-
Supporting InformationWefers et al. 10.1073/pnas.1218721110SI
Materials and MethodsSoftware for Transcription Activator-Like
Effector-Binding Site Detection.Software for the identification of
transcription activator-like (TAL)effector-binding sites is written
in Perl 5.8.8 programming languageandusesBioperl libraries for
parsing inputDNAsequences (1).TheTAL effector nuclease
(TALEN)–Designer software takes DNAsequences as input and searches
for 5′-to-3′ oriented homo- andheteromeric potential binding motifs
on opposite strands of theinput DNA sequences. The TAL
effector-binding sites have eithera length of 15 nt or a variable
length of 13–15 nt. TheWeb interface(www.talen-design.de) takes one
or multiple FASTA-formattedDNA sequences as input. The Web
interface is running on anApache Web Server 2.2.3 on SUSE Linux
Enterprise Server 10Service Pack 2 (SUSE LINUX Products GmbH,
Nürnberg, Ger-many). The user can select one of the search pattern
as describedabove and can customize the length of the spacer
between the TALeffector-binding sites in a range of 13–17 nt.
Potential binding sitesare only detected, if they are preceded by a
thymine in position -1of theDNA sequence on opposite strands.
Potential bindingmotifson the reverse complement of the input DNA
sequences are au-tomatically identified as well. Replied to the
user are the identifierand length of the input DNA sequences,
theDNA sequences of themotif identified with colored heteromeric
binding sites and thespacer in between, the total length of the TAL
effector-bindingmotif, start and end coordinates of the motif with
regard tothe input sequence where the motif has been identified,
and theguanine content of the TAL effector-binding motifs.The
software for genome-wide searches for TAL effector-
binding sites uses theEnsembl application program interface
(API)(Release 6; www.ensembl.org) to download genomic sequences
ofeach exon of themouse (Mus musculus) genome including 40 nt
ofintronic sequence flanking each of the exons. Input sequences
aresearched with two patterns consisting of 15 nt for each of the
twoTAL effector-binding sites preceded by a T and either a
fixed-length spacer of 15 nt or a more flexible spacer of 14 or 16
nt. Thebinding sites identified are stored as genomic coordinates
ina MySQL database (5.0.26). For display of the TAL
effector-binding sites, which allows user-specific searches, we
installeda local instance of the genome browser GBrowse Version 2.0
(2).Turning on and off genome browser tracks allows the user
todisplay the transcripts of each of the genomes and their
exon–intron structure and the identified TAL effector-binding
sites,respectively (Fig. S1).
Nuclease Expression Vectors. The Rab38-specific zinc-finger
nucle-ase (ZFN) expression vectors pCAG-Rab-ZFN-L and -R
weredescribed previously (3). For the expression of TALENs
inmammalian cells, we designed the generic expression
vectorpCAG-TALEN, which contains a CAG hybrid promoter regionand a
transcriptional unit comprising a sequence coding for theN-terminal
amino acids 1–176 of TAL nucleases, located up-stream of a pair of
BsmBI restriction sites. This N-terminal regionsincludes an ATG
start codon, a nuclear localization sequence,a FLAG Tag sequence, a
glycine-rich linker sequence, a segmentcoding for 110 aa of the TAL
protein AvrBs3, and the invariableN-terminal TAL repeat of the Hax3
TAL effector. Downstream ofthe central BsmBI sites, the
transcriptional unit contains 78 codons,including an invariable
C-terminal TAL repeat and 44 residuesderived from the TAL protein
AvrBs3, followed by the codingsequence of the FokI nuclease domain
and a polyadenylation signalsequence. DNA segments coding for
arrays of TAL repeats can beinserted into the BsmBI sites of
pCAG-TALEN in frame with the
up- and downstream coding regions to enable the expression
ofpredesigned TAL-Fok nuclease proteins. To derive TAL
elementDNA-binding domains, we used the TAL effectormotif (repeat)
11of the Xanthomonas Hax3 protein
(LTPEQVVAIASNIGGKQA-LETVQRLLPVLCQAHG) to recognize A, the TAL
effectormotif 5 (LTPQQVVAIASHDGGKQALETVQRLLPVLCQAHG)derived from
the Hax3 protein to recognize C, and the TALeffector motif 4
(LTPQQVVAIASNGGGKQALETVQRLLP-VLCQAHG) from the Xanthomonas Hax4
protein to recognizeT (bold letters indicate repeat variable
diresidues). To recognizea target G nucleotide, we used the TAL
effector motif 4 from theHax4 protein with replacement of the amino
acids 12 into N and 13intoN
(LTPQQVVAIASNNGGKQALETVQRLLPVLCQAHG).These elements were obtained
by gene synthesis (Genscript) andfurther amplified by PCR using
primers that include BsaI sitesoutside of the coding region. For a
15-bp TALEN target sequence,seven elements each are pooled in a
pair of reactions together withBsaI and T4DNA ligase to create and
ligate unique overhangs. Full-length ligation products were
recovered by gel extraction and in-serted by seamless cloning
(Gibson assembly; NewEngland Biolabs)into pCAG-TALEN opened with
BsmBI. The integrity of allTALENexpression vectors was confirmed
byDNA sequencing. Thecomplete sequence of the vector encoded TALEN
proteins andrepeat variable diresidue (RVD) arrays is shown below
[blue letters,nuclear localization sequence; red letters, FLAG tag;
green letters,AvrBs3 N- and C-terminal sequences; black letters,
TAL repeats(RVDs are underlined); orange letters, FokI nuclease
domain; grayletters, linker sequences]:
TALENRab38 (1).
