Embed Size (px)
Citation preview

Sulfation of Tyrosine Residues in Coagulation Factor V
By G.L. Hortin
Sulfation of human coagulation factor V was investigated by biosynthetically labeling the products of HepG2 cells with [36S]sulfate. There was abundant incorporation of the sulfate label into a product identified as factor V by immunoprecipitation, lability to proteases, affinity for the lectin jacalin, and sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Two or more sites in factor V incorpo- rated sulfate as indicated by labeling of different peptide chains of factor Va. The 150-Kd activation fragment of factor Va incorporated the greatest amounts of sulfate. This fragment of factor Va was bound selectively by jacalin-agarose, reflecting its content of 0-linked oligosac- charides. Analysis of an alkaline hydrolysate of sulfate-
OAGULATION FACTOR V, also termed proaccel- C erin, is an essential hemostatic component occurring in plasma as a single-chain glycoprotein with a mass of about 330,000 daltons.’ During activation of the clotting cascade, factor V is cleaved at three sites by thrombin, yielding factor Va,1-5 which is a potent cofactor in the activation of prothrom- bin by factor Xa. Factor Va combines with factor Xa, calcium ions, and a phospholipid membrane to form the prothrombinase complex, which activates prothrombin sev- eral orders of magnitude more rapidly than factor Xa alone.6-8 The cofactor action of factor Va is ended by cleavage by activated protein C.9 Factor V bears consider- able structural homology with factor VIII, the anti- hemophiliac factor, and enacts an analogous physiologic role in coagulation.’,’0-’-’ Hereditary deficiency of functional fac- tor V results in the bleeding diathesis, p a r a h e m ~ p h i l i a . ’ ~ ~ ’ ~
The complete primary sequence of the 2,196 amino acids in factor V has been determined by cloning of complemen- tary DNA sequences.’.’’,’’ However, analysis of purified factor V indicates that the polypeptide chain undergoes extensive posttranslational modification. Carbohydrate ac- counts for approximately 13% of the mass of human factor V,I6 with multiple N-linked and 0-linked oligosaccharides,l-16 and removal of sialic acids or oligosaccharides alters activity of the protein.” Another potential posttranslational modifica- tion of factor V, sulfation of tyrosine residues, is suggested by recent identification of this modification of the homologous protein, factor VIIII8 and by occurrence of amino acid sequences in factor V similar to known sites of sulfation in other protein^.'^,'^ The present study seeks to determine
From The Edward Mallinckrodt Department of Pediatrics,
Submitted October 27, 1989; accepted May 9, 1990. Supported by a grant from the National Institutes of Health. Address reprint requests to Glen L. Hortin, MD. PhD, Box 8116,
Department of Pediatrics. Washington University School of Medi- cine, 400 S Kingshighway, St Louis, M O 631 I O .
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U,S.C. section 1734 solely to indicate this fact.
Washington University School of Medicine. St Louis, MO.
0 I990 by The American Society of Hematology. 0006-4971/90/7605-0022$3.00/0
labeled factor Va by anion-exchange chromatography showed that the sulfate occurred partly in tyrosine sulfate residues and partly in alkaline-labile linkages. Sulfate groups are potentially important structural and functional ele- ments in factor V, and labeling with [36S]sulfate provides a useful approach for examining the biosynthesis and process- ing of this protein. The hypothesis is advanced that sites of sulfation in factor V and several other plasma proteins contribute to the affinity and specificity of thrombin for these molecules, just as it does for the interaction of thrombin with the potent inhibitor hirudin from leeches. 0 1990 by The American Society of Hematology.
whether human factor V also undergoes biosynthetic sul- fation. Identification of this modification has assumed in- creased significance with the recent observations that it can alter the function of macromolecules, such as the fourth component of complement” and fibronectin,” and that tyrosine sulfate residues occur a t thrombin-binding sites in heparin cofactor IIZ3 and h i r ~ d i n . ’ ~ . ~ ~
MATERIALS AND METHODS
[4,5-’H]Leucine (60 Ci/mmol) was obtained from Amersham (Arlington Heights, IL), and carrier-free [’SS]sulfate from ICN Radiochemicals (Irvine, CA). Rabbit antiserum to human factor V was purchased from Accurate (Westbury, NY), and antiserum to human fibrinogen from Calbiochem (San Diego, CA). Protein A-Sepharose was from Pharmacia (Piscataway, NJ). Factor V-defi- cient plasma, other reagents for assay of factor V activity, and thrombin were obtained from Sigma Chemical (St Louis, MO). Jacalin-agarose was purchased from Vector Laboratories (Burlin- game, CA). Hirudin purified from leeches (activity 10,000 U/mg) was from American Diagnostica (New York, NY). Other reagents were purchased from Sigma Chemical.
HepG2 cells, a human hepatoma-derived cell line,*’ were grown to confluence in Earle’s medium with 10% fetal calf serum and 4 mmol/L glutamine. Biosynthetic labeling of proteins was performed for 4 to 16 hours with 1 mCi [35S]sulfate or 0.25 mCi [’Hlleucine in 10 ml serum-free medium deficient in sulfate and leucine. There was no chase incubation. Procedures for immunoprecipitation, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) of samples under reducing conditions, and autoradiography have been described previously.*’
Lectin affinity chromatography of culture medium from HepG2 cells was performed at room temperature using a column of jacalin-agarose with 1 mL bed volume, 0.6 cm in diameter. Samples of 5 mL serum-free culture medium containing labeled products were passed through the column with gravity flow. The column was washed with 10 vol of 0.15 mol/L NaCl or 0.5 mol/L NaC1, each buffered with 20 mmol/L Hepes (pH 7.4). Specifically bound components were then eluted with 5 mL of 0.1 mol/L melibiose in the corresponding wash buffer. Factor V in the eluate was purified and concentrated by immunoprecipitation. Human plasma in acid- citrate-dextrose was separated from whole blood 1 day after collec- tion, and was fractionated at 4°C on a IO-mL column of jacalin- agarose in 0.5 mol/L NaCl with 20 mmol/L HEPES (PH 7.4). Fractions of 4 mL were collected, and specific elution was effected with 20 mmol/L a-methyl galactopyranoside in the same buffer. Factor V activity was determined with a clotting assayz9 using a Precision fibrometer.
