Embed Size (px)
Citation preview

5
STUDIUL SIMBIOZELOR MICORIZIENE ÎN UNELE ECOSISTEME DE PAJIŞTI DIN ZONA MUNŢILOR APUSENI –
rezumat teză de doctorat
THE STUDY OF MYCORRHIZAL SYMBIOSIS IN SOME GRASSLAND ECOSYSTEMS FROM APUSENI MOUNTAINS -
phd. thesis abstract
Vlad Stoian USAMV Cluj-Napoca
Teza de doctorat a fost realizată sub îndrumarea: Prof. univ. dr. Viorel FLORIAN
Prof. univ. dr. Carmen PUIA
INTRODUCERE
Pe parcursul perioadelor geologice, fungii micorizieni au evoluat odată cu plantele, fiind prezente ca parteneri simbiotici sistemele radiculare ale acestora. Până în anul 1885, care marchează descoperirea ştiinţifică a simbiozelor micoriziene de către cercetătorul germen B. Frank, acest tip de tip de asociaţii a trecut prin diferite stadii de transformare reuşind să se specializeze pe anumite nişe ecologice şi definitivându-şi ciclurile de viaţă în funcţie de cerinţele şi permisivitatea plantelor gazdă.
Studii mai recente (Remy et al., 1994) certifică existenţa fungilor micorizieni încă din Devonianul timpuriu, în urmă cu aproximativ 400 milioane de ani, fosilele plantelor de Aglaophyton major prezentând urme de arbusculi care sunt considerate structuri de transfer definitorii pentru tipul endomicorizian vezicular-arbuscular.
Aceste aspecte, completate cu o serie de studii cu privire la rolul şi beneficiile existenţei simbiozelor micoriziene au arătat influenţa majoră pe care o au acestea asupra creşterii şi dezvoltării plantelor, cât şi caracterul de reglare asupra proceselor interne din ecosistemele în care îşi duc existenţa.
Fungii vezicular-arbusculari au o contribuţie majoră la absobţia elementelor nutritive (în special fosfor) din sol cu ajutorul hifelor extraradiculare – mai ales din acele porţiuni de sol la care planta nu a avut acces. Hifele ciupercii acţionează similar cu perişorii absorbanţi de pe rădăcina plantei; deşi în urma comparării diametrului perişorilor absorbanţi (5-20 µm) cu cel al hifelor ciupercii (3-7 µm) perişorii

6
absorbanţi ar avea câştig de cauză, prin compararea lungimii şi densităţii hifelor ciupercii cu cea a perişorilor absorbanţi – ciuperca iese în avantaj, depăşind posibilităţile de extindere ale plantei de 10 până la 100 ori mai mult.
Hifele ciupercii au un important rol în ciclul nutrienţilor, în acest mod prevenind apariţia pierderilor din ecosistem, în special în perioada în care rădăcinile sunt inactive. Fungii micorizieni asigură transportul şi transferul carbonului primit de la plantele gazdă către alte microorganisme implicate în procesele biologice (procese de nutriţie, procese descompunere), prin acest fenomen asigurându-se o stabilitate în compoziţia microflorei din sol (Van der Heijden et al., 1998).
Ecosistemele mature sunt caracterizate de o mişcare permanentă şi ciclică a elementelor între partea biotică şi abiotică din care sunt compuse. Micorizele au rolul de a regla compoziţia şi funcţionarea comunităţilor de plante prin alocarea resurselor de hrană şi influenţarea creşterii plantelor cu care interacţionează.
În acest context şi pe baza identificării unui microclimat neperturbat în satul Gheţari (judeţul Alba), am propus un proiect de specializare prin doctorat intitulat “Studiul simbiozelor micoriziene în unele ecosisteme de pajişti din zona Munţilor Apuseni”. Obiectivele principale ale acestui proiect au fost studierea şi analizarea comportamentului fungilor micorizieni care colonizează în mod natural plantele din zona de experimentare sub aspectul influenţei unor factori tehnologici, respectiv identificarea nivelului perturbativ al acestor factori asupra parametrilor colonizării.
Pentru înţelegerea şi sprijinul acordat pe parcursul perioadei de doctorantură şi în faza de finalizare a tezei de doctorat îi aduc alese mulţumiri conducătorului de doctorat – profesor universitar doctor Carmen Emilia PUIA, care a preluat sarcina dificilă de mă îndruma după trecerea în nefiinţă a profesorului universitar doctor Viorel FLORIAN.
De asemenea le mulţumesc colegilor de la disciplina de Fitopatologie pentru ajutorul acordat pe tot parcursul perioadei de doctorantură, atât în ceea ce priveşte aspectele practice din teren şi laborator cât şi cele teoretice.
Mulţumiri deosebite le aduc pe această cale cadrelor didactice de la disciplina de Cultura pajiştilor şi a plantelor furajere care mi-au fost alături atât din punct de vedere material cât şi în ceea ce priveşte sfaturile bine intenţionate şi cu un mare grad de profesionalism.
Le adresez mulţumiri şi membrilor comisiei, pentru efortul de a analiza această lucrare şi de a întocmi referatele de apreciere şi nu în

7
ultimul rând celor care, în diverse modalităţi, mi-au oferit ajutorul la efectuarea experimentărilor şi redactarea lucrării.
Capitolul 1
CONSIDERAŢII GENERALE PRIVIND SIMBIOZELE MICORIZIENE
1.1. ISTORIC, ORIGINE ŞI EVOLUŢIE
Termenul de micoriză a fost folosit, pentru prima dată, de către
cercetătorul german B. FRANK, specialist în domeniul protecţiei speciilor forestiere, în anul 1885, care afirma că: „Anumite specii de pomi nu se hrănesc în mod regulat din sol, dar orice porţiune a întregului lor sistem radicular se află în simbioză cu un miceliu de ciupercă care serveşte ca mamă vitregă şi ia plata pentru hrănirea pomului cu hrană din sol” (B. FRANK, 1885, citat de www.biologie.uni-hamburg.de).
Fungii micorizieni sunt simbionţi în rădăcinile majorităţii plantelor evoluate. Aceste asociaţii variază ca structură şi funcţie, dar cele mai comune interacţiuni sunt de tip vezicular-arbuscular. Se estimează că procentul de plante terestre care formează acest tip de asociaţii depăşeşte 80% - aici fiind incluse şi multe specii cultivate (Harley and Harley, 1987).
1.2. BENEFICII ALE SIMBIOZELOR MICORIZIENE
1.2.1. Beneficii pentru plantă
La majoritatea tipurilor de micorize mişcarea carbohidraţilor,
produşi în timpul fotosintezei, se face de la planta gazdă (partenerul autotrof) la ciuperca simbiontă (partenerul heterotrof). Contribuţia fungilor vezicular-arbusculari la asimilarea nutrienţilor este absorbţia elementelor nutritive (în special fosfor) din sol, cu ajutorul hifelor extraradiculare – în special din acele porţiuni de sol la care planta nu a avut acces.
Azotul este un nemetal, necesar în formarea aminoacizilor, purinelor şi pirimidinelor, parte integrantă a multor enzime din sistemul plantei iar în mod indirect în formarea proteinelor şi a sintezei acizilor nucleici. Carenţa de azot are efecte asupra ritmului de creştere al plantelor, respectiv asupra clorofilei din frunze.

8
Studiile desfăşurate până în acest moment au confirmat faptul că plantele micorize posedă o mai mare rezistenţă în condiţii de stres hidric. Explicaţia acestui fenomen este extinderea mare pe care o are miceliul ciupercii în sol iar absorbţia apei se face similar cu absorbţia elementelor nutritive (Mohammadi Goltapeh et al., 2008).
1.2.2. Beneficii pentru ecosistem
Hifele ciupercii au un important rol în ciclul nutrienţilor, în acest mod prevenind apariţia pierderilor din ecosistem, în special în perioada în care rădăcinile sunt inactive.
Fungii micorizieni asigură transportul şi transferul carbonului primit de la plantele gazdă către alte microorganisme implicate în procesele biologice (procese de nutriţie, procese descompunere), prin acest fenomen asigurându-se o stabilitate în compoziţia microflorei din sol (Van der Heijden et al., 1998).
1.3. FACTORI CARE INFLUENŢEAZĂ FORMAREA
SIMBIOZELOR MICORIZIENE
Influenţa factorilor genetici ai gazdei este foarte importantă pentru iniţierea colonizării cu fungi vezicular-arbusculari iar alături de aceşti factori, structura şi morfologia rădăcinii are un rol important în micorizare. Un alt aspect este microflora din jurul rădăcinilor şi din sol, aceasta influenţând de asemenea formarea coloniilor de micorize.
Lumina. Sursa de energie a fungului simbiont se află în plantă şi depinde în mod direct de modul în care aceasta îşi desfăşoară procesul de fotosinteză şi de abilitatea ei de a transloca produsele fotosintezei către rădăcină (Varma, 2008).
Temperatura. Din punct de vedere al proceselor de germinare al sporilor, penetrarea rădăcinilor de către hife şi proliferarea acestora în interiorul celulelor corticale, temperatura poate fi un factor cu efect limitativ (Gavito et al., 2005).
Capitolul 2
TIPURI MORFOLOGICE DE MICORIZE
În funcţie de modul de dezvoltare la nivel radicular micorizele se
împart în două categorii: ectomicorize şi endomicorize.