MGPKKKRKVAAADYKDDDDKPGGGGSG-GGGVPASPAAQVDLRTLGYSQQQQEKIKPKVRSTVAQ-HHEALVGHGFTHAHIVALSQHPAALGTVAVKYQDMIA-ALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPP-LQSGLDTGQLLKIAKRGGVTAVEAVHAWRNALTGAPL-NLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTP-QQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPQQV-VAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIA-SNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIG-GKQALETVQRLLPVLCQAHGLTPQQVVAIASNNGGKQ-ALETVQRLLPVLCQAHGLTPQQVVAIASHDGGKQALE-TVQRLLPVLCQAHGLTPQQVVAIASNNGGKQALETVQ-RLLPVLCQAHGLTPQQVVAIASHDGGKQALETVQRLL-PVLCQAHGLTPQQVVAIASNGGGKQALETVQRLLPVL-CQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAH-GLTPQQVVAIASNGGGKQALETVQRLLPVLCQAHGLT-PQQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPQQ-VVAIASNGGGKQALETVQRLLPVLCQAHGLTPQQVVAI-ASNGGGRPALESIVAQLSRPDPALARSALTNDHLVALA-CLGGRPALDAVKKGLPHAPALIKRTNRRIPEGSDRLN-QLVKSELEEKKSELRHKLKYVPHEYIELIEIARNSTQD-RILEMKVMEFFMKVYGYRGKHLGGSRKPDGAIYTVG-SPIDYGVIVDTKAYSGGYNLPIGQADEMQRYVEENQ-TRNKHINPNEWWKVYPSSVTEFKFLFVSGHFKGNYKA-QLTRLNHITNCNGAVLSVEELLIGGEMIKAGTLTLEEV-RRKFNNGEINF
TALENRab38 (2).
