Embed Size (px)
Citation preview

Supramolecular insulin assembly II for a sustainedtreatment of type 1 diabetes mellitusSarika Guptaa, Tandrika Chattopadhyaya, Mahendra Pal Singha, and Avadhesha Suroliaa,b,1
aNational Institute of Immunology, Aruna Asaf Ali Marg, New Delhi 110067, India; and bMolecular Biophysics Unit, Indian Institute of Sciences,Bangalore 560012, India
Communicated by William J. Lennarz, State University of New York at Stony Brook, Stony Brook, NY, May 21, 2010 (received for review May 1, 2009)
Diabetes is a chronic disease requiring continuous medical supervi-sion and patient education to prevent acute secondary complica-tions. In this study, we have harnessed the inherent property ofinsulin to aggregate into an oligomeric intermediate on thepathway to amyloid formation, to generate a form that exhibitscontrolled and sustained release for extended periods. Administra-tion of a single dose of the insulin oligomer, defined here as thesupramolecular insulin assembly II (SIA-II), to experimental animalsrendered diabetic by streptozotocin or alloxan, released thehormone capable of maintaining physiologic glucose levels for>120 days for bovine and >140 days for recombinant human insulinwithout fasting hypoglycemia. Moreover, the novel SIA-IIdescribed here not only improved the glycemic control, but alsoreduced the extent of secondary diabetic complications.
protein folding ∣ hyperglycemia ∣ normoglycemia ∣ insulin-like growthfactor-1
Diabetes is a debilitating disease, responsible for extensivedeaths worldwide (http://www.who.int/diabetes/en/). The
economic impact of this is likely to be even more stupendous,because people may live for years with diabetes; their cause ofdeath is often recorded as cardiovascular diseases, stroke, andkidney failures, all precipitating largely because of diabetes(1). Diabetes is a chronic disease characterized by either theinability of the body to produce insulin (type 1) or the failureto respond to it (type 2) (2). Attempts to replicate physiologicalinsulin secretion, as a means of restoring the normal metabolicmilieu and thereby minimizing the risk of diabetic complications,have become an essential feature of treatment of this disease. Thecurrent advocacy of intensive insulin therapy regimens involvingmultiple daily s.c. injections places a heavy burden of complianceon patients and has prompted interest in developing alternative,less invasive therapies. To date, attempts to exploit the nasal,oral, gastrointestinal, and transdermal routes have been mostlyunsuccessful. The conventional regimen of multiple daily insulininjections for type 1 diabetes is effective in preventing postpran-dial blood glucose excursions, but it is limited by its inadequatecontrol of fasting hypoglycemia (3).
In the present study, we utilize the currently available knowl-edge of protein folding to develop a safe and long-acting supramo-lecular insulin assembly (SIA-I, SIA-II, and SIA-III). Insulinis a 51 residue polypeptide that exists in equilibrium in solutionas a mixture of different oligomeric states. In pancreatic secretoryvesicles, insulin is stored as ahexamer at physiological pH,whereasit interacts with its receptor as a monomer. Under denaturingconditions, such as low pH or in the presence of strong denatur-ants, insulin aggregates into amyloid fibrils. To test the hypothesisthat insulin released from supramolecular insulin assembly II(SIA-II) obtained by oligomerization around physiological pHachieved better glycemic control for a sustained period of timein animalmodel of diabetes, SIAwas developed and characterizedin detail during these studies for its use as an alternative treatmentto conventional insulin therapy. Insulin from two different sources(namely, bovine insulin and recombinant human insulin) wereseen for their ability to form SIA-II and exert a tight glycemic
control in various diabetic models, such as rat, mice, and rabbit.(Mice and rabbit data are given in SI Text). A single dose of insulinSIA-II when administered s.c. or i.m. was able to maintain a basallevel of insulin in streptozotocin (STZ)induced diabetic rat for aprolonged period of >120 and >140 days for bovine and recom-binant human (rH) insulin, respectively, simultaneously keeping atight glycemic control, thus affording a long-lasting treatmentagainst diabetes mellitus type 1 (DM-1) in experimental animals.
ResultsMonitoring of Insulin Fibril Formation by Thioflavin T (Th-T) Fluores-cence. Fibril formation by bovine and rH insulin was monitoredby Th-T fluorescence. As shown in Fig. 1A, an increase in Th-Tfluorescence with time was observed, which reached a maximumvalue at 48 h for the hormone incubated at pH 7.0, whereasfor pH 2.0, Th-T fluorescence attained a maximum value at20 h, indicating that the fibril formation is more rapid under acidicconditions. Three supramolecular insulin intermediates at pH 7.0SIA-I, SIA-II, and SIA-III formed at 6–8 h, 12–16 h, and 20–24 h,respectively, were selected. These intermediates were furthercharacterized structurally and release of insulin monomers fromthem was studied.
Release of Insulin Monomers from Its Amyloid and SIA-II Forms. Toprove our hypothesis that the SIA-II form of insulin can act asa reservoir for the sustained release of the hormone for a longtime, release of insulin monomers from its amyloid and variousintermediates (SIA-I, SIA-II, and SIA-III) through a 8-kDa cutoffmembrane was examined (Fig. S1A and B). Fully formed amyloidfibers did not release insulin, irrespective of the pH of fibriliza-tion. The pH 2.0 intermediates released insulin at a very slowrate, suggesting that these oligomers are sturdy and tightly asso-ciated. However, the pH 7.0 intermediate (termed SIA-II) re-leased insulin at an appreciable rate. A linear increase in therelease of insulin from SIA-II, as monitored by its absorbanceat 280 nm over a period of 15� 5 days, was observed (Fig. 1B).These observations were also corroborated by tyrosine fluores-cence, which increased in the dialysate with time. Therefore,SIA-II was chosen so as to achieve a constant in vivo release,for maintaining basal insulin levels in the body.