946 Blood, Vol 7 6 , No 5 (September 11, 1990: pp 946-952
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
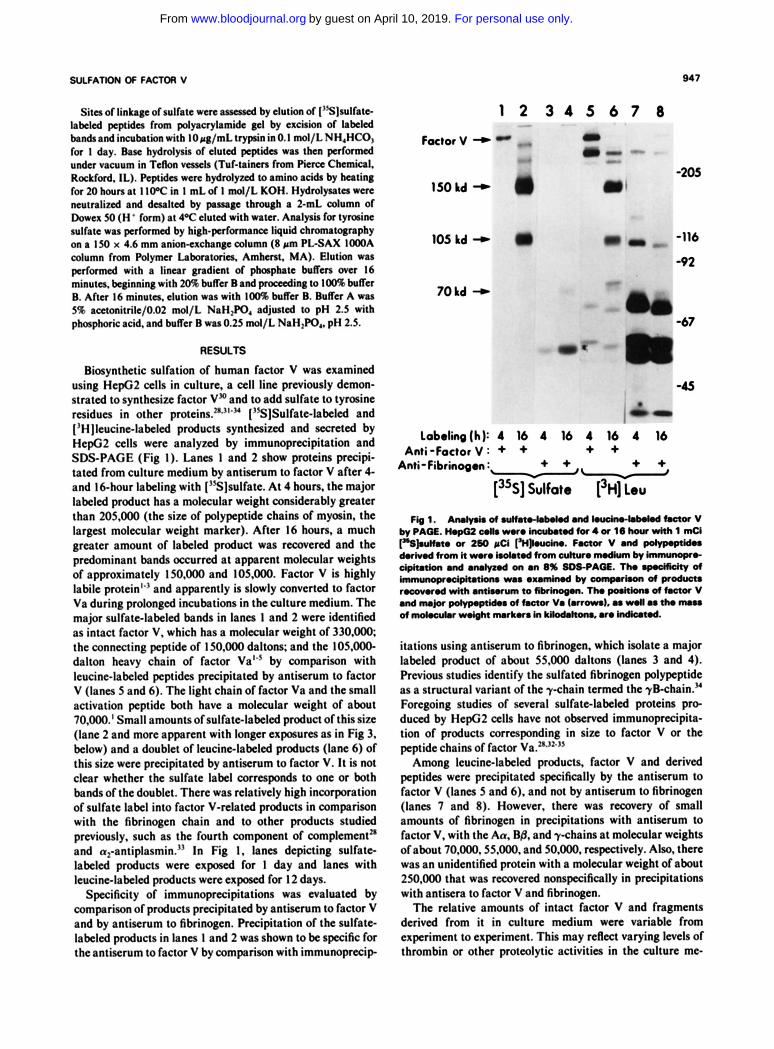
SULFATION OF FACTOR V 947
Site of linkage of sulfate were asscsed by elution of ["S]sulfata labeled peptides from polyacryhmide gel by excision of labeled bands and incubation with IOpglmL trypsin in 0.1 mol/LNH,HCO, for I day. Base hydrolysis of eluted peptides was then performed under vacuum in Teflon vessels (Tuf-tainm from Pierce Chemical, Rockford. IL). Peptides were hydrolyzed to amino acids by heating for 20 hours at I 10% in I mL of I mol/L KOH. H y d d p t ~ ~ were neutrali7xd and desalted by passage through a 2-mL column of Dowex 50 (H form) at 4OC eluted with water. Analysis for tyrosine sulfate was performed by high-performance liquid chromatography on a IS0 x 4.6 mm anion-exchange column (8 pm PL-SAX IOOOA wlumn from Polymer Laboratories, Amhmt. MA). Elution was performed with a linear gradient of phosphate buffers over 16 minutes. beginning with 20% buffer Band proceeding to IoOR, buffer B. After 16 minutes, elution was with 10096 buffer B. Buffer A was 5% acetonitrile/0.02 mol/L NaH,PO, adjusted to pH 2.5 with phosphoric acid. and buffer B was 0.25 mol/L NaH,PO,. pH 2.5.
RESULTS
Biosynthetic sulfation of human factor V was examined using HepG2 cells in culture. a cell line previously demon- strated to synthesize factor V" and to add sulfate to tyrosine residues in other pro~eins.~*'"" ['S]Sulfate-labeled and I'H ]leucinelabeled products synthesized and secreted by HepG2 cells were analyzed by immunoprecipitation and SDS-PAGE (Fig I). Lanes 1 and 2 show proteins precipi- tated from culture medium by antiserum to factor V after 4- and 16-hour labeling with ['%]sulfate. At 4 hours, the major labeled product has a molecular weight considerably greater than 205.000 (the size of polypeptide chains of myosin, the largest molecular weight marker). After 16 hours, a much greater amount of labeled product was recovered and the predominant bands occurred at apparent molecular weights of approximately 15O.OOO and 105,000. Factor V is highly labile protein'" and apparently is slowly converted to factor Va during prolonged incubations in the culture medium. The major sulfate-labeled bands in lanes 1 and 2 were identified as intact factor V. which has a molecular weight of 330.000, the connecting peptide of 150,000 daltons; and the 105.000- dalton heavy chain of factor Val" by comparison with leucinelabeled peptides precipitated by antiserum to factor V (lanes 5 and 6). The light chain of factor Va and the small activation peptide both have a molecular weight of about 70.000.' Small amounts of sulfatelabeled product of this si72 (lane 2 and more apparent with longer exposures as in Fig 3, below) and a doublet of leucinelabeled products (lane 6) of this size were precipitated by antiserum to factor V. It is not clear whether the sulfate label corresponds to one or both bands of the doublet. There was relatively high incorporation of sulfate label into factor V-related products in comparison with the fibrinogen chain and to other products studied previously. such as the fourth component of complement" and a,-antiplasmin." In Fig 1, lanes depicting sulfate labeled products were exposed for 1 day and lanes with leucine-labeled products were exposed for 12 days.