9
2.1. ECTOMICORIZE
Definiţie. Ectomicoriza este un tip de asociaţie micoriziană la care hifele ciupercii nu penetrează celulele rădăcinii dar acoperă rădăcina şi se dezvoltă între celule acesteia (Peterson, 2004).
2.2. ENDOMICORIZE
Endomicorizele nu formează o manta în jurul rădăcinilor şi nu au
în general o fază extraradiculară bine dezvoltată. Nu formează sporocarpi, dezvoltând de obicei o serie de spori care rămân în sol în jurul rădăcinilor. Faza intraradiculară constă în dezvoltarea hifelor printre celulele corticale pe care le şi penetrează în unele cazuri.
Principalele tipuri de endomicorize sunt cele orhidacee, ericoide, arbutoide, monotropoide şi vezicular-arbusculare.
Capitolul 3 STADIUL ACTUAL AL CUNOAŞTERII PRIVIND SIMBIOZELE
ENDOMICORIZIENE VEZICULAR-ARBUSCULARE
3.1. BIOLOGIE ŞI DEZVOLTARE GENERALĂ
Pe măsură ce sporii germinează, creşterea hifelor implicând unele
diviziuni nucleare, sunt consumate rezerve de carbohidraţi şi lipide şi se produc cantităţi limitate de ramificaţii – miceliu cenocitic capabil de anastomoză. Dacă gazda lipseşte sau plantele învecinate nu sunt compatibile pentru colonizare, creşterea stagnează şi în cele din urmă încetează ca urmare a lipsei moleculelor semnal din rădăcini, care au rolul de a stimula ramificarea hifelor (Giovannetti et al.,1993; Tamasloukht et al., 2003; Akiyama et al, 2006). Stoparea creşterii poate fi programată prin controlarea reacţiei citoplasmatice şi nucleare, respectiv a producerii septelor, fapt care permite sporilor şi miceliului asociat să îşi păstreze viabilitatea şi capacitatea de regenerare şi colonizare un timp îndelungat. După stabilizarea simbiozei cu planta gazdă, creşterea miceliului are loc atât în interiorul rădăcinii cât şi în sol, etapa următoare fiind formarea de noi spori polinucleaţi la capătul hifelor. Fungii vezicular-arbusculari sunt incapabili să îşi completeze ciclul de viaţă fără existenţa unei gazde cu care să stabilească o relaţie

10
simbiotică, de aceea sunt greu de cultivat în lipsa gazdelor (Hildebrandt et al., 2006).
Miceliul provenit dintr-un singur spor nu rămâne independent. Anastomoza dintre hife, conform studiilor recente (Giovannetti et al, 1999; de la Providencia et al., 2005), are loc chiar dacă acestea provin din spori sau rădăcini colonizate, rezultând o continuitate citoplasmatică şi o migrare nucleară. Frecvenţa anastomozei şi în consecinţă schimbul potenţialului genetic este mai mare în cadrul unui miceliu provenit dintr-un singur spor decât între miceliile provenite de la spori diferiţi în cadrul aceluiaşi izolat. Studierea anastomozei are implicaţii semnificative în înţelegerea geneticii fungilor vezicular-arbusculari precum şi în întemeierea şi menţinerea reţelei miceliene implicate în colonizarea plantelor şi transferul de elemente nutritive (Jakobsen, 2004).
3.3. PLANTE NEMICORIZANTE
Presupusele plante non-micorize fac parte din grupurile de muşchi, ferigi şi multe familii de angiosperme, distanţate din punct de vedere al relaţiilor una de alta. În plus, unele familii au în componenţă atât membri capabili de asocieri de tip micorizian cât şi membri non-micorizieni.
Evoluţia independentă a plantelor cu filogenie înrudită, respectiv pierderea statutului micorizian, se bazează pe transformările celulare şi fiziologice suferite în urma dezvoltării în condiţii de mediu diferite. Familii ca Brassicaceae, Chenopodiaceae, Caryophyllaceae, Polygonaceae, Juncaceae şi Proteaceae sunt recunoscute ca având membrii care nu formează asociaţii micoriziene (Smith and Read, 2008).
3.4. PLANTE MICORIZANTE
Gama de plante care pot fi parteneri pentru fungii vezicular-arbusculari este extrem de largă, de aceea Gerdeman (1968) a afirmat “simbioza este omniprezentă încât este mai uşor de alcătuit lista cu familiile de plante la care nu se ştie dacă apare decât alcătuirea unei liste la care a fost găsită”. Membrii celor mai multe familii de angiosperme şi gimnosperme împreună cu formele sporofite ale ferigilor şi muşchilor formează micorize arbusculare. Există excepţii deoarece au fost descoperite dovezi ale colonizării în rădăcinile unora dintre speciile din cadrul acestor familii, colonizarea fiind reprezentată atât de arbusculi şi vezicule cât şi de hife dispersate şi vezicule. Fiziologia acestor

11
interacţiuni nu a fost explorată complet dar se poate emite ipoteza că aceste simbioze pot varia de la utilizarea carbonului organic în mod parazit până la interacţiuni neutre până la simbioze mutualistice tipice (Koltai and Kapulnik, 2010).
Wang şi Qiu (2006) au urmărit 3617 specii de plante din 263 de familii pentru a identifica existenţa colonizării indiferent de tipul acesteia. Studiul a oferit dovezile necesare pentru următoarea afirmaţie: dintre plantele terestre, luate ca un întreg, 80% din specii şi 92% din familii au capacitatea potenţială de a forma cel puţin un tip de asociaţie micoriziană. Pentru angiosperme, cifrele sunt 84% şi 94%. Micorizele vezicular-arbusculare se prezintă ca un tip ancestral de simbioză colonizând marea majoritate a speciilor terestre și fiind prezente în toate liniile timpurii divergente ale grupurilor majore de plante. Celelalte tipuri de micorize apar în liniile cu o origine mai recentă. Micorizele orhidacee par să fi evoluat o singură dată spre deosebire de ectomicorize şi micorize ericoide care au probabil origini independente în mai multe linii de plante nerelaţionate între ele. Aceeaşi ipoteză se poate aplica şi organismelor micoheterotrofe (plante aclorofile aflate în simbioză cu diferiţi fungi, inclusiv specii din încrengătura Glomeromycota) şi plantelor non-micorize (Smith and Read, 2008).
3.6. FUNGI VEZICULAR-ARBUSCULARI. MORFOLOGIE ŞI
ANATOMIE
Gallaud (1905) a descris două tipuri de colonizare, diferenţiate de structurile anatomice interne pe care le produc ciupercile vezicular-arbusculare. Tipul de micoriză arbusculară considerat ca fiind “micoriza arbusculară tipică” este corespunzător tipului Arum descris de Gallaud. În aceste asociaţii ciuperca se răspândeşte relativ rapid în cortexul radicular prin hife intercelulare care se extind în lungul spaţiilor intercelulare ocupate de aer. Ramificaţiile laterale penetrează pereţii celulelor corticale şi se ramifică dichotomic în lumenul celulei pentru a produce arbusculi caracteristici foarte ramificați. Buclele hifale pot fi formate în timpul penetrării straturilor exoderme ale rădăcinii dar nu sunt o componentă fundamentală a colonizării corticale.
Tipul Paris, al doilea tip de micorize descris de Gallaud (1905), este caracterizat de o dezvoltare extensivă a hifelor intracelulare buclate în interiorul celulelor corticale colonizate; răspândirea hifelor se face direct, dintr-o celulă în alta. Ramificaţiile asemănătoare arbusculilor (ramificaţii arbusculare) cresc din aceste bucle dar creşterea intercelulară este foarte scăzută.

12
Capitolul 4
CADRUL NATURAL AL ZONEI DE EXPERIMENTARE
4.1. AŞEZARE GEOGRAFICĂ
Comuna Gârda de Sus este situată în partea de N-V a judeţului Alba în bazinul hidrografic superior al Arieşului Mare, principalul afluent al Arieşului fiind pârâul Gârda Seacă iar ca particularitate regională aparţine Ţării Moţilor (fig. 4.1). Este delimitată la nord de comuna Beliş (jud. Cluj), la vest de comuna Arieşeni, la est de comuna Scărişoara şi la sud de comunele Avram Iancu şi Poiana Vadului.
Teritoriul administrativ are o suprafaţă de 8.270 ha şi cuprinde 17 sate. Localităţile componente ale comunei Gârda de Sus sunt sate de munte cu vatră răsfirată, casele fiind împrăştiate pe aproximativ 60 km2, pe o diferenţă de nivel cuprinsă între 775 m şi 1.380 m. Zona delimitată de râul Arieş spre nord, datorită valorilor cadrului natural, a determinat ca această comună să fie cuprinsă în proporţie de 64,75% în Parcul Natural Apuseni.
4.3. REGIMUL CLIMATIC
În Munţii Apuseni, unde circulaţia aerului se face predominant din sectorul vestic, valorile medii anuale ale temperaturii aerului sunt în general mai scăzute pe versanţii vestici, în comparaţie cu cei estici, la aceeaşi altitudine, ca rezultat al proceselor de interacţiuni dintre suprafaţa pantelor şi aerul în mişcare. Media anuală a temperaturii aerului în zona Gheţari–Poiana Călineasa a fost prognozată în jurul valorii de 4oC de către INMH. Ianuarie este luna cu cele mai scăzute temperaturi medii multianuale.
Anul 2010 a fost caracterizat de o temperatură medie de 5.3°C, valoare foarte apropiată de cea a mediei pe anii 2001-2010 (5.2°C); cea mai călduroasă lună fiind august (16.3°C), iar cea mai rece ianuarie (-6.0°C).
La nivelul anului 2011, pe perioada ianuarie-septembrie, cea mai călduroasă lună a fost august (14,7°C), iar cea mai rece ianuarie (-4,9°C).
Pentru experimentele din teren s-au luat în calcul în special valorile din intervalul aprilie – septembrie, care în zona de