SIA-II intermediates were further characterized using Congo-red (CR) binding and FTIR, and its morphology was assessedby atomic force microscopy (AFM) and transmission electronmicroscopy (TEM).
Congo-Red Binding. CR binds specifically to the β-sheet rich struc-tures of amyloid and has been used routinely for their detection.Both bovine and rH SIA-II exhibited weak binding to CR (Fig. 1C and D), whereas fully grown fibers at both pH 2.0 and 7.0
Author contributions: S.G. and A.S. designed research; S.G., T.C., and M.P.S. performedresearch; S.G., T.C., M.P.S., and A.S. analyzed data; and S.G., T.C., and A.S. wrote the paper.
The authors declare no conflict of interest.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1005704107/-/DCSupplemental.
13246–13251 ∣ PNAS ∣ July 27, 2010 ∣ vol. 107 ∣ no. 30 www.pnas.org/cgi/doi/10.1073/pnas.1005704107

showed significant binding. Thus SIA-II is a nonfibrillar structureand is devoid of amyloid-like characteristics.
Fourier Transform Infrared Spectroscopy. SIA-I, SIA-II, and SIA-IIIwere also characterized using attenuated total reflection (ATR)-FTIR.Distinct spectra corresponding to each stage were observed(Fig. S1C). SIA-II has a sharp peak at 1;647 cm−1 and 1;645 cm−1
for bovine and rH insulin, respectively. In contrast, the fullyformed amyloid peaks at 1;630 cm−1 and 1;628 cm−1 for bovineand rH insulins, respectively. The FTIR spectra is in goodagreement with the CR binding data showing that the conforma-tion of SIA-II is still largely helical, albeit with an increase in thecontent of random coil structure.
Atomic Force Microscopy. Morphology of fibers formed at pH 2.0and 7.0 were assessed by AFM. Native insulins (both bovine andrH) at pH 7.0 and pH 2.0 show random distribution with a heightof 1.3� 0.21 nm, which correlates with the dimension of insulinmonomers (1.11 nm) and dimers (1.49 nm) [Fig. 1E(i)]. Inter-mediates of the fibrilization process at pH 7.0 are shown inFig. 1E(ii–iv) for bovine insulin and Fig. 1E(v–vii) for rH insulin.SIA-I represents elongated clusters having a pearl-like arrange-ment. In between, there are some elongated linear particleswith 12� 2 nm height, suggesting further association into higheroligomeric states. The SIA-II intermediate of bovine insulin isseen as a linear association of the above-mentioned elongatedclusters; however, in case of rH insulin, large elongated bead-likestructures with 34� 10 nm height were seen [Fig. 1E(iii and vi)].SIA-III (stage succeeding SIA-II) showed an increase in densityof higher oligomeric structures. Insulin amyloids are seen as fullygrown fibers at pH 7.0 [Fig. 1E(viii)]. Intermediates of the pH 2.0fibrilization process at 6–7 h reveal a lateral association of twofibrils of height 7.2–8.3 nm, and after 20 h, large twisted fibersof 10–12 nm width were observed [Fig. 1E(ix–x)].
TEM studies of the intermediates were performed to see thepresence of other possible assemblies and are shown in Fig. S1D.
Protease Cleavage. Proteolysis experiments were performed toexplore the resistance of SIA-II and SIA-III to cleavage by pro-teinase K. rH insulin and SIA-I were highly susceptible to cleavageby proteinase K as compared to SIA-II and SIA-III [Fig. 1F(i–iv)].
This demonstrates that both SIA-II and SIA-III adopt a higher-or-der oligomeric form, resistant to protease action.