Specificity of immunoprecipitations was evaluated by comparison of products precipitated by antiserum to factor V and by antiserum to fibrinogen. Precipitation of the sulfate labeled products in lanes 1 and 2 was shown to be specific for the antiserum to factor V by comparison with immunoprecip
1 2 3 4 5 6 7 8
150 kd - a 105 kd - 0
70kd +
-205
-116
-92 - - *
-
-67 b
-4s
Labol i i (h) : 4 16 4 16 4 Anti-Factor V : + + +
FIg1. A n l y r k d ~ . n d k u O i r w - l . k k d ~ V by PA=. HopG2 & m r o lnwbmodtor4a 16 hwr wkh 1 mCi ["SJsuhto or 260 pCi "no. Foamr V and po)yp.pcid.. duivod from it -0 hd.tod from cu)nno nwdkm, immunopro- dpitat)on and arulymd on an 8% SDS-PAGE. Tho rp.dfidy of immunoprodpitatlons m a o x a m i d by comporhon of prod- r.covrod with antisorum to fibrinogen. Tho p d t b of factor V
of m0I.cut.r wdght nurkrs in ki)od.kom. aro indknod. and nu@ mid.. of factor V. ~arrom). as d l as th. "
itations using antiserum to fibrinogen. which isolate a major labeled product of about 55.000 daltons (lanes 3 and 4). Previous studies identify the sulfated fibrinogen polypeptide as a structural variant of the y-chain termed the ykhain ." Foregoing studies of several sulfatelabeled proteins pro- duced by HepG2 cells have not observed immunoprecipita- tion of products corresponding in size to factor V or the peptide chains of factor Va."."'"
Among leucinelabeled products, factor V and derived peptides were precipitated specifically by the antiserum to factor V (lanes 5 and 6). and not by antiserum to fibrinogen (lanes 7 and 8). However. there was recovery of small amounts of fibrinogen in precipitations with antiserum to factor V, with the Aa, B& and y-chains at molecular weights of about 70.000.55.000. and 50.000. respectively. Also, there was an unidentified protein with a molecular weight of about 250.000 that was recovered nonspecifically in precipitations with antisera to factor V and fibrinogen.
The relative amounts of intact factor V and fragments derived from it in culture medium were variable from experiment to experiment. This may reflect varying levels of thrombin or other proteolytic activities in the culture me-
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
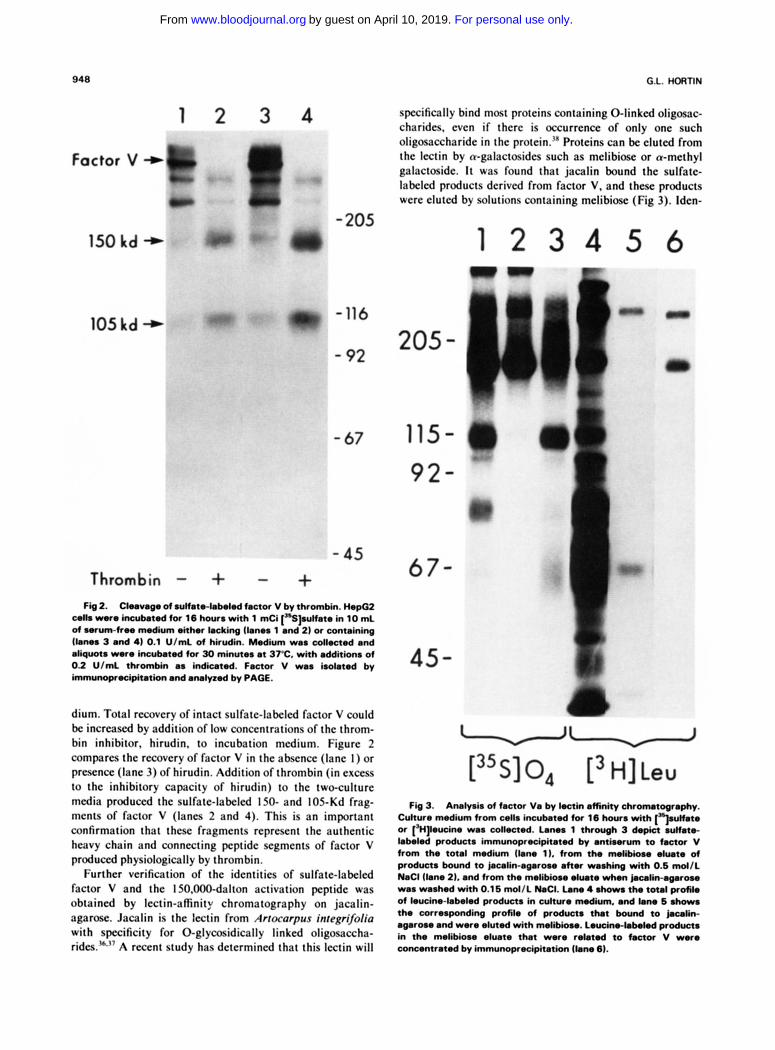
948
1
Factor V *
150 kd *
10s kd -
Thrombin -
G.L. HOFITlN
specifically bind most proteins containing 0-linked oligosac- charides. even if there is occurrence of only one such oligowxharide in the protein.'" Proteins can be eluted from the lectin by a-galactosides such as melibiose or a-methyl galactoside. It was found that jacalin bound the sulfate- labeled products derived from factor V, and these products were eluted by solutions containing melibiose (Fig 3). lden-
2 3 4
- 205 1 2 3 4 5 6
Fig 2. Clnvogo of.Unn.-bkkd factor V bythrombh. H.po2 colla wore hrmbeted tor 16 h a with 1 mCi ["Sptfate in 10 mL of wrumfrw medium &her lacking (bno% 1 and 2) or containing ~bma 3 and 4) 0.1 U/mL of hirudin. Medium waa cdlected and aliquot. were incubeted for 30 minutea at 37°C. with addition. of 0.2 U/mL thrombin a% indicated. Factor V waa isolated by immunoprecipitation and a ~ l v e d by PAGE.
dium. Total recovery of intact sulfate-labeled factor V could be increased by addition of low concentrations of the throm- bin inhibitor. hirudin. to incubation medium. Figure 2 compares the recovery of factor V in the absence (lane I ) or presence (lane 3) of hirudin. Addition of thrombin (in excess to the inhibitory capacity of hirudin) to the two-culture media produced the sulfate-labeled 150- and 105-Kd frag- ments of factor V (lanes 2 and 4). This is an important confirmation that these fragments represent the authentic heavy chain and connecting peptide segments of factor V produced physiologically by thrombin.