MGPKKKRKVAAADYKDDDDKPGGGGSG-GGGVPASPAAQVDLRTLGYSQQQQEKIKPKVRSTVAQ-HHEALVGHGFTHAHIVALSQHPAALGTVAVKYQDMIA-ALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPP-LQSGLDTGQLLKIAKRGGVTAVEAVHAWRNALTGAPL-NLTPQQVVAIASNNGGKQALETVQRLLPVLCQAHGLT-
Wefers et al. www.pnas.org/cgi/content/short/1218721110 1 of
6
http://www.talen-design.dehttp://www.ensembl.orgwww.pnas.org/cgi/content/short/1218721110
-
PQQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPQQ-VVAIASHDGGKQALETVQRLLPVLCQAHGLTPQQVVA-IASHDGGKQALETVQRLLPVLCQAHGLTPQQVVAIASHD-GGKQALETVQRLLPVLCQAHGLTPQQVVAIASNNGGK-QALETVQRLLPVLCQAHGLTPQQVVAIASNNGGKQAL-ETVQRLLPVLCQAHGLTPQQVVAIASNGGGKQALETV-QRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLL-PVLCQAHGLTPQQVVAIASNNGGKQALETVQRLLPVLC-QAHGLTPQQVVAIASNGGGKQALETVQRLLPVLCQAH-GLTPQQVVAIASNNGGKQALETVQRLLPVLCQAHGLTP-QQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPQQV-VAIASNNGGKQALETVQRLLPVLCQAHGLTPQQVVAIA-SNGGGRPALESIVAQLSRPDPALARSALTNDHLVALA-CLGGRPALDAVKKGLPHAPALIKRTNRRIPEGSDRLN-QLVKSELEEKKSELRHKLKYVPHEYIELIEIARNSTQDR-ILEMKVMEFFMKVYGYRGKHLGGSRKPDGAIYTVGS-PIDYGVIVDTKAYSGGYNLPIGQADEMQRYVEENQT-RNKHINPNEWWKVYPSSVTEFKFLFVSGHFKGNYKAQ-LTRLNHITNCNGAVLSVEELLIGGEMIKAGTLTLEEVR-RKFNNGEINF
Reporter Vectors. The nuclease reporter plasmid pTAL-Rep
con-tains BstBI and NruI sites for cloning of TALEN target
sites(underlined) as oligodeoxynucleotides (ODNs) (Metabion);
Rab38target sense oligonucleotide:
5′-CGGCCACCATGGTCGTATCA-AGCGCTATGTGCACCAAAACTTCTCCTCGCACTACCG-GGCCACCATTGGT-3′;
Rab38 target anti-sense
oligonucleotide:5′-ACCAATGGTGGCCCGGTAGTGCGAGGAGAAGTTTTG-GTGCACATAGCGCTTGATACGACCATGGTGGC-3′.The
integrity of all reporter plasmids was confirmed by DNA
sequencing.
Single-Stranded Oligodesoxynucleotide and Targeting Vector
Design.The single-stranded oligodesoxynucleotide ODNIDG-Cht,
targetingthe first exon of the mouse Rab38 gene, was synthesized by
Met-abion and had a length of 144 nt
(5′-CACCTCACAAGGAGC-ACCTGTACAAGCTGCTGGTGATCGGCGACCTGGTAG-TGGGCAAGACCAGCATTATTAAACGGTACGTGCATCA-AAATTTCTCCTCTCATTATCGAGCCACCATTGGTGTG-GACTTCGCGCTGAAGGTGC-3′).
The targeting vectorpRab38IDG-WT comprised two homology regions
encompassing918 and 2789 bp of genomic sequence flanking exon 1 of
themouse Rab38 gene.
Nuclease Assay in HEK293 Cells. HEK293 cells were grown inDMEM
supplied with 10% FCS (vol/vol) under standard con-ditions. A total
of 1 × 106 cells were transfected by coelec-troporation with 5 μg
of TALEN reporter, 5 μg of each TALENexpression plasmid, and 5 μg
of the luciferase expression plasmidpCMV-FLuc. Control
transfections were done with 5 μg ofTALEN reporter, 10 μg of
pBluescript plasmid DNA, and 5 μg ofluciferase expression plasmid
pCMV-FLuc. After transfection,cells were split into three wells and
treated as technical repli-
cates. Forty-eight hours after transfection, cells were lysed,
andwhole-protein samples were extracted. β-Galactosidase
activitywas determined using the β-galactosidase reporter gene
assay(Roche Diagnostics) following the manufacturer’s
instructions,and signals were detected using a Centro LB 960
luminometer(Berthold Technologies). In parallel, luciferase
activity wasmeasured as transfection control. Luciferase substrate
[25 mMglycylglycin, 15 mM MgSO4, 4 mM EGTA, 2 mM ATP, 1 mMDTT, 100
μM coenzyme A, 75 μM luciferin, 15 mM K2HPO4/KH2PO4 (pH 8.0)] was
added to the samples, and chemilumi-nescent signals were measured
in the luminometer. The triplicateβ-galactosidase values of each
transfected DNA mixture werenormalized in relation to the levels of
luciferase activity of thesame samples, and the mean value and SEM
of β-galactosidaseactivity were calculated. The values derived from
cotransfectionof TALEN expression and reporter plasmids were
comparedwith the transfection of reporter plasmid without TALEN
plas-mids and expressed as relative increase.