In Vivo Studies in STZ-Induced DM-1 Rat Model; Treatment with SIA-IIForm of Insulin.Optimization of bovine and rH insulin dosage wasdone using 2–8 units∕kg body weight, which has been reported tomaintain normoglycemia in diabetic rats. When administeredonce or twice daily, 4 units∕kg body weight was found to be idealand did not lead to sudden fasting (preprandial) and nonfasting(postprandial) hypoglycemia. STZ-induced diabetic rats weredivided into five groups, and the sixth group consisted of nondia-betic animals. Group I diabetic animals were treated with insulin(4 units∕kg, i.p. daily), group II—insulin amyloid, group III—SIA-II (s.c.), group IV—SIA-II (i.m.), and group V—PBS-treateddiabetic control rats. Insulin amyloid had no beneficial effect onthe glycemic status of diabetic rats (Fig. S2A), due to negligiblerelease of insulin monomers from the tightly packed fibrils. How-ever, SIA-II was effective in lowering blood glucose levels. Tostandardize the dosage 50, 100, 200, and 400 μg of the hormoneoligomer were injected either s.c. or i.m. in STZ-diabetic rats. Asingle dose of 50 and 100 μg of SIA-II maintained near-normo-glycemic levels up to 10 and 30 days for bovine insulin and 15 and45 days for rH insulin (Fig. 2A) compared to 135 and 160 days forbovine (Upper) and rH (Lower) insulin, respectively, when 200 μgSIA-II was used. Dosage of 400 μg resulted in sudden hypogly-cemia, suggesting an initial bolus release (above basal level) ofinsulin monomers at this dose (Fig. 2A). Thus 200 μg SIA-II (bothbovine and rH insulin) was chosen as a therapeutic dosage forthe detailed long-term prospective studies. Preprandial (over-night 8- to 10-h fasting) and postprandial (nonfasting/fed state,i.e., random-fed BGL taken at 11.00 AM). Blood glucose levels(BGL) were monitored till the physiological effect of SIA-II wasobserved. Treatment of STZ-induced diabetic rats with 200 μg ofSIA-II led to a significant reduction in the nonfasting BGL(Fig. 2B) without fasting hypoglycemia (Fig. 2C). No significantdifference was observed whether SIA-II was injected subcuta-neously or intramuscularly. Daily insulin dosage (4 units∕kg)led to reduced BGL, nonfasting value of 350–450 mg∕dL, andfasting value of 300–400 mg∕dL for bovine insulin and 350–400 mg∕dL (random-fed state) and 240–260 mg∕dL (fasting)for rH insulin. Twice daily injection of 4 units∕kg insulin (bovineand rH insulin) marginally improved the glycemic levels. While
Fig. 1. Formation of fibrils by insulin at 37 °C (A) Kinetics of fibril formation at pH 7.0 and 2.0, monitored with 50 μMTh-T for both bovine and rH insulin. (B) Invitro release of insulin from SIA-II (bovine and rH) intermediatemonitored by absorbance (A) at 280 nm and intrinsic tyrosine fluorescence (F). The Th-T intensityof solution inside the dialysis membrane at 0 h and 15 days is also given. (C andD) Congo-red binding studies with native insulin, SIA-I, SIA-II, SIA-III, and amyloidinsulin (rH and bovine). (E) Morphology of SIA-II intermediates and insulin fibrils using ATM: (i) insulin monomer, (ii and v) SIA-I, (iii and vi) SIA-II, (iv and vii)SIA-III intermediate, (viii) fully grown fibers at pH 7.0, 37 °C, (ix) SIA-II intermediate formed at 6 h, pH 2.0, and (x) amyloid fibril formed at 20 h, pH 2.0, 37 °C.(F) Twenty percent Coomassie stained gel showing a cleavage pattern of (i) rH insulin, (ii) SIA-I (6–8 h), (iii) SIA-II (12–16 h), and (iv) SIA-III (26 h) with differentdilutions of 2 mg∕mL proteinase K (lane 1, marker; lane 2, no proteinase K; lane 3, 1∶1;000; lane 4, 1∶2;000, and lane 5, 1∶5;000). Results are mean� SD of fivedifferent experiments performed in triplicate. B, bovine insulin; rH, recombinant human insulin.
Gupta et al. PNAS ∣ July 27, 2010 ∣ vol. 107 ∣ no. 30 ∣ 13247
APP
LIED
BIOLO
GICAL
SCIENCE
S

the daily dose of insulin was able to maintain glucose levels atmoderately high values, a single injection of SIA-II insulin wassufficient for achieving near-normoglycemic postprandial levels(201� 25.96 mg∕dL and 206� 38.68 mg∕dL in s.c. and i.m.,respectively) in diabetic rat for about 135 days without fastinghypoglycemia (124� 11.82 mg∕dL and 130� 12.9 mg∕dL ins.c. and i.m., respectively) where the oligomer was made of bovineinsulin. Administration of rH insulin SIA-II resulted in a moredesirable glycemic control with BGL of 160� 13.61 mg∕dL(fed state) and 105� 15 mg∕dL (fasting) for up to 160 days witha single injection (both s.c. and i.m.) (Fig. 2 B andC). BGL for theinitial 6 days for both bovine and rH SIA-II are given in SI Text(Fig. S2 C–F). Body weight, which is an indicator of normalhealth, was monitored over the entire period of treatment. Therewas an initial loss of body weight immediately after STZ injection(Fig. 3A and Fig. S2B), but progressive weight gains wereachieved in diabetic rats after treatment with insulin SIA-II,and the curve of SIA-II treated animals paralleled that of non-diabetic control. Thus, the striking efficacy of insulin SIA-IItherapy as a long-term treatment of DM-1 in diabetic animalsover that observed for the conventional therapy constitutes akey finding of this study.
Glucose Tolerance Test (GTT). GTT was performed to assess theinstantaneous release of insulin monomers from SIA-II in vivo,from the site of injection. The biological activity of the releasedmonomer was estimated by blood glucose disposal from theblood. Bovine insulin, SIA-II, and in vitro released monomers,significantly improved the glucose tolerance in treated animals,with their elevated BGL of 600 mg∕dL due to glucose infusionreturning to the prechallenged values within 1.5 h. However,in diabetic controls treated with PBS, BGL remained hypergly-cemic over the experimental period (Fig. 3B).