Further VCrificaliOn of the identities of sulfate-labeled factor V and the IS0.000-dalton activation peptide was obtained by lectin-afinity chromatography on jacalin- agarose. Jacalin is the lectin from Arrocclrpus integrvoliu with specificity for 0-glycosidically linked oligosaccha- rides."," A recent study has determined that this lectin will
45-
uu [35S] 0, [ 3 H] Leu
Fb3. AMI@% offactor Va b y k c t k , . m n i y v . Culture nwdium from cdla incubated for 16 hovr with [..lrulfate or ['H]kucine was collected. Lano% 1 through 3 depia sulfate- labeled producta immunoprecipitated by antiwum to factor V from the total medium (lane 1). from the mdibbw d w t e of products bound to jacalin-agsrou a h r waahing with 0.6 md/L NaCl (lane 21. and from the melibiom elute when j n c a l i n - ~ r o u was washed with 0.16 mol/L NaCI. Lane 4 .horn the total protik of leucine-labeled producta in culture medium. and lam 6 n h o w a the corresponding profile of products that bound to jacalin- agaroae and were duted with melibiou. Leucirw-bbdod product. in the melibiom eluate that were related to factor V were concentrated by immunoprocipkation (lane 6).
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
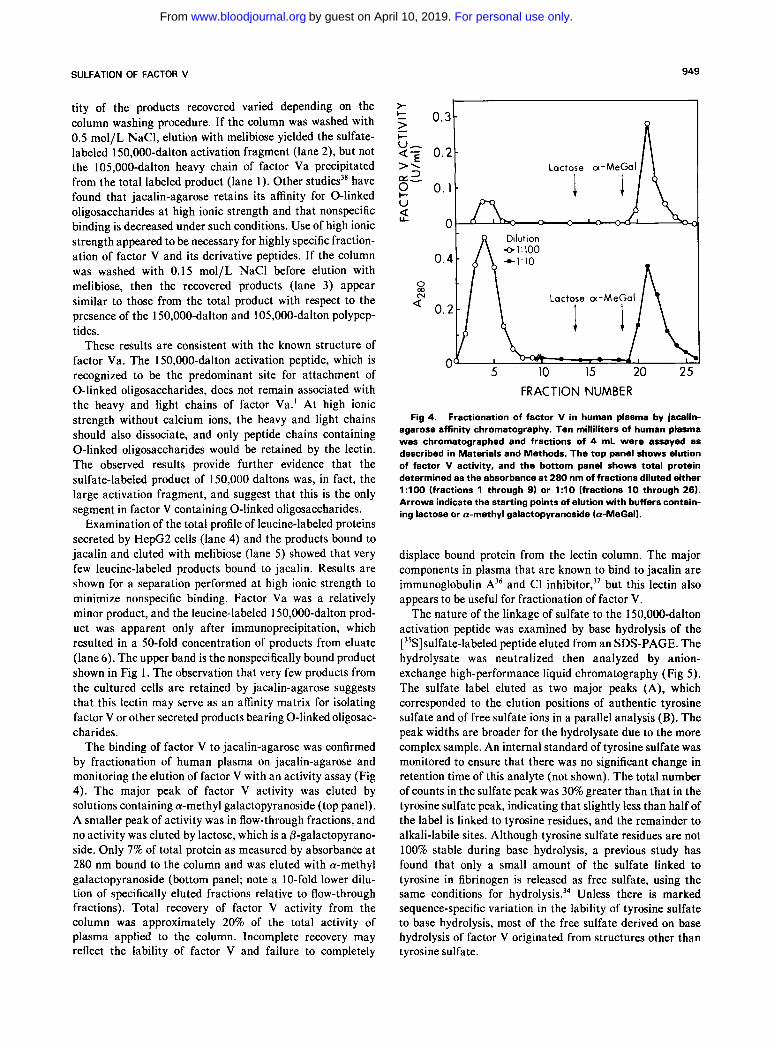
SULFATION OF FACTOR V 949
tity of the products recovered varied depending on the column washing procedure. If the column was washed with 0.5 mol/L NaCl, elution with melibiose yielded the sulfate- labeled 150,000-dalton activation fragment (lane 2), but not the 105,000-dalton heavy chain of factor Va precipitated from the total labeled product (lane 1). Other studies3’ have found that jacalin-agarose retains its affinity for 0-linked oligosaccharides a t high ionic strength and that nonspecific binding is decreased under such conditions. Use of high ionic strength appeared to be necessary for highly specific fraction- ation of factor V and its derivative peptides. If the column was washed with 0.15 mol/L NaCl before elution with melibiose, then the recovered products (lane 3) appear similar to those from the total product with respect to the presence of the 150.000-dalton and 105,000-dalton polypep- tides.
These results are consistent with the known structure of factor Va. The 150,000-dalton activation peptide, which is recognized to be the predominant site for attachment of 0-linked oligosaccharides, does not remain associated with the heavy and light chains of factor Va.’ At high ionic strength without calcium ions, the heavy and light chains should also dissociate, and only peptide chains containing 0-linked oligosaccharides would be retained by the lectin. The observed results provide further evidence that the sulfate-labeled product of 150,000 daltons was, in fact, the large activation fragment, and suggest that this is the only segment in factor V containing 0-linked oligosaccharides.
Examination of the total profile of leucine-labeled proteins secreted by HepG2 cells (lane 4) and the products bound to jacalin and eluted with melibiose (lane 5 ) showed that very few leucine-labeled products bound to jacalin. Results are shown for a separation performed at high ionic strength to minimize nonspecific binding. Factor Va was a relatively minor product, and the leucine-labeled 150,000-dalton prod- uct was apparent only after immunoprecipitation, which resulted in a 50-fold concentration of products from eluate (lane 6 ) . The upper band is the nonspecifically bound product shown in Fig 1. The observation that very few products from the cultured cells are retained by jacalin-agarose suggests that this lectin may serve as an affinity matrix for isolating factor V or other secreted products bearing 0-linked oligosac- charides.