13
experimentare sunt asigurate cu temperaturi peste 5°C, ceea ce reprezintă o perioadă de maximă vegetaţie a plantelor studiate.
Cele mai mici cantităţi medii lunare se observă în lunile februarie - martie, fiind la majoritatea staţiilor în luna februarie. Mediile lunare au coborât până la cca. 60 mm în zona muntoasă înaltă, 90 mm pe versanţii vestici şi 40 mm pe cei estici în Valea Arieşului. Precipitaţiile zilnice căzute la staţia meteo Gheţari prezintă o frecvenţă ridicată pe întreaga perioadă de vegetaţie. Anul 2008 a însumat o cantitate mică de precipitații: doar 909,2 mm. Cea mai scăzută a fost înregistrată în luna iunie (7,2 mm), iar cea mai ridicată a fost în luna iulie (130,4 mm) (tabelul 2.3.).
Capitolul 5
OBIECTIVELE CERCETĂRII. MATERIALE ŞI METODE
5.1. OBIECTIVELE CERCETĂRII
Proiectul propus pentru realizarea tezei de doctorat se intitulează: “Studiul simbiozelor micoriziene în unele ecosisteme de pajişti din zona Munţilor Apuseni”.
Pentru îndeplinirea obiectivelor sugerate de titlul proiectului s-a instalat un câmp experimental în satul Gheţari, judeţul Alba, unde s-a urmărit în principal studierea influenţei pe care o au micorizele asupra creşterii şi dezvoltării plantelor, respectiv efectul unor variante de fertilizare şi tratament asupra apariţiei şi dezvoltării asociaţiilor fungi-plantă.
Alături de experienţele amplasate în Munţii Apuseni pentru îndeplinirea obiectivelor propuse pe parcursul celor doi ani de cercetare au mai fost instalate o serie de experienţe în casa de vegetaţie a Universităţii de Ştiinţe agricole şi Medicină Veterinară Cluj, cu scopul de a studia comportamentul şi strategiile de colonizare a fungilor micorizieni în sistemele radiculare ale unor plante cultivate în cultură pură în mediu controlat.
Pentru realizarea cercetărilor întreprinse în cadrul acestei teme de doctorat au fost stabilite următoarele obiective:
1. studiul dinamicii populaţiilor de micorize vezicular-arbusculare din zona de experimentare (pajişte naturală subalpină);

14
2. specificitatea fungilor faţă de anumite specii sau familii botanice;
3. studiul corelaţiilor dintre factorii climatici, tehnologici (fertilizare, tratamente fitosanitare, tehnologii de întreţinere) asupra ciclului evolutiv şi capacităţii de colonizare al fungilor micorizieni;
4. efectul caracteristicilor solului, reliefului şi altitudinii asupra formării simbiozelor micoriziene;
5. comportamentul fungilor micorizieni în anumite secvenţe de vegetaţie;
6. comportamentul fungilor micorizieni într-un mediu controlat.
5.2. MATERIALELE ŞI METODELE UTILIZATE ÎN REALIZAREA
OBIECTIVELOR
Pentru realizarea obiectivelor a fost amplasată o parcelă experimentală pe teritoriul satului Gheţari, comuna Gârda de Sus, judeţul Alba. Câmpul experimental a fost împărţit în şapte variante, în trei repetiţii, urmând ca fiecare variantă să fie subdivizată în alte patru variante (fig. 5.1.).
Factorii experimentali luaţi în studiu au fost: Factorul A (ANUL) cu două graduări:
1. 2010 2. 2011
Factorul B (SECVENŢA DE RECOLTARE) cu două graduări: 1. 100 de zile de la fertilizare 2. 150 de zile de la fertilizare
Factorul C (FERTILIZAREA) cu şapte graduări: 1. Nefertilizat 2. Gunoi de grajd (10t/ha) 3. Gunoi de grajd (10t/ha) + N50P25K25
4. N50P25K25
5. Eurofertil mezocalc (120kg/ha) + N50 6. Eurofertil mezocalc (120kg/ha) 7. Cenuşă (100kg/ha)

15
Factorul D (TRATAMENTUL) cu patru graduări: 1. Netratat 2. Sulfat de zinc (1%) 3. Mulci 4. Fungicid (Botran 75 WP, 0.07%)
Pentru facilitarea efectuării observaţiilor şi recoltării probelor variantele de tratament au fost grupate în cadrul variantelor de fertilizare.
Ca şi material biologic au fost utilizate plante din flora spontană, existente în pajiştile selectate pentru efectuarea observaţiilor. Speciile selectate, pe baza studiilor asupra compoziţiei floristice, s-au evidenţiat prin gradul mare de acoperire vegetală şi dominanţa în cadrul tipului de pajişte luat în studiu. Astfel, gramineele au fost reprezentate de Festuca rubra ca specie dominantă, iar leguminoasele de Trifolium repens.
Pe parcursul anului s-a stabilit un protocol de recoltare a probelor, separat pe două secvenţe de vegetaţie. Prima secvenţă de vegetaţie coincide cu momentul cosirii parcelei - la 100 de zile de la aplicarea fertilizanţilor şi a tratamentelor; a doua secvenţă - la 150 de zile de la aplicarea fertilizanţilor şi tratamentelor şi considerată ca fiind momentul de începere a declinului creşterii şi dezvoltării plantelor, respectiv pregătirea pentru iernare.
În anul 2011 au fost amplasate o serie de experimente în mediul controlat din casa de vegetaţie a Universităţii de Ştiinţe agricole şi Medicină Veterinară Cluj cu scopul de a urmări parametri colonizării micoriziene diferenţiat pe secvenţe de vegetaţie, respectiv pentru a găsi anumite puncte de referinţă care să poată fi relaţionate cu datele din teren, pentru formarea unei imagini cât mai complete asupra funcţionării simbiozelor micoriziene.
Durata experimentelor a fost de 150 de zile, plantele utilizate fiind aceleaşi ca şi în câmpul experimental, respectiv Trifolium repens şi Festuca rubra.
Pentru primul experiment înfiinţat recoltarea variantelor s-a făcut progresiv, la un interval de 10 zile una faţă de cealaltă începând cu prima recoltare, programată la 10 zile de la răsărirea plantelor. Au fost amplasate 15 variante în trei repetiţii iar însămânţarea s-a efectuat pe un sol de tip terra rosa, identic cu cel din câmpul experimental
Un al doilea experiment compus din şapte variante a fost instalat pentru a analiza efectul fertilizanţilor asupra parametrilor colonizării în mediul controlat din fitotron. Fertilizarea s-a făcut utilizând acelaşi protocol experimental ca şi în cazul experienţelor din teren, solul fiind

16
identic cu cel din câmpul experimental amplasat în satul Gheţari. Recoltarea probelor de rădăcini s-a făcut în două secvenţe, la 100 şi 150 de zile de la răsărirea plantelor.
Probele de rădăcini de Festuca rubra şi Trifolium repens prelevate din teren şi din casa de vegetaţie au fost analizate pentru determinarea parametrilor colonizării micoriziene.
Experienţele de laborator au presupus o prelucrare probelor de rădăcini pentru a putea fi analizate microscopic.
Pentru analizarea microscopică a fungilor vezicular-arbusculari metoda clasică presupune folosirea unor substanţe periculoase (tripan albastru), cu efect cancerigen. De aceea, s-au folosit două o metode alternative la care s-au adus o serie de modificări, iar rezultatele au indicat o eficienţă bună a colorării (Stoian şi Florian, 2009a, 2009b).
Metoda I de colorare cuprinde patru etape distincte: 1. Etapa de curăţare – rădăcinile au fost fierte timp de 3-5
minute (în funcţie de specia de la care au fost colectate) într-o soluţie de 10% KOH (hidroxid de potasiu).
2. Etapa de clătire – rădăcinile se clătesc de câteva ori cu apă distilată.
3. Etapa de colorare – presupune fierberea rădăcinilor timp de 3 minute într-o soluţie de 5% cerneală (albastră, neagră sau roşie - Pelikan) şi 5% oţet alb (provenit din oţet alimentar cu 9% acid acetic).
4. Etapa de decolorare – în funcţie de tipul cernelii folosite rădăcinile au fost clătite timp de 10-20 de minute cu apă distilată sau timp de 3 minute cu oţet pur.
Modificările aduse metodei s-au datorat similarităţii dintre tăria hidroxidului de potasiu şi hidroxidul de sodiu, preferinţa pentru utilizarea acestuia din urmă fiind de natură practică, deoarece este mai uşor de găsit în comerţ.
Metoda II de colorare a fost adaptată dotărilor şi condiţiilor existente în laboratorul de la Gheţari, cele patru etape fiind modificate astfel:
1. Etapa de curăţare – rădăcinile sunt imersate într-o soluţie de 10% NaOH timp de 24-48 de ore (în funcţie de specie şi dezvoltarea sistemului radicular).
2. Etapa de clătire – rădăcinile se clătesc de câteva ori cu apă distilată.
3. Etapa de colorare – presupune imersarea timp de 24 de ore într-o soluţie de 5% cerneală (albastră, neagră sau roşie –

17
Pelikan) şi 5% oţet alb (provenit din oţet alimentar cu 9% acid acetic).
4. Etapa de decolorare – în funcţie de tipul cernelii folosite, rădăcinile au fost clătite timp de 10-20 de minute cu apă distilată, sau timp de 3 minute cu oţet pur.
Rezultatele au demonstrat că după un timp de curăţare adecvat se obţine o colorare excelentă a rădăcinilor, fapt care face posibilă identificare micorizelor.
Estimarea colonizării micoriziene s-a conform parametrilor propuşi de Trouvelot et al. (1986, citat de www2.dijon.inra.fr), notele acordate colonizării fiind în funcţie de dezvoltarea miceliului intraradicular şi abundenţa arbusculilor.
Parametrii propuşi de Trouvelot şi formula după care se calculează
sunt: - Frecvenţa colonizării micoriziene în sistemul radicular
- Intensitatea colonizării micoriziene în sistemul radicular
(n = numărul de fragmente, 5, 4, 3, 2, 1 – notele acordate
fragmentelor) - Intensitatea colonizării micoriziene în fragmentele de
rădăcină
- Abundenţa arbusculilor în părţile micorizate ale
fragmentelor de rădăcină
(mA3, mA2, mA1 sunt procentele de m, notate cu A3, A2 şi A1)
- Abundenţa arbusculilor în sistemul radicular
Pentru a crea o imagine cât mai completă asupra stării de
colonizare micoriziană a fost adaptată formula de calcul a gradului de atac folosită în aprecierea stării fitosanitare a culturilor agricole.