Serum Insulin Level.Optimum BGL were sustained for a period ofabout 135 days for bovine insulin and 160 days for rH insulin, witha single dose of the insulin SIA-II. From the day of injection andthroughout the experimental period, a sustained level of rH/
bovine insulin (0.5–1.2 ng∕mL) into the serum was observed(Fig. 3C) compared to endogenous level of rat insulin(0.08 ng∕mL) in PBS-treated diabetic rats. Serum bovine insulinlevels were also quantified for GTT (Fig. 3D), which were expect-edly negligible for both diabetic and nondiabetic controls, asno SIA-II was administered to these. In contrast, the values fornative insulin and in vitro released monomers from SIA-IIincreased to ∼0.9 ng∕mL in 30 min and then decreased in thenext 4.5 h. In SIA-II treatment (s.c.), insulin levels in serum
Fig. 2. In vivo efficacy of SIA-II (both bovine and rH) in glucose homeostasis. (A) Preprandial/fasting blood glucose profile of diabetic animals in response todifferent SIA-II dosages. (B) Postprandial BGL and (C) preprandial BGLs of nondiabetic rats (control) and diabetic rats treated with PBS-200 μL; insulin singledose-4 units∕kg body weight (b.wt) in the morning; insulin double dose- 4 IU∕kg b.wt in the morning and evening; SIA-II (SC, IM)- 200 μg of SIA-II administeredeither s.c. or i.m., monitored over a period of 135 days for bovine (Left) and 160 days for rH (Right) SIA-II in diabetic rats. n ¼ 10 animals in each group.P ¼< 0.001 STZ vs. insulin/SIA-II; insulin vs. SIA-II, P ¼ 0.462 fasting control vs. SIA-II.
Fig. 3. (A) Body weight profile of SIA-II treated rats. (B) Blood glucoseprofile of GTT. (C) Quantification of serum (bovine or recombinant human)insulin ELISA in STZ-treated rats in response to insulin (bovine or rH) SIA-IIinjected s.c. or i.m.. (D) Quantification of serum bovine insulin of GTT. Resultsare mean� SD of three different experiments. n ¼ 10 animals in each group.n ¼ 5 animals in each group. P ¼< 0.001 STZ vs. insulin/SIA-II/released, P ¼ 1.0insulin vs. released insulin from SIA-II, P ¼ 0.234 control vs. SIA-II.P ¼ 0.548 insulin level (s.c.) vs. (i.m.), P ¼< 0.05 STZ vs. insulin/ SIA-II,P ¼ 0.002 control vs. STZ.
13248 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1005704107 Gupta et al.
reached 0.8–0.9 ng∕mL in 30 min and were sustained over theperiod of study (Fig. 3D). Thus insulin is released instantaneouslyfrom the depot formed at the site of injection.
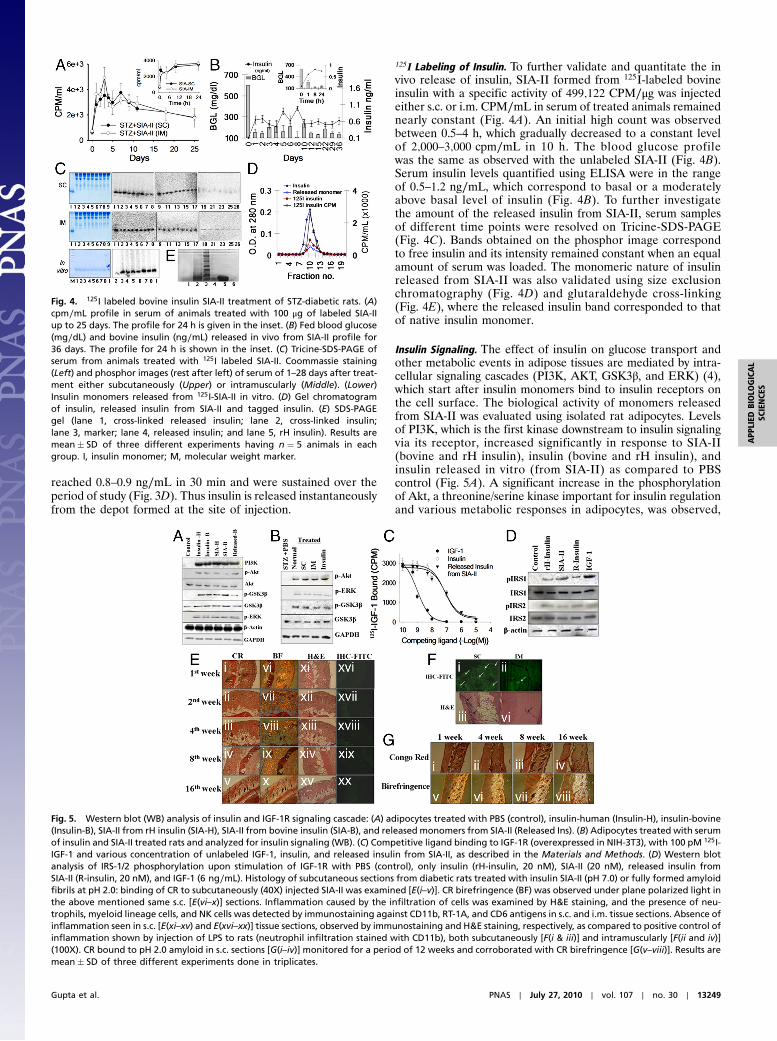
125I Labeling of Insulin. To further validate and quantitate the invivo release of insulin, SIA-II formed from 125I-labeled bovineinsulin with a specific activity of 499;122 CPM∕μg was injectedeither s.c. or i.m. CPM∕mL in serum of treated animals remainednearly constant (Fig. 4A). An initial high count was observedbetween 0.5–4 h, which gradually decreased to a constant levelof 2;000–3;000 cpm∕mL in 10 h. The blood glucose profilewas the same as observed with the unlabeled SIA-II (Fig. 4B).Serum insulin levels quantified using ELISA were in the rangeof 0.5–1.2 ng∕mL, which correspond to basal or a moderatelyabove basal level of insulin (Fig. 4B). To further investigatethe amount of the released insulin from SIA-II, serum samplesof different time points were resolved on Tricine-SDS-PAGE(Fig. 4C). Bands obtained on the phosphor image correspondto free insulin and its intensity remained constant when an equalamount of serum was loaded. The monomeric nature of insulinreleased from SIA-II was also validated using size exclusionchromatography (Fig. 4D) and glutaraldehyde cross-linking(Fig. 4E), where the released insulin band corresponded to thatof native insulin monomer.