Microinjection of One-Cell Embryos. The injection of TALENmRNA
and targeting molecules (vector plasmid or ODN, re-
spectively) was performed as described previously for ZFNs (3,
4).Briefly, TALEN mRNA was prepared by in vitro transcriptionfrom
linearized plasmid DNA using the mMessage mMachine T7Ultra kit and
the MEGAclear kit (Life Technologies). EachTALEN mRNA was then
diluted in injection buffer [10 mM Tris,0.1 mM EDTA (pH7.2)] to a
working concentration of 10 ng/μL.mRNAs were mixed and stored
together with the supercoiledtargeting vector or
oligodesoxynucleotide at −80 °C. The targetingvectors were
precipitated and dissolved in injection buffer toa working
concentration of 15 ng/μL. The targeting ODNs weredissolved in
water and diluted with injection buffer to a workingconcentration
of 15 ng/μL. For the targeting of wild-type alleles,zygotes were
obtained by mating of C57BL/6N males with super-ovulated FVB
females (Charles River) as described (3, 4). Fortargeting of the
Rab38cht allele, zygotes from matings of homo-zygous Rab38cht males
(5) (kindly provided byM. Seabra, ImperialCollege London, London,
UK) with FVB females were used.Zygotes were injected with the
mixture of the targeting constructand the TALENmRNAs in a two-step
procedure, as described (3,4). Briefly, a first aliquot of the
DNA/RNA mixture was injectedinto, whenever possible, the larger
(male) pronucleus to deliverthe DNA vector, as used for the
production of transgenic mice.Upon the withdrawal of the injection
needle from the pronucleus,a second aliquot of the DNA/RNA mixture
was injected into thecytoplasm to deliver the TALEN mRNAs directly
to the trans-lation machinery. Injections were performed using a
Leica mi-cromanipulator and microscope and an Eppendorf
FemtoJetinjection device. Injected zygotes were transferred into
pseudo-pregnant CD1 female mice, and viable adult mice were
obtained.All mice showed normal development and appeared
healthy.Micewere handled according to institutional guidelines
approved bythe animal welfare and use committee of the Regierung
von
TALEN RVD arrays
TALENTAL repeat diresidues (without last
half repeat)
TALENRab38 no. 1 NI NG HD NI NI NN HD NN HD NG NI NGNN NG↓ ↓ ↓ ↓
↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓
Target sequence (5′–3′) A- T- C- A- A- G- C- G- C- T- A- T- G-
T-TALENRab38 no. 2 NN NN HD HD HD NN NN NG NI NN NGNNHD NN
↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓ ↓Target sequence (5′–3′) G - G - C - C
- C - G - G - T - A - G - T - G - C - G -
Wefers et al. www.pnas.org/cgi/content/short/1218721110 2 of
6
www.pnas.org/cgi/content/short/1218721110
-
Oberbayern and housed in standard cages in a specific
pathogen-free facility on a 12 h light/dark cycle with ad libitum
access to foodand water.
Southern Blot Analysis. Southern blot analysis was performed
aspreviously described (4) using the Rab38 5′ probe.
Hybridization
probes were obtained by PCR amplification (P-Rab1:
5′-CTGG-AAACTAAAATTCAAGGTGTTATAC-3′; P-Rab2:
5′-TATT-CATTCACTTAACCATTTGTTC-3′) from C57BL/6N genomicDNA,
purified with the Qiaquick PCR Purification Kit (Qiagen),cloned
with the Strataclone PCR cloning kit (Agilent Technolo-gies), and
confirmed by sequencing.
1. Stajich JE, et al. (2002) The Bioperl toolkit: Perl modules
for the life sciences. GenomeRes 12(10):1611–1618.
2. Stein LD, et al. (2002) The generic genome browser: A
building block for a modelorganism system database. Genome Res
12(10):1599–1610.
3. Meyer M, Ortiz O, Hrabé de Angelis M, Wurst W, Kühn R (2012)
Modeling diseasemutations by gene targeting in one-cell mouse
embryos. Proc Natl Acad Sci USA109(24):9354–9359.
4. Meyer M, de Angelis MH, Wurst W, Kühn R (2010) Gene targeting
by homologousrecombination in mouse zygotes mediated by zinc-finger
nucleases. Proc Natl Acad SciUSA 107(34):15022–15026.
5. Loftus SK, et al. (2002) Mutation of melanosome protein RAB38
in chocolate mice. ProcNatl Acad Sci USA 99(7):4471–4476.