18
În lucrarea de faţă gradul de atac a fost denumit grad de colonizare micoriziană, formula de calcul fiind:
GC = (F*I ) / 100
Capitolul 6
REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIILE FESTUCA RUBRA ŞI TRIFOLIUM
REPENS CULTIVATE ÎN MEDIU CONTROLAT
6.1. REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIA FESTUCA RUBRA
Analiza grafică a relaţiei dintre frecvenţa, intensitatea colonizării
şi abundenţa arbusculilor în sistemul radicular al plantelor de Festuca rubra prezentată în figura 6.1., de-a lungul unui interval de vegetaţie de 150 de zile, scoate în evidenţă o serie de conexiuni între parametri colonizării şi secvenţa de vegetaţie în care au fost înregistraţi.
Secvenţa considerată ca martor în acest experiment reprezintă valorile parametrilor colonizării micoriziene la 10 zile de la răsărirea plantelor. Astfel frecvenţa a înregistrat o valoare de 91,11%, intensitatea colonizării a atins un nivel de 49,39% iar abundenţa arbusculilor în sistemul radicular un procent de 30,13%.
La 20 de zile valoarea frecvenţei colonizării urcă până la 100%, totodată crescând şi intensitatea şi abundenţa arbusculilor. Valorile ridicate ale acestor parametri indică o permisivitate crescută a plantei gazdă pentru colonizare, o capacitate mare de colonizare a ciupercii, respectiv stabilizarea interfeţei de transfer al elementelor nutritive între cei doi parteneri creată în prima secvenţă de vegetaţie.
La 30 de zile valorile înregistrate indică un regres al colonizării, frecvenţa scăzând la valoarea de 85,56% iar intensitatea până la o valoare de 14,53%, respectiv abundenţa arbusculilor la o valoare de 2,55%. Fenomenul se poate explica pe baza transferului intens din primele 20 de zile, fapt care a adus plantei necesarul de elemente nutritive pentru o dezvoltare mai rapidă a sistemului radicular, ciuperca nefiind capabilă de a păstra un ritm de colonizare egal cu cel al producţiei de rădăcini tinere.
Intervalul 40 – 70 de zile de la răsărire este marcat de o creştere a intensităţii colonizării la 40 de zile până la 60,61% urmată de o scădere

19
până la o valoare de 32,39% şi din nou o creştere până la 69,50% la 70 de zile. În mod similar fluctuează şi abundenţa arbusculilor în sistemul radicular, procentul fiind de 26,75% la începutul intervalului, urmând o scădere până la 0 (la 50 de zile) şi apoi o creştere până la 51,79% la finalul intervalului. În toată această perioadă, valoarea frecvenţei colonizării rămâne fixată la 100%.
Fig.6.1. Valorile frecvenţei (F%), intensităţii (I%) şi abundenţei
arbusculilor (A%) pe o perioadă de 150 zile
Apariţia valorii de 0% la 50 de zile de la răsărire în ceea ce priveşte procentul de arbusculi din sistem radicular se poate explica pe baza duratei scurte de viaţă acestor structuri de transfer relaţionată cu orientarea ciupercii înspre producerea de structuri colonizatoare pentru de a recupera spaţiul de colonizare din rădăcinile tinere pierdut în intervalul precedent.
Secvenţa de 80 de zile se evidenţiază printr-o nouă scădere a frecvenţei colonizării, o scădere a intensităţii până la o valoare de 3,20% şi absenţa arbusculilor.
În intervalul 90 – 110 zile se remarcă o nouă creştere a parametrilor colonizării, frecvenţa apropiindu-se de valoarea de 100%, intensitatea având o valoare de 45,73% şi o abundenţă a arbusculilor de 17,06% la 90 de zile. În ceea ce priveşte restul intervalului, intensitatea scade la 100 de zile şi creşte din nou la 110 zile în timp ce procentul de arbusculi se situează sub 1%. În acest moment ciuperca este orientată înspre producerea de vezicule cu rol de înmagazinare a substanţelor nutritive acumulate de la planta gazdă.
Perioada 120 – 140 de zile reprezintă un declin al colonizării micoriziene, atât intensitatea cât şi procentul de arbusculi având valori

20
reduse, explicaţia posibilă fiind pregătirea ciupercii pentru procesul de sporulaţie.
La finalul perioadei de vegetaţie se observă o nouă creştere a frecvenţei colonizării dar intensitatea şi procentul de arbusculi rămân la un nivel redus.
Analiza dendrogramei din figura 6.2. oferă date despre gradul de înrudire dintre parametri colonizării respectiv strategia de colonizare a sistemului radicular de către ciupercile simbionte.
Pentru simplificarea analizei modelului explicaţiile se referă doar la 2 clustere principale, compuse fiecare din câte 2 sub-clustere.
Sub-clusterul A1 este compus din secvenţele de vegetaţie corespunzătoare perioadei de 80, 120, 130 şi 140 de zile. Gruparea acestor secvenţe s-a făcut pe baza valorilor medii ale frecvenţei, corelate cu valori sub 5% ale intensităţii şi absenţa arbusculilor din sistemul radicular. Caracterul sub-clusterului astfel obţinut este unul de înmagazinare, strategia ciupercii fiind cea de formare a veziculelor şi de depozitare a substanţelor nutritive acumulate.
Sub-clusterul A2 este format prin gruparea secvenţelor de 30, 50, 100, 110 şi 150 de zile de la răsărirea plantelor luate în experiment. Valorile care caracterizează acest sub-cluster sunt peste 85% în cazul frecvenţei dar nu depăşesc 35% ca procent de intensitate iar abundenţa arbusculilor este foarte redusă. Aceste aspecte evidenţiază caracterul de colonizare superficială a acestei subunităţi, strategia ciupercii pe parcursul secvenţelor de vegetaţie componente fiind de extindere a sistemului de hife la nivelul superficial al endodermului radicular, acaparând aproape toată suprafaţa rădăcinilor tinere dezvoltate de planta gazdă dar penetrând foarte puţin profunzimea cortexului radicular.
Combinând caracteristicile sub-clusterelor A1 şi A2 se formează un cluster caracterizat de parametri indicatori ai unor secvenţe de vegetaţie pe parcursul cărora are loc o colonizare superficială a ţesuturilor cortexului radicular dar şi etapa de producere a veziculelor şi înmagazinare a substanţelor nutritive.
În ceea ce priveşte secvenţele de vegetaţie de 20 şi 70 de zile, acestea sunt caracterizate de valori de 100% ale frecvenţei, valori de 74,67% şi 69,50% ale intensităţii colonizării şi cele mai mari valori ale abundenţei arbusculilor. Corelând valorile acestor parametri se evidenţiază un caracter dual al sub-clusterului B1; strategia fungilor simbionţi pe parcursul celor două secvenţe fiind îndreptată atât spre colonizarea inter- şi intracelulară cât şi spre crearea unei interfeţe de schimb a elementelor nutritive, prin intermediul arbusculilor, în interiorul celulelor colonizate.
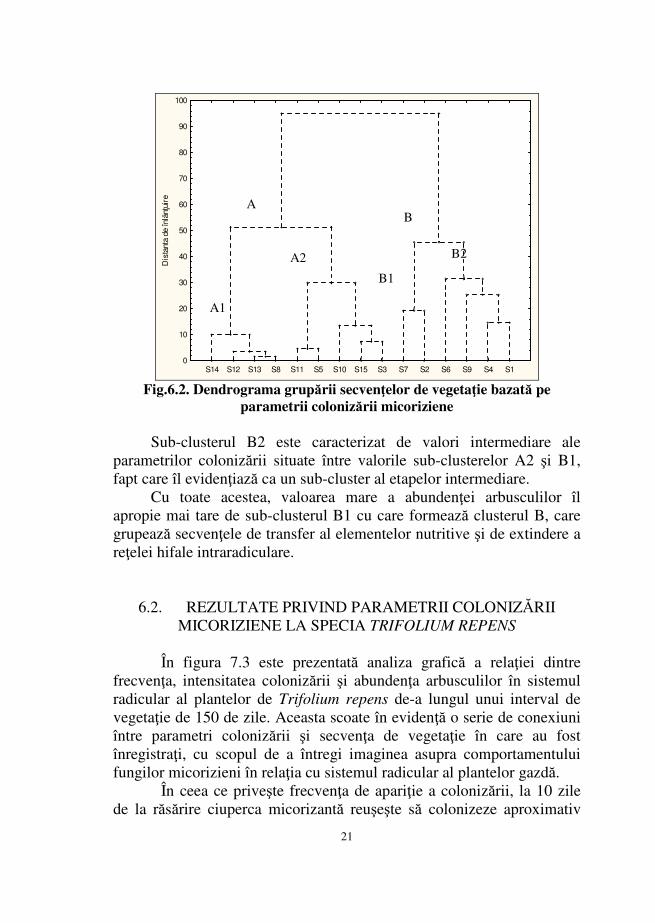
21
S14 S12 S13 S8 S11 S5 S10 S15 S3 S7 S2 S6 S9 S4 S1
0
10
20
30
40
50
60
70
80
90
100
Dis
tanta
de înlă
nţu
ire
Fig.6.2. Dendrograma grupării secvenţelor de vegetaţie bazată pe
parametrii colonizării micoriziene Sub-clusterul B2 este caracterizat de valori intermediare ale
parametrilor colonizării situate între valorile sub-clusterelor A2 şi B1, fapt care îl evidenţiază ca un sub-cluster al etapelor intermediare.
Cu toate acestea, valoarea mare a abundenţei arbusculilor îl apropie mai tare de sub-clusterul B1 cu care formează clusterul B, care grupează secvenţele de transfer al elementelor nutritive şi de extindere a reţelei hifale intraradiculare.
6.2. REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIA TRIFOLIUM REPENS
În figura 7.3 este prezentată analiza grafică a relaţiei dintre
frecvenţa, intensitatea colonizării şi abundenţa arbusculilor în sistemul radicular al plantelor de Trifolium repens de-a lungul unui interval de vegetaţie de 150 de zile. Aceasta scoate în evidenţă o serie de conexiuni între parametri colonizării şi secvenţa de vegetaţie în care au fost înregistraţi, cu scopul de a întregi imaginea asupra comportamentului fungilor micorizieni în relaţia cu sistemul radicular al plantelor gazdă.
În ceea ce priveşte frecvenţa de apariţie a colonizării, la 10 zile de la răsărire ciuperca micorizantă reuşeşte să colonizeze aproximativ
B
B1
A2
A1
A
B2