Insulin Signaling. The effect of insulin on glucose transport andother metabolic events in adipose tissues are mediated by intra-cellular signaling cascades (PI3K, AKT, GSK3β, and ERK) (4),which start after insulin monomers bind to insulin receptors onthe cell surface. The biological activity of monomers releasedfrom SIA-II was evaluated using isolated rat adipocytes. Levelsof PI3K, which is the first kinase downstream to insulin signalingvia its receptor, increased significantly in response to SIA-II(bovine and rH insulin), insulin (bovine and rH insulin), andinsulin released in vitro (from SIA-II) as compared to PBScontrol (Fig. 5A). A significant increase in the phosphorylationof Akt, a threonine/serine kinase important for insulin regulationand various metabolic responses in adipocytes, was observed,
Fig. 5. Western blot (WB) analysis of insulin and IGF-1R signaling cascade: (A) adipocytes treated with PBS (control), insulin-human (Insulin-H), insulin-bovine(Insulin-B), SIA-II from rH insulin (SIA-H), SIA-II from bovine insulin (SIA-B), and released monomers from SIA-II (Released Ins). (B) Adipocytes treated with serumof insulin and SIA-II treated rats and analyzed for insulin signaling (WB). (C) Competitive ligand binding to IGF-1R (overexpressed in NIH-3T3), with 100 pM 125I-IGF-1 and various concentration of unlabeled IGF-1, insulin, and released insulin from SIA-II, as described in the Materials and Methods. (D) Western blotanalysis of IRS-1/2 phosphorylation upon stimulation of IGF-1R with PBS (control), only insulin (rH-insulin, 20 nM), SIA-II (20 nM), released insulin fromSIA-II (R-insulin, 20 nM), and IGF-1 (6 ng∕mL). Histology of subcutaneous sections from diabetic rats treated with insulin SIA-II (pH 7.0) or fully formed amyloidfibrils at pH 2.0: binding of CR to subcutaneously (40X) injected SIA-II was examined [E(i–v)]. CR birefringence (BF) was observed under plane polarized light inthe above mentioned same s.c. [E(vi–x)] sections. Inflammation caused by the infiltration of cells was examined by H&E staining, and the presence of neu-trophils, myeloid lineage cells, and NK cells was detected by immunostaining against CD11b, RT-1A, and CD6 antigens in s.c. and i.m. tissue sections. Absence ofinflammation seen in s.c. [E(xi–xv) and E(xvi–xx)] tissue sections, observed by immunostaining and H&E staining, respectively, as compared to positive control ofinflammation shown by injection of LPS to rats (neutrophil infiltration stained with CD11b), both subcutaneously [F(i & iii)] and intramuscularly [F(ii and iv)](100X). CR bound to pH 2.0 amyloid in s.c. sections [G(i–iv)] monitored for a period of 12 weeks and corroborated with CR birefringence [G(v–viii)]. Results aremean� SD of three different experiments done in triplicates.
Fig. 4. 125I labeled bovine insulin SIA-II treatment of STZ-diabetic rats. (A)cpm∕mL profile in serum of animals treated with 100 μg of labeled SIA-IIup to 25 days. The profile for 24 h is given in the inset. (B) Fed blood glucose(mg∕dL) and bovine insulin (ng∕mL) released in vivo from SIA-II profile for36 days. The profile for 24 h is shown in the inset. (C) Tricine-SDS-PAGE ofserum from animals treated with 125I labeled SIA-II. Coommassie staining(Left) and phosphor images (rest after left) of serum of 1–28 days after treat-ment either subcutaneously (Upper) or intramuscularly (Middle). (Lower)Insulin monomers released from 125I-SIA-II in vitro. (D) Gel chromatogramof insulin, released insulin from SIA-II and tagged insulin. (E) SDS-PAGEgel (lane 1, cross-linked released insulin; lane 2, cross-linked insulin;lane 3, marker; lane 4, released insulin; and lane 5, rH insulin). Results aremean� SD of three different experiments having n ¼ 5 animals in eachgroup. I, insulin monomer; M, molecular weight marker.
Gupta et al. PNAS ∣ July 27, 2010 ∣ vol. 107 ∣ no. 30 ∣ 13249
APP
LIED
BIOLO
GICAL
SCIENCE
S

similar to the response obtained for free insulin. Phosphorylationof GSK3β on Ser-21 increased several fold in adipocytes treatedwith either free insulin or SIA-II compared to control (Fig. 5A).Therefore, both Akt and GSK3β showed rapid and pronouncedresponse to SIA-II, and the results were similar to responsesobserved on stimulation with free insulin (used as positive con-trol). A significant activation of ERK1/2 was observed comparedto PBS control in case of treated adipocytes. Similar activationof signaling mediators was observed, when serum from insulinor its SIA-II-treated animals was added to cultured adipo-cytes (Fig. 5B).