The binding of factor V to jacalin-agarose was confirmed by fractionation of human plasma on jacalin-agarose and monitoring the elution of factor V with an activity assay (Fig 4). The major peak of factor V activity was eluted by solutions containing a-methyl galactopyranoside (top panel). A smaller peak of activity was in flow-through fractions, and no activity was eluted by lactose, which is a P-galactopyrano- side. Only 7% of total protein as measured by absorbance a t 280 nm bound to the column and was eluted with a-methyl galactopyranoside (bottom panel; note a 1 0-fold lower dilu- tion of specifically eluted fractions relative to flow-through fractions). Total recovery of factor V activity from the column was approximately 20% of the total activity of plasma applied to the column. Incomplete recovery may reflect the lability of factor V and failure to completely
> I
-7 Lactose a-Me
L
Dilution
+1:10
0.2
FRACTION NUMBER
Fig 4. Fractionation of factor V in human plasma by jacalin- agarose affinity chromatography. Ten milliliters of human plasma was chromatographed and fractions of 4 mL were assayed as described in Materials and Methods. The top panel shows elution of factor V activity, and the bottom panel shows total protein determined as the absorbance at 280 nm of fractions diluted either 1:lOO (fractions 1 through 9) or 1 : l O (fractions 10 through 26). Arrows indicate the starting points of elution with buffers contain- ing lactose or a-methyl galactopyranoside (a-MeGal).
displace bound protein from the lectin column. The major components in plasma that are known to bind to jacalin are immunoglobulin A36 and C1 inhibi t~r ,~’ but this lectin also appears to be useful for fractionation of factor V.
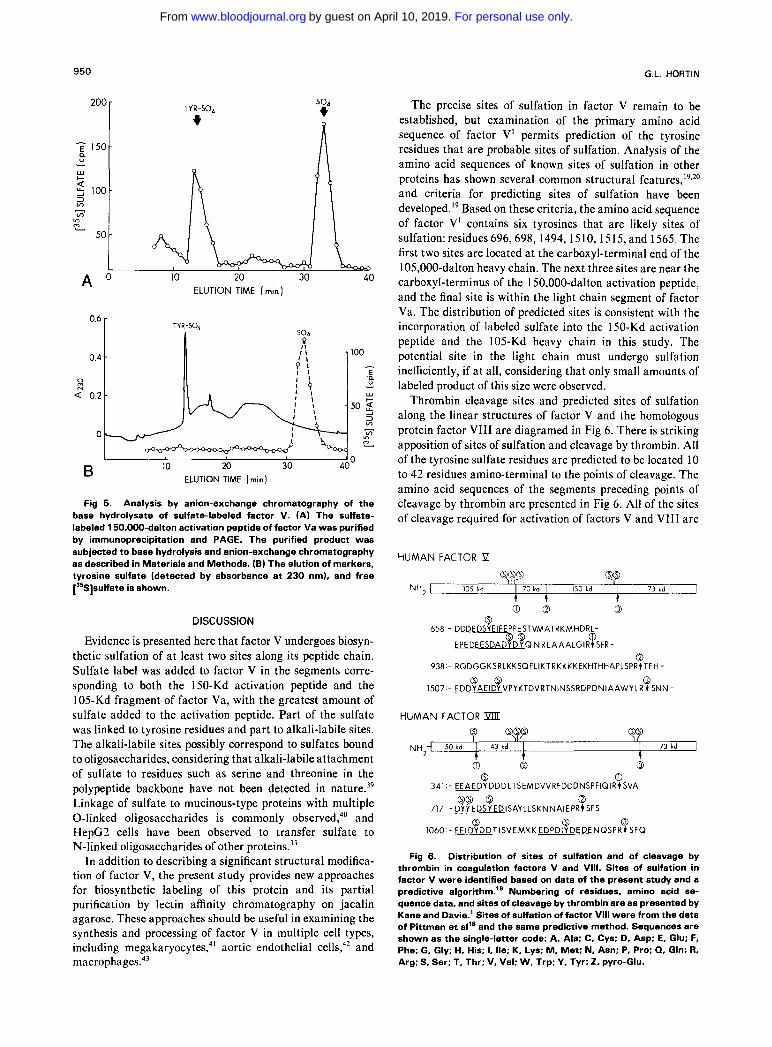
The nature of the linkage of sulfate to the 150,000-dalton activation peptide was examined by base hydrolysis of the [35S]sulfate-labeled peptide eluted from an SDS-PAGE. The hydrolysate was neutralized then analyzed by anion- exchange high-performance liquid chromatography (Fig 5) . The sulfate label eluted as two major peaks (A), which corresponded to the elution positions of authentic tyrosine sulfate and of free sulfate ions in a parallel analysis (B). The peak widths are broader for the hydrolysate due to the more complex sample. An internal standard of tyrosine sulfate was monitored to ensure that there was no significant change in retention time of this analyte (not shown). The total number of counts in the sulfate peak was 30% greater than that in the tyrosine sulfate peak, indicating that slightly less than half of the label is linked to tyrosine residues, and the remainder to alkali-labile sites. Although tyrosine sulfate residues are not 100% stable during base hydrolysis, a previous study has found that only a small amount of the sulfate linked to tyrosine in fibrinogen is released as free sulfate, using the same conditions for h y d r ~ l y s i s . ~ ~ Unless there is marked sequence-specific variation in the lability of tyrosine sulfate to base hydrolysis, most of the free sulfate derived on base hydrolysis of factor V originated from structures other than tyrosine sulfate.