22
jumătate din sistemul radicular (52,22%) dar cu o intensitate de doar 2,07% şi o abundenţă a arbusculilor de 0,11%. Parametri colonizării cresc începând cu secvenţa de vegetaţie de 20 de zile frecvenţa atingând valoarea de 100%, pe care o păstrează până la 70 de zile de la răsărire.
Pe parcursul aceluiaşi interval intensitatea colonizării creşte atinge valoarea de 76,17% la 20 de zile şi 71% la 30 de zile, scăzând apoi până la o valoare de 42,28% la 70 de zile de la răsărirea plantelor, în timp ce abundenţa arbusculilor creşte până la o valoare de 28,60% la 20 de zile, respectiv 35,84% la 30 de zile, dar scade până la 0,13% în timpul secvenţei de 50 de zile şi creşte din nou până la o valoare de 14,66% la 70 de zile.
Faţă de trendul descreşterii intensităţii colonizării, secvenţa de 60 de zile face o excepţie, valoarea fiind de 62,67% (mai mare decât la secvenţele de 50 şi 70 de zile), dar cu o valoare a abundenţei arbusculilor de doar 0,13%.
Momentul stabilirii colonizării în sistemul radicular este evidenţiat de valori reduse ale frecvenţei şi intensităţii colonizării, respectiv a abundenţei arbusculilor. O dată cu creşterea şi dezvoltarea plantelor, solicitarea de elemente nutritive provoacă o creştere continuă a sistemului radicular.
Acest aspect influenţează colonizarea micoriziană, crescând frecvenţa cu care fungii colonizează rădăcinile nou formate şi ajungând la o valoare de 100% a prezenţei hifelor miceliene pe suprafaţa endodermului radicular în intervalul 20 – 70 de zile de la răsărire.
Fig.6.3. Valorile frecvenţei (F%), intensităţii (I%) şi abundenţei arbusculilor
(A%) pe o perioadă de 150 zile
În ceea ce priveşte intensitatea colonizării în acelaşi interval de timp, valorile se încadrează între 40 şi 80%, descrescând de la valoarea înregistrată în secvenţa de 20 de zile, în timp ce abundenţa arbusculilor

23
în sistem creşte în secvenţele de vegetaţie 2 şi 3, fapt care evidenţiază etapa de maximă dezvoltare atât a sistemului radicular cât şi a partenerului micorizian. Procentul ridicat de arbusculi în sistem indică un proces intens de transfer al substanţelor nutritive între cei doi simbionţi.
Intervalul 80 – 120 de zile este marcat de o descreştere a frecvenţei de apariţie a colonizării, comparativ cu intervalul precedent, până la o valoare de 66,67% în cazul secvenţei de 80 de zile, crescând apoi până la valoarea de 100% pe intervalul 90 – 120 de zile. Intensitatea colonizării fluctuează, pornind de la o valoare de 8.87% la 80 de zile, crescând până la valoarea de 64,60% la 90 de zile, scăzând la o valoare de 20,11% (la 100 de zile) şi atingând valoarea de la 76,11% la 120 de zile de la răsărire, în timp ce abundenţa arbusculilor înregistrează valori foarte mari la secvenţele de 90 şi 120 de zile (36,69%, respectiv 52,07%), dar şi valori reduse la 80 şi 100 de zile (2,42%, respectiv 1,61%).
Fluctuaţia valorilor parametrilor colonizării indică o serie de transformări în ciclul biologic al ciupercii, producţia de structuri a ciupercii în timpul secvenţelor de 70, 90 şi 110 a fost orientată spre producerea de arbusculi pentru intensificarea transferului. Secvenţele de 80 şi 100 de zile au fost caracterizate de formarea de vezicule şi depozitarea în ele a substanţelor nutritive acumulate.
Secvenţele de vegetaţie de la 130 la 150 de zile marchează un declin al colonizării în sistemul radicular, valorile frecvenţei fiind peste 55%, dar sub 75%, în timp ce intensitatea colonizării se situează sub 5%, prezenţa arbusculilor nemaifiind semnalată. Acest aspect indică o nouă schimbare a structurilor produse de către ciuperca simbiontă, aceasta pregătindu-se de sporulaţie şi finalizarea unui ciclu de viaţă.
Dendrograma din figura 6.4. indică o grupare în clustere a secvenţelor de vegetaţie în funcţie de gradul de înrudire dintre ele, bazată pe distanţele euclidiene şi înlănţuirea completă dintre valorile frecvenţei, intensităţii colonizării şi abundenţa arbusculilor în sistemul radicular.
Din cele 15 secvenţe de vegetaţie analizate, s-au obţinut o serie de clustere bazate pe relaţiile dintre valorile fiecărei secvenţe, respectiv relaţiile de înrudire dintre secvenţe. Pentru simplificarea analizării modelului s-au ales doar clusterele mari, care conţin mai multe perechi de secvenţe înrudite, respectiv sub-clusterele lor care sunt componente subordonate primar.
Analiza modului de dezvoltare al fungilor micorizieni pe parcursul secvenţelor cuprinse de clusterul A îi imprimă acestuia un

24
caracter colonizator, fungii dezvoltându-se atât la suprafaţa cât şi în profunzimea sistemului radicular. Cantitatea redusă de arbusculi dovedeşte că interfaţa de transfer a elementelor nutritive între cei doi parteneri simbiotici este restrânsă ca şi suprafaţă, motiv care ne îndeamnă să credem că în perioada corespunzătoare acestor secvenţe între cei doi parteneri se creează un contact intim în special la nivel intercelular, contactul la nivel intracelular fiind foarte redus.
Clusterul B este compus din cinci secvenţe de vegetaţie. Pe baza analizei acestor secvenţe se poate concluziona că acest cluster are un caracter atât colonizator cât şi de transfer. Valorile frecvenţelor sunt de 100% la secvenţele corespunzătoare perioadelor de 20, 30, 110 şi 120 de zile, respectiv de 96,67% la secvenţa de 90 de zile de la răsărire. În ceea ce priveşte intensitatea colonizării sistemului radicular, valoarea înregistrată la secvenţa de 90 de zile este de 64,60%, valorile celorlalte secvenţe depăşind 70%. Din punct de vedere al transferului elementelor nutritive, procentul mare al abundenţei arbusculilor la toate secvenţele indică un proces intens de transfer între cei doi parteneri.
Contactul intim crescut dintre cei doi parteneri, atât la nivel intercelular cât şi intracelular, dovedeşte că pe perioadele corespunzătoare secvenţelor componente ale clusterului B ambii parteneri au nevoie de simbioză pentru o mai bună creştere şi dezvoltare.
S10 S7 S5 S6 S4 S11 S9 S3 S12 S2 S14 S8 S15 S13 S1
0
10
20
30
40
50
60
70
80
90
100
Dis
tanţ
a de
înlă
nţui
re
Fig.6.4. Dendrograma grupării secvenţelor de vegetaţie pe baza
parametrilor colonizării micoriziene