Insulin-Like Growth Factor 1 Receptor (IGF-1R) Binding Studies. Dueto the structural similarity of insulin with insulin-like growthfactor 1 (IGF-1), it is imperative to study the affinity of insulinreleased from the SIA-II toward binding to IGF-1R. Altered(i.e., increased) affinity may result in the activation of mitogenicresponse upon binding. To exclude such a possibility and confirmthat the released monomers from the SIA-II are unaltered insulinmonomers, the affinity of rH insulin, SIA-II, and IGF-1 to bind toIGF-1R was studied. NIH-3T3 cells overexpressing the humanIGF-1R were utilized to perform the competition binding assaywith both labeled (125I) and unlabeled rH insulin and IGF-1 andanalyzed by Scatchard plot. As evident from Fig. 5C, the ED50 ofIGF-1 binding to IGF-1R was determined to be 1.00� 0.216 nM,whereas that of insulin and insulin released from SIA-II was0.134� 0.021 μM and 0.175� 0.03 μM, respectively. Low con-centration of IGF-1 was able to displace 125IGF-1, in contrastto high concentration of insulin and released insulin. Thus theaffinity of released insulin from the SIA-II for IGF-1R is notaltered. This was also validated by Western blot, where thedifferential phosphorylation profile of IRS-1/2 was similar forboth insulin and insulin released from SIA-II (less phosphoryla-tion of IRS-1), and is distinct from that observed for IGF-1 (moreIRS-1 phosphorylation) (Fig. 5D). IRS-1 is preferentially phos-phorylated upon binding of IGF-1 to IGF-1R, whereas IRS-2phosphorylation is more when insulin binds to insulin receptor.Ligand binding to IGF-1R was completely obliterated in thepresence of IGF-1R antibody, but retained when antibody toinsulin receptor was used (Table S1).
CR, H&E, and Immunostaining of Subcutaneous and Muscle Tissues.Diabetic rats treated with SIA-II and amyloid formed at pH7.0 and 2.0 were monitored for the presence of the depot andthe residual amount of SIA-II or amyloid for 1, 2, 4, 8, and16 weeks using CR staining. Tissues obtained after 1 week ofSIA-II (pH 7.0) injection were found to bind CR, confirmingthe presence of higher amounts of SIA-II [Fig. 5E(i) andFig. S3A(i)]. The amount of bound CR thereafter decreased ina time-dependent manner [Fig. 5E(ii–iv) and Fig. S3A(ii–iv)]and was considerably lower after 16 weeks [Fig. 5E(v) andFig. S3A(v)], corroborated with a birefringence of bound CR[Fig. 5E(vi–x) and Fig. S3A(vi–x)]. The same tissues were alsochecked for inflammation caused, if any, due to insulin SIA-IIby H&E and immunostaining. As compared to the sections fromLPS-injected rats, where infiltration of proinflammatory cellswere visible in both subcutaneous [Fig. 5F(i) and (iii)] and muscletissues [Fig. 5F(ii) and (iv)], SIA-II-injected sections did not showsignificant inflammation even after 16 weeks in both immunos-tained [Fig. 5E(xvi–xx) and Fig. S3A(xi–xv)] and H&E [Fig. 5E(xi–xv)] sections, confirming the noninflammatory and nonimmu-nogenic nature of the SIA-II being used. As described earlier,amyloid fibrils formed at pH 2.0 bound to CR very efficientlyand its intensity reduced negligibly over the course of our study,validating the absence of insulin release from these mature fibers[Fig. 5G(i–viii)]. The possibility of deposition of higher oligomersof injected SIA-II in heart, kidney, and liver was ruled out by theinsignificant CR binding to these tissue sections even after
16 weeks of treatment with SIA-II [Fig. S3B(i–vi)], as these tissuesmaintained normal cellular morphology [Fig. S3B(vii–ix)].
Reduction of Secondary Complications Related to Hyperglycemia inType 1 Diabetes. Type 1 diabetic rats show a marked decreasein skeletal muscle (∼20–40%) and abdominal fat (>60%) (3,5). In contrast, SIA-II-treated diabetic animals had normal bodyweight and were healthy. Developments of cataract in insulin-treated diabetic rats as well as in untreated diabetic rats wereseen, whereas no cataract was observed in animals treated withSIA-II (Fig. S3C). Moreover, functions of liver and kidney, thetwo main organs of the body that typically and severely getaffected in diabetes, were almost normal in SIA-II-treated ratsas compared to their untreated or free insulin infused counter-parts (Table S2).
Absence of Insulin Degrading Enzyme (IDE) and Screening of Antibo-dies Against Insulin. Subcutaneous resistance to insulin in DM-1diabetes is a rare syndrome (6), defined as the lack of biologicalactivity of subcutaneously injected insulin; nevertheless, efficacyof intravenously infused insulin is retained. It has been reportedthat IDE degrades insulin specifically (7) and the partiallydegraded insulin is reabsorbed into the circulation with increasedimmunogenicity but no biological activity. To check the develop-ment of such a resistance in insulin SIA-II-treated animals,biological activity of IDE and the presence of antiinsulin antibo-dies in serum were tested. IDE activity was found to be negligibleand comparable to that of normal rats in tissue homogenate ofSIA-II treated animals (Fig. S4A). Similarly antiinsulin antibodieswere not observed even after 16 weeks of treatment, supportingthe nonimmunogenecity of insulin SIA-II and the absence of IDEinduction (Fig. S4B).