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
950 G.L. HORTIN
200
y 150 0. U
w c - a 5 100 3 In
In - VI 2
50
10 20 30 40 ELUTION TIME (min )
A '
TYR-SO, + 200
150 0. U
w c - a 5 100 3 In
In - VI 2
50
10 20 30 40 ELUTION TIME (min )
A '
IO 20 30 40" ELUTION TIME (min)
B
Fig 5. Analysis by anion-exchange chromatography of the base hydrolysate of sulfate-labeled factor V. (A) The sulfate- labeled 150,000-dalton activation peptide of factor Va was purified by immunoprecipitation and PAGE. The purified product was subjected to base hydrolysis and anion-exchange chromatography as described in Materials and Methods. (B) The elution of markers, tyrosine sulfate (detected by absorbance at 230 nm), and free [35S]sulfate is shown.
DISCUSSION
Evidence is presented here that factor V undergoes biosyn- thetic sulfation of a t least two sites along its peptide chain. Sulfate label was added to factor V in the segments corre- sponding to both the 150-Kd activation peptide and the 105-Kd fragment of factor Va, with the greatest amount of sulfate added to the activation peptide. Part of the sulfate was linked to tyrosine residues and part to alkali-labile sites. The alkali-labile sites possibly correspond to sulfates bound to oligosaccharides, considering that alkali-labile attachment of sulfate to residues such as serine and threonine in the polypeptide backbone have not been detected in nature." Linkage of sulfate to mucinous-type proteins with multiple 0-linked oligosaccharides is commonly ob~erved,~' and HepC2 cells have been observed to transfer sulfate to N-linked oligosaccharides of other protein^.^'
In addition to describing a significant structural modifica- tion of factor V, the present study provides new approaches for biosynthetic labeling of this protein and its partial purification by lectin affinity chromatography on jacalin agarose. These approaches should be useful in examining the synthesis and processing of factor V in multiple cell types, including megakaryocyte^,^' aortic endothelial cells,42 and macrophage^.^^
The precise sites of sulfation in factor V remain to be established, but examination of the primary amino acid sequence of factor V' permits prediction of the tyrosine residues that are probable sites of sulfation. Analysis of the amino acid sequences of known sites of sulfation in other proteins has shown several common structural features,'9J0 and criteria for predicting sites of sulfation have been developed.'' Based on these criteria, the amino acid sequence of factor V' contains six tyrosines that are likely sites of sulfation: residues 696,698, 1494, 1510, 1515, and 1565. The first two sites are located at the carboxyl-terminal end of the 105,000-dalton heavy chain. The next three sites are near the carboxyl-terminus of the 150,000-dalton activation peptide, and the final site is within the light chain segment of factor Va. The distribution of predicted sites is consistent with the incorporation of labeled sulfate into the 150-Kd activation peptide and the 105-Kd heavy chain in this study. The potential site in the light chain must undergo sulfation inefficiently, if a t all, considering that only small amounts of labeled product of this size were observed.
Thrombin cleavage sites and predicted sites of sulfation along the linear structures of factor V and the homologous protein factor VI11 are diagramed in Fig 6. There is striking apposition of sites of sulfation and cleavage by thrombin. All of the tyrosine sulfate residues are predicted to be located I O to 42 residues amino-terminal to the points of cleavage. The amino acid sequences of the segments preceding points of cleavage by thrombin are presented in Fig 6. All of the sites of cleavage required for activation of factors V and VI11 are
HUMAN FACTOR Il
090 8 8
NH2-1 105 kd I 70 kd I 150 kd I 73 id I 4 4
0 658 - DDDEDSYEIFEPPESTVMATRKMHDRL-
S @ 0 E PE DE ESDADVD Y Q N R LA A A L G I R t SFR -
a 938 - RQDGGKSRLKKSQFLlKTRKKKKEKHTHHAPLSPRtTFH -
0 0 0 1507 - EDDYAEIDY VPYKTDVRTNINSSRDPDNIAAWY LR t SNN-
HUMAN FACTOR Pm Q Q S S s s
'I I 73 kd I 9' 1 43 kd 1 t
0 Q 0 0 0
341 - EEAEDYDDDLTSEMDVVRFDDDNSPFlQlRt SVA
QO 0 0 717 - DYY EDSY EDISAYLLSKNNAIEPR~SFS
0 0 0 1060 - EEIDYDDTISVEMKK EDPDlYDEDENQSPRt SFQ
Fig 6. Distribution of sites of sulfation and of cleavage by thrombin in coagulation factors V and VIII. Sites of sulfation in factor V were identified based on data of the present study and a predictive algorithm.'' Numbering of residues, amino acid se- quence data, and sites of cleavage by thrombin are as presented by Kane and Davie.' Sites of sulfation of factor Vlll were from the data of Pittman et all' and the same predictive method. Sequences are shown as the single-letter code: A, Ala; C, Cys; D, Asp: E, Glu: F. Phe: G, Gly; H. His; 1, Ile; K, Lys; M, Met; N, Asn; P, Pro: Q, Gln; R. Arg: S, Ser; 1, Thr; V, Val; W, Trp: Y. Tyr; 2. pyro-Glu.
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

SULFATION OF FACTOR V 95 1
preceded by clusters of acidic amino acid residues (under- lined) embodying predicted tyrosine sulfate residues. Cleav- age a t the one site, following Arg,,,, in factor V, that is not preceded by a cluster of acidic amino acids apparently is not essential for activation of factor V to factor Va.1*44
Functional significance of the tyrosine sulfate residues and the surrounding clusters of acidic amino acids is suggested by identification of an anion-binding site on thrombin’s surface that participates in interactions with fibrinogen and hir~din.~’ .~’ This functional element, termed the “anion- binding exosite,” is located a significant distance from the enzyme’s catalytic ~ite:~.~’ such that a minimal-length pep- tide segment of approximately 10 amino acid residues would be required to span the intervening space. Interactions of the anion-binding exosite with peptide ligands have been best defined with regard to a carboxyl-terminal segment of hirudin. The final 12 residues of hirudin contain multiple acidic residues, which contribute to binding to the anion- binding exosite. Sulfation of the tyrosine residue near the carboxyl-terminus of hirudin increases its affinity for throm- bin 10-f0ld,’~ and sulfation has similar effects on the affinity for thrombin of synthetic peptides corresponding to carboxyl- terminal segments of h i r ~ d i n . ~ ” ~ ’ There has been extensive study of the relationship of peptide structure to this binding site on and it is observed that considerable sequence variation is permitted with retention of affinity for thrombin. Recent work has provided evidence that the anion-binding exosite also binds to an acidic segment in the plasma proteinase inhibitor, heparin cofactor 11, that con- tains two tyrosine sulfate residue^.'^.^^ The acidic segment of heparin cofactor I1 does not have a high degree of sequence homology with the carboxyl-terminus of hirudin. Thus, the thrombin’s anion-binding exosite may be able to interact with a variety of polyanionic peptide segments, such as those that occur in factors V and VI11 preceding points of cleavage by thrombin. A surprising number of proteins that interact with thrombin are recognized to contain acidic regions with tyrosine sulfate residues, including coagulation factors V and
VIII,” hirudin,24 heparin cofactor II,3z fibrinogens (within B fibrinopeptides) of many animal specie^,^' human fibrinogen (within yB chain),34 and vitronectin.” Thus, thrombin’s interactions with many proteins may be directed, a t least in part, by binding to these sites.