25
Analiza secvenţelor componente ale clusterului C oferă indicii asupra fenomenului de regresie al colonizării simbiontului fungal faţă de dezvoltarea sistemului radicular al plantei gazdă.
Frecvenţa de colonizare redusă, cu valori cuprinse în intervalul 50 – 75%, intensitatea colonizării cu o valoare sub 10% şi procentul redus sau chiar absenţa arbusculilor din sistemul radicular indică o colonizare incipientă în cazul secvenţei de 10 zile, un proces de depozitare a elementelor nutritive în vezicule la 80 de zile, respectiv o pregătire pentru sporulare şi formare de noi vezicule în cazul secvenţelor cuprinse în intervalul 130 – 150 de zile de la răsărire.
6.3. REZULTATE COMPARATIVE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIILE FESTUCA RUBRA ŞI
TRIFOLIUM REPENS
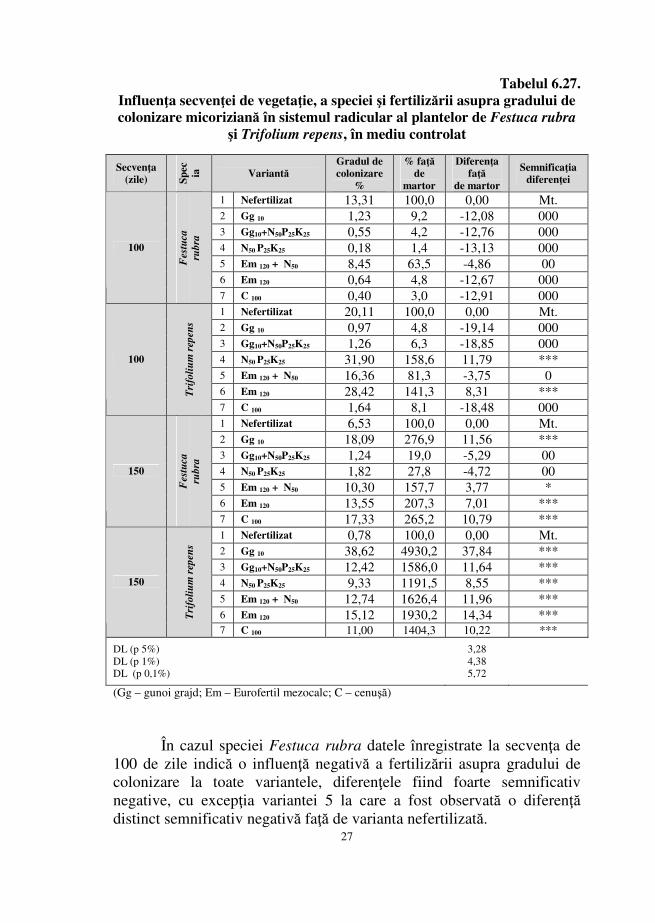
Analiza gradului de colonizare al speciilor Festuca rubra şi Trifolium repens prezentată în tabelul 6.24. evidenţiază un comportament al fungilor micorizieni similar cu cel observat în urma analizei intensităţii colonizării.
Valorile gradului de colonizare la specia Festuca rubra au un caracter fluctuant pe parcursul unei perioade de vegetaţie de 150 de zile, crescând şi descrescând cu diferenţe statistice foarte semnificativ pozitive şi foarte semnificativ negative pe parcursul intervalului 20 – 70 de zile, intervalul imediat următor fiind marcat doar de scăderi faţă de secvenţa martor.
În ceea ce priveşte comportamentul simbiozei la nivelul sistemului radicular al speciei Trifolium repens, pe durata intervalului 20 – 120 de zile se înregistrează valori mai crescute faţă de valoarea secvenţei martor, diferenţele statistice fiind foarte semnificativ pozitive, cu excepţia valorii obţinute la secvenţa 80.
Analiza cu ajutorul testului Duncan a influenţei secvenţelor asupra gradului de colonizare a sistemelor radiculare ale celor două specii indică cea mai favorabilă secvenţă ca fiind cea de 20 de zile la specia Trifolium repens, lipsa influenţei la secvenţele de 90 şi 110 zile la specia Festuca rubra respectiv secvenţa de 150 de zile care are cea mai redusă favorabilitate a influenţei.
Influenţa fertilizării asupra gradului de colonizare micoriziană a sistemelor radiculare ale speciilor Festuca rubra şi Trifolium repens pe parcursul secvenţelor de vegetaţie de 100 şi 150 de zile de la răsărire este analizată statistic în tabelul 6.27.

26
Tabelul 6.24. Influenţa secvenţei de vegetaţie şi a speciei asupra gradului de colonizare
micoriziană în sistemul radicular al plantelor de Festuca rubra şi Trifolium
repens, în mediu controlat
Specia Număr de
zile
Grad colonizare
%
% faţă de
martor
Diferenţa faţă
de martor
Semnificaţia diferenţei
10 45,04 100,0 0,00 Mt. 20 74,67 165,8 29,62 *** 30 12,48 27,7 -32,56 000 40 60,61 134,6 15,57 *** 50 32,39 71,9 -12,65 000 60 68,89 152,9 23,85 *** 70 69,50 154,3 24,46 *** 80 2,25 5,0 -42,79 000 90 37,83 84,0 -7,22 0
100 13,31 29,5 -31,73 000 110 34,23 76,0 -10,82 000 120 0,99 2,2 -44,05 000 130 1,87 4,1 -43,18 000 140 0,81 1,8 -44,23 000
Festuca
rubra
150 6,53 14,5 -38,51 000 10 1,15 100,0 0,00 Mt. 20 76,17 6623,5 75,02 *** 30 71,00 6174,2 69,85 *** 40 59,39 5164,1 58,24 *** 50 46,89 4077,4 45,74 *** 60 62,67 5449,3 61,52 *** 70 42,28 3676,2 41,13 *** 80 7,16 622,9 6,01 * 90 62,50 5434,8 61,35 ***
100 20,11 1749,0 18,96 *** 110 71,97 6258,0 70,82 *** 120 76,11 6618,3 74,96 *** 130 0,81 70,1 -0,34 - 140 3,41 296,8 2,26 -
Trifolium
repens
150 0,78 68,1 -0,37 -
DL (p 5%) DL (p 1%) DL (p 0,1%)
5,70 7,59 9,88
27
Tabelul 6.27. Influenţa secvenţei de vegetaţie, a speciei şi fertilizării asupra gradului de colonizare micoriziană în sistemul radicular al plantelor de Festuca rubra
şi Trifolium repens, în mediu controlat
Secvenţa (zile) Sp
ec ia
Variantă Gradul de colonizare
%
% faţă de
martor
Diferenţa faţă
de martor
Semnificaţia diferenţei
1 Nefertilizat 13,31 100,0 0,00 Mt. 2 Gg 10 1,23 9,2 -12,08 000 3 Gg10+N50P25K25 0,55 4,2 -12,76 000 4 N50 P25K25 0,18 1,4 -13,13 000 5 Em 120 + N50 8,45 63,5 -4,86 00 6 Em 120 0,64 4,8 -12,67 000
100
Fes
tuca
rub
ra
7 C 100 0,40 3,0 -12,91 000 1 Nefertilizat 20,11 100,0 0,00 Mt. 2 Gg 10 0,97 4,8 -19,14 000 3 Gg10+N50P25K25 1,26 6,3 -18,85 000 4 N50 P25K25 31,90 158,6 11,79 *** 5 Em 120 + N50 16,36 81,3 -3,75 0 6 Em 120 28,42 141,3 8,31 ***
100
Tri
foli
um
rep
ens
7 C 100 1,64 8,1 -18,48 000 1 Nefertilizat 6,53 100,0 0,00 Mt. 2 Gg 10 18,09 276,9 11,56 *** 3 Gg10+N50P25K25 1,24 19,0 -5,29 00 4 N50 P25K25 1,82 27,8 -4,72 00 5 Em 120 + N50 10,30 157,7 3,77 * 6 Em 120 13,55 207,3 7,01 ***
150
Fes
tuca
rub
ra
7 C 100 17,33 265,2 10,79 *** 1 Nefertilizat 0,78 100,0 0,00 Mt. 2 Gg 10 38,62 4930,2 37,84 *** 3 Gg10+N50P25K25 12,42 1586,0 11,64 *** 4 N50 P25K25 9,33 1191,5 8,55 *** 5 Em 120 + N50 12,74 1626,4 11,96 *** 6 Em 120 15,12 1930,2 14,34 ***
150
Tri
foli
um
rep
ens
7 C 100 11,00 1404,3 10,22 ***
DL (p 5%) DL (p 1%) DL (p 0,1%)
3,28 4,38 5,72
(Gg – gunoi grajd; Em – Eurofertil mezocalc; C – cenuşă)
În cazul speciei Festuca rubra datele înregistrate la secvenţa de 100 de zile indică o influenţă negativă a fertilizării asupra gradului de colonizare la toate variantele, diferenţele fiind foarte semnificativ negative, cu excepţia variantei 5 la care a fost observată o diferenţă distinct semnificativ negativă faţă de varianta nefertilizată.

