. (Paris). 21:627–633.
2. Selkoe, D. J. 2003. Folding proteins in fatal ways.Nature. 426:900–904.
3. Glenner, G. G., H. R. Keiser,., R. A. DeLellis. 1968. Amyloid. VI. Acomparison of two morphologic components of human amyloiddeposits. J. Histochem. Cytochem. 16:633–644.
4. Westermark, P., M. D. Benson, ., J. D. Sipe. 2007. A primer ofamyloid nomenclature. Amyloid. 14:179–183.
5. Herbert, M. A., D. V. Milford,., F. Raafat. 1995. Secondary amyloido-sis from long-standing bacterial endocarditis. Pediatr. Nephrol. 9:33–35.
6. Gertz, M. A., and R. A. Kyle. 1991. Secondary systemic amyloidosis:response and survival in 64 patients. Medicine (Baltimore).70:246–256.
7. Ferrario, F., and M. P. Rastaldi. 2006. Renal amyloidosis (part I).J. Nephrol. 19:123–125.
8. Wegmann, S., M. Miesbauer,., D. J. Muller. 2008. Observing fibrillarassemblies on scrapie-infected cells. Pflugers Arch. 456:83–93.
9. Bousset, L., F. Briki,., R. Melki. 2003. The native-like conformationof Ure2p in fibrils assembled under physiologically relevant conditionsswitches to an amyloid-like conformation upon heat-treatment of thefibrils. J. Struct. Biol. 141:132–142.
10. Croixmarie, V., F. Briki, ., A. Sanson. 2005. A cylinder-shapeddouble ribbon structure formed by an amyloid hairpin peptide derivedfrom the b-sheet of murine PrP: an x-ray and molecular dynamicssimulation study. J. Struct. Biol. 150:284–299.
11. Kayed, R., E. Head, ., C. G. Glabe. 2003. Common structure ofsoluble amyloid oligomers implies common mechanism of pathogen-esis. Science. 300:486–489.
12. Shirahama, T., and A. S. Cohen. 1967. High-resolution electron micro-scopic analysis of the amyloid fibril. J. Cell Biol. 33:679–708.
13. Bonar, L., A. S. Cohen, and M. M. Skinner. 1969. Characterization ofthe amyloid fibril as a cross-b protein. Proc. Soc. Exp. Biol. Med.131:1373–1375.
14. Glenner, G. G., E. D. Eanes, ., J. D. Termine. 1974. b-pleated sheetfibrils. A comparison of native amyloid with synthetic protein fibrils.J. Histochem. Cytochem. 22:1141–1158.
15. Carulla, N., G. L. Caddy,., C. M. Dobson. 2005. Molecular recyclingwithin amyloid fibrils. Nature. 436:554–558.
16. Sunde, M., L. C. Serpell,., C. C. Blake. 1997. Common core structureof amyloid fibrils by synchrotron x-ray diffraction. J. Mol. Biol.273:729–739.
17. Makin, O. S., E. Atkins, ., L. C. Serpell. 2005. Molecular basis foramyloid fibril formation and stability. Proc. Natl. Acad. Sci. USA.102:315–320.
18. Inoue, S., M. Kuroiwa,., R. Kisilevsky. 1998. A high resolution ultra-structural comparison of isolated and in situ murine AA amyloid fibrils.Amyloid. 5:99–110.
19. Pras, M., M. Schubert,., E. C. Franklin. 1968. The characterization ofsoluble amyloid prepared in water. J. Clin. Invest. 47:924–933.
20. O’Leary, T. J., and I. W. Levin. 1985. Secondary structure of endocrineamyloid: infrared spectroscopy of medullary carcinoma of the thyroid.Lab. Invest. 53:240–242.
21. Choo, L. P., D. L. Wetzel, ., H. H. Mantsch. 1996. In situ character-ization of b-amyloid in Alzheimer’s diseased tissue by synchrotronFourier transform infrared microspectroscopy. Biophys. J. 71:1672–1679.
22. Muller, M., M. Burghammer, and C. Riekel. 2001. Combined scanningmicrodiffraction and micro small-angle scattering at the microfocusbeamline ID13 (ESRF). Nucl. Instrum. Methods Phys. Res. A.467–468:958–961.
23. Leccia, E., A. Gourrier, ., F. Briki. 2010. Hard a-keratin degradationinside a tissue under high flux x-ray synchrotron micro-beam: a multi-scale time-resolved study. J. Struct. Biol. 170:69–75.
24. Hammersley, A. P. 1997. ‘‘FIT2D: An Introduction and Overview’’.ESRF Internal Report, ESRF97HA02T.
25. Kreplak, L., J. Doucet,., F. Briki. 2004. New aspects of the a-helix tob-sheet transition in stretched hard a-keratin fibers. Biophys. J.87:640–647.
26. Rafik, M. E., F. Briki, ., J. Doucet. 2006. In vivo formation steps ofthe hard a-keratin intermediate filament along a hair follicle: evidencefor structural polymorphism. J. Struct. Biol. 154:79–88.
27. Jahn, T. R., O. S. Makin, ., L. C. Serpell. 2010. The common archi-tecture of cross-b amyloid. J. Mol. Biol. 395:717–727.
28. Serpell, L. C., M. Sunde, ., P. E. Fraser. 2000. The protofilamentsubstructure of amyloid fibrils. J. Mol. Biol. 300:1033–1039.
29. Inoue, S., M. Kuroiwa, ., R. Kisilevsky. 1998. Ultrastructure offamilial amyloid polyneuropathy amyloid fibrils: examination withhigh-resolution electron microscopy. J. Struct. Biol. 124:1–12.
30. Shirahama, T., and A. S. Cohen. 1980. Redistribution of amyloiddeposits. Am. J. Pathol. 99:539–550.
31. Shirahama, T., and A. S. Cohen. 1969. Ultrastructural studies of renalperitubular amyloid experimentally induced in guinea pigs. 3. Bloodand lymphatic capillaries. Exp. Mol. Pathol. 11:300–322.
32. Glenner, G. G., E. D. Eanes, and D. L. Page. 1972. The relation of theproperties of Congo red-stained amyloid fibrils to the � conformation.J. Histochem. Cytochem. 20:821–826.
33. Khurana, R., V. N. Uversky, ., A. L. Fink. 2001. Is Congo red anamyloid-specific dye? J. Biol. Chem. 276:22715–22721.
34. Miller, L. M., Q. Wang, ., J. Miklossy. 2006. Synchrotron-basedinfrared and x-ray imaging shows focalized accumulation of Cu andZn co-localized with b-amyloid deposits in Alzheimer’s disease.J. Struct. Biol. 155:30–37.
Biophysical Journal 101(2) 486–493





