Wefers et al. www.pnas.org/cgi/content/short/1218721110 3 of
6
www.pnas.org/cgi/content/short/1218721110
-
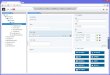
Fig. S1. Screenshot of the TALEN-Designer browser
(www.talen-design.de). (A) Region overview showing the different
transcripts of mouse PR domain zincfinger protein 13 gene (Prdm13)
(Ensembl genome database, www.ensembl.org. Accession no.
ENSMUSG00000040478). The potential TAL effector-bindingsites
identified with search patterns of high and medium stringency are
displayed as separated genome browser tracks, respectively. (B)
Detailed view of exon 2of Prdm13 with a selection of TAL effector
sites showing the binding domain as a green box and the spacer as a
thin black line. The genomic coordinates andthe sequence of pattern
tal2434667 as identified on Prdm13 exon 2 sequence
(ENSMUSE00000731842) is displayed upon activating a hyperlink. In
the FASTA-formatted sequence panel, the two DNA-binding domains are
highlighted by a gray background.
Wefers et al. www.pnas.org/cgi/content/short/1218721110 4 of
6
http://www.talen-design.dewww.ensembl.orgwww.pnas.org/cgi/content/short/1218721110
-
Fig. S2. Screenshot of the TALEN-Designer browser. Distribution
of TALEN target sites in the first exon (202 bp) of the Rab38 gene
(red bar), searching forbipartite 15-bp target sequences (red
boxes), separated by a spacer regions of 14–16 bp (thin line).
Fig. S3. Comparison of TALEN activities. TALENs directed against
sequences within 20 different genes were constructed and tested by
the HEK293 reporterassay for specific nuclease activity against
their target sequences cloned into pTALEN-Rep. The activities
obtained from transfection of the reporter in thepresence or
absence of TALEN expression vectors were compared and expressed as
an index of nuclease activity. These values (filled columns) are
comparedwith the value obtained from transfection of the
Rab38-specific ZFN (ZFNRab38) (red column) (Fig. 1D).
Wefers et al. www.pnas.org/cgi/content/short/1218721110 5 of
6
www.pnas.org/cgi/content/short/1218721110
-
Fig. S4. Coat color of homozygous Rab38IDG-Cht and Rab38wt
littermates. (A) Pup Rc6-4 carrying the Rab38IDG-Cht allele (Fig.
2D) was mated with C57BL/6 wild-type mice, and the derived
heterozygous offspring were intercrossed. Pups derived from these
matings were genotyped by ApaLI digestion of PCR productsamplified
from Rab38, identifying homozygous Rab38wt and heterozygous and
homozygous Rab38IDG-Cht mice. Compared with the black fur of a
Rab38wt/wt
littermate (pup 159), homozygous Rab38IDG-Cht/IDG-Cht mutants
(pup 160) exhibit the dark brown coat color characteristic for
chocolate mutants (5). (B) TheRab38 genotype of pups 159 and 150
was confirmed by sequence analysis of PCR products.
Table S1. Overview of founders and Rab38 alleles obtained from
embryo microinjections
Microinjection experiment No. of pupsFounder
IDNo. of Rab38NHEJ alleles
No. of Rab38HR alleles
No. of founders withNHEJ allele(s)
No. of founders withHR allele(s)
TALENRab38 and ODNIDG-Cht 117 Rc6 1 2 1 1Rc7 2 1 1 1Rc1 1 0 1
0Rc2 1 0 1 0Rc3 1 0 1 0Rc4 1 0 1 0Rc5 1 0 1 0
TALENRab38 and pRab38IDG-WT 50 Rw2 0 1 0 1Rw1 2 0 1 0
∑ 167 9 founders 10 4 8 (4.8%) 3 (1.8%)
NHEJ, nonhomologous end-joining.
Wefers et al. www.pnas.org/cgi/content/short/1218721110 6 of
6
www.pnas.org/cgi/content/short/1218721110


![CRISPR/Cas9-mediated genome editing induces exon skipping ... · HeLa cells can cause skipping of exon 3, exon 4, or exons 3, 4, and 5 [18]. We also detected infrequent exon skipping](https://img.dokumen.tips/doc/110x75/60db8f117fb86d112c69c947/crisprcas9-mediated-genome-editing-induces-exon-skipping-hela-cells-can-cause.jpg)