28
Influenţa negativă a fertilizării este regăsită şi la specia Trifolium repens, dar numai la variantele 2, 3 şi 6. Diferenţele statistice faţă de martor fiind foarte semnificativ negative la variantele 2 şi 3, respectiv semnificativ negativă la varianta 6.
Secvenţa de vegetaţie de 150 de zile prezintă o serie de modificări în ceea ce priveşte gradul de colonizare la ambele specii.
La Festuca rubra se observă o continuitate a influenţei negative a fertilizării la variantele 3 şi 4 dar valorile înregistrate indică o diferenţă mai scăzută, asigurată statistic ca fiind distinct semnificativ negativă faţă de valoarea observată la varianta nefertilizată. La specia Trifolium repens se remarcă o influenţă benefică a fertilizării, valorile înregistrate la toate variantele având diferenţe foarte semnificativ pozitive faţă de varianta martor
Analiza datelor înregistrate cu ajutorul testului Duncan reliefează ierarhizarea variantelor de fertilizare şi secvenţa în care au fost aplicate în funcţie de influenţa pe care au avut-o asupra gradului de colonizare al celor două specii.
Cea mai puţin favorabilă variantă de fertilizare în combinaţie cu secvenţa la care a fost aplicată este fertilizarea cu NPK din prima secvenţă de vegetaţie la nivelul speciei Festuca rubra. Cea mai favorabilă variantă de fertilizare a fost indicată cea cu gunoi de grajd, la nivelul celei de-a doua secvenţe de vegetaţie la specia Trifolium repens.
Capitolul 7
REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIILE FESTUCA RUBRA ŞI TRIFOLIUM
REPENS ÎN CONDIŢIILE EXPERIMENTALE DE LA GHEŢARI
7.1.REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIA FESTUCA RUBRA ÎN PERIOADA 2010-
2011 În tabelul 7.19. se prezintă influenţa fertilizării efectuată în anii
2010 şi 2011 asupra gradulului colonizării micoriziene în sistemul radicular al plantelor de Festuca rubra.
Comparaţia dintre valorile gradului de colonizare înregistrat în cei doi ani de studiu dezvăluie o influenţă negativă a fertilizării la variantele 4 şi 6 în 2010, respectiv o influenţă benefică a fertilizării la asupra variantelor 3, 5 şi 7, diferenţele înregistrate faţă de varianta nefertilizată fiind asigurate statistic ca distinct semnificativ pozitive în
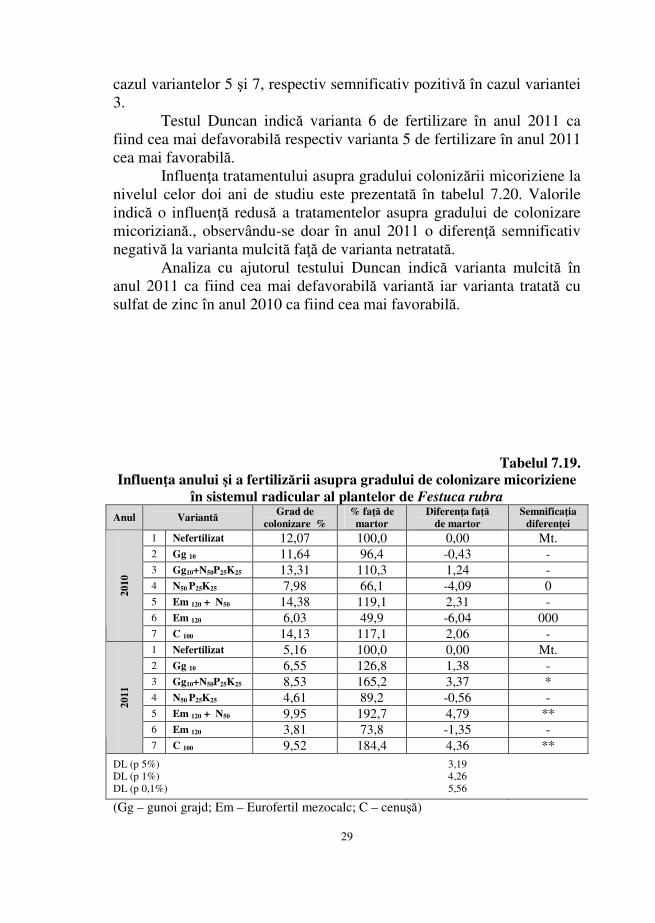
29
cazul variantelor 5 şi 7, respectiv semnificativ pozitivă în cazul variantei 3.
Testul Duncan indică varianta 6 de fertilizare în anul 2011 ca fiind cea mai defavorabilă respectiv varianta 5 de fertilizare în anul 2011 cea mai favorabilă.
Influenţa tratamentului asupra gradului colonizării micoriziene la nivelul celor doi ani de studiu este prezentată în tabelul 7.20. Valorile indică o influenţă redusă a tratamentelor asupra gradului de colonizare micoriziană., observându-se doar în anul 2011 o diferenţă semnificativ negativă la varianta mulcită faţă de varianta netratată.
Analiza cu ajutorul testului Duncan indică varianta mulcită în anul 2011 ca fiind cea mai defavorabilă variantă iar varianta tratată cu sulfat de zinc în anul 2010 ca fiind cea mai favorabilă.
Tabelul 7.19. Influenţa anului şi a fertilizării asupra gradului de colonizare micoriziene
în sistemul radicular al plantelor de Festuca rubra Anul Variantă
Grad de colonizare %
% faţă de martor
Diferenţa faţă de martor
Semnificaţia diferenţei
1 Nefertilizat 12,07 100,0 0,00 Mt. 2 Gg 10 11,64 96,4 -0,43 - 3 Gg10+N50P25K25 13,31 110,3 1,24 - 4 N50 P25K25 7,98 66,1 -4,09 0 5 Em 120 + N50 14,38 119,1 2,31 - 6 Em 120 6,03 49,9 -6,04 000
2010
7 C 100 14,13 117,1 2,06 - 1 Nefertilizat 5,16 100,0 0,00 Mt. 2 Gg 10 6,55 126,8 1,38 - 3 Gg10+N50P25K25 8,53 165,2 3,37 * 4 N50 P25K25 4,61 89,2 -0,56 - 5 Em 120 + N50 9,95 192,7 4,79 ** 6 Em 120 3,81 73,8 -1,35 -
2011
7 C 100 9,52 184,4 4,36 ** DL (p 5%) DL (p 1%) DL (p 0,1%)
3,19 4,26 5,56
(Gg – gunoi grajd; Em – Eurofertil mezocalc; C – cenuşă)

30
Tabelul 7.20. Influenţa anului şi a tratamentului asupra gradului colonizării micoriziene
în sistemul radicular al plantelor de Festuca rubra Anul
Variantă
Grad de colonizare
%
% faţă de
martor
Diferenţa faţă
de martor
Semnificaţia diferenţei
Testul Duncan
1 Netratat 10,82 100,0 0,00 Mt. C 2 Sulfat de zinc1% 11,78 108,9 0,96 - C 3 Mulci 11,37 105,0 0,54 - C
2010
4 Botran 0.07% 11,47 106,0 0,64 - C 1 Netratat 7,69 100,0 0,00 Mt. B 2 Sulfat de zinc1% 6,82 88,7 -0,87 - AB 3 Mulci 4,90 63,7 -2,79 0 A
2011
4 Botran 0.07% 8,09 105,2 0,40 - B DL (p 5%) DL (p 1%) DL (p 0,1%)
2,29 3,02 3,89
DS
2,29 – 2,66
7.2. REZULTATE PRIVIND PARAMETRII COLONIZĂRII MICORIZIENE LA SPECIA TRIFOLIUM REPENS ÎN PERIOADA
2010-2011
În tabelul 7.40. se prezintă influenţa fertilizării efectuate în anii 2010 şi 2011 asupra gradulului colonizării micoriziene în sistemul radicular al plantelor de Trifolium repens.
Comparaţia dintre valorile gradului de colonizare înregistrat în cei doi ani de studiu dezvăluie o influenţă negativă a fertilizării la variantele 2, 3, 5 şi 6 în 2010, cu diferenţe foarte semnificativ negative, respectiv o influenţă benefică asupra variantei 4, diferenţa înregistrată faţă de varianta nefertilizată fiind asigurată statistic ca distinct semnificativ pozitive.
Testul Duncan indică cea mai defavorabilă variantă ca fiind varianta 6 de fertilizare în anul 2011 iar cea mai favorabilă varianta 4 în anul 2010.
În tabelul 7.41. este prezentată influenţa tratamentului asupra gradului colonizării micoriziene la nivelul celor doi ani de studiu. Varianta mulcită a influenţat în mod negativ gradul de colonizare micoriziană al nivelul ambilor ani, diferențele înregistrate fiind asigurate statistic ca foarte semnificativ negative.

31
Tabelul 7.40. Influenţa anului şi a fertilizării asupra gradului de colonizare
micoriziene în sistemul radicular al plantelor de Trifolium repens
Anul Variantă Grad de
colonizare %
% faţă de
martor
Diferenţa faţă de
martor
Semnificaţia diferenţei
Testul Duncan
1 Nefertilizat 18,92 100,0 0,00 Mt. HI
2 Gg 10 9,81 51,8 -9,11 000 C
3 Gg10+N50P25K25 16,52 87,3 -2,40 000 G
4 N50 P25K25 20,52 108,5 1,61 ** J
5 Em 120 + N50 6,78 35,9 -12,13 000 B
6 Em 120 11,25 59,5 -7,66 000 D
2010
7 C 100 18,78 99,3 -0,14 - HI
1 Nefertilizat 19,13 100,0 0,00 Mt. I
2 Gg 10 6,03 31,5 -13,10 000 AB
3 Gg10+N50P25K25 13,13 68,7 -5,99 000 E
4 N50 P25K25 17,88 93,5 -1,24 0 H
5 Em 120 + N50 5,97 31,2 -13,16 000 AB
6 Em 120 5,36 28,0 -13,76 000 A
2011
7 C 100 14,69 76,8 -4,44 000 F
DL (p 5%) DL (p 1%) DL (p 0,1%)
1,06 1,42 1,85
DS
1,07 – 1,28
(Gg – gunoi grajd; Em – Eurofertil mezocalc; C – cenuşă)
Tabelul 7.41.
Influenţa anului şi a tratamentului asupra gradului colonizării micoriziene în sistemul radicular al plantelor de Trifolium repens
Anul Variantă Grad de
colonizare %
% faţă de
martor
Diferenţa faţă
de martor
Semnificaţia Diferenţei
Testul Duncan
1 Netratat 13,54 100,0 0,00 Mt. D
2 Sulfat de zinc1% 13,46 99,4 -0,09 - D
3 Mulci 11,83 87,3 -1,72 000 C 2010
4 Botran 0.07% 19,79 146,1 6,25 *** F
1 Netratat 10,69 100,0 0,00 Mt. B
2 Sulfat de zinc1% 11,68 109,3 1,00 * C
3 Mulci 9,17 85,8 -1,52 000 A 2011
4 Botran 0.07% 15,43 144,4 4,75 *** E
DL (p 5%) DL (p 1%) DL (p 0,1%)
0.77 1.02 1.31
DS
0,77–0,90
Tratamentul cu sulfat de zinc nu a influenţat gradul de colonizare
în anul 2010, influenţa începând să se facă simţită abia în anul 2011,