Location of ligands for thrombin’s anion-binding exosite within substrates and inhibitors of thrombin offers the potential for substantial increases in the efficiency and specificity of interactions with thrombin. The local concentra- tion of thrombin would be increased and the enzyme’s catalytic site may be aligned with appropriate points of cleavage or reactive sites of inhibitors. A more unexpected consequence of interaction of the anion-binding exosite with peptide ligands is that there is a substantial conformational change of the thrombin molecule.25 The conformational change produces functional changes a t the catalytic site that are reflected by altered kinetic parameters for chromogenic substrate^^',^^ and increased affinity for the amino-terminal segment of hirudin.’* These results suggest that conforma- tional changes of thrombin induced by interactions with polyanionic peptide segments of a natural substrate or inhibitor may produce in situ increases in the enzyme’s catalytic efficiency.
The proposed role for polyanionic sequences, and particu- larly tyrosine sulfate residues, in the interactions of factors V and VI11 and other proteins with thrombin targets a number of sites for further investigation by site-directed mutagenesis, by use of inhibitors of sulfation, and by production of model synthetic peptides. Understanding these interactions will be important for determining the molecular basis for thrombin’s specificity and for delineating the mechanism of a new family of anticoagulants based on the carboxyl-terminal sequence of
ACKNOWLEDGMENT
I thank James Graham and Barbara Benutto for technical assistance.
REFERENCES
1. Kane WH, Davie EW: Blood coagulation factors V and VIII: Structural and functional similarities and their relationship to hemorrhagic and thrombotic disorders. Blood 71 539, 1988
2. Dahlback B: Human coagulation factor V purification and thrombin-catalyzed activation. J Clin Invest 66583, 1980
3. Suzuki K, Dahlback B, Stenflo J: Thrombin-catalyzed activa- tion of human coagulation factor V. J Biol Chem 257:6556, 1982
4. Nesheim ME, Foster WB, Mann KG: Characterization of factor V intermediates. J Biol Chem 259:3187, 1984
5. Dahlback B: A new model for coagulation factor V suggesting a unique mechanism of activation. Scand J Clin Lab Invest 148:47, 1988 (suppl 191)
6. Miletich JP, Jackson CM, Majerus PW: Properties of the factor Xa binding site on human platelets. J Biol Chem 253:6908, 1978
7. Krishnaswamy S, Church WR, Nesheim ME, Mann KG: Activation of human prothrombin by human prothrombinase: Influ- ence of factor Va on the reaction mechanism. J Biol Chem 262:3291, 1987
8. Tracy PB, Mann KG: Abnormal formation of the prothrombi-
nase complex: Factor V deficiency and related disorders. Human Pathol 18:162, 1987
9. Walker FJ, Sexton PW, Esmon CT: The inhibition of blood coagulation by activated protein C through the selective inactivation of activated factor V. Biochim Biophys Acta 571:333,1979
10. Kane WH, Davie EW: Cloning of a cDNA coding for human factor V, a coagulation factor homologous to factor VI11 and ceruloplasmin. Proc Natl Acad Sci USA 83:6800, 1986
1 1 . Jenny RJ, Pittman DD, Toole JJ, Kriz RW, Aldape RA, Hewick RM, Kaufman RJ, Mann KG: Complete cDNA and derived amino acid sequence of human factor V. Proc Natl Acad Sci USA 84:4846, 1987
12. Toole JJ, Knopf JL, Wozney JM, Sulzman LA, Buecker JL, Pittman DD, Kaufman RJ, Brown E, Shoemaker C, Orr EC, Amphlett GW, Foster WB, Coe ML, Knutson GJ, Fass DN, Hewick RM: Molecular cloning of a cDNA encoding human antihemophilic factor. Nature 312:342, 1984
13. Vehar GA, Keyt B, Eaton D, Rodriguez H, OBrien DPO, Rotblat F, Oppermann H, Keck R, Wood WI, Harkins RN, Tuddenham EGD, Lawn RM, Capon DJ: Structure of human factor VIII. Nature 312:337, 1984
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom

952 G.L. HORTIN
14. Seeler RA: Parahemophilia. Factor V deficiency. Med Clin North Am 56:119, 1972
15. Chiu HC, Whitaker E, Colman RW: Heterogeneity of human factor V deficiency. Evidence for the existence of antigen-positive variants. J Clin Invest 72:493, 1983
16. Kane WH, Majerus PW: Purification and characterization of human coagulation factor V. J Biol Chem 256:1002, 1981
17. Bruin T, Sturk A, Ten Cate JW, Cath M: The function of the human factor V carbohydrate moiety in blood coagulation. Eur J Biochem 170:305, 1987
18. Pittman DD, Wasley LC, Murray BL, Wang JH, Kaufman RJ: Analysis of structural requirements for factor VI11 function using site directed mutagenesis. Thromb Haemost 58:344, 1987 (abstr)
19. Hortin G, Folz R, Gordon JI, Strauss AW: Characterization of sites of tyrosine sulfation in proteins and criteria for predicting their occurrence. Biochem Biophys Res Commun 141:326,1986
20. Huttner WB: Tyrosine sulfation and the secretory pathway. Ann Rev Physiol50:363, 1988
21. Hortin GL, Farries TC, Graham JP, Atkinson JP: Sulfation of tyrosine residues increases activity of the fourth component of complement. Proc Natl Acad Sci USA 86:1338, 1989