DiscussionIn DM-1, maintenance of normal glycemic levels throughout theday is a great concern, which is not fulfilled efficiently by theexisting therapies. Patients with DM-1 need insulin once or twicea day to sustain normoglycemia. In addition to this, postprandialglucose homeostasis is maintained through regular insulin injec-tions before each meal. Intensive insulin therapy delays the onsetand/or slows the progression of secondary complications. Long-acting insulin analogues, such as glargine and detemir, achievemore constant glucose levels but suffered a setback due to induc-tion of fasting hypoglycemia (8). Therefore, a formulation thatreleases insulin in a controlled manner for longer periods of timewithout any burst or rapid release would free the patients fromthe need to administer multiple doses of insulin.
We employed the remarkable properties exhibited by theinsulin oligomer, such as increased stability, protease resistance,and longer shelf life, to develop an insulin prodrug—supramole-cular insulin assembly II, which releases the hormone in acontrolled fashion for extended periods of time and can beharnessed for therapeutic purposes. SIA-II is a prodrug, whereneither any chemical modification was done for its formationin vitro nor is chemical cleavage required for achieving itstherapeutic effect in vivo. The intrinsic property of the protein tounfold and oligomerize has been harnessed here to achieve aglucose regulatory effect. Moreover, the period of sustainedrelease is 100 times greater than any of the studies reportedearlier (9). SIA-II selected by us was observed to be a linear asso-ciation of elongated oligomers by AFM, which released insulinmonomers at a constant rate. The SIA-II and SIA-III were resis-tant to protease cleavage by proteinaseK, in comparison to insulinand SIA-I, which were more susceptible. The chosen intermediatedid not show significant CR binding and corresponded to a sharpIR band position at 1;647–1;645 cm−1, unlike the amyloid state ofprotein.
13250 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1005704107 Gupta et al.

The biological effectiveness of insulin is generally assessed byits ability to regulate glycemic levels in the blood. Resultantdecrease in the blood glucose concentration represents the mostnoticeable and therapeutically the most important effect of insu-lin. As hypothesized, SIA-II treatment reduced the severity ofhyperglycemia and a single dose of 200 μg maintained near-nor-moglycemic levels (160� 13.6 mg∕dL) in diabetic animals forabout 120–140 days. To determine whether improvement inglycemic control by SIA-II was accompanied with increased riskof fasting hypoglycemia, treated animals were subjected to weeklyextended fasting of 18–20 h (Fig. S5 A and B). These fasted ani-mals were able to maintain their fasting BGL within the normalrange (60–110 mg∕dL). Thus in comparison to conventionalinsulin therapy, SIA-II treatment achieved a tightly regulatedglycemic control without fasting hypoglycemia. GTT conductedto determine the onset of the action of insulin SIA-II in caseof induced severe hyperglycemia demonstrated no lag phase inthe release of bioactive insulin monomer from the SIA-II depot.Continuous subcutaneous insulin infusion using insulin pumptherapy had been developed earlier to overcome the problemof multiple daily injections and was fairly successful in maintain-ing BGL (10). However, the extra cost of the pump, pumpmalfunctions, bacterial infections in subcutaneous catheters,delayed and unpredictable absorption from the subcutaneoussite, variability of insulin sensitivity, diabetic gastroparesis, andneed of trained personnel to supervise added to the woes of thistherapy (11, 12).
To assess the in vivo release of human recombinant insulin fromSIA-II, ELISA was performed. A sustained release of insulin(0.5–0.9 ng∕mL) sufficient for maintaining normoglycemia wasseen, which correlated well with the duration of the treatment.Radiolabeling of insulin with 125I was done to validate the releasekinetics of monomers from insulin. Amount of insulin quantifiedby ELISA in the serum of treated rats was comparable to thelabeled insulin released in the serum upon administration of499;122 CPM∕μg of SIA-II. Constant release of insulin, with noburst phase, from the depot formed at the site of injection followszero-order kinetics, a beneficial requirement for the sustainedand extended period of treatment seen with SIA-II. Radioactiveserum samples resolved onto Tricine-SDS-PAGE showed bandscorresponding to insulin monomer, which was also confirmedby size exclusion chromatography and glutaraldehyde cross-link-ing studies. These monomers triggered insulin signaling cascadeefficaciously by the activation of intracellular mediators (PI3K,Akt, GSK3β, and ERK1/2) in cultured adipocytes, suggesting thatthe monomers released from SIA-II are biologically active.Storage did not alter the blood glucose lowering effect of SIA-II. Slow release of insulin was again ascertained by histologicalstudies of tissues from rats injected with SIA-II, which alsodemonstrated the formation of a depot at the site of injection fromwhich insulin is released over a period of time and resulted in adecrease of the residual SIA-II. No inflammation was observedat the site of injection even after 16 weeks.
The SIA-II therapy conferred profound physiological benefitsin diabetic animals, which partially reflected an improved
glycemic control as well as markedly reduced urea and creatinineconcentrations due to improved liver and kidney functions, res-pectively. Absence of antiinsulin antibodies and inability to elicitIDE in or around the site of injection are important factors inprolonged beneficial antidiabetic affect.