32
când s-a înregistrat o diferenţă semnificativ pozitivă faţă de varianta martor.
Aplicarea fungicidului a avut un efect benefic asupra gradului de colonizare micoriziană pe parcursul ambilor ani de studiu, diferenţele înregistrate fiind foarte semnificativ pozitive faţă de varianta netratată.
Analiza cu ajutorul testului Duncan a indicat cel mai favorabil tratament ca fiind fungicidul aplicat în anul 2010 iar mulcirea ca fiind cea mai defavorabilă variantă.
CONCLUZII
Analiza clusterizată a strategiei colonizării fungilor micorizieni în sistemul radicular al plantelor de Festuca rubra, pe baza valorilor frecvenţei, intensităţii şi abundenţei arbusculelor, indică existenţa a două etape de colonizare. O etapă marcată de un proces de înmagazinare a substanţelor nutritive, producere de vezicule şi colonizare superficială şi o a doua etapă caracterizată de procese de transfer al elementelor nutritive şi extinderea reţelei hifale intraradiculare.
Gradul de colonizare în sistemul radicular al speciei Festuca rubra, la nivelul unui interval de 150 de zile de la răsărirea plantelor, atinge valoarea maximă la 20 de zile si valoarea minimă la 140 de zile.
Pe durata de 150 de zile a perioadei de vegetaţie, analizată pe secvenţe de 10 zile, valoarea maximă a abundenţei arbusculilor la specia Trifolium repens a fost atinsă la secvenţa de 110 zile, aspect care indică un transfer sporit de elemente nutritive comparativ cu valoarea minimă înregistrată la secvenţa de 140 de zile de la răsărirea plantelor care indică o lipsă a acestui transfer, fenomenul de maxim şi minim al transferului arbuscular având loc în etapa finală de dezvoltare a plantelor.
Analiza clusterizată a strategiei colonizării fungilor micorizieni în sistemul radicular al speciei Trifolium repens, în mediu controlat, pe baza valorilor frecvenţei, intensităţii şi abundenţei arbusculilor observate pe parcursul unei perioade de vegetaţie de 150 de zile indică existenţa unui cluster A de colonizare, un cluster B de transfer suprapus peste o colonizare abundentă respectiv un cluster C în care s-a identificat fenomenul de reducere a permisivităţii plantei gazdă pentru agenţii de colonizare.
Valorile gradului de colonizare în sistemul radicular al speciei Trifolium repens înregistrate pe o perioadă de vegetaţie de 150 de zile, cultivată în mediu controlat, indică un maxim la nivelul secvenţei de 20

33
de zile respectiv un minim la nivelul secvenţei de 150 de zile de la răsărirea plantelor.
Comparaţia dintre valorile gradului de colonizare înregistrate la specia Trifolium repens pe o perioadă de vegetaţie de 150 de zile, în mediu controlat, sub aspectul influenţei fertilizării indică cea mai favorabilă variantă de fertilizare ca fiind cea cu eurofertil mezocalc în timp ce varianta fertilizată cu cenuşă a fost evidenţiată ca fiind cea mai defavorabilă.
Analiza influenţei complexului an, tratament şi fertilizare asupra frecvenţei colonizării sistemului radicular al plantelor de Festuca rubra, observate în câmpul experimental, indică combinaţia anul 2010 x fungicid x lipsa fertilizării ca fiind cea mai favorabilă combinaţie ca şi influenţă, respectiv combinaţia anul 2011 x sulfat de zinc x gunoi de grajd ca având cea mai defavorabilă influenţă.
Observaţiile efectuate asupra complexului de factori reprezentând anii de experimentare, tratamentele şi fertilizările la nivelul influenţei asupra intensităţii colonizării în sistemul radicular al speciei Festuca rubra au indicat combinaţia condiţiilor climatice ale anului 2011 x tratamentul cu sulfat de zinc x feretilizarea cu gunoi de grajd ca fiind cea mai defavorabilă variantă comparativ cu combinaţia condiţiile climatice ale anului 2010 x tratament cu fungicid x lipsa fertilizării care a avut cea mai favorabilă influenţă.
Complexul de factori reprezentat de anii de experimentare, tratamentele şi fertilizările privit din prisma influenţei asupra gradului de colonizare al sistemului radicular al plantelor de Festuca rubra a reliefat în urma analizei comparative influenţa cea mai favorabilă a combinaţiei condiţiilor climatice din anul 2010 x tratamentul cu fungicid x lipsa fertilizării respectiv cea mai defavorabilă influenţă regăsită la combinaţia condiţiilor anului 2011 x tratamentul cu sulfat de zinc x fertilizarea cu gunoi de grajd.
Analiza influenţei complexului an, tratament şi fertilizare asupra frecvenţei colonizării sistemului radicular al plantelor de Trifolium repens, observate în câmpul experimental, indică combinaţia anul 2010 x fungicid x lipsa fertilizării ca fiind cea mai favorabilă combinaţie respectiv combinaţia anul 2011 x sulfat de zinc x gunoi de grajd ca având cea mai defavorabilă influenţă.
Observaţiile efectuate asupra complexului de factori reprezentând anii de experimentare, tratamentele şi fertilizările la nivelul influenţei asupra intensităţii colonizării în sistemul radicular al speciei Trifolium repens au indicat combinaţia condiţţilor climatice ale anului 2011 x tratamentul cu fungicid x feretilizarea cu eurofertil mezocalc ca

34
fiind cea mai defavorabilă variantă comparativ cu combinaţia condiţiile climatice ale anului 2010 x tratament cu sulfat de zinc x fertilizarea cu cenuşă care a avut cea mai favorabilă influenţă.
Complexul de factori reprezentat de anii de experimentare, tratamentele şi fertilizările privit din prisma influenţei asupra gradului de colonizare al sistemului radicular al plantelor de Trifolium repens a reliefat în urma analizei comparative influenţa cea mai favorabilă a combinaţiei condiţiilor climatice din anul 2010 x tratamentul cu sulfat de zinc x fertilizarea cu cenuşă respectiv cea mai defavorabilă influenţă regăsită la combinaţia condiţiilor anului 2011 x tratamentul cu fungicid x fertilizarea cu eurofertil mezocalc.
BIBLIOGRAFIE SELECTIVĂ
1. Akiyama K., H. Hayashi, 2006, Strigolactones: chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 97:925–931.
2. de la Providencia I. E., F. A. de Souza, F. Fernandez, N. S. Delmas, S. Declerck, 2005, Arbuscular mycorrhizal fungi reveal distinct patterns of anastomosis formation and hyphal healing mechanisms between different phylogenic groups. New Phytologist 165, 261–271.
3. Gallaud I., 1905, Études sur les mycorrhizes endotrophes. Rev Gén Bot 17:5–48; 66–83, 123–135; 223–239; 313–325; 425–433; 479–500
4. Gavito M. E., P. A. Olsson, H. Rouhier, 2005, Temperature constraints on the growth and functioning of root organ cultures with arbuscular the mycorrhizal fungi. New Phytologist 168,179–188.
5. Gerdemann J. W., 1968, Vesicular-arbuscular mycorrhiza and plant growth. Annual Review of Phytopathology 6, 397–418.
6. Giovannetti M., C. Sbrana, L. Avio, A. S. Citernesi, C. Logi, 1993, Differential hyphal morphogenesis in arbuscular mycorrhizal fungi during pre-infection stages. New Phytologist. 125: 587–593.
7. Giovannetti M., D. Azzolini, A. S. Citernesi. 1999. Anastomosis formation and nuclear and protoplasmic exchange in arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 65:5571–5575.
8. Harley J. L., E. L. Harley, 1987, A check-list of mycorrhizan British Flora. New Phytologist. 105: 1–102.
9. Hildebrandt U., F. Ouziad, F-J. Marner, H. Bothe, 2006, The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiology Letters 254, 258–267.

35
10. Jakobsen I., 1999, Transport of phosphorus and carbon in arbuscular mycorrhizas. în: Varma A., B. Hock (eds.) Mycorrhiza: structure, function, molecular biology, 2nd edn. Springer, Heidelberg, pp 535–542.
11. Koltai H., Y. Kapulnik, 2010, Arbuscular Mycorrhizas: Physiology and Function, 2nd edn. Springer.323 pp.
12. Mohammadi Goltapeh E., Y. Rezaee Danesh, R. Prasad, A. Varma, 2008, Mycorrhizal Fungi: What We Know and What Should We Know?. In: Varma A., (eds.) Mycorrhiza: State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics, 3rd. edn. Springer, Heidelberg. pp 3 – 28.
13. Peterson R. L., H. B. Massicotte, L. H. Melville, 2004, Mycorrhizas: Anatomy and Cell Biology. CABI Publishing, Wallingford. 196 pp.
14. Smith S. E., D. Read, 2008, Mycorrhizal Symbiosis, 3ed edn. Elsevier. 800 pp.
15. Stoian H. V., V. Florian, 2009a, Mycorrhiza – Benefits, Influence, Diagnostic Method. Bulletin UASMV Agriculture. 66 (1)/2009.
16. Stoian H. V., V. Florian, 2009b, Mycorrhiza – Method of Quantification. Bulletin UASMV Agriculture. 66 (1)/2009.
17. Tamasloukht M., N. Sejalon-Delmas, A. Kluever, A. Jauneau, C. Roux, G. Becard, P. Franken, 2003, Root factors induce mitochondrial-related gene expression and fungal respiration during the developmental switch from asymbiosis to presymbiosis in the arbuscular mycorrhizal fungus Gigaspora rosea. Plant Physiol. 131:1468–1478.
18. Van der Heijden M. G. A., J. N. Klironomos, M. Ursic, P. Moutoglis, R. Streitwolf-Engel, T. Boller, A. Wiemken, I. R. Sanders, 1998, Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:69–72.
19. Varma A., (ed), 2008, Mycorrhiza: State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics, 3rd edn. Springer, Heidelberg.797 pp.
20. Wang B., Y. L. Qiu, 2006, Phylogenetic distribution and evolution of mycorrhizae in land plants. Mycorrhiza 16: 299–636.
21. *** www2.dijon.inra.fr
22. *** www.biologie.uni-hamburg.de





