22. Suiko M, Liu M-C: Change in binding affinities of 3Y1 secreted fibronectin upon desulfation of tyrosine-0-sulfate. Biochem Biophys Res Commun 1541094,1988
23. Hortin GL, Tollefsen DM, Benutto BM: Antithrombin activ- ity of a peptide corresponding to residue 54-75 of heparin cofactor 11. J Biol Chem 264:13979,1989
24. Braun PJ, Dennis S, Hofsteenge J, Stone SR: Use of site-directed mutagenesis to investigate the basis for the specificity of hirudin. Biochemistry 27:6517, 1988
25. Mao SJT, Yates MT, Owen TJ, Krstenansky JL: Interaction of hirudin with thrombin: Identification of a minimal binding domain of hirudin that inhibits clotting activity. Biochemistry 27:8 170, 1988
26. Maraganore JM, Chao B, Joseph ML, Jablonski J, Ramachan- dran KL: Anticoagulant activity of synthetic hirudin peptides. J Biol Chem 264:8692,1989
27. Knowles BB, Howe CC, Aden D P Human hepatocellular carcinoma cell lines secrete the major plasma proteins and hepatitis B surface antigen. Science 209:497, 1980
28. Hortin G, Sims H, Strauss AW: Identification of the site of sulfation of the fourth component of human complement. J Biol Chem 261:1786,1986
29. Ware AG, Murphy RC, Seegers WH: The function of Ac-globulin. Science 106:6 18, 1947
30. Wilson DB, Salem HH, Mruk JS, Maruyama I, Majerus PW: Biosynthesis of coagulation factor V by a human hepatocellular carcinoma cell line. J Clin Invest 73:654, 1984
3 1. Liu M-C, Yu S, Sy J, Redman CM, Lipmann F Tyrosine sulfation of proteins from the human hepatoma cell line HepG2. Proc Natl Acad Sci USA 82:7160, 1985
32. Hortin G, Tollefsen DM, Strauss AW: Identification of two sites of sulfation of human heparin cofactor 11. J Biol Chem 261:15827,1986
33. Hortin G, Fok KF, Toren PC, Strauss AW: Sulfation of a tyrosine residue in the plasmin-binding domain of a,-antiplasmin. J Biol Chem 262:3082, 1987
34. Hortin GL: Sulfation of a gamma-chain variant of human fibrinogen. Biochem Int 19:1355, 1989
35. Hortin G, Green ED, Baenziger JU, Strauss AW: Sulphation of proteins secreted by a human hepatoma-derived cell line: Sul- phation of N-linked oligosaccharides on a,HS-glycoprotein. Bio- chem J 235:407, 1986
36. Skea DL, Christopoulous P, Plaut AG, Underdown BJ: Studies on the specificity of the IgA-binding lectin, jacalin. Mol Immunol25:1, 1988
37. Pilatte Y, Hammer CH, Frank MM, Fries L F A new simplified procedure for C1 inhibitor purification: A novel use for jacalin-agarose. J Immunol Methods 120:37, 1989
38. Hortin GL, Trimpe BL: Lectin affinity chromatography of proteins bearing 0-linked oligosaccharides: Applications of jacalin- agarose. Anal Biochem 1990 (in press)
39. Huttner WB: Determination and Occurrence of tyrosine 0-sulfate in proteins. Methods Enzymol 107:200, 1984
40. Yosizawa Y: Sulphated glycoproteins, in Gottschalk A (ed): Glycoproteins. Amsterdam, The Netherlands, Elsevier, 1972, p 1000
41. Chiu HC, Schick PK, Colman RW: Biosynthesis of factor V in isolated guinea pig megakaryocytes. J Clin Invest 75:339, 1985
42. Cerveny TJ, Fass DN, Mann KG: Synthesis of coagulation factor V by cultured aortic endothelium. Blood 63:1467, 1984
43. Osterud B, Bogwald J, Lindahl U, Seljelid R: Production of blood coagulation factor V and tissue thromboplastin by macroph- ages in vitro. FEBS Lett 127:154, 1981
44. Monkovic DD, Tracy PB: Activation of human factor V by factor Xa and thrombin. Biochemistry 29:1118, 1990
45. Fenton JW 11: Thrombin interactions with hirudin. Semin Thromb Hemost 15:265, 1989
46. Chang J-Y: The hirudin-binding site of human a-thrombin. J Biol Chem 264:7141, 1989
47. Bode W, Mayr I, Baumann U, Huber R, Stone SR, Hof- steenge J: The refined A1.9 A crystal structure of human cu-throm- bin: Interaction with D-Phe-Pro-Arg chloromethylketone and signif- icance of the Tyr-Pro-Pro-Trp insertion segment. EMBO J 8:3467, 1989
48. Hortin GL, Benutto BM, Trimpe BL: Synthetic peptides as molecular probes of thrombin’s interactions with substrates and inhibitors. J Cell Biochem 14C:243, 1990 (suppl)
49. Ragg H, Ulshofer T, Gerewitz J: On the activation of human leuserpin-2, a thrombin inhibitor, by glycosaminoglycans. J Biol Chem 265521 I , 1990 50. Dayhoff MO: Atlas of Protein Sequence and Structure. vol5.
Washington, DC, National Biomedical Research Foundation, 1972, p D212
51. Jenne D, Hille A, Stanley KK, Huttner WB: Sulfation of two tyrosine residues in human complement S-protein (vitronectin). Eur J Biochem 185:391, 1989
52. Dennis S, Wallace A, Hofsteenge J, Stone SR: Use of fragments of hirudin to investigate thrombin-hirudin interaction. Eur J Biochem 188:61,1990
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom
1990 76: 946-952
GL Hortin Sulfation of tyrosine residues in coagulation factor V
http://www.bloodjournal.org/content/76/5/946.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 10, 2019. by guest www.bloodjournal.orgFrom





