The report presented here shows that insulin released fromSIA-II is equivalent to soluble insulin as far as its biologicalfunction is concerned. A significant difference lies in the durationof action. Wheras for the standard insulin injection, it is only6–10 h (7), for SIA-II the duration of action is markedly enhancedup to 135 days for bovine and 160 days for rH insulin. This may beattributed to the remarkable stability of insulin in the SIA-II state,which forms a depot at the site of injection for supply of thephysiologically most relevant form of insulin, namely, insulinmonomers, for a longer period of time. In contrast, any peptideor protein drug administered is cleared and/or degraded from thebody at a faster rate, thereby decreasing the window period of itsefficacy. Studies with SIA-II as a therapeutic for diabetes type 1was also carried out in other model systems such as in mice andrabbit and also using the Alloxan for induction of diabetes apartfrom STZ (Fig. S5 C and D). Similar beneficial outcomes werenoted in these models of diabetes also (Fig. S6 A–D). In sum-mary, we have demonstrated the efficacy as well as the feasibilityof insulin SIA-II treatment for the significant improvement ofBGL without causing fasting hypoglycemia in STZ-induceddiabetic animal model. Unlike intensive insulin therapy, by whichBGLs are controlled by an increased frequency of insulin injec-tion with concomitant risk of hypoglycemia (3), the significantlyimproved glycemic control using insulin SIA-II therapy is accom-plished without multiple insulin injections, which displays noadverse side effects.
Materials and MethodsInsulin Fibrilization. Bovine or recombinant human insulin (Sigma), 2 mg∕mL(28 units∕mg), was dissolved in PBS at pH 7.0 or hydrochloric acid in water(pH 2.0) and incubated at 37 °C for 7 days with constant agitation at180 rpm. The process of oligomerization was monitored by turbidity at600 nm and by Th-T fluorescence.
In Vitro Monomer Release Kinetics. Aliquots of 200 μL were withdrawn atdifferent time points from the insulin fibrilization reaction at pH 7.0 and2.0, 37 °C. Insulin oligomers produced at different time points were isolatedby centrifugation at 10,000 rpm for 10 min. The pellet obtained was washedwith PBS and resuspended in 1 mL PBS in an Eppendorf tube, cap wasremoved, and the Eppendorf was sealed with 8-kDa cutoff dialysis mem-brane. The Eppendorf tube was suspended in 20 mL PBS containing0.02% sodium azide in a 50-mL Falcon tube and the kinetics of insulin releaseinto PBS was monitored spectrophotometrically at 280 nm and by its intrinsic(tyrosine) fluorescence for 15 days. The amount of insulin released per hourwas calculated.
A complete and detailed description of the methods is included in SI Text.
ACKNOWLEDGMENTS. The authors thank Dr. Ira Surolia and Dr. Shiv Pillai(Massachusetts General Hospital) for a critical reading of the manuscript.
1. Brown A, et al. (2010) Intensive glycemic control and cardiovascular disease: Anupdate. Nat Rev Cardiol doi: 10.1038/nrcardio.2010.35.
2. Burn Paul (2010) Type 1 diabetes. Nat Rev Drug Discov 9:187–188.3. Smith JF, et al. (2006) Characterization of the nanoscale properties of individual
amyloid fibrils. Proc Natl Acad Sci USA 103:15806–15811.4. Paul Bevan (2001) Insulin signalling. J Cell Sci 114:1429–1430.5. Nathan DM, et al. (2005) Intensive diabetes treatment and cardiovascular disease in
patients with type 1 diabetes. N Engl J Med 353:2643–53.6. Paulsen EP, et al. (1979) Insulin resistance caused by massive degradation of subcuta-
neous insulin. Diabetes 28:640–645.7. Duckworth WC, et al. (1998) Insulin degradation: Progress and potential. Endocr Rev
19:608–624.
8. Hirsch B (2005) Insulin Analogues. New Eng J Med 352:174–183.
9. Gershonov E, et al. (2000) A novel approach for a water-soluble long-acting insulin
prodrug: Design, preparation, and analysis of [(2-sulfo)-9-fluorenylmethoxycarbo-
nyl](3)-insulin. J Med Chem 43:2530–7.
10. Pickup JC, Renard E (2008) Long-acting insulin analogs versus insulin pump therapy for
the treatment of type 1 and type 2 diabetes. Diabetes Care 31:S140–S145.
11. Nowakowska M, et al. (2007) Bacterial strains colonizing subcutaneous catheters of
personal insulin pumps. Pol J Microbiol 56:239–43.
12. Shalitin S, Phillip M (2008) Hypoglycemia in type 1 diabetes: A still unresolved problem
in the era of insulin analogs and pump therapy. Diabetes Care 31(Suppl 2):S121–S122.
Gupta et al. PNAS ∣ July 27, 2010 ∣ vol. 107 ∣ no. 30 ∣ 13251
APP
LIED
BIOLO
GICAL
SCIENCE
S








![Supramolecular anion recognition in water: synthesis of ... · Supramolecular anion recognition in water: synthesis of hydrogen-bonded supramolecular frameworks ... (TP) 2] n taken](https://img.dokumen.tips/doc/110x75/5b9ce37509d3f2321b8d8473/supramolecular-anion-recognition-in-water-synthesis-of-supramolecular-anion.jpg)




![7. Supramolecular structures - Acclab h55.it.helsinki.fiknordlun/nanotiede/nanosc7nc.pdf · 7. Supramolecular structures [Poole-Owens 11.5] Supramolecular structures are large molecules](https://img.dokumen.tips/doc/110x75/5f071ded7e708231d41b63bf/7-supramolecular-structures-acclab-h55it-knordlunnanotiedenanosc7ncpdf.jpg)















